 Siming Yu1
Siming Yu1 Xingxing Yuan
Xingxing Yuan Dandan Guo
Dandan Guo- 1Department of Nephrology II, First Affiliated Hospital of Heilongjiang University of Chinese Medicine, Harbin, China
- 2School of Graduate Studies, Heilongjiang University of Chinese Medicine, Harbin, China
- 3Department of Gastroenterology, Heilongjiang Academy of Traditional Chinese Medicine, Harbin, China
- 4Department of Cardiology, Second Affiliated Hospital of Heilongjiang University of Chinese Medicine, Harbin, China
Diabetes, a multifaceted metabolic disorder, poses a significant global health burden with its increasing prevalence and associated complications, such as diabetic nephropathy, diabetic retinopathy, diabetic cardiomyopathy, and diabetic angiopathy. Recent studies have highlighted the intricate interplay between N6-methyladenosine (m6A) and non-coding RNAs (ncRNAs) in key pathways implicated in these diabetes complications, like cell apoptosis, oxidative stress, and inflammation. Thus, understanding the mechanistic insights into how m6A dysregulation impacts the expression and function of ncRNAs opens new avenues for therapeutic interventions targeting the m6A-ncRNAs axis in diabetes complications. This review explores the regulatory roles of m6A modifications and ncRNAs, and stresses the role of the m6A-ncRNA axis in diabetes complications, providing a therapeutic potential for these diseases.
1 Introduction
Diabetes mellitus, a metabolic disorder characterized by chronic hyperglycemia, represents a significant global health burden due to various long-term complications including diabetic nephropathy (DN), diabetic retinopathy (DR), diabetic cardiomyopathy (DC), and diabetic angiopathy (DA) (1, 2). The pathophysiology of these complications is complex and multifaceted, involving biochemical, molecular, and cellular changes, among which post-transcriptional modifications of non-coding RNAs (ncRNAs) have emerged as crucial areas of interest (3). One of abundant and biologically significant modifications in RNA is N6-methyladenosine (m6A), affecting multiple aspects of RNA metabolism, such as stability, splicing, and translation (4). The dynamic and reversible modification of RNA by m6A is mediated by a set of proteins known as methyltransferases, demethylases, and binding proteins, which recognize and bind to m6A sites (5). The m6A modification landscape interacts significantly with ncRNAs, a diverse class of RNAs including microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs), playing critical roles in regulating gene expression at the transcriptional and post-transcriptional levels (6, 7).
Emerging evidence underscores a pivotal role of the m6A-ncRNAs axis in regulating the pathological processes underlying diabetes complications. For instance, m6A modification influences the maturation, stability, and function of miRNAs, thereby participating in influencing cell survival in hyperglycemic state (8). Similarly, it also affects the structure and function of lncRNAs and circRNAs, which are involved in crucial regulatory pathways impacting inflammation, fibrosis, and endothelial dysfunction, which are key processes in the progression of diabetes complications (9, 10). The interplay between m6A modifications and ncRNAs opens new vistas for understanding the molecular mechanisms underpinning diabetes complications. However, the precise mechanisms by which m6A modifications affect ncRNA function in the context of diabetes are not fully elucidated, and the therapeutic potential of modulating this axis is still largely unexplored. Hence, this review concisely summarizes the regulatory mechanisms of m6A modifications and ncRNAs, and emphasizes current knowledge and recent discoveries concerning the role of the m6A-ncRNA axis in diabetes complications, aiming to provide a detailed understanding of this axis as a critical component in the pathogenesis of diabetes complications and its potential for therapeutic intervention.
2 The regulation of RNA m6A modification
The m6A modification is defined as methylation of the N6 position of adenosine and occurs in the common sequence DRACH (D = G/A/U, R = G/A, and H = A/C/U) (11, 12), which usually locates in the 3′-untranslated regions (UTRs), coding sequences, around stop codon regions, and 5′-UTRs (13, 14). This process orchestrates intricate regulatory networks on RNA, covering the reversible and dynamic interplay among methylases (writers), demethylases (erasers) and m6A-binding proteins (readers) (15).
The writers are at the forefront of m6A regulation and are responsible for the targeted deposition of m6A marks on RNA transcripts (16, 17). The core writer complex, comprised of methyltransferase-like 3 (METTL3) and METTL14, ensures the precise installation of m6A marks, underscoring the significance of this dynamic complex (16). Beyond the core writers, accessory proteins play pivotal roles in modulating m6A modification. Wilms tumor 1-associated protein (WTAP) has been identified as a key cofactor, facilitating the interaction between METTL3 and METTL14 and influencing their substrate specificity (18). Moreover, Vir-like m6A methyltransferase-associated (VIRMA, also known as KIAA1429), RNA‐binding motif protein 15 (RBM15) and its paralog RBM15B have been recognized as additional components of the m6A writer complex, expanding the regulatory landscape (19). Currently, novel methyltransferases, such as METTL4, METTL5, METTL16, and zinc finger CCCH domain‐containing protein 13 (ZC3H13), has been identified as modifiers to affect RNA metabolism in diabetes and its complications (20–22), showcasing their adaptability to diverse RNA substrates.
Equally pivotal in the m6A regulatory network are the erasers that specifically remove m6A marks. Two prominent erasers, fat mass and obesity-associated protein (FTO) and alkB homolog 5 (ALKBH5), serve as demethylases to catalyze RNA demethylation (23, 24). FTO, initially identified in the context of obesity-related processes, mediates the demethylation of m6A sites, resulting in the alteration of RNA structure and function (23, 25). ALKBH5, a member of the alkB family, specifically obliterates m6A marks in the coding region and 3’UTR of single-stranded RNA, and directly eliminate methyl groups from m6A-methylated adenosine (26). These erasers contribute to the dynamic turnover of m6A modifications, influencing various cellular processes, including mRNA stability and translation efficiency, contributing to glucose metabolic disorders in multiple organs and tissues, such as the liver, kidney, and blood vessels (27, 28).
Completing the triumvirate are the readers that recognize and interpret the m6A marks, transducing them into functional outcomes. Increasing m6A reader proteins have been discovered in mammals, including YTH domain‐containing family proteins (YTHDF1–3), YTH domain‐containing proteins (YTHDC1–2), insulin-like growth factor 2 mRNA-binding proteins (IGF2BP1–3), and heterogeneous nuclear ribonucleoproteins (HNRNPs) (29). YTHDF1 has been implicated in promoting the translation efficiency of m6A-modified mRNAs (30). YTHDF2, conversely, facilitates the degradation of m6A-modified transcripts through interactions with the RNA decay machinery, influencing mRNA stability (31). YTHDF3 cooperates with YTHDF1 and YTHDF2 to fine-tune the functions of m6A modification on gene regulation (32). Besides, YTHDC1, a nuclear m6A reader, mediates RNA splicing in the nuclear speckle and transports m6A methylated RNA from the nucleus to induce protein expression (33). YTHDC2 increases the translation efficiency of the substrate by targeting the conserved m6A motif (34). As another class of m6A readers, IGF2BPs are required for the stability of m6A-modified transcripts and enhance their translation, participating in activation of oncogenic pathways and resistance to targeted therapies (35). The HNRNP family of readers incorporates HNRNPC, HNRNPG and HNRNPA2B1, which bind to m6A sites of RNAs with high affinity. Both HNRNPC and HNRNPG affect the localization and splicing of RNAs in the nucleus (36), while HNRNPA2B1 regulates m6A modification via facilitating the processing of miRNA primers and miRNA precursors (37).
Conclusively, the dynamic interplay among m6A writers, erasers, and readers regulates fundamental cellular processes, including RNA processing, stability, and translation. In diabetes and its complications, the dysregulation of m6A modifiers is instrumental in determining the cellular RNA methylation landscape, ultimately affecting gene expression and biological processes, such as pancreatic β-cell dysfunction, insulin resistance, and abnormal lipid metabolism (5, 22, 38). As vital gene regulators, ncRNAs have also been implicated in the occurrence and progression of metabolic diseases. The intricate interplay between m6A modifiers and ncRNAs in the cellular milieu highlights the complexity of m6A modification in these diseases. Thus, targeting m6A modifiers holds substantial promise for the development of innovative therapeutics, provided that ongoing and future studies continue elucidating the mechanisms of these regulators in diabetes and its complications.
3 Regulatory mechanisms of ncRNAs
NcRNAs mainly contains miRNAs, lncRNAs, and circRNAs, playing a crucial role in the occurrence and development of diabetes complications (3, 39). NcRNAs are involved in the expression regulation of target genes via a variety of molecular mechanisms.
MiRNAs, a class of small endogenous and single-stranded ncRNAs with a length of 20–24 nucleotides, participate in regulating gene expression and affecting the development of various diseases (40). The biosynthesis of miRNAs is mediated by primary miRNA transcripts (pri-miRNAs) that are transcribed by RNA polymerase II, and pri-miRNAs are subsequently converted into precursor mRNAs (pre-mRNAs) by the Drosha–DGCR8 complex in the nucleus. Pre-miRNAs are next delivered into the cytoplasm and are further catalyzed to generate a nucleotide duplex by the RNase III enzyme Dicer, ultimately one strand of the duplex is processed by the RNA-induced silencing complex to produce a mature miRNA (41). By matching with the complementary sequences at the 3′-UTR of mRNAs of target genes, miRNAs suppress mRNAs translation or trigger their degradation, and thus regulating the post-transcriptional process of gene expression (42). In diabetes complications, the aberrant expression of miRNAs affects various biological process including cell proliferation, apoptosis, inflammation, angiogenesis, and tissue fibrosis, thereby acting as novel biomarkers and potential therapeutic targets (43, 44).
LncRNAs, a kind of transcripts of with lengths over 200 nucleotides and are transcribed by RNA polymerases, with no or limited protein-coding potential (45). Each lncRNA has distinct structures and functions, and its dynamic secondary and tertiary structures are required for their interchangeably interaction with DNA, RNAs, and proteins (45). LncRNAs have been implicated in gene expression regulation at various levels, ranging from transcription to post-translation. At the transcriptional level, lncRNAs are confirmed to modulate gene expression through interacting with chromatin-modifying enzymes and transcription factors (46, 47). Besides, at the post-transcriptional level, lncRNAs induce the inactivation or even degradation of mRNA by hindering the pre-mRNA chain splicing or directly binding to mRNA; meanwhile, they function as miRNA sponges, which is also referred to a competing endogenous RNAs (ceRNAs) mechanism, leading to the loss of regulatory function of miRNA on mRNA (48). Also, at the translation and post-translation level, lncRNAs act as protein scaffolds to mediate the assembly of translational components and form a specific spatial structure to activate or inhibit functions of proteins (49). Moreover, lncRNAs interact with signaling molecules and further master the activation of downstream signal transduction pathways (50). Abnormally expressed lncRNAs are observed in patients with various diabetes complications, which are considered to be potential diagnostic and prognostic biomarkers (51). In preclinical settings, dysregulation of lncRNA expression in animal models of diabetes complications is extensively engaged in multiple pathological processes, such as inflammation, oxidative stress, fibrosis, and microvascular dysfunction (52). Clarifying the regulatory mechanisms of lncRNAs in the pathogenesis of diabetes complications may provide new therapeutic avenues for this disease.
CircRNAs are a family of single-stranded covalently RNA molecules with the length of 100nt ~ over 4 kb, in which the closed loop is formed by the link between 5’ and 3’ terminal nucleotide sequences during splicing (53). With the help of the ligated 5′ and 3′ ends, circRNAs are able to avoid degradation mediated by exoribonuclease and are steadier compared to linear transcripts such as miRNAs and lncRNAs (54), indicating a potential for diagnostic or prognostic markers of various diseases. Like these linear RNAs, circRNAs are also derived from pre-mRNAs and are transcribed by RNA polymerase II (RNA Pol II), which are further regulated by the cis-regulatory elements and trans-acting factors, suggesting a competition with linear RNAs during their biogenesis (55). In fact, circRNAs have been verified to modulate gene expression via various mechanisms. First, they serve as miRNA sponges directly targeting miRNAs by competing with mRNA, thus modulating gene expression. Besides, they function as transcription regulators of target genes through interacting with RNA Pol II by a mechanism of cis-regulation. Finally, they serve as protein scaffolds to block protein function and inactivate related signaling pathways (56, 57). Increasing studies have proved the aberrant expression of circRNAs in patients with diabetes complications, implying its feasibility as a diagnostic or prognostic biomarker for these diseases (58, 59). Experimentally, circRNAs are susceptible to high glucose (HG)-induced tissue injury, and resultant dysregulation of their expression levels and functions contribute to the pathological process under diabetic conditions (60, 61). Therefore, circRNAs exhibit potential roles as novel biomarkers and treatment tools in diabetes complications.
To sum up, miRNAs directly target mRNAs to regulate the gene translational process. Both lncRNAs and circRNAs function as ceRNAs to sponge miRNAs, thus affecting the miRNAs-mediated gene regulation. They also directly interact with proteins, influencing protein functions and their downstream signaling pathways. The expression of ncRNAs is dysregulated in diabetes complications where ncRNAs are engaged in various biological processes, such as insulin secretion, vascular dysfunction, thermal hyperalgesia, tissue fibrosis, and cellular autophagy. As concluded above, m6A modification also participates in the onset and progression of diabetes complications through affecting these pathological processes. As a main target of m6A methylation, the modification of ncRNAs may play a fundamental role in their regulation, protein interactome and subsequent downstream effector functions (7, 62), exerting a crucial role in the initiation and progression of diabetes complications (Figure 1). In this context, further investigating roles of m6A-regulated ncRNA might provide potential diagnostic or prognostic markers for diabetes complications, as well as promising therapeutic targets to tackle this disease.
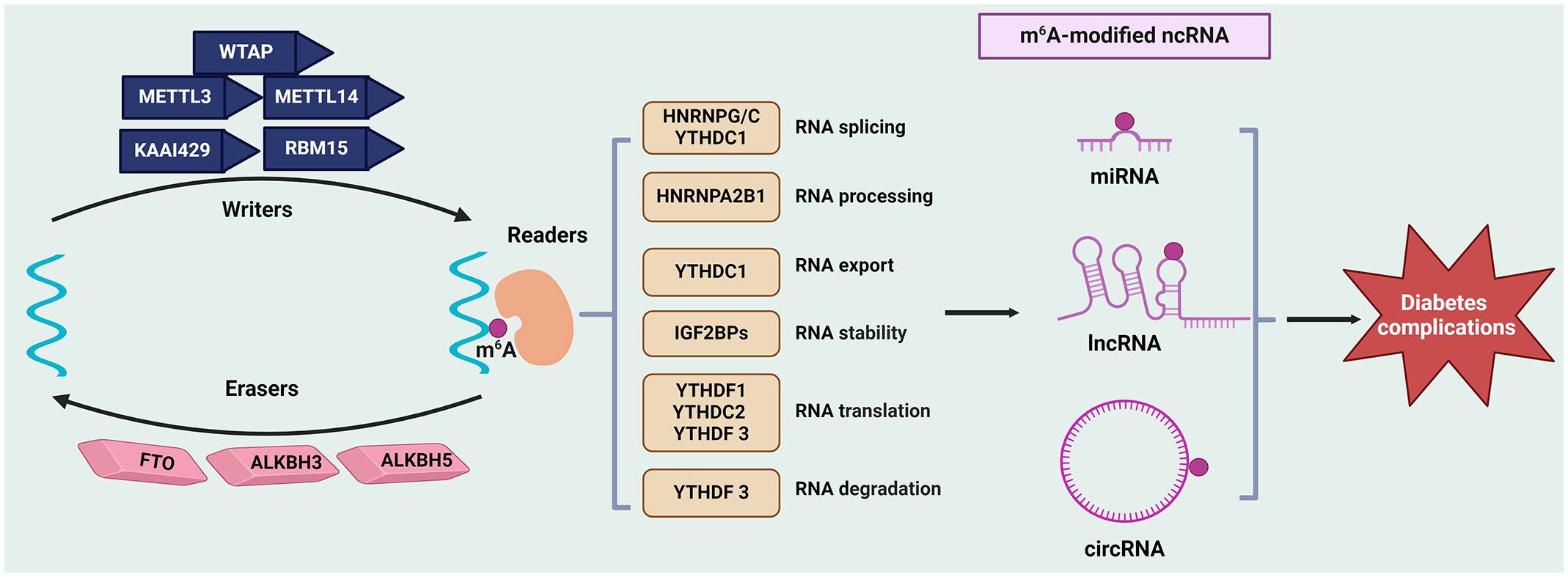
Figure 1 Regulation of ncRNAs modifications by m6A in diabetes complications. The m6A modification is mediated by writers (such as METTLs, WTAP, KAAI429, and RBM15), erasers (FTO, ALKBH5, and ALKBH3), and readers (like YTHDFs, YTHDCs, IGF2BPs and HNRNPs). This modification affects the metabolism ncRNAs (include miRNAs, lncRNAs, and circRNAs), such as their stability, splicing, and translation, thus regulating gene expression and further influencing the initiation and progression of diabetes complications. ALKBH, AlkB Homolog; circRNAs, circular RNAs; FTO, Fat Mass and Obesity-Associated Protein; HNRNPs, Heterogeneous nuclear ribonucleoproteins; lncRNAs, long non-coding RNAs; METTL, methyltransferase‐like; miRNAs, microRNAs; ncRNAs, non-coding RNAs; m6A, N6-methyladenosine; RBM15, RNA‐binding motif protein 15; KAAI429, Vir‐like m6A methyltransferase associated; WTAP, Wilms’ tumor 1-associating protein; YTHDF, YTH domain‐containing family.
4 The m6A-modified ncRNAs in the pathogenesis of diabetes complications
4.1 Diabetic nephropathy
DN stands as a prominent microvascular complication of diabetes mellitus, leading to end-stage renal disease and heightened mortality rates among affected individuals (63). Podocytes, a kind of epithelial cells that located outside the glomerular capillaries, act as a crucial component of the glomerular filtration barrier, thus podocyte injury is regarded as the core event in the occurrence and progression of DN (64). Recent research endeavors have shed light on the intricate molecular mechanisms underlying DN pathogenesis, with a particular focus on the involvement of ncRNAs and their m6A modifications. For example, METTL14 is found to promote renal tubular epithelial cell apoptosis and endoplasmic reticulum stress during DN progression by regulating the m6A modification of lncRNA TUG1. This interaction facilitates the activation of MAPK/ERK signaling, further exacerbating DN pathogenesis (65). Additionally, the upregulation of lncRNA ENST00000436340, mediated by FTO-induced m6A modification, promotes podocyte injury by facilitating the association of polypyrimidine tract binding protein 1 (PTBP1) with the Rab GTPases RAB3B mRNA, leading to cytoskeleton rearrangement and inhibition of glucose transporter 4 (GLUT4) translocation (66). Further analysis of macrophage M1-related lncRNAs in the process of m6A modification identifies several lncRNAs including LINC00342, LINC00667, and LNC00963 are regulated by FTO, RBM15, and WTAP, which is required for the immune regulation of macrophage M1 during DN progression (67). Also, m6A modification of circRNAs has emerged as a critical regulatory mechanism in DN progression. It is reported that METTL3 reduces the expression of circRNA 0000953 by upregulating its methylation level through YTHDF2, which inhibits podocyte autophagy via the miR665–3p/ATG4b (autophagy-related gene 4b) axis, thereby exacerbating podocyte injury and albuminuria in DN mice (68). Similarly, circRNA UBXN7 promotes macrophage infiltration and renal fibrosis via interactions with IGF2BP2-dependent specificity protein 1 (SP1) mRNA stability, suggesting its potential as a prognostic indicator and therapeutic target for DN management (9). These findings underscore the circRNA-mediated regulatory networks orchestrated by m6A modifications in DN pathogenesis.
Collectively, m6A-modified ncRNAs have emerged as key players in modulating various aspects of DN progression, including podocyte injury, macrophage infiltration, renal fibrosis, and inflammatory responses. Targeting these m6A-related modifiers and their associated regulatory networks may hold the key to mitigating the progression of DN and improving patient outcomes. Future research endeavors aimed at unraveling the complexities of m6A-modified ncRNAs in DN pathogenesis are warranted to facilitate the development of novel therapeutic strategies for this debilitating condition.
4.2 Diabetic retinopathy
DR represents a major microvascular complication of diabetes mellitus and a leading cause of vision loss. Recent advances in molecular biology have unveiled the critical roles of m6A modifications and ncRNAs in the pathophysiology of DR (69), which have been identified as key regulators in various biological processes, including angiogenesis, inflammation, and cell death (70). Research has elucidated multiple mechanisms through which m6A modifications of ncRNAs influence DR progression. For example, a study identified that METTL3 overexpression attenuates HG-induced pyroptosis in retinal pigment epithelial cells by regulating the miR-25–3p/PTEN/AKT signaling cascade. This process enhances cell viability and reduces oxidative stress responses, crucial for preventing DR exacerbation (8). Another aspect of METTL3 roles in DR is demonstrated through the induction of mature miR-4654, which facilitates HG-mediated apoptosis and oxidative stress in lens epithelial cells via decreasing the expression of antioxidant superoxide dismutase 2 (SOD2) (71). In addition, METTL3 mediates m6A modifications enhancing the stability of lncRNA SNHG7, which inhibits the endothelial-mesenchymal transition via downregulating the RNA-binding protein KHSRP and the transcriptional coactivator MKL1, thus mitigating DR progression (72). Another lncRNA, NEAT1 is upregulated by WTAP in DR, and further aggravates cell apoptosis and inflammation through activating the nod-like receptor pyrin domain 3 (NLRP3) inflammasome (73). Additional findings have highlighted the role of circRNA FAT1 in DR. This circRNA is confirmed to enhance autophagy and further decrease pyroptosis in retinal pigment epithelial cells under diabetic conditions through its interaction with YTHDF2 (74). Hence, the diversity of these mechanisms underscores the complexity of regulatory roles of m6A on ncRNAs and their broad therapeutic potential in DR.
In sum, the detailed mechanisms by which m6A modification controls ncRNA function and stability provide a deeper understanding of DR at a molecular level, suggesting that targeting specific m6A writers, erasers, or readers could offer new strategies for treatment. Future research should focus on elucidating more comprehensive pathways involving m6A-modified ncRNAs, validating their roles in vivo, and exploring their potential in clinical applications.
4.3 Diabetic cardiomyopathy
DC represents a severe and often irreversible cardiac dysfunction linked to diabetes, primarily characterized by fibrosis, apoptosis, and metabolic disturbances (75). It is reported that the expression of lncRNA Airn is decreased in HG-treated cardiac fibroblasts, which contributes to myocardial fibrosis and cardiac dysfunction. The anti-fibrotic effect of lncRNA Airn is mechanistically linked to its interaction with IGF2BP2, protecting p53 mRNA from degradation in an METTL3-dependent manner, thereby leading to cell cycle arrest in fibroblasts and reduced cardiac fibrosis (76). Similarly, the role of METTL14 is explored in its capacity to suppress pyroptosis and ameliorate DC progression by downregulating the lncRNA TINCR, which in turn upregulates the expression of NLRP3 by increasing its mRNA stability (77). Conversely, circRNA CDR1as is verified to promote cardiomyocyte apoptosis in DC. Mechanistically, ALKBH5 cooperates with YTHDF2 to enhance the expression of transcription factor FOXO3, which is responsible for the upregulation of circRNA CDR1as, thus inhibiting the ubiquitination of mammalian sterile 20-like kinase 1 (MST1) and further activating of the Hippo signaling pathway (78). These findings provide insights into the regulatory mechanisms of m6A modification on ncRNAs in DC.
In brief, these studies underscore the significant regulatory roles of m6A-modified ncRNAs in the pathophysiology of DC. The intricate molecular interactions point to a promising frontier in cardiac research where ncRNAs and epigenetic modifications intersect to offer novel insights and therapeutic strategies in the fight against DC. Future research should aim to expand the repertoire of ncRNAs and m6A modifications associated with DC, exploring their interactions and the downstream pathways they regulate. This could pave the way for the development of multi-target drugs that modulate several pathways simultaneously, offering a more holistic approach to managing or potentially curing DC.
4.4 Diabetic angiopathy
DA, a critical complication of diabetes, manifests predominantly as vascular dysfunction leading to morbidity and mortality among diabetic patients (79). Emerging research elucidates the significant role of m6A modifications in ncRNAs, which pivotally influence vascular cell behavior and disease progression in diabetic conditions. For instance, one such study focused on human umbilical vein endothelial cells, treated with HG and TNF-α to simulate diabetic endothelial dysfunction, revealed extensive changes in the m6A landscape of lncRNAs, which is involved in the regulation of metabolic processes and hypoxia-inducible factor 1 (HIF-1) signaling pathway, thus determining endothelial cell survival and function (80). Concurrently, another study investigated the role of circRNA YTHDC2 in vascular smooth muscle cells, a vital player in vascular integrity and remodeling (10). In this preclinical setting, circRNA YTHDC2 is overexpressed and its depletion suppresses cell proliferation and migration, thereby alleviating vascular remodeling and atherosclerosis. Furthermore, the stability of circRNA YTHDC2 is regulated via YTHDC2-mediated m6A modification; subsequently, circRNA YTHDC2 is verified to inhibit the expression of ten-eleven translocation 2 (TET2), a regulator of DNA demethylation, which prompts a dedifferentiated phenotype in vascular smooth muscle cells, contributing to diabetic vascular complications (10).
In conclusion, m6A modifications affect ncRNA function and resultant vascular cell behavior in diabetes. The intricate network of m6A-modified ncRNAs modulating key signaling pathways and cellular behaviors presents a promising therapeutic frontier (Figure 2). By targeting specific ncRNAs and related m6A modifications, it might be possible to mitigate or even reverse the pathological vascular remodeling characteristic of DA. Of interest, in type 2 diabetes patients, it has been demonstrated that HG enhances FTO mRNA expression and further decreases the m6A level, which is responsible for the upregulation of methyltransferases, suggesting that glucose participates in the dynamic regulation of m6A (81). However, whether the dynamic of HG-mediated m6A modification affects downstream ncRNAs in diabetes complications should be taken into account in future investigation.
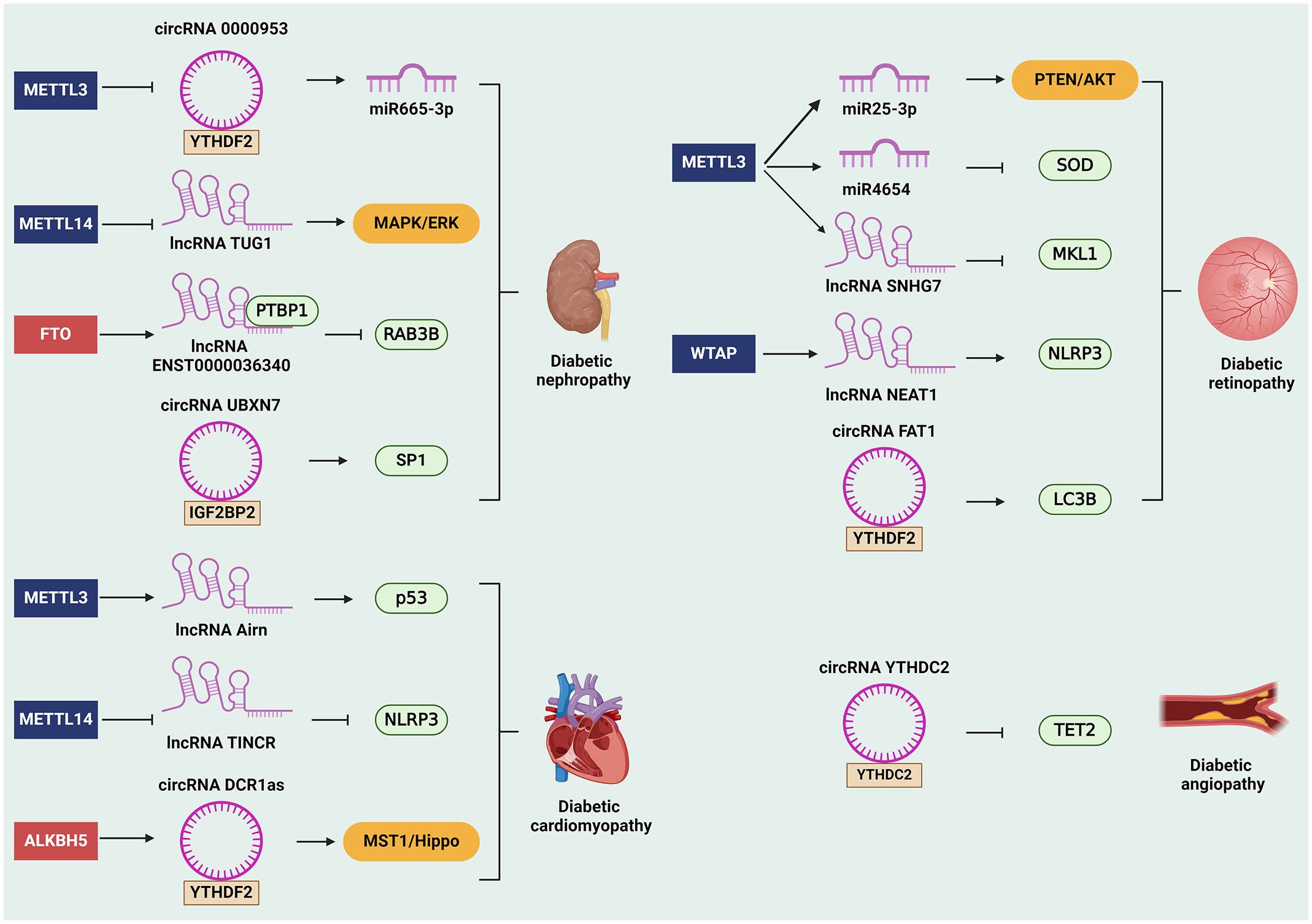
Figure 2 The role of m6A-modified ncRNAs in diabetes complications. The m6A-modified ncRNAs are involved in the occurrence and development in diabetes complications. For instance, m6A- modified ncRNAs can affect diabetic nephropathy by regulating downstream genes and signaling pathways, such as miR665–3p, RAB3B, SP1, and MAPK/ERK signaling pathway. Besides, m6A-related ncRNAs promote diabetic retinopathy by regulating SOD, MKL1, NLRP3, LC3B, and PTEN/AKT signaling pathway. Moreover, m6A-related ncRNAs facilitate the progression of diabetic cardiomyopathy via modulating the expression of p53 and NLRP3. Likewise, m6A-related ncRNAs can also affect the development of diabetic angiopathy through inhibiting the expression of TET2. ALKBH5, alkB homolog 5; circRNAs, circular RNAs; FTO, fat mass and obesity-associated protein; IGF2BP2, insulin-like growth factor 2 mRNA-binding protein 2; lncRNAs, long non-coding RNAs; m6A, N6-methyladenosine; METTL, methyltransferase-like; miRNAs, microRNAs; MST1, mammalian sterile 20-like kinase 1; ncRNAs, non-coding RNAs; NLRP3, nod-like receptor pyrin domain 3; PTBP1, polypyrimidine tract binding protein 1; SP1, specificity protein 1; SOD2, superoxide dismutase 2; TET2, ten-eleven translocation 2; WTAP, Wilms tumor 1-associated protein; YTHDF2, YTH domain‐containing family protein 2; YTHDC2, YTH domain‐containing protein 2.
5 Conclusions and future perspectives
This review synthesizes the current knowledge regarding the regulation of m6A modification on ncRNAs in diabetes and its complications, as well as emphasizes the roles of m6A-modified ncRNAs on these diseases, shedding light on novel mechanisms and therapeutic avenues. The dysregulation of m6A modifications in ncRNAs, including miRNAs, lncRNAs, and circRNAs, has emerged as a key driver in the pathophysiology of diabetic nephropathy, retinopathy, cardiomyopathy, and angiopathy. By modulating the expression and function of critical genes involved in insulin signaling, oxidative stress, inflammation, and cellular metabolism, aberrant m6A-ncRNAs axis contributes to the progression of diabetes complications. Therefore, further understanding of the molecular intricacies underlying m6A-mediated ncRNA regulation provides a foundation for the development of targeted interventions aimed at restoring homeostasis and ameliorating diabetes complications. It should be noted that therapeutic potentials of targeting the m6A-ncRNAs axis in diabetes complications lies in its ability to modulate gene expression at the post-transcriptional level with precision and specificity. Strategies targeting m6A writers (e.g., METTL3, METTL14), erasers (e.g., FTO, ALKBH5), and readers (e.g., YTHDF1–3, YTHDC1–2) offer promising avenues for therapeutic intervention. Small molecule inhibitors, RNA-based therapeutics, and CRISPR-based approaches hold immense potential in restoring the dysregulated m6A modification patterns in diabetic tissues, thereby attenuating disease progression and improving clinical outcomes.
However, several challenges and opportunities for future research warrant consideration. Firstly, the tissue-specific regulation of m6A modifications in ncRNAs underscores the importance of understanding context-dependent effects on diabetes complications. Comprehensive profiling of m6A landscapes across different tissues and cell types will enhance our understanding of spatial and temporal dynamics in diabetic pathogenesis. Secondly, elucidating the crosstalk between m6A-modified ncRNAs and other epigenetic mechanisms, such as DNA methylation and histone modifications, may uncover synergistic or antagonistic interactions driving diabetes complications. Hence, harnessing the regulatory potential of m6A modifications in ncRNAs might reshape the landscape of diabetes care, advancing towards precision medicine tailored to the individual needs of diabetic patients.
Author contributions
SY: Writing – original draft, Conceptualization. CL: Writing – review & editing. XL: Writing – review & editing. ZH: Writing – review & editing. YL: Writing – review & editing. XY: Writing – original draft, Conceptualization. DG: Writing – original draft, Conceptualization.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research is supported by several fundings: Natural Science Foundation Project of Heilongjiang Province (No. LH2019H112), Traditional Chinese Medicine Research Project of Heilongjiang Province (No. ZHY2020-121).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
DC, diabetic cardiomyopathy; DN, diabetic nephropathy; DR, diabetic retinopathy; FTO, fat mass and obesity-associated protein; HNRNPs, heterogeneous nuclear ribonucleoproteins; HG, high glucose; IGF2BPs, insulin-like growth factor 2 mRNA-binding proteins; lncRNAs, long non-coding RNAs; m6A, N6-methyladenosine; METTL3, methyltransferase-like 3; miRNAs, microRNAs; MST1, mammalian sterile 20-like kinase 1; ncRNAs, non-coding RNAs; NLRP3, nod-like receptor pyrin domain 3; PTBP1, polypyrimidine tract binding protein 1; RBM15, RNA‐binding motif protein 15; RNA Pol II, RNA polymerase II; SP1, specificity protein 1; SOD2, superoxide dismutase 2; TET2, ten-eleven translocation 2; UTRs, untranslated regions; VIRMA, Vir-like m6A methyltransferase-associated; WTAP, Wilms tumor 1-associated protein; YTHDFs, YTH domain‐containing family proteins; YTHDCs, YTH domain‐containing proteins; ZC3H13, zinc finger CCCH domain‐containing protein 13.
References
1. Cole JB, Florez JC. Genetics of diabetes mellitus and diabetes complications. Nat Rev Nephrol. (2020) 16:377–90. doi: 10.1038/s41581-020-0278-5
2. Demir S, Nawroth PP, Herzig S, Ekim Üstünel B. Emerging targets in type 2 diabetes and diabetic complications. Adv Sci. (2021) 8:e2100275. doi: 10.1002/advs.202100275
3. Chang W, Li M, Song L, Miao S, Yu W, Wang J. Noncoding RNAs from tissue-derived small extracellular vesicles: Roles in diabetes and diabetic complications. Mol Metab. (2022) 58:101453. doi: 10.1016/j.molmet.2022.101453
4. Cheng C, Yu F, Yuan G, Jia J. Update on N6-methyladenosine methylation in obesity-related diseases. Obes (Silver Spring). (2024) 32:240–51. doi: 10.1002/oby.23932
5. Wu Y, Zeng Y, Ren Y, Yu J, Zhang Q, Xiao X. Insights into RNA N6-methyladenosine in glucose and lipid metabolic diseases and their therapeutic strategies. Endocrinology. (2023) 165:170. doi: 10.1210/endocr/bqad170
6. Chen Y, Lin Y, Shu Y, He J, Gao W. Interaction between N(6)-methyladenosine (m(6)A) modification and noncoding RNAs in cancer. Mol Cancer. (2020) 19:94. doi: 10.1186/s12943-020-01207-4
7. Dai F, Wu Y, Lu Y, An C, Zheng X, Dai L, et al. Crosstalk between RNA m(6)A modification and non-coding RNA contributes to cancer growth and progression. Mol Ther Nucleic Acids. (2020) 22:62–71. doi: 10.1016/j.omtn.2020.08.004
8. Zha X, Xi X, Fan X, Ma M, Zhang Y, Yang Y. Overexpression of METTL3 attenuates high-glucose induced RPE cell pyroptosis by regulating miR-25-3p/PTEN/Akt signaling cascade through DGCR8. Aging (Albany NY). (2020) 12:8137–50. doi: 10.18632/aging.v12i9
9. Lin Z, Lv D, Liao X, Peng R, Liu H, Wu T, et al. CircUBXN7 promotes macrophage infiltration and renal fibrosis associated with the IGF2BP2-dependent SP1 mRNA stability in diabetic kidney disease. Front Immunol. (2023) 14:1226962. doi: 10.3389/fimmu.2023.1226962
10. Yuan J, Liu Y, Zhou L, Xue Y, Lu Z, Gan J. YTHDC2-mediated circYTHDC2 N6-methyladenosine modification promotes vascular smooth muscle cells dysfunction through inhibiting ten-eleven translocation 2. Front Cardiovasc Med. (2021) 8:686293. doi: 10.3389/fcvm.2021.686293
11. Zhang H, Shi X, Huang T, Zhao X, Chen W, Gu N, et al. Dynamic landscape and evolution of m6A methylation in human. Nucleic Acids Res. (2020) 48:6251–64. doi: 10.1093/nar/gkaa347
12. Desrosiers R, Friderici K, Rottman F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc Natl Acad Sci U.S.A. (1974) 71:3971–5. doi: 10.1073/pnas.71.10.3971
13. Wu J, Frazier K, Zhang J, Gan Z, Wang T, Zhong X. Emerging role of m(6) A RNA methylation in nutritional physiology and metabolism. Obes Rev. (2020) 21:e12942. doi: 10.1111/obr.12942
14. Ke S, Alemu EA, Mertens C, Gantman EC, Fak JJ, Mele A, et al. A majority of m6A residues are in the last exons, allowing the potential for 3' UTR regulation. Genes Dev. (2015) 29:2037–53. doi: 10.1101/gad.269415.115
15. Benak D, Benakova S, Plecita-Hlavata L, Hlavackova M. The role of m(6)A and m(6)Am RNA modifications in the pathogenesis of diabetes mellitus. Front Endocrinol. (2023) 14:1223583. doi: 10.3389/fendo.2023.1223583
16. Wang X, Feng J, Xue Y, Guan Z, Zhang D, Liu Z, et al. Structural basis of N(6)-adenosine methylation by the METTL3-METTL14 complex. Nature. (2016) 534:575–8. doi: 10.1038/nature18298
17. Huang J, Yin P. Structural insights into N(6)-methyladenosine (m(6)A) modification in the transcriptome. Genomics Proteomics Bioinf. (2018) 16:85–98. doi: 10.1016/j.gpb.2018.03.001
18. Ping XL, Sun BF, Wang L, Xiao W, Yang X, Wang WJ, et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. (2014) 24:177–89. doi: 10.1038/cr.2014.3
19. Yue Y, Liu J, Cui X, Cao J, Luo G, Zhang Z, et al. VIRMA mediates preferential m(6)A mRNA methylation in 3'UTR and near stop codon and associates with alternative polyadenylation. Cell Discovery. (2018) 4:10. doi: 10.1038/s41421-018-0019-0
20. Zhang H, Gu Y, Gang Q, Huang J, Xiao Q, Ha X. N6-methyladenosine RNA modification: an emerging molecule in type 2 diabetes metabolism. Front Endocrinol (Lausanne). (2023) 14:1166756. doi: 10.3389/fendo.2023.1166756
21. Li YL, Li L, Liu YH, Hu LK, Yan YX. Identification of metabolism-related proteins as biomarkers of insulin resistance and potential mechanisms of m(6)A modification. Nutrients. (2023) 15:1839. doi: 10.3390/nu15081839
22. Li YL, Zhang Y, Chen N, Yan YX. The role of m(6)A modification in type 2 diabetes: A systematic review and integrative analysis. Gene. (2024) 898:148130. doi: 10.1016/j.gene.2024.148130
23. Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol. (2011) 7:885–7. doi: 10.1038/nchembio.687
24. Ueda Y, Kitae K, Ooshio I, Fusamae Y, Kawaguchi M, Jingushi K, et al. A real-time PCR-based quantitative assay for 3-methylcytosine demethylase activity of ALKBH3. Biochem Biophys Rep. (2016) 5:476–81. doi: 10.1016/j.bbrep.2016.02.007
25. Gerken T, Girard CA, Tung YC, Webby CJ, Saudek V, Hewitson KS, et al. The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science. (2007) 318:1469–72. doi: 10.1126/science.1151710
26. Zheng G, Dahl JA, Niu Y, Fedorcsak P, Huang CM, Li CJ, et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell. (2013) 49:18–29. doi: 10.1016/j.molcel.2012.10.015
27. Sun YH, Zhao TJ, Li LH, Wang Z, Li HB. Emerging role of N(6)-methyladenosine in the homeostasis of glucose metabolism. Am J Physiol Endocrinol Metab. (2024) 326:E1–e13. doi: 10.1152/ajpendo.00225.2023
28. Huang J, Yang F, Liu Y, Wang Y. N6-methyladenosine RNA methylation in diabetic kidney disease. BioMed Pharmacother. (2024) 171:116185. doi: 10.1016/j.biopha.2024.116185
29. Yang Y, Hsu PJ, Chen YS, Yang YG. Dynamic transcriptomic m(6)A decoration: writers, erasers, readers and functions in RNA metabolism. Cell Res. (2018) 28:616–24. doi: 10.1038/s41422-018-0040-8
30. Zhang Z, Theler D, Kaminska KH, Hiller M, de la Grange P, Pudimat R, et al. The YTH domain is a novel RNA binding domain. J Biol Chem. (2010) 285:14701–10. doi: 10.1074/jbc.M110.104711
31. Wang F, Bai J, Zhang X, Wang D, Zhang X, Xue J, et al. METTL3/YTHDF2 m6A axis mediates the progression of diabetic nephropathy through epigenetically suppressing PINK1 and mitophagy. J Diabetes Investig. (2024) 15:288–99. doi: 10.1111/jdi.14113
32. Shi H, Wang X, Lu Z, Zhao BS, Ma H, Hsu PJ, et al. YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. (2017) 27:315–28. doi: 10.1038/cr.2017.15
33. Kasowitz SD, Ma J, Anderson SJ, Leu NA, Xu Y, Gregory BD, et al. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PloS Genet. (2018) 14:e1007412. doi: 10.1371/journal.pgen.1007412
34. Wojtas MN, Pandey RR, Mendel M, Homolka D, Sachidanandam R, Pillai RS. Regulation of m(6)A transcripts by the 3'→5' RNA helicase YTHDC2 is essential for a successful meiotic program in the mammalian germline. Mol Cell. (2017) 68:374–87.e312. doi: 10.1016/j.molcel.2017.09.021
35. Bell JL, Wächter K, Mühleck B, Pazaitis N, Köhn M, Lederer M, et al. Insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs): post-transcriptional drivers of cancer progression? Cell Mol Life Sci. (2013) 70:2657–75. doi: 10.1007/s00018-012-1186-z
36. Zhao Y, Shi Y, Shen H, Xie W. m(6)A-binding proteins: the emerging crucial performers in epigenetics. J Hematol Oncol. (2020) 13:35. doi: 10.1186/s13045-020-00872-8
37. Alarcón CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 is a mediator of m(6)A-dependent nuclear RNA processing events. Cell. (2015) 162:1299–308. doi: 10.1016/j.cell.2015.08.011
38. Ren Y, Li Z, Li J, Liang R, Wang Z, Bai Y, et al. m(6) A mRNA methylation: Biological features, mechanisms, and therapeutic potentials in type 2 diabetes mellitus. Obes Rev. (2023) 24:e13639. doi: 10.1111/obr.13639
39. Feng X, Yang X, Zhong Y, Cheng X. The role of ncRNAs-mediated pyroptosis in diabetes and its vascular complications. Cell Biochem Funct. (2024) 42:e3968. doi: 10.1002/cbf.3968
40. Fan B, Luk AOY, Chan JCN, Ma RCW. MicroRNA and diabetic complications: A clinical perspective. Antioxid Redox Signal. (2018) 29:1041–63. doi: 10.1089/ars.2017.7318
41. Szostak J, Gorący A, Durys D, Dec P, Modrzejewski A, Pawlik A. The role of microRNA in the pathogenesis of diabetic nephropathy. Int J Mol Sci. (2023) 24:6214. doi: 10.3390/ijms24076214
42. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. (2009) 136:215–33. doi: 10.1016/j.cell.2009.01.002
43. Ismail A, El-Mahdy HA, Eldeib MG, Doghish AS. miRNAs as cornerstones in diabetic microvascular complications. Mol Genet Metab. (2023) 138:106978. doi: 10.1016/j.ymgme.2022.106978
44. Roy D, Modi A, Khokhar M, Sankanagoudar S, Yadav D, Sharma S, et al. MicroRNA 21 emerging role in diabetic complications: A critical update. Curr Diabetes Rev. (2021) 17:122–35. doi: 10.2174/1573399816666200503035035
45. Mattick JS, Rinn JL. Discovery and annotation of long noncoding RNAs. Nat Struct Mol Biol. (2015) 22:5–7. doi: 10.1038/nsmb.2942
46. Kim LK, Park SA, Nam EJ, Kim YT, Heo TH, Kim HJ. LncRNA SNHG4 modulates EMT signal and antitumor effects in endometrial cancer through transcription factor SP-1. Biomedicines. (2023) 11:1018. doi: 10.3390/biomedicines11041018
47. Butler AA, Johnston DR, Kaur S, Lubin FD. Long noncoding RNA NEAT1 mediates neuronal histone methylation and age-related memory impairment. Sci Signal. (2019) 12:9277. doi: 10.1126/scisignal.aaw9277
48. Shelke V, Kale A, Sankrityayan H, Anders HJ, Gaikwad AB. Long non-coding RNAs as emerging regulators of miRNAs and epigenetics in diabetes-related chronic kidney disease. Arch Physiol Biochem. (2024) 130:230–41. doi: 10.1080/13813455.2021.2023580
49. Vijayalalitha R, Archita T, Juanitaa GR, Jayasuriya R, Amin KN, Ramkumar KM. Role of long non-coding RNA in regulating ER stress response to the progression of diabetic complications. Curr Gene Ther. (2023) 23:96–110. doi: 10.2174/1566523222666220801141450
50. Lu X, Tan Q, Ma J, Zhang J, Yu P. Emerging role of lncRNA regulation for NLRP3 inflammasome in diabetes complications. Front Cell Dev Biol. (2021) 9:792401. doi: 10.3389/fcell.2021.792401
51. Leung A, Natarajan R. Long noncoding RNAs in diabetes and diabetic complications. Antioxid Redox Signal. (2018) 29:1064–73. doi: 10.1089/ars.2017.7315
52. Geng M, Liu W, Li J, Yang G, Tian Y, Jiang X, et al. LncRNA as a regulator in the development of diabetic complications. Front Endocrinol. (2024) 15:1324393. doi: 10.3389/fendo.2024.1324393
53. Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. (2019) 20:675–91. doi: 10.1038/s41576-019-0158-7
54. Zhang Z, Yang T, Xiao J. Circular RNAs: promising biomarkers for human diseases. EBioMedicine. (2018) 34:267–74. doi: 10.1016/j.ebiom.2018.07.036
55. Tu C, Wang L, Wei L, Jiang Z. The role of circular RNA in Diabetic Nephropathy. Int J Med Sci. (2022) 19:916–23. doi: 10.7150/ijms.71648
56. Shu H, Zhang Z, Liu J, Chen P, Yang C, Wu Y, et al. Circular RNAs: An emerging precise weapon for diabetic nephropathy diagnosis and therapy. BioMed Pharmacother. (2023) 168:115818. doi: 10.1016/j.biopha.2023.115818
57. Zhou WY, Cai ZR, Liu J, Wang DS, Ju HQ, Xu RH. Circular RNA: metabolism, functions and interactions with proteins. Mol Cancer. (2020) 19:172. doi: 10.1186/s12943-020-01286-3
58. Wan H, Zhao S, Zeng Q, Tan Y, Zhang C, Liu L, et al. CircRNAs in diabetic cardiomyopathy. Clin Chim Acta. (2021) 517:127–32. doi: 10.1016/j.cca.2021.03.001
59. Zhang JR, Sun HJ. Roles of circular RNAs in diabetic complications: From molecular mechanisms to therapeutic potential. Gene. (2020) 763:145066. doi: 10.1016/j.gene.2020.145066
60. Fan W, Pang H, Xie Z, Huang G, Zhou Z. Circular RNAs in diabetes mellitus and its complications. Front Endocrinol. (2022) 13:885650. doi: 10.3389/fendo.2022.885650
61. Zhou HR, Kuang HY. Circular RNAs: Novel target of diabetic retinopathy. Rev Endocr Metab Disord. (2021) 22:205–16. doi: 10.1007/s11154-021-09646-0
62. Coker H, Wei G, Brockdorff N. m6A modification of non-coding RNA and the control of mammalian gene expression. Biochim Biophys Acta Gene Regul Mech. (2019) 1862:310–8. doi: 10.1016/j.bbagrm.2018.12.002
63. Naaman SC, Bakris GL. Diabetic nephropathy: update on pillars of therapy slowing progression. Diabetes Care. (2023) 46:1574–86. doi: 10.2337/dci23-0030
64. Zuo Y, Wang C, Sun X, Hu C, Liu J, Hong X, et al. Identification of matrix metalloproteinase-10 as a key mediator of podocyte injury and proteinuria. Kidney Int. (2021) 100:837–49. doi: 10.1016/j.kint.2021.05.035
65. Zheng Y, Zhang Z, Zheng D, Yi P, Wang S. METTL14 promotes the development of diabetic kidney disease by regulating m(6)A modification of TUG1. Acta Diabetol. (2023) 60:1567–80. doi: 10.1007/s00592-023-02145-5
66. Hu J, Wang Q, Fan X, Zhen J, Wang C, Chen H, et al. Long noncoding RNA ENST00000436340 promotes podocyte injury in diabetic kidney disease by facilitating the association of PTBP1 with RAB3B. Cell Death Dis. (2023) 14:130. doi: 10.1038/s41419-023-05658-7
67. Li C, Su F, Liang Z, Zhang L, Liu F, Fan W, et al. Macrophage M1 regulatory diabetic nephropathy is mediated by m6A methylation modification of lncRNA expression. Mol Immunol. (2022) 144:16–25. doi: 10.1016/j.molimm.2022.02.008
68. Liu X, Jiang L, Zeng H, Gao L, Guo S, Chen C, et al. Circ-0000953 deficiency exacerbates podocyte injury and autophagy disorder by targeting Mir665-3p-Atg4b in diabetic nephropathy. Autophagy. (2023) 20:1072–97. doi: 10.1080/15548627.2023.2286128
69. Kumari N, Karmakar A, Ahamad Khan MM, Ganesan SK. The potential role of m6A RNA methylation in diabetic retinopathy. Exp Eye Res. (2021) 208:108616. doi: 10.1016/j.exer.2021.108616
70. Chang X, Zhu G, Cai Z, Wang Y, Lian R, Tang X, et al. miRNA, lncRNA and circRNA: Targeted Molecules Full of Therapeutic Prospects in the Development of Diabetic Retinopathy. Front Endocrinol. (2021) 12:771552. doi: 10.3389/fendo.2021.771552
71. Guo M, Su F, Chen Y, Su B. Methyltransferase METTL3-mediated maturation of miR-4654 facilitates high glucose-induced apoptosis and oxidative stress in lens epithelial cells via decreasing SOD2. Chem Biol Drug Des. (2024) 103:e14491. doi: 10.1111/cbdd.14491
72. Cao X, Song Y, Huang LL, Tian YJ, Wang XL, Hua LY. m(6)A transferase METTL3 regulates endothelial-mesenchymal transition in diabetic retinopathy via lncRNA SNHG7/KHSRP/MKL1 axis. Genomics. (2022) 114:110498. doi: 10.1016/j.ygeno.2022.110498
73. Guo C, Yu M, Liu J, Jia Z, Liu H, Zhao S. Molecular mechanism of Wilms tumour 1-associated protein in diabetes-related dry eye disease by mediating m6A methylation modification of lncRNA NEAT1. J Drug Target. (2024) 32:200–12. doi: 10.1080/1061186X.2023.2300682
74. Huang C, Qi P, Cui H, Lu Q, Gao X. CircFAT1 regulates retinal pigment epithelial cell pyroptosis and autophagy via mediating m6A reader protein YTHDF2 expression in diabetic retinopathy. Exp Eye Res. (2022) 222:109152. doi: 10.1016/j.exer.2022.109152
75. Tan Y, Zhang Z, Zheng C, Wintergerst KA, Keller BB, Cai L. Mechanisms of diabetic cardiomyopathy and potential therapeutic strategies: preclinical and clinical evidence. Nat Rev Cardiol. (2020) 17:585–607. doi: 10.1038/s41569-020-0339-2
76. Peng T, Liu M, Hu L, Guo D, Wang D, Qi B, et al. LncRNA Airn alleviates diabetic cardiac fibrosis by inhibiting activation of cardiac fibroblasts via a m6A-IMP2-p53 axis. Biol Direct. (2022) 17:32. doi: 10.1186/s13062-022-00346-6
77. Meng L, Lin H, Huang X, Weng J, Peng F, Wu S. METTL14 suppresses pyroptosis and diabetic cardiomyopathy by downregulating TINCR lncRNA. Cell Death Dis. (2022) 13:38. doi: 10.1038/s41419-021-04484-z
78. Shao Y, Li M, Yu Q, Gong M, Wang Y, Yang X, et al. CircRNA CDR1as promotes cardiomyocyte apoptosis through activating hippo signaling pathway in diabetic cardiomyopathy. Eur J Pharmacol. (2022) 922:174915. doi: 10.1016/j.ejphar.2022.174915
79. Xiang H, Song R, Ouyang J, Zhu R, Shu Z, Liu Y, et al. Organelle dynamics of endothelial mitochondria in diabetic angiopathy. Eur J Pharmacol. (2021) 895:173865. doi: 10.1016/j.ejphar.2021.173865
80. Shan L, Guo M, Dai Y, Wei L, Zhang W, Gao J. Comprehensive analysis of m6A modification lncRNAs in high glucose and TNF-α induced human umbilical vein endothelial cells. Med (Baltimore). (2023) 102:e33133. doi: 10.1097/MD.0000000000033133
Keywords: N6-methyladenosine, non-coding RNA, diabetic nephropathy, diabetic retinopathy, diabetic cardiomyopathy, diabetic angiopathy
Citation: Yu S, Li C, Lu X, Han Z, Li Y, Yuan X and Guo D (2024) The m6A-ncRNAs axis in diabetes complications: novel mechanism and therapeutic potential. Front. Endocrinol. 15:1426380. doi: 10.3389/fendo.2024.1426380
Received: 01 May 2024; Accepted: 10 June 2024;
Published: 24 June 2024.
Edited by:
Jean-François Tanti, INSERM U1065 Centre Méditerranéen de Médecine Moléculaire, FranceReviewed by:
Valerie Grandjean, INSERM U1065 Centre Méditerranéen de Médecine Moléculaire, FranceCopyright © 2024 Yu, Li, Lu, Han, Li, Yuan and Guo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dandan Guo, eWNoMjAyMzEyQHNpbmEuY29t; Xingxing Yuan, eXVhbnhpbmd4aW5nODBAMTYzLmNvbQ==