 Éverton L. Vogt
Éverton L. Vogt Jorge F. A. Model
Jorge F. A. Model Anapaula S. Vinagre
Anapaula S. Vinagre- Laboratório de Metabolismo e Endocrinologia Comparada (LAMEC), Departamento de Fisiologia, Federal University of Rio Grande do Sul (UFRGS), Porto Alegre, Brazil
Organotins (OTs) are considered some of the most toxic chemicals introduced into aquatic environments by anthropogenic activities. They are widely used for agricultural and industrial purposes and as antifouling additives on boat hull’s paints. Even though the use of OTs was banned in 2008, elevated levels of OTs can still be detected in aquatic environments. OTs’ deleterious effects upon wildlife and experimental animals are well documented and include endocrine disruption, immunotoxicity, neurotoxicity, genotoxicity, and metabolic dysfunction. Crustaceans are key members of zooplankton and benthic communities and have vital roles in food chains, so the endocrine-disrupting effects of tributyltin (TBT) on crustaceans can affect other organisms. TBT can disrupt carbohydrate and lipid homeostasis of crustaceans by interacting with retinoid X receptor (RXR) and crustacean hyperglycemic hormone (CHH) signaling. Moreover, it can also interact with other nuclear receptors, disrupting methyl farnesoate and ecdysteroid signaling, thereby altering growth and sexual maturity, respectively. This compound also interferes in cytochrome P450 system disrupting steroid synthesis and reproduction. Crustaceans are also important fisheries worldwide, and its consumption can pose risks to human health. However, some questions remain unanswered. This mini review aims to update information about the effects of OTs on the metabolism, growth, and reproduction of crustaceans; to compare with known effects in mammals; and to point aspects that still needs to be addressed in future studies. Since both macrocrustaceans and microcrustaceans are good models to study the effects of sublethal TBT contamination, novel studies should be developed using multibiomarkers and omics technology.
Introduction
Organotins (OTs) are organometallic compounds in which an atom of tin (Sn) is covalently bounded to one or more organic chains (1). They are considered some of the most toxic chemicals introduced into aquatic environments by anthropogenic activities (1–3). OT’s deleterious effects upon wildlife and experimental animals are well documented and include endocrine disruption, immunotoxicity, neurotoxicity, genotoxicity, and metabolic dysfunction including obesity (2, 4). Butyltins (BTs) and phenyltins, the major species of OTs, are widely used for agricultural purposes (insecticides, fungicides), in PVC industry, as industrial catalysts, and as additives on boat hull’s paints to avoid encrustations by barnacles, mussels, algae, and other aquatic invertebrates (1–3, 5, 6). Therefore, large quantities of OTs have been released into aquatic ecosystems, either directly as wastewater treatment plants or indirectly as hull’s residues, posing serious environmental risks to non-target species (5, 6). Even though the use of OTs was banned in 2008, as determined by the International Marine Organization in 2001 (7), high levels of OTs can still be detected in different matrices such as surface water, clays, quartz, amorphous silica, natural soils, sediments, and organisms (5, 6, 8–10). OT levels vary in the different matrices and in different geographical regions, since environmental factors (e.g., pH, salinity, temperature) as well as the properties of the matrices can affect their adsorption (5). Recent studies in Europe revealed that OTs are still being released into the environment as outgoing water from boat wash pads, historic paint layers of hulls, and abandoned boats (11).
Marine sediment invertebrates, such as mollusks, ascidians, and crustaceans, can accumulate OTs (6, 8, 12–15). Since mollusks and crustaceans are important fisheries worldwide, many studies on OT accumulation and toxicity were developed in these animals (16, 17). Marine bivalves (mussels, clams, and oysters) tend to accumulate higher OT levels than fishes or crustaceans (13, 14, 16). Tributyltin (TBT) and triphenyltin, the most toxic forms of OTs, are well-recognized endocrine-disrupting chemicals of mollusks causing imposex or masculinization of females in more than 200 species (4, 13, 18, 19). Fishes and marine mammals can be contaminated either by drinking or by ingesting OTs-contaminated invertebrates. Therefore, the consumption of contaminated seafood (fishes, clams, mussels, oysters, crabs, and shrimps) can pose risks to human health (4, 6, 12, 20–22).
Crustaceans form a large and diverse clade of arthropods, whose members are usually free-living aquatic animals, with some terrestrial (isopods), parasitic (fish lice, tongue worms), and sessile (barnacles) species (17, 23, 24). Small crustacean species or microcrustaceans (water flees, brine shrimps, and copepods) and larval forms of larger species of decapods (crabs, lobsters) are major constituents of the zooplankton and have a vital role in the trophic transfer of nutrients and xenobiotics (17, 22, 25, 26). Decapod crustaceans, important worldwide fisheries, are usually marine, with few freshwater (crayfishes) and terrestrial (land crabs) species (17). Since decapods live on the sea floor, they can accumulate OTs dissolved in the water, in their food, or on the sediment (8, 27, 28). However, there is still little information about the mechanisms of OTs’ effects in crustaceans. This mini review aims to update information about the effects of OTs on the metabolism, growth, and reproduction of crustaceans; to compare with known effects in mammals, and to point aspects that still needs to be addressed in future studies.
OTs Effects on the Metabolism
The main neuroendocrine center of crustaceans is the X organ–sinus gland system, located inside decapods’ eyestalk (Figure 1) (29, 30). This system is the functional counterpart of the vertebrate hypothalamus–pituitary axis, controlling many processes such as metabolism, growth, color, and reproduction (17, 29, 31, 32). It secretes neuropeptides, amines (serotonin, melatonin, and catecholamines), and opioids (enkephalins) (29, 32, 33). The most abundant neuropeptide is crustacean hyperglycemic hormone (CHH), which forms a protein family with gonad-inhibiting hormone (GIH), molt-inhibiting hormone (MIH), and mandibular organ-inhibiting hormone (MOIH). As vertebrate pituitary trophic hormones, these neuropeptides regulate other endocrine glands: gonads, androgenic gland, mandibular organ (MO), and Y organ, controlling the synthesis and secretion of other hormones (29, 32, 34).
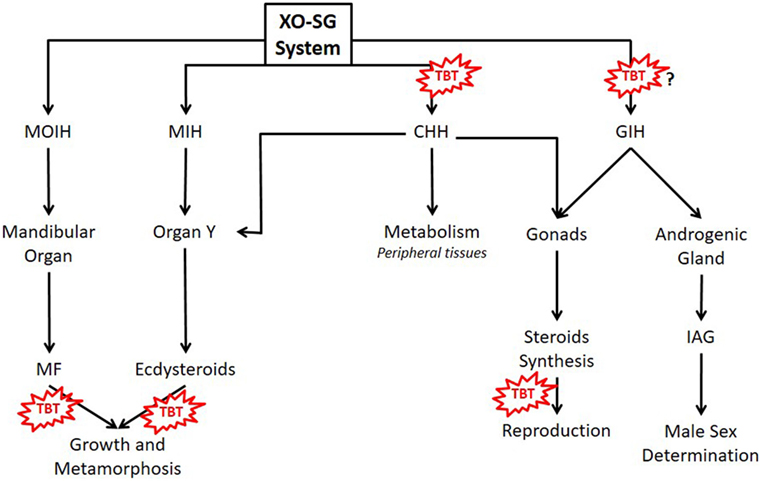
Figure 1. Main hormones controlling metabolism, growth, and reproduction of crustaceans and possible TBT’s action sites. MF, methyl farnesoate; MIH, molt-inhibiting hormone; MOIH, mandibular organ inhibitory hormone; CHH, crustacean hyperglycemic hormone; GIH, gonad-inhibiting hormone; IAG, insulin-like androgenic gland hormone; TBT, tributyltin; XO-SG, X organ–sinus gland system.
Both macrocrustaceans and microcrustaceans are considered good animal models to study xenobiotics’ ecological and toxicological effects (16, 25, 26, 35–37). Acute toxicity assays of xenobiotics, useful to assess environmental risks, usually evaluate endpoints parameters such as mortality, egg hatching, development, growth, and reproduction (16, 25, 37, 38). These endpoints are usually expressed as median-lethal or median-effect concentrations (LC50 and EC50) and no-observed-effect-level, which can be compared with predicted environmental concentrations in exposure media for purposes of risk assessment (17, 19, 39). Decapod crustaceans exhibit higher LC50 values to TBT than mysidacid shrimps, copepods, amphipods, and branchiopods (16, 26, 35, 40). This higher tolerance to TBT of decapods can be related to a faster rate of TBT elimination and/or activation (16). However, larval forms of decapods are highly sensitive to TBT (41). The LC50 for TBT of the shrimp Penaeus japonicus increased progressively during initial larval stages (nauplius to mysis) and sharply after metamorphosis (41). When the larvae were exposed to hyperosmotic or hypo-osmotic stress, the osmoregulatory capacity was compromised by TBT (41).
Organotins can enter crustacean’s hemolymph from water, sediment, or food via gills and stomach (28, 42). Once inside the animal, their fate depends on the processes of accumulation, biotransformation (metabolism), and elimination (16, 28, 42, 43). In the hermit crab Clibanarius vittatus, assimilation of a single dose of TBT from food was higher than from water, and the levels of TBT in the tissues decreased progressively after 15 days, reaching null values after 75 days (44). In this study, dibutyltin (DBT) was also detected indicating an active metabolism of TBT (44). The hepatopancreas of crustaceans is an important metabolic organ that accumulates functions equivalent to vertebrate pancreas and liver: digestive enzyme synthesis, uptake and storage of nutrients, and xenobiotic’s metabolism (42, 45–49). According to their physicochemical properties, xenobiotics can be metabolized in two distinct phases: phase I—oxidation, reduction, and hydrolysis of the substance by the cytochrome P-450 (CYP) system family of proteins; and phase II—conjugation of polar groups to become soluble (28, 42, 50). Crustaceans’ hepatopancreas have an active CYP-dependent monoxygenase system that oxidizes TBT to a series of hydroxylated derivatives that are dealkylated to form DBT and/or monobutyltin (MBT) (42, 50–53). When blue crabs Callinectes sapidus were fed with TBT-contaminated food, TBT levels in the whole abdomen peaked to 0.12 µg g−1 after 4 days of feeding, while DBT and MBT peaked to 0.39 and 0.35 µg g−1 after 8 and 12 days of feeding, respectively (54). In another study in which C. sapidus were fed TBT-contaminated food, TBT levels were higher in hepatopancreas compared to gills and muscle (43). In a third study in which C. sapidus was fed TBT-contaminated food, the respiration rate, the expression of P-450 3A (CYP3A), and heat shock proteins (HSPs) in the hepatopancreas increased, indicating that the crabs were stressed by TBT (51). An active heat shock response, specially with increased HSP70 expression, occurs when crustaceans are exposed to many types of environmental stress such as heat (55–58), metals (59, 60), and salinity alterations (61, 62). Therefore, increased expression of HSPs could be a useful indicator of BTs/TBT contamination that should be studied in other crustacean species (Figure 2).
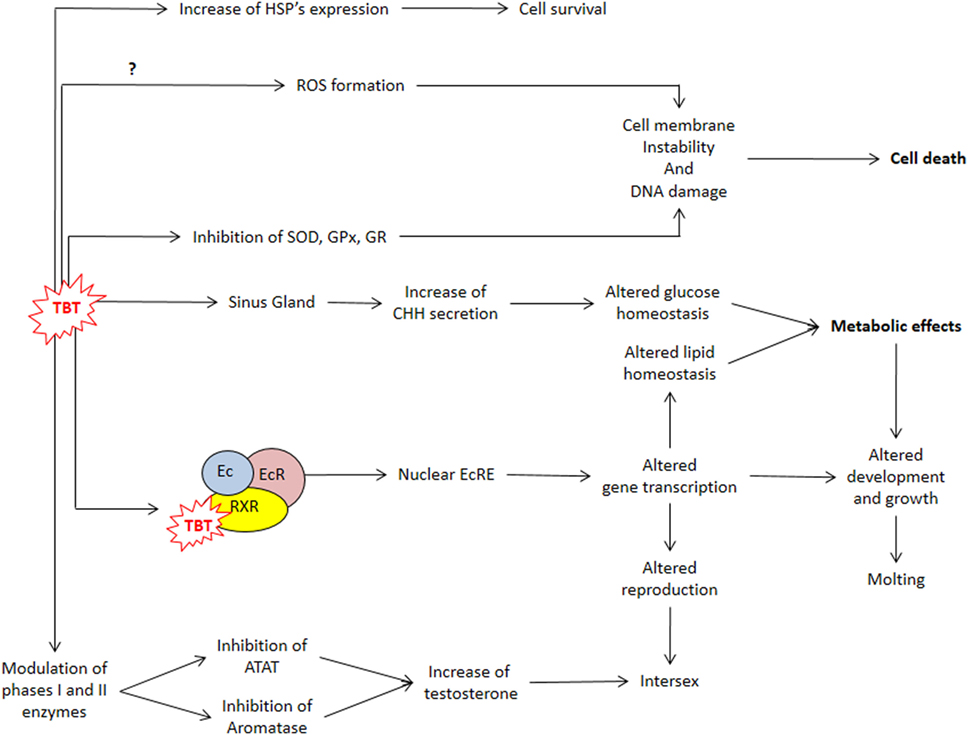
Figure 2. TBT actions impairing metabolism, cell protection, and reproduction. ATAT, acyl-CoA:testosterone acyltransferase; CHH, crustacean hyperglycemic hormone; Ec, ecdysteroid; EcR, ecdysteroid receptor; EcRE, ecdysteroid responsive element; GPx, glutathione peroxidase; GR, glutathione reductase; HSP, heat shock protein gene; ROS, reactive oxygen species; RXR, retinoid X receptor; SOD, superoxide dismutase; TBT, tributyltin.
Reactive oxygen species (ROS), byproducts of cellular respiratory chain, are kept at physiological levels by a balance between oxidant and antioxidant agents (63, 64). Liver phase I metabolism also generates ROS as byproducts, leading to oxidative stress (OS) (37). Many drugs, pesticides, and metals induce OS in crustaceans, either by altering the expression and activity of antioxidant enzymes such as catalase, superoxide dismutase (SOD), and glutathione peroxidase (GPx) or by decreasing non-enzymatic antioxidants such as glutathione (37, 65, 66). In mammals, BTs increase ROS by decreasing the concentration and activity of SOD, GPx, and glutathione reductase (GR), while simultaneously increasing lipid peroxidation in liver, testis, and kidney (67). Since decapod crustaceans, such as the green crab Carcinus maenas, C. sapidus, and Macrobrachium rosenbergii, are considered good sentinel species, OS biomarkers should be monitored in bioassays with sublethal concentrations of BTs.
Stressed animals usually develop hyperglycemia. In vertebrates, it is considered a secondary response to the increase in catecholamine and corticosteroids’ blood levels (68, 69). In crustaceans, the main hormone responsible for triggering hyperglycemia during stress is CHH (29, 34, 70, 71). Injection of 10 μmoles of tripalmitin, fentin, and fenbutatin increased glucose levels in the hemolymph of the crab Oziotelphusa senex senex (72). Since this effect did not occur in the eyestalk-ablated crabs, it is possible that OTs injection caused CHH secretion (72). In M. rosenbergii, the treatment with TBT (10, 100, and 1000 ng L−1) dissolved in water for 90 days also increased glucose levels in the hemolymph (73). Therefore, synthesis, release, and secretion of CHH and its signaling are processes that could be disrupted as the result of OTs exposure and needs to be further investigated.
In mammals, TBT disrupts both glucose and lipid homeostasis: increases body weight, inflammation, adipogenesis, and blood glucose and insulin levels (2, 74, 75). These effects are mediated by alterations in insulin signaling cascade and of nuclear receptors such as estrogen receptor, peroxisome proliferator-activated receptor γ (PPARγ), and retinoid X receptor (RXR) (2, 74, 75). RXR can form both homodimers or heterodimers with many other nuclear receptors, including PPARs, and therefore bind to DNA response elements inducing the transcription of genes involved in xenoprotection, lipid homeostasis, and development (19, 76). Since TBT is recognized as a potent agonist of RXR, this binding can be considered a key step of TBT’s mechanism of action (19, 77).
The main sites of glycogen and lipid storage in decapod crustaceans are the hepatopancreas, gonads, and muscle, and these energetic reserves fluctuate in distinct species according to seasonality, reproductive stage, molt cycle, type, and regularity of the diet (46, 49, 78). These metabolites are distinctively mobilized during diverse types of stresses, reflecting homeostasis alterations that can be used as biomarkers of health and stress condition (31, 37, 46, 47, 79). In the freshwater prawn M. rosenbergii, TBT (10, 100, and 1,000 ng L−1) treatment reduced hepatosomatic index (HIS) and the content of proteins, glycogen, and lipids in the hepatopancreas in a dose-dependent manner (73). In the cladoceran Daphnia magna, lipids are stored in spherical lipid droplets scattered throughout the body, and treatment with 0.036 or 0.36 µg L−1 increased lipid fluorescent stain (80). In female D. magna, both doses of TBT decreased the levels of triglycerides, cholesteryl esters, and phosphocolines and increased diacylglycerol levels and altered the expression of many genes, including RXR (Figure 2) (80).
OTs Effects on Growth
Crustacean growth, as in other ecdysozoans, occurs by the recapitulated molting process (81). Molting is regulated by a negative feedback mechanism involving CHH, MIH, and ecdysteroids (Figure 1) (81, 82). Ecdysone and 25-deoxyecdysone, inactive ecdysteroids, are secreted by the Y-organ and converted to 20-hydroxyecdysone (20-HE) and ponasterone A, the active forms, in peripheral tissues (33, 81). Ecdysteroids bind to arthropod ecdysteroid receptor (EcR) that complex with RXR (22, 80). The heterodimer EcR:RXR binds to ecdysteroid response element regulating the transcription of genes involved in development, growth, reproduction, and the genes involved in the pathways of ecdysone synthesis (17, 22, 80). Incomplete ecdysis leading to death occurs when D. magna is exposed to exogenous 20-HE (22). TBT alone do not alter the incidence of incomplete ecdysis; however, when in combination with 20-HE, this incidence is increased. Therefore, TBT synergizes with 20-HE leading to mortality associated with molting (22). In TBT-treated daphnids, the expression of RXR and EcR increase, disrupting the ecdysteroids’ pathways (22, 80). In the brown shrimp Cangron cangron, it was demonstrated that TBT fits in the ligand binding pocket of RXR, affecting the expression of RXR and EcR and probably of downstream genes (83). This genomic action of TBT was also demonstrated in the larvae of an insect Chironomus riparius, where TBT also increased the expression of RXR, EcR, as well as estrogen-related receptor gene and E74 (84).
Besides ecdysteroids, the sesquiterpenoids methyl farnesoate (MF) and juvenile hormone are also important during arthropod’s growth and metamorphosis (85). MF, synthesized in the MOs, is the main sesquiterpenoid of crustaceans (Figure 1) (86). The major function of MF in crustaceans is regulation of reproductive maturation (86). MF binds to methoprene-tolerant (MET), which forms a heterodimer with steroid receptor coactivator (SRC), activating the transcription of downstream genes, such as sex-determining genes involved in oocyte maturation (87). In D. magna, TBT also affected the expression of genes related to MF signaling pathway such as MET and SRC (80). Considering that TBT may also affect MF signaling in other crustaceans, and therefore alter their growth and development, serious impact on both planktonic and benthic communities can be expected.
OTs Effects on Reproduction
Imposex in female gastropods is one of the better-known effects caused by TBT on invertebrates. Imposex is characterized by the formation of male sexual organs such as penis and vas deferens in these females (19, 86). Although some studies show an early sexual reversal (intersex) in crustaceans exposed to TBT, these changes are less marked than those occurring in mollusks (31, 88). Nevertheless, other detrimental effects on the reproductive system of different species of crustaceans were found in both females and males (27, 88–90). The mechanism by which TBT causes these damages is still unclear, and there are different possible sites of action (80, 86, 89).
Unlike mollusks, when female crustaceans are exposed to TBT, there is no formation of complete male sex organs (31). Nevertheless, in M. rosenbergii, the treatment with TBT (10, 100, and 1000 ng L−1) for 45 days altered ovarian morphology and induced spermatogonia and ovotestis (with spermatocytes and structures similar to seminiferous tubules) (88). In the hermit crab C. vittatus, TBT induced several degrees of ovarian disorganization with follicular atresia and irregular oocytes although there was no formation of male sexual structures (27). Besides damage to reproductive organs, TBT may impair reproductive rates in further generations. Juvenile female D. magna exposed to TBT (100 and 1,000 ng L−1) produced smaller newborn neonates than those of unexposed females and suffered a higher mortality during their adulthood, which resulted in lower reproductive output and fitness. The reproductive rates of exposed female’s first clutch were also lower than control (80).
Although the main described effect of TBT is the masculinization of females, it also causes damage to male reproductive organs. In M. rosenbergii, exposure to TBT (10, 100, and 1,000 ng L−1) for 45 or 90 days caused several damages to the gametes and to the gonadal tissue itself. The gonadosomatic index of the testes reduced, and the seminiferous tubules architecture was compromised by an increase in connective tissue and immature cells (spermatogonia and spermatocytes) (73, 90). Spermatozoa count and length reduced (73, 90). The activity of the antioxidant enzymes SOD, GPx, and GR reduced in the testes, while DNA damage increased (89). These results are in line with studies in mammals such as the hamster Mesocricetus auratus, where TBT also caused alterations in testicular histology and reduction in spermatogenesis and in enzymatic and non-enzymatic antioxidants (67).
Since sex steroids are the major regulators of vertebrate reproduction, many steroidogenic enzymes and steroid receptors seem to have co-evolved (91, 92). However, the role of vertebrate-type sex steroids on invertebrate reproduction is not well determined (19). In mollusks, TBT-induced imposex correlates with increased free testosterone (T) levels, probably induced by inhibition of acyl-CoA:testosterone acyltransferase, which conjugates T with fatty acids, and/or CYPs, reducing T clearance (19, 93). The stimulatory effects of steroids on crustacean reproduction are well recognized; however, it was only with the development of modern omics technology that genes of steroidogenic enzymes and putative steroid receptors were identified (31, 39, 94–98). In female M. rosenbergii, TBT reduced 17β-estradiol in the hemolymph and ovary and increased T levels in the ovary (88), while in males, TBT reduced T levels in testis (73, 90) (Figure 2) (53, 94). In crustaceans, an alternative action proposed was that TBT could block T excretion, but results are still inconclusive (18, 93, 99, 100).
The synthesis and release of steroids in crustaceans is controlled mainly by GIH and CHH, released from the ES-SG system (Figure 1) (32, 39). As already mentioned, OTs can stimulate CHH release and probably also interfere with other peptides of the CHH family such as GIH (72). Gonad-stimulating hormone, released from the brain and thoracic ganglion, monoamines, and MF also participate in the control of crustacean reproduction (32, 33, 39). GIH and MIH also regulate a peptide hormone called insulin-like androgenic gland hormone, synthesized by the androgenic gland, which is responsible for male sexual differentiation (39, 97). Therefore, there are many sites where TBT may affect the neuroendocrine regulation of crustacean’s reproduction.
Conclusion
Crustaceans form a large group of aquatic animals that are important from both the economic and the ecological perspectives. They are important members of zooplankton and benthic communities and have vital roles in food chains, so the endocrine-disrupting effects of TBT on crustaceans can affect other organisms. They are also important fisheries worldwide. Therefore, human consumption of TBT-contaminated crustaceans can pose risks to human health. In summary, TBT can disrupt carbohydrate and lipid homeostasis of crustaceans by interacting with RXR and CHH signaling and can interact with other nuclear receptors, such as EcR, MET, and SRC, disrupting MF and ecdysteroid signaling, thereby altering growth and sexual maturity, respectively. This compound also interferes in cytochrome P450 system disrupting steroid synthesis and reproduction. Both macrocrustaceans and microcrustaceans are good models to study the effects of sublethal TBT contamination, usually found in natural environments. Multibiomarkers studies focusing on TBT’s effects on molecular, biochemical, cellular, morphological, physiological, and behavioral endpoints can be developed with crustaceans. The recent advances in omics technology, with the development of transcriptomes, lipidomes, and proteomes, are providing a novel set of information. The knowledge of the genes involved in the growth, development, and reproduction of crustaceans will certainly provide novel insights about TBT effects.
Author Contributions
EV wrote Sections “Introduction,” “OTs Effects on the Metabolism,” and “OTs Effects on Growth.” JM wrote Sections “OTs Effects on Reproduction” and “Conclusion” and elaborated figures. AV reviewed the manuscript.
Conflict of Interest Statement
There is no potential conflict of interest including any financial, personal or other relationships with other people or organizations related to this manuscript.
The reviewer EV and handling Editor declared their shared affiliation.
Acknowledgments
The authors gratefully acknowledge support from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the fellowship to EV and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the fellowship to JM.
Funding
This work was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
1. Hoch M. Organotin compounds in the environment: a review. Appl Geochem (2001) 16:719–43. doi:10.1016/S0883-2927(00)00067-6
2. Graceli JB, Sena GC, Lopes PFI, Zamprogno GC, da Costa MB, Godoi AFL, et al. Organotins: a review of their reproductive toxicity, biochemistry, and environmental fate. Reprod Toxicol (2013) 36:40–52. doi:10.1016/j.reprotox.2012.11.008
3. Gao J-M, Wu L, Chen Y-P, Zhou B, Guo J-S, Zhang K, et al. Spatiotemporal distribution and risk assessment of organotins in the surface water of the Three Gorges Reservoir Region, China. Chemosphere (2017) 171:405–14. doi:10.1016/j.chemosphere.2016.12.089
4. Podratz PL, Merlo E, Sena GC, Morozesk M, Bonomo MM, Matsumoto ST, et al. Accumulation of organotins in seafood leads to reproductive tract abnormalities in female rats. Reprod Toxicol (2015) 57:29–42. doi:10.1016/j.reprotox.2015.05.003
5. Fang L, Xu C, Li J, Borggaard OK, Wang D. The importance of environmental factors and matrices in the adsorption, desorption, and toxicity of butyltins: a review. Environ Sci Pollut Res (2017) 24(10):9159–73. doi:10.1007/s11356-017-8449-z
6. Lee CC, Hsu YC, Kao YT, Chen HL. Health risk assessment of the intake of butyltin and phenyltin compounds from fish and seafood in Taiwanese population. Chemosphere (2016) 164:568–75. doi:10.1016/j.chemosphere.2016.08.141
7. International Convention on the Control of Harmful Anti-fouling Systems on Ships (AFS). (2001). Available from: http://www.imo.org/en/About/Conventions/ListOfConventions/Pages/International-Convention-on-the-Control-of-Harmful-Anti-fouling-Systems-on-Ships-(AFS).aspx
8. Sant’Anna BS, Santos DM, Marchi MRR, Zara FJ, Turra A. Surface-sediment and hermit-crab contamination by butyltins in southeastern Atlantic estuaries after ban of TBT-based antifouling paints. Environ Sci Pollut Res (2014) 21:6516–24. doi:10.1007/s11356-014-2521-8
9. Lam NH, Jeong HH, Kang SD, Kim DJ, Ju MJ, Horiguchi T, et al. Organotins and new antifouling biocides in water and sediments from three Korean Special Management Sea Areas following ten years of tributyltin regulation: contamination profiles and risk assessment. Mar Pollut Bull (2017) 121(1–2):302–12. doi:10.1016/j.marpolbul.2017.06.026
10. Moreira LB, Castro ÍB, Hortellani MA, Sasaki ST, Taniguchi S, Petti MAV, et al. Effects of harbor activities on sediment quality in a semi-arid region in Brazil. Ecotoxicol Environ Saf (2017) 135:137–51. doi:10.1016/j.ecoenv.2016.09.020
11. Lagerström M, Strand J, Eklund B, Ytreberg E. Total tin and organotin speciation in historic layers of antifouling paint on leisure boat hulls. Environ Pollut (2017) 220:1333–41. doi:10.1016/j.envpol.2016.11.001
12. Jadhav S, Bhosale D, Bhosle N. Baseline of organotin pollution in fishes, clams, shrimps, squids and crabs collected from the west coast of India. Mar Pollut Bull (2011) 62:2213–9. doi:10.1016/j.marpolbul.2011.06.023
13. Park MS, Kim YD, Kim BM, Kim YJ, Kim JK, Rhee JS. Effects of antifouling biocides on molecular and biochemical defense system in the gill of the pacific oyster Crassostrea gigas. PLoS One (2016) 11:e0168978. doi:10.1371/journal.pone.0168978
14. Martinović R, Kolarević S, Kračun-Kolarević M, Kostić J, Jokanović S, Gačić Z, et al. Comparative assessment of cardiac activity and DNA damage in haemocytes of the Mediterranean mussel Mytilus galloprovincialis in exposure to tributyltin chloride. Environ Toxicol Pharmacol (2016) 47:165–74. doi:10.1016/j.etap.2016.09.019
15. Cristale J, Dos Santos DM, Sant’Anna BS, Sandron DC, Cardoso S, Turra A, et al. Tributyltin in crustacean tissues: analytical performance and validation of method. J Braz Chem Soc (2012) 23:39–45. doi:10.1590/S0103-50532012000100007
16. Tang CH, Hsu TC, Tsai CW, Wang WH. Characterization of the planktonic shrimp, Acetes intermedius, as a potential biomonitor for butyltin. J Environ Monit (2009) 11:92–9. doi:10.1039/b807864e
17. LeBlanc GA. Crustacean endocrine toxicology: a review. Ecotoxicology (2007) 16:61–81. doi:10.1007/s10646-006-0115-z
18. Oberdörster E, Rittschof D, McClellan-Green P. Testosterone metabolism in imposex and normal Ilyanassa obsoleta: comparison of field and TBTA Cl-induced imposex. Mar Pollut Bull (1998) 36:144–51. doi:10.1016/S0025-326X(97)00174-4
19. Lagadic L, Katsiadaki I, Biever R, Guiney PD, Karouna-Renier N, Schwarz T, et al. Tributyltin: Advancing the Science on Assessing Endocrine Disruption with an Unconventional Endocrine-Disrupting Compound. New York, NY: Springer New York (2017). p. 1–63.
20. Santos MM, Enes P, Reis-Henriques MA, Kuballa J, Castro LFC, Vieira MN. Organotin levels in seafood from Portuguese markets and the risk for consumers. Chemosphere (2009) 75:661–6. doi:10.1016/j.chemosphere.2008.12.066
21. Sousa ACA, Coelho SD, Pastorinho MR, Taborda-Barata L, Nogueira AJA, Isobe T, et al. Levels of TBT and other selected organotin compounds in duplicate diet samples. Sci Total Environ (2017) 574:19–23. doi:10.1016/j.scitotenv.2016.09.037
22. Wang YH, Kwon G, Li H, LeBlanc GA. Tributyltin synergizes with 20-hydroxyecdysone to produce endocrine toxicity. Toxicol Sci (2011) 123:71–9. doi:10.1093/toxsci/kfr154
23. Rota-Stabelli O, Kayal E, Gleeson D, Daub J, Boore JL, Telford MJ, et al. Ecdysozoan mitogenomics: evidence for a common origin of the legged invertebrates, the Panarthropoda. Genome Biol Evol (2010) 2:425–40. doi:10.1093/gbe/evq030
24. Edgecombe GD, Legg DA. Origins and early evolution of arthropods. Palaeontology (2014) 57:457–68. doi:10.1111/pala.12105
25. Sun PY, Foley HB, Handschumacher L, Suzuki A, Karamanukyan T, Edmands S. Acclimation and adaptation to common marine pollutants in the copepod Tigriopus californicus. Chemosphere (2014) 112:465–71. doi:10.1016/j.chemosphere.2014.05.023
26. U’Ren S. Acute toxicity of bis (tributyltin) oxide to a marine copepod. Mar Pollut Bull (1983) 14:303–6. doi:10.1016/0025-326X(83)90540-4
27. Sant’Anna BS, dos Santos DM, de Marchi MRR, Zara FJ, Turra A. Effects of tributyltin exposure in hermit crabs: Clibanarius vittatus as a model. Environ Toxicol Chem (2012) 31:632–8. doi:10.1002/etc.1724
28. Lee RF. Metabolism and accumulation of xenobiotics within hepato-pancreas cells of the blue crab, Callinectes sapidus. Mar Environ Res (1989) 28:93–7. doi:10.1016/0141-1136(89)90190-6
29. Webster SG, Keller R, Dircksen H. The CHH-superfamily of multifunctional peptide hormones controlling crustacean metabolism, osmoregulation, moulting, and reproduction. Gen Comp Endocrinol (2012) 175:217–33. doi:10.1016/j.ygcen.2011.11.035
30. Fanjul-Moles ML. Biochemical and functional aspects of crustacean hyperglycemic hormone in decapod crustaceans: review and update. Comp Biochem Physiol C Toxicol Pharmacol (2006) 142:390–400. doi:10.1016/j.cbpc.2005.11.021
31. Rodríguez EM, Medesani DA, Fingerman M. Endocrine disruption in crustaceans due to pollutants: a review. Comp Biochem Physiol A Mol Integr Physiol (2007) 146:661–71. doi:10.1016/j.cbpa.2006.04.030
32. Swetha CH, Sainath SB, Ramachandra Reddy P, Sreenivasula Reddy P. Reproductive endocrinology of female crustaceans: perspective and prospective. J Mar Sci Res Dev (2011) s3:1–13. doi:10.4172/2155-9910.S3-001
33. Girish BP, Swetha CH, Reddy PS. Induction of ecdysteroidogenesis, methyl farnesoate synthesis and expression of ecdysteroid receptor and retinoid X receptor in the hepatopancreas and ovary of the giant mud crab, Scylla serrata by melatonin. Gen Comp Endocrinol (2015) 21(7–218):37–42. doi:10.1016/j.ygcen.2015.05.007
34. Katayama H, Ohira T, Nagasawa H. Crustacean peptide hormones: structure, gene expression and function. Aqua BioScience Monogr (2013) 6:49–90. doi:10.5047/absm.2013.00602.0049
35. Sun PY, Foley HB, Bao VWW, Leung KMY, Edmands S. Variation in tolerance to common marine pollutants among different populations in two species of the marine copepod Tigriopus. Environ Sci Pollut Res (2015) 22:16143–52. doi:10.1007/s11356-015-4846-3
36. Trapp J, Armengaud J, Salvador A, Chaumot A, Geffard O. Next-generation proteomics: toward customized biomarkers for environmental biomonitoring. Environ Sci Technol (2014) 48:13560–72. doi:10.1021/es501673s
37. Rodrigues ET, Pardal MÂ. The crab Carcinus maenas as a suitable experimental model in ecotoxicology. Environ Int (2014) 70:158–82. doi:10.1016/j.envint.2014.05.018
39. Nagaraju GPC. Reproductive regulators in decapod crustaceans: an overview. J Exp Biol (2011) 214:3–16. doi:10.1242/jeb.047183
40. Oberdörster E, Rittschof D, LeBlanc GA. Alteration of [14C]-testosterone metabolism after chronic exposure of Daphnia magna to tributyltin. Arch Environ Contam Toxicol (1998) 34:21–5. doi:10.1007/s002449900281
41. Lignot JH, Pannier F, Trilles JP, Charmantier G. Effects of tributyltin oxide on survival and osmoregulation of the shrimp Penaeus japonicus (crustacea, decapoda). Aquat Toxicol (1998) 41:277–99. doi:10.1016/S0166-445X(97)00088-X
42. Lee RF. Metabolism of tributyltin by aquatic organisms. Organotin Environ Fate Eff (1996):369–82. doi:10.1007/978-94-009-1507-7_18
43. Vannuci-Silva M, Menegario AA, Franchi M, Brossi-Garcia AL, De Souza JM, De Araújo MAG, et al. Bioaccumulation of tributyltin by blue crabs. J Braz Chem Soc (2013) 24:1642–8. doi:10.5935/0103-5053.20130209
44. Sant’Anna BS, Dos Santos DM, Sandron DC, De Souza SC, De Marchi MRR, Zara FJ, et al. Hermit crabs as bioindicators of recent tributyltin (TBT) contamination. Ecol Indic (2012) 14:184–8. doi:10.1016/j.ecolind.2011.08.010
45. Vinagre AS, Da Silva RSM. Effects of fasting and refeeding on metabolic processes in the crab Chasmagnathus granulata (Dana, 1851). Can J Zool (2002) 80:1413–21. doi:10.1139/z02-122
46. Antunes F, do Amaral AP, Ribarcki FP, Wiilland Ede F, Zancan DM, Vinagre AS. Seasonal variations in the biochemical composition and reproductive cycle of the ghost crab Ocypode quadrata (Fabricius, 1787) in Southern Brazil. J Exp Zool (2010) 313:280–91. doi:10.1002/jez.593
47. Vinagre AS, Chung JS. Effects of Starvation on Energy Metabolism and Crustacean Hyperglycemic Hormone (CHH) of the Atlantic Ghost Crab Ocypode quadrata (Fabricius, 1787) Supplementary File 1: Alignment of Crustacean Hyperglycemic Hormone Sequences of Crab Species (2016).
48. Brouwer M, Lee RF. In: Guillory V, Perry H, Vanderkooy S, editors. Effects of Environmental Contaminants on the Blue Crab Callinectes sapidus. Lafayette, Louisiana: Gulf States Marine Fisheries Commission (1996). p. 1–17.
49. Sarapio E, Santos JT, Model JFA, De Fraga LS, Vinagre AS, Martins TL, et al. Glyceroneogenesis in the hepatopancreas of the crab Neohelice granulata: diet, starvation and season effects. Comp Biochem Physiol B Biochem Mol Biol (2017) 211:1–7. doi:10.1016/j.cbpb.2017.02.004
50. James MO. Isolation of cytochrome P450 from hepatopancreas microsomes of the spiny lobster, Panulirus argus, and determination of catalytic activity with NADPH cytochrome P450 reductase from vertebrate liver. Arch Biochem Biophys (1990) 282:8–17. doi:10.1016/0003-9861(90)90080-I
51. Oberdörster E, Rittschof D, McClellan-Green P. Induction of cytochrome P450 3A and heat shock protein by tributyltin in blue crab, Callinectes sapidus. Aquat Toxicol (1998) 41:83–100. doi:10.1016/S0166-445X(97)00067-2
52. Snyder M. Cytochrome P450 enzymes in aquatic invertebrates: recent advances and future directions. Aquat Toxicol (2000) 48:529–47. doi:10.1016/S0166-445X(00)00085-0
53. James MO, Boyle SM. Cytochromes P450 in crustacea. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol (1998) 121:157–72. doi:10.1016/S0742-8413(98)10036-1
54. Rice SD, Short JW, Stickle WB. Uptake and catabolism of tributyltin by blue crabs fed TBT contaminated prey. Mar Environ Res (1989) 27:137–45. doi:10.1016/0141-1136(89)90005-6
55. Cascella K, Jollivet D, Papot C, Léger N, Corre E, Ravaux J, et al. Diversification, evolution and sub-functionalization of 70kDa heat-shock proteins in two sister species of antarctic krill: differences in thermal habitats, responses and implications under climate change. PLoS One (2015) 10:e0121642. doi:10.1371/journal.pone.0121642
56. Cottin D, Foucreau N, Hervant F, Piscart C. Differential regulation of hsp70 genes in the freshwater key species Gammarus pulex (Crustacea, Amphipoda) exposed to thermal stress: effects of latitude and ontogeny. J Comp Physiol B Biochem Syst Environ Physiol (2015) 185:303–13. doi:10.1007/s00360-014-0885-1
57. Frenkel L, Dimant B, Portiansky EL, Maldonado H, Delorenzi A. Both heat shock and water deprivation trigger Hsp70 expression in the olfactory lobe of the crab Chasmagnathus granulatus. Neurosci Lett (2008) 443:251–6. doi:10.1016/j.neulet.2008.07.072
58. Xiu Y, Feng J, Lu W, Liu D, Wu T, Zhu H, et al. Identification of a novel cognate cytosolic Hsp70 gene (MnHsc70-2) from oriental river prawn Macrobrachium nipponense and comparison of its expressions with the first cognate Hsc70 (MnHsc70-1) under different stresses. Cell Stress Chaperones (2014) 19:949–61. doi:10.1007/s12192-014-0519-2
59. Mazzei V, Giannetto A, Brundo MV, Maisano M, Ferrante M, Copat C, et al. Metallothioneins and heat shock proteins 70 in Armadillidium vulgare (Isopoda, Oniscidea) exposed to cadmium and lead. Ecotoxicol Environ Saf (2015) 116:99–106. doi:10.1016/j.ecoenv.2015.03.007
60. Qian Z, Liu X, Wang L, Wang X, Li Y, Xiang J, et al. Gene expression profiles of four heat shock proteins in response to different acute stresses in shrimp, Litopenaeus vannamei. Comp Biochem Physiol C Toxicol Pharmacol (2012) 156:211–20. doi:10.1016/j.cbpc.2012.06.001
61. Fu W, Zhang F, Liao M, Liu M, Zheng B, Yang H, et al. Molecular cloning and expression analysis of a cytosolic heat shock protein 70 gene from mud crab Scylla serrata. Fish Shellfish Immunol (2013) 34:1306–14. doi:10.1016/j.fsi.2013.02.027
62. Bao XN, Mu CK, Zhang C, Wang YF, Song WW, Li RH, et al. mRNA expression profiles of heat shock proteins of wild and salinity-tolerant swimming crabs, Portunus trituberculatus, subjected to low salinity stress. Genet Mol Res (2014) 13:6837–47. doi:10.4238/2014.August.29.5
63. Espinosa-Diez C, Miguel V, Mennerich D, Kietzmann T, Sánchez-Pérez P, Cadenas S, et al. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol (2015) 6:183–97. doi:10.1016/j.redox.2015.07.008
64. Niforou K, Cheimonidou C, Trougakos IP. Molecular chaperones and proteostasis regulation during redox imbalance. Redox Biol (2014) 2:323–32. doi:10.1016/j.redox.2014.01.017
65. Rodrigues AP, Lehtonen KK, Guilhermino L, Guimarães L. Exposure of Carcinus maenas to waterborne fluoranthene: accumulation and multibiomarker responses. Sci Total Environ (2013) 443:454–63. doi:10.1016/j.scitotenv.2012.10.077
66. Aguirre-Martínez GV, Del Valls TA, Martín-Díaz ML. Identification of biomarkers responsive to chronic exposure to pharmaceuticals in target tissues of Carcinus maenas. Mar Environ Res (2013) 8(7–88):1–11. doi:10.1016/j.marenvres.2013.02.011
67. Kanimozhi V, Palanivel K, Akbarsha MA, Kadalmani B. Tributyltin-mediated hepatic, renal and testicular tissue damage in male Syrian hamster (Mesocricetus auratus): a study on impact of oxidative stress. Springerplus (2016) 5:1523. doi:10.1186/s40064-016-3186-1
68. Fabbri E, Moon TW. Adrenergic signaling in teleost fish liver, a challenging path. Comp Biochem Physiol B Biochem Mol Biol (2016) 199:74–86. doi:10.1016/j.cbpb.2015.10.002
69. Massarsky A, Trudeau VL, Moon TW. β-blockers as endocrine disruptors: the potential effects of human β-blockers on aquatic organisms. J Exp Zool Part A Ecol Genet Physiol (2011) 315 A:251–65. doi:10.1002/jez.672
70. Prymaczok NC, Pasqualino VM, Viau VE, Rodríguez EM, Medesani DA. Involvement of the crustacean hyperglycemic hormone (CHH) in the physiological compensation of the freshwater crayfish Cherax quadricarinatus to low temperature and high salinity stress. J Comp Physiol B (2016) 186:181–91. doi:10.1007/s00360-015-0954-0
71. Chung JS, Webster SG. Binding sites of crustacean hyperglycemic hormone and its second messengers on gills and hindgut of the green shore crab, Carcinus maenas: a possible osmoregulatory role. Gen Comp Endocrinol (2006) 147:206–13. doi:10.1016/j.ygcen.2006.01.002
72. Nagaraju GPC, Basha MR, Reddy PS. Organotin-induced hyperglycemia in the crab, Oziotelphusa senex senex fabricius. Zeitschrift fur Naturforsch (2001) 56:315–7. doi:10.1515/znc-2001-3-425
73. Revathi P, Iyapparaj P, Vasanthi LA, Munuswamy N, Krishnan M. Ultrastructural changes during spermatogenesis, biochemical and hormonal evidences of testicular toxicity caused by TBT in freshwater prawn Macrobrachium rosenbergii (De Man, 1879). Environ Toxicol (2014) 29:1171–81. doi:10.1002/tox.21848
74. Li B, Guo J, Xi Z, Xu J, Zuo Z, Wang C. Tributyltin in male mice disrupts glucose homeostasis as well as recovery after exposure: mechanism analysis. Arch Toxicol (1961) 91:3261–9. doi:10.1007/s00204-017-1961-6
75. Bertuloso BD, Podratz PL, Merlo E, de Araújo JFP, Lima LCF, de Miguel EC, et al. Tributyltin chloride leads to adiposity and impairs metabolic functions in the rat liver and pancreas. Toxicol Lett (2015) 235:45–59. doi:10.1016/j.toxlet.2015.03.009
76. Evans RM, Mangelsdorf DJ. Nuclear receptors, RXR, and the big bang. Cell (2014) 157:255–66. doi:10.1016/j.cell.2014.03.012
77. Nakanishi T. Endocrine disruption induced by organotin compounds; organotins function as a powerful agonist for nuclear receptors rather than an aromatase inhibitor. J Toxicol Sci (2008) 33:269–76. doi:10.2131/jts.33.269
78. Vinagre AS, Da Silva RS. Effects of starvation on the carbohydrate and lipid metabolism in crabs previously maintained on a high protein or carbohydrate-rich diet. Camp Biochem Physiol (1992) 102:579–83. doi:10.1016/0300-9629(92)90213-A
79. Filiciotto F, Vazzana M, Celi M, Maccarrone V, Ceraulo M, Buffa G, et al. Behavioural and biochemical stress responses of Palinurus elephas after exposure to boat noise pollution in tank. Mar Pollut Bull (2014) 84:104–14. doi:10.1016/j.marpolbul.2014.05.029
80. Jordão R, Casas J, Fabrias G, Campos B, Piña B, Lemos MFL, et al. Obesogens beyond vertebrates: lipid perturbation by tributyltin in the crustacean Daphnia magna. Environ Health Perspect (2015) 123:813–9. doi:10.1289/ehp.1409163
81. Techa S, Chung JS. Ecdysteroids regulate the levels of molt-inhibiting hormone (MIH) expression in the blue crab, Callinectes sapidus. PLoS One (2015) 10:e0117278. doi:10.1371/journal.pone.0117278
82. Lv J, Zhang L, Liu P, Li J. Transcriptomic variation of eyestalk reveals the genes and biological processes associated with molting in Portunus trituberculatus. PLoS One (2017) 12(4):e0175315. doi:10.1371/journal.pone.0175315
83. Verhaegen Y, Parmentier K, Swevers L, Renders E, Rougé P, De Coen W, et al. The heterodimeric ecdysteroid receptor complex in the brown shrimp Crangon crangon: EcR and RXR isoform characteristics and sensitivity towards the marine pollutant tributyltin. Gen Comp Endocrinol (2011) 172:158–69. doi:10.1016/j.ygcen.2011.02.019
84. Morales M, Martínez-Paz P, Ozáez I, Martínez-Guitarte JL, Morcillo G. DNA damage and transcriptional changes induced by tributyltin (TBT) after short in vivo exposures of Chironomus riparius (Diptera) larvae. Comp Biochem Physiol C Toxicol Pharmacol (2013) 158:57–63. doi:10.1016/j.cbpc.2013.05.005
85. Miyakawa H, Toyota K, Sumiya E, Iguchi T. Comparison of JH signaling in insects and crustaceans. Curr Opin Insect Sci (2014) 1:81–7. doi:10.1016/j.cois.2014.04.006
86. Matthiessen P, Gibbs PE. Critical appraisal of the evidence for tributyltin-mediated endocrine disruption in mollusks. Environ Toxicol Chem (1998) 17:37–43. doi:10.1002/etc.5620170106
87. Miyakawa H, Toyota K, Hirakawa I, Ogino Y, Miyagawa S, Oda S, et al. A mutation in the receptor Methoprene-tolerant alters juvenile hormone response in insects and crustaceans. Nat Commun (2013) 4:1856–7. doi:10.1038/ncomms2868
88. Revathi P, Iyapparaj P, Vasanthi LA, Munuswamy N, Prasanna VA, Suganya T, et al. TBT effects on the development of intersex (Ovotestis) in female fresh water prawn Macrobrachium rosenbergii. Biomed Res Int (2014) 2014:412619. doi:10.1155/2014/412619
89. Rani KU, Musthafa MS, War M, Al-Sadoon MK, Paray BA, Shareef THMA, et al. Impact of tributyltin on antioxidant and DNA damage response in spermatozoa of freshwater prawn Macrobrachium rosenbergii. Environ Sci Pollut Res (2015) 22:20000–6. doi:10.1007/s11356-015-5202-3
90. Revathi P, Iyapparaj P, Arockia Vasanthi L, Munuswamy N, Arun Prasanna V, Pandiyarajan J, et al. Influence of Short Term Exposure of TBT on the Male Reproductive Activity in Freshwater Prawn Macrobrachium rosenbergii (De Man). Bull Environ Contam Toxicol (2014) 93:446–51. doi:10.1007/s00128-014-1332-4
91. Thornton JW. Evolution of vertebrate steroid receptors from an ancestral estrogen receptor by ligand exploitation and serial genome expansions. Proc Natl Acad Sci U S A (2001) 98:5671–6. doi:10.1073/pnas.091553298
92. Baker ME, Nelson DR, Studer RA. Origin of the response to adrenal and sex steroids: roles of promiscuity and co-evolution of enzymes and steroid receptors. J Steroid Biochem Mol Biol (2015) 151:12–24. doi:10.1016/j.jsbmb.2014.10.020
93. Gooding MP, LeBlanc GA. Biotransformation and disposition of testosterone in the eastern mud snail Ilyanassa obsoleta. Gen Comp Endocrinol (2001) 122:172–80. doi:10.1006/gcen.2001.7630
94. Thongbuakaew T, Siangcham T, Suwansa-Ard S, Elizur A, Cummins SF, Sobhon P, et al. Steroids and genes related to steroid biosynthesis in the female giant freshwater prawn, Macrobrachium rosenbergii. Steroids (2016) 107:149–60. doi:10.1016/j.steroids.2016.01.006
95. Gao J, Wang X, Zou Z, Jia X, Wang Y, Zhang Z. Transcriptome analysis of the differences in gene expression between testis and ovary in green mud crab (Scylla paramamosain). BMC Genomics (2014) 15:585. doi:10.1186/1471-2164-15-585
96. Meng XL, Liu P, Jia FL, Li J, Gao BQ. De novo transcriptome analysis of Portunus trituberculatus ovary and testis by RNA-Seq: identification of genes involved in gonadal development. PLoS One (2015) 10:e0128659. doi:10.1371/journal.pone.0128659
97. Chandler JC, Aizen J, Fitzgibbon QP, Elizur A, Ventura T. Applying the power of transcriptomics: understanding male sexual development in decapod crustacea. Integr Comp Biol (2016) 56:1144–56. doi:10.1093/icb/icw007
98. Janer G, LeBlanc GA, Porte C, Wu X, Chen H, Liu Z, et al. Immunorecognition and distribution of progesterone receptors in the chinese mitten crab Eriocheir sinensis during ovarian development. Ann N Y Acad Sci (2014) 33:35–43. doi:10.2983/035.033.0105
99. Verslycke T, Vercauteren J, Devos C, Moens L, Sandra P, Janssen CR. Cellular energy allocation in the estuarine mysid shrimp Neomysis integer (Crustacea: Mysidacea) following tributyltin exposure. J Exp Mar Bio Ecol (2003) 288:167–79. doi:10.1016/S0022-0981(03)00006-6
Keywords: crustaceans, organotins, endocrine disruption, growth, metabolism, reproduction
Citation: Vogt ÉL, Model JFA and Vinagre AS (2018) Effects of Organotins on Crustaceans: Update and Perspectives. Front. Endocrinol. 9:65. doi: 10.3389/fendo.2018.00065
Received: 06 December 2017; Accepted: 13 February 2018;
Published: 27 February 2018
Edited by:
Leandro Miranda-Alves, Universidade Federal do Rio de Janeiro, BrazilReviewed by:
Taisen Iguchi, National Institute for Basic Biology, JapanGabriela Hollmann, Universidade Federal do Rio de Janeiro, Brazil
Copyright: © 2018 Vogt, Model and Vinagre. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Éverton L. Vogt, ZXZ2b2d0QGdtYWlsLmNvbQ==