 Lindsey Gillson1*†
Lindsey Gillson1*† Estelle Razanatsoa1†
Estelle Razanatsoa1† Andriantsilavo Hery Isandratana Razafimanantsoa1†
Andriantsilavo Hery Isandratana Razafimanantsoa1† Malika Virah-Sawmy2†
Malika Virah-Sawmy2† Anneli Ekblom3†
Anneli Ekblom3†- 1Plant Conservation Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, South Africa
- 2Sensemakers Collective, Berlin, Germany
- 3Department of Archaeology and Ancient History, University of Uppsala, Uppsala, Sweden
Planting trees is proposed as an important climate mitigation tool, but can be detrimental to biodiversity and livelihoods if not carefully planned and managed, with landscape history and livelihoods in mind. In Madagascar, deforestation is of concern, and a threat to forest-adapted biota. However, much of Madagascar’s landscape harbours ancient mosaic and open ecosystems that are home to unique suites of flora and fauna and provide a wide range of ecosystem services. Though guidelines for ecologically and socially responsible reforestation are emerging, the potential role of landscape history and palaeoecology has been generally underemphasised. Here, using Madagascar as a case study, we argue that forest restoration projects need a sound understanding of landscape history that includes a greater integration of palaeoecological data. This would help establish the former composition and extent of forests and also investigate the antiquity of open and mosaic ecosystems. When economic interests are strong, information from palaeoecology and environmental history can help reduce biases when identifying appropriate locations and suites of species for forestation. Furthermore, a reflective approach to landscape history can contribute to restoration projects that integrate cultural and livelihood considerations. A transdisciplinary approach that considers local needs and cultural context can facilitate the design and implementation of restoration projects that share benefits equitably. Underpinning this ambition is a more comprehensive consideration of ecosystem service benefits in a changing climate that includes accurate carbon storage calculations, as well as other ecosystem services including water provision, soil formation and erosion prevention, grazing resources, medicine and cultural components.
1 Introduction
Madagascar is a biodiversity hotspot (Myers et al., 2000; Antonelli et al., 2022) and a focus of conservation effort for several decades (Waeber et al., 2016). The island is one of the countries that have received the most external carbon mitigation and afforestation funding (Neudert et al., 2017). Yet, assessment of the last decades shows a discouraging lack of success in terms of both biodiversity protection and local livelihoods (Tsayem Demaze, 2014; Neudert et al., 2017). Analysis of historical national tree cover maps and satellite imagery show deforestation and forest fragmentation in several areas of Madagascar since the 1950s, with an increasing rate since 2005 (Vieilledent et al., 2018). In March 2019, the island’s new presidency announced a reforestation plan at the One Planet summit in Nairobi, committing to reforest at least 40,000 hectares per year for the next five years (Mandimbisoa, 2021). The goal is to restore 4 million ha of “degraded forests” in the island by 2030 within the northwest, east and central highlands regions (Lacroix et al., 2016; Ranjatson et al., 2019).
While reforestation of degraded forest areas can have biodiversity and carbon benefits if carefully managed, such initiatives will also have negative consequences for both people and biodiversity if they extend beyond formerly forested areas, or if inappropriate species are chosen (Bond et al., 2019; Vetter, 2020). Inadequate restoration may result when there are large economic interests at stake, for example for carbon or mining projects. Furthermore, the benefits of tree-planting risks being over-estimated if only above-ground biomass is considered, as most carbon is below ground (e.g. see Dass et al., 2018; Razafindrakoto et al., 2018; Veldman et al., 2019). In recent years, guidelines have emerged that advocate more ecologically sound and socially inclusive approaches to tree-planting, and that consider ecological relevance, drivers of forest loss, and social dimensions (e.g. see Brancalion and Holl, 2020; Di Sacco et al., 2021). However, though several such guidelines highlight the importance of distinguishing reforestation of degraded forests from the afforestation of landscapes in areas not formerly forested (Veldman et al., 2015; Boissière et al., 2021; Di Sacco et al., 2021), there has been little attempt to address how that distinction is established. Where afforestation is defined, it usually refers to tree plantations on land that did not harbor forest in the past 50 years (Bredemeier and Dohrenbusch, 2009). Yet, this baseline in the 1970s is unlikely to be an accurate reflection of the long-term history and ecological character of landscapes. Many landscapes have been radically transformed at least since the “great acceleration” of human impact in the 1950s, and most likely since the dramatic changes of the C18th and C19th including, industrialization and agricultural intensification, and European colonization (Dietl et al., 2015; Barak et al., 2016; Falk et al., 2019; Gillson et al., 2021).
Tree planting in areas that were not historically forested may negatively impact biodiversity and ecosystem services provided by ancient open and mosaic landscapes (Bond et al., 2019; Vetter, 2020; García-Dory et al., 2021; Lehmann et al., 2021). Afforestation projects commonly use exotic species and compromise other ecosystem services including water provision, grazing/forage, and biodiversity (Stickler et al., 2009; Bond et al., 2019; García-Dory et al., 2021). Loss of access to such ecosystem services is of particular concern given that Madagascar is one of the world’s poorest countries, with almost 80% living below the poverty line $1.90 a day (Neugarten et al., 2016; Baumann, 2021). Most rural communities in Madagascar are directly dependent on ecosystem services, and multiple livelihood streams are essential in buffering environmental shocks (Huff, 2014). While carbon markets are frequently touted as a win-win solution for both climate and people, payments for ecosystem services may not reach the most vulnerable and can furthermore create a dependence on short-term external payments that are not sustainable and would erode social-ecological resilience and adaptive capacity in the longer-term (Bulte et al., 2008; Gross-Camp et al., 2012; Kronenberg and Hubacek, 2013). It is therefore vital that attempts to mitigate climate change consider local livelihoods and biodiversity and integrate more accurate carbon storage calculations alongside consideration of other ecosystem services.
Here, we advocate for the integration of palaeoecological approaches that can be used to explore former forest extent and history over timescales that are ecologically relevant, using case studies from Madagascar (Virah-Sawmy, 2009; Razafimanantsoa, 2022). Such palaeoecological methods include the analysis of fossil pollen from lake sediments, the analysis of charcoal to reconstruct fire history, stable isotopes to investigate vegetation and climate, dung fungal spores to reconstruct changes in the abundance of herbivores, diatoms to examine water quality, and geochemistry to investigate the interacting effects of climate and disturbance. We argue that determining appropriate reference conditions for restoration requires establishing the extent and composition of vegetation over time-periods that are relevant to landscape history and allow the impact of environmental change to be assessed over timescales of decades to centuries and even thousands of years. In Madagascar, human settlement (around 2000 years ago, Douglass et al., 2019), the Medieval Climate Anomaly (around 1000 years ago) and European colonization since the 19th Century, (Rid, 2010) provide critical reference conditions, that can help to distinguish ancient open and mosaic ecosystems from formerly forested areas. Research around such milestones in history, also contributes to disentangling anthropogenic and environmental drivers of land-cover change. This information is valuable in exploring the resilience of social-ecological systems, providing an underpinning for explorations of future sustainability under different scenarios of climate and land-use change.
2 The importance of landscape history– where and how should reforestation take place
There is no doubt that deforestation has taken place in some areas of Madagascar particularly in the last 50 years (Zaehringer et al., 2015; Vieilledent et al., 2018; Suzzi-Simmons, 2023). However, the idea that the whole island was once entirely covered by forest has been disproved by a growing number of palaeoecological studies, which suggest that open, mosaic and fire-adapted ecosystems are an integral part of many Malagasy landscapes outside of the mesic northeast (Figure 1).

Figure 1 Mosaic landscapes of Madagascar (A) next to Tsaramandroso village in the Northwest (Photo: A. Razafimanantsoa, 2011) (B) Landscape between Morondava and Malaimbandy, Southwest (Photo: Andriantsaralaza, S. 2021) (C) Tampoketsa grassland – forest mosaic of the Central Highlands of Madagascar (Photo: Kull, July 2019) (D) Littoral Forests, South East Efatsy Manombo, SE of Madagascar (Photo: Onjalalaina, 2020).
The “whole island forest” was an idea based on sparse data and anecdotal evidence from the 18th century (McConnell and Kull, 2014). Nevertheless, deforestation rates are still measured relative to a the presumed former 100% forest cover, leading to dramatic over-estimation of deforestation rates in areas that harbor ancient open and mosaic landscapes (Huff and Orengo, 2020). Such calculations assume unforested landscapes are degraded, without investigation the landscape history. This approach leads to neglect of the conservation of open and mosaic landscapes and the ecosystem services that they provide (Raveloaritiana et al., 2023) (see case study 1). Such assumptions can also have negative consequences for communities who are blamed for forest destruction or poor land stewardship (Seagle, 2012) (see case study 2). It is therefore essential to distinguish the extent of former forest cover prior to human arrival in Madagascar, the historical accounts of forest cover and its trajectory in recent centuries, in order that appropriate locations for reforestation can be identified.
To accurately calculate deforestation rates, we need to understand the former extent of forest cover in the early and mid-Holocene. Prior to human settlement and expansion c 2500–1350 years ago (Crowley, 2010; Douglass et al., 2019), closed canopy rainforest occurred only in the mesic northeast of the island, and eastern coastal strip (Straka, 1996; Moat and Smith, 2007). In the north-west, where rainfall is lower and more seasonal, tropical dry forests predominated, interspersed with succulents and heathlands (shrublands dominated by the family Ericaceae) (Figure 2). In the arid south-western part of the island, rainfall is too low to support true forests (ie. those that are fire intolerant and with a closed or near closed canopy) and open and arid-adapted landscapes including the spiny thickets existed here (Figure 2). Between these mesic and arid extremes, for example in the Highlands of Central Madagascar, there were mosaic ecosystems where true forest occurred only in patches (Figure 2) within matrices of ancient fire-prone grasslands, heathlands and woodland or savannas (e.g. (Burney et al., 2003; Virah-Sawmy et al., 2009b). These mosaic landscapes need particular consideration in reforestation planning, so that forests do not encroach on ancient open landscapes elements that harbor unique biodiversity (Bond et al., 2008; Willis et al., 2008). Specifically, fossil pollen data from lake sediments be used to identify whether forest distribution was once continuous, or was always patchy. This information can inform landscape management and restoration plans, a point illustrated by a case study from the Central Highlands of Madagascar.
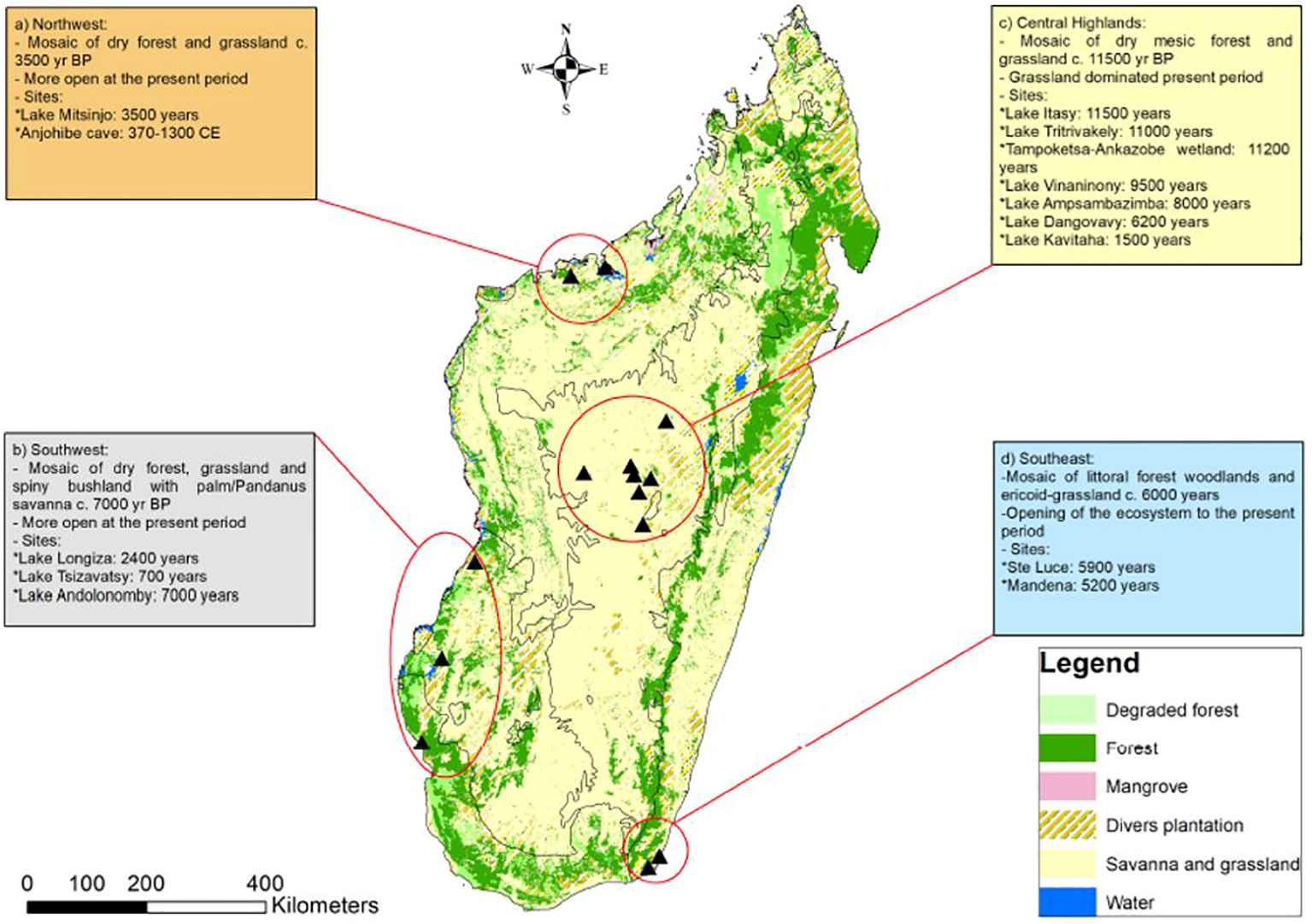
Figure 2 Summary of the Holocene vegetation reconstruction in Madagascar based on published vegetation records. The time span covered by each record is (A) Matsumoto and Burney, 1994; Burns et al., 2016; Voarintsoa et al., 2017; (B) Burney, 1993; Virah-Sawmy et al., 2016; Razanatsoa et al., 2021a; Razanatsoa et al., 2022; (C) Burney, 1987a; Burney, 1987b; Straka, 1996; Razafimanantsoa, 2022; Burney, 1987c; MacPhee et al., 1985; (D) Virah-Swamy et al., 2009a; 2009b; Virah-Sawmy, 2009. Map of main vegetation types based and simplified from Moat and Smith (2007).
2.1 Case study 1: Central Highlands
The Central Highlands are one of the main focuses for afforestation schemes in Madagascar. They are currently dominated by grasslands with patches of forest that are largely confined to ravines. This landscape is assumed to be highly degraded, however, palaeoecological investigations in the Central Highlands suggest the long-term presence of mosaic landscapes, where forest patches of variable extent co-existed with more open landscape elements including grasslands, heathlands and savannas (Burney, 1987b; Gasse and Van Campo, 2001; Razafimanantsoa, 2022). Though rainfall in the Central Highlands is high enough to support forests, open landscapes are likely maintained by fire and herbivory, as occurs in much of sub-Saharan Africa (Bond et al., 2005; Archibald et al., 2019). In such mosaic landscapes, co-existence of different vegetation types in the same climatic zone is mediated by topography, local hydrology and fire-vegetation-herbivory feedbacks, with forest elements generally confined to ravines which have greater water availability and are somewhat protected from fire (Bond and Keeley, 2005; Bond et al., 2005; Lehmann et al., 2011). Once established, fire sensitive forest elements can maintain themselves by shading out the herbaceous layer, which excludes fire except in extreme fire events or in the case of deliberate clearance by people (Staver et al., 2011; Archibald et al., 2019). Modeling experiments that predict forest cover based solely on rainfall are therefore likely to over-estimate past and present forest cover (Bond and Keeley, 2005; Bond et al., 2005) whereas those including fire more accurately represent the occurrence of savanna and grasslands throughout much of sub-Saharan Africa, including all but the eastern-most coastal strip in Madagascar (Bond and Keeley, 2005; Bond et al., 2005; Lehmann et al., 2011).
Fire- and grazing- adapted traits are common in many highlands species, strongly suggesting the presence of open vegetation elements, including grassland, heathland and savannas (e.g. Burney, 1987a; Burney, 1987b; Straka, 1996; Razafimanantsoa, 2022). Some of the grasslands, especially on the western flank, are highly biodiverse ancient grasslands, probably originating from the Neogene period (Bond et al., 2008; Vorontsova et al., 2016; Hackel et al., 2018; Vorontsova et al., 2020), while others are likely to be of anthropogenic origin. A recent pollen record from Tampoketsa-Ankazobe wetland, in the eastern slopes of the Highlands demonstrated the abundance of ericoid shrubland during the early Holocene, (Razafimanantsoa, 2022) (see Figure 3) confirming previous studies showing extensive presence of heathlands (Burney, 1987c; Straka, 1996) and supported by the high diversity of Ericaceae species in Madagascar (Grubb, 2003). While attention usually focuses on forest loss, the conversion of heathlands to grasslands warrants further attention and conservation/restoration effort (Silander et al., 2023).
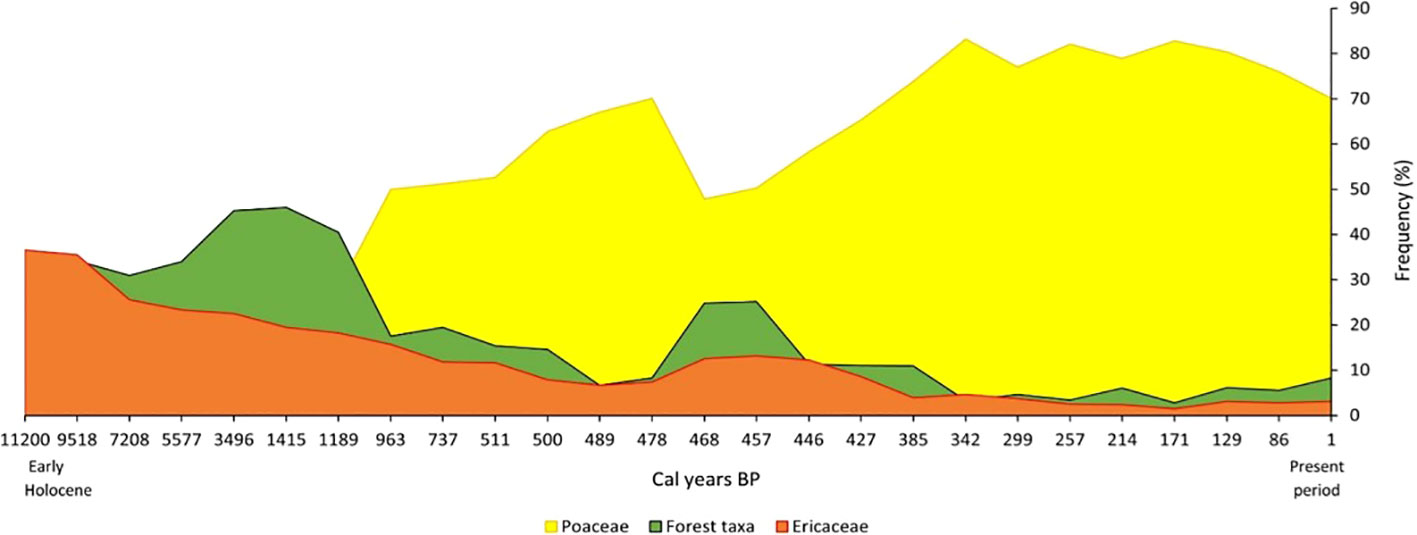
Figure 3 Summary pollen diagram of the dominant taxa in Tampoketsa-Ankazobe site, Central Highlands Madagascar during the Holocene (Adapted from Razafimanantsoa, 2022).
The palaeoecological and evolutionary data highlight the importance of considering landscape-specific histories in designing restoration in the Highlands. Though grasslands dominate the Highlands today, it is likely that heathland, woodland, savanna elements and forest patches were more extensive in the past and co-existed alongside grasslands in some areas (Burney, 1987a; Burney, 1987b; Straka, 1996; Razafimanantsoa, 2022) (see Figure 3). A further concern of tree planting initiatives is the selection of species, and how this selection process takes place. Many forest plantation projects in the Highlands use alien species that have negative effects on soil and water and creating novel landscapes with reduced ecosystem service provision. Eucalyptus has now infiltrated the remaining vestige of indigenous Tapia savanna woodland (Uapaca bojeri), leading to increased fire intensity and decreased regeneration of native species including the Tapia seedlings (Baohanta et al., 2012; Rakotondrasoa et al., 2012).
In assessing degradation and forest loss, the distinction is critical between true forests from wooded savannas. The latter maintain a near continuous herbaceous layer, with scattered tree cover, and which are fire and grazing adapted. In the Central Highlands, ancient Tapia (Uapaca) woodlands are now classified as savannas based on their functional traits (Solofondranohatra et al., 2018; Salmona et al., 2020). Native savanna trees such as the “olive tree” Noronhia lowryi originated in the Miocene period, c. 6.5 million years ago, concurrent with the global spread of savannas (Salmona et al., 2020). Classifying all savannas and heathlands as degraded forests (see for example Joseph and Seymour, 2020; Joseph and Seymour, 2021) is ecologically inaccurate and can lead to further errors in estimating forest loss and in encouraging tree planting schemes that replace unique open and mosaic ecosystems and associated ecosystem services (Raveloaritiana et al., 2023). Forest restoration (i.e. restoring previously forested areas with indigenous species) should be emphasized over afforestation of landscapes whose history is uncertain and which may contain ancient grasslands, heathlands and savannas. In south-east of Madagascar, for example, fast-growing native species such as Treblus mauritianus, Syzygium bernieri, Treculia madagascariensis and Uapaca thouarsii (Manjaribe et al., 2013) provide crucial ecosystem services as well as maintaining biodiversity.
The false dichotomy between forest-grassland (Joseph and Seymour, 2020; Joseph and Seymour, 2021) has diverted attention away from the loss and conversion of heathlands and savannas in the highlands, which are also ancient elements of the vegetation. Distinguishing ancient grasslands and mosaic ecosystems from derived anthropogenic ones is only one strand of a comprehensive conservation approach to the highlands that includes the conservation and restoration of ancient mosaic landscapes, reforestation in appropriate areas, as well as fire management that reflects the history and ecology of the region (Razafimanantsoa, 2022).
3 Balancing trade-offs between biodiversity, carbon storage a resource management
The primary motivation for most forest plantation projects is their perceived benefit to carbon storage to mitigate climate change. For example, the global carbon mitigation frameworks of the Clean Development Mechanism (CDM) and Reduced Emissions from Deforestation and Degradation (REDD and REDD+) have created new economic incentives for low income nations to engage in afforestation in Africa (Leach and Scoones, 2015; Bastin et al., 2019). However, in Madagascar, there is limited evidence for the success of these projects (Neudert et al., 2017), attributed to the choice of species, the way ecosystem service benefits are calculated, and to whom, and how, benefits are accrued.
Carbon calculations are typically incomplete or inaccurate and other ecosystem services, including cultural ones, are seldom considered. A particular problem is tendency to use above ground biomass as the main indicator of carbon stocks, which can lead to the underestimation of carbon stored in open and mosaic ecosystems. In Madagascar, as elsewhere, soil carbon pools hold three times higher than above-ground (Powers et al., 2011; Grinand et al., 2017; Ramifehiarivo et al., 2017; Dass et al., 2018; Razafindrakoto et al., 2018; Veldman et al., 2019; Yang et al., 2019).
Payments for avoided deforestation can in theory provide a more stable revenue than other forms of livelihood such as charcoal production or swidden agriculture (Neudert et al., 2018). However, communities are receiving fewer benefits than expected from REDD projects due to structural and institutional challenges locally, and also governance failure (Neudert et al., 2017). They therefore rely on multiple coping strategies to buffer environmental shocks.
At the same time, there are competing interests for example from agriculture or mining and these financial interests are backed by powerful organisations that may overpower local voices and concerns over biodiversity loss. While the benefits of carbon storage and export revenue are derived globally and nationally, the costs of biodiversity protection, including loss of agricultural areas, loss of access to non-timber forest products, and loss of income derived from charcoal production, are born locally (Scales, 2014; Neudert et al., 2017).
For long-term sustainability, tangible compensations to local communities based on principles of co-management and self-determinacy are vital (Neudert et al., 2017; Bond et al., 2019; Temperton et al., 2019; Razanatsoa et al., 2021b). Collaborations are needed that consider landscape history and social context in restoration projects that improve food security and sustainability of livelihoods, while also conserving the biodiversity on which ecosystem services depend. Participatory and ground-up approaches can help in selecting species that are locally appropriate and therefore more likely to survive as well as providing benefits beyond carbon storage, and material, cultural and educational benefits that contribute to the sustainability of conservation projects (Razanatsoa et al., 2021b), especially where competing interests are at play and valuable resources at stake (Virah-Swamy et al., 2009a; 2009b). Case study 2 from the Fort Dauphin area of southeast Madagascar illustrates these points.
3.1 Case study 2: littoral forests of southeast Madagascar
The littoral forests are Madagascar’s most endangered and range-restricted vegetation type, and are of high conservation value due to their scarcity and abundance of endemic species (Watson et al., 2004; Ingram and Dawson, 2005; Consiglio et al., 2006; Dawson and Ingram, 2008; Virah-Sawmy et al., 2009b). They are also used by communities for a range ecosystem services including oil, firewood, timber, medicine, animal food, human food, fibres and oil, as well as for spiritual purposes (Dawson and Ingram, 2008; Virah-Sawmy et al., 2014b). Their distribution closely coincides with the occurrence of ilmenite, and the forest patches have therefore been targeted for mining operations by QMM, a collaboration between Rio Tinto and the Malagasy Government. In assessing the environmental impact of mining operations, high rates of deforestation over recent decades were calculated relative to a presumed 100% former forest cover (Huff and Orengo, 2020). Based on this calculation, QMM argued that the rate of littoral forest destruction by local communities was unsustainable and would far exceed that of the planned mining operations (Bidaud et al., 2015). This argument was key in the progress and approval of the mining plans (Consiglio et al., 2006).
An alternative explanation for the patchy distribution of the littoral forest is that its extent is controlled mainly by local environmental factors such as soil and topography, and that it already existed in a patchy or mosaic form before human occupation. This hypothesis was tested by analyzing fossil pollen from within and outside forest patches. The results showed the presence of ancient fire adapted heathlands and open woodlands, pre-dating human arrival in Madagascar (Virah-Sawmy et al., 2009a; Virah-Sawmy et al., 2009b). Furthermore, the work demonstrated the ability of the forests to recover from natural disturbances including climatic perturbations and storm surges. The results suggest that littoral forest was always fragmented, its distribution confined to localized areas defined by local topographic, edaphic and hydrological factors (Virah-Sawmy, 2009; Virah-Sawmy et al., 2009a; Virah-Sawmy et al., 2009b). Forest loss has accelerated in recent decades, exacerbated by road construction during the exploratory phases of mining operations (Bidaud et al., 2015). However, the forest losses prior to mining operations, labeled as ‘catastrophic’, were inferred from current distribution and in incorrect assumption of 100% historical forest cover. The falsely high deforestation rates were used to obscure the negative impacts of mining, without conservation of the natural mosaic composition of the landscape.
The findings have implications for how littoral forests are managed and how the impacts of communities are perceived. Though QMM planned for mining operations to have Net Positive Impact (NPI), the success of this policy is disputed, and the ilmenite operations have been criticized for their impact on biodiversity and communities (Virah-Sawmy et al., 2014a; Huff and Orengo, 2020). First, NPI is calculated relative to the projected rate of forest destruction in the absence of mining, which as explained above is erroneously high due to an assumed former 100% forest cover and associated high rate of forest destruction. Any slowing of this perceived deforestation rate (“avoided deforestation”) is presented as a net positive impact i.e. forest destruction can still occur, but if the rate is calculated to be lower than the current rate calculated relative to past 100% forest cover, this is counted as a positive impact. Second, the effect of other mining impacts included construction of a dam, deforestation, afforestation with alien species, destruction of culturally important sites, loss of access to reeds and fisheries were not properly accounted for. Villagers displaced from their land were poorly compensated, if at all, and were moved to areas with soils unsuitable for farming and dislocated from cultural heritage (Huff and Orengo, 2020). Communities suffered loss of access to resources such reed beds and fisheries, either directly through prohibition of access, or indirectly through pollution of water and dam construction. Education programs instigated by QMM have also been criticized, doing little more than reinstate practices such as basket weaving that were only disrupted because of mining operations (Huff, 2014; Huff and Orengo, 2020). Rio Tinto retracted its NPI policy in 2016 on the grounds that it is not always achievable (de Silva et al., 2019).
Around Fort Dauphin, the meagre compensation, low land price and loss of access to resources were accepted by poor communities, who were struggling to survive. The high levels of poverty and urgent need for employment and economic interests and pressures by the national government and international companies mean implementing strict conservation of all forests is not an option (Virah-Swamy et al., 2009a; 2009b). Instead, Virah-Sawmy and Ebeling (2010) advocate for a more environmentally sensitive and inclusive approach to conservation and landscape management that considers biodiversity alongside local needs cultural context and landscape history Virah-Sawmy and Ebeling 2009a; 2009b. Palaecological information can help acknowledge or hopefully reduce biases in the way we manage and restore landscapes, especially when economic interests are at stake and local communities can easily be marginalized. Nevertheless, the spectre of “green-washing” remains ever-present in conservation mechanisms that are embedded in neo-liberalism (Adams, 2017).
4 Discussion – toward a transdisciplinary approach
Though deforestation and forest fragmentation in Madagascar are of great concern (Vieilledent et al., 2018), the misrepresentation of ancient open or mosaic ecosystems as ‘degraded’ forests or of anthropogenic origin has hampered conservation protection, alienated local communities and also enabled the destruction of wildlife habitats due to mining endeavors and afforestation schemes (Bond et al., 2008; Virah-Sawmy et al., 2014a; Bond et al., 2019; Huff and Orengo, 2020). Such schemes have also contributed to the disenfranchisement of communities and the loss of livelihoods (Huff and Orengo, 2020). A transdisciplinary approach is needed that considers landscape history, assesses ecosystem services holistically and accurately, and includes stakeholders and communities in decision-making and planning (Figure 4) (Fischer et al., 2020; Edrisi and Abhilash, 2021; Robinson et al., 2021).
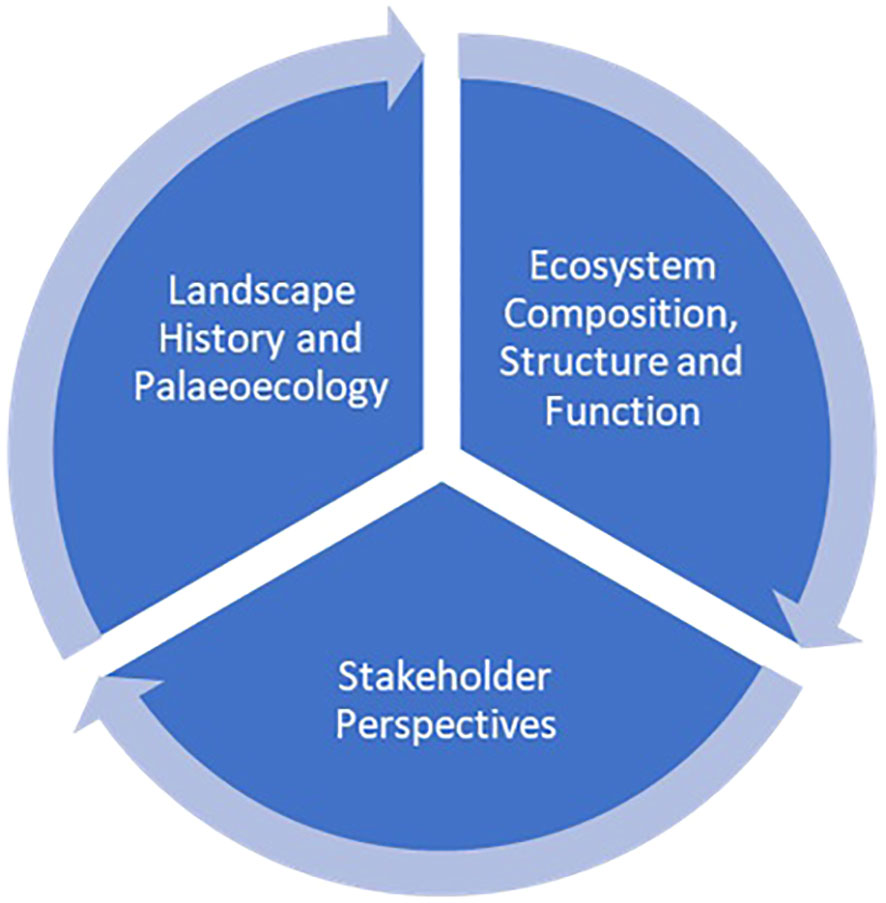
Figure 4 An integrated assessment of landscapes that considers landscape history, current ecological knowledge of pattern and process, and stakeholder perspectives on the history, culture and ecosystem services that landscapes provide.
The perceptions, management and afforestation initiatives of the Malagassy landscapes do not originate from local communities, but are based on anecdotal “baselines”, reinforced by narratives entangled in degradation and economic interests (Pollini, 2010; Hajdu and Fischer, 2017). For example, the assumption that all open ecosystems in Madagascar are degraded forests continues to underpin calculations of deforestation rates and debates in the literature (Huff and Orengo, 2020; Joseph and Seymour, 2020; Joseph and Seymour, 2021). However, the dichotomy between forest versus degraded forest is an artificial one that does not consider the spectrum of open and mosaic landscapes including savannas, heathlands and shrubland. Classifying such landscapes as degraded forest leads to over-estimation of deforestation rates and neglect of conservation of open and mosaic ecosystems including ancient grasslands, heathlands and savanna woodlands (Bond et al., 2019).
Current debates over whether open ecosystems are ancient or anthropogenic in Madagascar (Joseph and Seymour, 2020; Joseph and Seymour, 2021), are missing a key challenge for an informed and bottom up approach to conservation and restoration, which is to distinguish which open landscapes are ancient or anthropogenic, their historic nature and their current value to communities in ecosystem service provision (Vetter, 2020; Lehmann et al., 2021). The current polarization of forested versus degraded land obfuscates a sound ecological discussion of the ecological function of open and mosaic landscapes and their longevity and importance as biodiversity hotspots. Though principles for considering landscape history in restoration ecology and reforestation are emerging (Di Sacco et al., 2021), there have as yet been few attempts to integrate long-term data sets into restoration plans. In fact, a range of ecological and palaeoecological techniques are available to explore these questions, yet they remain underutilized (Virah-Sawmy et al., 2009b; Crowley et al., 2021; Gillson et al., 2021).
Palaeoecology can provide important information on the former extent of forest cover and the antiquity of mosaic and open ecosystems. Palaeoecological data can also guide the species used in reforestation, providing information on former forest composition that could be used to select indigenous instead of exotic species (Wingard et al., 2017). Though Pinus, Eucalyptus and Acacia, are favored because of their high survival rate and rapid growth compared with many native species (Carriere and Randriambanona, 2007; Verhaegen et al., 2011; Kull et al., 2019; Randriambanona et al., 2019), these plantations are low in biodiversity and negatively impact soil and water provision provided by open and mosaic ecosystems (Charles and Dukes, 2008; Stickler et al., 2009; Bond, 2019; Kull et al., 2019).
It is of course essential that the benefits of tree-planting on carbon storage are accurately calculated. The benefits of afforestation with non-native species are likely to be over-estimated, when only above-ground biomass, the most commonly used measurement in the REDD and REDD+ initiative, is used (Leach and Scoones, 2015; Panfil and Harvey, 2016; Bastin et al., 2019). Carbon calculations based on above-ground biomass have underestimated carbon storage in grasslands, savannas and heathlands, despite their potential as carbon sinks (Veldman et al., 2019), favoring the accelerated loss of high-conservation-value ecosystems with low above-ground biomass and exacerbating loss of biodiversity elsewhere through displaced agricultural expansion (Bond et al., 2019). In addition, it is critical that REDD+ and other carbon storage and afforestation schemes consider the wider range of ecosystem services provided by mosaic and open ecosystems, and their importance in local livelihoods (Lehmann et al., 2021). Some land-uses including agroforestry generally are able to maintain good soil carbon stocks (Rakotovao et al., 2020). Taviala (agroforestry) can help to maintain forest cover and also provides food and other resources as well as opportunities for producing high value cash crops such as vanilla and silk (Virah-Sawmy et al., 2015; Reuter et al., 2016; Rakotonarivo et al., 2017; Hending et al., 2018; Hewson et al., 2019; Hending et al., 2020; Schüßler et al., 2020; Wurz et al., 2022). The benefits of agroforestry to biodiversity are greater in forest-derived than fallow-derived land (Osen et al., 2021) Multiple livelihood streams from Taviala can buffer households against environmental shocks and improve the security of land-tenure, as well as providing habitat for biodiversity (Rakotonarivo et al., 2017; Schüßler et al., 2020).
Protected area expansion is another approach that can potentially mitigate carbon emissions while at the same time maintaining essential ecosystem services that bolster livelihoods, especially during times of environmental and economic stress (Waeber et al., 2016; Lehmann et al., 2021). However, it is important that local communities are consulted in the designation and management of such areas. In Madagascar, sacred groves protected by local taboos were incorporated into protected area planning (Virah-Sawmy et al., 2014b; Nopper et al., 2017; Schüßler et al., 2020), so that conservation planning reinforced and did not erode local culture and traditions.
Palaeoecology will be only one strand in developing a more nuanced, ecologically sound and locally appropriate approach to landscape restoration (Gillson et al., 2021). Local communities are often the ones who benefit the least from afforestation, resource extraction, and even some forms of conservation (Neudert et al., 2017).
Understanding the importance of cultural landscapes in maintaining diverse ecosystem services and buffering livelihoods against environmental shocks will require close engagement with communities. This will require a transdisciplinary approach that incorporates more accurate calculation of carbon stocks, a better understanding and reflexivity of landscape history, consideration of process (how decisions are made) and impacts of access to resources and livelihoods (Sayer et al., 2013; Fischer et al., 2020). A more inclusive approach to restoration is needed that considers desired futures and acceptable trade-offs between ecosystem services and resource provision (Fischer et al., 2020; Di Sacco et al., 2021; Gillson et al., 2021). Social processes are needed that aim to facilitate constructive debate among interest groups toward a common understanding and resolution of complex objectives in restoration, especially when there are competing powerful economic interests at stake (Sayer et al., 2013; Fischer et al., 2020). Palaeoecological information has the potential to be an important source of data that can help acknowledge or reduce biases in the way we perceive, manage and restore landscapes (Gillson, 2015).
5 Conclusions
Though motivated by legitimate concerns to halt deforestation and mitigate global emissions, poorly planned and implemented tree-planting projects risk - and have already resulted in - exacerbated degradation either directly through loss or replacement of indigenous vegetation or indirectly through social and economic displacement (Fairhead et al., 2012; Larson et al., 2013).
In Madagascar, accumulated evolutionary, comparative and palaeoecological knowledge suggest the antiquity of open and mosaic ecosystems, establishing them as valid conservation targets (Burney et al., 2003; Bond et al., 2008; Virah-Sawmy et al., 2009b; Vorontsova et al., 2016; Hackel et al., 2018; Vorontsova et al., 2020). Mosaic landscapes of forested and open ecosystems provide higher biodiversity and a wider range of ecosystem services than forested landscapes alone (Raveloaritiana et al., 2023) – and this is especially true where afforestation schemes use invasive alien species. The carbon storage capacity of open and mosaic landscapes has been underestimated and needs to be considered in restoration projects. Looking ahead, it is vital for both biodiversity and local communities that carbon storage schemes look beyond afforestation to consider more complex, nuanced and authentic landscape mosaics that reflect landscape history and better serve both biodiversity and people. Understanding the history and culture of landscapes requires a multi-disciplinary approach underpinned by biological science but that considers local context, landscape history and stakeholder needs to co-design restoration, conservation and carbon mitigation projects that contribute to social-ecological resilience.
Author contributions
LG: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing. ER: Conceptualization, Visualization, Writing – review & editing. AR: Conceptualization, Data curation, Formal Analysis, Visualization, Writing – review & editing. MV: Writing – review & editing. AE: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The authors gratefully acknowledge the following sources of funding which contributed to this work: National Research Foundation (NRF) Competitive Programme for Rated Researchers (Grant Number 118538), NRF/African Origins Platform (Grant Number 117666), NRF/Global Change Grand Challenge/SASSCAL (Grant number 118589), and the University of Cape Town Vice Chancellor’s Future Leaders Fund.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adams W. (2017). Sleeping with the enemy? Biodiversity conservation, corporations and the green economy. J. Pol. Ecol. 24 (1), 243–257. doi: 10.2458/v24i1.20804
Antonelli A., Smith R. J., Perrigo A. L., Crottini A., Hackel J., Testo W., et al. (2022). Madagascar’s extraordinary biodiversity: Evolution, distribution, and use. Science 378, eabf0869. doi: 10.1126/science.abf0869
Archibald S., Bond W. J., Hoffmann W., Lehmann C., Staver C., Stevens N. (2019). “Distribution and determinants of savannas,” in Savanna woody plants and large herbivores. Eds. Scogings P. F., Sankaran M. (Chichester: Wiley), 1–24.
Baohanta R., Thioulouse J., Ramanankierana H., Prin Y., Rasolomampianina R., Baudoin E., et al. (2012). Restoring native forest ecosystems after exotic tree plantation in Madagascar: combination of the local ectotrophic species Leptolena bojeriana and Uapaca bojeri mitigates the negative influence of the exotic species Eucalyptus camaldulensis and Pinus patula. Biol. Invasions 14, 2407–2421. doi: 10.1007/s10530-012-0238-5
Barak R. S., Hipp A. L., Cavender-Bares J., Pearse W. D., Hotchkiss S. C., Lynch E. A., et al. (2016). Taking the long view: integrating recorded, archeological, paleoecological, and evolutionary data into ecological restoration. Int. J. Plant Sci. 177, 90–102. doi: 10.1086/683394
Bastin J.-F., Finegold Y., Garcia C., Mollicone D., Rezende M., Routh D., et al. (2019). The global tree restoration potential. Science 365, 76–79. doi: 10.1126/science.aax0848
Baumann F. (2021). The next frontier—Human development and the anthropocene: UNDP human development report 2020. Environment: Sci. Policy Sustain. Dev. 63, 34–40. doi: 10.1080/00139157.2021.1898908
Bidaud C., Hrabanski M., Meral P. (2015). Voluntary biodiversity offset strategies in Madagascar. Ecosystem Serv. 15, 181–189. doi: 10.1016/j.ecoser.2015.02.011
Boissière M., Atmadja S., Guariguata M. R., Kassa H., Sist P. (2021). Perspectives on the socio-economic challenges and opportunities for tree planting: A case study of Ethiopia. For. Ecol. Manage. 497, 119488. doi: 10.1016/j.foreco.2021.119488
Bond W. J. (2019). Open ecosystems: ecology and evolution beyond the forest edge (Oxford: Oxford University Press).
Bond W. J., Keeley J. E. (2005). Fire as a global 'herbivore': the ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 20, 387–394. doi: 10.1016/j.tree.2005.04.025
Bond W. J., Silander J. A. Jr., Ranaivonasy J., Ratsirarson J. (2008). The Antiquity of Madagascar's grasslands and the rise of C4 biomes. J. Biogeography 35, 1743–1758. doi: 10.1111/j.1365-2699.2008.01923.x
Bond W. J., Stevens N., Midgley G. F., Lehmann C. E. (2019). The trouble with trees: afforestation plans for Africa. Trends Ecol. Evol. 34, 963–965. doi: 10.1016/j.tree.2019.08.003
Bond W. J., Woodward F. I., Midgley G. F. (2005). The global distribution of ecosystems in a world without fire. New Phytol. 165, 525–538. doi: 10.1111/j.1469-8137.2004.01252.x
Brancalion P. H., Holl K. D. (2020). Guidance for successful tree planting initiatives. J. Appl. Ecol. 57, 2349–2361. doi: 10.1111/1365-2664.13725
Bredemeier M., Dohrenbusch A. (2009). Afforestation and reforestation. Biodiversity: Structure Function-Volume II 2, 219.
Bulte E. H., Lipper L., Stringer R., Zilberman D. (2008). Payments for ecosystem services and poverty reduction: concepts, issues, and empirical perspectives. Environ. Dev. Economics 13, 245–254. doi: 10.1017/S1355770X08004348
Burney D. A. (1987a). “Pre-settlement vegetation changes at Lake Tritrivakely, Madagascar,” Palaeoecology of Africa and the surrounding islands 18, 357–381.
Burney D. A. (1987b). Late Holocene vegetational change in central Madagascar. Quaternary Res. 28, 130–143. doi: 10.1016/0033-5894(87)90038-X
Burney D. (1987c). Late Quaternary Stratigraphic Charcoals Records from Madagascar. Quaternary Res. 28:274–280.
Burney D. A., Robinson G. S., Burney L. P. (2003). Sporormiella and the late Holocene extinctions in Madagascar. Proc. Natl. Acad. Sci. 100, 10800–10805. doi: 10.1073/pnas.1534700100
Burns S. J., Godfrey L. R., Faina P., McGee D., Hardt B., Ranivoharimanana L., et al. (2016). Rapid human-induced landscape transformation in madagascar at the end of the first millennium of the common era. Quaternary Sci. Rev. 134, 92–99.
Carriere S. M., Randriambanona H. (2007). Biodiversité introduite et autochtone: antagonisme ou complémentarité? Le cas de l'eucalyptus à Madagascar. BOIS FORETS Des. TROPIQUES 292, 5–21. doi: 10.19182/bft2007.292.a20349
Charles H., Dukes J. S. (2008). “Impacts of invasive species on ecosystem services,” in Biological invasions (Berlin, Heidelberg: Springer), 217–237.
Consiglio T., Schatz G. E., McPherson G., Lowry P. P., Rabenantoandro J., Rogers Z. S., et al. (2006). Deforestation and plant diversity of Madagascar's littoral forests. Conserv. Biol. 20, 1799–1803. doi: 10.1111/j.1523-1739.2006.00562.x
Crowley B. E. (2010). A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quaternary Sci. Rev. 29, 2591–2603. doi: 10.1016/j.quascirev.2010.06.030
Crowley B. E., Godfrey L. R., Hansford J. P., Samonds K. E. (2021). Seeing the forest for the trees—and the grasses: revisiting the evidence for grazer-maintained grasslands in Madagascar's Central Highlands. Proc. R. Soc. B 288, 20201785. doi: 10.1098/rspb.2020.1785
Dass P., Houlton B. Z., Wang Y., Warlind D. (2018). Grasslands may be more reliable carbon sinks than forests in California. Environ. Res. Lett. 13, 074027. doi: 10.1088/1748-9326/aacb39
Dawson T. P., Ingram J. C. (2008). Sustainable livelihoods and forest resources in Madagascar: A multi-scale analysis using remote sensing. Environ. Sci. 5, 129–143. doi: 10.1080/15693430801912188
de Silva G. C., Regan E. C., Pollard E. H. B., Addison P. F. E. (2019). The evolution of corporate no net loss and net positive impact biodiversity commitments: Understanding appetite and addressing challenges. Business Strategy Environ. 28, 1481–1495. doi: 10.1002/bse.2379
Dietl G. P., Kidwell S. M., Brenner M., Burney D. A., Flessa K. W., Jackson S. T., et al. (2015). Conservation paleobiology: leveraging knowledge of the past to inform conservation and restoration. Annu. Rev. Earth Planetary Sci. 43, 79–103. doi: 10.1146/annurev-earth-040610-133349
Di Sacco A., Hardwick K. A., Blakesley D., Brancalion P. H., Breman E., Cecilio Rebola L., et al. (2021). Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Global Change Biol. 27, 1328–1348. doi: 10.1111/gcb.15498
Douglass K., Hixon S., Wright H. T., Godfrey L. R., Crowley B. E., Manjakahery B., et al. (2019). A critical review of radiocarbon dates clarifies the human settlement of Madagascar. Quaternary Sci. Rev. 221, 105878. doi: 10.1016/j.quascirev.2019.105878
Edrisi S. A., Abhilash P. (2021). Need of transdisciplinary research for accelerating land restoration during UN decade on ecosystem restoration. Restor. Ecol. 29 (8), e13531. doi: 10.1111/rec.13531
Fairhead J., Leach M., Scoones I. (2012). Green Grabbing: a new appropriation of nature? J. Peasant Stud. 39, 237–261. doi: 10.1080/03066150.2012.671770
Falk D. A., Watts A. C., Thode A. E. (2019). Scaling ecological resilience. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00275
Fischer J., Riechers M., Loos J., Martin-Lopez B., Temperton V. M. (2020). Making the UN Decade on Ecosystem Restoration a social-ecological endeavour. Trends Ecol. Evol. 36 (1), 20–28. doi: 10.1016/j.tree.2020.08.018
García-Dory F., Houzer E., Scoones I. (2021). Livestock and climate justice: Challenging mainstream policy narratives. IDS Bulletin 53 (4), 47–64. doi: 10.19088/1968-2021.128
Gasse F., Van Campo E. (2001). Late Quaternary environmental changes from a pollen and diatom record in the soutehrn tropics (Lake Tritivakely, Madagascar). Palaeogeography Palaeoclimatology Palaeoecol. 167, 287–308. doi: 10.1016/S0031-0182(00)00242-X
Gillson L. (2015). Biodiversity Conservation and Environmental Change: Using palaeoecology to manage dynamic landscapes in the Anthropocene (Oxford, U.K: Oxford University Press).
Gillson L., Dirk C., Gell P. (2021). Using long-term data to inform a decision pathway for restoration of ecosystem resilience. Anthropocene 36, 100315. doi: 10.1016/j.ancene.2021.100315
Grinand C., Le Maire G., Vieilledent G., Razakamanarivo H., Razafimbelo T., Bernoux M. (2017). Estimating temporal changes in soil carbon stocks at ecoregional scale in Madagascar using remote-sensing. Int. J. Appl. Earth observation geoinformation 54, 1–14. doi: 10.1016/j.jag.2016.09.002
Gross-Camp N. D., Martin A., McGuire S., Kebede B., Munyarukaza J. (2012). Payments for ecosystem services in an African protected area: exploring issues of legitimacy, fairness, equity and effectiveness. Oryx 46, 24–33. doi: 10.1017/S0030605311001372
Hackel J., Vorontsova M. S., Nanjarisoa O. P., Hall R. C., Razanatsoa J., Malakasi P., et al. (2018). Grass diversification in Madagascar: in situ radiation of two large C3 shade clades and support for a Miocene to Pliocene origin of C4 grassy biomes. J. Biogeography 45, 750–761. doi: 10.1111/jbi.13147
Hajdu F., Fischer K. (2017). Problems, causes and solutions in the forest carbon discourse: a framework for analysing degradation narratives. Climate Dev. 9, 537–547. doi: 10.1080/17565529.2016.1174663
Hending D., Andrianiaina A., Maxfield P., Rakotomalala Z., Cotton S. (2020). Floral species richness, structural diversity and conservation value of vanilla agroecosystems in Madagascar. Afr. J. Ecol. 58, 100–111. doi: 10.1111/aje.12682
Hending D., Andrianiaina A., Rakotomalala Z., Cotton S. (2018). The use of vanilla plantations by lemurs: encouraging findings for both lemur conservation and sustainable agroforestry in the Sava region, northeast Madagascar. Int. J. Primatology 39, 141–153. doi: 10.1007/s10764-018-0022-1
Hewson J., Razafimanahaka J. H., Wright T. M., Mandimbiniaina R., Mulligan M., Jones J. P., et al. (2019). Land change modelling to inform strategic decisions on Forest cover and CO 2 emissions in eastern Madagascar. Environ. Conserv. 46, 25–33. doi: 10.1017/S0376892918000358
Huff A. (2014). Weathering the'Long Wounded Year': livelihoods, nutrition and changing political ecologies in the Mikea Forest Region, Madagascar. J. Political Ecol. 21, 83–107.
Huff A., Orengo Y. (2020). Resource warfare, pacification and the spectacle of ‘green’development: Logics of violence in engineering extraction in southern Madagascar. Political Geogr. 81, 102195. doi: 10.1016/j.polgeo.2020.102195
Ingram J. C., Dawson T. P. (2005). Climate change impacts and vegetation response on the island of Madagascar. Philos. Trans. R. Soc. A: Mathematical Phys. Eng. Sci. 363, 55–59. doi: 10.1098/rsta.2004.1476
Joseph G. S., Seymour C. L. (2020). Madagascan highlands: originally woodland and forest containing endemic grasses, not grazing-adapted grassland. Proc. R. Soc. B 287, 20201956. doi: 10.1098/rspb.2020.1956
Joseph G. S., Seymour C. L. (2021). The unlikely’ Antiquity of Madagascar's grasslands’: Disproportionately forest-limited endemic fauna support anthropogenic transformation from woodland. J. Biogeogr. 48 (8), 2111–2115. doi: 10.1111/jbi.14132
Kronenberg J., Hubacek K. (2013). Could payments for ecosystem services create an “ecosystem service curse”? Ecol. Soc. 18 (1), 10. doi: 10.5751/ES-05240-180110
Kull C. A., Harimanana S. L., Andrianoro A. R., Rajoelison L. G. (2019). Divergent perceptions of the ‘neo-Australian’forests of lowland eastern Madagascar: Invasions, transitions, and livelihoods. J. Environ. Manage. 229, 48–56. doi: 10.1016/j.jenvman.2018.06.004
Lacroix E., Carodenuto S., Richter F., Pistorius T., Tennigkeit T. (2016). Restauration des paysages forestiers Evaluation des potentialités dans le contexte des engagements de Bonn 2.0 et de la Déclaration de New York sur les forêts. Méthodologie résultats pour Madagascar. Mars. Available at: https://afr100.org/sites/default/files/2022-11/06-MDG_RPF_MEOR_Madagascar_Final.pdf.
Larson A. M., Brockhaus M., Sunderlin W. D., Duchelle A., Babon A., Dokken T., et al. (2013). Land tenure and REDD+: The good, the bad and the ugly. Global Environ. Change 23, 678–689. doi: 10.1016/j.gloenvcha.2013.02.014
Leach M., Scoones I. (2015). “Political ecologies of carbon in Africa,” in Carbon conflicts and forest landscapes in Africa, Leach M., Scoones I. (eds) (London: Routledge), 21–62.
Lehmann C. E. R., Archibald S. A., Hoffmann W. A., Bond W. J. (2011). Deciphering the distribution of the savanna biome. New Phytol. 191, 197–209. doi: 10.1111/j.1469-8137.2011.03689.x
Lehmann C. E., Solofondranohatra C. L., Vorontsova M. S. (2021). Beyond ancient versus anthropogenic for Madagascar's grassy ecosystems. A Reply to: Crowley et al.(2021). Proc. R. Soc. B 288, 20210388. doi: 10.1098/rspb.2021.0388
MacPhee R., Burney D., Wells N. (1985). Early holocene chronology and environment of ampasambazimba, a malagasy subfossil lemur site. Int. J. Primatology 6, 463–489.
Mandimbisoa R. (2021). Une vitrine présidentielle de reboisement à Analamanga. (Antananarivo: https://Madagascar-Tribune.com)
Manjaribe C., Frasier C. L., Rakouth B., Louis E. E. (2013). Ecological restoration and reforestation of fragmented forests in Kianjavato, Madagascar. Int. J. Ecol, 726275. doi: 10.1155/2013/726275
Matsumoto K., Burney D. A. (1994). Late holocene environments at lake mitsinjo, northwestern madagascar. Holocene 4, 16–24.
McConnell W. J., Kull C. A. (2014). “Deforestation in Madagascar: Debates over the island's forest cover and challenges of measuring forest change,” in Conservation and environmental management in Madagascar, Ed, Scales I. R. (New York and London: Routledge), 67–104.
Moat J., Smith P. (2007). Atlas of the Vegetation of Madagascar (London, UK: The Royal Botanic Gardens, Kew. Kew Publishing).
Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kents J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Neudert R., Ganzhorn J. U., WÄTzold F. (2017). Global benefits and local costs – The dilemma of tropical forest conservation: A review of the situation in Madagascar. Environ. Conserv. 44, 82–96. doi: 10.1017/S0376892916000552
Neudert R., Olschofsky K., Kübler D., Prill L., Köhl M., Wätzold F. (2018). Opportunity costs of conserving a dry tropical forest under REDD+: The case of the spiny dry forest in southwestern Madagascar. For. Policy Economics 95, 102–114. doi: 10.1016/j.forpol.2018.07.013
Neugarten R. A., Honzák M., Carret P., Koenig K., Andriamaro L., Cano C. A., et al. (2016). Rapid assessment of ecosystem service co-benefits of biodiversity priority areas in Madagascar. PLoS One 11, e0168575. doi: 10.1371/journal.pone.0168575
Nopper J., Ranaivojaona A., Riemann J. C., Rödel M.-O., Ganzhorn J. U. (2017). One forest is not like another: The contribution of community-based natural resource management to reptile conservation in Madagascar. Trop. Conserv. Sci. 10, 1940082917693234. doi: 10.1177/1940082917693234
Osen K., Soazafy M. R., Martin D. A., Wurz A., März A., Ranarijaona H. L. T., et al. (2021). Land-use history determines stand structure and tree diversity in vanilla agroforests of northeastern Madagascar. Appl. Vegetation Sci. 24, e12563. doi: 10.1111/avsc.12563
Panfil S. N., Harvey C. A. (2016). REDD+ and biodiversity conservation: A review of the biodiversity goals, monitoring methods, and impacts of 80 REDD+ projects. Conserv. Lett. 9, 143–150. doi: 10.1111/conl.12188
Pollini J. (2010). Environmental degradation narratives in Madagascar: From colonial hegemonies to humanist revisionism. Geoforum 41, 711–722. doi: 10.1016/j.geoforum.2010.04.001
Powers J. S., Corre M. D., Twine T. E., Veldkamp E. (2011). Geographic bias of field observations of soil carbon stocks with tropical land-use changes precludes spatial extrapolation. Proc. Natl. Acad. Sci. 108, 6318–6322. doi: 10.1073/pnas.1016774108
Rakotonarivo O. S., Jacobsen J. B., Larsen H. O., Jones J. P., Nielsen M. R., Ramamonjisoa B. S., et al. (2017). Qualitative and quantitative evidence on the true local welfare costs of forest conservation in Madagascar: are discrete choice experiments a valid ex ante tool? World Dev. 94, 478–491. doi: 10.1016/j.worlddev.2017.02.009
Rakotondrasoa O. L., Malaisse F., Rajoelison G. L., Razafimanantsoa T. M., Rabearisoa M. R., Ramamonjisoa B. S., et al. (2012). Tapia forest, endemic ecosystem to Madagascar: ecology, functions, causes of degradation and transformation: a review. Biotechnologie Agronomie Société Environnement 16, 541–552.
Rakotovao N. H., Ramifehiarivo N., Grinand C., Razakamanarivo H. R., Albrecht A., Razafimbelo T. M. (2020). Impact of agroecological practices on soil organic carbon sequestration using synchronic and diachronic approaches in Madagascar. J. Agric. Rural Dev. Tropics Subtropics (JARTS) 121, 113–125.
Ramifehiarivo N., Brossard M., Grinand C., Andriamananjara A., Razafimbelo T., Rasolohery A., et al. (2017). Mapping soil organic carbon on a national scale: Towards an improved and updated map of Madagascar. Geoderma Regional 9, 29–38. doi: 10.1016/j.geodrs.2016.12.002
Randriambanona H., Randriamalala J. R., Carrière S. M. (2019). Native forest regeneration and vegetation dynamics in non-native Pinus patula tree plantations in Madagascar. For. Ecol. Manage. 446, 20–28. doi: 10.1016/j.foreco.2019.05.019
Ranjatson P., Rebecca M., Randrianasolo R., Mananga J., Razafimbelo N., Steven L. (2019). “Tenure security and forest landscape restoration: Results from exploratory research in Boeny, Madagascar,” in World bank conference on land and poverty (Washington DC: The World Bank), 1–28. Available at: https://events.globallandscapesforum.org/wp-content/uploads/sites/2/2019/10/PMcLain1901.pdf
Raveloaritiana E., Wurz A., Osen K., Soazafy M. R., Grass I., Martin D. A., et al. (2023). Complementary ecosystem services from multiple land uses highlight the importance of tropical mosaic landscapes. Ambio 52, 1558–1574. doi: 10.1007/s13280-023-01888-3
Razafimanantsoa A. H. I. (2022). Holocene vegetation history and environmental change in the forest-grassland mosaic of the Central Highlands of Madagascar (Cape Town: Doctoral. University of Cape Town).
Razafindrakoto M., Andriamananjara A., Razafimbelo T., Hewson J., Andrisoa R. H., Jones J. P. G., et al. (2018). “Chapter 3 - organic carbon stocks in all pools following land cover change in the rainforest of Madagascar,” in Soil Management and Climate Change. Eds. Muñoz MÁ., Zornoza R. (Cambridge, Massachusetts, United States: Academic Press), 25–37.
Razanatsoa E., Andriantsaralaza S., Holmes S. M., Rakotonarivo O. S., Ratsifandrihamanana A. N., Randriamiharisoa L., et al. (2021b). Fostering local involvement for biodiversity conservation in tropical regions: Lessons from Madagascar during the COVID-19 pandemic. Biotropica 53 (4), 994–1003. doi: 10.1111/btp.12967
Razanatsoa E., Gillson L., Virah-Sawmy M., Woodborne S. (2021a). Pollen records of the 14th and 20th centuries AD from Lake Tsizavatsy in southwest Madagascar. Palaeoecology of Africa 35, 309–315. doi: 10.1201/9781003162766-20
Razanatsoa E., Gillson L., Virah-Sawmy M., Woodborne S. (2022). Synergy between climate and human land-use maintained open vegetation in southwest Madagascar over the last millennium. The Holocene 32 (1–2), 57–69. doi: 10.1177/09596836211041731
Reuter K. E., Randell H., Wills A. R., Sewall B. J. (2016). The consumption of wild meat in Madagascar: drivers, popularity and food security. Environ. Conserv. 43, 273. doi: 10.1017/S0376892916000059
Rid T. (2010). The nineteenth century origins of counterinsurgency doctrine. J. Strategic Stud. 33, 727–758. doi: 10.1080/01402390.2010.498259
Robinson J. M., Gellie N., MacCarthy D., Mills J. G., O'Donnell K., Redvers N. (2021). Traditional ecological knowledge in restoration ecology: a Call to listen deeply, to engage with, and respect indigenous voices. Restor. Ecol. 29, e13381. doi: 10.1111/rec.13381
Salmona J., Olofsson J. K., Hong-Wa C., Razanatsoa J., Rakotonasolo F., Ralimanana H., et al. (2020). Late Miocene origin and recent population collapse of the Malagasy savanna olive tree (Noronhia lowryi). Biol. J. Linn. Soc. 129, 227–243. doi: 10.1093/biolinnean/blz164
Sayer J., Sunderland T., Ghazoul J., Pfund J.-L., Sheil D., Meijaard E., et al. (2013). Ten principles for a landscape approach to reconciling agriculture, conservation, and other competing land uses. Proc. Natl. Acad. Sci. 110, 8349–8356. doi: 10.1073/pnas.1210595110
Scales I. R. (2014). The future of conservation and development in Madagascar: time for a new paradigm? Madagascar Conserv. Dev. 9, 5–12. doi: 10.4314/mcd.v9i1.2
Schüßler D., Mantilla-Contreras J., Stadtmann R., Ratsimbazafy J. H., Radespiel U. (2020). Identification of crucial stepping stone habitats for biodiversity conservation in northeastern Madagascar using remote sensing and comparative predictive modeling. Biodiversity Conserv. 29, 2161–2184. doi: 10.1007/s10531-020-01965-z
Seagle C. (2012). Inverting the impacts: Mining, conservation and sustainability claims near the Rio Tinto/QMM ilmenite mine in Southeast Madagascar. J. Peasant Stud. 39, 447–477. doi: 10.1080/03066150.2012.671769
Silander J. A. Jr., Bond W. J., Ratsirarson J. (2023). The grassy ecosystems of Madagascar in context: Ecology, evolution, and conservation. Plants People Planet. doi: 10.1002/ppp3.10417
Solofondranohatra C. L., Vorontsova M. S., Hackel J., Besnard G., Cable S., Williams J., et al. (2018). Grass functional traits differentiate forest and savanna in the Madagascar central highlands. Front. Ecol. Evol. 6, 184. doi: 10.3389/fevo.2018.00184
Staver A. C., Archibald S., Levin S. A. (2011). The global extent and determinants of Savanna and forest as alternative biome states. Science 334, 230–232. doi: 10.1126/science.1210465
Stickler C. M., Nepstad D. C., Coe M. T., McGrath D. G., Rodrigues H. O., Walker W. S., et al. (2009). The potential ecological costs and cobenefits of REDD: a critical review and case study from the Amazon region. Global Change Biol. 15, 2803–2824. doi: 10.1111/j.1365-2486.2009.02109.x
Straka H. (1996). “Histoire de la végétation de Madagascar oriental dans les derniers 100 millénaires,” in BiogeHographie de Madagascar (Paris: ORSTOM), 37.
Suzzi-Simmons A. (2023). Status of deforestation of Madagascar. Global Ecol. Conserv. 42, e02389. doi: 10.1016/j.gecco.2023.e02389
Temperton V. M., Buchmann N., Buisson E., Durigan G., Kazmierczak Ł., Perring M. P., et al. (2019). Step back from the forest and step up to the Bonn Challenge: how a broad ecological perspective can promote successful landscape restoration. Restor. Ecol. 27, 705–719. doi: 10.1111/rec.12989
Tsayem Demaze M. (2014). L’enrôlement de Madagascar dans la REDD+: domestiquer une opportunité internationale. VertigO: la revue électronique en sciences de l’environnement. (Open Edition Journals) 14 (1).
Veldman J. W., Aleman J. C., Alvarado S. T., Anderson T. M., Archibald S., Bond W. J., et al. (2019). Comment on “The global tree restoration potential”. Science 366, eaay7976. doi: 10.1126/science.aay7976
Veldman J. W., Overbeck G., Negreiros D., Mahy G., Le Stradic S., Fernandes G. W., et al. (2015). Tyranny of trees in grassy biomes. Science 347, 484–485. doi: 10.1126/science.347.6221.484-c
Verhaegen D., Randrianjafy H., Montagne P., Danthu P., Rabevohitra R., Tassin J., et al. (2011). Reconstructing the history of Eucalyptus genus introduction in Madagascar. Bois Forêts Des. Tropiques 309 (3), 18–25. doi: 10.19182/bft2011.309.a20462
Vetter S. (2020). With power comes responsibility–A rangelands perspective on forest landscape restoration. Front. Sustain. Food Syst. 4, 225. doi: 10.3389/fsufs.2020.549483
Vieilledent G., Grinand C., Rakotomalala F. A., Ranaivosoa R., Rakotoarijaona J.-R., Allnutt T. F., et al. (2018). Combining global tree cover loss data with historical national forest cover maps to look at six decades of deforestation and forest fragmentation in Madagascar. Biol. Conserv. 222, 189–197. doi: 10.1016/j.biocon.2018.04.008
Virah-Sawmy M. (2009). Ecosystem management in Madagascar during global change. Conserv. Lett. 2, 163–170. doi: 10.1111/j.1755-263X.2009.00066.x
Virah-Sawmy M., Ebeling J. (2010). The difficult road toward real-world engagement: conservation science and mining in southern Madagascar. Conserv. Lett. 3, 288–289. doi: 10.1111/j.1755-263X.2010.00126.x
Virah-Sawmy M., Ebeling J., Taplin R. (2014a). Mining and biodiversity offsets: A transparent and science-based approach to measure “no-net-loss”. J. Environ. Manage. 143, 61–70. doi: 10.1016/j.jenvman.2014.03.027
Virah-Sawmy M., Gardner C., Ratsifandrihamanana A. (2014b). “The Durban vision in practice. Experiences in the participatory governance of Madagascar's new protected areas,” in Conservation and Environmental Management in Madagascar. Ed. Scales I. (New York and London: Routledge), 216–251.
Virah-Sawmy M., Gillson L., Gardner C., Anderson A., Clark G., Haberle S. (2015). A landscape vulnerability framework for identifying integrated conservation and adaptation pathways to climate change: the case of Madagascar’s spiny forest. Landscape Ecol. 31, 637–654. doi: 10.1007/s10980-015-0269-2
Virah-Sawmy M., Gillson L., Willis K. J. (2009a). How does spatial heterogeneity influence resilience to climatic changes? Ecol. dynamics southeast Madagascar. Ecol. Monogr. 79, 557–574. doi: 10.1890/08-1210.1
Virah-Sawmy M., Willis K. J., Gillson L. (2009b). Threshold response of Madagascar's littoral forest to sea-level rise. Global Ecol. Biogeography 18, 98–110. doi: 10.1111/j.1466-8238.2008.00429.x
Virah-Sawmy M., Gillson L., Gardner C. J., Anderson A., Clark G., Haberle S. (2016). A landscape vulnerability framework for identifying integrated conservation and adaptation pathways to climate change: the case of madagascar’s spiny forest. Landscape Ecol. 31, 637–654.
Voarintsoa N. R. G., Wang L., Railsback L. B., Brook G. A., Liang F., Cheng H., et al. (2017). Multiple proxy analyses of a U/Th-dated stalagmite to reconstruct paleoenvironmental changes in northwestern madagascar between 370 CE and 1300 CE. Palaeogeogr. Palaeoclimatology Palaeoecol. 469, 138–155.
Vorontsova M. S., Besnard G., Forest F., Malakasi P., Moat J., Clayton W. D., et al. (2016). Madagascar's grasses and grasslands: anthropogenic or natural? Proc. R. Soc. B: Biol. Sci. 283, 20152262. doi: 10.1098/rspb.2015.2262
Vorontsova M. S., Lowry P. P., Andriambololonera S. R., Wilmé L., Rasolohery A., Govaerts R., et al. (2020). Inequality in plant diversity knowledge and unrecorded plant extinctions: An example from the grasses of Madagascar. Plants People Planet. 3 (1), 45–60.
Waeber P. O., Wilmé L., Mercier J.-R., Camara C., Lowry P. P. (2016). How effective have thirty years of internationally driven conservation and development efforts been in Madagascar? PLoS One 11, e0161115. doi: 10.1371/journal.pone.0161115
Watson J. E. M., Whittaker R. J., Dawson T. P. (2004). Avifaunal responses to habitat fragmentation in the threatened littoral forests of south-eastern Madagascar. J. Biogeography 31, 1791–1807. doi: 10.1111/j.1365-2699.2004.01142.x
Willis K. J., Gillson L., Virah-Sawmy M. (2008). Nature or nurture: the ambiguity of C4 grasslands in Madagascar. J. Biogeography 35, 1741–1742. doi: 10.1111/j.1365-2699.2008.01985.x
Wingard G. L., Bernhardt C. E., Wachnicka A. H. (2017). The role of paleoecology in restoration and resource management—the past as a guide to future decision-making: review and example from the Greater Everglades Ecosystem, USA. Front. Ecol. Evol. 5, 11. doi: 10.3389/fevo.2017.00011
Wurz A., Tscharntke T., Martin D. A., Osen K., Rakotomalala A. A., Raveloaritiana E., et al. (2022). Win-win opportunities combining high yields with high multi-taxa biodiversity in tropical agroforestry. Nat. Commun. 13, 4127. doi: 10.1038/s41467-022-30866-8
Yang Y., Tilman D., Furey G., Lehman C. (2019). Soil carbon sequestration accelerated by restoration of grassland biodiversity. Nat. Commun. 10, 1–7. doi: 10.1038/s41467-019-08636-w
Keywords: ecosystem services, livelihoods, palaeoecology, reforestation, restoration
Citation: Gillson L, Razanatsoa E, Razafimanantsoa AHI, Virah-Sawmy M and Ekblom A (2023) The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar. Front. Conserv. Sci. 4:1286459. doi: 10.3389/fcosc.2023.1286459
Received: 31 August 2023; Accepted: 24 November 2023;
Published: 20 December 2023.
Edited by:
Maria M. Romeiras, University of Lisbon, PortugalReviewed by:
Matthew E. Aiello-Lammens, Pace University, United StatesDirk Hölscher, University of Göttingen, Germany
Bruno Salomon Ramamonjisoa, University of Antananarivo, Madagascar
Copyright © 2023 Gillson, Razanatsoa, Razafimanantsoa, Virah-Sawmy and Ekblom. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lindsey Gillson, bGluZHNleS5naWxsc29uQHVjdC5hYy56YQ==
†ORCID: Lindsey Gillson, orcid.org/0000-0001-9607-6760
Estelle Razanatsoa, orcid.org/0000-0002-7219-1411
Andriantsilavo Hery Isandratana Razafimanantsoa, orcid.org/0000-0002-9933-6991
Malika Virah-Sawmy, orcid.org/0000-0003-3646-5646
Anneli Ekblom, orcid.org/0000-0001-9248-5516