 Grant S. Joseph
Grant S. Joseph Colleen L. Seymour
Colleen L. Seymour- 1FitzPatrick Institute of African Ornithology, Department of Biological Sciences, University of Cape Town, Cape Town, South Africa
- 2Kirstenbosch Research Centre, South African National Biodiversity Institute, Cape Town, South Africa
A Commentary on
The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar
By Gillson L, Razanatsoa E, Razafimanantsoa AHI, Virah-Sawmy M and Ekblom A (2023). Front. Conserv. Sci. 4:1286459. doi: 10.3389/fcosc.2023.1286459
Introduction
Currently, 80% of Madagascar is treeless grassland. Identifying lost savanna woodland and grassland, forest, and heathland before pastoralism was introduced circa 0.5–1 ka (Hixon et al., 2021) informs conservation planning/restoration on the island. Gillson et al. (2023; hereafter G2023) warn that binary classification of “all savannas and heathlands as degraded forests is ecologically inaccurate”, setting a “false dichotomy between forest-grassland”, and “has diverted attention away from the loss and conversion of heathlands and savannas.”
We agree, and were surprised to see these statements (italicised) attributed to us (Joseph and Seymour, 2020, 2021; henceforth J&S20,21), two years after we debunked outdated “forest-grassland” dichotomies for Madagascar’s Central Highlands (MCH). We concluded: “This interdisciplinary review challenges the century-old extreme perspectives … Evidence supports neither (1) a forested MCH with secondary grasslands … nor (2) MCH characterised by vast natural treeless grassland … Findings support a more wooded, more ericoid-rich past, with grasses coexisting with trees as woodland understory and as components of ericoid systems. At fine scales, a complex mosaic … seems likely, including smaller treeless grasslands”. We postulated an eight-habitat mosaic (not two), with savanna >30%, and 10-fold more heathland than today (Joseph et al., 2021). We clearly (1) opposed and disproved the dichotomy, and (2) have never found “all savannas and heathlands” to be degraded forest.
Trees/grass/savannas and fire
Savanna biomes are characterised by a continuous C4 grass layer, ranging from grass-dominated systems to tree-dense savanna woodland. Although both are savanna, woodland is functionally distinct from treeless grassland. Annually, >100,000 km2 of MCH grassland burns anomalously relative to comparable ecoregions globally (Joseph et al., 2023). Most (95%–99%) fires are human-lit, and most severe where human densities are lowest (Kull, 2003). Globally, savanna tree-cover >45% limits fire, but tree-cutting/frequent fires convert woodland (Trapnell, 1959) and heathland (Gil-Romera et al., 2019) to fire-adapted grassland lacking trees, which can expand into regions where climate supports forest (Staver et al., 2011).
Wakeling et al. (2015), citing Bond et al. (2008), specified Madagascar’s “treeless grassland” as systems where “trees are absent”, and despite annual human-lit fires (Kull, 2003), consider them ancient, non-anthropogenic systems, formed before human settlement. This view proposes woodland as naturally <5% (Solofondranohatra et al., 2018); with grassland-limited fauna supporting their antiquity (Bond et al., 2008); that human-lit fires and cattle are surrogates for natural fires/endemic grazers, and that obligate grazing lawns coevolved with extinct C4-feeding giant tortoises and “pure” C4-grazing hippos (Bond et al., 2008; Solofondranohatra et al., 2020). We provide an alternative: Figure 1 details evidence for humans extirpating tree-adapted, forest-feeding megafauna that were never pure C4-feeders, transforming (by cutting/burning) a mosaic of larger woodlands/forests/ericoids and smaller grasslands circa 1 ka, to derived treeless grassland, maintained by introduced cattle and human-lit fire, where fire-adapted cosmopolitan grasses outcompete endemic grasses.
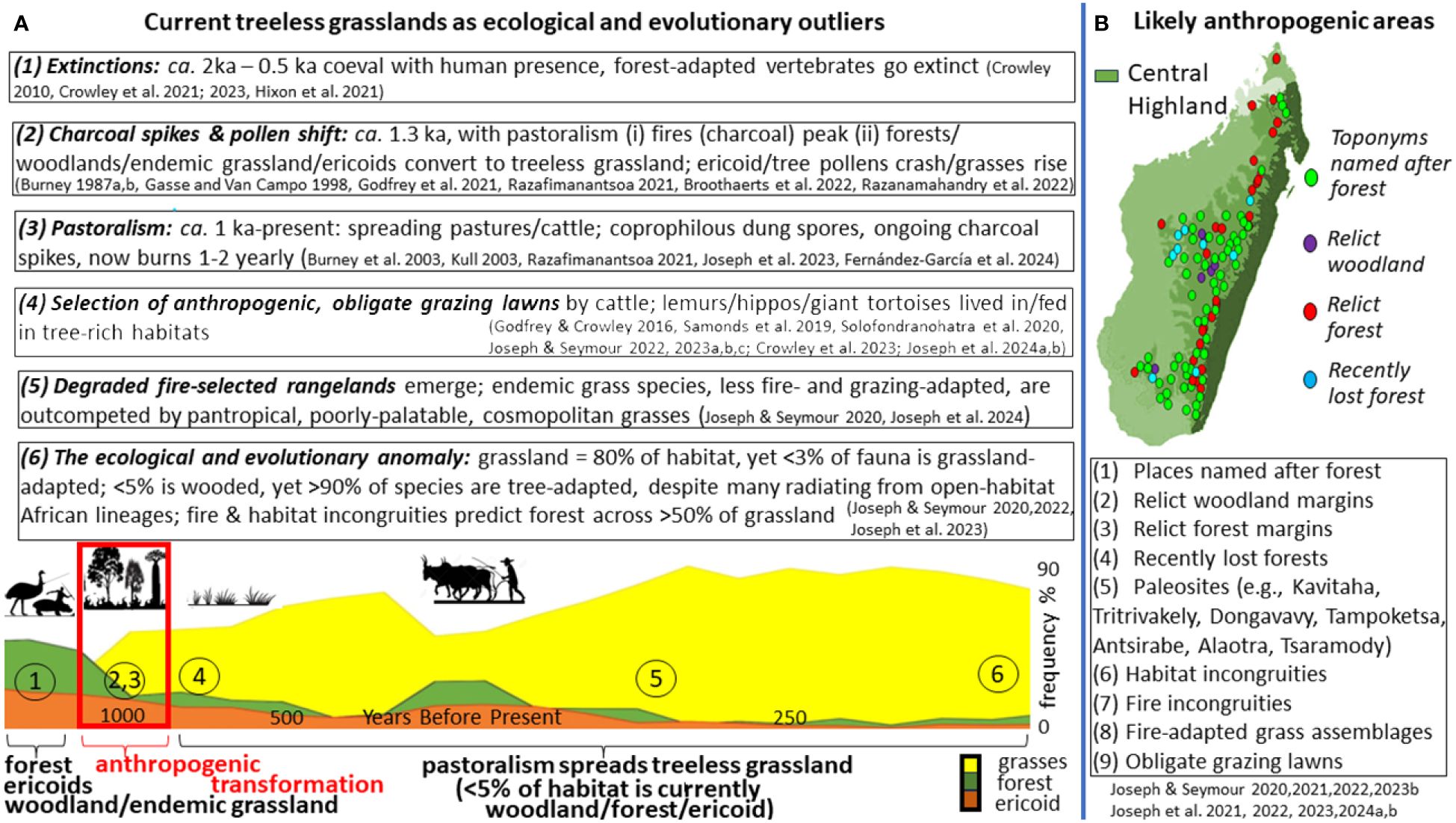
Figure 1 Ecological and evolutionary anomalies across regularly burned, tree-limited grassland support anthropogenic habitat transformation from forest/woodland/ericoids/endemic grasslands. (A) Grass pollens dominate palaeosites following pastoralism (e.g., Tampoketsa-Ankazobe, adapted from G2023; see also Godfrey et al., 2019). (B) Areas of likely anthropogenic transformation (>50% of treeless grasslands; box); grasslands named after forest. Cited in figure: Burney (1987a, 1987b); Gasse and Van Campo (1998); Bureny et al. (2003); Kull (2003); Crowley (2010); Godfrey and Crowley (2016); Samonds et al. (2019); Solofondranohatra et al. (2020); Crowley et al. (2021, 2023); Crowley (2021); Hixon et al. (2021); Razafimanantsoa (2021); Broothaerts et al. (2022); Joseph and Seymour (2020, 2021, 2022, 2023a, 2023b, 2023c); Razanamahandry et al. (2022); Joseph et al. (2021, 2022, 2023, 2024a, 2024b); Fernández-García et al. (2024).
Woodlands are savannas; ericoid-habitats are heathlands
Woodlands, with their C4-grass layer fall within the savanna biome. They are distinct from forest (closed-canopy biomes, where shade limits C4-grasses). For G2023 to ascribe (1) a binary “forest-grassland” dichotomy, and (2) “all savannas and heathlands” being “degraded forests” to J&S20,21, requires two errors.
The first is conflating woodland (savanna) and forest, creating an ecologically irrational dichotomy (woodland/forest vs. grassland). This is strange, because G2023 understand woodland as savanna, stating: “Tapia … woodlands are now classified as savannas”. They also clearly regard savannas and forests as separate categories (“savannas and heathlands as degraded forests is … inaccurate”). J&S20 unambiguously describe savanna: “there is no debating the ancient existence of grassy woodland, nor of grassy patches vacillating in size with prevailing fire and climate conditions”. J&S20 define “open canopy” Tapia with C4-grass understories as akin to Miombo (savanna woodland), but distinct from closed-canopy forest. Similarly, J&S21 emphasise “forest-limited endemic fauna support anthropogenic transformation from woodland”. If tree-rich savanna woodland had not been anthropogenically transformed, aspects of today’s grasslands would likely have harboured tree-adapted fauna, which are now limited to forest. Equally strange is G2023’s missing our nine vegetation studies, citing instead papers documenting faunal radiation (J&S20,21 find >90% of species are adapted to tree-rich habitat; <3% are grassland-limited).
The second error is disregarding our reference to ericoids. J&S20,21 describe transformation of grassy woodlands/smaller grasslands/forests/ericoids to treeless grassland; J&S21 details pre-human heathland presence, noting “even today these areas harbour ericoids”. J&S20,21 are not ecologically inaccurate: woodlands with C4-grass understories are savannas; ericoid habitats are heathlands.
Preserving ecological accuracy
Where disturbance incongruities exist, re-evaluation is needed (Lehmann et al., 2011), because incongruities suggest anthropogenic transformation (Thompson et al., 2021; Joseph et al., 2022). G2023 assert J&S20,21 miss “a key challenge for an informed and bottom up approach to conservation and restoration, which is to distinguish which open landscapes are ancient or anthropogenic, their historic nature and their current value to communities in ecosystem service provision”. Yet our findings (1) report nine ways of identifying regions/sites likely degraded by anthropogenic impacts (involving >50% of grasslands; Figure 1A); (2) promote indigenous knowledge/history (much grassland is named after forest; Figure 1B), and (3) highlight anthropogenic, island-wide ecosystem breakdown.
Regarding ecosystem services: ongoing tree-removal/fire-setting initiated >1 ka causes nutrient-rich soil loss (Brosens, 2022), silting lakes, lowering water quality (Bakoariniaina et al., 2006), and reducing marine and terrestrial diversity (Raharimahefa and Kusky, 2010). These degrade rangelands/grasslands/forests and threaten livelihoods (Styger et al., 2007; Neugarten et al., 2016; Joseph et al., 2023). Despite foreign, paternalistic promotion of burning practices, many pastoralists are aware that this exacerbates landscape degradation/poverty/armed conflict/gender inequality (malnutrition is pervasive; UNICEF, 2021) and strive for economic diversification (Joseph et al., 2024b).
Conclusions
The binary classification/ecological inaccuracy raised by G2023 is not held by mainstream researchers, who support pre-settlement habitat mosaics (Joseph et al., 2021; Lehmann et al., 2022; Crowley et al., 2023). The “key challenge” of identifying anthropogenically-impacted areas, and their value/risk to people, is not “missing” (G2023), but well underway. Conservation lacks resources: framing research around already-resolved debates (there is no dichotomy) and mis-citations undermines transdisciplinary cooperation and introduces unnecessary distractions.
Author contributions
GJ: Conceptualization, Writing – original draft, Writing – review & editing. CS: Conceptualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bakoariniaina L. N., Kusky T., Raharimahefa T. (2006). Disappearing Lake Alaotra: Monitoring catastrophic erosion, waterway silting, and land degradation hazards in Madagascar using Landsat imagery. J. Afr. Earth Sci. 44, 241–252. doi: 10.1016/j.jafrearsci.2005.10.013
Bond W. J., Silander J. A., Ranaivonasy J., Ratsirarson J. (2008). The antiquity of Madagascar’s grasslands and the rise of C4 grassy biomes. J. Biogeogr. 35, 1743–1758. doi: 10.1111/j.1365-2699.2008.01923.x
Broothaerts N., Razanamahandry V. F., Brosens L., Campforts B., Jacobs L., Razafimbelo T., et al. (2022). Vegetation changes and sediment dynamics in the Lake Alaotra region, central Madagascar. Holocene, 095968362211453. doi: 10.1177/09596836221145376
Brosens L. (2022). Under pressure: lavaka in the changing environment of Madagascar. PhD. Leuven (Netherlands: University of Leuven) 33.
Burney D. (1987a). “Pre-settlement vegetation changes at Lake Tritrivakely, Madagascar,” in Paleoecology of Africa and of the surroundings islands. Ed. Coetzee J. A., (Amsterdam, Netherlands: AA Balkema publishers) 375–381.
Burney D. A. (1987b). Late Holocene vegetational change in central Madagascar. Quat. Res. 28, 130–143. doi: 10.1016/0033-5894(87)90038-X
Burney D. A., Robinson G. S., Burney L. P. (2003). Sporormiella and the late Holocene extinctions in Madagascar. Proc. Natl. Acad. Sci. U.S.A. 100, 10800–10805. doi: 10.1073/pnas.1534700100
Crowley B. E. (2010). A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quat. Sci. Rev. 29, 2591–2603. doi: 10.1016/j.quascirev.2010.06.030
Crowley B. E., Godfrey L. R., Hansford J. P., Samonds K. E. (2021). Seeing the forest for the trees-and the grasses: revisiting the evidence for grazer-maintained grasslands in Madagascar’s Central Highlands. Proc. R. Soc. B. 288, 20201785. doi: 10.1098/rspb.2020.1785
Crowley B. E., Godfrey L. R., Samonds K. E. (2023). What can hippopotamus isotopes tell us about past distributions of C4 grassy biomes on Madagascar? Plants People Planet. 5, 997–1010. doi: 10.1002/ppp3.10402
Fernández-García V., Franquesa M., Kull C. A. (2024). Madagascar’s burned area from Sentinel-2 imagery (2016–2022): Four times higher than from lower resolution sensors. Sci. Total Env. 914, 169929. doi: 10.1016/j.scitotenv.2024.169929
Gasse F., Van Campo E. (1998). A 40,000-yr pollen and diatom record from Lake Tritrivakely, Madagascar, in the southern tropics. Quat. Res. 49, 299–311. doi: 10.1006/qres.1998.1967
Gillson L., Razanatsoa E., Razafimanantsoa A. H. I., Virah-Sawmy M., Ekblom A. (2023). The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar. Front. Cons. Sci. 4. doi: 10.3389/fcosc.2023.1286459
Gil-Romera G., Adolf C., Benito B. M., Bittner L., Johansson M. U., Grady D. A., et al. (2019). Long-term fire resilience of the Ericaceous Belt, Bale Mountains, Ethiopia. Biol. Lett. 15, 20190357. doi: 10.1098/rsbl.2019.0357
Godfrey L. R., Crowley B. E. (2016). Madagascar’s ephemeral palaeo-grazer guild: Who ate the ancient C4 grasses? Proc. R. Soc B 283, 5–8. doi: 10.1098/rspb.2016.0360
Godfrey L. R., Scroxton N., Crowley B. E., Burns S. J., Sutherland M. R., Pérez V. R., et al. (2019). A new interpretation of Madagascar’s megafaunal decline: The “Subsistence Shift Hypothesis” J. Hum. Evol. 130, 126–140. doi: 10.1016/j.jhevol.2019.03.002
Hixon S. W., Douglass K. G., Crowley B. E., Rakotozafy L. M. A., Clark G., Anderson A., et al. (2021). Late Holocene spread of pastoralism coincides with endemic megafaunal extinction on Madagascar. Proc. R. Soc B 288 (1955), 20211204. doi: 10.1098/rspb.2021.1204
Joseph G. S., Rakotoarivelo A. R., Pedrono M., Seymour C. L. (2024a). Can rewilding with giant tortoises increase woody habitat and limit fire across Madagascar’s grasslands? Plants People Planet. doi: 10.1002/ppp3.10474
Joseph G. S., Rakotoarivelo A., Seymour C. L. (2021). How expansive were Malagasy Central Highland forests, ericoids, woodlands and grasslands? A multidisciplinary approach to a conservation conundrum. Biol. Conserv. 261, 1–13. doi: 10.1016/j.biocon.2021.109282
Joseph G. S., Rakotoarivelo A. R., Seymour C. L. (2022). Tipping points induced by palaeo-human impacts can explain presence of savannah in Malagasy and global systems where forest is expected. Proc. R. Soc B 289, 1–10. doi: 10.1098/rspb.2021.2771
Joseph G. S., Rakotoarivelo A. R., Seymour C. L. (2024b). A review of Madagascar’s derived grasslands: Low palatability following anthropogenic fires may threaten food security. Plants People Planet 6, 67–78. doi: 10.1002/ppp3.10408
Joseph G. S., Seymour C. L. (2020). Madagascan highlands: originally woodland and forest containing endemic grasses, not grazing-adapted grassland. Proc. R. Soc B 287, 2020195620201956. doi: 10.1098/rspb.2020.1956
Joseph G. S., Seymour C. (2021). The unlikely “antiquity of Madagascar’s grasslands”: disproportionately forest-limited endemic fauna support anthropogenic transformation from woodland. J. Biogeogr. 48, 2111–2115. doi: 10.1111/jbi.14132
Joseph G. S., Seymour C. L. (2022). Are Madagascar’s obligate grazing-lawns ancient and evolved with endemic herbivores, or recently selected by introduced cattle? Biol. Lett. 18, 1–3. doi: 10.1098/rsbl.2022.0212
Joseph G. S., Seymour C. L. (2023a). Dispersal limitation and fire feedbacks maintain mesic savannas in Madagascar: Comment. Ecology 104, 1–4. doi: 10.1002/ecy.4045
Joseph G. S., Seymour C. L. (2023b). Why have Madagascar’s mammals, despite being closely-related to African open-habitat specialists, failed to radiate into open grasslands? J. Biogeogr. 50, 622–627. doi: 10.1111/jbi.14550
Joseph G. S., Seymour C. L. (2023c). Anthropogenic disturbance, not topographic dispersal barriers, limit forest and maintain Madagascar’s derived savannas. Bull. Ecol. Soc. Am. 104, e2087. doi: 10.1002/bes2.2087
Joseph G. S., Seymour C. L., Rakotoarivelo A. R. (2023). Fire incongruities can explain widespread landscape degradation in Madagascar’s forests and grasslands. Plants People Planet. doi: 10.1002/ppp3.10471
Kull C. (2003). “Fire and the management of highland vegetation,” in The Natural History of Madagascar. Eds. Goodman S., Benstead J. P. (The University of Chicago Press, Chicago), 153–157.
Lehmann C. E. R., Archibald S. A., Hoffmann W. A., Bond W. J. (2011). Deciphering the distribution of the savanna biome. New Phytol. 191, 197–209. doi: 10.1111/j.1469-8137.2011.03689.x
Lehmann C. E. R., Solofondranohatra C., Morton J., Phelps L., Ralimanana H., Razanatsoa J., et al. (2022). “The grassy ecosystems of Madagascar,” in The New Natural History of Madagascar. Ed. Goodman S. M. (Princeton University Press, Princeton, NJ), 152–168. doi: 10.2307/j.ctv2ks6tbb.27
Neugarten R. A., Honzak M., Carret P., Koenig K., Andriamaro L., Cano C. A., et al. (2016). Rapid assessment of ecosystem service co-benefits of biodiversity priority areas in Madagascar. PloS One 11, 1–25. doi: 10.1371/journal.pone.0168575
Raharimahefa T., Kusky T. M. (2010). Environmental monitoring of Bombetoka bay and the Betsiboka estuary, Madagascar, using multi-temporal satellite data. J. Earth Sci. 21, 210–226. doi: 10.1007/s12583-010-0019-y
Razafimanantsoa A. H. I. (2021). Holocene vegetation history and environmental change in the forest-grassland mosaic of the Central Highlands of Madagascar. PhD. Leuven (South Africa: University of Cape Town).
Razanamahandry V. F., Dewaele M., Govers G., Brosens L., Campforts B., Jacobs L., et al. (2022). Stable isotope profiles of soil organic carbon in forested and grassland landscapes in the Lake Alaotra basin (Madagascar): insights in past vegetation changes. Biogeosci 19, 3825–3841. doi: 10.5194/bg-19-3825-2022
Samonds K. E., Crowley B. E., Rasolofomanana T. R. N., Andriambelomanana M. C., Andrianavalona H. T., Ramihangihajason T. N., et al. (2019). A new late Pleistocene subfossil site (Tsaramody, Sambaina basin, central Madagascar) with implications for the chronology of habitat and megafaunal community change on Madagascar’s Central Highlands. J. Quat. Sci. 34, 379–392. doi: 10.1002/jqs.3096
Solofondranohatra C., Vorontsova M. S., Hackel J., Besnard G., Cable S., Williams J., et al. (2018). Grass functional traits differentiate forest and savanna in the Madagascar central highlands. Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00184
Solofondranohatra C., Vorontsova M. S., Hempson G. P., Hackel J., Cable S., Vololoniaina J., et al. (2020). Fire and grazing determined grasslands of central Madagascar represent ancient assemblages. Proc. R Soc B 287, 20200598. doi: 10.1098/rspb.2020.0598
Staver A. C., Archibald S., Levin S. A. (2011). The global extent and determinants of savanna and forest as alternative biome states. Sci. (1979) 334, 230–232. doi: 10.1126/science.1210465
Styger E., Rakotondramasy H. M., Pfeffer M. J., Fernandes E. C. M., Bates D. M. (2007). Influence of slash-and-burn farming practices on fallow succession and land degradation in the rainforest region of Madagascar. Agric. Ecosyst. Environ. 119, 257–269. doi: 10.1016/j.agee.2006.07.012
Thompson J. C., Wright D. K., Ivory S. J., Choi J. H., Nightingale S., Mackay A., et al. (2021). Early human impacts and ecosystem reorganization in southern-central Africa. Sci. Adv. 7, eabf9776. doi: 10.1126/sciadv.abf9776
Trapnell C. G. (1959). Ecological results of woodland burning experiments in Northern Rhodesia. J. Ecol. 47, 129–168. doi: 10.2307/2257252
UNICEF (2021) UNICEF MADAGASCAR. Available online at: https://www.unicef.org/Madagascar/en/programme/nutrition (Accessed October 6, 2021).
Keywords: anthropogenic fire, vegetation mosaic, Tapia, soil erosion, biodiversity hotspot, woodland, heathlands, treeless grassland
Citation: Joseph GS and Seymour CL (2024) Commentary: The role of palaeoecology in reconciling biodiversity conservation, livelihoods, and carbon storage in Madagascar. Front. Conserv. Sci. 5:1389498. doi: 10.3389/fcosc.2024.1389498
Received: 21 February 2024; Accepted: 25 March 2024;
Published: 10 April 2024.
Edited by:
Richard T. Corlett, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Thomas Edward Marler, University of Guam, GuamCopyright © 2024 Joseph and Seymour. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Grant S. Joseph, a2Fyb29ncmFudEBnbWFpbC5jb20=