 Jing Zhou
Jing Zhou Wang Hou
Wang Hou Huilin Zhong
Huilin Zhong Dan Liu
Dan Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol., 19 August 2024
Sec. Clinical Microbiology
Volume 14 - 2024 | https://doi.org/10.3389/fcimb.2024.1401448
This article is part of the Research TopicPathogenesis, Diagnosis, and Treatments of SARS-CoV-2 Co-infection with Influenza Viruses or Other Respiratory PathogensView all 9 articles
The lungs, as vital organs in the human body, continuously engage in gas exchange with the external environment. The lung microbiota, a critical component in maintaining internal homeostasis, significantly influences the onset and progression of diseases. Beneficial interactions between the host and its microbial community are essential for preserving the host’s health, whereas disease development is often linked to dysbiosis or alterations in the microbial community. Evidence has demonstrated that changes in lung microbiota contribute to the development of major chronic lung diseases, including chronic obstructive pulmonary disease (COPD), idiopathic pulmonary fibrosis (IPF), asthma, and lung cancer. However, in-depth mechanistic studies are constrained by the small scale of the lung microbiota and its susceptibility to environmental pollutants and other factors, leaving many questions unanswered. This review examines recent research on the lung microbiota and lung diseases, as well as methodological advancements in studying lung microbiota, summarizing the ways in which lung microbiota impacts lung diseases and introducing research methods for investigating lung microbiota.
The lung microbiota is an important component of the human microbiota (Kovaleva et al., 2019). It consists of the entire microbial community in the lungs, including bacteria, viruses, and fungi, forming a biological system that interacts with the host’s lung microenvironment in cellular signaling pathways and metabolic products, influencing each other. Studies have shown that the lung microbiota is established at birth, and the respiratory microbial community continues to develop during the first two years of life (Chu et al., 2017). The establishment of the microbiota is a crucial factor in the formation of a mature lung immune system and in protecting the lungs from harmful inflammatory responses (El Tekle and Garrett, 2023). It is involved in the normal development of the respiratory tract, regulating respiratory immunity, and maintaining respiratory health by preventing the spread of pathogens (Hou et al., 2022). Any dynamic system can be dysregulated by internal or external factors, and the same is true for lung microbiota. The imbalance in the lung microbiota may trigger or exacerbate respiratory diseases such as chronic obstructive pulmonary disease, asthma, and lung cancer (Figure 1) (Maddi et al., 2019). Investigating the mechanisms of lung microbiota and its microenvironment changes in the occurrence and development of lung diseases is of significant importance in exploring new potential therapeutic targets.
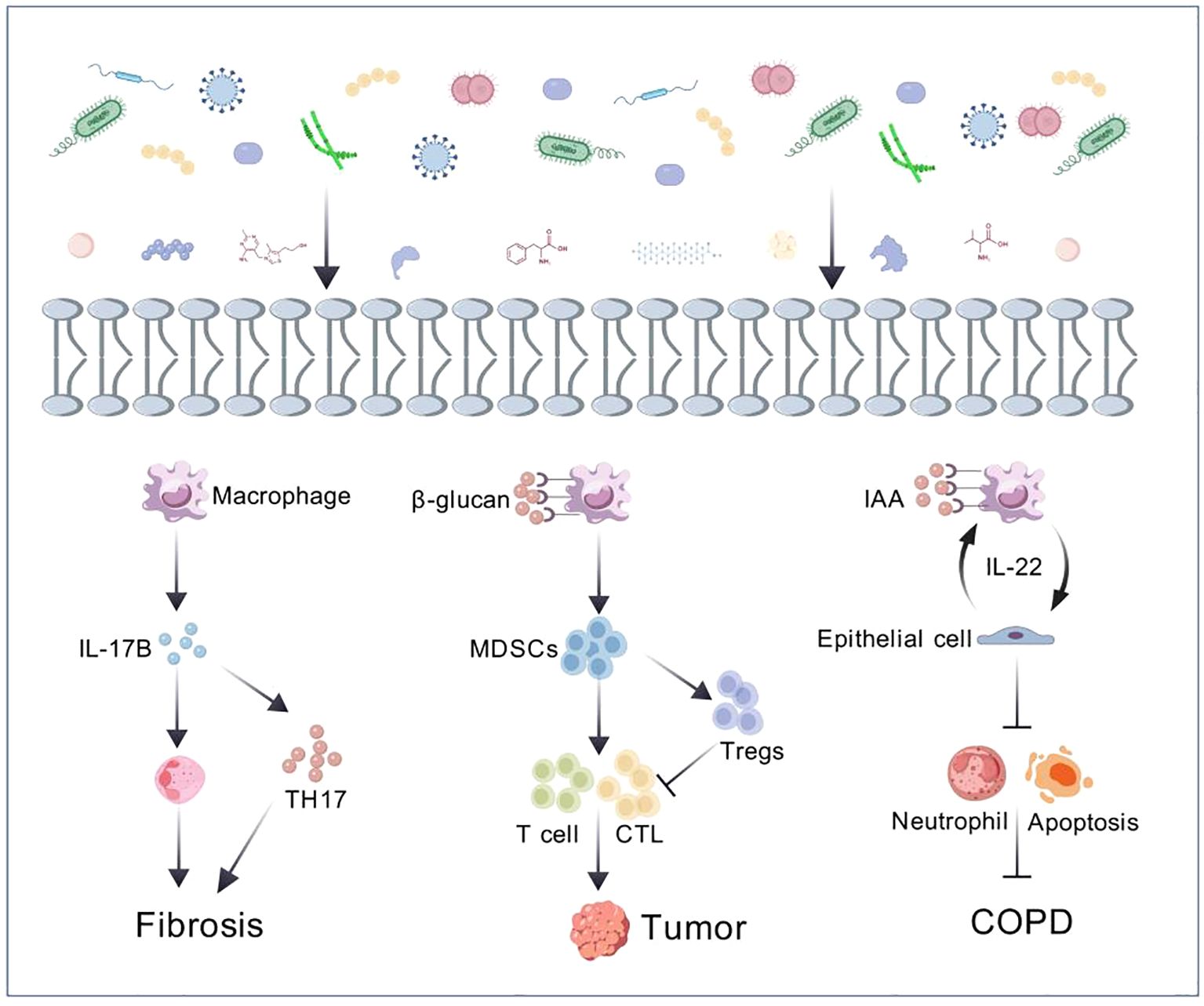
Figure 1. The crosstalk between microbiome and host (Yang et al., 2019; Liu et al., 2023; Yi et al., 2022).
Although research on the lung microbiota is still in its early stages compared to the well-studied gut microbiota, studies have found that the microbial community in lung tissue also plays a functional role in the progression of lung diseases (Meng et al., 2023; Natalini et al., 2023a). Furthermore, on account of the unique characteristics of the lung microbiota (significantly smaller scale than the gut microbiota but highly diverse and more sensitive to environmental influences and greatly affected by oral and upper respiratory tract microbiota), the development of research techniques targeting the lung microbiota is essential for advancing research. To comprehensively understand the role of local microbiota in diseases, this review reports on the microbial characteristics of several chronic lung diseases, the mechanisms by which the microbiota promotes the occurrence and development of diseases, and recent cutting-edge research methodologies related to lung microbiota.
The lung microbiota are mainly obtained from the exchange of nasopharynx, oropharynx and ambient air (Tsay et al., 2021). The lung microbiota of healthy individuals is mainly composed of Streptococcus (Firmicutes), Fusobacterium (Fusobacteria), Haemophilus (Proteobacteria), Bacteroides (Bacteroidetes), Pseudomonas (Proteobacteria), Prevotella (Bacteroidetes), and Neisseria (Proteobacteria) (Charlson et al., 2011; Yang et al., 2018; Zhu and Chang, 2023). In addition to bacteria, fungi and viruses also contribute to the lung’s microbial environment. A study on post-antibiotic mouse colonies suggests that the presence of fungi can influence the composition of lung bacteria and the host’s response (Erb Downward et al., 2013). There is relatively less reporting on viruses in the microbiota, but viruses are also present in the blood of healthy individuals. Viruses exhibit high specificity to their hosts and are relatively stable (Minot et al., 2011; Abeles et al., 2014). Under normal circumstances, the lung microbiota is in a balanced state of migration and elimination, with various symbiotic microorganisms in relatively balanced and stable quantities (Spijkerman et al., 2012; Mika et al., 2015). When the lung microbiota is imbalanced, it can affect the immune microenvironment by releasing metabolic products, inducing inflammatory processes, producing bacterial toxins that alter the stability of the host’s genome, and increasing levels of carcinogenic microbial metabolites, thereby leading to the occurrence and development of diseases (Table 1) (Mao et al., 2018; Tsay et al., 2021; Campbell et al., 2022). For example, environmental exposures such as cigarette smoke, PM2.5 and air pollutants can lead to increased respiratory symptoms and lung damage. Elevated levels of Atopobium, Actinomyces and Prevotella have been reported in smokers compared to non-smokers). In addition, fungal taxa and especially Cladosporium are associated with PM2.5 concentrations (Lin et al., 2023). Exposure to PM2.5 can lead to lung inflammation and oxidative stress (Wang S. M. et al., 2022).
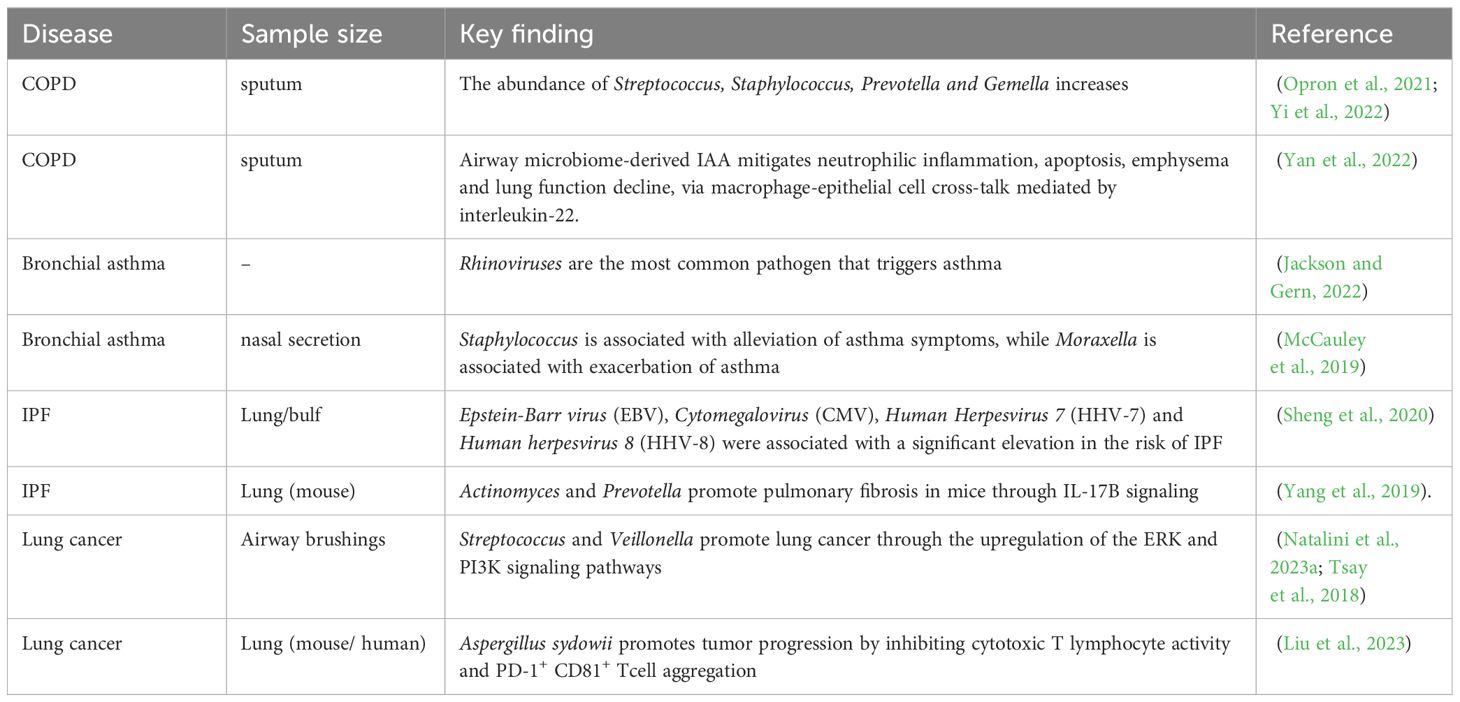
Table 1. Summary of key findings on the lung microbiota in chronic pulmonary diseases.
Chronic obstructive pulmonary disease (COPD) is defined by persistent inflammation in the airways, parenchymal part of lung tissue, and pulmonary vessels, and it is progressive and irreversible, making it one of the major contributor of death from chronic lung diseases (Devadoss et al., 2019). COPD has several risk factors, including smoking, genetic factors, environmental pollution, and infection (Labaki and Rosenberg, 2020). Colonization and infection of airway bacteria are the main triggering factors for acute exacerbations of COPD (Meldrum et al., 2024). On one hand, bacteria release bacterial products such as oligosaccharide lipids or other soluble bacterial toxins, causing damage to airway epithelial cells. On the other hand, it can cause local inflammatory reactions, with inflammatory cells releasing cytokines and increasing elastase activity, disrupting the balance of elastase/anti-elastase systems, thereby promoting the progression of COPD and leading to irreversible lung damage (Leung et al., 2017; Pathak et al., 2020; Isaacs et al., 2023). Compared to healthy individuals, the relative abundance of Actinobacteria decreases in COPD patients, while the relative abundance of Haemophilus increases, which shows a positive correlation with interleukin-8 (IL-8) levels in sputum (Wang Z. et al., 2021). Another study indicates that the presence of a microbial community in sputum dominated by Proteobacteria in COPD patients is associated with poorer lung function and disease progression (Dicker et al., 2021). Furthermore, clinical phenotypes of COPD can be distinguished by respiratory microbiota and can better predict patient response to antibiotic therapy (Wang Z. et al., 2021). For example, patients with a neutrophilic inflammatory phenotype are often accompanied by bacterial infections and require antibiotic treatment, while patients with an eosinophilic inflammatory phenotype often show no signs of infection. The alpha diversity of the lower respiratory tract microbiota decreases in COPD patients after glucocorticoid treatment, with an increase in Moraxella and Haemophilus abundance and a decrease in Streptococcus abundance, while the use of antibiotics shows opposite results (Wang et al., 2016). The respiratory microbiota undergoes significant changes during acute exacerbations of COPD compared to stable periods, with decreased microbial diversity and increased abundance of Proteobacteria, particularly Haemophilus and Moraxella, and a significant decrease in Staphylococcus (Sun et al., 2020; Zheng et al., 2022). Viral infections are among the factors contributing to acute exacerbations, and nasal virus infection in COPD patients can enhance neutrophil elastase-mediated antimicrobial peptide degradation. This virus-induced increase in secondary bacterial infections leads to an increase in Haemophilus abundance and microbial dysbiosis in the lungs (Zheng et al., 2022).
Respiratory viruses and fungi are associated with exacerbation of COPD. Rhinovirus is the most common type of viral infection that exacerbates COPD (Stolz et al., 2019). In addition, Influenza Virus and Respiratory Syncytial Virus (RSV) are frequently detected in the respiratory tract of COPD patients (Simon et al., 2023). COPD patients have high expression of ACE2 (the receptor for SARS-CoV-2), making them more susceptible to COVID-19 (Higham et al., 2020). Multiple studies have shown that virus-induced COPD exacerbations may be related to interferon IFN (Garcia-Valero et al., 2019; Collinson et al., 2021). In addition to viruses, the role of fungi in COPD patients is gradually being recognized, and fungal sensitization is prevalent in COPD patients and associated with poor outcomes (Tiew et al., 2020). For example, a prospective multicenter study from Singapore found that Aspergillus sensitization is associated with COPD exacerbations (Tiew et al., 2023).
It has been reported that most COPD exacerbations are caused by bacterial or viral infections (Kim et al., 2024). Due to the downregulation of pattern recognition receptors (PRRs) such as Toll-like receptors (TLRs) and Nod-like receptors (NLRs) on the airway epithelial cells of COPD patients, the recognition ability of bacterial pathogens is impaired, leading to delayed and insufficient immune responses (Sidletskaya et al., 2020). Bacterial infections in COPD patients can induce oxidative stress, produce reactive oxygen species (ROS), and impaired phagocytic function (Yamasaki and van Eeden, 2018; Singh et al., 2021). Most respiratory viruses target airway epithelial cells, causing epithelial barrier disruption, microvascular dilatation, edema, and immune cell infiltration, which can lead to increased levels of CD8+ T cells, neutrophils, eosinophils, TNF-n and IFN-n in COPD patients (Paats et al., 2012). Additionally, COPD is characterized by specific fungal genera such as Aspergillus, Curvularia and Penicillium (Tiew et al., 2021). Environmental exposure is the main source of fungal allergens and Aspergillus can form biofilms on the airway epithelial cells of COPD patients to resist host immunity and antifungal therapy. It can also lead to impairment of neutrophil function and increased apoptosis, resulting in disease progression (Tiew et al., 2020).
Bronchial asthma (asthma) is a common respiratory system disease characterized by recurrent wheezing, shortness of breath, chest tightness, or coughing. Colonization or infection of microorganisms in the upper respiratory tract and lower respiratory tract can lead to the onset of asthma (Gon and Hashimoto, 2018). The mechanism involved mainly includes promoting IgE synthesis and histamine release, leading to a hypersensitive state of the body, promoting the release of various cytokines. This process triggers numerous allergic responses, such as eosinophilic inflammation, transformation of immunoglobulin (IgG) into IgE, promotion of B cell proliferation, goblet cell transformation, and the consequent mucus secretion, exacerbating airway inflammation and damage, leading to airway spasm, edema, and exudation. Changes in the local microbial community lead to local immune dysfunction, resulting in the occurrence of asthma (Whetstone et al., 2022). Studies have shown that changes in the microbiota in asthma have a significant impact on the pathophysiology of the disease (Barcik et al., 2020; Santos et al., 2021). For example, the abundance of neutrophils in the sputum of asthma patients is related to the levels of specific taxa, including Moraxella (Ma et al., 2021). Researchers have detected a variety of microorganisms in the lower respiratory tract microbiota of asthmatic children, including Bacteroides, Faecalibacterium, Roseburia, Moraxella, Staphylococcus, and Streptococcus (Goldman et al., 2018; Al Bataineh et al., 2020). Among these microorganisms, Staphylococcus is associated with alleviation of asthma symptoms, while Moraxella is associated with exacerbation of asthma (McCauley et al., 2019). In addition, an increase in Proteobacteria and an elevation of non-Proteobacteria such as Pseudomonas, Clostridium, and members of the family Enterobacteriaceae have been observed in the airways of asthma patients (Azim et al., 2021), and are significantly associated with the expression of Th17-related genes, which may lead to recruitment of neutrophils (Wilburn et al., 2023). Rhinovirus is the most common pathogen triggering asthma, followed by Human Bocavirus and Human Metapneumovirus (10-25% positivity) (Coverstone et al., 2019). CDHR3 has been found to be highly expressed in differentiated bronchial epithelial cells and acts as a receptor for Rhinovirus C to increase the risk of respiratory disease (Bonnelykke et al., 2018). These studies indicate that changes in the microbial community are not only related to asthma but may also play a role in the changes of asthma symptoms.
Idiopathic Pulmonary Fibrosis (IPF) is the most common and prevalent type of pulmonary fibrosis. Activation of cells in the alveolar region leads to the release of a large number of cytokines and growth factors, promoting the recruitment, proliferation and differentiation of lung fibroblasts into myofibroblasts, resulting in progressive lung parenchymal damage. This process leads to irreversible decline in lung function and even respiratory failure (Moss et al., 2022). One multicenter study showed that Human Herpesvirus 7 (HHV-7), Human Herpesvirus 8 (HHV-8), Epstein-Barr virus (EBV), and Cytomegalovirus (CMV) were associated with a significantly increased risk of IPF (Sheng et al., 2020). Testing of the lower airways of IPF patients has revealed an increased abundance of Haemophilus, Veillonella, Streptococcus, and Neisseria (Zhang T. et al., 2023). It has been reported a positive correlation between the concentration of IL-6 in the alveoli of IPF patients and the relative abundance of Firmicutes, while the concentration of IL-12p70 in the alveoli was negatively correlated with the relative abundance of Proteobacteria (O'Dwyer et al., 2019). Researchers have found that peptides secreted by Staphylococcus induce apoptosis of lung epithelial cells and collagen deposition, leading to acute exacerbation of IPF and further inhibition of these apoptotic peptides can improve acute exacerbation of pulmonary fibrosis (D'Alessandro-Gabazza et al., 2020). Another study demonstrated that Actinomyces and Prevotella promote pulmonary fibrosis in mice through IL-17B signaling (Yang et al., 2019). In a mouse model of bleomycin-induced pulmonary fibrosis, germ-free mice have a higher mortality rate compared to conventional mice, demonstrating the complex relationship between lung microbiota changes and IPF-related inflammatory activity. In conclusion, we can find that microorganisms may promote or inhibit IPF through certain key signaling pathways.
The incidence and mortality of lung cancer are among the highest globally (Leiter et al., 2023). Approximately 90% of lung cancer cases are attributed to risk factors such as smoking, tobacco smoke, air pollution and other carcinogens (Qi et al., 2023; Xue et al., 2023). Lung cancer patients show decreased alpha diversity and altered bacterial composition. Researchers have found a transition in dominant bacterial taxa from Firmicutes to Bacteroidetes in saliva and bronchoalveolar lavage samples of lung cancer patients (Xie et al., 2022) and a correlation between intratumoral bacteria and tumor type and subtype, patient smoking status and immune therapy response (Nejman et al., 2020). Chronic airway inflammation can increase susceptibility to lung cancer, suggesting that airway dysbiosis may be one of its pathogenic mechanisms (Goto, 2022). Studies have found enrichment of the airway commensal bacteria Megasphaera and Veillonella in the bronchoalveolar lavage fluid (BALF) of lung adenocarcinoma patients (Guo et al., 2022). Other researchers have found an abundance of Streptococcus and Veillonella in the lower respiratory tract of lung cancer patients, leading to upregulation of the ERK and PI3K signaling pathways, promoting lung cancer cell proliferation (Tsay et al., 2018). Bacterial metabolites such as reactive oxygen and nitrogen species can directly cause DNA damage and disrupt multiple signaling pathways, creating a pro-carcinogenic environment (Dong et al., 2021; Ho et al., 2021). In addition to bacteria, fungi such as Blastomyces (Li and Saxena, 2022) and Aspergillus sydowii (A. Sydowii) (Liu et al., 2023) have also been found in lung tumor tissues. A recent study revealed that the intratumoral fungus Aspergillus sydowii promoted lung cancer progression through IL-1ughsionlTE expansion and activation of myeloid-derived suppressor cells (MDSCs), and the enrichment of Aspergillus was closely associated with poorer prognosis in lung cancer patients (Liu et al., 2023). The relationship between microorganisms and their microenvironment with tumors is very close and a more comprehensive understanding of the character of the microbiota in lung cancer is essential. Given that the lung microbiome is associated with the prognosis of lung cancer patients and can promote lung cancer progression through key signaling pathways, it can serve as a critical diagnostic and preventive biomarker for lung cancer staging, genotyping and risk stratification (Poore et al., 2020).
Some studies have shown that microorganisms interact with the host through metabolites to regulate signaling pathways. For example, the indole IAA produced by Lactobacillus alleviates neutrophil inflammation, cell apoptosis, emphysema and lung function decline through IL-22-mediated macrophage-epithelial cell interaction (Yan et al., 2022). Additionally, Lactobacillus can metabolize dietary tryptophan into indole, thereby inhibiting tumor immunity and promoting the growth of pancreatic ductal adenocarcinoma (Hezaveh et al., 2022). Given that host-microbiota interactions are bidirectional, microbial-derived metabolites may interact with host macromolecules and affect their responses. Short-chain fatty acids (SCFAs), such as acetate, propionate and butyrate, reduce tumor necrosis factor TNF-o production by inhibiting histone deacetylase (HDAC) and suppressing the transcription factor NF-to (Chambers et al., 2018). Metabolomic changes can predict asthma outcomes, as researchers have found a positive correlation between 5’-AMP, uracil and niacinamide with asthma exacerbations using non-targeted sputum metabolomics (Liu et al., 2022). A study revealed that metabolites related to lipid peroxidation in urine samples are linked to the severity of asthma, lung function and eosinophilic inflammation in non-obese asthmatic individuals (Wang C. et al., 2021). Additionally, in a mouse model, fecal microbiota from patients with COPD was demonstrated to play a role in the onset of COPD (Li et al., 2021) and COPD patients and healthy individuals exhibit distinct microbial and metabolic features in fecal samples (Bowerman et al., 2020). NMR analysis of urine from pneumonia patients indicates that specific metabolic profiles can be used to differentiate pneumococcal pneumonia from pneumonia caused by other bacterial strains (Green et al., 2023). NMR analysis of urine from pneumonia animals infected with either Streptococcus pneumoniae or methicillin-resistant Staphylococcus aureus (MRSA) revealed different metabolic profiles (Green et al., 2023). These results suggest that metabolomics has potential in the diagnosis and monitoring of pneumonia.
All parts of the human body are colonized by microorganisms, with the gut harboring the highest density. The gut microbiota, comprising tens of trillions of symbiotic bacteria, fungi, archaea and viruses (Zhou et al., 2021), has garnered significant attention. The emerging concept of the gut-lung axis underscores the intricate interplay between lung and gut microbiota (Anand and Mande, 2018). Clinical studies indicate that lung diseases may be associated with gut microbiota (Li et al., 2021) and alterations in lung microbiota can lead to changes in the composition and metabolism of gut microbiota (Bowerman et al., 2020). Conversely, translocated gut microbiota and their products can influence pulmonary immunity (Özçam and Lynch, 2024). For instance, fecal microbiota from COPD patients has been shown to contribute to COPD development, with gut microbial-derived lipopolysaccharides (LPS) exacerbating COPD progression in mice (Li et al., 2021). Bowerman et al. identified a disease-related network linking Streptococcus parasanguinis_B with COPD-associated metabolites, such as N-acetylglutamate and its analogue, providing valuable insights for COPD biomarker discovery (Bowerman et al., 2020). Additionally, fecal microbiota transplantation in healthy mice has been demonstrated to attenuate emphysema development by inhibiting inflammation both locally and systemically, and by altering gut microbiota composition (Jang et al., 2020).
Currently, amplicon sequencing and metagenomics Next-Generation sequencing (mNGS), as well as targeted sequencing (tNGS), are the predominant sequencing technology used in microbiome research (Han et al., 2022). Amplicon sequencing targeting 16S rRNA region of bacteria gene or ITS region of fungi (Gao et al, 2023). Primarily involves PCR amplification of a partial region followed by high-throughput sequencing to detect sequence variations and abundance information. When studying lung microbes, the V4 or V3V4 region sequence is most often selected. For fungi, often a portion of ITS1 or ITS2 sequence was characterized. This method reveals the types, relative abundance, and evolutionary relationships of microorganisms in environmental samples (Liu et al., 2021). Due to its high specificity, sensitivity, throughput and simple data analysis process, amplicon sequencing is favored by many researchers and remains irreplaceable in microbiota studies (Zhang W. et al., 2023). However, this method has limitations as it is not applicable for sequencing viral genomes due to the lack of a conserved gene similar to the 16S rRNA gene in these organisms. Additionally, limited universal primers, methodological constraints, and high host contamination restrict the accurate reflection of actual microbial community structures in samples (Table 2) (Weinroth et al., 2022).

Table 2. Comparison of microbial sequencing method.
In the clinical setting, mNGS is applied to infectious disease diagnosis, respiratory microbiome analysis, human host response analysis to infections, drug resistance prediction, colonization and infection differentiation, as well as the identification of tumor-related viruses and their genomic integration sites in various syndromes and sample types (Diao et al., 2022; Ibañez-Lligoña et al., 2023). The main advantage of mNGS is its unbiased sampling, as it maps the obtained sequence information to microbial resource databases, overcoming the limitations of targeted detection methods by characterizing all microorganisms in the human body system, including viruses, fungi, bacteria and parasites, in a single test (Qin et al., 2022). mNGS can provide a comprehensive view of the microbial community structure and function. However, mNGS also has limitations, as most bacteria can only be identified at the genus level, and contamination may occur during sampling or DNA extraction processes (Wensel et al., 2022).
Targeted sequencing (tNGS), also known as pathogen-targeted sequencing, detects known pathogenic microbial information in samples through multiplex PCR. tNGS specifically amplifies target genes of interest, thereby avoiding the influence of the host and sampling process, making it a highly sensitive and cost-effective optimization method (Sibandze et al., 2022). tNGS technology is used for the detection of known pathogens and drug-resistant genes (Wang S. et al., 2022; Yi et al., 2024)]. However, this technology faces challenges in its development and application, including low detection rates due to non-specific amplification, primer dimer interference, and amplification bias, as well as the need for improvement in the recognition and detection of new pathogens and rare specimen capabilities (Li et al., 2021).
Metatranscriptomics can assess the gene expression of microorganisms to explain their composition and function in the environment, such as the lungs, oral cavity and gut. It allows us to understand the interactions between the microbe and the host (Ren et al., 2019). It overcomes the limitations of polymerase chain reaction (PCR) amplification and is not limited to the analysis of specific bacteria, making it particularly advantageous in characterizing host-microbe gene expression. However, it also has its limitations, such as the possibility of host RNA contamination and RNA degradation during processing (Gao et al., 2023). Therefore, it requires extra caution. In the analysis process, even using the same database, different macro-genomic analysis methods can sometimes produce different results.
Microbial metabolomics can provide accurate information about the actual physiological status of microorganisms, identifying immunomodulatory metabolites to reflect the health status of the environment or evidence of ecological imbalance (Liu et al., 2022). NMR, GC-MS, and LC-MS are several commonly used tools in microbial metabolomics (Ye et al., 2022). Sample pretreatment techniques based on NMR metabolomics are relatively simple, allowing for objective and non-destructive sample evaluation and identification, with stable and strong repeatability ofdetection results (Traverso et al., 2018; Valentino et al., 2020). NMR is better suited for analyzing compounds that are difficult to ionize and require derivatization. However, due to limited sensitivity, MS is a better choice for achieving higher sensitivity and separation efficiency. LC-MS and GC-MS can detect thousands of different metabolites in various metabolic areas at the micromolar to millimolar leve l (Misra, 2020). Currently, mass spectrometry ismainly used in clinical microbiological identification (Bauermeister et al., 2022). LC-MS is primarily used for the analysis and detection of stable compounds and large molecular compounds (including proteins, peptides, and polymers). Compared to LC-MS, GC-MS can relatively easily identify a larger proportion of metabolites, as well as separate, identify, and quantify molecules in mixed samples, making it the preferred tool for the analysis of volatile small molecule metabolites (Weisskopf et al., 2021).
To thoroughly understand the role of microorganisms in the human body and their impact on human health, it is necessary to use a combination of omics tools. Multi-omics integration is an inevitable trend in future research, and it is hoped that through multi-omics, we can gain a deeper understanding of the role of lung microorganisms in respiratory diseases and develop more effective disease treatment strategies.
Existing research indicates that the human microbiome plays a crucial role in the development and progression of human diseases, with the changes in the microbiome and its metabolites having a significant impact on the pathophysiology of diseases. Therefore, in order to conduct a more comprehensive study of the microbiome, we have listed several of the most mainstream microbiome sequencing methods. These sequencing tools can help us identify the presence of microorganisms, understand the dynamic changes of microbiota in diseases, assess their functions and their direct impact on the host. Furthermore, combined with metabolomics and other multi-omics methods for joint analysis, they can deepen our understanding of the molecular mechanisms underlying microbiome-related diseases.
Despite some achievements in certain aspects, the lung microbiome still faces challenges. Sampling is a key aspect in lung microbiome research. Compared to the skin and gut, the biomass in the lungs is low, making sampling and detection difficult (Sulaiman et al., 2021). Additionally, the upper respiratory tract serves as the entry point connecting with the external environment and is typically the first point of contact for inhaled pollutants and pathogens, thus sample data from the upper respiratory tract may influence disease prognosis (Kumpitsch et al., 2019; Tiotiu et al., 2020). Another significant limitation is that existing metabolomic analysis techniques cannot distinguish whether certain metabolites, such as histamine, originate from the host or from microorganisms if both produce the same metabolite (Yamauchi and Ogasawara, 2019; Krell et al., 2021). Furthermore, to better understand the relationship between the role of the respiratory microbiome and disease progression, more longitudinal studies are crucial.
In the clinical medical field, microbiota transplantation has been applied clinically, but only in the gastrointestinal tract (Waller et al., 2022). The impact of altering the respiratory microbiota on clinical treatment is unknown, but it will be a direction for future research. With the continuous advancement of technology, high-throughput sequencing techniques will continue to have a key role in microbiome research. Recently, two innovative technologies, 2bRAD-M simplified metagenome sequencing and MobiMicrobe high-throughput single-cell genome sequencing, have emerged (Sun et al., 2022; Zheng et al., 2022). These technologies can effectively handle low biomass, severe degradation, and high host-contaminated samples, overcoming the limitations of mainstream technologies and offering unique core advantages. In future research, these technologies will demonstrate their strengths and value in development and application, enabling tailored microbial intervention strategies for different individuals and applying them to the prevention and treatment of clinical diseases. This will give us a comprehensive understanding of the microbial communities in the human body.
JZ: Writing – original draft, Writing – review & editing. WH: Writing – original draft, Writing – review & editing. HZ: Writing – original draft, Writing – review & editing. DL: Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (82173182), Science and Technology Program of Sichuan (2023NSFSC1939), and 1.3.5 Project for Disciplines of Excellence, West China Hospital, Sichuan University (ZYJC 21054).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abeles, S. R., Robles-Sikisaka, R., Ly, M., Lum, A. G., Salzman, J., Boehm, T. K., et al. (2014). Human oral viruses are personal, persistent and gender-consistent. Isme J. 8, 1753–1767. doi: 10.1038/ismej.2014.31
Al Bataineh, M. T., Hamoudi, R. A., Dash, N. R., Ramakrishnan, R. K., Almasalmeh, M. A., Sharif, H. A., et al. (2020). Altered respiratory microbiota composition and functionality associated with asthma early in life. BMC Infect. Dis. 20, 697. doi: 10.1186/s12879-020-05427-3
Anand, S., Mande, S. S. (2018). Diet, microbiota and gut-lung connection. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02147
Azim, A., Green, B., Lau, L., Rupani, H., Jayasekera, N., Bruce, K., et al. (2021). Peripheral airways type 2 inflammation, neutrophilia and microbial dysbiosis in severe asthma. Allergy 76, 2070–2078. doi: 10.1111/all.14732
Barcik, W., Boutin, R. C. T., Sokolowska, M., Finlay, B. B. (2020). The role of lung and gut microbiota in the pathology of asthma. Immunity 52, 241–255. doi: 10.1016/j.immuni.2020.01.007
Bauermeister, A., Mannochio-Russo, H., Costa-Lotufo, L. V., Jarmusch, A. K., Dorrestein, P. C. (2022). Mass spectrometry-based metabolomics in microbiome investigations. Nat. Rev. Microbiol. 20, 143–160. doi: 10.1038/s41579-021-00621-9
Bonnelykke, K., Coleman, A. T., Evans, M. D., Thorsen, J., Waage, J., Vissing, N. H., et al. (2018). Cadherin-related family member 3 genetics and rhinovirus C respiratory illnesses. Am. J. Respir. Crit. Care Med. 197, 589–594. doi: 10.1164/rccm.201705-1021OC
Bowerman, K. L., Rehman, S. F., Vaughan, A., Lachner, N., Budden, K. F., Kim, R. Y., et al. (2020). Disease-associated gut microbiome and metabolome changes in patients with chronic obstructive pulmonary disease. Nat. Commun. 11, 5886. doi: 10.1038/s41467-020-19701-0
Campbell, C. D., Barnett, C., Sulaiman, I. (2022). A clinicians' review of the respiratory microbiome. Breathe (Sheff) 18, 210161. doi: 10.1183/20734735.0161-2021
Chambers, E. S., Preston, T., Frost, G., Morrison, D. J. (2018). Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr. Nutr. Rep. 7, 198–206. doi: 10.1007/s13668-018-0248-8
Charlson, E. S., Bittinger, K., Haas, A. R., Fitzgerald, A. S., Frank, I., Yadav, A., et al. (2011). Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 184, 957–963. doi: 10.1164/rccm.201104-0655OC
Chu, D. M., Ma, J., Prince, A. L., Antony, K. M., Seferovic, M. D., Aagaard, K. M. (2017). Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 23, 314–326. doi: 10.1038/nm.4272
Collinson, N., Snape, N., Beagley, K., Fantino, E., Spann, K. (2021). COPD is associated with elevated IFN-a production by bronchial epithelial cells infected with RSV or hMPV. Viruses-Basel 13, 911. doi: 10.3390/v13050911
Coverstone, A. M., Wang, L. Y., Sumino, K. (2019). Beyond respiratory syncytial virus and rhinovirus in the pathogenesis and exacerbation of asthma the role of metapneumovirus, bocavirus and influenza virus. Immunol. Allergy Clinics North America 39, 391. doi: 10.1016/j.iac.2019.03.007
D'Alessandro-Gabazza, C. N., Kobayashi, T., Yasuma, T., Toda, M., Kim, H., Fujimoto, H., et al. (2020). A Staphylococcus pro-apoptotic peptide induces acute exacerbation of pulmonary fibrosis. Nat. Commun. 11, 1539. doi: 10.1038/s41467-020-15344-3
Devadoss, D., Long, C., Langley, R. J., Manevski, M., Nair, M., Campos, M. A., et al. (2019). Long noncoding transcriptome in chronic obstructive pulmonary disease. Am. J. Respir. Cell Mol. Biol. 61, 678–688. doi: 10.1165/rcmb.2019-0184TR
Diao, Z., Han, D., Zhang, R., Li, J. (2022). Metagenomics next-generation sequencing tests take the stage in the diagnosis of lower respiratory tract infections. J. Adv. Res. 38, 201–212. doi: 10.1016/j.jare.2021.09.012
Dicker, A. J., Huang, J. T. J., Lonergan, M., Keir, H. R., Fong, C. J., Tan, B., et al. (2021). The sputum microbiome, airway inflammation, and mortality in chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 147, 158–167. doi: 10.1016/j.jaci.2020.02.040
Dong, Q., Chen, E. S., Zhao, C., Jin, C. (2021). Host-microbiome interaction in lung cancer. Front. Immunol. 12. doi: 10.3389/fimmu.2021.679829
El Tekle, G., Garrett, W. S. (2023). Bacteria in cancer initiation, promotion and progression. Nat. Rev. Cancer 23, 600–618. doi: 10.1038/s41568-023-00594-2
Erb Downward, J. R., Falkowski, N. R., Mason, K. L., Muraglia, R., Huffnagle, G. B. (2013). Modulation of post-antibiotic bacterial community reassembly and host response by Candida albicans. Sci. Rep. 3, 2191. doi: 10.1038/srep02191
Gao, J. Y., Yi, X. Z., Wang, Z. (2023). The application of multi-omics in the respiratory microbiome: Progresses, challenges and promises. Comput. Struct. Biotechnol. J. 21, 4933–4943. doi: 10.1016/j.csbj.2023.10.016
Garcia-Valero, J., Olloqueuqi, J., Montes, J. F., Rodríguez, E., Martín-Satué, M., Texids, L., et al. (2019). Deficient pulmonary IFN-o expression in COPD patients. PloS One 14, e0219349. doi: 10.1371/journal.pone.0219349
Goldman, D. L., Chen, Z., Shankar, V., Tyberg, M., Vicencio, A., Burk, R. (2018). Lower airway microbiota and mycobiota in children with severe asthma. J. Allergy Clin. Immunol. 141, 808–811.e807. doi: 10.1016/j.jaci.2017.09.018
Gon, Y., Hashimoto, S. (2018). Role of airway epithelial barrier dysfunction in pathogenesis of asthma. Allergol Int. 67, 12–17. doi: 10.1016/j.alit.2017.08.011
Goto, T. (2022). Microbiota and lung cancer. Semin. Cancer Biol. 86, 1–10. doi: 10.1016/j.semcancer.2022.07.006
Green, A. E., Pottenger, S., Monshi, M. S., Barton, T. E., Phelan, M., Neill, D. R. (2023). Airway metabolic profiling during Streptococcus pneumoniae infection identifies branched chain amino acids as signatures of upper airway colonisation. PloS Pathog. 19, e1011630. doi: 10.1371/journal.ppat.1011630
Guo, H., Zhao, L., Zhu, J., Chen, P., Wang, H., Jiang, M., et al. (2022). Microbes in lung cancer initiation, treatment, and outcome: Boon or bane? Semin. Cancer Biol. 86, 1190–1206. doi: 10.1016/j.semcancer.2021.05.025
Han, D., Zhen, H., Liu, X., Zulewska, J., Yang, Z. (2022). Organelle 16S rRNA amplicon sequencing enables profiling of active gut microbiota in murine model. Appl. Microbiol. Biotechnol. 106, 5715–5728. doi: 10.1007/s00253-022-12083-x
Han, J. R., Zhang, B., Zhang, Y. N., Yin, T. Y., Cui, Y. Y., Liu, J. M., et al. (2023). Gut microbiome: decision-makers in the microenvironment of colorectal cancer. Front. Cell. Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1299977
Hezaveh, K., Shinde, R. S., Klötgen, A., Halaby, M. J., Lamorte, S., Ciudad, M. T., et al. (2022). Tryptophan-derived microbial metabolites activate the aryl hydrocarbon receptor in tumor-associated macrophages to suppress anti-tumor immunity. Immunity 55, 324–340.e328. doi: 10.1016/j.immuni.2022.01.006
Higham, A., Mathioudakis, A., Vestbo, J., Singh, D. (2020). COVID-19 and COPD: a narrative review of the basic science and clinical outcomes. Eur. Respir. Rev. 29, 200199. doi: 10.1183/16000617.0199-2020
Ho, L. J., Yang, H. Y., Chung, C. H., Chang, W. C., Yang, S. S., Sun, C. A., et al. (2021). Increased risk of secondary lung cancer in patients with tuberculosis: A nationwide, population-based cohort study. PloS One 16, e0250531. doi: 10.1371/journal.pone.0250531
Hou, K., Wu, Z. X., Chen, X. Y., Wang, J. Q., Zhang, D., Xiao, C., et al. (2022). Microbiota in health and diseases. Signal Transduct Target Ther. 7, 135. doi: 10.1038/s41392-022-00974-4
Ibañez-Lligoña, M., Colomer-Castell, S., González-Sánchez, A., Gregori, J., Campos, C., Garcia-Cehic, D., et al. (2023). Bioinformatic tools for NGS-based metagenomics to improve the clinical diagnosis of emerging, re-emerging and new viruses. Viruses 15, 587–602. doi: 10.3390/v15020587
Isaacs, B., Chung, M., Wu, B. G., Tsay, J., Barnett, C. R., Kwok, B., et al. (2023). Profiling the functional microbiome in mild COPD. Am. J. Respir. Crit. Care Med. 207, A6139–A6139. doi: 10.1164/ajrccm-conference.2023.C108
Jackson, D. J., Gern, J. E. (2022). Rhinovirus infections and their roles in asthma: etiology and exacerbations. J. Allergy Clin. Immunology-in Pract. 10, 673–681. doi: 10.1016/j.jaip.2022.01.006
Jang, Y. O., Lee, S. H., Choi, J. J., Kim, D. H., Choi, J. M., Kang, M. J., et al. (2020). Fecal microbial transplantation and a high fiber diet attenuates emphysema development by suppressing inflammation and apoptosis. Exp. Mol. Med. 52, 1128–1139. doi: 10.1038/s12276-020-0469-y
Kim, S. J., Kim, T., Choi, H., Shin, T. R., Kim, H. I., Jang, S. H., et al. (2024). Respiratory pathogen and clinical features of hospitalized patients in acute exacerbation of chronic obstructive pulmonary disease after COVID 19 pandemic. Sci. Rep. 14, 10462. doi: 10.1038/s41598-024-61360-4
Kovaleva, O. V., Romashin, D., Zborovskaya, I. B., Davydov, M. M., Shogenov, M. S., Gratchev, A. (2019). Human lung microbiome on the way to cancer. J. Immunol. Res. 2019, 1394191. doi: 10.1155/2019/1394191
Krell, T., Gavira, J. A., Velando, F., Fernández, M., Roca, A., Monteagudo-Cascales, E., et al. (2021). Histamine: A bacterial signal molecule. Int. J. Mol. Sci. 22, 6312–6329. doi: 10.3390/ijms22126312
Kumpitsch, C., Koskinen, K., Schöpf, V., Moissl-Eichinger, C. (2019). The microbiome of the upper respiratory tract in health and disease. BMC Biol. 17, 87. doi: 10.1186/s12915-019-0703-z
Labaki, W. W., Rosenberg, S. R. (2020). Chronic obstructive pulmonary disease. Ann. Intern. Med. 173, Itc17–itc32. doi: 10.7326/aitc202008040
Leiter, A., Veluswamy, R. R., Wisnivesky, J. P. (2023). The global burden of lung cancer: current status and future trends. Nat. Rev. Clin. Oncol. 20, 624–639. doi: 10.1038/s41571-023-00798-3
Leung, J. M., Tiew, P. Y., Mac Aogáin, M., Budden, K. F., Yong, V. F., Thomas, S. S., et al. (2017). The role of acute and chronic respiratory colonization and infections in the pathogenesis of COPD. Respirology 22, 634–650. doi: 10.1111/resp.13032
Li, B., Xu, L., Guo, Q., Chen, J., Zhang, Y., Huang, W., et al. (2021). GenSeizer: a multiplex PCR-based targeted gene sequencing platform for rapid and accurate identification of major mycobacterium species. J. Clin. Microbiol. 59. doi: 10.1128/jcm.00584-20
Li, N., Dai, Z., Wang, Z., Deng, Z., Zhang, J., Pu, J., et al. (2021). Gut microbiota dysbiosis contributes to the development of chronic obstructive pulmonary disease. Respir. Res. 22, 274–289. doi: 10.1186/s12931-021-01872-z
Li, S., Han, X., Ma, J., Huang, G. H., Yang, S. T., Wang, C. M. (2024). Study on mNGS Technique in Diagnosing Pneumocystis jirovecii Pneumonia in Non-HIV-Infected Patients. Infect. Drug Resist. 17, 1397–1405. doi: 10.2147/Idr.S450878
Li, X., Saxena, D. (2022). The tumor mycobiome: A paradigm shift in cancer pathogenesis. Cell 185, 3648–3651. doi: 10.1016/j.cell.2022.09.013
Lin, L. F., Yi, X. Z., Liu, H. Y., Meng, R. L., Li, S. Q., Liu, X. M., et al. (2023). The airway microbiome mediates the interaction between environmental exposure and respiratory health in humans. Nat. Med. 29, 1750. doi: 10.1038/s41591-023-02424-2
Liu, N. N., Yi, C. X., Wei, L. Q., Zhou, J. A., Jiang, T., Hu, C. C., et al. (2023). The intratumor mycobiome promotes lung cancer progression via myeloid-derived suppressor cells. Cancer Cell 41, 1927. doi: 10.1016/j.ccell.2023.08.012
Liu, Y., Zhang, X., Zhang, L., Oliver, B. G., Wang, H. G., Liu, Z. P., et al. (2022a). Sputum metabolomic profiling reveals metabolic pathways and signatures associated with inflammatory phenotypes in patients with asthma. Allergy Asthma Immunol. Res. 14, 393–411. doi: 10.4168/aair.2022.14.4.393
Liu, Y., Zhao, N., Xu, Q., Deng, F., Wang, P., Dong, L., et al. (2022b). MBL binding with ahR controls th17 immunity in silicosis-associated lung inflammation and fibrosis. J. Inflamm. Res. 15, 4315–4329. doi: 10.2147/jir.S357453
Liu, Y. T., Zhang, H., Lu, W. T., Jiang, T. (2023). Integrating metabolomics, 16S rRNA sequencing, network pharmacology, and metorigin to explore the mechanism of Cinnamomi Cortex in treating chronic atrophic gastritis rats. Phytomedicine 121, 155084. doi: 10.1016/j.phymed.2023.155084
Liu, Y. X., Qin, Y., Chen, T., Lu, M., Qian, X., Guo, X., et al. (2021). A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 12, 315–330. doi: 10.1007/s13238-020-00724-8
Ma, M., Li, G., Qi, M., Jiang, W., Zhou, R. (2021). Inhibition of the inflammasome activity of NLRP3 attenuates HDM-induced allergic asthma. Front. Immunol. 12. doi: 10.3389/fimmu.2021.718779
Maddi, A., Sabharwal, A., Violante, T., Manuballa, S., Genco, R., Patnaik, S., et al. (2019). The microbiome and lung cancer. J. Thorac. Dis. 11, 280–291. doi: 10.21037/jtd.2018.12.88
Mao, Q., Jiang, F., Yin, R., Wang, J., Xia, W., Dong, G., et al. (2018). Interplay between the lung microbiome and lung cancer. Cancer Lett. 415, 40–48. doi: 10.1016/j.canlet.2017.11.036
McCauley, K., Durack, J., Valladares, R., Fadrosh, D. W., Lin, D. L., Calatroni, A., et al. (2019). Distinct nasal airway bacterial microbiotas differentially relate to exacerbation in pediatric patients with asthma. J. Allergy Clin. Immunol. 144, 1187–1197. doi: 10.1016/j.jaci.2019.05.035
Meldrum, O. W., Donaldson, G. C., Narayana, J. K., Xaverius Ivan, F., Jaggi, T. K., Mac Aogáin, M., et al. (2024). Accelerated lung function decline and mucus-microbe evolution in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 210 (3), 298–310. doi: 10.1164/rccm.202306-1060OC
Meng, Y., Mao, Y., Tang, Z., Qiu, X., Bajinka, O., Tan, Y., et al. (2023). Crosstalk between the lung microbiome and lung cancer. Microb. Pathog. 178, 106062. doi: 10.1016/j.micpath.2023.106062
Mika, M., Mack, I., Korten, I., Qi, W., Aebi, S., Frey, U., et al. (2015). Dynamics of the nasal microbiota in infancy: a prospective cohort study. J. Allergy Clin. Immunol. 135, 905–912.e911. doi: 10.1016/j.jaci.2014.12.1909
Minot, S., Sinha, R., Chen, J., Li, H., Keilbaugh, S. A., Wu, G. D., et al. (2011). The human gut virome: inter-individual variation and dynamic response to diet. Genome Res. 21, 1616–1625. doi: 10.1101/gr.122705.111
Misra, B. B. (2020). The connection and disconnection between microbiome and metabolome: A critical appraisal in clinical research. Biol. Res. Nurs. 22, 561–576. doi: 10.1177/1099800420903083
Moss, B. J., Ryter, S. W., Rosas, I. O. (2022). Pathogenic mechanisms underlying idiopathic pulmonary fibrosis. Annu. Rev. Pathol. 17, 515–546. doi: 10.1146/annurev-pathol-042320-030240
Natalini, J. G., Singh, S., Segal, L. N. (2023a). The dynamic lung microbiome in health and disease. Nat. Rev. Microbiol. 21, 222–235. doi: 10.1038/s41579-022-00821-x
Nejman, D., Livyatan, I., Fuks, G., Gavert, N., Zwang, Y., Geller, L. T., et al. (2020). The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 368, 973–980. doi: 10.1126/science.aay9189
O'Dwyer, D. N., Ashley, S. L., Gurczynski, S. J., Xia, M., Wilke, C., Falkowski, N. R., et al. (2019). Lung microbiota contribute to pulmonary inflammation and disease progression in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 199, 1127–1138. doi: 10.1164/rccm.201809-1650OC
Opron, K., Begley, L. A., Erb-Downward, J. R., Freeman, C., Madapoosi, S., Alexis, N. E., et al. (2021). Lung microbiota associations with clinical features of COPD in the SPIROMICS cohort. NPJ Biofilms Microbiomes 7, 14. doi: 10.1038/s41522-021-00185-9
Özçam, M., Lynch, S. V. (2024). The gut-airway microbiome axis in health and respiratory diseases. Nat. Rev. Microbiol. 22 (8), 492–506. doi: 10.1038/s41579-024-01048-8
Paats, M. S., Bergen, I. M., Hoogsteden, H. C., van der Eerden, M. M., Hendriks, R. W. (2012). Systemic CD4+ and CD8+ T-cell cytokine profiles correlate with GOLD stage in stable COPD. Eur. Respir. J. 40, 330–337. doi: 10.1183/09031936.00079611
Pathak, U., Gupta, N. C., Suri, J. C. (2020). Risk of COPD due to indoor air pollution from biomass cooking fuel: a systematic review and meta-analysis. Int. J. Environ. Health Res. 30, 75–88. doi: 10.1080/09603123.2019.1575951
Poore, G. D., Kopylova, E., Zhu, Q., Carpenter, C., Fraraccio, S., Wandro, S., et al. (2020). Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature 579, 567–574. doi: 10.1038/s41586-020-2095-1
Qi, J., Li, M., Wang, L., Hu, Y., Liu, W., Long, Z., et al. (2023). National and subnational trends in cancer burden in Chin–20: an analysis of national mortality surveillance data. Lancet Public Health 8, e943–e955. doi: 10.1016/s2468-2667(23)00211-6
Qin, Z., Zou, Y., Huang, Z., Yu, N., Deng, Z., Chen, Z., et al. (2022). Metagenomic next-generation sequencing contributes to the diagnosis of mixed pulmonary infection: a case report. Ann. Clin. Microbiol. Antimicrob. 21, 52. doi: 10.1186/s12941-022-00545-z
Ren, L., Zhang, R., Rao, J., Xiao, Y., Zhang, Z., Yang, B., et al. (2019). Transcriptionally active lung microbiome and its association with bacterial biomass and host inflammatory status. Eur. J. Immunol. 49, 1329–1330. doi: 10.1128/mSystems.00199-18
Santos, A., Pitto, H., Chaves-Loureiro, C., Rocha, S. M., Taborda-Barata, L. (2021). Metabolic phenotypes in asthmatic adults: relationship with inflammatory and clinical phenotypes and prognostic implications. Metabolites 11, 534–564. doi: 10.3390/metabo11080534
Sheng, G. H., Chen, P., Wei, Y. Q., Yue, H. H., Chu, J. J., Zhao, J. P., et al. (2020). Viral infection increases the risk of idiopathic pulmonary fibrosis A meta-analysis. Chest 157, 1175–1187. doi: 10.1016/j.chest.2019.10.032
Sibandze, D. B., Kay, A., Dreyer, V., Sikhondze, W., Dlamini, Q., DiNardo, A., et al. (2022). Rapid molecular diagnostics of tuberculosis resistance by targeted stool sequencing. Genome Med. 14, 52. doi: 10.1186/s13073-022-01054-6
Sidletskaya, K., Vitkina, T., Denisenko, Y. (2020). The role of toll-like receptors 2 and 4 in the pathogenesis of chronic obstructive pulmonary disease. Int. J. Chron. Obstruct. Pulmon. Dis. 15, 1481–1493. doi: 10.2147/Copd.S249131
Simon, S., Joean, O., Welte, T., Rademacher, J. (2023). The role of vaccination in COPD: influenza, SARS-CoV-2, pneumococcus, pertussis, RSV and varicella zoster virus. Eur. Respir. Rev. 32, 230034–230050. doi: 10.1183/16000617.0034-2023
Singh, R., Belchamber, K. B. R., Fenwick, P. S., Chana, K., Donaldson, G., Wedzicha, J. A., et al. (2021). Defective monocyte-derived macrophage phagocytosis is associated with exacerbation frequency in COPD. Respir. Res. 22, 113. doi: 10.1186/s12931-021-01718-8
Spijkerman, J., Prevaes, S. M., van Gils, E. J., Veenhoven, R. H., Bruin, J. P., Bogaert, D., et al. (2012). Long-term effects of pneumococcal conjugate vaccine on nasopharyngeal carriage of S. pneumoniae, S. aureus, H. influenzae and M. catarrhalis. PloS One 7, e39730. doi: 10.1371/journal.pone.0039730
Stolz, D., Papakonstantinou, E., Grize, L., Schilter, D., Strobel, W., Louis, R., et al. (2019). Time-course of upper respiratory tract viral infection and COPD exacerbation. Eur. Respir. J. 54, 1900407–1900423. doi: 10.1183/13993003.00407-2019
Sulaiman, I., Wu, B. G., Li, Y., Tsay, J. C., Sauthoff, M., Scott, A. S., et al. (2021). Functional lower airways genomic profiling of the microbiome to capture active microbial metabolism. Eur. Respir. J. 58, 2003434–2003457. doi: 10.1183/13993003.03434-2020
Sun, Z., Huang, S., Zhu, P., Tzehau, L., Zhao, H., Lv, J., et al. (2022). Species-resolved sequencing of low-biomass or degraded microbiomes using 2bRAD-M. Genome Biol. 23, 36. doi: 10.1186/s13059-021-02576-9
Sun, Z., Zhu, Q. L., Shen, Y., Yan, T., Zhou, X. (2020). Dynamic changes of gut and lung microorganisms during chronic obstructive pulmonary disease exacerbations. Kaohsiung J. Med. Sci. 36, 107–113. doi: 10.1002/kjm2.12147
Tiew, P. Y., Dicker, A. J., Keir, H. R., Poh, M. E., Pang, S. L., Mac Aogáin, M., et al. (2021). A high-risk airway mycobiome is associated with frequent exacerbation and mortality in COPD. Eur. Respir. J. 57, 2002050. doi: 10.1183/13993003.02050-2020
Tiew, P. Y., Narayana, J. K., Quek, M. S. L., Ang, Y. Y., Ko, F. W. S., Poh, M. E., et al. (2023). Sensitisation to recombinant Aspergillus fumigatus allergens and clinical outcomes in COPD. Eur. Respir. J. 61, 2200507–2200521. doi: 10.1183/13993003.00507-2022
Tiew, P. Y., San Ko, F. W., Pang, S. L., Matta, S. A., Sio, Y. Y., Poh, M. E., et al. (2020). Environmental fungal sensitisation associates with poorer clinical outcomes in COPD. Eur. Respir. J. 56, 2000418. doi: 10.1183/13993003.00418-2020
Tiotiu, A. I., Novakova, P., Nedeva, D., Chong-Neto, H. J., Novakova, S., Steiropoulos, P., et al. (2020). Impact of air pollution on asthma outcomes. Int. J. Environ. Res. Public Health 17, 6212–6241. doi: 10.3390/ijerph17176212
Traverso, A., Wee, L., Dekker, A., Gillies, R. (2018). Repeatability and reproducibility of radiomic features: A systematic review. Int. J. Radiat. Oncol. Biol. Phys. 102, 1143–1158. doi: 10.1016/j.ijrobp.2018.05.053
Tsay, J. J., Wu, B. G., Sulaiman, I., Gershner, K., Schluger, R., Li, Y., et al. (2021). Lower airway dysbiosis affects lung cancer progression. Cancer Discov. 11, 293–307. doi: 10.1158/2159-8290.Cd-20-0263
Tsay, J. C. J., Wu, B. G., Badri, M. H., Clemente, J. C., Shen, N., Meyn, P., et al. (2018). Airway microbiota is associated with upregulation of the PI3K pathway in lung cancer. Am. J. Respir. Crit. Care Med. 198, 1188–1198. doi: 10.1164/rccm.201710-2118OC
Valentino, G., Graziani, V., D'Abrosca, B., Pacifico, S., Fiorentino, A., Scognamiglio, M. (2020). NMR-based plant metabolomics in nutraceutical research: an overview. Molecules 25, 1444–1466. doi: 10.3390/molecules25061444
Waller, K. M. J., Leong, R. W., Paramsothy, S. (2022). An update on fecal microbiota transplantation for the treatment of gastrointestinal diseases. J. Gastroenterol. Hepatol. 37, 246–255. doi: 10.1111/jgh.15731
Wang, C., Jiang, S., Zhang, S., Ouyang, Z., Wang, G., Wang, F. (2021). Research progress of metabolomics in asthma. Metabolites 11, 567–587. doi: 10.3390/metabo11090567
Wang, S., Zhao, C., Yin, Y., Chen, F., Chen, H., Wang, H. (2022). A practical approach for predicting antimicrobial phenotype resistance in staphylococcus aureus through machine learning analysis of genome data. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.841289
Wang, S. M., Zhou, Q. X., Tian, Y. Z., Hu, X. A. (2022). The lung microbiota affects pulmonary inflammation and oxidative stress induced by PM exposure. Environ. Sci. Technol. 56, 12368–12379. doi: 10.1021/acs.est.1c08888
Wang, Z., Bafadhel, M., Haldar, K., Spivak, A., Mayhew, D., Miller, B. E., et al. (2016). Lung microbiome dynamics in COPD exacerbations. Eur. Respir. J. 47, 1082–1092. doi: 10.1183/13993003.01406-2015
Wang, Z., Locantore, N., Haldar, K., Ramsheh, M. Y., Beech, A. S., Ma, W., et al. (2021). Inflammatory endotype-associated airway microbiome in chronic obstructive pulmonary disease clinical stability and exacerbations: A multicohort longitudinal analysis. Am. J. Respir. Crit. Care Med. 203, 1488–1502. doi: 10.1164/rccm.202009-3448OC
Weinroth, M. D., Belk, A. D., Dean, C., Noyes, N., Dittoe, D. K., Rothrock, M. J., et al. (2022). Considerations and best practices in animal science 16S ribosomal RNA gene sequencing microbiome studies. J. Anim. Sci. 100, 1–18. doi: 10.1093/jas/skab346
Weisskopf, L., Schulz, S., Garbeva, P. (2021). Microbial volatile organic compounds in intra-kingdom and inter-kingdom interactions. Nat. Rev. Microbiol. 19, 391–404. doi: 10.1038/s41579-020-00508-1
Wensel, C. R., Pluznick, J. L., Salzberg, S. L., Sears, C. L. (2022). Next-generation sequencing: insights to advance clinical investigations of the microbiome. J. Clin. Invest. 132, 154944–154964. doi: 10.1172/jci154944
Whetstone, C. E., Ranjbar, M., Omer, H., Cusack, R. P., Gauvreau, G. M. (2022). The role of airway epithelial cell alarmins in asthma. Cells 11, 1105–1131. doi: 10.3390/cells11071105
Wilburn, A. N., McAlees, J. W., Haslam, D. B., Graspeuntner, S., Schmudde, I., Laumonnier, Y., et al. (2023). Delayed microbial maturation durably exacerbates th17-driven asthma in mice. Am. J. Respir. Cell Mol. Biol. 68, 498–510. doi: 10.1165/rcmb.2022-0367OC
Xie, Y., Xie, F., Zhou, X., Zhang, L., Yang, B., Huang, J., et al. (2022). Microbiota in tumors: from understanding to application. Adv. Sci. (Weinh) 9, e2200470. doi: 10.1002/advs.202200470
Xue, C., Chu, Q., Zheng, Q., Yuan, X., Su, Y., Bao, Z., et al. (2023). Current understanding of the intratumoral microbiome in various tumors. Cell Rep. Med. 4, 100884. doi: 10.1016/j.xcrm.2022.100884
Yamasaki, K., van Eeden, S. F. (2018). Lung macrophage phenotypes and functional responses: role in the pathogenesis of COPD. Int. J. Mol. Sci. 19, 582. doi: 10.3390/ijms19020582
Yamauchi, K., Ogasawara, M. (2019). The role of histamine in the pathophysiology of asthma and the clinical efficacy of antihistamines in asthma therapy. Int. J. Mol. Sci. 20, 1733–1749. doi: 10.3390/ijms20071733
Yan, Z., Chen, B., Yang, Y., Yi, X., Wei, M., Ecklu-Mensah, G., et al. (2022). Multi-omics analyses of airway host-microbe interactions in chronic obstructive pulmonary disease identify potential therapeutic interventions. Nat. Microbiol. 7, 1361–1375. doi: 10.1038/s41564-022-01196-8
Yang, D., Chen, X., Wang, J., Lou, Q., Lou, Y., Li, L., et al. (2019). Dysregulated lung commensal bacteria drive interleukin-17B production to promote pulmonary fibrosis through their outer membrane vesicles. Immunity 50, 692–706.e697. doi: 10.1016/j.immuni.2019.02.001
Yang, X., Li, H., Ma, Q., Zhang, Q., Wang, C. (2018). Neutrophilic asthma is associated with increased airway bacterial burden and disordered community composition. BioMed. Res. Int. 2018, 9230234. doi: 10.1155/2018/9230234
Ye, D., Li, X., Shen, J., Xia, X. (2022). Microbial metabolomics: From novel technologies to diversified applications. TrAC Trends Anal. Chem. 148, 116540. doi: 10.1016/j.trac.2022.116540
Yi, X., Lu, H., Liu, X., He, J., Li, B., Wang, Z., et al. (2024). Unravelling the enigma of the human microbiome: Evolution and selection of sequencing technologies. Microb. Biotechnol. 17, e14364. doi: 10.1111/1751-7915.14364
Yi, X. Z., Gao, J. Y., Wang, Z. (2022). The human lung microbiome-A hidden link between microbes and human health and diseases. Imeta 1, e33. doi: 10.1002/imt2.33
Zhang, T., Zhang, M., Yang, L., Gao, L., Sun, W. (2023). Potential targeted therapy based on deep insight into the relationship between the pulmonary microbiota and immune regulation in lung fibrosis. Front. Immunol. 14. doi: 10.3389/fimmu.2023.1032355
Zhang, W., Fan, X., Shi, H., Li, J., Zhang, M., Zhao, J., et al. (2023). Comprehensive Assessment of 16S rRNA Gene Amplicon Sequencing for Microbiome Profiling across Multiple Habitats. Microbiol. Spectr. 11, e0056323. doi: 10.1128/spectrum.00563-23
Zheng, W., Zhao, S., Yin, Y., Zhang, H., Needham, D. M., Evans, E. D., et al. (2022). High-throughput, single-microbe genomics with strain resolution, applied to a human gut microbiome. Science 376, eabm1483. doi: 10.1126/science.abm1483
Zhou, C. B., Zhou, Y. L., Fang, J. Y. (2021). Gut microbiota in cancer immune response and immunotherapy. Trends Cancer 7, 647–660. doi: 10.1016/j.trecan.2021.01.010
Keywords: lung microbiota, chronic pulmonary diseases, microbial metabolomics, chronic obstructive pulmonary disease (COPD), microbial genomics
Citation: Zhou J, Hou W, Zhong H and Liu D (2024) Lung microbiota: implications and interactions in chronic pulmonary diseases. Front. Cell. Infect. Microbiol. 14:1401448. doi: 10.3389/fcimb.2024.1401448
Received: 06 May 2024; Accepted: 31 July 2024;
Published: 19 August 2024.
Edited by:
Zifeng Yang, Guangzhou Institute of Respiratory Health, ChinaReviewed by:
Zhang Wang, South China Normal University, ChinaCopyright © 2024 Zhou, Hou, Zhong and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dan Liu, bGl1ZGFuMTA5NjVAd2Noc2N1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.