
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Anim. Sci. , 28 January 2025
Sec. Product Quality
Volume 6 - 2025 | https://doi.org/10.3389/fanim.2025.1527127
 María Sofía Castaño Ledesma1†
María Sofía Castaño Ledesma1† Gustavo Adolfo Palma1,2†
Gustavo Adolfo Palma1,2† Claudio Darío Borsarelli1,2*†
Claudio Darío Borsarelli1,2*† María Sumampa Coria1,2*†
María Sumampa Coria1,2*†The impact of corn silage supplementation and post mortem aging on the antioxidant status in longissimus thoracis et lumborum (LTL) muscle was assessed using micro-spectroscopic techniques. A total of 30 Braford steers were utilized in the study. Of these, 15 animals were supplemented with corn silage for 120 days, while the remaining animals were fed exclusively on pasture. LTL steak samples were aged for 2, 7, 14, and 21 days at a temperature of 2 ± 1°C, under conditions of darkness and vacuum. Several colorimetric assays were employed to evaluate the antioxidant capacity of both enzymatic and non-enzymatic origin, as well as the levels of protein and lipid oxidation. The content of heme pigment was determined by a spectrophotometric assay, while the fluorescence fingerprint of beef was evaluated by fluorescence spectroscopy excitation-emission matrices (FEEM). Additionally, the muscle microstructure was analyzed via scanning electron microscopy (SEM). The results demonstrated that the finishing system did not exert any discernible influence on the rates of lipid oxidation, antioxidant capacity, or muscle microstructure. However, the carbonyl content and total myoglobin exhibited higher levels in the corn silage-supplemented animals. Conversely, during aging period, data revealed that lipid degradation proceeded at a faster rate than protein oxidation, accompanied by a notable enhancement in antioxidant capacity within the hydrophilic meat extracts. In contrast, the lipophilic extracts demonstrated a reduction in both enzymatic and non-enzymatic antioxidant capabilities with the progression of post mortem aging. The aforementioned alterations were accompanied by a reduction in the muscle microstructure during the aging process. The results demonstrate that finishing steers with or without corn silage over 120 days results in comparable and satisfactory beef quality after 21 days of vacuum aging. It can be concluded that 21 days of storage compensates the antioxidant status obtained by feeding strategies in fresh meat. Furthermore, SEM and FEEM techniques allowed for a precise evaluation of the microstructure and oxidative status, suggesting that these methods could be employed in future assays.
The oxidative stability of post-mortem muscle is dependent on the composition and concentration of substrates, as well as the balance of pro- and antioxidants, which is influenced by a number of endogenous and exogenous factors. This encompasses a range of catalysts, including iron, oxygen, light, temperature, and microorganisms, as well as antioxidants present in muscle tissue, including vitamin E, provitamin A, vitamin C, glutathione, and antioxidant enzymes (Huff Lonergan et al., 2010; Bekhit et al., 2013; Domínguez et al., 2019, 2022). In vivo, cellular defense mechanisms against oxidative stress include enzymatic scavenging of reactive species, sequestration of prooxidant metals by nutrients or transport and storage proteins, attenuation of cellular damage by expression of heat shock proteins, and consumption of free radicals by antioxidant molecules acting as a proton or electron donors (Nimse and Pal, 2015). Conversely, during the post-mortem conversion of muscle into meat, circumstances may be established that permit a shift in the equilibrium between antioxidant and pro-oxidant capabilities, thereby increasing the likelihood of oxidative damage (Huff Lonergan et al., 2010; Bekhit et al., 2013; Sabow et al., 2016).
It is important to highlight that lipids and proteins in meat are the primary targets of oxidative stress processes, which result in irreversible chemical modifications that reduce the nutritional value, shelf life, and sensory qualities of the meat. These modifications make meat undesirable for human consumption (Haak et al., 2008; Lund et al., 2008; Huff Lonergan et al., 2010; Bekhit et al., 2013; Domínguez et al., 2019, 2022). It is therefore of the utmost importance to mitigate oxidative deterioration in meat to maintain its quality and safety for consumption. Indeed, there is substantial evidence demonstrating that the implementation of an appropriate animal feeding strategy can enhance meat quality (Ripoll et al., 2014; Adeyemi et al., 2016a; Terevinto et al., 2019; Coria et al., 2020, 2022), as well as, that storage with vacuum packaging can prevent the modification of lipids, proteins, and the oxidative stability in beef (Vincenti et al., 2009; Beriain et al., 2009; Adeyemi et al., 2016b).
Argentine meat has a longstanding tradition of production on natural and cultivated pastures. However, feedlot systems are becoming more prevalent among producers, particularly when low-cost corn is available (Descalzo and Sancho, 2008). The composition of antioxidants, pro-oxidants, and fatty acids differs between meat produced on pasture or grain (Pouzo et al., 2016). While carcasses from grain-finished animals are generally heavier and present better conformation scores (Rosa et al., 2014), cattle fed on corn finishing diets tend to increase the proportion of omega-6 polyunsaturated fatty acids (PUFA), such as linoleic acid (Descalzo et al., 2007). Indeed, diets based on corn can result in increased energy intake, which in turn promotes elevated metabolic activity and ROS production, thereby contributing to oxidative damage (Ponnampalam et al., 2022). It is important to note that lipids containing PUFA are particularly susceptible to attack by free radicals, which can subsequently result in the degradation of lipids and proteins (Domínguez et al., 2019, 2022; Terevinto et al., 2019). Conversely, animals that have been pasture-finished produce meat with a higher content of omega-3 PUFA and conjugated linoleic acid. Additionally, these animals are rich in carotenoids, vitamin E, and polyphenols, which are natural antioxidants that reduce oxidative stress in animal tissues and enhance the nutritional quality of the meat (Descalzo and Sancho, 2008).
The quality and shelf life of meat are typically evaluated subjectively through analysis or tests that are laborious, time-consuming, destructive, and do not provide an immediate response (Wu et al., 2022). In recent decades, the advancement of non-destructive analytical techniques, including spectroscopy and microscopy, has facilitated the rapid, precise, and sensitive analysis of food matrices (Nawrocka and Lamorsk, 2013; Qin et al., 2017). To illustrate, fluorescence spectroscopy (FS) has been employed in meat analysis to predict surface spoilage (Yoshimura et al., 2014), assess freshness, and differentiate fat, connective tissue, and myofibers (Liu et al., 2019). Scanning electron microscopy (SEM) combined with energy dispersive X-ray spectroscopy (EDX) was employed to characterize the microstructure of bovine muscle (Palka, 2003; Coria et al., 2023). In a previous study, Coria et al. (2020) evaluated the effect of corn silage supplementation on some meat quality parameters. Their findings demonstrated that beef derived from steers reared exclusively on pasture exhibited reduced hardness, as assessed by Warner-Bratzler shear force (WBSF), and a lower intramuscular fat content, than meat derived from corn silage-supplemented Braford steers. Furthermore, in both finishing systems, the aging time resulted in a notable increase in pH and lightness values after 7 and 21 days, respectively, accompanied by a linear decline in WBSF values (Coria et al., 2022).
In light of the aforementioned findings, the present study extends the research of Coria et al. (2020, 2022) by examining the impact of pasture supplementation with corn silage (PS) on the post-mortem pro- and antioxidant status of the longissimus thoracis et lumborum (LTL) muscle in Bradford steers. This is compared to the effects of natural pasture (P) on the same muscle during aging under vacuum-packed storage conditions, using micro-spectroscopic techniques.
This study was conducted with animals from a commercial breeding herd in Santiago del Estero, northwestern Argentina (27°17′34.3″S - 62°15′14.1″W). Animal handling and experimental procedures followed the Animal Welfare Procedures Manual of the Argentine National Animal Health Service (SENASA, 2016). The meat samples used in the present study were obtained from animals given diets as described by Coria et al. (2022). Briefly, 30 two-year-old Braford steers were randomly divided into two experimental groups of 15 animals each and fed as follows for 120 days before slaughter: one group was fed ad libitum pasture (P) and the second group was fed ad libitum pasture-supplemented with corn silage (PS) (Table 1). Both groups had ad libitum access to water and were weighed after the trial. The mean weights for the P and PS groups were 455 ± 16 Kg and 473 ± 18 Kg, respectively (p = 0.05) (Coria et al., 2020). After assignment to dietary strategies, animals were randomly assigned to three paddocks of 5 animals each and slaughtered at an average age of 26 months. Following slaughter, the carcasses were weighed, with the result expressed as Hot Carcass Weight (HCW). The HCW for the control group was 264 ± 15 kg, and for the supplemented group, it was 298 ± 17 kg (p < 0.05) (Coria et al., 2020). The degree of fattening was determined following the Argentine Bovine Carcass Typification System (Junta Nacional de Carne, Resolucion J-378/73 de la SAGPSyA). Based on a visual assessment of the quantity and distribution of subcutaneous fat, the fattening could assume values between 1 and 4. Higher values indicate a greater thickness of this deposit. The control group carcass exhibited fattening degree values of 1 ± 0.0, while the supplemented group showed values of 1.7 ± 0.5 (p <0.05) (Coria et al., 2020). Following a 48-hour post-slaughter cooling period between 1°C and 5°C, the block of LTL muscle between the 9th and 13th rib was removed from each left half carcass. Four subsamples of approximately 10 cm and 300 g were taken with an electric saw (Coria et al., 2022).
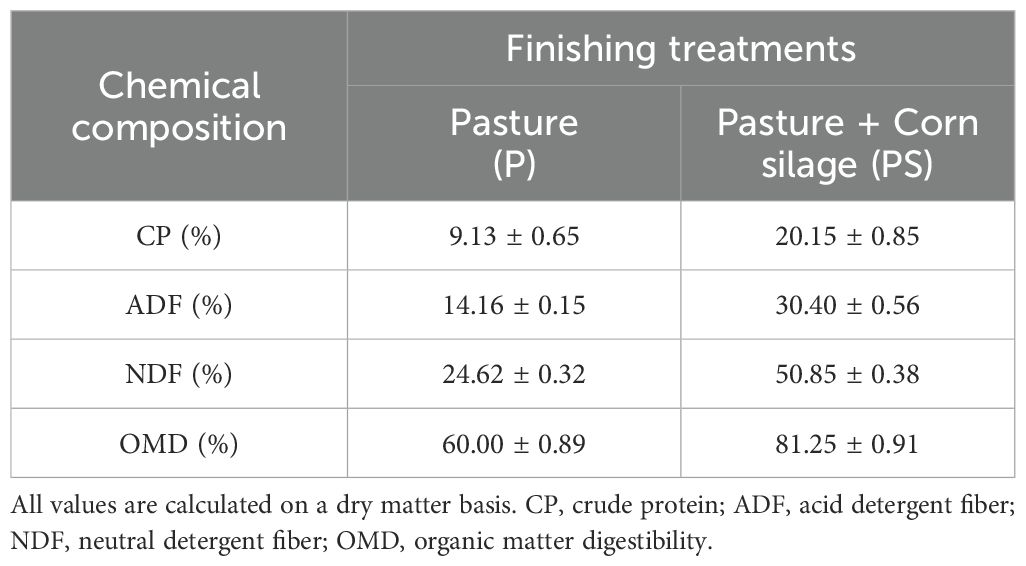
Table 1. Mean and standard deviation values for the chemical composition of finishing diets fed to Braford steers over 120 days.
Samples were vacuum-packed (Dise SA., thickness 90 μm, oxygen permeability 50 cm3 m-2 day-1, CO2 permeability 140 cm3 m-2 day-1, water vapor permeability 6 to 8 g m-2 day-1) and aged in the dark at 2 ± 1°C for 2, 7, 14, or 21 days and then frozen at -80°C until analysis. Before thawing, 50 grams of each subsample was obtained, processed with a grinder, divided into 10 tubes for the different tests, and stored at -18°C until further analysis. The intramuscular fat content was previously determined in duplicate according to the Soxhlet AOAC method (Coria et al., 2022), by continuous distillation with hexane from 5 g of meat. The results were expressed as a percentage of fat (grams of fat per 100 grams of meat), yielding values of 2.22 ± 0.14 for the P group and 3.47 ± 1.24 for the PS group (p < 0.05). The analysis was conducted before the meat reached six months of storage.
The Ferrous Oxidation-Xylenol orange (FOX) assay described by Grau et al. (2000) was used to measure lipid hydroperoxides (LHP) in subsamples of raw meat aged 2, 7, 14, and 21 days. Absorbance was measured at 592 nm after 30 min using a NanoDrop 2000c UV-Vis spectrophotometer (Thermo Scientific, USA). The LHP content was expressed as µmoles of cumene hydroperoxide (CHP) equivalent per gram of sample and was determined using a calibration curve with authentic CHP as the standard.
2-Thiobarbituric acid reactive substances (TBARs) assay was performed according to the method of Duan et al. (2010) with some modifications. Briefly, 1 g of subsamples of raw meat aged 2, 7, 14, and 21 days, was homogenized with 2 mL of 10% (w/v) trichloroacetic acid and centrifuged at 4,000 ×g for 30 min at 4°C. Then, 2 mL of 10 mM thiobarbituric acid (TBA) was added and the solution was heated in a boiling water bath for 20 min. The absorbance of the mixture was measured at 532 nm using the NanoDrop 2000c UV-Vis spectrophotometer. The assay was calibrated with a solution of known concentration of 1,1,3,3-tetramethoxypropane (TMP), an analog of malondialdehyde (MDA), and TBARS values were expressed as mg MDA per Kg of the sample.
Protein oxidation was measured in raw meat aged 2, 7, 14, and 21 days by the formation of the side chain carbonyl group (C=O) as determined by derivatization with 2,4-dinitrophenylhydrazine (DNPH) according to Oliver et al. (1987). The protein concentration was calculated at 280 nm using bovine serum albumin (BSA) as standard, and the carbonyl content was measured by the increase in absorbance at 370 nm and expressed as nanomoles of carbonyl per gram of sample using the molar absorption coefficient of 22,000 M-1 cm-1 for the protein hydrazones (Wang et al., 2022).
The reducing power of the LTL muscle samples was characterized by measuring the total phenolic content (TPC) only in the hydrophilic fraction of raw meat samples aged 2, 7, 14, and 21 days using the Folin-Ciocalteu (FC) method (Singleton and Rossi, 1965; Echegaray et al., 2021). Total phenolic content was determined by measuring the absorbance at 620 nm using a microplate reader. Gallic acid (GA) was used for calibration and TPC values were expressed as mg GA equivalents per gram of sample.
The antioxidant capacity of the meat samples was determined by analyzing both the hydrophilic and lipophilic extracts obtained by homogenizing 1 g of raw meat aged 2, 7, 14, and 21 days in 2 mL of 50 mM phosphate buffer (pH 7) and 2 mL of acetone:ethanol: distilled water (5:4:1), respectively. Both homogenates were centrifuged at 18,000 ×g for 30 minutes at 4°C, and each supernatant was evaluated by reaction with the colored radical solutions described as follows. The 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical scavenging capacity was determined according to Re et al. (1999) measuring the absorbance at 734 nm after 120 min with the NanoDrop 2000c UV-Vis spectrophotometer. A calibration curve was also constructed using Trolox (TX) as a reference and the results were expressed as µmoles of TX per gram of sample. The 1,1-diphenyl-2-picrylhydrazyl radical (DPPH) scavenging capacity was determined according to Brand-Williams et al. (1995). Absorbance was measured at 492 nm in a microplate reader (Infinite F50, Tecan). The radical scavenging capacity was also expressed as µmoles of TX equivalent per gram of sample using a calibration curve.
Determinations were made in raw samples aged 2, 7, 14, and 21 days. Catalase activity (CAT) was measured by recording H2O2 depletion monitored by absorbance at 240 nm after 3 min, according to the method of Aebi (1984). Enzymatic activity was calculated using the molar extinction coefficient of H2O2 (39.4 M-1cm-1) and results were expressed as millimoles.min-1.mg-1 protein.
Total superoxide dismutase activity (SOD) was determined by measuring the inhibition of pyrogallol autoxidation as proposed by Gatellier et al. (2004). The enzymatic reaction was monitored by the increase in absorbance at 340 nm after 5 minutes. One unit of SOD activity was taken as the activity that inhibited the reaction by 50%.
The activity of glutathione peroxidase activity (GPx) was measured at room temperature by recording the oxidation of NADPH by the decrease in absorbance of the incubation mixture at 340 nm after 3 min (DeVore and Greene, 1982). The GPx activity was expressed as the nanomoles of oxidized NADPH.min-1.mg-1 protein.
The proportions of deoxymyoglobin (%DMb), oxymyoglobin (%OxMb), and metmyoglobin (%MMb) were calculated based on the absorbance data (Wyrwisz et al., 2016) on samples of raw meat aged 2, 7, 14, and 21 days, free of external fat and connective tissue. The surface layers excised from the samples with a sharp blade were used (Faustman and Cassens, 1990). One gram of each sample was then finely minced in 9 ml of ice-cold 40 mM phosphate buffer (pH 6.8). After a 1 h blooming period at 4°C, the homogenate was centrifuged (15,000 xg) for 45 min at 4°C. The supernatant was filtered through a Whatman No. 1 filter paper. The absorbance was measured at 503, 525, 557, 582, and 700nm with a NanoDrop 2000c UV-Vis spectrophotometer (Thermo Scientific, USA). The values of %DMb, %OxMb, %MMb, and total myoglobin (Mb) were calculated using the following equations: (Tang et al., 2004; AMSA, 2012).
The structure of the LTL muscle was analyzed by scanning electron microscopy (SEM) following the protocol proposed by Coria et al. (2023). Briefly, 3 samples of raw meat aged 2, 7, 14, and 21 days per group (P or PS) cut longitudinally to the muscle fibers were fixed with Karnovsky’s solution (2.5% paraformaldehyde and 1.5% glutaraldehyde) in 0.1M dibasic phosphate (pH 7.2) at room temperature. The samples were then rinsed twice with distilled water and dehydrated with increasing ethanol gradient solutions (e.g., 30, 50, 50, 70, 90, and 100%) for 10 minutes in each solution at room temperature. The samples were then placed in 100% acetone for 10 minutes, dried with CO2 at critical point conditions (Denton Vacuum, DCP-1), mounted in a holder with double-sided adhesive tape, and gold plated twice for 10 and 20 minutes. The ultrastructure was evaluated using a Phenom ProX scanning electron microscope (SEM, Thermofisher, USA) with an accelerating voltage of 5 kV and a working distance of 2 mm. SEM images were obtained at 1000× for fiber magnification and 15000× for sarcomere structure evaluation. Five randomly selected representative fields per sample were analyzed.
Both hydrophilic and lipophilic extracts of the LTL aged 2, 7, 14, and 21 days muscle samples were also analyzed using an Agilent Cary Eclipse fluorometer in the 3D fluorescence scanning mode (Rey et al., 2021). The FEEMs were recorded with a front-face cuvette configuration at room temperature and a scanning interval between spectra of 5 nm, with both excitation and emission slits fixed at 5 nm.
Data were checked for normality using Infostat software (Di Rienzo et al., 2016). Meat trait data were analyzed using mixed models. Aging time, feeding treatment (P or PS), and their interaction were included as fixed effects and paddock as a random effect in the statistical model. The choice of residual covariance structure was based on the magnitude of the Akaike Information Criterion (the lowest being better). If no significant interactions were observed, the data were reanalyzed for main effects only. If a fixed effect was significant, the least significant difference Fisher test was performed to determine differences between individual treatment means. For all assays, the significance level was set at 0.05.
Figure 1, Supplementary Table S1 illustrate the variation in molecular markers of protein and lipid oxidation in LTL muscle from Braford steers subjected to two distinct feeding regimes (P and PS, respectively) and stored in vacuum package at 2°C for up to 21 days. Protein oxidation, as quantified by the increase in carbonyl residue (-C=O) content, exhibited a gradual increase with aging days (Figure 1A). However, protein carbonylation is approximately 25% higher in animals fed PS than in those fed P (12.2 ± 6.5 vs 8.8 ± 6.7, p < 0.001), as presented in Supplementary Table S1 of the Supplementary Material. This finding is consistent with the 48% increase in carbonyl content observed by Insani et al. (2008) in the psoas major muscle of British Indicus steers finished in grain instead of grass. Furthermore, the triceps brachii muscle from Aberdeen Angus steers fed pasture exhibited lower protein oxidation compared to those fed pasture and concentrate or exclusively concentrate (Terevinto et al., 2015). It is therefore reasonable to hypothesize that the observed increases in protein carbonyl content are associated with lipid oxidation. This is based on the fact that proteins react with oxidized lipid by-products, such as aldehydes or reducing sugars (Domínguez et al., 2022). In this context, the content of lipid hydroperoxides demonstrated minimal variation throughout the aging process. In contrast, a pronounced increase in secondary lipid degradation products, such as malondialdehyde (MDA), was observed (Figures 1B, C, respectively). Consequently, the observed protein carbonylation is strongly indicative of a correlation between lipid degradation and meat aging (Domínguez et al., 2019, 2022). In contrast, no notable discrepancies were identified in the oxidative status of lipids between the two dietary regimens. To enhance the precision of the comparison between the effects of aging on the oxidative status of meat, the mean value between feeding strategies for each parameter during the aging period was calculated (Supplementary Table S1). Subsequently, the aforementioned values (Y) were employed to calculate the relative change in each oxidative marker, as follows: [(Yt – Y0)/Y0], where Y0 and Yt represent the values at the initial and a given day of aging, as illustrated in Figure 1D. The comparative oxidative analysis indicates that lipid degradation occurs at approximately twice the rate of protein carbonylation, whereas the rate of lipid peroxidation remains relatively constant throughout the aging process. Although the formation of MDA in meat is known to produce a rancid odor even at low concentrations, in the present case, the amount of MDA produced even after 21 days of storage is an order of magnitude lower than the accepted rancidity limit of 2.5 mg MDA.kg-1 meat (Campo et al., 2006; Sales et al., 2020).
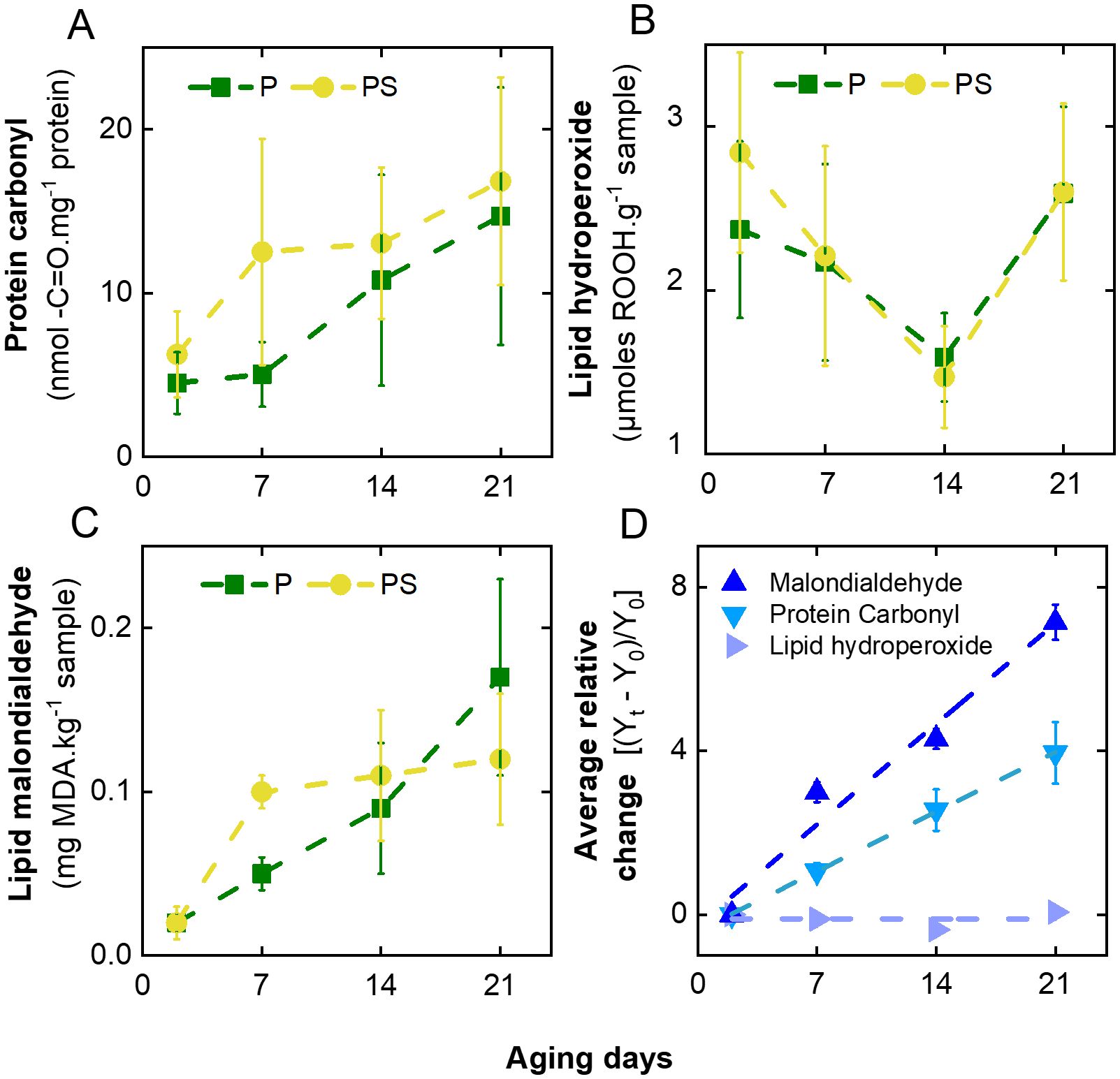
Figure 1. Variation of molecular markers of oxidation with days of aging of LTL muscle from Braford steers aged in vacuum packages at 2°C and fed ad libitum grass (P) or grass-supplemented with corn silage (PS). (A) Protein carbonyls (-C=O), (B) lipid hydroperoxides (ROOH), (C) lipid degradation malondialdehyde (MDA), and (D) mean values during aging for relative change in each molecular marker of meat oxidation.
On the other hand, Insani et al. (2008) observed that samples of psoas major muscle from British x Indicus steers aged 9 days exhibited heigher levels of lipid and protein oxidation in comparison to fresh meat. Furthermore, they noted a higher carbonyl concentration in supplemented animals than in pasture-finished animals. Additionally, elevated levels of protein, carbonyl groups, and MDA were observed in steaks aged for 14 days in the biceps femoris muscle of Aberdeen Angus steers, compared to steaks aged for 2 days (Terevinto et al., 2015). This follows the established phenomenon of oxidative stress occurring in proteins and lipids during post-mortem aging, which results in the expected increase in protein and lipid oxidation (Echegaray et al., 2021).
Figure 2, Supplementary Table S1 illustrate the variation in reducing and antioxidant capacities of LTL muscle samples during the specified storage conditions. The results show that corn silage supplementation did not adversely affect the overall antioxidant status of meat. However, a comparison of TPC, determined by the Folin-Ciocalteu assay and the dye radical scavenging tests, during the aging period of the meat samples shows a different behavior depending on the type of solvent extraction. In the case of the hydrophilic fraction of the meat, the TPC value exhibited a slight decrease with the passage of aging days (Figure 2A), while the total antioxidant capacity for the scavenging of DPPH• and ABTS•+ radicals increased (Figures 2B, C). As the TPC is a measure of the electron-donating capacity of the sample, the scavenging mechanism of both dye radicals involves a mixture of reactions based on hydrogen atom transfer (HAT) and electron transfer (ET) (Echegaray et al., 2021). It can be hypothesized that the aging of meat may result in the formation of hydrophilic hydrogen donor compounds, as opposed to electron donors. Conversely, the radical scavenging capacity of the lipophilic fraction was observed to diminish with the passage of aging days (Figures 2B, C). This finding follows the results obtained, which indicated a higher radical-mediated lipid degradation reaction than protein carbonylation production (Figure 1D). The aforementioned outcomes follow those previously documented by Feidt et al. (1996) who demonstrated that beef, the liberation of free amino acids in beef increases with storage duration and that the concentration of hydrophilic antioxidants increases at a greater rate than that of lipophilic antioxidants as a consequence of cell-matrix fragmentation and protein breakdown.
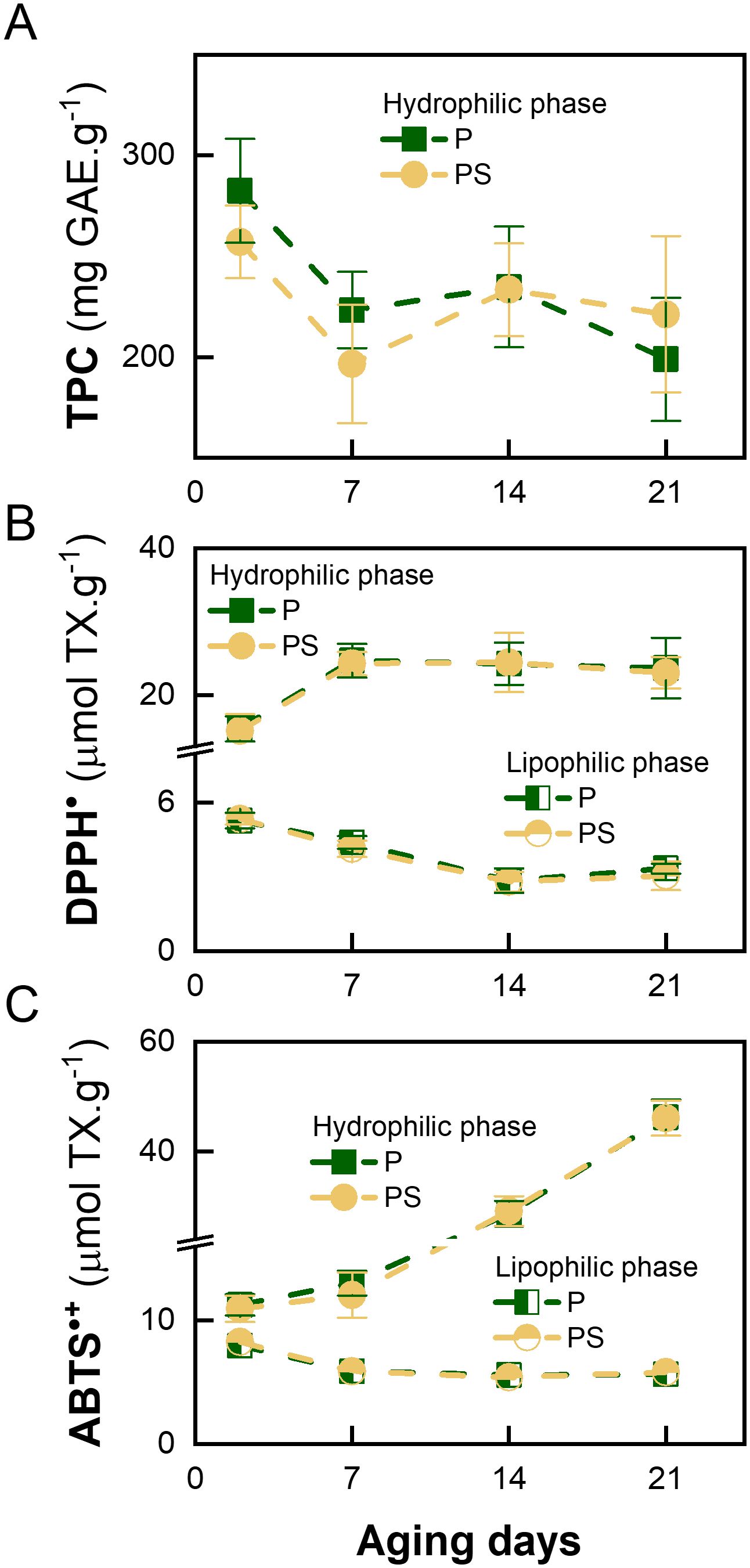
Figure 2. Variation in antioxidant capacity with days of aging of LTL muscle from Braford steers stored in a vacuum package at 2°C fed ad libitum grass (P) or grass supplemented with corn silage (PS). (A) Total phenolic content (TPC) expressed as gallic acid equivalents (GAE), (B) DPPH: 2,2-diphenyl-1-picrylhydrazyl radical scavenging assay expressed as Trolox equivalents (TX) and (C) ABTS: 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) radical scavenging assay expressed as TX.
Moreover, Gobert et al. (2010) demonstrated that the antioxidant capacity of the hydrophilic extract remained constant after 12 days of aging of longissimus thoracis and semitendinosus muscles from Norman cull cows. Also, it was reported that, during the aging of meat in the longissimus dorsi and psoas major muscles of Aberdeen Angus steers, the TPC remained almost constant over 30 days. In contrast, the antioxidant capacity of lipophilic extracts decreased, as evaluated by DPPH• and ABTS•+ scanning assays (Pirotti Corrales, 2019).
Figure 3, Supplementary Table S1 demonstrate that the enzymatic activity of the antioxidant enzymes GPx, SOD, and CAT is largely independent of the dietary regimen of the animals. The activity of GPx remained relatively stable throughout the aging process. Conversely, SOD and CAT exhibited a gradual decline with storage time, particularly after 14 days of aging. This suggests that SOD and CAT are coupled enzymes and that a similar decline in the activity of both enzymes is to be expected during the aging period (Insani et al., 2008). Similarly, Pastsart et al. (2013) observed a reduction in SOD activity at 10 days post-mortem in the longissimus dorsi and femoral biceps muscles of young Belgian Blue bulls. A comparable outcome was documented by Renerre et al. (1996) for the longissimus lumborum, tensor fasciae latae, psoas major, and diaphragm muscles in Francaise Frisonne Pie Noire bulls aged for 8 days.
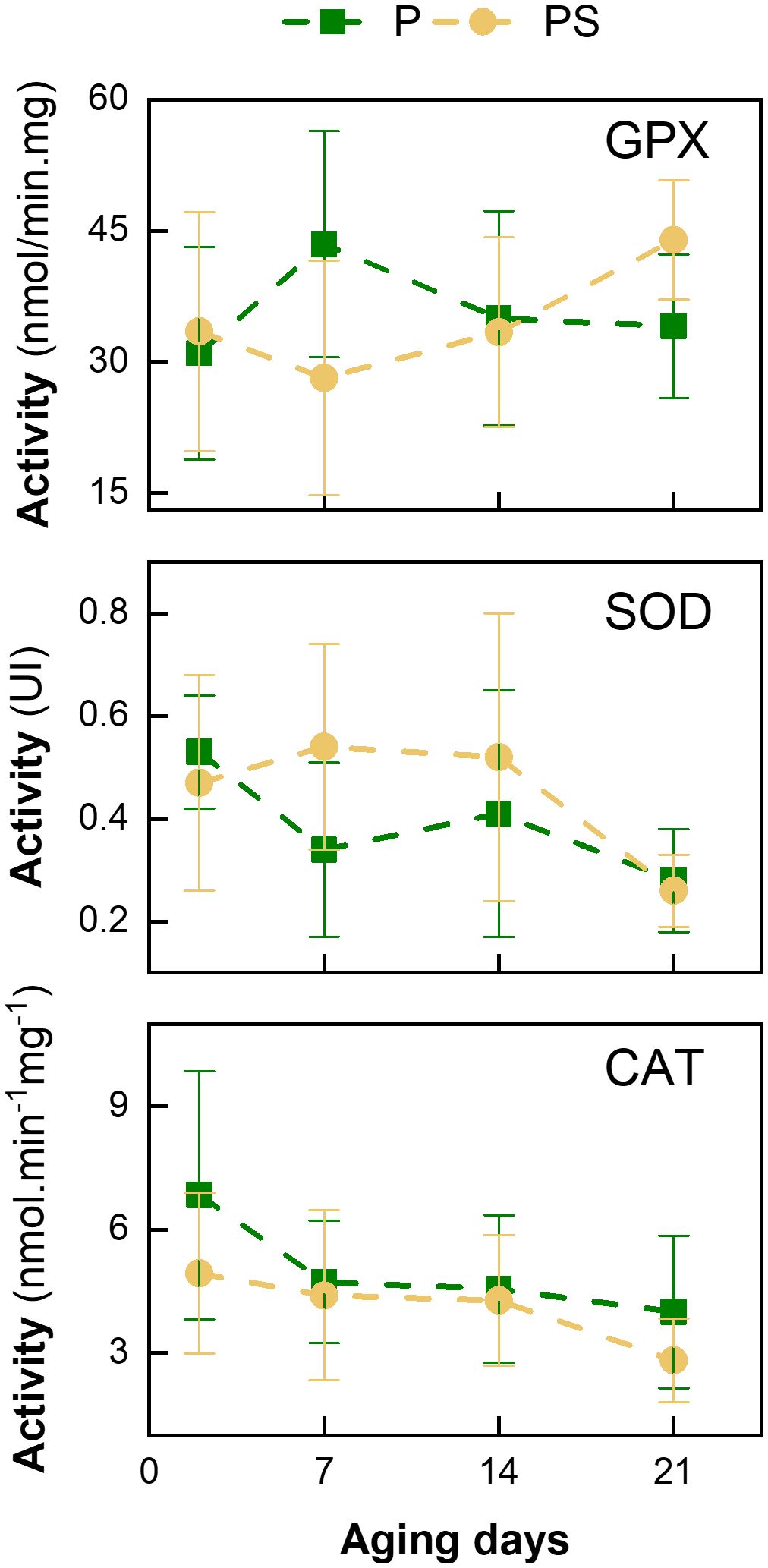
Figure 3. Variation in enzymatic activity of glutathione peroxidase (GPx), superoxide dismutase (SOD), and catalase (CAT) with days of aging of LTL muscle from Braford steers stored in a vacuum package at 2°C fed ad libitum grass (P) or grass supplemented with corn silage (PS).
This can be explained by the inactivation of the SOD enzyme, which was previously described by Bekhit et al. (2013). Furthermore, following the present findings, Daun et al. (2001) observed no differences in GPx activity in steer psoas major and longissimus dorsi muscles over a 14-day aging period. These authors demonstrated that the concentration of the cofactor (selenium) did not decline sufficiently over the 14-day aging period to result in a reduction in GPx activity. This suggests that this enzyme plays a pivotal role in the oxidative defense of the muscle.
Figure 4 illustrates the fluorescence excitation-emission matrix (FEEM) of the hydrophilic and lipophilic extracts derived from the LTL muscle of Braford steers, contrasting the impact of aging between 2- and 21-day samples across both feeding regimens. In the case of the hydrophilic extracts, a sole fluorescence peak was discerned at excitation/emission wavelengths of 285 nm/332 nm. This fluorescence signature is characteristic of partially buried tryptophan (Trp) residues in proteins (Andersen et al., 2018; Liu et al., 2019), given that the fluorescence maximum of Trp in aqueous media is red-shifted to 345 nm (Supplementary Figure S1). In the presence of oxidizing agents, Trp residues can undergo several different degradation pathways, with the final oxidized product dependent on the nature of the oxidant precursor (Bellmaine et al., 2020). The majority of oxygenated Trp degradation products exhibit a red-shifted fluorescent emission band. For example, kynurenine and N-formylkynurenine display emission maximums at 480 nm and 434 nm, respectively (Fukunaga et al., 1982). Furthermore, the formation of advanced Maillard products exhibits a FEEM peak with excitation/emission wavelengths at 320 nm/380 nm, as previously observed in Japanese black cattle meat stored in sealed plastic bags (Liu et al., 2019). In contrast, in the present case, the constant FEEM fingerprint of the hydrophilic extracts, irrespective of the feeding system and aging period, provides compelling evidence that the buried Trp residues in the extracted proteins were effectively protected from any oxidative damage under the vacuum packing storage condition. Accordingly, the slight increase in carbonyl content observed during the aging process (Figure 1D) may be attributed to the oxidation of other protein residues exposed to the media and yield no fluorescent derivatives.
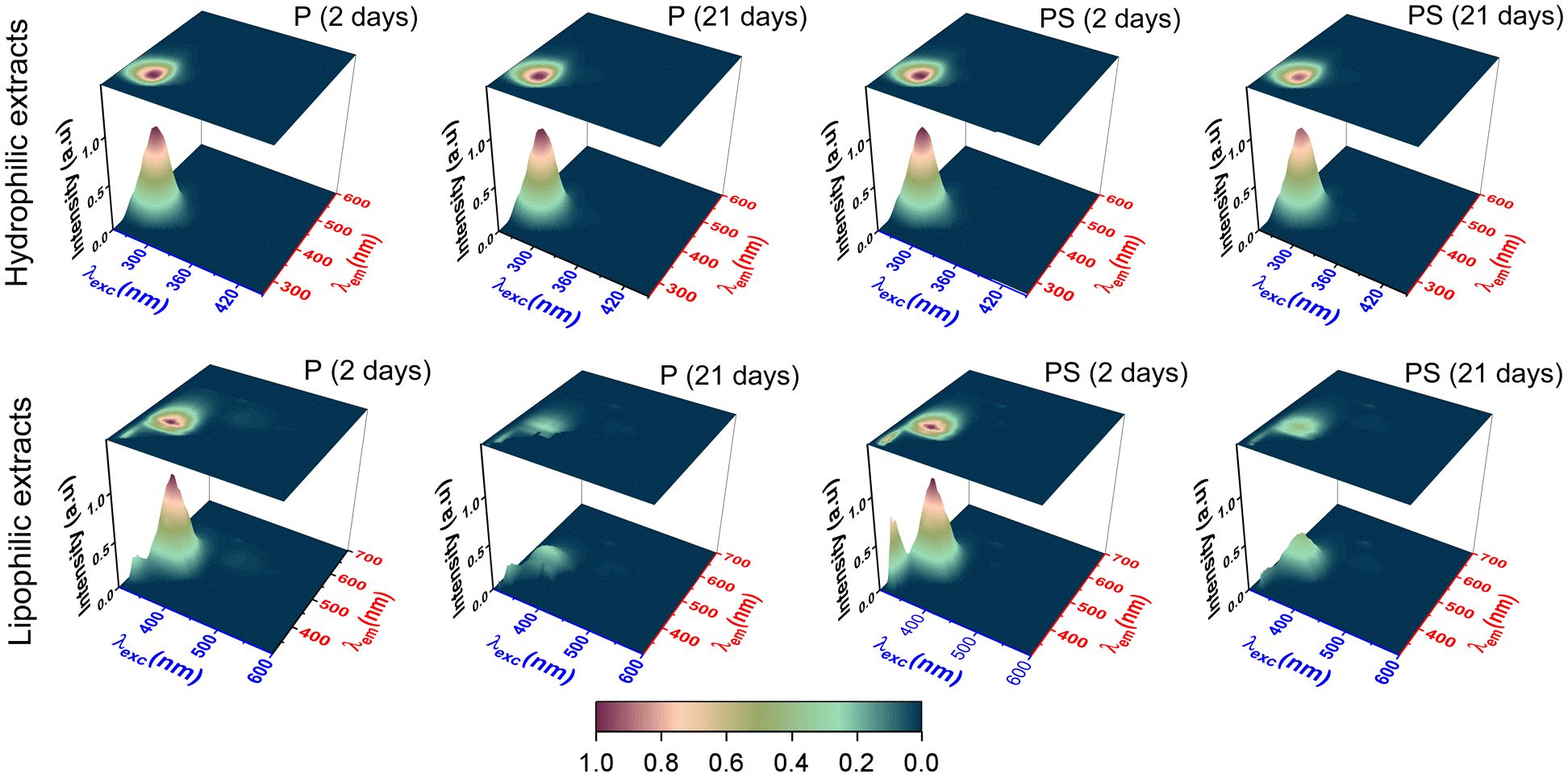
Figure 4. Fluorescence excitation-emission matrix (FEEM) spectra of hydrophilic (top panel) and lipophilic (bottom panel) extracts of samples of longissimus thoracis et lumborum (LTL) muscle from Braford steers fed ad libitum on pasture (P) and pasture supplemented with corn silage (PS), vacuum-packed at 2°C for 2 and 21 days.
Conversely, the FEEM fingerprint of the lipophilic extract from the LTL muscle of Braford steers displayed a distinctive 3D fluorescence topography. The grass-fed and corn silage-supplemented animals exhibited a prominent FEEM peak at excitation/emission wavelengths of 355/434 nm. This observed FEEM corresponded to that of the neat vitamin D (see Supplementary Figure S1). However, with the progression of aging from 2 to 21 days, the intensity of this FEEM band demonstrated a notable decline, reaching approximately 74% and 59% reductions in the grass-fed and supplemented animals, respectively. This result suggests the potential degradation of vitamin D during the storage period, which correlates with the decrease in the antioxidant capacity of the lipid extracts with the days of aging (Figures 2B, C). The loss of vitamin D, together with other lipid-soluble antioxidants, may also be responsible for the increased degradation of lipids (Figures 1C, D). Interestingly, Liu et al. (2019) reported that for n-hexane extracts of adipose tissue from Japanese black cattle meat stored in sealed plastic bags for 24 days, a newly formed peak at 385 nm/470 nm, which was attributed to conjugated Schiff base compounds formed by auto-oxidation reactions within the meat. Therefore, the method of post-mortem meat preservation plays a role in the type and amount of lipophilic off-flavor compounds, which can be easily monitored by the evolution of FEEM fingerprints (Islam et al., 2020).
Figure 5 illustrates the variation in the percentage of heme pigments from the LTL muscle of Braford steers obtained from both feeding systems during the aging period (Supplementary Table S2). It can be observed that there was a concomitant decrease in metmyoglobin (% MMb, Figure 5A) and an increase in oxymyoglobin (%OxMb, Figure 5B) with aging, while deoxymyoglobin (%DMb, Figure 5C) remained constant. Similar behavior was observed for the longissimus dorsi Pirenaica bulls at 14 days of age (Beriain et al., 2009), in semimembranosus muscle after 1 h of blooming in Holstein-Friesian × Simmental crossbred bulls at 21 days of age (Wyrwisz et al., 2016), and also in longissimus thoracis Nellore and Aberdeen Angus bulls during 21 days of aging (Aroeira et al., 2017). The elevated initial values of %MMb observed in the present study may be attributed to prolonged storage in frozen conditions, as previously described by Henriot et al. (2020). In strip loins frozen for up to six months, %MMb has been shown to increase from approximately 25% to 35%. These authors have proposed that the observed increase in MMb is due to a reduction in metmyoglobin-reducing enzymes. On the other hand, the increase in %OxMb during aging has been previously explained by Aroeira et al. (2017) as a reduction in the oxygen consumption rate, which is related to residual post-mortem mitochondrial respiration. In this sense, it has been suggested that during the early storage period under aerobic conditions at low temperatures, the depth of oxygen penetration in meat increases due to a decrease in tissue oxygen consumption by mitochondria and better oxygen diffusion. The combined effect of these processes is that oxygen penetrates more deeply into the meat, oxygenating the myoglobin. This is the mechanism by which the percentage of %OxMb increases during the aging process (Beriain et al., 2009).
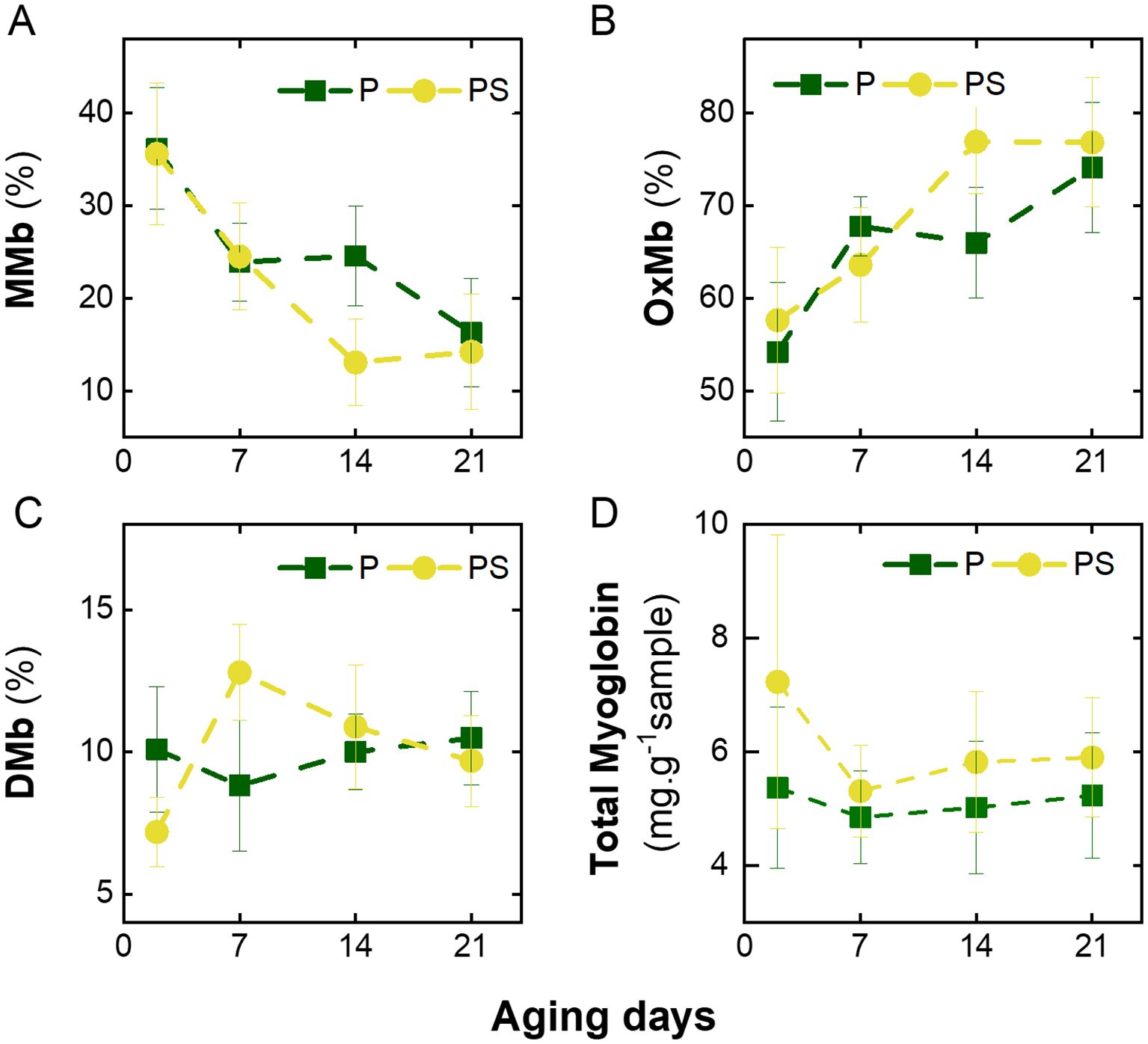
Figure 5. Variation in heme pigments with days of aging of LTL muscle from Braford steers stored in vacuum package at 2°C fed ad libitum grass (P) or grass supplemented with corn silage (PS). (A) Proportion of Metmyoglobin (%MMb), (B) oxymyoglobin (%OxMb), (C) deoxymyoglobin (%DMb), and (D) total myoglobin (Mb).
The PS animals exhibited higher total myoglobin (Mb) concentrations in the LTL muscle than the P group (p < 0.001), with this difference becoming particularly significant at the onset of the aging process (Figure 5D, Supplementary Table S2). Similarly, higher values of Mb concentration were observed in the longissimus thoracis muscle from Spanish Brown Swiss - Limousine bulls that were fed concentrate ad libitum (in the form of a meal and barley straw throughout the finishing period) compared to those that were fed a concentrated diet restricted in quantity (Cerdeño et al., 2006). The authors proposed that concentrated feeding resulted in enhanced protein synthesis, including that of myoglobin protein. Subsequently, in the current case, the total Mb content decreased at day 7 and remained constant during 21 days of aging for both feeding diets (Figure 5D). This behavior was similar to that observed in the longissimus dorsi muscle from Chinese Yellow female cattle stored in a refrigerator for 10 days (Fu et al., 2017). Coria et al. (2022) found with the same samples, an increase in lightness and wyellowness during aging, that can be attributed to protein degradation, which weakens protein structures and enhances light scattering, thereby increasing meat lightness (Beriain et al., 2009). Furthermore, previous research has indicated that muscles with lower levels of total Mb demonstrate higher lightness (Onopiuk et al., 2016). However, it is noteworthy that the discrepancies in total Mb content observed in the present study did not mirror the changes in yellowness values previously documented (Coria et al., 2022). These findings could imply that the presence of multiple pigments, such as carotenoids, deposited in muscle from pasture may counteract the color effect by reducing %Mb.
The results collectively indicate that finishing strategies exerted no discernible influence on heme pigments and, consequently, meat color. Furthermore, it was previously postulated that when %MMb pigment reached 50% on the surface, meat was deemed unacceptable by all consumers (Van den Oord and Wesdorp, 1971). It is noteworthy that in the present study, this heme pigment exhibited values below 36%, suggesting that fresh or aged beef under vacuum packaging could be visually acceptable for consumers. Indeed, several factors can affect the determination of heme pigments in meat. These include the type of muscle, sample preparation, aging temperature (Madhavi and Carpenter, 1993), packaging method, oxygen penetration (Henriot et al., 2020), blooming time (Beriain et al., 2009), analytical techniques employed, storage of samples (Aroeira et al., 2017; Fu et al., 2017), and the equations used for their determination (Tang et al., 2004), among other factors, contribute to the observed variability.
The use of scanning electron microscopy (SEM) allowed the direct morphological high-resolution characterization of the microstructure of the muscle tissue, thereby enabling the measurement of the length of the sarcomeres and the diameter of the muscle fibers (Figure 6). The images obtained indicate that there were no discernible differences in the microstructure of the muscle from animals fed with pasture (Figures 6A–C, E–G) and silage-supplemented (Figures 6I–K, M–O) for up to 14 days of aging. This is evidenced by the identification of fibers adhered to each other and the sarcomeres delimited by the Z-disks. However, the 21-day-aged samples exhibited a notable distinction in the separation of fibers and a less discernible differentiation between sarcomeres across both feeding systems (Figures 6D, H, L, P). Similarly, Palka (2003) observed that the myofibrillar structure of the bull semitendinosus muscle became less distinct after 12 days of aging. They noted that gaps between myofibrils became more visible and that Z-disks were less pronounced. It is important to note that oxidative stress can result in the direct chemical oxidation of numerous contractile proteins, which can lead to alterations in their structural conformation and functional activity, explaining the results obtained in the present work (Steinberg, 2013).

Figure 6. Scanning electron microscopy (SEM) images were obtained at a magnification of 1000x (i.e., white bars of 50 µm) for 2, 7, 14, and 21 days of aging, respectively (panels A through D and I through L), while for the sarcomere structure, a magnification of 15000x (i.e., white bars of 5 µm) was selected for the same aging days (panels E through H and M through P). The LTL muscle samples from Braford steers stored in a vacuum package at 2°C and fed grass (P) are presented in figures A–H, while those fed grass supplemented with corn silage (PS) are presented in panels I–P.
The present study aims to provide comprehensive insights regarding the impact of corn-silage supplementation on the post-mortem oxidative status and heme pigment stability during aging under vacuum packaging of the longissimus thoracis et lumborum muscle in Braford steers. The addition of corn silage to the diet had a minimal impact on the oxidation of lipids and antioxidant capacity when compared to the control group that was fed a diet based on pasture. However, the supplementation resulted in elevated protein carbonyl and total myoglobin levels, without affecting the proportion of heme pigments. Irrespective of the feeding strategy employed, post-mortem aging exerted a pronounced influence on oxidative processes, with lipid degradation occurring at a faster rate than protein oxidation. Moreover, hydrophilic antioxidant capacity increased during the aging process, presumably due to the release of antioxidant peptides, whereas lipophilic antioxidant activity declined, correlating with vitamin D degradation. The findings demonstrate that a 21-day vacuum aging period effectively balances oxidative changes, yielding satisfactory heme pigment contents and, consequently, color, regardless of the feeding system employed. Furthermore, the utilization of analytical techniques, such as fluorescence excitation-emission matrices (FEEM) and scanning electron microscopy (SEM), proved invaluable in capturing the subtleties of changes in the oxidative status and microstructure. These methodologies provide invaluable tools for future research aimed at optimizing meat quality.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by Comite Institucional para el Uso y Cuidado de Animales de Experimentación. The study was conducted in accordance with the local legislation and institutional requirements.
MSCL: Formal analysis, Investigation, Methodology, Writing – review & editing. GAP: Conceptualization, Funding acquisition, Project administration, Resources, Writing – review & editing. CDB: Conceptualization, Data curation, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing. MSC: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Universidad Nacional de Santiago del Estero (UNSE) (23/A289 and 23/A308), Agencia Nacional de Promoción de la investigación, el Desarrollo Tecnológico y la Innovación (PICT 2020-00062 and PICT 2019-02052) and Consejo Nacional de Investigaciones Científicas y Técnicas (PUE 2018 0035).
The authors would like to acknowledge Dr. Eduardo A. Parellada for the technical assistance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2025.1527127/full#supplementary-material
Adeyemi K. D., Sabow A. B., Abubakar A., Samsudin A. A., Sazili A. Q. (2016a). Effects of dietary oil blend on fatty acid composition, oxidative stability and physicochemical properties of Longissimus thoracis et lumborum muscle in goats. Anim. Sci. J. 87, 1421–1432. doi: 10.1111/asj.12597
Adeyemi K. D., Shittu R. M., Sabow A. B., Ebrahimi M., Sazili A. Q. (2016b). Influence of diet and postmortem ageing on oxidative stability of lipids, myoglobin and myofibrillar proteins and quality attributes of gluteus medius muscle in goats. PLoS One 11, 1–15. doi: 10.1371/journal.pone.0154603
Aebi H. (1984). Catalase in vitro. Methods Enzymology 105, 121–126. doi: 10.1016/S0076-6879(84)05016-3
Andersen P. V., Wold J. P., Gjerlaug-Enger E., Veiseth-Kent E. (2018). Predicting post-mortem meat quality in porcine longissimus lumborum using Raman, near infrared and fluorescence spectroscopy. Meat Sci. 145, 94–100. doi: 10.1016/j.meatsci.2018.06.016
Aroeira C. N., de Almeida Torres Filho R., Fontes P. R., de Lemos Souza Ramos A., de Miranda Gomide L. A., Ladeira M. M., et al. (2017). Effect of freezing prior to aging on myoglobin redox forms and CIE color of beef from Nellore and Aberdeen Angus cattle. Meat Sci. 125, 16–21. doi: 10.1016/j.meatsci.2016.11.010
Bekhit A. E. D. A., Hopkins D. L., Fahri F. T., Ponnampalam E. N. (2013). Oxidative processes in muscle systems and fresh meat: Sources, markers, and remedies. Compr. Rev. Food Sci. Food Saf. 12, 565–597. doi: 10.1111/1541-4337.12027
Bellmaine S., Schnellbaecher A., Zimmer A. (2020). Reactivity and degradation products of tryptophan in solution and proteins. Free Radical Biol. Med. 160, 696–718. doi: 10.1016/j.freeradbiomed.2020.09.002
Beriain M. J., Goñi M. V., Indurain G., Sarriés M. V., Insausti K. (2009). Predicting Longissimusdorsi myoglobin oxidation in aged beef based on early post-mortem colour measurements on the carcass as a colour stability index. Meat Sci. 81, 439–445. doi: 10.1016/J.MEATSCI.2008.09.009
Brand-Williams W., Cuvelier M. E., Berset C. (1995). Use of a free radical method to evaluate antioxidant activity. LWT - Food Sci. Technol. 28, 25–30. doi: 10.1016/S0023-6438(95)80008-5
Campo M. M., Nute G. R., Hughes S. I., Enser M., Wood J. D., Richardson R. I. (2006). Flavour perception of oxidation in beef. Meat Sci. 72, 303–311. doi: 10.1016/j.meatsci.2005.07.015
Cerdeño A., Vieira C., Serrano E., Lavín P., Mantecó A. R. (2006). Effects of feeding strategy during a short finishing period on performance, carcass and meat quality in previously-grazed young bulls. Meat Sci. 72, 719–726. doi: 10.1016/j.meatsci.2005.10.002
Coria M. S., Castaño Ledesma M. S., Gómez Rojas J. R., Grigioni G., Palma G. A., Borsarelli C. D. (2023). Prediction of tenderness in bovine longissimus thoracis et lumborum muscles using Raman spectroscopy. Anim. Bioscience 36, 1435–1444. doi: 10.5713/ab.22.0451
Coria M. S., Pighin D., Grigioni G., Palma G. A. (2022). Feeding strategies and ageing time alter calpain system proteins activities and meat quality of Braford steers. Anim. Bioscience 35, 272. doi: 10.5713/AB.21.0227
Coria M. S., Reineri P. S., Pighin D., Barrionuevo M. G., Carranza P. G., Grigioni G., et al. (2020). Feeding strategies alter gene expression of the calpain system and meat quality in the longissimus muscle of Braford steers. Asian-Australasian J. Anim. Sci. 33, 753–762. doi: 10.5713/ajas.19.0163
Daun C., Johansson M., Önning G., Åkesson B. (2001). Glutathione peroxidase activity, tissue and soluble selenium content in beef and pork in relation to meat ageing and pig RN phenotype. Food Chem. 73, 313–319. doi: 10.1016/S0308-8146(00)00303-4
DeVore V. R., Greene B. E. (1982). Glutathione peroxidase in post-rigor bovine semitendinosus muscle. J. Food Sci. 47, 1406–1409. doi: 10.1111/J.1365-2621.1982.TB04949.X
Descalzo A. M., Sancho A. M. (2008). A review of natural antioxidants and their effects on oxidative status, odor and quality of fresh beef produced in Argentina. Meat Science 79, 423–436. doi: 10.1016/j.meatsci.2007.12.006
Descalzo A. M., Rossetti L., Grigioni G., Irurueta M., Sancho A. M., Carrete J., et al. (2007). Antioxidant status and odour profile in fresh beef from pasture or grain-fed cattle. Meat Science 75, 299–307. doi: 10.1016/j.meatsci.2006.07.015
Di Rienzo J. A., Casanoves F., Balzarini M. G., Gonzalez L., Tablada M., Robledo C. W. (2016). InfoStat versión 2012. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available at: http://www.infostat.com.ar.
Domínguez R., Pateiro M., Gagaoua M., Barba F. J., Zhang W., Lorenzo J. M. (2019). A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 8, 429. doi: 10.3390/antiox8100429
Domínguez R., Pateiro M., Munekata P. E. S., Zhang W., Garcia-Oliveira P., Carpena M., et al. (2022). Protein oxidation in muscle foods: A comprehensive review. Antioxidants 11, 60. doi: 10.3390/antiox11010060
Duan J., Cherian G., Zhao Y. (2010). Quality enhancement in fresh and frozen lingcod (Ophiodon elongates) fillets by employment of fish oil incorporated chitosan coatings. Food Chem. 119, 524–532. doi: 10.1016/J.FOODCHEM.2009.06.055
Echegaray N., Pateiro M., Munekata P. E. S., Lorenzo J. M., Chabani Z., Farag M. A., et al. (2021). Measurement of antioxidant capacity of meat and meat products: Methods and applications. Molecules 26, 3880. doi: 10.3390/molecules26133880
Faustman C., Cassens R. G. (1990). The biochemical basis for discoloration in fresh meat: a Review. Journal of Muscle Foods 1, 217–243. doi: 10.1111/j.1745-4573.1990.tb00366.x
Feidt C., Petit A., Bruas-Reignier F., Brun-Bellut J. (1996). Release of free amino-acids during ageing in bovine meat. Meat Sci. 44, 19–25. doi: 10.1016/S0309-1740(96)00088-5
Fu Q. Q., Liu R., Zhou G. H., Zhang W. G. (2017). Effects of packaging methods on the color of beef muscles through influencing myoglobin status, metmyoglobin reductase activity and lipid oxidation. J. Food Process. Preservation 41. doi: 10.1111/jfpp.12740
Fukunaga Y., Katsuragi Y., Izumi T., Sakiyama F. (1982). Fluorescence characteristic of kynurenine and N’-formylkynurenine. Their use as reporters of the environment of tryptophan 62 in hen egg-white lysozyme. J. Biochem. 92, 129–141. doi: 10.1093/oxfordjournals.jbchem.a133909
Gatellier P., Mercier Y., Renerre M. (2004). Effect of diet finishing mode (pasture or mixed diet) on antioxidant status of Charolais bovine meat. Meat Sci. 67, 385–394. doi: 10.1016/j.meatsci.2003.11.009
Gobert M., Gruffat D., Habeanu M., Parafita E., Bauchart D., Durand D. (2010). Plant extracts combined with vitamin E in PUFA-rich diets of cull cows protect processed beef against lipid oxidation. Meat Sci. 85, 676–683. doi: 10.1016/j.meatsci.2010.03.024
Grau A., Codony R., Rafecas M., Barroeta A. G., Guardiola F. (2000). Lipid hydroperoxide determination in dark chicken meat through a ferrous oxidation-xylenol orange method. J. Agric. Food Chem. 48, 4136–4143. doi: 10.1021/JF991054Z
Haak L., Raes K., Van Dyck S., De Smet S. (2008). Effect of dietary rosemary and α-tocopheryl acetate on the oxidative stability of raw and cooked pork following oxidized linseed oil administration. Meat Sci. 78, 239–247. doi: 10.1016/j.meatsci.2007.06.005
Henriot M. L., Herrera N. J., Ribeiro F. A., Hart K. B., Bland N. A., Calkins C. R. (2020). Impact of myoglobin oxygenation level on color stability of frozen beef steaks. Journal of Animal Science 98, skaa193.
Huff Lonergan E., Zhang W., Lonergan S. M. (2010). Biochemistry of postmortem muscle - lessons on mechanisms of meat tenderization. Meat Sci. 86, 184–195. doi: 10.1016/j.meatsci.2010.05.004
Insani E. M., Eyherabide A., Grigioni G., Sancho A. M., Pensel N. A., Descalzo A. M. (2008). Oxidative stability and its relationship with natural antioxidants during refrigerated retail display of beef produced in Argentina. Meat Sci. 79, 444–452. doi: 10.1016/j.meatsci.2007.10.017
Islam K., Mahbub S. B., Clement S., Guller A., Anwer A. G., Goldys E. M. (2020). Autofluorescence excitation-emission matrices as a quantitative tool for the assessment of meat quality. J. Biophotonics 13, 1–8. doi: 10.1002/jbio.201900237
Liu H., Saito Y., Al Riza D. F., Kondo N., Yang X., Han D. (2019). Rapid evaluation of quality deterioration and freshness of beef during low temperature storage using three-dimensional fluorescence spectroscopy. Food Chem. 287, 369–374. doi: 10.1016/j.foodchem.2019.02.119
Lund M. N., Hviid M. S., Claudi-Magnussen C., Skibsted L. H. (2008). Effects of dietary soybean oil on lipid and protein oxidation in pork patties during chill storage. Meat Sci. 79, 727–733. doi: 10.1016/j.meatsci.2007.11.008
Madhavi D. L., Carpenter C. E. (1993). Aging and Processing Affect Color, Metmyoglobin Reductase and Oxygen Consumption of Beef Muscles. Journal of Food Science 58, 939–942. doi: 10.1111/j.1365-2621.1993.tb06083.x
Nawrocka A., Lamorsk J. (2013). Determination of Food Quality by Using Spectroscopic Methods. Adv. Agrophysical Res. (InTech). doi: 10.5772/52722
Nimse S. B., Pal D. (2015). Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 5, 27986–28006. doi: 10.1039/c4ra13315c
Oliver C. N., Ahn B. W., Moerman E. J., Goldstein S., Stadtman E. R. (1987). Age-related changes in oxidized proteins. J. Biol. Chem. 262, 5488–5491. doi: 10.1016/S0021-9258(18)45598-6
Onopiuk A., Półtorak A., Wierzbicka A. (2016). Influence of post-mortem muscle glycogen content on the quality of beef during aging. J. Veterinary Res. (Poland) 60, 301–307. doi: 10.1515/jvetres-2016-0046
Palka K. (2003). The influence of post-mortem ageing and roasting on the microstructure, texture and collagen solubility of bovine semitendinosus muscle. Meat Sci. 64, 191–198. doi: 10.1016/S0309-1740(02)00179-1
Pastsart U., De Boever M., Claeys E., De Smet S. (2013). Effect of muscle and post-mortem rate of pH and temperature fall on antioxidant enzyme activities in beef. Meat Sci. 93, 681–686. doi: 10.1016/j.meatsci.2012.11.008
Pirotti Corrales F. D. (2019). Estatus antioxidante de carne bovina Uruguaya: Influencia del sistema de producción, tipo de músculo y tiempo de maduración. Magister Thesis.. (Montevideo, Uruguay: Universidad Nacional de la República). Available at: https://www.colibri.udelar.edu.uy/jspui/handle/20.500.12008/27460.
Ponnampalam E. N., Kiani A., Santhiravel S., Holman B. W. B., Lauridsen C., Dunshea F. R. (2022). The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality—Invited Review. Animals 12, 332279. doi: 10.3390/ani12233279
Pouzo L. B., Descalzo A. M., Zaritzky N. E., Rossetti L., Pavan E. (2016). Antioxidant status, lipid and color stability of aged beef from grazing steers supplemented with corn grain and increasing levels of flaxseed. Meat Science 111, 1–8. doi: 10.1016/j.meatsci.2015.07.026
Qin J., Kim M. S., Chao K., Schmidt W. F., Cho B. K., Delwiche S. R. (2017). Line-scan Raman imaging and spectroscopy platform for surface and subsurface evaluation of food safety and quality. Journal of Food Engineering 198, 17–27. doi: 10.1016/j.jfoodeng.2016.11.016
Re R., Pellegrini N., Proteggente A., Pannala A., Yang M., Rice-Evans C. (1999). Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med. 9–10, 1231–1237. doi: 10.1016/S0891-5849(98)00315-3
Renerre M., Dumont F., Gatellier P. (1996). Antioxidant enzyme activities in beef in relation to oxidation of lipid and myoglobin. Meat Sci. 43, 111–121. doi: 10.1016/0309-1740(96)84583-9
Rey V., Abatedaga I., Vera C., Vieyra F. E. M., Borsarelli C. D. (2021). Photosensitized formation of soluble bionanoparticles of lysozyme. ChemistrySelect 6, 13443–13451. doi: 10.1002/slct.202103215
Ripoll G., Blanco M., Albertí P., Panea B., Joy M., Casasús I. (2014). Effect of two Spanish breeds and diet on beef quality including consumer preferences. J. Sci. Food Agric. 94, 983–992. doi: 10.1002/jsfa.6348
Rosa H. J. D., Rego O. A., Silva C. C. G., Alves S. P., Alfaia C. M. M., Prates J. A. M., et al. (2014). Effect of corn supplementation of grass finishing of Holstein bulls on fatty acid composition of meat lipids. Journal of Animal Science 92, 3701–3714. doi: 10.2527/jas.2013-6982
Sabow A. B., Sazili A. Q., Aghwan Z. A., Zulkifli I., Goh Y. M., Ab Kadir M. Z. A., et al. (2016). Changes of microbial spoilage, lipid-protein oxidation and physicochemical properties during post mortem refrigerated storage of goat meat. Anim. Sci. J. 87, 816–826. doi: 10.1111/asj.12496
Sales L. A., Rodrigues L. M., Silva D. R. G., Fontes P. R., Torres Filho R de A., Ramos A de L. S., et al. (2020). Effect of freezing/irradiation/thawing processes and subsequent aging on tenderness, color, and oxidative properties of beef. Meat Sci. 163, 108078. doi: 10.1016/j.meatsci.2020.108078
SENASA (2016). Manual de Bienestar Animal: un enfoque práctico para el buen manejo de especies domésticas durante su tenencia producción concentración transporte faena. Available online at: https://biblioteca.senasa.gob.ar/items/show/3895. (Accessed January 14, 2025)
Singleton V. L., Rossi J. A. (1965). Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enology Viticulture 16, 144–158. doi: 10.5344/ajev.1965.16.3.144
Steinberg S. F. (2013). Oxidative stress and sarcomeric proteins. Circ. Res. 112, 393–405. doi: 10.1161/CIRCRESAHA.111.300496
Tang J., Faustman C., Hoaglands T. A. (2004). Krzywicki Revisited : Equations for spectrophotometric determination of myoglobin redox forms in auqueous meat extracts. J. Food Sci. 69, 717–720. doi: 10.1111/j.1365-2621.2004.tb09922.x
Terevinto A., Cabrera M. C., Saadoun A. (2015). Catalase, SOD and GPx activities in triceps brachii muscle from aberdeen angus steers finished on pasture, pasture and concentrate, or concentrate. Am. J. Food Nutr. 3, 118–124. doi: 10.12691/ajfn-3-5-2
Terevinto A., Cabrera M. C., Saadoun A. (2019). Oxidative stability, fatty acid composition and health lipid indices of longissimus dorsi muscle from Aberdeen angus steers produced in different feeding systems. Ciencia Rural 49, 1–11. doi: 10.1590/0103-8478cr20190537
Van den Oord A. H. A., Wesdorp J. J. (1971). Analysis of pigments in intact beef samples: A simple method for the determination of oxymyoglobin and ferric myoglobin in intact beef samples using reflectance spectrophotometry. Int. J. Food Sci. Technol. 6, 1–13. doi: 10.1111/j.1365-2621.1971.tb01587.x
Vincenti F., Carbone K., Zilio D. M., Ronchi B., Gigli S., Iacurto M. (2009). Effects of dietary supplementation with mineral/vitamin mix on beef meat colour and oxidation parameters. Ital. J. Anim. Sci. 8, 558–560. doi: 10.4081/ijas.2009.s2.558
Yoshimura M., Sugiyama J., Tsuta M., Fujita K., Shibata M., Kokawa M., et al. (2014). Prediction of Aerobic Plate Count on Beef Surface Using Fluorescence Fingerprint. Food and Bioprocess Technology 7, 1496–1504. doi: 10.1007/s11947-013-1167-8
Wang Z., Liu X., Ojangba T., Zhang L., Yu Q., Han L. (2022). Storage and packaging effects on the protein oxidative stability, functional and digestion characteristics of yak rumen smooth muscle. Foods 11. doi: 10.3390/foods11142099
Wu X., Liang X., Wang Y., Wu B., Sun J. (2022). Non-destructive techniques for the analysis and evaluation of meat quality and safety: A review. Foods 11, 1–30. doi: 10.3390/foods11223713
Keywords: beef, corn supplementation, antioxidants, oxidation, meat color, fluorescence
Citation: Castaño Ledesma MS, Palma GA, Borsarelli CD and Coria MS (2025) Comparative analysis of feeding strategies and post mortem aging time on the oxidative status and color of the longissimus thoracis et lumborum muscle in Braford steers. Front. Anim. Sci. 6:1527127. doi: 10.3389/fanim.2025.1527127
Received: 12 November 2024; Accepted: 03 January 2025;
Published: 28 January 2025.
Edited by:
Eva Maria Santos, Autonomous University of the State of Hidalgo, MexicoReviewed by:
Maria Rubio, National Autonomous University of Mexico, MexicoCopyright © 2025 Castaño Ledesma, Palma, Borsarelli and Coria. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María Sumampa Coria, c3VtaWNvcmlhQGdtYWlsLmNvbQ==; Claudio Darío Borsarelli, Y2Rib3JzYXJlbGxpQGdtYWlsLmNvbQ==
†ORCID: María Sofía Castaño Ledesma, orcid.org/0000-0001-8578-5748
Gustavo Adolfo Palma, orcid.org/0000-0002-9175-1322
Claudio Darío Borsarelli, orcid.org/0000-0003-0120-645X
María Sumampa Coria, orcid.org/0000-0002-3472-8775
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.