
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PROTOCOLS article
Front. Physiol. , 19 June 2018
Sec. Systems Biology Archive
Volume 9 - 2018 | https://doi.org/10.3389/fphys.2018.00646
This article is part of the Research Topic Logical Modeling of Cellular Processes: From Software Development to Network Dynamics View all 23 articles
 Aurélien Naldi1*
Aurélien Naldi1* Céline Hernandez1Wassim Abou-Jaoudé1
Céline Hernandez1Wassim Abou-Jaoudé1 Pedro T. Monteiro2
Pedro T. Monteiro2 Claudine Chaouiya3*
Claudine Chaouiya3* Denis Thieffry1*
Denis Thieffry1*The logical formalism is well adapted to model large cellular networks, in particular when detailed kinetic data are scarce. This tutorial focuses on this well-established qualitative framework. Relying on GINsim (release 3.0), a software implementing this formalism, we guide the reader step by step toward the definition, the analysis and the simulation of a four-node model of the mammalian p53-Mdm2 network.
The logical formalism is becoming increasingly popular to model cellular networks (Naldi et al., 2015; Abou-Jaoudé et al., 2016). Here, we focus on the framework developed by René Thomas and colleagues, which includes the use of multi-valued variables when functionally justified, along with sophisticated logical rules or parameters (Thomas, 1991; Thomas et al., 1995).
This approach has been applied to the study of a wide range of networks controlling, for example, the lysis-lysogeny decision of the bacteriophage λ (Thieffry and Thomas, 1995), the specification of flower organs in arabidopsis (Mendoza et al., 1999; Azpeitia et al., 2014), the segmentation of drosophila embryo (Sánchez and Thieffry, 2001; Sánchez and Thieffry, 2003; Sánchez et al., 2008; Mbodj et al., 2016), the specification of compartments in drosophila imaginal disks (González et al., 2006, 2008), drosophila egg shell patterning (Fauré et al., 2014), the control of cell cycle in yeast and mammals (Fauré et al., 2006, 2009; Traynard et al., 2016), the specification of immune cells from common progenitors (Mendoza and Méndez, 2015; Collombet et al., 2017), the differentiation of T-helper lymphocytes (Naldi et al., 2010; Abou-Jaoudé et al., 2015; Martinez-Sanchez et al., 2015), neuronal differentiation (Coolen et al., 2012), as well as cancer cell fate decisions (Sahin et al., 2009; Calzone et al., 2010; Grieco et al., 2013; Flobak et al., 2015; Remy et al., 2015), etc.
In order to ease access to logical modeling by biologists, this protocol proposes a stepwise introduction to the framework, relying on its implementation into the software GINsim (release 3.0). The following section introduces the biological system used as an illustration. Next, in section 3, we proceed with the stepwise construction and analysis of a logical model. Section 4 covers potential troubleshooting. The article then ends with some conclusions and prospects.
The transcription factor p53 plays an essential role in the control of cell proliferation in mammals by regulating a large number of genes involved notably in growth arrest, DNA repair, or apoptosis (Vogelstein et al., 2000). Its level is tightly regulated by the ubiquitin ligase Mdm2. More precisely, nuclear Mdm2 down-regulates the level of active p53, both by accelerating p53 degradation through ubiquitination (Brooks and Gu, 2006) and by blocking the transcriptional activity of p53 (Oliner et al., 1993; Coutts et al., 2007). In turn, p53 activates Mdm2 transcription (Barak et al., 1993) and down-regulates the level of nuclear Mdm2 by inhibiting Mdm2 nuclear translocation through inactivation of the kinase Akt (Mayo and Donner, 2002). Finally, high levels of p53 promote damage repair by inducing the synthesis of DNA repair proteins (Gatz and Wiesmüller, 2006).
Given its key role in DNA repair and cell fate control, various groups have modeled this network using different formalisms, including ordinary differential equations (Ciliberto et al., 2005; Zhang et al., 2011), stochastic models (Puszynski et al., 2008; Ouattara et al., 2010; Sun and Cui, 2014), hybrid deterministic and stochastic models (Iwamoto et al., 2014), as well as logical models (Abou-Jaoudé et al., 2009; Choi et al., 2012).
In this protocol, we rely on a refined version of a logical model presented by Abou-Jaoudé et al. (2009), involving the protein p53, the ubiquitin ligase Mdm2 in the cytoplasm, the ubiquitin ligase Mdm2 in the nucleus, and DNA damage (see Figure 1).
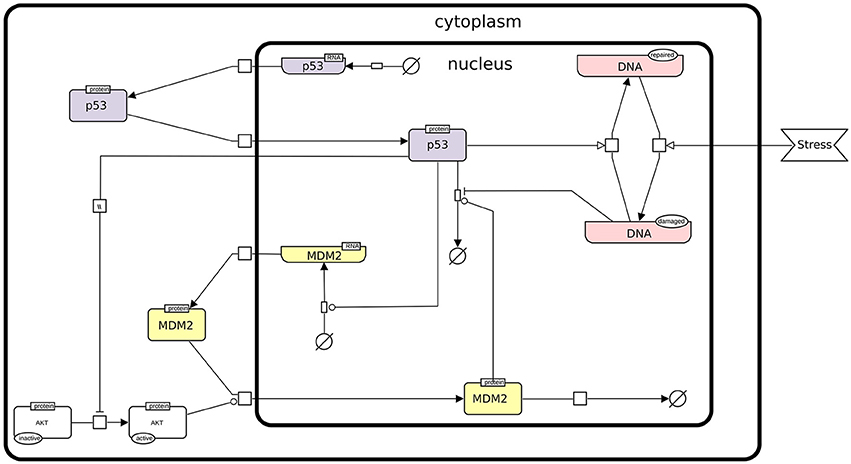
Figure 1. The p53-Mdm2 network. This figure describes the interactions between p53, Mdm2, and DNA damage. An external stress induces a damage to the DNA, which promotes Mdm2 degradation. The level of p53 can then increase and activate DNA repair mechanisms. In parallel, p53 inhibits Mdm2 translocation from the cytoplasm to the nucleus through the inactivation of AKT. However, in the nucleus, high level of p53 activates Mdm2 transcription, while Mdm2 induces the degradation of p53, thereby forming a negative feedback circuit. This figure has been drawn according to the Systems Biology Graphical Notation (SBGN) specifications (Le Novère et al., 2009).
In this section, referring to the p53-Mdm2 network defined above, we introduce the different steps required for the definition of a logical model and for the analysis of its dynamical properties with the software GINsim, release 3.0.
The GINsim software supports the definition, the simulation and the analysis of regulatory graphs, based on the (multi-valued) logical formalism. GINsim is freely available from its dedicated website (http://ginsim.org), along with documentation and a model repository. For this tutorial, we use the recent release 3.0, which is available for all platforms with version 8 of the Java Virtual Machine.
To get started with GINsim, download the corresponding Java ARchive (JAR file), with dependencies included, from the download section of GINsim website (http://ginsim.org/downloads). On your computer, double-click on the file icon to start the application or launch it with the command: java -jar GINsim-♯version.jar in a terminal. Further instructions, troubleshooting and options are documented on the website.
Upon launch, GINsim displays a window enabling the creation of a new model, the import of a model in a supported format, or the opening of a previously defined model (if any). By clicking on the New model button, a window enabling the edition of a new logical regulatory graph opens.
To edit a graph, use the toolbox located just on the top of the window (below the menu bar, see Figure 2). Passing slowly with the mouse on each of the editing tools displays a message explaining the function of each tool. Clicking on the E icon enables further edition of an existing node or arc upon selection, while the garbage can icon serves to delete selected arcs and nodes. Clicking once on one of the remaining icons enables the drawing of a single node or arc. Clicking twice on one of these tools locks the corresponding editing mode, enabling the drawing of several nodes or arcs without clicking repeatedly on the same tool.
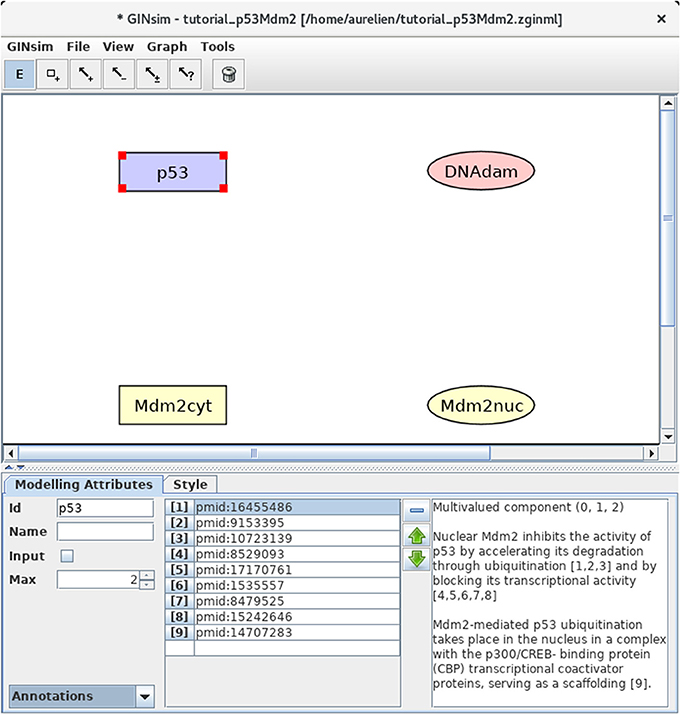
Figure 2. GINsim main window displaying the nodes of the p53-Mdm2 logical regulatory graph. The upper part of the window displays five scrolling menus. These menus provide access to classical file management options, as well as exports into various formats. The central area displays the regulatory graph (here the nodes of the p53-Mdm2 model), while the other area contains two tabs: the Modeling Attributes tab (selected here) and the Style tab, corresponding to the selected node, here p53. The graphical appearance of the nodes have been modified using the Style tab. The Edit button on the top is selected and emphasized in blue, enabling the edition of the attributes of the selected node, including its id and name, its maximal level (Max, here set to 2), and also the insertion of annotations in the form of free text (bottom right) or of links to relevant database entries (bottom middle).
First, we need to define four nodes for the four key regulatory factors of the model: p53, Mdm2cyt, Mdm2nuc, and DNA damage (DNAdam). Each node has a unique identifier and a maximal level, specifying a range of possible functional qualitative levels, as listed in Table 1. To define all the nodes in a row, first double-click on the node addition tool (symbol is a square with a plus sign) to lock this mode, then click four times on the panel to create the four nodes, with default identifiers and a maximal level of 1. Next, click on the E icon to stop adding nodes, and select each node to change its ID and maximal level (when required) in the bottom edition panel. Figure 2 illustrates this step.

Table 1. Regulatory nodes and maximal levels for the p53-Mdm2 model.
Next, we need to define the arcs representing the regulatory interactions between the factors considered in the model. An arc is defined by its source and target nodes, a sign, and a threshold, as described in Table 2 and illustrated in Figure 3. In the non-Boolean case, a node may have distinct actions on a target node, depending on its activity level (e.g., from Mdm2cyt onto Mdm2nuc). In this case, one arc is drawn, which encompasses multiple interactions, each with its own threshold. An interaction is then active when the level of its source is equal or above its threshold, but below the threshold of the next interaction. Add each arc between each relevant pair of nodes by selecting the relevant tool (addition of positive, negative, dual, or unknown interaction) and dragging a line from the source to the target node. Next, use the edition panel to specify multiple interactions with their thresholds, and possibly change their signs.
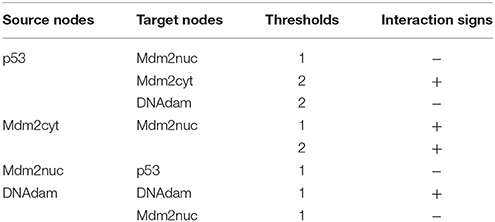
Table 2. Interactions and thresholds for the p53-Mdm2 model.
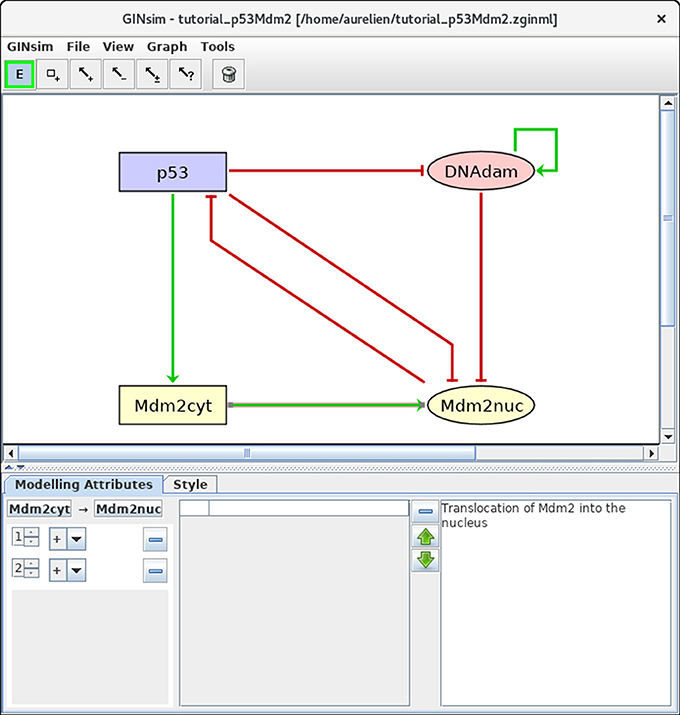
Figure 3. Regulatory arc management in GINsim. To add an arc, the corresponding arc button must be pushed (push twice to add several arcs in one go), allowing the drawing of an arc between a source node and its target. Once an arc has been defined, it can be further edited by selecting it after locking the E button. The sign and threshold of the interaction(s) associated with an arc are defined within the Modeling Attributes tab, as shown here for the arc from Mdm2cyt onto Mdm2nuc. The additional interaction with threshold level 2 was created by clicking on the + button displayed when additional thresholds are available.
We can now define the rules governing the evolution of the regulatory node levels. For each node, specify the logical rules listed in Table 3. For this, select a node and the Formulae view in the drop-down list at the bottom left of the GINsim window. Click on the little arrow in the main bottom panel, expand the tree view and then click on the E button, to enter a formula. Figure 4 illustrates this step. Note that the definition of adequate logical rules (or parameters, see Note 1) is necessary to ensure the desired effects of each interaction on the target nodes. Per default, GINsim assigns a null target value to each node devoid of explicit rule.

Table 3. Logical rules for the nodes of the p53-Mdm2 model.
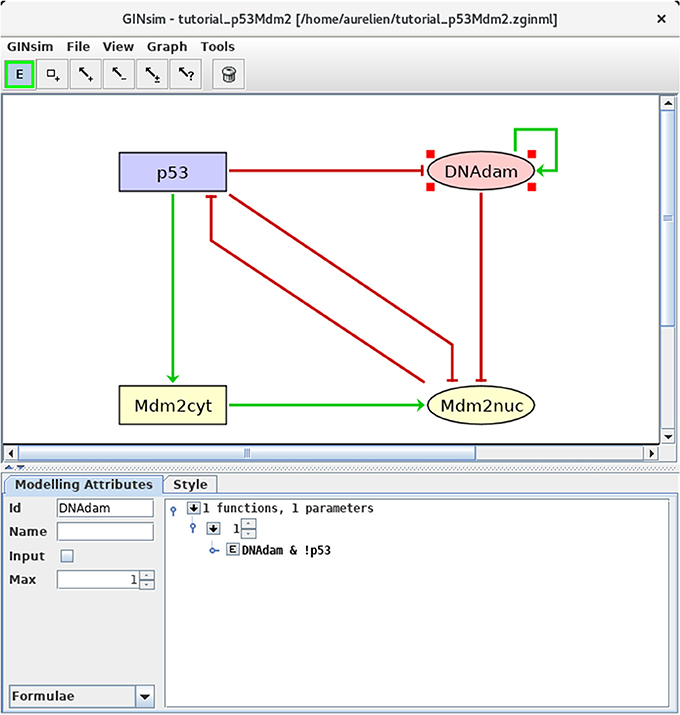
Figure 4. Defining logical rules for the regulatory nodes. This screenshot shows the Modeling Attributes associated with the selected node DNAdam. The maximal level is set to 1. After selecting Formulae with the bottom-left scrolling menu, the user can enter logical formulae by clicking on the little arrows in the main bottom. The target level (set to 1 per default) can be changed in the case of a multi-valued node. By clicking on the E button, one can directly write a formula, using literals (these should exactly match the IDs of nodes regulating the selected node, i.e., p53 or DNAdam in the present case) and the Boolean operators !, & and |, denoting NOT, AND and (inclusive) OR, respectively (following the usual priority ordering; parentheses can be used to define complex formulae). Note that several rows can be used in association with a single target value; these rows are then combined with OR operators. Here, the formula DNAdam & !p53 associated with the target value 1 implies that DNAdam will be maintained at a level 1 if already present, but only in the absence of p53.
To keep track of supporting data and modeling assumptions, the user can add textual annotations and hyperlinks to relevant database entries, at the level of the model itself, as well as for each individual node or arc (see Figure 2 for an illustration). While the annotation panel is always visible when editing an arc, it requires to select the Annotations view (in the bottom left drop-down list) when editing a node.
The layout and graphical appearance of nodes and arcs of the graph can be changed according to the user taste. For this, select a node or an arc, along with the Style tab. The user can further change the default style or define new styles. To change the graph layout, drag a node to change its position or drag an arc to create a new intermediate point. An existing intermediate point can be moved or deleted using right-click.
Selecting the Modeling Attributes tab, with no object selected in the main window, verify that the order of the nodes is: p53, Mdm2cyt, Mdm2nuc, DNAdam. If this is not the case, modify the node order accordingly, using the arrows close to the node list at the left of the Modelling attribute tab. Using this node order will ease the comparison of your results with the Figures hereafter.
The model along with simulations settings (see hereafter) can be saved into a compressed archive (with a zginml extension) by using the Save option in the File menu. Save the model regularly during its encoding, as there is no undo functionality.
The qualitative state of a logical model is defined by the activity levels of its nodes. At a given state, the rules associated with each node define its target level. When the current level of a node is different from its target level, it is called to update toward this target level, resulting in a transition to another state. Several nodes can be called for update at a given state.
Two main strategies are then commonly used. Under the synchronous updating, all concerned nodes change their levels simultaneously in a unique transition toward a single successor state. In contrast, the asynchronous updating generates a successor state for each single node update. If the current state involves k updating calls, it will thus have k successors, each differing from the current state by the level of a single node (see Note 2 for additional explanations). The introduction of priority classes allows to define subtler updating schedules (see Note 3 and Fauré et al., 2006).
The resulting state transitions define another type of graph called state transition graph (STG), which represents the dynamical behavior of the logical model (i.e., the regulatory graph + logical rules). In this graph, the nodes correspond to logical states, while the arcs represent state transitions induced by the rules along with the updating scheme. Using the default level layout of GINsim for state transition graphs, it is easy to spot the stable states, defined as nodes with no outgoing arcs, displayed at the bottom. More complex attractors, defined as terminal strongly connected components (SCCs, maximal sets of nodes that are mutually reachable) denote oscillatory behaviors, which are harder to grasp visually.
Beyond the identification of attractors, we are particularly interested in knowing which of them can be reached from specific initial conditions. Such questions can be addressed by verifying the existence of trajectories (i.e., sequences of transitions), e.g., from initial states to attractor states.
Selecting the Run Simulation option in the Tools menu opens a panel enabling the construction of the dynamics (see Figure 5).
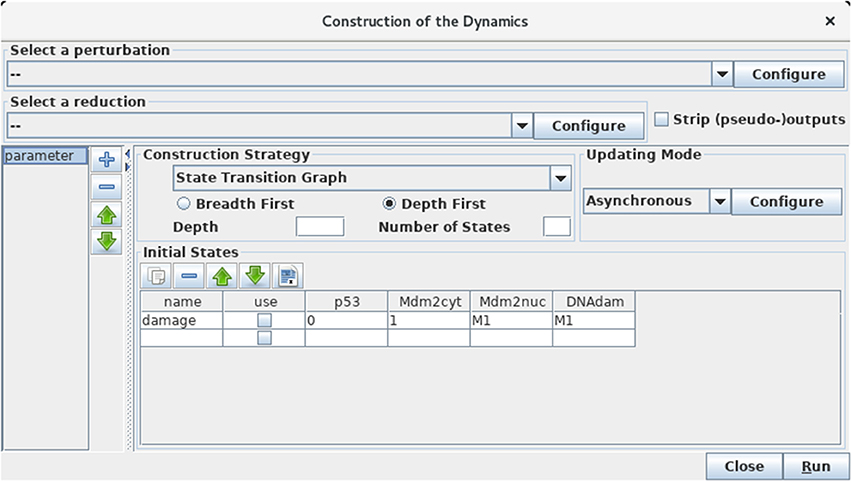
Figure 5. Launching of the construction of a state transition graph. This panel is obtained when selecting Run Simulation from the Tools scrolling menu in GINsim main window. The default simulation settings are shown, i.e., the construction of a state transition graph using the asynchronous updating, with no specified initial state (meaning that all states are considered in the simulation). Hitting the Run button will generate the corresponding state transition graph, which can be displayed in a new window (see Figure 6). In the table under Initial States, one can define one or several initial states from which the dynamics will be constructed (just type the desired values in a row along with an optional name). Each row of the table defines a single pattern of states, and the check-boxes allow to select the states to be used for a simulation. The levels are specified for each node in the corresponding table cell. Nodes for which values are left free are denoted by stars (*). Initial states can be reordered, deleted and duplicated using the buttons just above the table. Here, a unique initial state has been defined, but not selected for simulation: the state 0111 (i.e., with p53 set to 0, and the three other nodes set to 1). Note that M1 emphasizes the fact that the value 1 is the maximal level for Mdm2nuc and for DNAdam. Several parameter configurations can be created and stored using the + button on the left side.
The boxes on the top of the panel labeled by Select a perturbation and Select a reduction permit to define (by clicking on the Configure buttons) and select (using the scrolling menus) model perturbations and reductions (see below).
The bottom left panel enables the definition and the recording of different parameter settings, which greatly facilitates the reproduction of simulation results. One can create, delete and reorder parameter settings by using the buttons on the right of the panel listing the parameter settings.
Regarding the construction strategy, a scrolling menu enables the choice between the generation of a state transition graph (STG), its compression into a strongly connected components graph (SCC), or its further compression into a hierarchical transition graph (HTG) (for more details about these STG compressions, see Bérenguier et al., 2013). Using another scrolling menu, the user can select the synchronous or asynchronous updating, or define or select predefined priority classes (see Note 3 for more details on priority classes).
Finally, the Initial State box enables the definition and/or the selection of initial state(s), from which the construction of the dynamics will be performed. Initial states can be combined with defined sets of Fixed inputs (defined in the panel just below). If no initial state is selected or specified, all the states will be considered in the simulation, leading to the construction of a full STG. As the number of possible states doubles with each additional (Boolean) node, the computation of the full STG is discouraged for models involving more than 15 nodes.
Let us first consider the construction of the asynchronous dynamics. Before launching the simulation, check that the default settings are specified as in Figure 5: state transition graph, asynchronous updating, no perturbation selected, no initial state selected. To ease comparisons with the figures enclosed in this protocol, verify that the order of the nodes is: p53, Mdm2cyt, Mdm2nuc, DNAdam in any panel listing the four components. If the order is different, it can be modified by using the green arrows displayed on the right of the list of nodes in Modeling Attributes panel, when no component or arc is selected.
Clicking on the Run button launches the simulation, i.e., the computation of the state transition graph (STG). A dialog indicates that the result is available, allowing to display the STG or to perform other actions on it. In the default level layout, the nodes with no incoming arc are placed at the top, whereas the nodes with no outgoing arc (i.e., stable states) are placed at the bottom. Stable states are further emphasized with a specific graphical attribute. In this new window, nodes can be rearranged, either manually or by selecting a predefined layout in the View menu. Outgoing transitions are displayed when selecting a state, as shown in Figure 6. Graphical settings can be modified after selecting the Style tab. Note that the scrolling menus propose various options, including path search functions, etc.
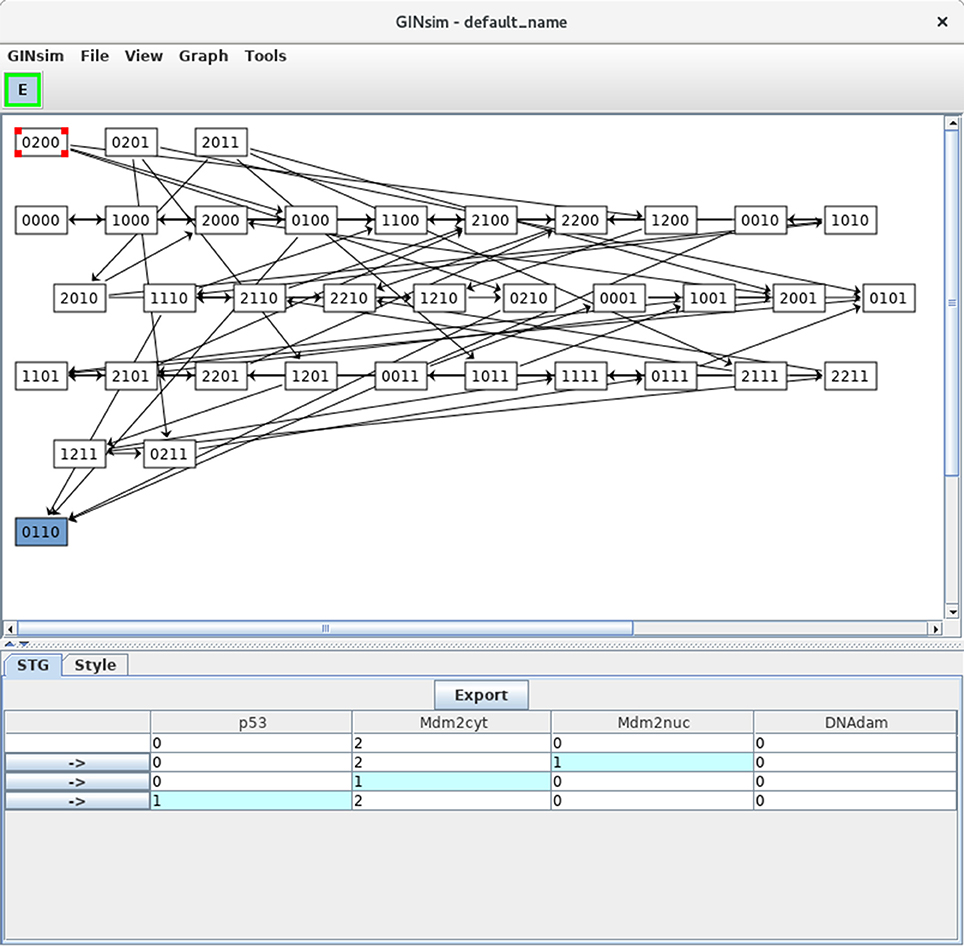
Figure 6. Asynchronous state transition graph for the p53-Mdm2 model. This STG has been generated with the simulation parameters shown in Figure 5. The unique stable state 0110 lays at the bottom. The selected state 0200 is shown in the STG tab, with its successors.
In Figure 6, the state 0200 (i.e., with high level of Mdm2cyt, and the other three nodes OFF) is selected, from which three unitary transitions are enabled by the logical rules (Table 3): increase of Mdm2nuc from 0 to 1, decrease of Mdm2cyt from 2 to 1, and increase of p53 from 0 to 1. The selected state and its three successor states are shown in the bottom panel. It is possible to follow a transition path by clicking on a rightwards arrow button in the bottom panel, which switches the selection to the corresponding state. When the selected state also connects to predecessors states, these are also shown, preceded by leftwards arrows.
Note that a unique stable state was obtained, 0110 (following the order defined above, this vector states that p53 = 0, Mdm2cyt = 1, Mdm2nuc = 1 and DNAdam = 0), which corresponds to the cell rest state (no p53, medium levels of cytoplasmic and nuclear Mdm2, no DNA damage).
Select the Compute stable states option in the Tools menu of the main window to verify that the unique stable state of this model is indeed 0110 (see Figure 7).
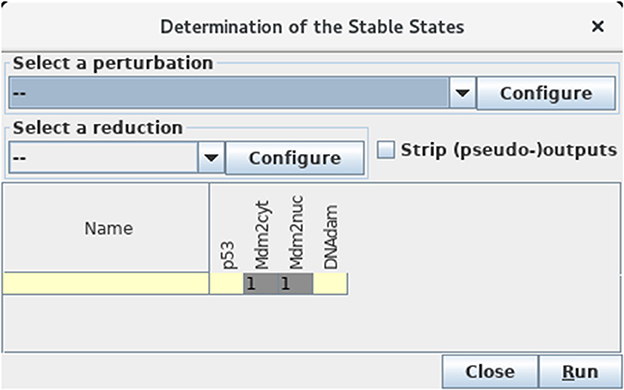
Figure 7. Determination of stable states. This window appears upon selection of Compute stable states with the Tools scrolling menu. After hitting the Run button, GINsim returns all stable states using an efficient algorithm. In the wild type case, we obtain a unique stable state 0110 as shown (yellow and gray cells denote levels 0 and 1, respectively).
This calculation uses an algorithm bypassing the construction of the STG, which is particularly useful for large models (for more details, see Naldi et al., 2007).
If another (or no) stable state is obtained, check carefully the maximum level of each node, the threshold associated with each interaction, as well as each logical rule, as there must be a mistake somewhere…
For comparison, let us now build the state transition graph of the model using the synchronous updating strategy. Select Run simulation in the Tools menu of the main window, then select the Synchronous option with the scrolling menu under Updating Mode in Figure 5, and launch the simulation by clicking on the Run button.
The resulting STG (after a manual improvement of the layout) is shown in shown in Figure 8. Naturally, the stable state 0110 is preserved (bottom left), but two cyclic attractors (bottom middle and right) are now obtained. Transitions representing single and multiple node updates are denoted by solid and dotted arcs, respectively.
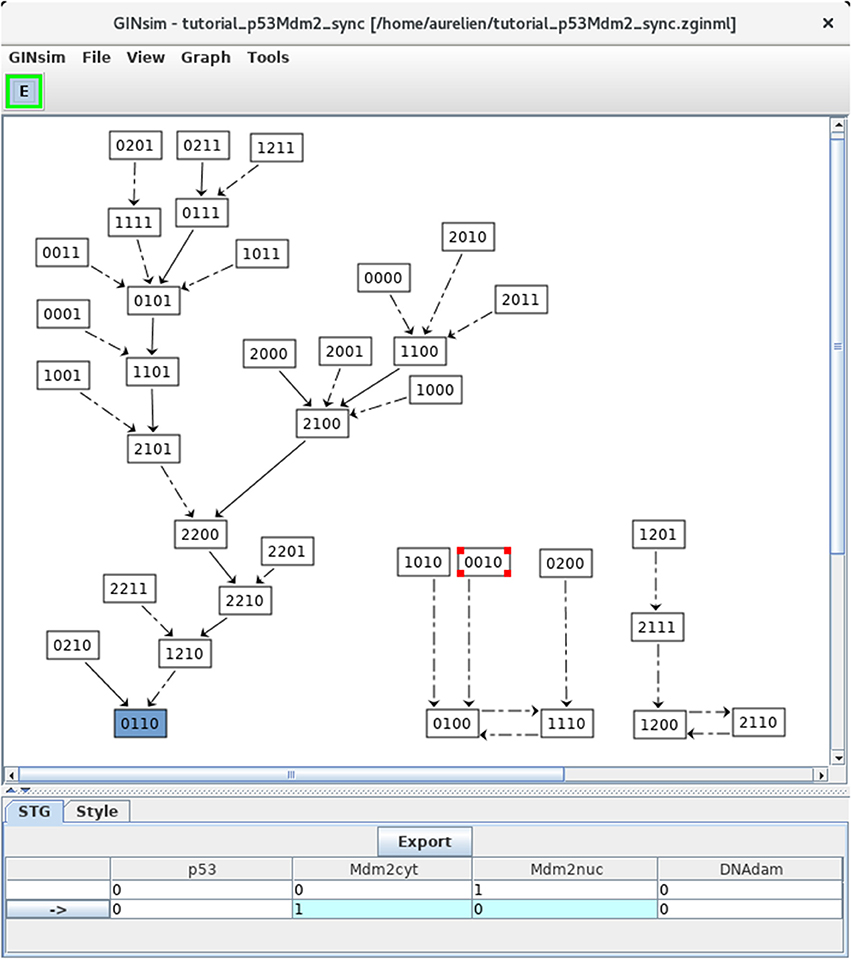
Figure 8. Synchronous state transition graph for the p53-Mdm2 model. This STG has been generated with the simulation parameters shown in Figure 5 (without specifying any initial state, but using the synchronous updating scheme). Note that the layout has been manually rearranged for sake of clarity. The STG is composed of three non connected subgraphs. On the left, we find back the resting stable state 0110, which can be reached from 26 other states. On the right, we see that the synchronous updating further generates two two-states cyclic attractors, which can be reached from three or two other states, respectively. Solid and dotted arrows denote single and multiple transitions, respectively.
Note that the selected state 0010 leads to the state 0100 through simultaneous changes of Mdm2cyt and Mdm2nuc, as shown in the bottom panel (blue cells).
When the size of the model increases, the state transition graph (STG) quickly becomes hard to visualize. To ease its analysis, a compression (or compaction) can be performed by grouping sets of states into hyper-nodes. The arcs connecting the resulting nodes then still correspond to state transitions. In particular, by lumping states that belong to the same strongly connected component (SCC, in the graph-theoretical sense), an acyclic graph is obtained. Interestingly, the resulting SCC graph preserves the reachability properties of the original graph. However, in many situations, the SCC graph results only in a moderate STG compression.
To increase STG compression and ease the interpretation of the dynamics, we have recently introduced another acyclic graph, called hierarchical transition graph, which further merges linear chains of states (in addition to cycles) into single nodes (Bérenguier et al., 2013). The resulting graph preserves the attractors and other important dynamical properties, but does not fully conserve reachability properties.
Selecting the corresponding option with the Construction Strategy scrolling menu allows to compress the dynamics by using the hierarchical transition graph (HTG) representation. Figure 9 shows the resulting HTG, with all other simulation parameters maintained as shown in Figure 5.
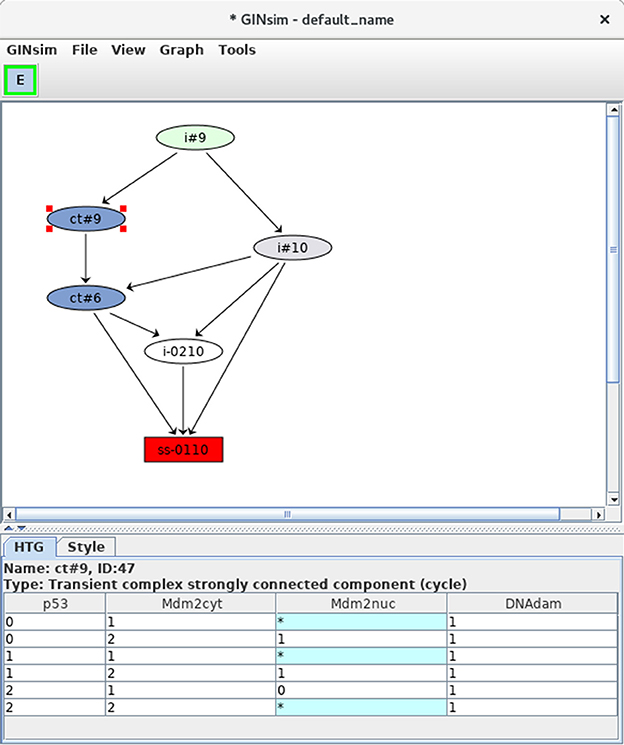
Figure 9. Hierarchical transition graph. The hierarchical transition graph for the complete asynchronous dynamics of the p53-Mdm2 model is shown. It has been obtained by selecting the construction of Hierarchical Transition Graph in the corresponding scrolling menu when launching the simulation. Note that the layout has been manually improved. The blue nodes correspond to the two non trivial strongly connected components of the STG, and the unique stable state is shown in red at the bottom. The blue node labeled by ct#9 has been selected; this transient cyclic component encompasses nine states from the STG (as indicated by the #9 in its name), which are listed in the bottom. The * denotes all possible values for the corresponding node. Hence the first row in the table listing the states encompassed by the hypernode ct#9 corresponds to two states: 0101 and 0111.
Although relatively modest in this case (six nodes in the HTG, to be compared with 36 nodes for the original STG), this compression can be much more impressive in cases with long alternative trajectories (see e.g., Bérenguier et al., 2013; Grieco et al., 2013). However, the computation of the HTG relies on that of the STG, with the compression done progressively. Hence, HTG computation may become intractable for large networks.
At the bottom of the HTG shown in Figure 9, note again the stable state 0110 (red box). In addition, two blue nodes representing strongly connected components can now be clearly seen, each labeled by ct, for cyclic transient, as both nodes are the sources of outgoing transitions.
The first of these cyclic components (ct#9) is selected and the corresponding states are listed in the bottom panel (where a star stands for all possible values for the corresponding node, which compresses the list of states). This cyclic component contains nine states, all with the DNAdam node set to 1, p53 oscillating between the values 0 and 2, Mdm2cyt oscillating between 1 and 2, and Mdm2nuc oscillating between 0 and 1. Hence, this cyclic component captures large oscillations of p53 in the presence of DNA damage.
The second cyclic component (ct#6) contains six states, with DNAdam now set to 0, with p53 and Mdm2cyt both oscillating between the values 1 and 2, and Mdm2nuc oscillating between the values 0 and 1. Hence, this cyclic component captures smaller transient p53 oscillations observed just after DNA repair.
In brief, starting from initial conditions with DNAdam = 1, the system first goes through an unspecified number of large p53 activity oscillations, followed by DNA repair (DNAdam taking the value 0) along with transient smaller p53 oscillations, and finally the return to the rest state 0110.
Several complementary analyses can be perfomed with GINsim. Hereafter, we illustrate three main functionalities: the encoding of perturbations, an algorithm enabling the analysis of the roles of regulatory circuits, along with a model reduction tool. Further information regarding GINsim functionalities can be found in the user manual and documentation available online.
Common perturbations are easily specified within the logical framework:
• A gene knock-down is specified by driving and constraining the level of the corresponding regulatory node to the value 0.
• Ectopic expression is specified by driving and constraining the level of the corresponding node to its highest value (or possibly to a range of values greater than zero, in the case of a multi-valued node).
• Multiple perturbations can be defined by combining several such constraints.
• More subtle perturbations can be defined by more sophisticated rewriting of node rules (i.e., to change the effect of a given regulatory arc).
Various perturbations can thus be defined to account for experimental observations or to generate predictions regarding the dynamical role of specific regulatory factors or interactions.
Define a mutant corresponding to an ectopic expression of DNAdam (see Figure 10). Such a perturbation can be encoded before the computation of stable states or of a state transition graph. Verify that the resting stable state 0110 is not stable anymore for this perturbation. Note the striking change of attractor for this perturbation, which now corresponds to ample oscillations of p53, along with oscillations of both nuclear and cytoplasmic Mdm2 forms in the presence of DNA damage.
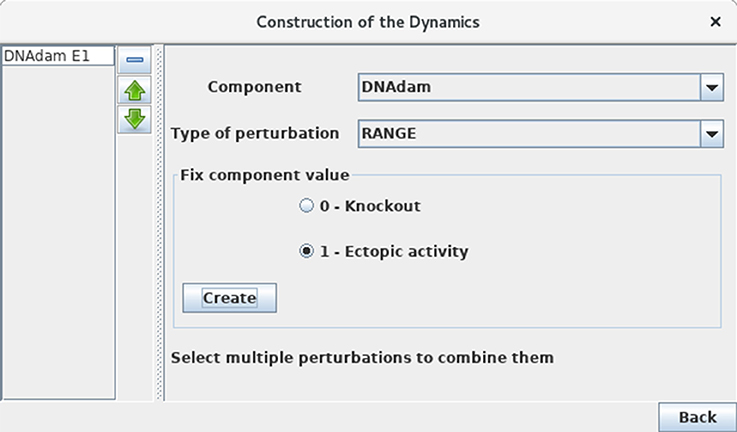
Figure 10. Perturbation specification. This window can be activated from the simulation launching window (Figure 5) and various other windows, including the Compute stable states window. It enables the specification of various kinds of model perturbations, including loss-of-function and gain-of-function mutants. The figure illustrates the specification of a simple blockade of the level of DNAdam to level 1.
Regulatory circuits are responsible for the emergence of dynamical properties, such as multistationarity or sustained oscillations (see Note 4). In this respect, GINsim implements specific algorithms to:
• Identify all the circuits of a regulatory graph (possibly considering constraints such as maximum length, consideration or exclusion of some nodes, etc.).
• Determine the functionality contexts of these circuits, using a computational method presented in Naldi et al. (2007).
To further identify and analyse the circuits of the model regulatory graph (see subsection 3.2), select the Analyse Circuits option of the Tools scrolling menu in the main window, then click on the Search Circuits button. Verify that the regulatory graph contains four circuits, among which three are functional (i.e., have a non-empty functionality context). For each functional circuit, one can verify its sign and functionality context (depending on the rules), by clicking on the Functionality Analysis button. As shown in Figure 11, the positive circuit defined by the cross inhibitions between p53 and Mdm2nuc is functional when Mdm2cyt = 1 and DNAdam = 0. Indeed, the inhibition of Mdm2nuc by p53 is not functional in the presence of DNAdam or of a high level of Mdm2cyt, or in the absence of Mdm2cyt.
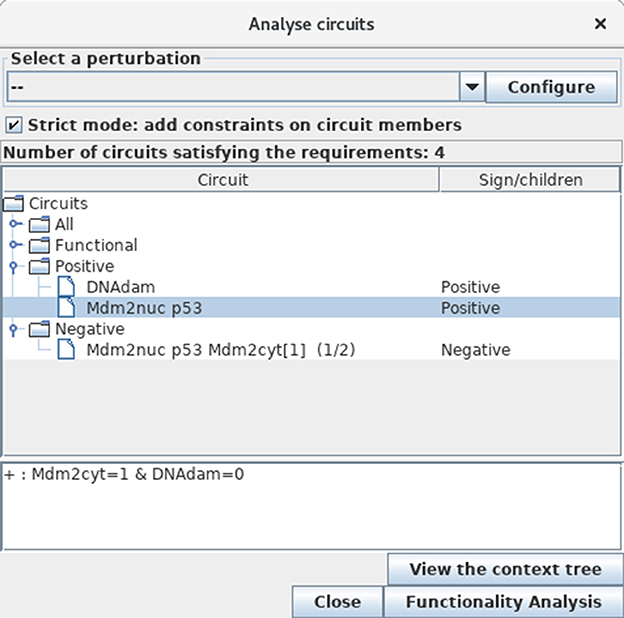
Figure 11. Circuit analysis for the p53-Mdm2 logical model. This window appears after first selecting the Analyse Circuits option of the Tools scrolling menu in the main window, then clicking on the Search Circuits button, and finally launching the Functionality Analysis option. Among the four circuits found in the regulatory graph, three are functional: one is negative, while the other two are positive. The selected circuit (involving p53 and Mdm2nuc) is positive and functional when the level of Mdm2cyt is medium (equal to 1) in the absence of DNA damage (DNAdam = 0).
When models increase in size, it quickly becomes difficult to cope with the size of the corresponding STG. One solution consists in simplifying or reducing the model before simulation. In this respect, GINsim implements a method to reduce a model on the fly, i.e., just before the simulation. The modeler can specify the nodes to be reduced, and the logical rules associated with their targets are then recomputed taking into account the (indirect) effects of their regulators. This construction of reduced models preserves crucial dynamical properties of the original model, including stable states and more complex attractors (Naldi et al., 2011).
Although our application is of limited size, we can still illustrate the use of GINsim model reduction functionality. Selecting the Reduce Model option in the Tools scrolling menu launches the reduction interface. Click on the + icon to define a reduction, then select the node Mdm2cyt for reduction, as shown in Figure 12. Clicking on the Run button generates a logical model encompassing only the three remaining nodes, where Mdm2nuc is the target of a dual interaction from p53. The logical rule associated with Mdm2nuc is consistently modified to take into account the former indirect effect of p53 through Mdm2cyt.
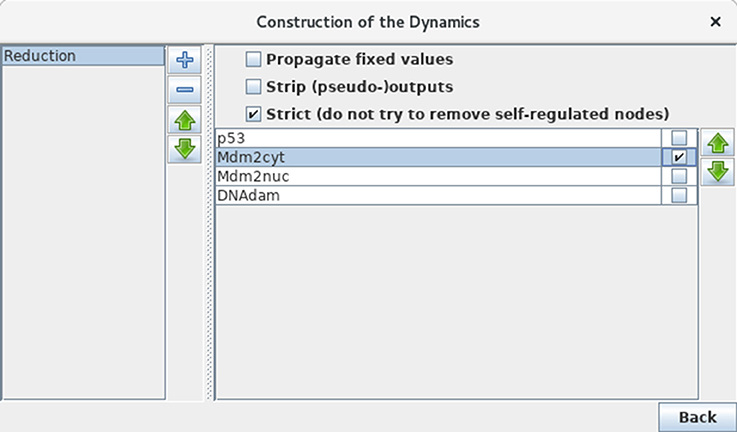
Figure 12. Model reduction. This window appears following the selection of Reduce model from the Tools scrolling menu in the main GINsim window. Here, only Mdm2cyt has been selected for reduction. By hitting the Run Button, a reduced model is generated, provided that no self-regulated node is affected. Alternatively, one can close the window after the definition of one or several reduction(s) (the + button on the left enable to create new reductions) and then select a predefined reduction directly when performing simulations or other kinds of analyses.
Now that a reduction has been defined, it can be selected when launching a simulation or computing stable states, without generating the reduced graph. Perform a complete asynchronous simulation to get the full state transition graph and verify that the number of states is now lower by a factor of three (12 states instead of 36) compared to Figure 6. Compute the HTG keeping the same parameter settings (asynchronous updating and full state space as initial condition). Although very much compressed, the resulting STG still captures the two kinds of p53 transient oscillatory behavior, ample in presence of DNA damage, smaller after DNA repair.
The online documentation includes a troubleshooting page (see http://doc.ginsim.org) providing some solutions to common problems. The graphical interface can have some refresh issues after long or complex modeling sessions. Such issues are usually resolved after saving the model and restarting the GINsim software. For other issues, we encourage users to send a message describing their problem to the GINsim forum or directly to the GINsim team (see http://ginsim.org/contact). Because some issues are difficult to reproduce, the user should provide log traces (using the GINsim/support/export log files menu option), after launching GINsim from the command line to catch additional error messages.
A few hints to solve issues that may arise in the course of this tutorial are provided below.
Some nodes can be defined as input nodes using a check-box in the node property panel. These input nodes can have neither incoming interactions nor regulatory rules. Indeed, input nodes have an implicit rule specifying that they maintain their current activity levels (i.e., they are maintained constant). Therefore, all regulatory interactions and rules must be removed before setting a node as an input. Likewise, the input status must be removed before adding any new regulator or rule. The model p53-Mdm2 has no input: the input check-box should be unselected for all the nodes.
In case of unexpected dynamical results (e.g., stable states, trajectories, etc.), verify successively the structure of the regulatory graph, the maximal levels of the nodes, the thresholds of the regulatory interactions with multi-valued sources and finally the regulatory rules. GINsim further provides a tool to Compute interaction functionality, which facilitates the identification of inconsistencies between the structure of the regulatory graph and the regulatory rules (see Note 5). To delete an invalid logical formula, select it (without editing it) and use the delete key or the contextual menu.
The logical formalism is particularly useful to model regulatory networks for which precise quantitative information is barely available, or yet to have a first glance of the dynamical properties of a complex model.
For this protocol, we have considered a network comprising four regulatory factors, and we have followed the different steps enabling the delineation of a consistent logical model. Despite its limited size, this model yields relatively complex dynamics, including several transient oscillatory patterns and a stable state. It further served as a reference to illustrate advanced functions, such as model reduction or regulatory circuit analysis.
Large signaling networks have been handled with GINsim (e.g., Calzone et al., 2010; Naldi et al., 2010; Abou-Jaoudé et al., 2015), in which input nodes denote external signals, which are not regulated and often maintained constant. Such Input nodes can be specified as such in GINsim to enforce the maintenance of the levels specified at initial states. As the reduction of input and output nodes or cascades have a marginal impact on the dynamics (Abou-Jaoudé et al., 2016), such reductions are facilitated in GINsim.
Furthermore, a novel functionality Assess Attractor Reachability in the Tool menu enables to evaluate the reachability of attractors based on stochastic simulation algorithms (for more details, see Mendes et al., 2014).
Taking advantage of the multiple export formats supported by GINsim, it is also possible to use complementary tools, including stochastic simulation software (e.g., MaBoSS, see Stoll et al., 2017), model checking tools (e.g., NuSMV, see Abou-Jaoudé et al., 2015; Abou-Jaoudé et al., 2016; Traynard et al., 2016), or yet various graph visualization and analysis packages (see Note 6 for a list of export options).
As mentioned in the introduction, various logical models for different cellular processes have been proposed during the last decades, many of them available in the repository included along with GINsim on the dedicated website (http://ginsim.org). The interested reader can thus download the model of his choice and play with it, reproduce some of the results reported in the corresponding publication, or modify and extend it according to his own research aims.
1. Logical parameters constitute an alternative way of defining regulatory rules. For each node, each combination of incoming interactions then defines a logical parameter. This includes the situation in the absence of any specific activation or inhibition, or basal level. As a large fraction of the parameters are usually set to zero, this is the default value in GINsim (i.e., any parameter lacking an explicitly assigned value is set to 0). Consult the online documentation for details on parameters definition (http://doc.ginsim.org/lrg-parameters.html).
2. Transitions between states of the state transition graphs amount to the update of one (in the asynchronous case) or several (in the synchronous case) regulatory nodes. GINsim further support a complete updating mode, considering all possible (single or multiple) transitions enabled by the rules, as well as a sequential updating mode, which updates nodes sequentially following the predefined order node. In any case, the update (increase or decrease) of a node is unitary (current value +1 or −1). Obviously, this remark applies only for multi-valued nodes (for which the maximal level is greater than 1).
3. Priority classes allow to refine the updating schemes applied to construct the state transition graphs (Fauré et al., 2006). GINsim users can group nodes into different classes and assign a priority rank to each of them. In case of concurrent updating transitions (i.e., calls for level changes for several regulatory nodes in the same state), GINsim updates the node(s) belonging to the class with the highest ranking. For each priority class, the user can further specify the desired updating assumption, which then determines the treatment of concurrent transition calls inside that class. When several classes have the same rank, concurrent transitions are treated under an asynchronous assumption (no priority).
4. A regulatory circuit is defined as a sequence of interactions forming a simple closed directed path. The sign of a circuit is given by the product of the signs of its interactions. Consequently, a circuit is positive if it has an even number of inhibitions, it is negative otherwise. R. Thomas proposed that positive circuits are necessary to generate multistationarity, whereas negative circuits are necessary to generate stable oscillations (see Thieffry, 2007 and references therein). External regulators might prevent the functioning of a circuit imbedded in a more complex network. Naldi et al. (2007) proposed a method to determine the functionality context of a circuit in terms of constraints on the levels of its external regulator. A circuit functionality context can be interpreted as the part of the state space where the circuit is functional, i.e., generates the expected dynamical property (Comet et al., 2013).
5. The Compute interaction functionality option of the Tools scrolling menu allows to check if the signs of the interactions (graphically defined) comply with the regulatory rules. Inconsistencies arise when, for instance, a positive interaction has been drawn, while the regulatory rule of the target node defines an inhibitory effect or no effect at all. This is a convenient tool to check model inconsistencies. Note however that such inconsistencies do not prevent (inconsistent) model simulation or analysis.
6. GINsim allows the user to export logical regulatory graphs (or state transition graphs) toward various formats, facilitating the use of other software:
• SBML-qual, the qualitative extension of the popular model exchange format (Chaouiya et al., 2013).
• MaBoSS, a C++ software for simulating continuous/discrete time Markov processes, applied on a Boolean networks (https://maboss.curie.fr/).
• BoolSim (http://www.vital-it.ch/software/genYsis/).
• GNA, a software for the piecewise linear modeling of regulatory networks (http://ibis.inrialpes.fr/article122.html).
• NuSMV, a symbolic model-checking tool (http://nusmv.fbk.eu/).
• Integrated Net Analyzer (INA) supporting the analysis of Place/Transition Nets (Petri Nets) and Colored Petri nets (http://www2.informatik.hu-berlin.de/~starke/ina.html).
• Snoopy, a tool to design and animate hierarchical graphs, among others Petri nets (http://www-dssz.informatik.tu-cottbus.de/DSSZ/Software/Snoopy).
• Graphviz, an open source graph visualization software offering main graph layout programs (http://www.graphviz.org/).
• Cytoscape, a popular open source software platform for visualizing molecular interaction networks (http://www.cytoscape.org/).
• Scalable Vector Graphics (SVG) format, an XML standard for describing two-dimensional graphics (http://www.w3.org/Graphics/SVG/).
While AN and PM have been the main developers of GINsim over the last years, all authors of this manuscript have taken part in various practical tutorials introducing the usage of GINsim to biologists, which served as a basis for the preparation of this method article. All authors have further participated in the writing of the manuscript and in the preparation of the Figures. AN, CH, WA-J, and PM should be considered as co-first authors, and CC and DT as co-last authors. All authors reviewed the content of this article and agreed to endorse it.
CC and PM acknowledge support from the Fundação para a Ciência e a Tecnologia, through grants PTDC/BEX-BCB/0772/2014 and PTDC/EEI-CTP/2914/2014. DT acknowledges support from the French Plan Cancer, in the context of the projects CoMET (2014–2017) and SYSTAIM (2015–2019), as well as from the French Agence Nationale pour la Recherche, in the context of the project SCAPIN [ANR-15-CE15-0006-01].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors acknowledge numerous constructive comments from GINsim users over the years, in particular insightful feedback from Laurence Calzone, Samuel Collombet, Karla Corral, Adrien Fauré, Swann Floc'hlay, Asmund Floback, Anna Niarakis, Elisabeth Remy, Otoniel Rodriguez, and Gautier Stoll.
Abou-Jaoudé, W., Monteiro, P. T., Naldi, A., Grandclaudon, M., Soumelis, V., Chaouiya, C., et al. (2015). Model checking to assess T-Helper cell plasticity. Front. Bioeng. Biotechnol. 2:86. doi: 10.3389/fbioe.2014.00086
Abou-Jaoudé, W., Ouattara, D. A., and Kaufman, M. (2009). From structure to dynamics: frequency tuning in the p53-Mdm2 network I. logical approach. J. Theor. Biol. 258, 561–577. doi: 10.1016/j.jtbi.2009.02.005
Abou-Jaoudé, W., Traynard, P., Monteiro, P. T., Saez-Rodriguez, J., Helikar, T., Thieffry, D., et al. (2016). Logical modeling and dynamical analysis of cellular networks. Front. Genet. 7:94. doi: 10.3389/fgene.2016.00094
Azpeitia, E., Davila-Velderrain, J., Villarreal, C., and Alvarez-Buylla, E. R. (2014). “Gene regulatory network models for floral organ determination,” in Flower Development, eds J. L. Riechmann and F. Wellmer (New York, NY: Humana Press), 441–469.
Barak, Y., Juven, T., Haffner, R., and Oren, M. (1993). mdm2 expression is induced by wild type p53 activity. EMBO J. 12, 461–468.
Bérenguier, D., Chaouiya, C., Monteiro, P. T., Naldi, A., Remy, E., and Thieffry, D. (2013). Dynamical modeling and analysis of large cellular regulatory networks. Chaos 23:025114. doi: 10.1063/1.4809783
Brooks, C. L., and Gu, W. (2006). p53 ubiquitination: Mdm2 and beyond. Mol. Cell 21, 307–315. doi: 10.1016/j.molcel.2006.01.020
Calzone, L., Tournier, L., Fourquet, S., Thieffry, D., Zhivotovsky, B., Barillot, E., et al. (2010). Mathematical modelling of cell-fate decision in response to death receptor engagement. PLoS Comput. Biol. 6:e1000702. doi: 10.1371/journal.pcbi.1000702
Chaouiya, C., Bérenguier, D., Keating, S. M., Naldi, A., van Iersel, M. P., Rodriguez, N., et al. (2013). SBML qualitative models: a model representation format and infrastructure to foster interactions between qualitative modelling formalisms and tools. BMC Syst. Biol. 7:135. doi: 10.1186/1752-0509-7-135
Choi, M., Shi, J., Jung, S. H., Chen, X., and Cho, K. H. (2012). Attractor landscape analysis reveals feedback loops in the p53 network that control the cellular response to DNA damage. Sci. Signal. 5:ra83. doi: 10.1126/scisignal.2003363
Ciliberto, A., Novak, B., and Tyson, J. J. (2005). Steady states and oscillations in the p53/Mdm2 network. Cell Cycle 4, 488–493. doi: 10.4161/cc.4.3.1548
Collombet, S., van Oevelen, C., Sardina Ortega, J. L., Abou-Jaoudé, W., Di Stefano, B., Thomas-Chollier, M., et al. (2017). Logical modeling of lymphoid and myeloid cell specification and transdifferentiation. Proc. Natl. Acad. Sci. U.S.A. 114, 5792–5799. doi: 10.1073/pnas.1610622114
Comet, J.-P., Noual, M., Richard, A., Aracena, J., Calzone, L., Demongeot, J., et al. (2013). On circuit functionality in boolean networks. Bull. Math. Biol. 75, 906–919. doi: 10.1007/s11538-013-9829-2
Coolen, M., Thieffry, D., Drivenes, Ø., Becker, T. S., and Bally-Cuif, L. (2012). miR-9 controls the timing of neurogenesis through the direct inhibition of antagonistic factors. Dev. Cell 22, 1052–1064. doi: 10.1016/j.devcel.2012.03.003
Coutts, A. S., Boulahbel, H., Graham, A., and La Thangue, N. B. (2007). Mdm2 targets the p53 transcription cofactor JMY for degradation. EMBO Reports 8, 84–90. doi: 10.1038/sj.embor.7400855
Fauré, A., Naldi, A., Chaouiya, C., and Thieffry, D. (2006). Dynamical analysis of a generic boolean model for the control of the mammalian cell cycle. Bioinformatics 22, 124–31. doi: 10.1093/bioinformatics/btl210
Fauré, A., Naldi, A., Lopez, F., Chaouiya, C., Ciliberto, A., and Thieffry, D. (2009). Modular logical modelling of the budding yeast cell cycle. Mol. Biosyst. 5, 1787–1796. doi: 10.1039/b910101m
Fauré, A., Vreede, B. M., Sucena, E., and Chaouiya, C. (2014). A discrete model of drosophila eggshell patterning reveals cell-autonomous and juxtacrine effects. PLoS Comput. Biol. 10:e1003527. doi: 10.1371/journal.pcbi.1003527
Flobak, Å., Baudot, A., Remy, E., Thommesen, L., Thieffry, D., Kuiper, M., et al. (2015). Discovery of drug synergies in gastric cancer cells predicted by logical modeling. PLoS Comput. Biol. 11:e1004426. doi: 10.1371/journal.pcbi.1004426
Gatz, S. A., and Wiesmüller, L. (2006). p53 in recombination and repair. Cell Death Differ. 13, 1003–1016. doi: 10.1038/sj.cdd.4401903
González, A., Chaouiya, C., and Thieffry, D. (2006). Dynamical analysis of the regulatory network defining the dorsal-ventral boundary of the drosophila wing imaginal disc. Genetics 174, 1625–1634. doi: 10.1534/genetics.106.061218
González, A., Chaouiya, C., and Thieffry, D. (2008). Logical modelling of the role of the Hh pathway in the patterning of the Drosophila wing disc. Bioinformatics 24, i234–i240. doi: 10.1093/bioinformatics/btn266
Grieco, L., Calzone, L., Bernard-Pierrot, I., Radvanyi, F., Kahn-Perlès, B., and Thieffry, D. (2013). Integrative modelling of the influence of MAPK network on cancer cell fate decision. PLoS Comput. Biol. 9:21003286. doi: 10.1371/annotation/90e5e4be-952b-42b8-b56d-46baae3479ed
Iwamoto, K., Hamada, H., Eguchi, Y., and Okamoto, M. (2014). Stochasticity of intranuclear biochemical reaction processes controls the final decision of cell fate associated with DNA damage. PLoS ONE 9:e101333. doi: 10.1371/journal.pone.0101333
Le Novère, N., Hucka, M., Mi, H., Moodie, S., Schreiber, F., Sorokin, A., et al. (2009). The systems biology graphical notation. Nat. Biotechnol. 27, 735–741. doi: 10.1038/nbt.1558
Martinez-Sanchez, M. E., Mendoza, L., Villarreal, C., and Alvarez-Buylla, E. R. (2015). A minimal regulatory network of extrinsic and intrinsic factors recovers observed patterns of CD4+ T cell differentiation and plasticity. PLoS Comput. Biol. 11:e1004324. doi: 10.1371/journal.pcbi.1004324
Mayo, L. D., and Donner, D. B. (2002). The PTEN, Mdm2, p53 tumor suppressor-oncoprotein network. Trends Biochem. Sci. 27, 462–467. doi: 10.1016/S0968-0004(02)02166-7
Mbodj, A., Gustafson, E. H., Ciglar, L., Junion, G., Gonzalez, A., Girardot, C., et al. (2016). Qualitative dynamical modelling can formally explain mesoderm specification and predict novel developmental phenotypes. PLOS Comput. Biol., 12:e1005073. doi: 10.1371/journal.pcbi.1005073
Mendoza, L., and Méndez, A. (2015). A dynamical model of the regulatory network controlling lymphopoiesis. Biosystems 137, 26–33. doi: 10.1016/j.biosystems.2015.09.004
Mendes, N. D., Monteiro, P. T., Carneiro, J., Remy, E., and Chaouiya, C. (2014). Quantification of reachable attractors in asynchronous discrete dynamics. arXiv:1411.3539
Mendoza, L., Thieffry, D., and Alvarez-Buylla, E. R. (1999). Genetic control of flower morphogenesis in Arabidopsis thaliana: a logical analysis. Bioinformatics 15, 593–606. doi: 10.1093/bioinformatics/15.7.593
Naldi, A., Carneiro, J., Chaouiya, C., and Thieffry, D. (2010). Diversity and plasticity of th cell types predicted from regulatory network modelling. PLoS Comput. Biol. 6:e1000912. doi: 10.1371/journal.pcbi.1000912
Naldi, A., Monteiro, P., Müssel, C., Kestler, H. A., Thieffry, D., Xenarios, I., et al. (2015). Cooperative development of logical modelling standards and tools with CoLoMoTo. Bioinformatics 31, 1154–1159. doi: 10.1093/bioinformatics/btv013
Naldi, A., Remy, E., Thieffry, D., and Chaouiya, C. (2011). Dynamically consistent reduction of logical regulatory graphs. Theor. Comput. Sci. 412, 2207–2218. doi: 10.1016/j.tcs.2010.10.021
Naldi, A., Thieffry, D., and Chaouiya, C. (2007). Decision diagrams for the representation of logical models of regulatory networks. Lect. Notes Comput. Sci. 4695, 233–247. doi: 10.1007/978-3-540-75140-3_16
Oliner, J. D., Pietenpol, J. A., Thiagalingam, S., Gyuris, J., Kinzler, K. W., and Vogelstein, B. (1993). Oncoprotein MDM2 conceals the activation domain of tumour suppressor p53. Nature 362, 857–860. doi: 10.1038/362857a0
Ouattara, D. A., Abou-Jaoude, W., and Kaufman, M. (2010). From structure to dynamics: frequency tuning in the p53-Mdm2 network. II Differential and stochastic approaches. J. Theor. Biol. 264, 1177–1189. doi: 10.1016/j.jtbi.2010.03.031
Puszynski, K., Hat, B., and Lipniacki, T. (2008). Oscillations and bistability in the stochastic model of p53 regulation. J. Theor. Biol. 254, 452–465. doi: 10.1016/j.jtbi.2008.05.039
Remy, E., Rebouissou, S., Chaouiya, C., Zinovyev, A., Radvanyi, F., and Calzone, L. (2015). A modeling approach to explain mutually exclusive and co-occurring genetic alterations in ladder tumorigenesis. Cancer Res. 75, 4042–4052. doi: 10.1158/0008-5472.CAN-15-0602
Sahin, O., Fröhlich, H., Löbke, C., Korf, U., Burmester, S., Majety, M., et al. (2009). Modeling ERBB receptor-regulated G1/S transition to find targets for de novo trastuzumab resistance. BMC Syst. Biol. 3:1. doi: 10.1186/1752-0509-3-1
Sánchez, L., Chaouiya, C., and Thieffry, D. (2008). Segmenting the fly embryo: a logical analysis of the segment polarity cross-regulatory module. Int. J. Dev. Biol. 52, 1059–1075. doi: 10.1387/ijdb.072439ls
Sánchez, L., and Thieffry, D. (2001). A logical analysis of the drosophila gap-gene system. J. Theor. Biol. 211, 115–141. doi: 10.1006/jtbi.2001.2335
Sánchez, L., and Thieffry, D. (2003). Segmenting the fly embryo: a logical analysis of the pair-rule cross-regulatory module. J. Theor. Biol. 224, 517–537. doi: 10.1016/S0022-5193(03)00201-7
Stoll, G., Caron, B., Viara, E., Dugourd, A., Zinovyev, A., Naldi, A., et al. (2017). MaBoSS 2.0: an environment for stochastic Boolean modeling. Bioinformatics 33, 2226–2228. doi: 10.1093/bioinformatics/btx123
Sun, T., and Cui, J. (2014). A plausible model for bimodal p53 switch in DNA damage response. FEBS Lett. 588, 815–821. doi: 10.1016/j.febslet.2014.01.044
Thieffry, D. (2007). Dynamical roles of biological regulatory circuits. Brief. Bioinform. 8, 220–225. doi: 10.1093/bib/bbm028
Thieffry, D., and Thomas, R. (1995). Dynamical behaviour of biological regulatory networks, II. Immunity control in bacteriophage lambda. Bull. Math. Biol. 57, 277–297.
Thomas, R. (1991). Regulatory networks seen as asynchronous automata: a logical description. J. Theor. Biol. 153, 1–23. doi: 10.1016/S0022-5193(05)80350-9
Thomas, R., Thieffry, D., and Kaufman, M. (1995). Dynamical behaviour of biological regulatory networks i. biological role of feedback loops and practical use of the concept of the loop-characteristic state. Bull. Math. Biol. 57, 247–276. doi: 10.1007/BF02460618
Traynard, P., Fauré, A., Fages, F., and Thieffry, D. (2016). Logical model specification aided by model-checking techniques: application to the mammalian cell cycle regulation. Bioinformatics 32, i772–i780. doi: 10.1093/bioinformatics/btw457
Vogelstein, B., Lane, D., and Levine, A. J. (2000). Surfing the p53 network. Nature 408, 307–310. doi: 10.1038/35042675
Keywords: regulatory network, logical model, discrete dynamics, regulatory circuit, p53-Mdm2 network
Citation: Naldi A, Hernandez C, Abou-Jaoudé W, Monteiro PT, Chaouiya C and Thieffry D (2018) Logical Modeling and Analysis of Cellular Regulatory Networks With GINsim 3.0. Front. Physiol. 9:646. doi: 10.3389/fphys.2018.00646
Received: 05 March 2018; Accepted: 11 May 2018;
Published: 19 June 2018.
Edited by:
Theodore J. Perkins, University of Ottawa, CanadaReviewed by:
Elena S. Dimitrova, Clemson University, United StatesCopyright © 2018 Naldi, Hernandez, Abou-Jaoudé, Monteiro, Chaouiya and Thieffry. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aurélien Naldi, YXVyZWxpZW4ubmFsZGlAZW5zLmZy
Claudine Chaouiya, Y2hhb3VpeWFAaWdjLmd1bGJlbmtpYW4ucHQ=
Denis Thieffry, dGhpZWZmcnlAZW5zLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.