 Jinping Liu
Jinping Liu Lirong Chang2†
Lirong Chang2† Hui Li
Hui Li Yan Wu
Yan Wu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Neurosci. , 08 February 2019
Sec. Neurodegeneration
Volume 13 - 2019 | https://doi.org/10.3389/fnins.2019.00043
This article is part of the Research Topic Forms of Degeneration and Dysregulation in the Central Nervous System View all 8 articles
In Alzheimer’s disease (AD), early synaptic dysfunction is associated with the increased oligomeric amyloid-beta peptide, which causes NMDAR-dependent synaptic depression and spine elimination. Memantine, low-affinity NMDAR channel blocker, has been used in the treatment of moderate to severe AD. However, clear evidence is still deficient in demonstrating the underlying mechanisms and a relationship between NMDARs dysfunction and AD. This review focuses on not only changes in expression of different NMDAR subunits, but also some unconventional modes of NMDAR action.
Alzheimer’s disease (AD) is an age-related neurodegenerative disease. Clinically, this disorder is characterized by global cognitive dysfunction, especially memory loss, behavior and personality changes. AD progression has been associated with a gradual damage in function and structure in the hippocampus and neocortex, the vulnerable brain areas used for memory and cognition (Mota et al., 2014). The neuropathologic hallmarks of AD include extracellular amyloid plaques, being composed of the amyloid beta (Aβ) peptide and intraneuronal neurofibrillary tangles (NFTs), consisting of abnormally hyperphosphorylated tau protein (Ferreira et al., 2010). Nevertheless, damage and destruction of synapses seem to better correlate with memory loss than histopathologic markers. Synapse loss can be caused by the failure of live neurons to maintain functional axons and dendrites or by neuron death (Bloom, 2014; Avila et al., 2017; Morris et al., 2018).
Synaptic dysfunction may be due to perturbed synaptic Ca2+ handling in response to over activation of glutamate receptors, namely, the N-methyl-D-aspartate receptors (NMDARs) (Mota et al., 2014). Glutamate is the primary excitatory neurotransmitter in the brain, acting at ionotropic and metabotropic glutamate receptors. Ionotropic glutamate receptors (iGluRs), responsible for fast neuronal communication at excitatory synapses, comprise three subfamilies: α-amino-3-hydroxy-5-methyl-4-isoxasolepropionic acid (AMPA) receptors, kainate receptors, and NMDARs (Traynelis et al., 2010). However, excessive stimulation of glutamatergic signaling results in excitotoxicity (Rothman and Olney, 1986). Besides acute effects, many studies indicate a role for glutamate excitotoxicity in delayed slowly evolving neurodegeneration (Choi, 1988; Epstein et al., 1994). The toxicity is principally mediated by excessive Ca2+ entry, primarily through NMDARs (Choi, 1987; Tymianski et al., 1993), since NMDARs have a much higher permeability for calcium ions compared to other iGluRs (Choi, 1992; Wang and Reddy, 2017).
N-methyl-D-aspartate receptor plays a pivotal role in the synaptic transmission and synaptic plasticity thought to underlie learning and memory, which is not only central to the development and function of the nervous system, but also to neurotoxicity. NMDAR activation has been recently implicated in AD related to synaptic dysfunction (Paoletti et al., 2013; Kodis et al., 2018). The pathological level of Ca2+ signaling leads to gradual loss of synaptic function and ultimate neuronal cell death, which correlates clinically with the progressive decline in cognition/memory and the development of pathological neural anatomy seen in AD patients. And this, in turn, rationalizes the clinical trial of memantine, an NMDAR antagonist, as a symptomatological and neuroprotective treatment for AD (Danysz and Parsons, 2003; Wenk, 2006; Wang and Reddy, 2017). Memantine, a non-competitive NMDA receptor antagonist, is approved for use in moderate to severe AD. It has been widely prescribed to provide symptomatic relief and enhance life quality in AD, even if it did not improve excessive agitation (Fox et al., 2012), and hippocampal or total brain atrophy (Wilkinson et al., 2012). However, in the brain areas mainly affected in AD, NMDARs are mainly composed by GluN2A and GluN2B subunits. Furthermore, taking into account the fact that extrasynaptic GluN2B-containing NMDARs have been associated with excitotoxicity in AD (Hardingham and Bading, 2010), the use of a selective GluN2B subunit antagonists might be an interesting strategy to prevent synaptic dysfunction in AD.
N-methyl-D-aspartate receptor is a ligand of glutamate, which is the primary excitatory neurotransmitter in the human brain. Most NMDAR subtypes are unique in that their opening requires the coincidence of both presynaptic glutamate release and a strong postsynaptic membrane depolarization to relieve Mg2+ block of the channel (Mayer et al., 1984; Nowak et al., 1984). NMDARs are permeable to Na+, K+, and high permeability to Ca2+, which acts as a second messenger to modify synapses (Lynch et al., 1983). NMDARs are essential mediators of brain plasticity and are capable of converting specific patterns of neuronal activity into long-term changes in synapse structure and function that are thought to underlie higher cognitive functions (Traynelis et al., 2010). Activation of NMDARs leads to cytosolic free intracellular calcium ([Ca2+]i) increase (MacDermott et al., 1986), required for long-term potentiation (LTP) and long-term depression (LTD) (Muller et al., 2009), and, more generally, for synaptic plasticity (MacDonald et al., 2006; Lau et al., 2009). NMDARs are also thought to be involved in a process called excitotoxicity. Abnormal NMDAR activity is associated with seizure, ischemic stroke (Choi et al., 1988; Villmann and Becker, 2007), and neurodegenerative disorders (Benarroch, 2011), such as Alzheimer’s (Wenk, 2006), Huntington’s (Fan and Raymond, 2007), and Parkinson’s disease (Bonuccelli and Del Dotto, 2006).
N-methyl-D-aspartate receptors are also expressed across a wide spectrum of non-neuronal cells, including central and peripheral glial cells, endothelium, bone, kidney, pancreas, and others (Hogan-Cann and Anderson, 2016). Astrocytes express functional NMDARs capable of responding to neuronal glutamatergic input (Dzamba et al., 2013) and neuroinflammatory processes (Sofroniew, 2009; Ting et al., 2009). Potential roles for endothelial NMDARs is in the area of blood–brain barrier (BBB) function. Uncontrolled glutamate levels in brain are toxic to neurons, damage endothelial function and disrupt BBB integrity (András et al., 2007; Basuroy et al., 2013). Osteoblast NMDARs stimulated precursor differentiation and contributed to increased bone mineralization, leading to bone matrix deposition (Hinoi et al., 2003; Li et al., 2011). Renal NMDAR activity stimulated vasodilation in the glomerulus, thereby influencing renal blood flow, filtration, and reabsorption in the proximal tubule (Deng and Thomson, 2009; Anderson et al., 2011; Sproul et al., 2011). In the pancreas, NMDARs are expressed by insulin-producing islet β cells, and influence β cell function and survival (Inagaki et al., 1995; Molnár et al., 1995; Marquard et al., 2015). In the lung, airway smooth muscle cells express NMDARs, which may be involved in inflammatory bronchiole hyper-reactivity (Antošová and Strapková, 2013; Anaparti et al., 2015).
Seven different NMDAR subunits have been identified: the GluN1 subunit, four distinct GluN2 subunits (GluN2A, GluN2B, GluN2C, and GluN2D), and a pair of GluN3 subunits (GluN3A and GluN3B) (Paoletti, 2011). Functional NMDARs are heterotetramers composed of two glycine or D-serine-binding GluN1 subunits and two glutamate-binding GluN2 (GluN2A-D) subunits or, in some cases, glycine-binding GluN3 (GluN3A/B) subunits (Köhr, 2006). All NMDAR subunits share a common membrane topology that contain four discrete domains (Lee et al., 2014; Karakas and Furukawa, 2014): The extracellular amino-terminal domain (ATD) is involved in subunit assembly and allosteric regulation, and contributes to control of ion channel open probability and deactivation speeds. The extracellular ligand-binding domain (LBD) is formed by two discontinuous segments (S1 and S2), and binds agonists and antagonists to control ion channel opening. The transmembrane domain (TMD) contains three transmembrane segments (M1, M3, and M4) and a re-entrant pore loop (M2) which is part of the channel pore, containing a critical asparagine residue that determines calcium permeability of the channel and mediates the magnesium blockade (Paoletti et al., 2013; Karakas and Furukawa, 2014). The intracellular carboxyl-terminal domain (CTD) interacts with multiple cytosolic proteins (Sanz-Clemente et al., 2012). GluN2 LBDs contain the glutamate binding site, while GluN1 (or GluN3) LBDs contain the Glycine/D-serine coagonist binding site (Hackos and Hanson, 2017). In general, activation of NMDARs containing GluN1/GluN2 requires binding of both glycine and glutamate at the extracellular LBDs in addition to release of magnesium block by membrane depolarization at the TMD (Regan et al., 2015). Positively charged residues on the surface of the GluN2A LBD assist glutamate binding via a ‘guided-diffusion’ mechanism. On the other hand, Glycine binds to the GluN1 LBD via an ‘unguided-diffusion’ mechanism, whereby glycine finds its binding site primarily by random thermal fluctuations (Yu and Lau, 2018). A recent study show, through cryoelectron microscopy, that the presence of the exon 5 motif, at the ATD of the GluN1 subunit, alters the local architecture of heterotetrameric GluN1/GluN2 NMDA receptors and creates contacts with the LBDs of the GluN1 and GluN2 subunits. The unique interactions established by the exon 5 motif are essential to the stability of the ATD/LBD and LBD/LBD interfaces that are critically involved in controlling proton sensitivity and deactivation (Regan et al., 2018). The most widely expressed NMDARs contain the obligatory subunit GluN1 plus either GluN2B or GluN2A or a mixture of the two; therefore, in next sections we will focus on these two subunits.
The defining features of AD include conspicuous changes in both brain histology and behavior. AD brain is characterized microscopically by extracellular amyloid plaques and intraneuronal NFTs. Accumulating evidence indicated that soluble forms of Aβ and tau work together, independently of their accumulation into plaques and tangles, to drive healthy neurons into the diseased state and that hallmark toxic properties of Aβ require tau (Bloom, 2014). The cognitive impairment of AD is closely related to synaptic plasticity, in which NMDAR plays a critical role (Parameshwaran et al., 2008). Excitatory glutamatergic neurotransmission via NMDAR is critical for synaptic plasticity and survival of neurons. However, excessive NMDAR activity causes excitotoxicity and promotes cell death, underlying a potential mechanism of neurodegeneration occurred in AD (Wang and Reddy, 2017). The major factors that affect NMDAR signaling in AD include glutamate availability and the modulation of NMDAR channel functions (Wang and Reddy, 2017).
In AD, Aβ cascade hypothesis posits an initiating event of amyloidosis, with subsequent tau accumulation preceding downstream brain atrophy and cognitive decline (Hardy, 2002). From amyloid-PET and florbetapir-PET studies, it has been known that Aβ deposition occurs selectively first in the cortex, beginning in temporobasal and frontomedial areas, and successively affecting primary sensory-motor areas and the medial temporal lobe. It was followed by hippocampal regions, then striatum, basal forebrain, thalamus, and finally brainstem nuclei and cerebellum (Grothe et al., 2017; Rice and Bisdas, 2017; Whitesell et al., 2018). Deposition of Aβ in the medial parietal cortex appears to be the first stage in the development of AD, although tau aggregates in the medial temporal lobe precede Aβ deposition in cognitively healthy older people (Jagust, 2018).
Aβ is produced by endoproteolysis of the parental amyloid precursor protein (APP), which is achieved by the sequential cleavage of APP by groups of enzymes termed β- and γ-secretases (LaFerla et al., 2007). Aβ is produced as a monomer, but readily aggregates to form multimeric complexes. Human Aβ can exist in diverse assembly states, including monomers, dimers, trimers, tetramers, dodecamers, higher-order oligomers and protofibrils, as well as mature fibrils, which can form microscopically visible amyloid plaques in brain tissues (Glabe, 2008). The original amyloid hypothesis postulated that accumulation of Aβ in the brain is the primary influence driving AD pathogenesis. Cell studies and animal experiments confirmed that oligomeric, soluble Aβ rather than insoluble amyloid plaques exert the toxic effect (Lacor et al., 2007; Shankar et al., 2007). Thus, according to the modified amyloid cascade hypothesis, soluble oligomeric assemblies of Aβ induce the neurodegenerative triad (Wu et al., 2010).
The Aβ peptide was first identified as a component of extracellular amyloid plaques in the mid-1980s. Now, there have been a large number of studies have provided evidence for the presence of intracellular Aβ within neurons (LaFerla et al., 2007). Immunogold electron microscopy has been carried out to demonstrate that Aβ1-42 can be found in multivesicular bodies (MVBs) of neurons in the human brain, where it is associated with synaptic pathology. Studies also show that Aβ produced intracellularly in the endosome compartments (Kinoshita, 2003; Rogaeva et al., 2007), ER (endoplasmic reticulum) (Busciglio et al., 1993; Cook et al., 1997), and trans-Golgi network in neurons (Hartmann et al., 1997). The secreted Aβ, which forms the extracellular Aβ pool, could be taken up by cells and internalized into intracellular pools. Aβ binds to the α7 nicotinic acetylcholine receptor (α7nAChR) with high affinity, resulted in receptor internalization and accumulation of Aβ intracellularly (Wang et al., 2000; Nagele et al., 2002). In addition to nicotinic receptors, Aβ internalization has been reported through LRP (low-density lipoprotein receptor related protein) (Bu et al., 2006), RAGE (scavenger receptor for advanced glycation end products) (Deane et al., 2003), and NMDAR (Caldeira et al., 2013). Aβ uptake was completely blocked by NMDAR antagonists, evidencing an involvement of this receptor in the re-uptake of the peptide (Bi et al., 2002).
Aβ oligomerization initiates within cells rather than in the extracellular space (Walsh et al., 2000). Much evidence suggests that Aβ oligomers are more potent than Aβ fibrils in eliciting abnormalities in synaptic functions. Moreover, oligomers of Aβ1-42 are considered the most synaptotoxic forms, responsible for early cognitive deficits in AD (Palop and Mucke, 2010). Aβ leads to aberrant excitatory network activity and compensatory inhibitory responses involving learning and memory circuits (Palop et al., 2007). Intracellular Aβ within the MVBs is mechanistically linked to cytosolic proteasome inhibition, lead to higher Aβ levels and the buildup of tau protein (Almeida, 2006; LaFerla et al., 2007). Low (picomolar range) concentrations of Aβ markedly potentiate synaptic transmission, whereas higher concentrations (low nanomolar range) of Aβ cause synaptic depression by acting on α7nAChR activation (Dineley et al., 2002; Puzzo et al., 2008).
Accumulation of intracellular Aβ in mitochondria is associated with the multitude of mitochondrial defects described in AD (Manczak et al., 2006). Aβ is transported into mitochondria via the translocase of the outer membrane machinery and is located in mitochondria cristae (Hansson Petersen et al., 2008). In AD postmortem brain and cellular and animal AD models, mitochondrial dysfunction can be triggered by Aβ. Aβ-induced mitochondrial dysfunction was reported to be related to the interaction of Aβ with different mitochondrial proteins, including proteins of the outer mitochondrial membrane, intermembrane space, inner mitochondrial membrane, and the matrix, impairing oxidative phosphorylation and mitochondrial dynamics and increasing reactive oxygen species (ROS) production (Pagani and Eckert, 2011).
Aβ also induce ER stress. In mature hippocampal cultures, Aβ induce NADPH oxidase (NOX)-mediated superoxide production downstream of GluN2B and impairs ER and cytosolic Ca2+ homeostasis (Costa et al., 2012). In rat brain endothelial cells (RBE4 cell line), Aβ increased the levels of several markers of ER stress-induced unfolded protein response (UPR), affected the Ca2+ homeostasis (Fonseca et al., 2013). In turn, ER stress mediated Aβ production or neurotoxicity. Increased the ER stress promoted Aβ production was observed in RGC-5 cells, a cell line identical to the photoreceptor cell line 661W (Liu et al., 2014). Knockdown of syntaxin 5 expression enhanced the secretion of Aβ peptides under condition without ER stress in NG108-15 cells (Suga et al., 2015). In PC12 cells, the ER stress response factor X-box binding protein 1 (XBP1) prevents Aβ-introduced accumulation of [Ca2+]i (Casas-Tinto et al., 2011).
Aβ was shown to be produced in high amounts in synaptic terminals of the hippocampal dentate gyrus of transgenic mice and deposited in extracellular plaques (Lazarov et al., 2002). Indeed, studies in AD postmortem brain tissue samples and AD animal models support a role for disruption of synaptic Ca2+ regulation in the neurotoxic action of Aβ, which may serve as a trigger for synaptic deterioration driving the cognitive loss in AD (Camandola and Mattson, 2011; Mota et al., 2014). Cognitive function ultimately depends on synaptic plasticity where LTP is associated with synapse growth and LTD is associated with synapse loss. Aβ is associated with the inhibition of LTP (Walsh et al., 2002) and the promotion of LTD (Li et al., 2009; Hunter et al., 2018). During the induction of LTP, the strong and prolonged release of glutamate from the presynaptic terminal activates AMPA receptors and the subsequent depolarization removes the Mg2+ blockade of the NMDAR channel and allows the influx of Ca2+. This strong activation of NMDARs triggers a Ca2+/calmodulin-dependent protein kinase II (CaMKII)-mediated signaling cascade that eventually leads to an enhanced synaptic strength. On the contrary, a modest activation of NMDARs causes a modest increase in postsynaptic Ca2+ and triggers phosphatases-mediated LTD (Luscher and Malenka, 2012). Activation of synaptic NMDARs and large increases in [Ca2+]i are required for LTP, whereas internalization of synaptic NMDARs, activation of perisynaptic NMDARs and lower increases in [Ca2+]i are necessary for LTD. LTP induction promotes recruitment of AMPA receptors and growth of dendritic spines, whereas LTD induces spine shrinkage and synaptic loss (Kullmann and Lamsa, 2007). Pathologically elevated Aβ may indirectly cause a partial block of NMDARs and shift the activation of NMDAR-dependent signaling cascades toward pathways involved in the induction of LTD and synaptic loss (Kamenetz et al., 2003; Shankar et al., 2007). This model is consistent with the fact that Aβ impairs LTP (Walsh et al., 2002; Cleary et al., 2005) and enhances LTD (Kim et al., 2001; Hsieh et al., 2006; Li et al., 2009). Although the mechanisms underlying Aβ-induced LTD have not yet been fully elucidated, they may involve receptor desensitization (Liu, 2004) or internalization and subsequent collapse of dendritic spines (Snyder et al., 2005; Hsieh et al., 2006).
Aβ modulated NMDA-induced responses and vice versa; pre-exposure to Aβ decreased NMDA-evoked [Ca2+]i rise and pre-exposure to NMDA decreased Aβ response. In addition, simultaneous exposure to Aβ plus NMDA synergistically increased [Ca2+]i levels, an effect mediated by GluN2B-containing NMDARs (Ferreira et al., 2012). Accumulating evidence demonstrates that glutamate receptors are dysregulated by Aβ oligomers, resulting in disruption of glutamatergic synaptic transmission which parallels early cognitive deficits. In theory, there are several potential roles for the NMDA receptor in Aβ-related mechanisms (Malinow, 2012): first, NMDA receptor function may be an important downstream target of Aβ; second, NMDA receptors may be necessary in the actions of Aβ on synaptic transmission and plasticity; third, NMDA receptor may be a receptor for Aβ, or indirectly; and fourth, NMDA receptor activity may control the formation of Aβ.
Expression of NMDAR subunits differentially distribute throughout the brain and change strikingly during development. The glycine/D-serine binding GluN1 subunit is an obligatory subunit in all NMDA receptor subtypes. In accordance with the widespread central nervous system (CNS) distribution of NMDARs, the GluN1 subunit is ubiquitously expressed from embryonic stage E14 to adulthood (Monyer et al., 1994). The four different glutamate binding GluN2 subunits (GluN2A-D) have pronounced differences in both developmental and regional expression levels and endow NMDA receptors with strikingly different pharmacological and functional properties. In the embryonic brain, only GluN2B and GluN2D subunits are expressed, and the latter is mostly found in caudal regions. During the first two postnatal weeks, GluN2A expression rises steadily to become widely and abundantly expressed in virtually every CNS area in the adult. Meantime, GluN2D expression drops markedly, and in the adult, it is expressed at low levels mostly in the diencephalon and mesencephalon. GluN2B expression is maintained at high levels following birth, peaks around the first postnatal week and becomes progressively restricted to the forebrain. Expression of GluN2C appears late in development, about postnatal day 10, and mainly confined to the cerebellum and the olfactory bulb (Paoletti et al., 2013).
In the adult CNS, particularly in higher brain structures, such as the hippocampus and cortex, GluN2A and GluN2B are the predominant subunits, indicating that they have central roles in synaptic function and plasticity (Monyer et al., 1994; Takai et al., 2003). GluN2C- or GluN2D- containing receptors appear to give rise to ‘low-conductance’ openings with a lower sensitivity to extracellular Mg2+, which may affect the Ca2+ influx generated by synaptic activation of NMDAR (Momiyama et al., 1996). Misra et al. (2000) results indicate that GluN1/GluN2D receptors do not contribute to the excitatory postsynaptic current (EPSC) and appear to be restricted to the extrasynaptic membrane. The possibility still exists that the GluN2D subunit is present at the synapse but is preferentially co-assembled with other GluN2 subunits, such as triheteromeric assemblies (GluN1/GluN2B/GluN2D) (Dunah et al., 1998; Cull-Candy et al., 2001). GluN3A and GluN3B were cloned based on similarity to GluN1 and GluN2 subunits and were the last NMDA receptor subunits to be discovered (Low and Wee, 2010). GluN3A expression is low embryonically, peaks during early postnatal life, and diminishes to lower levels in adulthood. This expression profile is observed in many regions of the brain (Bendová et al., 2009; Roberts et al., 2009). Conversely, GluN3B levels are low around postnatal day 10 and gradually increase into adulthood within the neocortex, hippocampus, striatum, cerebellum, brainstem, and spinal cord (Wee et al., 2008). GluN3A and GluN3B are expressed by multiple neuronal cell types, including interneurons, pyramidal cells, motor neurons, trigeminal neurons, retinal ganglion and amacrine cells (Pachernegg et al., 2012). Although GluN3A is present in oligodendrocytes, it does not seem to be expressed in astrocytes (Matsuda et al., 2003; Káradóttir et al., 2005; Salter and Fern, 2005). The GluN3 subunits bind glycine and D-serine (Yao, 2006; Yao et al., 2008), but the functional properties and physiological roles of GluN3-containing NMDA receptors remain elusive. GluN3A acts in a dominant-negative manner to suppress receptor activity. GluN3A containing NMDARs display striking regional and temporal expression specificity, and, unlike most other NMDAR subtypes, they have a low conductance, are only modestly permeable to Ca2+, and pass current at hyperpolarized potentials in the presence of magnesium. While glutamate activates triheteromeric NMDARs composed of GluN1/GluN2/GluN3A subunits, glycine is sufficient to activate diheteromeric GluN1/GluN3A-containing receptors (Henson et al., 2010). About GluN3A is expressed in the right place at the right time to regulate spine and synapse development, there are two hypotheses, if the influence of GluN3A is to serve as a ‘synaptic brake’ to limit synapse/spine formation or if it serves as a ‘synaptic marker’ to promote the elimination of spines (Henson et al., 2010). Roberts et al. (2009) found that prolonging GluN3A expression prevents glutamatergic synapse maturation by limiting synapse potentiation and growth, and decreasing spine density, accompanied by major impairments in learning and memory processes. Whereas knocking out endogenous GluN3A conversely accelerates synaptic maturation events.
In line with the large number of subunits and their overlapping expression in several brain regions, many different NMDAR subtypes coexist in the CNS. All NMDAR subtypes are thought to combine two copies of the obligatory GluN1 subunit plus two copies of GluN2 and/or GluN3 subunits. Diheteromeric GluN1/GluN2B and GluN1/GluN2A receptors represent an important fraction of juvenile and adult NMDARs. Triheteromeric GluN1/GluN2A/GluN2B receptors also populate many regions in the adult brain, particularly in the hippocampus and cortex, with estimates of abundance ranging from 15% to >50% of the total receptor population (Al-Hallaq et al., 2007). They may maintain some signaling properties of the GluN2B subunit while having the kinetic properties of GluN1/2A NMDARs (Sun et al., 2017). The sensitivity of GluN1/GluN2A/GluN2B triheteromers to the agonists, glutamate and glycine, is intermediate to that of diheteromers. The sensitivity to glycine, conferred by the GluN1 sub-units, is close to that of GluN1/GluN2B receptors, whereas the sensitivity to glutamate, conferred by the GluN2 sub-units, is closer to that of GluN1/GluN2A receptors (Hansen et al., 2014; Stroebel et al., 2014). Triheteromeric GluN1/GluN2A/GluN2C receptors and GluN1/GluN2B/GluN2D receptors have also been described. Triheteromeric GluN1/GluN2A/GluN2C receptors are the predominant NMDARs in cerebellar granule cells and propose that co-expression of GluN2A and GluN2C in cerebellar granule cells occludes cell surface expression of diheteromeric GluN1/GluN2C receptors. For many agonists, potentiators, and inhibitors as well as endogenous ligands, GluN1/GluN2A/GluN2C NMDARs had intermediate properties compared with diheteromeric receptors (Bhattacharya et al., 2018). Similarly, triheteromeric GluN1/GluN2A/GluN2D and GluN1/GluN2B/GluN2D complexes populate the rat thalamus and midbrain (Dunah et al., 1998), as well as the human spinal cord (Sundström et al., 1997). GluN3A and GluN3B subunits can co-assemble with GluN2 subunits to form triheteromeric GluN1/GluN2/GluN3 receptors that are involved in synapse maturation during brain development (Paoletti, 2011; Paoletti et al., 2013; Pérez-Otaño et al., 2016).
N-methyl-D-aspartate receptors dysregulation evoked by Aβ and the consequent loss of Ca2+ homeostasis are thought to be related to the early cognitive deficits observed in AD. Many researches have demonstrated that Aβ oligomers and NMDAR contribute to the synaptic dysfunction in AD (Roselli et al., 2005; Liu et al., 2010; Chang et al., 2016; Li et al., 2016). The effect of Aβ on synapses is to produce depression of glutamatergic synaptic transmission (Kamenetz et al., 2003) and reduce surface glutamate receptors and other synaptic components in cultured hippocampal neurons (Snyder et al., 2005; Liu et al., 2010). NMDAR antagonist D-VAP can blocked the Aβ-induced depression of glutamatergic transmission (Kamenetz et al., 2003; Parameshwaran et al., 2008). Aβ accumulation may activate NMDARs at early stages of AD (Parameshwaran et al., 2008), and that Aβ oligomers can evoke an immediately [Ca2+]i rise through activation of GluN2B-containing NMDARs in cultured cortical neurons (Ferreira et al., 2012). Conversely, Aβ-induced activation of GluN2A-containing NMDARs in non-neuronal cells, namely in Xenopus laevis oocytes (Texidó et al., 2011) and HEK293cells (Domingues et al., 2007), was previously reported.
The effects of Aβ on the mRNA and protein levels of NMDAR subunits have been extensively studied in vivo and in vitro, although the alterations of the NMDARs remain inconclusive. Some reported that levels of both GluN1 mRNA (Ułas and Cotman, 1997; Hynd et al., 2001) and GluN1 protein (Sze et al., 2001; Jacob et al., 2007) were significantly lower in AD patients. Conversely, other studies evidenced that levels of GluN1 mRNA were unchanged (Bi and Sze, 2002; Hynd et al., 2004). Moreover, levels of GluN2A and GluN2B mRNA were decreased in AD brain (Hynd et al., 2004), especially in hippocampus and the entorhinal cortex of postmortem (Bi and Sze, 2002). Whereas levels of GluN2C and GluN2D mRNA did not differ from that in controls (Hynd et al., 2004). The levels of GluN2A and GluN2B protein were also decreased in AD brain (Sze et al., 2001; Hynd et al., 2004). Other study found that mRNA expression and protein levels for GluN1 and GluN2B were significantly reduced, while GluN2A mRNA expression and protein levels were unchanged in hippocampus (Mishizen-Eberz et al., 2004).
Costa et al. (2012) data show that Aβ oligomers binding to mature hippocampal cells was prevented when the extracellular termini of GluN1 and GluN2B, but not of GluN2A, was blocked, indicating that Aβ oligomers are able to bind NMDARs extracellularly. These were consistent with the results that NMDARs may be direct or indirect targets for Aβ, by the obligatory GluN1 subunit, affecting the activity of these receptors (De Felice et al., 2007; Malinow, 2012). In the AD brain and human cortical neurons, excitatory synapses containing the GluN2B subunit of the NMDARs appear to be the main sites of Aβ oligomers accumulation. In addition, some studies report that Aβ oligomers may bind to EphB2, a surface tyrosine kinase that binds the NMDARs for maintaining its integrity. EphB2 degradation results in a decrease in surface localization of NMDA receptor subunits like GluN2B (Shi et al., 2016), the loss of NMDAR function and reduced LTP (Dalva et al., 2000).
Some results suggest a circuit in which Aβ facilitates NMDARs activation, which in turn control the formation of Aβ. NMDARs activation (Lesne, 2005), and more particularly extrasynaptic activation (Bordji et al., 2010), increased production and secretion of Aβ in primary cultures of cortical neurons, which was preceded by a shift from APP695 to Kunitz protease inhibitory domain (KPI) containing amyloid-β precursor protein (KPI-APP), isoforms exhibiting an important high amyloidogenic potential, followed by a shift from α-secretase to β-secretase-mediated APP processing (Hoey et al., 2009). An in vivo study indicates that the level of NMNDAR activation may control the effect on Aβ production: low levels of NMDAR activation increase Aβ production while higher levels reduce Aβ production (Verges et al., 2011). However, a study using in vivo microdialysis techniques demonstrated that the neuronal activity dependence of Aβ secretion lies in the presynaptic compartment suggesting no NMDAR dependence (Cirrito et al., 2005). The role of NMDAR in production of Aβ is likely complex and requires more investigation.
N-methyl-D-aspartate receptors mediate many forms of synaptic plasticity. These tetrameric receptors consist of two obligatory GluN1 subunits and two regulatory subunits, usually a combination of GluN2A and GluN2B. Among the six regulatory subunits of NMDARs, GluN2A and GluN2B have been the most extensively studied because they are broadly expressed in the brain, predominate in the postnatal cortex, and are believed to play important roles in synaptic plasticity. In the neonatal neocortex GluN2B-containing NMDARs predominate, and sensory experience facilitates a developmental switch in which GluN2A levels increase relative to GluN2B (Yashiro and Philpot, 2008). However, GluN1/GluN2B carry about twofold more charge for a single synaptic event than GluN1/GluN2A channels (Erreger et al., 2005). More studies showed that GluN2A-dominated synapses are more likely to induce LTD than GluN2B-dominated synapses, while GluN2B-dominated synapses have a greater capacity to be potentiated. GluN2A co-immunoprecipitates with neuronal nitric oxide (NO) synthase more effectively than GluN2B (Al-Hallaq et al., 2007). Although this interaction is likely indirect, the association raises the interesting possibility that NO-mediated presynaptic forms of LTP and LTD may be preferentially linked to GluN2A-mediated signaling pathways (Haghikia et al., 2007). Activated CaMKII binds strongly to GluN2B, which is required for LTP induction (Barria and Malinow, 2005). However, both GluN2A-containing and GluN2B-containing NMDARs are capable of supporting bidirectional synaptic plasticity (Yashiro and Philpot, 2008). GluN2A and GluN2B are present as either diheteromers (GluN1/GluN2A or GluN1/GluN2B) or triheteromers (GluN1/GluN2A/GluN2B) (Yashiro and Philpot, 2008). In isolated GluN2A-only or GluN2B-only synapses, GluN1/GluN2A diheteromeric channels exhibit faster rising and decaying currents than GluN1/GluN2B diheteromeric channels (Prybylowski et al., 2002). GluN1/GluN2A/GluN2B triheteromeric channels appear to exhibit intermediate decay time courses between the two diheteromeric channel types (Vicini et al., 1998).
Proteins interacting with NMDAR subunits are therefore important determinates for the direction of synaptic plasticity. The differential interaction of GluN2A and GluN2B subunits to membrane-associated guanylate kinase (MAGUKs) is controversial. It was once believed that GluN2A preferentially bound to postsynaptic density protein-95 (PSD-95), while GluN2B preferentially bound synapse-associated protein 102 (SAP102) (Sans et al., 2000; Krapivinsky et al., 2003). GluN2B interacts directly with Ras-guanine nucleotide releasing factor 1 (Ras-GRF1) (Krapivinsky et al., 2003), and synaptic Ras GTPase activating protein (RasGAP), presumably through SAP102 (Kim et al., 1998). The unique associations are likely to affect the induction of plasticity (Zhu et al., 2002). Moreover, these interactions were thought to control distinct synaptic localization of GluN2A and GluN2B (Townsend et al., 2003). However, a recent biochemical study using a serial immunoprecipitation suggests that MAGUK proteins such as PSD-95 and SAP102 interact with diheteromeric GluN1/GluN2A and GluN1/GluN2B receptors at comparable levels (Al-Hallaq et al., 2007). Additional studies are needed to clarify the association of NMDAR subunits with MAGUK family members and what effects these associations may have on receptor localization and on plasticity signaling pathways (Yashiro and Philpot, 2008).
Our results suggested that enhancement of GluN2A activity and/or the reduction of GluN2B activity may be used in order to halt the early Aβ-mediated synaptic dysfunction (Liu et al., 2010). GluN2A and GluN2B subunits have opposite roles in regulating [Ca2+]i homeostasis. In rat cerebral cortical neurons, Aβ, like NMDA, increase [Ca2+]i levels mainly through activation of GluN2B. Conversely, GluN2A antagonism potentiates [Ca2+]i rise induced by a high concentration of Aβ (1 μM) (Ferreira et al., 2012). Simultaneous exposure to Aβ and NMDA affects the response to GluN2B-composed NMDARs, largely causing mitochondrial depolarization and mitochondrial Ca2+ (mitCa2+) retention (Ferreira et al., 2015). Aβ-induced ER stress and hippocampal dysfunction were prevented by ifenprodil, an antagonist of GluN2B subunits, while the GluN2A antagonist NVP-AAM077 only slightly attenuated Aβ-induced neurotoxicity (Costa et al., 2012). Moreover, exposure to Aβ caused a decrease in total and polymerized levels of beta-III tubulin and polymerized alpha-tubulin in mature hippocampal cells, which could be prevented by MK-801, memantine, and ifenprodil, suggesting a role for GluN2B-containing NMDARs in Aβ toxicity (Mota et al., 2012).
GluN2A and GluN2B are the major subunits of functional NMDARs. Some studies have suggested that GluN2A and GluN2B may differentially mediate NMDAR function at synaptic and extrasynaptic locations and play opposing roles in excitotoxicity (Thomas et al., 2006; Harris and Pettit, 2007). In the postsynaptic membrane of excitatory neurons, the density of NMDARs is higher in dendritic spines, within the postsynaptic density (PSD), considered synaptic NMDARs, than in the dendritic shaft and somatic membrane (Köhr, 2006). At the PSD, the receptors form a large macromolecular NMDAR complex (NRC), containing a vast collection of scaffolding, adaptor and effector proteins that are involved in activation of downstream signaling cascades and regulation of NMDAR function, membrane stability and trafficking (Sheng and Lee, 2000). Extrasynaptic NMDARs are localized at sites further from the PSD, on the spine neck, the dendritic shaft or soma (Newpher and Ehlers, 2008). Perisynaptic NMDARs are located on the plasma membrane (PM) within 200–300 nm of the PSD. The perisynaptic region may contain mobile receptors that are in transit to and from the PSD (Zhang and Diamond, 2006; Petralia et al., 2009). Synaptic and extrasynaptic NMDARs are gated by different endogenous co-agonists: D-serine for synaptic NMDARs and glycine for extrasynaptic NMDARs. Both co-agonists are of glial origin (Papouin et al., 2012). It was proposed that GluN2A- and GluN2B-containing receptors are predominantly found synaptically and extrasynaptically, respectively. For NMDAR redistribution plays a larger role during development, when there is a synaptic switch from GluN2B to GluN2A-containing receptors (Barria and Malinow, 2002). In addition, it has been found that synaptic and extrasynaptic NMDA receptors have opposing effects in determining the fate of neurons (Vizi et al., 2013). Calcium entry through synaptic GluN2A-containing NMDARs induces activity of cAMP response element binding protein (CREB) and gene expression of CREB-evoked brain-derived neurotrophic factor (BDNF), which is involved in the neuroprotective action of synaptic glutamatergic transmission. In contrast, Ca2+ entry through GluN2B-containing NMDARs, which are expressed extrasynaptically, triggers the CREB shut-off pathway (Leveille et al., 2008). There is some evidence that synaptic NMDARs support LTP, while extrasynaptic NMDARs mediate LTD in the mature brain (Massey, 2004). Specific NMDAR subunits are not confined to particular subcellular locations on PM. Some evidences shown that GluN2A- and GluN2B-subtypes are present at both synaptic and extrasynaptic sites (Thomas et al., 2006; Harris and Pettit, 2007; Martel et al., 2009; Zhou et al., 2013a). These suggest that any difference in signaling by synaptic and extrasynaptic NMDARs must be due to intracellular signaling pathways rather than subunit identity or mobility (Harris and Pettit, 2007). And some researchers thought that these results were partially refuted on the basis that these experiments were carried out on developing hippocampal cultured neurons; the same results may not be found under in vivo conditions (Newpher and Ehlers, 2008).
Certain interactions of NMDAR subunits with distinct signaling molecules may occur at synaptic but not at extrasynaptic sites (Köhr, 2006). Extrasynaptic NMDARs are exposed to ambient glutamate, whether this glutamate concentration is high enough to tonically activate extrasynaptic NMDARs remains controversial. Although microdialysis studies report that ambient glutamate concentrations in vivo are high enough to activate extrasynaptic NMDARs (Nyitrai et al., 2006), a study suggests that glutamate transporters regulate ambient glutamate concentrations at a level that is too low to cause significant receptor activation (Herman and Jahr, 2007). While, some reports that glutamate that is released into the extracellular space mainly from glial processes (Fellin et al., 2004) may result in the persistent activation of extrasynaptic GluN2B receptors, which are of high affinity and are sensitive to low concentrations of glutamate (Vizi, 2000).
Activation of synaptic NMDARs and large increases in [Ca2+]i are required for LTP, whereas internalization of synaptic NMDARs, activation of extrasynaptic NMDARs and lower increases in [Ca2+]i are necessary for LTD. LTP induction promotes recruitment of AMPARs and growth of dendritic spines, whereas LTD induces spine shrinkage and synaptic loss (Kullmann and Lamsa, 2007). Importantly, glutamate spillover from synapses or glutamate released from astrocytes activates extrasynaptic NMDARs (Fellin et al., 2004). Extrasynaptic NMDARs are activated not only at pathological situations (Hardingham et al., 2002), but also by bursts of activity that can occur under physiological situations (Harris and Pettit, 2008). Retinal ganglion cells express only extrasynaptic NMDARs and are invulnerable to NMDA neurotoxicity (Ullian et al., 2004). Synaptic NMDARs can also cause neurotoxicity (Sattler et al., 2000; Sinor et al., 2000) and can induce LTD (Malenka and Bear, 2004). Furthermore, Zhou et al. (2013b) demonstrate that activation of synaptic or extrasynaptic NMDAR alone stimulated pro-survival but not pro-death signaling, for they had overlapping but not opposing effects on genomic responses. Low-dose NMDA preferentially activated synaptic NMDAR and stimulated the extracellular signal-regulated kinase 1/2 (ERK1/2)-CREB-BDNF pro-survival signaling, while higher doses progressively activated increasing amount of extrasynaptic NMDAR along with synaptic NMDAR and triggered cell death program. While, Liu et al. (2007) suggested that the subunit composition of NMDARs rather than their cellular location determines the final effect of the activation of the NMDARs by glutamate.
N-methyl-D-aspartate receptors are highly mobile at both synaptic and extrasynaptic membranes. Importantly, the surface mobility of NMDARs appears to change with development (Harris and Pettit, 2007; Köhr, 2007) in a subunit composition-specific manner (Groc et al., 2006). GluN2A-containing NMDARs are less mobile than GluN2B-containing NMDARs, and the synaptic residency time of GluN2B-containing NMDARs decreases over development. This decrease in synaptic dwell time of GluN2B containing NMDARs is mediated by Reelin, an extracellular matrix protein (Groc et al., 2007). Regulatory mechanisms underlying synaptic and extrasynaptic NMDAR distribution include: protein–protein interactions, phosphorylation, palmitoylation, ubiquitination, role of proteases, such as calpain (Gladding and Raymond, 2011).
Aβ-induced synaptic depression may result from an initial increase in synaptic activation of NMDARs by glutamate, followed by synaptic NMDAR desensitization, NMDAR internalization, and activation of extrasynaptic or perisynaptic GluN2B-enriched NMDARs, which have a key role in LTD induction (Palop and Mucke, 2010). On one hand, Aβ downregulates the synaptic NMDAR response by promoting NMDAR endocytosis, leading to either neurotoxicity or neuroprotection. On the other hand, Aβ enhances the activation of extrasynaptic NMDARs by decreasing neuronal glutamate uptake and inducing glutamate spillover, subsequently causing neurotoxicity (Wang et al., 2013). Aβ oligomers bind with high affinity to cell surface cellular prion protein (PrPC). PrPC and Fyn are enriched in PSD. Metabotropic glutamate receptor, mGluR5, has ability to couple Aβ-PrPC with Fyn, controls the transmission of neurotoxic signals to intracellular substrates (Um et al., 2013; Haas et al., 2014). Aβ engagement of PrPC activates Fyn to phosphorylate the GluN2B subunit of NMDARs, which was coupled to an initial increase and then a loss of surface NMDARs (Um et al., 2012). Pretreatment with the 6D11 antibody to PrPC could prevented the Aβ-induced synaptic dysfunction (Um et al., 2012). Aβ also binds to the α7nAChR, resulting in increased Ca2+ influx, calcineurin activation and dephosphorylation and activation of STEP61. Increased STEP61 activity leads to Fyn inactivation and reduced NMDAR exocytosis, as well as enhanced GluN2B Y1472 dephosphorylation and increased NMDAR internalization (Kurup et al., 2010).
Tau is a major component of the NFT that represent a pathological hallmark of AD. In the healthy brain, tau is an exclusively axonal protein, engaged in the assembly and stability of microtubules. In contrast, in the AD brain, tau is hyperphosphorylated and forms fibrils that appear as neuropil threads in dendrites and as NFTs in the somatodendritic compartment and axons. It had provided strong evidence cerebral amyloid deposition precedes cerebral tau pathology in familial autosomal dominant AD, While, the appearance of NFTs precedes Aβ pathology in the vast majority of affected regions in sporadic AD (Morris et al., 2018). Roberson et al. (2007) first ruled out the possibility that tau reduction altered Aβ levels or aggregation, and uncoupled Aβ from downstream pathogenic mechanisms (Ittner et al., 2010).
Tau not only contributes to axonal structure by maintaining microtubule stability, but also is important in the regulation of synaptic function. Tau is required for Fyn-mediated NMDAR activation in the PSD (Ittner et al., 2010), and tau has been shown to be essential for the induction of LTD (Kimura et al., 2014) as well as BDNF-dependent morphological plasticity (Chen et al., 2012). Fyn, a member of the Src family of tyrosine kinases (Thomas and Brugge, 1997), can phosphorylate tau at its tyrosine 18 residue to generate pY18-tau (Lee, 2004) and can bind to tau through one or more proline-rich (PxxP) motifs in tau (Lee et al., 1998; Reynolds et al., 2008; Usardi et al., 2011). Fyn phosphorylates the NMDAR subunit GluN2B at Y1472 (Tezuka et al., 1999), which strengthens the interaction between NMDARs and PSD-95 in the PSD (Rong et al., 2001) and enhances the activity of GluN2B-containing NMDARs (Groveman et al., 2012). Some experiments indicated that tau is normally highly enriched in axons relative to dendrites (Binder et al., 1985), but in response to Aβ is extensively redistributed into the somato-dendritic compartment (Delacourte et al., 1990; Zempel et al., 2010). Excess Fyn accompanies the excess tau in AD dendrites and upregulates NMDA receptor activity there, flooding the dendrites with harmful levels of calcium. This calcium-driven excitotoxicity can damage postsynaptic sites and cause neuron death. Some results confirmed that glutamate-induced excitotoxicity is inhibited by reduction of tau (Roberson et al., 2007; Ittner et al., 2010) and exacerbated by overexpression of tau (Decker et al., 2016; Maeda et al., 2016). In turn, glutamate-induced excitotoxicity increased tau expression (Sindou et al., 1992; Esclaire et al., 1997) and phosphorylation (Sindou et al., 1994). Recently, it has been reported that activation of extrasynaptic NMDA receptors induces tau overexpression, with a simultaneous neuronal degeneration and decreased neuronal survival (Sun et al., 2016).
Traditionally, NMDARs are thought to be located at the postsynaptic membrane, while recent anatomical and physiological evidence demonstrates that they may also exist at presynaptic terminals. Presynaptic NMDARs (preNMDARs) can regulate presynaptic glutamate release, and reshape synaptic transmission and plasticity (Corlew et al., 2007; Paoletti et al., 2013; Banerjee et al., 2016; Dore et al., 2017). Therefore, preNMDAR subunit composition is critical for modulating where and how preNMDARs influence glutamate release. The subunit composition of preNMDARs shows strong variability, depending on the brain region, all four GluN2 subunits (Glu2A-D) as well as GluN3A can be incorporated (Bouvier et al., 2015). The presynaptic GluN2B subunit has been found in many brain regions, such as the hippocampus (Jourdain et al., 2007; McGuinness et al., 2010; Berg et al., 2013), the cerebellum (Rossi et al., 2012), the entorhinal cortex (Yang et al., 2006, 2008), somatosensory (Brasier and Feldman, 2008) and visual cortex (Banerjee et al., 2016). Although appears and peaks later during development (Monyer et al., 1994; Henson et al., 2010), GluN2A subunit can also be incorporated into preNMDAR sites (Bouvier et al., 2015). At cerebellar parallel fiber-Purkinje cell synapses, preNMDARs are predominately GluN1/GluN2A diheteromeric (Bidoret et al., 2009). Furthermore, GluN2B and GluN3A subunits, likely combining into triheteromeric GluN1/GluN2B/GluN3A, are essential preNMDARs in the developing L2/3 visual cortex (Li et al., 2009; Larsen et al., 2014).
It has been reported that preNMDARs may regulate both spontaneous and evoked release. Although the two forms of release were initially thought to employ the same machinery, the emerging evidence indicates that preNMDARs control evoked and spontaneous release by distinct mechanisms. PreNMDARs may control spontaneous release independently of Mg2+ and Ca2+ while regulating evoked release in a frequency-dependent manner by relying on the more conventional Mg2+-dependent pathway (Kunz et al., 2013; Paoletti et al., 2013; Abrahamsson et al., 2017; Bouvier et al., 2018). Abrahamsson et al. (2017) showed that preNMDARs in L5 pyramidal cells regulate evoked and spontaneous release via RIM1αβ and JNK2-dependent pathway, respectively.
In addition, the activation of preNMDARs is necessary for the induction of LTD (Duguid, 2013; Paoletti et al., 2013), but it is noteworthy that preNMDARs roles shift over development. Induction of LTD in visual cortex pyramidal cells of young mice (before postnatal 20 days) requires activation of presynaptic, whereas in older mice, the induction of LTD requires postsynaptic NMDARs activation (Corlew et al., 2007). Contrast to this results, Carter and Jahr (2016) show that in the somatosensory cortex of 2- to 3-week-old rats and mice, it is postsynaptic, not preNMDARs that required for LTD induction. This contradictory result may be due to different brain regions observed. Not only being required for the induction of LTD, preNMDARs also involve in LTP induction (McGuinness et al., 2010). Park et al. (2014) reported that preNMDARs play a pivotal role in the induction of LTP at mouse corticostriatal synapses. Activation of preNMDARs induces BDNF secretion via enhancing Ca2+ signals in the axonal terminals, indicating that preNMDARs are equally essential as postsynaptic NMDARs in LTP induction (Park et al., 2014).
Bell et al. (2007) found that subjects with mild cognitive impairment (MCI) displayed a paradoxical elevation in glutamatergic presynaptic bouton density, which then depletes and drops with disease progression. These results pointed out that dystrophic neurite generation and reduced presynaptic bouton densities detrimentally influence neurotransmission and cognitive function in later stages of AD. Although some advances have been made, much work still need to clarify the precise functions and molecular mechanisms on preNMDAR.
While neuronal NMDARs have been widely studied, NMDARs are also expressed in many non-neuronal cells, including astrocytes. Compared to neuronal receptors, astrocytic NMDA receptors are poorly understood. The emerging evidence suggests that astrocytic NMDARs have distinct structural and functional properties, including weak susceptibility to Mg2+ blockade and less Ca2+ permeability (Hogan-Cann and Anderson, 2016). NMDAR expression and function in astrocytes has been demonstrated in cultured astrocytes and mouse neocortex (Lalo, 2006; Palygin et al., 2011; Montes and Aguilera, 2015; Li et al., 2016). In human primary astrocytes, all seven identified NMDAR subunits (GluN1, GluN2A-D, GluN3A-B) were found. Increasing data demonstrate that astrocytes express NMDARs with a triheteromeric configuration combining GluN1, GluN2C or D, and a GluN3 subunit (Burzomato et al., 2010; Henson et al., 2012). Glutamate and QUIN both could activate astrocytic NMDARs, which in turn enhances Ca2+ influx and induces signaling cascade (Lee et al., 2010). It is established that astrocytic functional NMDARs are able to respond to neuronal glutamatergic input, which accompany with dynamic intracellular Ca2+ rise triggering gliotransmitter-mediated regulation of synapses function (Kato et al., 2006; Lalo, 2006; Hogan-Cann and Anderson, 2016). Astrocytic NMDARs may also be involved in neuroinflammatory processes, and contribute to morphological transformations characteristic of reactive astrogliosis and mediate the release of proinflammatory cytokines (Sofroniew, 2009; Ting et al., 2009; Gerard and Hansson, 2012). Notably, astrocytic NMDARs maybe contribute to AD due to their roles in facilitating glutamate excitotoxicity (Lee et al., 2010; Mota et al., 2014). Our research found that Aβ-induced early synaptotoxicity can be exacerbated after treatment with antagonism of astrocytic GluN2A and GluN2B, and nerve growth factor (β-NGF) is may be as a mediator in the synaptoprotection of astrocytic GluN2 activation (Li et al., 2016). In co-cultures system, pre-treatment of astrocytes with 1 μM or 10 μM NMDA to activate GluN2A or GluN2B, before exposure to Aβ1-40, was found to prevent the Aβ introduced PSD-95 and synaptophysin decreased. While blockade of astrocytic GluN2A with TCN-201 or GluN2B with ifenprodil respectively both aggravated the synaptotoxic effects of Aβ.
Additionally, NMDARs are also expressed by oligodendrocyte lineage cells, as mediators of intracellular Ca2+ accumulation, leading to reduced oligodendrocyte survival and white matter damage (Káradóttir et al., 2005; Salter and Fern, 2005; Micu et al., 2006). The dominant force behind NMDA-induced currents in mature oligodendrocytes is actually elevated extracellular K+ released upon activation of neuronal or astrocytic NMDARs. Increased in oligodendrocytic Ca2+ would be gated by transient receptor potential cation channel (TRP) A1 (Hamilton et al., 2016). Oligodendrocyte precursor cell (OPC) NMDARs may also contribute to myelination. Activation of NMDARs in OPC cultures increased migration (Wang et al., 1996), myelin basic protein expression (Wake et al., 2011) and differentiation (Li et al., 2013).
Rather surprisingly, a recent study observed that NMDARs were present in primary cultures of microglia from mice cortex and hippocampus (Franco et al., 2018), and exposure to NMDAR co-agonists resulted in evoked inward currents and intracellular Ca2+ increases sensitive to inhibition by the non-competitive NMDAR channel blocker, MK801 (Murugan et al., 2011; Kaindl et al., 2012). NMDAR activation in microglia leads to significant ERK1/2 phosphorylation. ERKs phosphorylation, namely, NMDAR interact with and signaling to mitogen-activated protein kinase (MAPK) depends on CaM and NCS1 in neurons, whereas the signaling via NMDAR in microglia only depends on CaM. NMDAR function was potentiated in microglia from the APPSw,Ind transgenic mice, indicating that the NCS1–NMDAR interaction is relevant for receptor function in the microglia of the AD mouse model (Franco et al., 2018).
It was thought that NMDAR-dependent synaptic plasticity was controlled entirely by Ca2+ influx, and the increased cytoplasmic Ca2+ acts as a second messenger in the postsynaptic neuron (Dore et al., 2017). More recently provocative emerging evidence assumes that, upon binding of glutamate, NMDARs can produce long-term changes in synaptic function in the absence of calcium conductance (Chung, 2013; Nabavi et al., 2013; Birnbaum et al., 2015; Stein et al., 2015). In other words, NMDAR can act as a metabotropic receptor and signal metabotropically, without the need for Ca2+ influx through the channel (Kessels et al., 2013; Nabavi et al., 2013; Stein et al., 2015; Hell et al., 2016; Sanders et al., 2018).
A conventional view proposes that NMDARs triggers LTP via a high level of Ca2+ influx, while metabotropic receptor arises from synaptic depression induced by low frequency stimulation (LFS), which is called as NMDAR-dependent LTD (Malenka, 1994; Thiels et al., 1996). Recent results demonstrated that glutamate binding alone is sufficient to induce conformational changes of NMDARs, triggering p38MAPK signaling cascades, and in turn to induce LTD (Nabavi et al., 2013; Carter and Jahr, 2016; Hell et al., 2016). It is noteworthy that Ca2+ rise could induce LTP; while keeping at the baseline level of intracellular Ca2+ is necessary for metabotropic NMDAR activation, which leads to synaptic depression (Chung, 2013).
Consistent with this metabotropic signaling, some researchers found preNMDARs may control spontaneous release without need of Mg2+ and Ca2+, while evoked release was sensitive to Mg2+ (Kunz et al., 2013; Abrahamsson et al., 2017). PreNMDARs promote transmitter release partly via protein kinase C signaling (Kunz et al., 2013). These data suggest that preNMDARs may signal metabotropically, and further support the emerging propose that evoked and spontaneous release are through distinct machinery (Kavalali, 2015; Dore et al., 2017). Although mounting findings support an ion flux-independent mechanism for NMDAR-dependent LTD (Stein et al., 2015; Carter and Jahr, 2016; Wong and Gray, 2018), it has been challenged by some recent discoveries (Babiec et al., 2014; Volianskis et al., 2015; Sanderson et al., 2016). It is more possible that two distinct forms of NMDAR-dependent LTD co-exist indeed: one requires ion-flux and the other does not (Dore et al., 2017).
In addition, some studies demonstrated that astrocytic NMDARs may also act through metabotropic signaling pathways (Jimenez-Blasco et al., 2015; Montes and Aguilera, 2015), in which a phospholipase C-mediated endoplasmic reticulum Ca2+ rise and activation of protein kinase Cδ may be involved (Jimenez-Blasco et al., 2015), but much work remains to clarify the complex mechanism (Hogan-Cann and Anderson, 2016).
Early synaptic dysfunction in AD is associated with increased levels of Aβ oligomers, which causes a rapid NMDAR-dependent synaptic depression and spine loss (Shankar et al., 2007; Dore et al., 2017). While, some studies showed that Aβ-induced NMDAR-dependent synaptic depression did not require ion flux through the receptor (Kessels et al., 2013; Tamburri et al., 2013; Birnbaum et al., 2015), and was blocked by AP-5, but not MK-801, suggesting a metabotropic effect of NMDARs contributes to Aβ-induced synaptic dysfunction. It may be a common mechanism between metabotropic NMDAR-dependent LTD and Aβ-induced synaptic depression (Hell et al., 2016; Foster et al., 2017).
The current therapeutic arsenal of AD comprise of two classes of medications: the cholinesterase inhibitors (ChEIs), which include galantamine and rivastigmine (both approved for use in mild to moderate AD) and donepezil (approved for use in mild to severe AD); and the non-competitive NMDA receptor antagonist memantine (approved for use in moderate to severe AD). Combining ChEIs and memantine could offer greater benefits on behavior, cognition, and global outcomes (Patel and Grossberg, 2011; Deardorff and Grossberg, 2016). However, these drugs provide only symptomatic benefits in AD (Salomone et al., 2012). Any CNS disorder in which neuronal loss is related to glutamate-induced excitotoxicity has the potential to be treated by the inhibition of NMDARs. Phencyclidine, ketamine, MK-801 (dizocilpine) and memantine all target NMDARs. But due to severe side effects of phencyclidine, ketamine, and MK801, such as high affinity for and long dwell time on the receptor, their clinical application is hindered (Ellison, 1995). By contrast, memantine has lower affinity and is well tolerated, thus is used to treat moderate to severe AD (Reisberg et al., 2003; Pierson et al., 2014; Song et al., 2018). Inspiringly, riluzole, a new inhibitor of glutamate release and postsynaptic glutamate receptor signaling, is now in a phase II trial in mild AD patients (Graham et al., 2017).
Memantine has an inhibitory effect on NMDAR-mediated excitotoxicity (Parsons et al., 2007), strong voltage dependency, and rapid unblocking kinetic properties. Similar to Mg2+, it blocks the NMDAR and does not leave it in the presence of tonic pathological over-activation of the receptor, while allowing the transmission of transient physiological signals crucial for memory and learning processes (Danysz and Parsons, 2012). It has been shown that memantine increases the release of neurotrophic factors from astroglia (Wu et al., 2009), which may account for its survival effect, and exerts a neuroprotective effect. To date, memantine is the only NMDAR antagonist that is clinically approved and preferentially acts as an antagonist of non-synaptic NMDARs (Lipton, 2006). Extrasynaptically located NMDARs have a relevant role in neurodegeneration. Activation of extrasynaptic receptors may not only regulate expression of tau (Paterlini et al., 1998), but also induce transcriptional inactivation of CREB (Rönicke et al., 2011; Grochowska et al., 2017) and also contribute to early synaptic dysfunction (Franco et al., 2018). The low-affinity antagonist memantine will be the focus of growing interest in targeting extrasynaptic GluN2B-containing NMDARs. In addition, selective enhancement of synaptic activity by low doses of NMDA, or reduction of extrasynaptic activity by memantine, halts Aβ-induced neurotoxicity. The concomitant blockade of GluN2A-containing receptors impaired the activation of protective pathways. In contrast, the selective inhibition of GluN2B-containing NMDARs may be beneficial because it prevents neuronal cell death but leaves the protective pathways intact. Therefore, it is almost certain that in the future, neuroprotective drugs for AD should aim at both the enhancement of synaptic activity and the disruption of extrasynaptic GluN2B-containing NMDAR-dependent death signaling (Wang et al., 2013), the non-selective NMDA antagonists cannot be effective as neuroprotective agents.
The first GluN2B-selective drug is ifenprodil, which inhibits GluN2B receptor-mediated currents with an IC50 of 0.34 μM, whereas its affinity for GluN2A receptors is 400-fold lower, with an IC50 of 146 μM (Williams, 1993). Ifenprodil binds to the N-terminal lysine/isoleucine/valine-binding protein (LIVBP)-like domain of GluN2B and acts as a non-competitive antagonist in a use-dependent and voltage-independent manner. There have been several second generation ifenprodil analogs, including CP-101,606 (traxoprodil) and Ro 25-6981 (Table 1), that exhibit a much greater selectivity for GluN2B subunits. Indole-2-carboxamide derivatives, such as RG-13579 and RG-1103, are a new group of GluN2B-selective antagonists. RG-13579 and RG-1103 bind GluN2B-containing receptors 1,600- and 13,600-fold more selectively, respectively, than they bind GluN2A-containing receptors (Nagy, 2004). Inhibition of the interactions of GluN2B with Fyn and GluN2B tyrosine phosphorylation may contribute to the CP-101,606-induced downregulation of NMDAR function (Kong et al., 2015). In vivo [3H]MK-801 binding study shows that NMDAR activity in the rodent forebrain can be inhibited completely by channel blockers, AZD6765 (lanicemine) and MK-801, but only partially (∼60%) by GluN2B receptor antagonists, CP-101,606, MK-0657 (CERC-301), EVT-101, Ro 25-6981 and radiprodil, at doses that completely occupied GluN2B receptors (Fernandes et al., 2015). Graef et al. (2015) demonstrated that a single dose of either the non-selective NMDA receptor blocker ketamine or the selective GluN2B antagonist CP-101,606 can enhance ex vivo hippocampal LTP in rats 24 h after treatment.
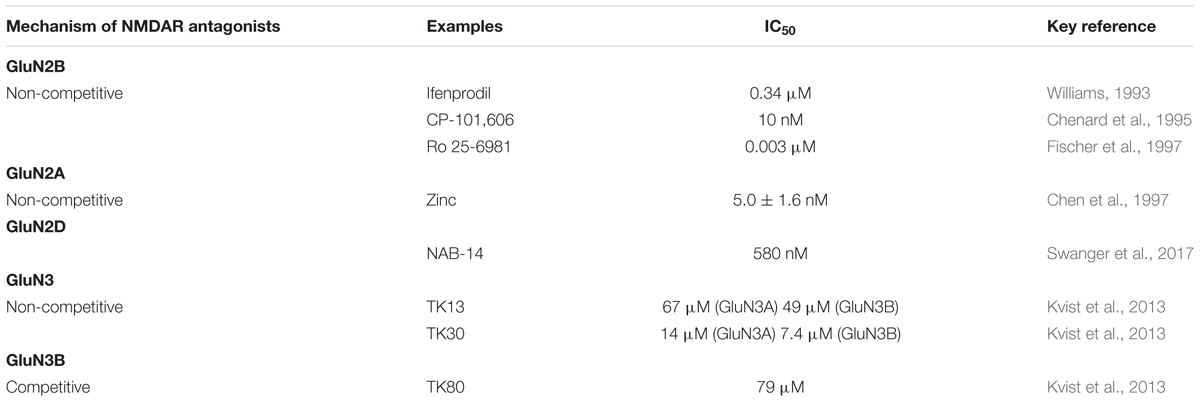
Table 1. Various classes of NMDAR antagonists.
Zinc binds to the leucine/isoleucine/valine binding protein (LIVBP)-like domain of GluN2A, displays a greater than 50-fold selectivity for GluN1/GluN2A over GluN1/GluN2B receptors (Paoletti et al., 1997). GluN2A-selective negative allosteric modulator (NAM) bound LBD heterodimer, corresponding to active and inhibited receptor states reveal a molecular switch in the modulatory binding site that mediate the allosteric inhibition (Yi et al., 2016). Zinc and ifenprodil bind with high affinity to the ATDs of GluN2A and GluN2B, respectively (Zhu and Paoletti, 2015). In hippocampal synapses, zinc decreased the EPSC peak and prolonged the deactivation. Ifenprodil, in contrast, decreased the peak but did not prolong the deactivation (Tovar and Westbrook, 2017).
Binding sites of GluN2A-NTD (N-terminal domain) and GluN2B-NTD retain selective high affinity for their ligand, zinc and ifenprodil. However, each ligand produces only partial inhibition, and maximal inhibition requires occupancy of both GluN2-NTDs by their respective ligands (Hatton and Paoletti, 2005). With regard to the triheteromeric NMDARs, such as GluN1/GluN2A/GluN2B, a single GluN2A or GluN2B subunit is sufficient to confer high affinity to zinc or ifenprodil, but the maximal level of inhibition is greatly reduced compared with diheteromeric NMDARs (Paoletti, 2011). Similarly, GluN1/Glu2A/GluN2C receptors are inhibited by zinc with high potency but low efficacy. Therefore, interactions between homologous NTDs determine the unique pharmacological properties of triheteromeric NMDARs (Paoletti, 2011). The sensitivity of triheteromeric NMDARs to zinc or ifenprodil provides an example of intermediate pharmacology, with reduced potency and efficacy (Hansen et al., 2014; Stroebel et al., 2014).
A recent work identifies a new series of NAMs to study GluN2C and GluN2D function in brain circuits. The prototypical compound, NAB-14, is >800-fold selective for recombinant GluN2C/GluN2D over GluN2A/GluN2B in Xenopus oocytes and has an IC50 value of 580 nM at recombinant GluN2D-containing receptors expressed in mammalian cells, which inhibition reside in the GluN2D M1 transmembrane helix. NAB-14 inhibits GluN2D-mediated synaptic currents in rat subthalamic neurons and mouse hippocampal interneurons, but has no effect on synaptic transmission in hippocampal pyramidal neurons, which do not express GluN2C or GluN2D (Swanger et al., 2017). Perszyk et al. (2016) data provide evidence that hippocampal interneurons contain synaptic NMDARs possessing a GluN2D subunit, which can influence interneuron function and signal processing. These works suggest that selectively modulating brain circuits involving the GluN2C and GluN2D subunits is likely to be developed into clinical therapies (Swanger et al., 2017).
GluN3A subunit was cloned almost two decades ago (Ciabarra et al., 1995; Sucher et al., 1995), followed by cloning of the GluN3B subunit in the beginning of this century (Andersson et al., 2001; Nishi et al., 2001). The GluN3 subunits appears to function as modulatory subunits that reduce the susceptibility of NMDA receptors to Mg2+-blockage and reduce Ca2+-permeability (Perez-Otano et al., 2001; Cavara et al., 2010). Furthermore, GluN3 subunits are involved in synapse maturation (Henson et al., 2012), synaptic plasticity (Larsen et al., 2011), and neuroprotective (Martínez-Turrillas et al., 2012). Overexpression of GluN3A might attenuate NMDAR mediated cell death by reducing Ca2+ permeability of existing NMDARs (Nakanishi et al., 2009). Consistent with the idea that GluN3A might offer neuroprotective benefits, high levels of GluN3A expression during early brain development might explain why excitotoxicity is not more prevalent at ages before the maturation of inhibitory circuitry. GluN3A dysfunction may contribute to neurological disorders involving NMDARs, and the subunit offers an attractive therapeutic target given its distinct pharmacological and structural properties. The study of GluN3A pharmacology is still in its infancy. However, it is clear that GluN3A shares more commonalities with GluN1 than it does with GluN2 subunits. The GluN3A NTD constitutes a crucial regulatory determinant of GluN1/GluN3A receptor function. A hemophilic intersubunit interaction amongst the GluN3A NTDs and the transition region between GluN3A NTD and LBD constitute major structural determinants underlying the low efficacy of glycine-gated GluN1/GluN3A receptors (Mesic et al., 2016). Glycine binds GluN3A with much higher affinity than it binds GluN1, rodent GluN3A, Kd = 40 nM, 650 times less than that for GluN1 (Yao, 2006), and human GluN3A, Kd = 535 nM (Nilsson et al., 2007). D-serine also binds human and rat GluN3A with high affinity (Yao, 2006; Nilsson et al., 2007). Very little is known about how GluN3A-containing receptors can be pharmacologically blocked. Kvist et al. (2013) have succeeded in identifying one novel competitive antagonist, identified 6-hydroxy-[1,2,5] oxadiazolo [3,4-b] pyrazin-5(4H)-one (TK80), with preference for the GluN3B subunit. In addition, they serendipitously identified two novel antagonists, [2-hydroxy-5-((4-(pyridin-3-yl) thiazol-2-yl) amino] benzoic acid (TK13) and 4-(2,4-dichlorobenzoyl)-1H-pyrrole-2-carboxylic acid (TK30), that appear to be non-competitive GluN3 antagonists (Kvist et al., 2013). When GluN1/GluN3A diheteromers are expressed in heterologous systems, they exhibit little electrophysiological block by the classic NMDAR antagonists APV, MK-801, or memantine (Chatterton et al., 2002). Interestingly, GluN1/GluN2/GluN3A triheteromeric receptors may also be blocked by antagonists targeting GluN2 subunits, as receptors composed of GluN1/GluN2B/GluN3A triheteromers and GluN1/GluN2B diheteromers are similarly blocked by high concentrations of the GluN2B antagonist ifenprodil (Smothers and Woodward, 2003). One might predict that, compared to GluN1/GluN2B diheteromers, GluN1/GluN2B/GluN3A triheteromers might be less sensitive to lower concentrations of ifenprodil where GluN2B specificity is greater, as previous studies have shown that the magnitude of ifenprodil block depends on the number of GluN2B subunits contained within the NMDAR complex (Hatton and Paoletti, 2005).
Death-associated protein kinase 1 (DAPK1), Ca/calmodulin (CaM)-dependent protein kinase II α (CaMKIIα), and protein kinase A (PKA) are able to phosphorylate NMDARs. A novel approach for preventing neurotoxicity is to inhibit the phosphorylation of GluN2B-containing NMDARs by CaMKIIα. The CaMKIIα antagonist KN-93, a methoxybenzene sulfonyl derivative that competitively inhibits calmodulin binding to CaM kinase with a Ki of 0.37 μM (Sumi et al., 1991), reduces the effect of CaMKIIα-mediated GluN2B phosphorylation (Farinelli et al., 2012). Protein phosphatase 1 (PP1), highly abundant protein in nerve cells, which co-localizes with the GluN2B subunits of NMDARs, is able to dephosphorylate the Ser1303 residue of GluN2B-containing NMDARs and prevent CaMKII from phosphorylating the receptors, thereby preventing Ca2+ overload inhibiting neurodegeneration (Raveendran et al., 2009; Farinelli et al., 2012; Vizi et al., 2013). Additionally, in the culture cortical neurons, treatment with 17-β-estradiol (E2) and estradiol receptor, the G-protein-coupled receptor 30 (GPR30) agonist G1 (Ki of 11 nM) attenuated the excitotoxicity induced by NMDA exposure. The short-term activation of GPR30 does not affect the expression of either GluN2A- or GluN2B-containing NMDARs; however, it depresses GluN2B subunit phosphorylation at Ser-1303 by inhibiting the dephosphorylation of DAPK1. The neuroprotection mediated by GPR30 is dependent on G-protein-coupled signals and ERK1/2 activation (Liu et al., 2012). Studies have shown that tricyclic desipramine and the serotonin-selective reuptake inhibitor (SSRI) fluoxetine also inhibit NMDA-induced currents in cortical cell cultures. Fluoxetine is a selective inhibitor of GluN2B-containing NMDARs (concentration that causes 50% inhibition, IC50 = 9.74 μM), whereas desipramine inhibits both GluN1/GluN2A (IC50 = 5.10 μM) and GluN1/GluN2B (IC50 = 3.61 μM) subtypes (Kiss et al., 2012). Szasz et al. (2007) observed that inhibition by desipramine (IC50 = 3.13 μM) was voltage-dependent, and the drug was unable to bind to NMDARs that were blocked by Mg2+, whereas the inhibition by fluoxetine (IC50 = 10.51 μM) was voltage-independent, and an association with the NMDAR was still possible in the presence of a Mg2+ block (Yux et al., 1996; Bolo, 2000).
A recent work suggest that the drugs targeting NMDA receptor function or downstream signaling cascades, can restore network and may be effective in treating AD (Zong et al., 2016). At the synapse, the NMDA receptor subunits combined with the post-synaptic related proteins (such as PSD-95) to form a macromolecular complex, which is central for mediating receptor-activated signaling cascades; thus a strategy uncoupling the NMDA receptor from PSD-95 would eliminate excitotoxicity (Morimoto, 2018). It is known that Fyn phosphorylating the GluN2 facilitate interaction of the NMDAR complex with PSD-95 (Nakazawa et al., 2001), linking NMDARs to excitotoxic downstream pathway (Salter and Kalia, 2004). It has been demonstrated that disruption of this interaction prevents excitotoxicity without affecting synaptic NMDA currents (Aarts et al., 2002). Some studies have suggested that agonist binding to NMDARs can signal cascade independently of Ca2+ influx through the receptor. These findings may lead to new pharmacological approaches to target specific synaptic signaling pathway in AD (Hell et al., 2016). Therefore, there is an urgent need to develop novel pharmacological compounds selectively targeting specific receptor subunits and the various signaling pathways (Paoletti et al., 2013).
As Alzheimer’s progresses, brain cells die and synapses are lost, causing cognitive symptoms to worsen. All of the prescription medications currently approved to treat Alzheimer’s symptoms in early to moderate stages are from a class of drugs called ChEIs. The non-competitive NMDA receptor antagonist memantine and a combination of memantine and donepezil are approved by the United States Food and Drug Administration (FDA) for treatment of moderate to severe AD. While current medications cannot stop the damage Alzheimer’s causes to brain cells, they may help lessen or stabilize symptoms for a limited time. The researches focusing mainly on Aβ amyloid deposits and tau protein aggregates forming NFTs have yielded disappointing results. As such, there is a need to develop effective and safe disease modifying treatments that directly target AD pathology and alter the course of AD progression (Ruthirakuhan et al., 2016). It has been motivating researchers to study drugs in earlier stages of the disease, particularly in pre-clinical AD, even before MCI, instead of mild-to-moderate AD trials (Khoury et al., 2017).
AD, related to glutamate-induced excitotoxicity, has been treated by the inhibition of NMDARs. However, clinical trials that have applied high affinity NMDA antagonists have so far failed. This failure has been due to a lack of efficacy and unexpected paradoxical side effects (Ellison, 1995). These negative results can be explained by the opposing role of the different subtypes of NMDARs, and all subtypes are inhibited by the administered drugs. The heteromeric nature of NMDARs provides a wide variety of receptor subtypes, allowing for a rich diversity in receptor signaling properties. While synaptic GluN2A-containing NMDARs activation is involved in neuroprotection, the stimulation of extrasynaptic GluN2B-containing NMDARs triggers cell destruction pathways. The findings that GluN2A- and GluN2B-containing NMDARs can be selectively inhibited by antagonists may be useful in determining pharmacological targets for drug development. In addition, the sensitivity of triheteromeric NMDARs to zinc or ifenprodil with reduced potency and efficacy (Hansen et al., 2014; Stroebel et al., 2014). It will require the generation of NMDAR antagonists that are highly efficient and highly selective (Köhr, 2006). Modern research is trying to discover effective disease-modifying therapies, which specifically target the pathophysiologic signaling, to delay onset and progression of the disease (Tayeb et al., 2012). In future, continuous and sustained efforts are still needed before NMDAR-targeted interventions can transform into effective treatments (Paoletti et al., 2013).
JL and LC wrote most part of the review. YS and HL searched the references and participated in the writing. YW designed the review and revised it.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Aarts, M., Liu, Y., Liu, L., Besshoh, S., Arundine, M., Gurd, J. W., et al. (2002). Treatment of ischemic brain damage by perturbing NMDA receptor-PSD-95 protein interactions. Science 298, 846–850. doi: 10.1126/science.1072873
Abrahamsson, T., Chou, C. Y. C., Li, S. Y., Mancino, A., Costa, R. P., Brock, J. A., et al. (2017). Differential regulation of evoked and spontaneous release by presynaptic NMDA receptors. Neuron 96, 839.e5–855.e855. doi: 10.1016/j.neuron.2017.09.030
Al-Hallaq, R. A., Conrads, T. P., Veenstra, T. D., and Wenthold, R. J. (2007). NMDA di-heteromeric receptor populations and associated proteins in rat hippocampus. J. Neurosci. 27, 8334–8343. doi: 10.1523/JNEUROSCI.2155-07.2007
Almeida, C. G. (2006). Beta-amyloid accumulation impairs multivesicular body sorting by inhibiting the ubiquitin-proteasome system. J. Neurosci. 26, 4277–4288. doi: 10.1523/JNEUROSCI.5078-05.2006
Anaparti, V., Ilarraza, R., Orihara, K., Stelmack, G. L., Ojo, O. O., Mahood, T. H., et al. (2015). NMDA receptors mediate contractile responses in human airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 308, L1253–L1264. doi: 10.1152/ajplung.00402.2014
Anderson, M., Suh, J. M., Kim, E. Y., and Dryer, S. E. (2011). Functional NMDA receptors with atypical properties are expressed in podocytes. Am. J. Physiol. Cell. Physiol. 300, C22–C32. doi: 10.1152/ajpcell.00268.2010
Andersson, O., Stenqvist, A., Attersand, A., and von Euler, G. (2001). Nucleotide sequence, genomic organization, and chromosomal localization of genes encoding the human NMDA receptor subunits NR3A and NR3B. Genomics 78, 178–184. doi: 10.1006/geno.2001.6666
András, I. E., Deli, M. A., Veszelka, S., Hayashi, K., Hennig, B., and Toborek, M. (2007). The NMDA and AMPA KA receptors are involved in glutamate-induced alterations of occludin expression and phosphorylation in brain endothelial cells. J. Cerebr. Blood Flow Mettab. 27, 1431–1443. doi: 10.1038/sj.jcbfm.9600445
Antošová, M., and Strapková, A. (2013). Study of the interaction of glutamatergic and nitrergic signalling in conditions of the experimental airways hyperreactivity. Pharmacol. Rep. 65, 650–657. doi: 10.1016/S1734-1140(13)71042-9
Avila, J., Llorens-Martín, M., Pallas-Bazarra, N., Bolós, M., Perea, J. R., Rodríguez-Matellán, A., et al. (2017). Cognitive decline in neuronal aging and alzheimer’s disease: role of NMDA receptors and associated proteins. Front. Neurosci. 11:626. doi: 10.3389/fnins.2017.00626
Babiec, W. E., Guglietta, R., Jami, S. A., Morishita, W., Malenka, R. C., and O’Dell, T. J. (2014). Ionotropic NMDA receptor signaling is required for the induction of long-term depression in the mouse hippocampal CA1 region. J. Neurosci. 34, 5285–5290. doi: 10.1523/JNEUROSCI.5419-13.2014
Banerjee, A., Larsen, R. S., Philpot, B. D., and Paulsen, O. (2016). Roles of presynaptic NMDA receptors in neurotransmission and plasticity. Trends Neurosci. 39, 26–39. doi: 10.1016/j.tins.2015.11.001
Barria, A., and Malinow, R. (2002). Subunit-specific NMDA receptor trafficking to synapses. Neuron 35, 345–353. doi: 10.1016/S0896-6273(02)00776-6
Barria, A., and Malinow, R. (2005). NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKII. Neuron 48, 289–301. doi: 10.1016/j.neuron.2005.08.034
Basuroy, S., Leffler, C. W., and Parfenova, H. (2013). CORM-A1 prevents blood-brain barrier dysfunction caused by ionotropic glutamate receptor-mediated endothelial oxidative stress and apoptosis. Am. J. Physiol. Cell Physiol. 304, C1105–C1115. doi: 10.1152/ajpcell.00023.2013
Bell, K. F. S., Bennett, D. A., and Cuello, A. C. (2007). Paradoxical upregulation of glutamatergic presynaptic boutons during mild cognitive impairment. J. Neurosci. 27, 10810–10817. doi: 10.1523/JNEUROSCI.3269-07.2007
Benarroch, E. E. (2011). NMDA receptors: recent insights and clinical correlations. Neurology 76, 1750–1757. doi: 10.1212/WNL.0b013e31821b7cc9
Bendová, Z., Sumová, A., and Mikkelsen, J. D. (2009). Circadian and developmental regulation of N-methyl-D-aspartate-receptor 1 mRNA splice variants and N-methyl-D-aspartate-receptor 3 subunit expression within the rat suprachiasmatic nucleus. Neuroscience 159, 599–609. doi: 10.1016/j.neuroscience.2009.01.016
Berg, L. K., Larsson, M., Morland, C., and Gundersen, V. (2013). Pre- and postsynaptic localization of NMDA receptor subunits at hippocampal mossy fibre synapses. Neuroscience 230, 139–150. doi: 10.1016/j.neuroscience.2012.10.061
Bhattacharya, S., Khatri, A., Swanger, S. A., DiRaddo, J. O., Yi, F., Hansen, K. B., Yuan, H., and Traynelis, S. F. (2018). Triheteromeric GluN1/GluN2A/GluN2C NMDARs with unique single-channel properties are the dominant receptor population in cerebellar granule cells. Neuron 99, 315.e5–328.e5. doi: 10.1016/j.neuron.2018.06.010
Bi, H., and Sze, C. (2002). N-methyl-D-aspartate receptor subunit NR2A and NR2B messenger RNA levels are altered in the hippocampus and entorhinal cortex in Alzheimer’s disease. J. Neurol. Sci. 200, 11–18. doi: 10.1016/S0022-510X(02)00087-4
Bi, X., Gall, C. M., Zhou, J., and Lynch, G. (2002). Uptake and pathogenic effects of amyloid beta peptide 1–42 are enhanced by integrin antagonists and blocked by NMDA receptor antagonists. Neuroscience 112, 827–840. doi: 10.1016/S0306-4522(02)00132-X
Bidoret, C., Ayon, A., Barbour, B., and Casado, M. (2009). Presynaptic NR2A-containing NMDA receptors implement a high-pass filter synaptic plasticity rule. Proc. Natl. Acad. Sci. U.S.A. 106, 14126–14131. doi: 10.1073/pnas.0904284106
Binder, L. I., Frankfurter, A., and Rebhun, L. I. (1985). The distribution of tau in the mammalian central nervous system. J. Cell Biol. 101, 1371–1378. doi: 10.1083/jcb.101.4.1371
Birnbaum, J. H., Bali, J., Rajendran, L., Nitsch, R. M., and Tackenberg, C. (2015). Calcium flux-independent NMDA receptor activity is required for Aβ oligomer-induced synaptic loss. Cell Death Dis. 6:e1791. doi: 10.1038/cddis.2015.160
Bolo, N. (2000). Brain pharmacokinetics and tissue distribution in vivo of fluvoxamine and fluoxetine by fluorine magnetic resonance spectroscopy. Neuropsychopharmacoloy 23, 428–438. doi: 10.1016/S0893-133X(00)00116-0
Bonuccelli, U., and Del Dotto, P. (2006). New pharmacologic horizons in the treatment of Parkinson disease. Neurology 67, S30–S38. doi: 10.1212/WNL.67.7_suppl_2.S30
Bordji, K., Becerril-Ortega, J., Nicole, O., and Buisson, A. (2010). Activation of extrasynaptic, but not synaptic, NMDA receptors modifies amyloid precursor protein expression pattern and increases amyloid-ß production. J. Neurosci. 30, 15927–15942. doi: 10.1523/JNEUROSCI.3021-10.2010
Bouvier, G., Bidoret, C., Casado, M., and Paoletti, P. (2015). Presynaptic NMDA receptors: roles and rules. Neuroscience 311, 322–340. doi: 10.1016/j.neuroscience.2015.10.033
Bouvier, G., Larsen, R. S., Rodríguez-Moreno, A., Paulsen, O., and Sjöström, P. J. (2018). Towards resolving the presynaptic NMDA receptor debate. Curr. Opin. Neurobiol. 51, 1–7. doi: 10.1016/j.conb.2017.12.020
Brasier, D. J., and Feldman, D. E. (2008). Synapse-specific expression of functional presynaptic NMDA receptors in rat somatosensory cortex. J. Neurosci. 28, 2199–2211. doi: 10.1523/JNEUROSCI.3915-07.2008
Bu, G., Cam, J., and Zerbinatti, C. (2006). LRP in amyloid-beta production and metabolism. Ann. N. Y. Acad. Sci. 1086, 35–53. doi: 10.1196/annals.1377.005
Burzomato, V., Frugier, G., Pérez-Otaño, I., Kittler, J. T., and Attwell, D. (2010). The receptor subunits generating NMDA receptor mediated currents in oligodendrocytes. J. Physiol. 588, 3403–3414. doi: 10.1113/jphysiol.2010.195503
Busciglio, J., Gabuzda, D. H., Matsudaira, P., and Yankner, B. A. (1993). Generation of beta-amyloid in the secretory pathway in neuronal and nonneuronal cells. Proc. Natl. Acad. Sci. U.S.A. 90, 2092–2096. doi: 10.1073/pnas.90.5.2092
Caldeira, G. L., Ferreira, I. L., and Rego, A. C. (2013). Impaired transcription in Alzheimer’s disease: key role in mitochondrial dysfunction and oxidative stress. J. Alzheimers Dis. 34, 115–131. doi: 10.3233/JAD-121444
Camandola, S., and Mattson, M. P. (2011). Aberrant subcellular neuronal calcium regulation in aging and Alzheimer’s disease. BBA-Mol. Cell Res. 1813, 965–973. doi: 10.1016/j.bbamcr.2010.10.005
Carter, B. C., and Jahr, C. E. (2016). Postsynaptic, not presynaptic NMDA receptors are required for spike-timing-dependent LTD induction. Nat. Neurosci. 19, 1218–1224. doi: 10.1038/nn.4343
Casas-Tinto, S., Zhang, Y., Sanchez-Garcia, J., Gomez-Velazquez, M., Rincon-Limas, D. E., and Fernandez-Funez, P. (2011). The ER stress factor XBP1s prevents amyloid-β neurotoxicity. Hum. Mol. Genet. 20, 2144–2160. doi: 10.1093/hmg/ddr100
Cavara, N. A., Orth, A., Hicking, G., Seebohm, G., and Hollmann, M. (2010). Residues at the tip of the pore loop of NR3B-containing NMDA receptors determine Ca2+ permeability and Mg2+ block. BMC Neurosci. 11:133. doi: 10.1186/1471-2202-11-133
Chang, L., Zhang, Y., Liu, J., Song, Y., Lv, A., Li, Y., et al. (2016). Differential regulation of N-methyl-D-aspartate receptor subunits is an early events in the actions of soluble amyloid-β1-40 oligomers on hippocampal neurons. J. Alzheimers Dis. 51, 197–212. doi: 10.3233/JAD-150942
Chatterton, J. E., Awobuluyi, M., Premkumar, L. S., Takahashi, H., Talantova, M., Shin, Y., et al. (2002). Excitatory glycine receptors containing the NR3 family of NMDA receptor subunits. Nature 415, 793–798. doi: 10.1038/nature715
Chen, N., Moshaver, A., and Raymond, L. A. (1997). Differential sensitivity of recombinant N-methyl-D-aspartate receptor subtypes to zinc inhibition. Mol. Pharmacol. 51:1015. doi: 10.1124/mol.51.6.1015
Chen, Q., Zhou, Z., Zhang, L., Wang, Y., Zhang, Y., Zhong, M., et al. (2012). Tau protein is involved in morphological plasticity in hippocampal neurons in response to BDNF. Neurochem. Int. 60, 233–242. doi: 10.1016/j.neuint.2011.12.013
Chenard, B. L., Bordner, J., Butler, T. W., Chambers, L. K., Collins, M. A., De Costa, D. L., et al. (1995). (1S,2S)-1-(4-Hydroxyphenyl)-2-(4-hydroxy-4-phenylpiperidino)-1-propanol: a potent new neuroprotectant which blocks N-Methyl-D-aspartate responses. J. Med. Chem. 38, 3138–3145. doi: 10.1021/jm00016a017
Choi, D. W. (1987). Ionic dependence of glutamate neurotoxicity. J. Neurosci. 7, 369–379. doi: 10.1523/JNEUROSCI.07-02-00369.1987
Choi, D. W. (1988). Glutamate neurotoxicity and diseases of the nervous system. Neuron 1, 623–634. doi: 10.1016/0896-6273(88)90162-6
Choi, D. W., Koh, J. Y., and Peters, S. (1988). Pharmacology of glutamate neurotoxicity in cortical cell culture: attenuation by NMDA antagonists. J. Neurosci. 8, 185–196. doi: 10.1523/JNEUROSCI.08-01-00185.1988
Chung, C. (2013). NMDA receptor as a newly identified member of the metabotropic glutamate receptor family: clinical implications for neurodegenerative diseases. Mol. Cells 36, 99–104. doi: 10.1007/s10059-013-0113-y
Ciabarra, A. M., Sullivan, J. M., Gahn, L. G., Pecht, G., Heinemann, S., and Sevarino, K. A. (1995). Cloning and characterization of chi-1: a developmentally regulated member of a novel class of the ionotropic glutamate receptor family. J. Neurosci. 15, 6498–6508. doi: 10.1523/JNEUROSCI.15-10-06498.1995
Cirrito, J. R., Yamada, K. A., Finn, M. B., Sloviter, R. S., Bales, K. R., May, P. C., et al. (2005). Synaptic activity regulates interstitial fluid Amyloid-β levels in vivo. Neuron 48, 913–922. doi: 10.1016/j.neuron.2005.10.028
Cleary, J. P., Walsh, D. M., Hofmeister, J. J., Shankar, G. M., Kuskowski, M. A., Selkoe, D. J., et al. (2005). Natural oligomers of the amyloid-β protein specifically disrupt cognitive function. Nat. Neurosci. 8, 79–84. doi: 10.1038/nn1372
Cook, D. G., Forman, M. S., Sung, J. C., Leight, S., Kolson, D. L., Iwatsubo, T., et al. (1997). Alzheimer’s A beta(1-42) is generated in the endoplasmic reticulum/intermediate compartment of NT2N cells. Nat. Med. 3, 1021–1023. doi: 10.1038/nm0997-1021
Corlew, R., Wang, Y., Ghermazien, H., Erisir, A., and Philpot, B. D. (2007). Developmental switch in the contribution of presynaptic and postsynaptic NMDA receptors to long-term depression. J. Neurosci. 27, 9835–9845. doi: 10.1523/JNEUROSCI.5494-06.2007
Costa, R. O., Lacor, P. N., Ferreira, I. L., Resende, R., Auberson, Y. P., Klein, W. L., et al. (2012). Endoplasmic reticulum stress occurs downstream of GluN2B subunit of N-methyl-D-aspartate receptor in mature hippocampal cultures treated with amyloid-β oligomers. Aging Cell 11, 823–833. doi: 10.1111/j.1474-9726.2012.00848.x
Cull-Candy, S., Brickley, S., and Farrant, M. (2001). NMDA receptor subunits: diversity, development and disease. Curr. Opin. Neurobiol. 11, 327–335. doi: 10.1016/s0959-4388(00)00215-4
Dalva, M. B., Takasu, M. A., Lin, M. Z., Shamah, S. M., Hu, L., Gale, N. W., et al. (2000). EphB Receptors interact with NMDA receptors and regulate excitatory synapse formation. Cell 103, 945–956. doi: 10.1016/S0092-8674(00)00197-5
Danysz, W., and Parsons, C. G. (2003). The NMDA receptor antagonist memantine as a symptomatological and neuroprotective treatment for Alzheimer’s disease: preclinical evidence. Int. J. Geriatr. Psychol. 18, S23–S32. doi: 10.1002/gps.938
Danysz, W., and Parsons, C. G. (2012). Alzheimer’s disease, β-amyloid, glutamate, NMDA receptors and memantine – Searching for the connections. Br. J. Pharmacol. 167, 324–352. doi: 10.1111/j.1476-5381.2012.02057.x
De Felice, F. G., Velasco, P. T., Lambert, M. P., Viola, K., Fernandez, S. J., Ferreira, S. T., et al. (2007). Abeta oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J. Biol. Chem. 282, 11590–11601. doi: 10.1074/jbc.M607483200
Deane, R., Du Yan, S., Submamaryan, R. K., LaRue, B., Jovanovic, S., Hogg, E., et al. (2003). RAGE mediates amyloid-β peptide transport across the blood-brain barrier and accumulation in brain. Nat. Med. 9, 907–913. doi: 10.1038/nm890
Deardorff, W. J., and Grossberg, G. T. (2016). Pharmacotherapeutic strategies in the treatment of severe Alzheimer’s disease. Expert Opin. Pharmacother. 17, 1789–1800. doi: 10.1080/14656566.2016.1215431
Decker, J. M., Krüger, L., Sydow, A., Dennissen, F. J., Siskova, Z., Mandelkow, E., et al. (2016). The Tau/A152T mutation, a risk factor for frontotemporal-spectrum disorders, leads to NR2B receptor-mediated excitotoxicity. EMBO Rep. 17, 552–569. doi: 10.15252/embr.201541439
Delacourte, A., Flament, S., Dibe, E. M., Hublau, P., Sablonnière, B., Hémon, B., et al. (1990). Pathological proteins Tau 64 and 69 are specifically expressed in the somatodendritic domain of the degenerating cortical neurons during Alzheimer’s disease. Acta Neuropathol. 80, 111–117. doi: 10.1007/BF00308912
Deng, A., and Thomson, S. C. (2009). Renal NMDA receptors independently stimulate proximal reabsorption and glomerular filtration. Am. J. Physiol. Renal Physiol. 296, 976–982. doi: 10.1152/ajprenal.90391.2008
Dineley, K. T., Bell, K. A., Bui, D., and Sweatt, J. D. (2002). β-amyloid peptide activates α7 nicotinic acetylcholine receptors expressed in xenopus oocytes. J. Biol. Chem. 277, 25056–25061. doi: 10.1074/jbc.M200066200
Domingues, A., Almeida, S., Da, C. E. S. E., Oliveira, C. R., and Rego, A. C. (2007). Toxicity of beta-amyloid in HEK293 cells expressing NR1/NR2A or NR1/NR2B N-methyl-D-aspartate receptor subunits. Neurochem. Int. 50, 872–880. doi: 10.1016/j.neuint.2007.03.001
Dore, K., Stein, I. S., Brock, J. A., Castillo, P. E., Zito, K., and Sjöström, P. J. (2017). unconventional NMDA receptor signaling. J. Neurosci. 37, 10800–10807. doi: 10.1523/JNEUROSCI.1825-17.2017
Duguid, I. C. (2013). Presynaptic NMDA receptors: are they dendritic receptors in disguise? Brain Res. Bull. 93, 4–9. doi: 10.1016/j.brainresbull.2012.12.004
Dunah, A. W., Luo, J., Wang, Y. H., Yasuda, R. P., and Wolfe, B. B. (1998). Subunit composition of N-methyl-D-aspartate receptors in the central nervous system that contain the NR2D subunit. Mol. Pharmacol. 53, 429–437. doi: 10.1124/mol.53.3.429
Dzamba, D., Honsa, P., and Anderova, M. (2013). NMDA receptors in glial cells: pending questions. Curr. Neuropharmacol. 11, 250–262. doi: 10.2174/1570159X11311030002
Ellison, G. (1995). The N-methyl-D-aspartate antagonists phencyclidine, ketamine and dizocilpine as both behavioral and anatomical models of the dementias. Brain Res. Brain Res. Rev. 20, 250–267. doi: 10.1016/0165-0173(94)00014-G
Epstein, F. H., Lipton, S. A., and Rosenberg, P. A. (1994). Excitatory amino acids as a final common pathway for neurologic disorders. New Engl. J. Med. 330, 613–622. doi: 10.1056/NEJM199403033300907
Erreger, K., Dravid, S. M., Banke, T. G., Wyllie, D. J. A., and Traynelis, S. F. (2005). Subunit-specific gating controls rat NR1/NR2A and NR1/NR2B NMDA channel kinetics and synaptic signalling profiles. J. Physiol. 563, 345–358. doi: 10.1113/jphysiol.2004.080028
Esclaire, F. O., Lesort, M., Blanchard, C., and Hugon, J. (1997). Glutamate toxicity enhances tau gene expression in neuronal cultures. J. Neurosci. Res. 49, 309–318. doi: 10.1002/(SICI)1097-4547(19970801)49:3<309::AID-JNR6<3.0.CO;2-G
Fan, M., and Raymond, L. (2007). N-Methyl-D-aspartate (NMDA) receptor function and excitotoxicity in Huntington’s disease. Prog. Neurobiol. 81, 272–293. doi: 10.1016/j.pneurobio.2006.11.003
Farinelli, M., Heitz, F. D., Grewe, B. F., Tyagarajan, S. K., Helmchen, F., and Mansuy, I. M. (2012). Selective regulation of NR2B by protein phosphatase-1 for the control of the NMDA receptor in neuroprotection. PLoS One 7:e34047. doi: 10.1371/journal.pone.0034047
Fellin, T., Pascual, O., Gobbo, S., Pozzan, T., Haydon, P. G., and Carmignoto, G. (2004). Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors. Neuron 43, 729–743. doi: 10.1016/j.neuron.2004.08.011
Fernandes, A., Wojcik, T., Baireddy, P., Pieschl, R., Newton, A., Tian, Y., et al. (2015). Inhibition of in vivo [3H]MK-801 binding by NMDA receptor open channel blockers and GluN2B antagonists in rats and mice. Eur. J. Pharmacol. 766, 1–8. doi: 10.1016/j.ejphar.2015.08.044
Ferreira, I. L., Bajouco, L. M., Mota, S. I., Auberson, Y. P., Oliveira, C. R., and Rego, A. C. (2012). Amyloid beta peptide 1–42 disturbs intracellular calcium homeostasis through activation of GluN2B-containing N-methyl-D-aspartate receptors in cortical cultures. Cell Calcium 51, 95–106. doi: 10.1016/j.ceca.2011.11.008
Ferreira, I. L., Ferreiro, E., Schmidt, J., Cardoso, J. M., Pereira, C. M. F., Carvalho, A. L., et al. (2015). Aβ and NMDAR activation cause mitochondrial dysfunction involving ER calcium release. Neurobiol. Aging 36, 680–692. doi: 10.1016/j.neurobiolaging.2014.09.006
Ferreira, I. L., Resende, R., Ferreiro, E., Rego, A. C., and Pereira, C. F. (2010). Multiple defects in energy metabolism in Alzheimer’s disease. Curr. Drug Targets 11:1193. doi: 10.2174/1389450111007011193
Fischer, G., Mutel, V., Trube, G., Malherbe, P., Kew, J. N., Mohacsi, E., et al. (1997). Ro 25-6981, a highly potent and selective blocker of N-methyl-D-aspartate receptors containing the NR2B subunit, characterization in vitro. J. Pharmacol. Exp. Ther. 283, 1285–1292.
Fonseca, A. C. R. G., Ferreiro, E., Oliveira, C. R., Cardoso, S. M., and Pereira, C. F. (2013). Activation of the endoplasmic reticulum stress response by the amyloid-beta 1–40 peptide in brain endothelial cells. BBA-Mol. Basis Dis. 1832, 2191–2203. doi: 10.1016/j.bbadis.2013.08.007
Foster, T. C., Kyritsopoulos, C., and Kumar, A. (2017). Central role for NMDA receptors in redox mediated impairment of synaptic function during aging and Alzheimer’s disease. Behav. Brain Res. 322, 223–232. doi: 10.1016/j.bbr.2016.05.012
Fox, C., Crugel, M., Maidment, I., Auestad, B. H., Coulton, S., Treloar, A., et al. (2012). Efficacy of memantine for agitation in alzheimer’s dementia: a randomised double-blind placebo controlled trial. PLoS One 7:e35185. doi: 10.1371/journal.pone.0035185
Franco, R., Aguinaga, D., Reyes, I., Canela, E. I., Lillo, J., Tarutani, A., et al. (2018). N-Methyl-D-aspartate receptor link to the MAP kinase pathway in cortical and hippocampal neurons and microglia is dependent on calcium sensors and is blocked by α-synuclein, tau, and phospho-tau in non-transgenic and transgenic APPSw, ind mice. Front. Mol. Neurosci. 11:273. doi: 10.3389/fnmol.2018.00273
Gerard, F., and Hansson, E. (2012). Inflammatory activation enhances NMDA-triggered Ca2+ signalling and IL-1beta secretion in primary cultures of rat astrocytes. Brain Res. 1473, 1–8. doi: 10.1016/j.brainres.2012.07.032
Glabe, C. G. (2008). Structural classification of toxic amyloid oligomers. J. Biol. Chem. 283, 29639–29643. doi: 10.1074/jbc.R800016200
Gladding, C. M., and Raymond, L. A. (2011). Mechanisms underlying NMDA receptor synaptic/extrasynaptic distribution and function. Mol. Cell Neurosci. 48, 308–320. doi: 10.1016/j.mcn.2011.05.001
Graef, J. D., Newberry, K., Newton, A., Pieschl, R., Shields, E., Luan, F., et al. (2015). Effect of acute NR2B antagonist treatment on long-term potentiation in the rat hippocampus. Brain Res. 1609, 31–39. doi: 10.1016/j.brainres.2015.03.019
Graham, W. V., Bonito-Oliva, A., and Sakmar, T. P. (2017). Update on Alzheimer’s disease therapy and prevention strategies. Annu. Rev. Med. 68, 413–430. doi: 10.1146/annurev-med-042915-103753
Groc, L., Choquet, D., Stephenson, F. A., Verrier, D., Manzoni, O. J., and Chavis, P. (2007). NMDA receptor surface trafficking and synaptic subunit composition are developmentally regulated by the extracellular matrix protein reelin. J. Neurosci. 27, 10165–10175. doi: 10.1523/JNEUROSCI.1772-07.2007
Groc, L., Heine, M., Cousins, S. L., Stephenson, F. A., Lounis, B., Cognet, L., et al. (2006). NMDA receptor surface mobility depends on NR2A-2B subunits. Proc. Natl. Acad. Sci. U.S.A. 103, 18769–18774. doi: 10.1073/pnas.0605238103
Grochowska, K. M., Yuanxiang, P., Bär, J., Raman, R., Brugal, G., Sahu, G., et al. (2017). Posttranslational modification impact on the mechanism by which amyloid-β induces synaptic dysfunction. EMBO Rep. 18, 962–981. doi: 10.15252/embr.201643519
Grothe, M. J., Barthel, H., Sepulcre, J., Dyrba, M., Sabri, O., and Teipel, S. J. (2017). In vivo staging of regional amyloid deposition. Neurology 89, 2031–2038. doi: 10.1212/WNL.0000000000004643
Groveman, B. R., Feng, S., Fang, X., Pflueger, M., Lin, S., Bienkiewicz, E. A., et al. (2012). The regulation of N-methyl-D-aspartate receptors by Src kinase. FEBS J. 279, 20–28. doi: 10.1111/j.1742-4658.2011.08413.x
Haas, L. T., Kostylev, M. A., and Strittmatter, S. M. (2014). Therapeutic molecules and endogenous ligands regulate the interaction between brain cellular prion protein (PrPC) and metabotropic glutamate receptor 5 (mGluR5). J. Biol. Chem. 289, 28460–28477. doi: 10.1074/jbc.M114.584342
Hackos, D. H., and Hanson, J. E. (2017). Diverse modes of NMDA receptor positive allosteric modulation: mechanisms and consequences. Neuropharmacology 112, 34–45. doi: 10.1016/j.neuropharm.2016.07.037
Haghikia, A., Mergia, E., Friebe, A., Eysel, U. T., Koesling, D., and Mittmann, T. (2007). Long-term potentiation in the visual cortex requires both nitric oxide receptor guanylyl cyclases. J. Neurosci. 27, 818–823. doi: 10.1523/JNEUROSCI.4706-06.2007
Hamilton, N. B., Kolodziejczyk, K., Kougioumtzidou, E., and Attwell, D. (2016). Proton-gated Ca(2+)-permeable TRP channels damage myelin in conditions mimicking ischaemia. Nature 529, 523–527. doi: 10.1038/nature16519
Hansen, K. B., Ogden, K. K., Yuan, H., and Traynelis, S. F. (2014). Distinct functional and pharmacological properties of triheteromeric GluN1/GluN2A/GluN2B NMDA receptors. Neuron 81, 1084–1096. doi: 10.1016/j.neuron.2014.01.035
Hansson Petersen, C. A., Alikhani, N., Behbahani, H., Wiehager, B., Pavlov, P. F., Alafuzoff, I., et al. (2008). The amyloid -peptide is imported into mitochondria via the TOM import machinery and localized to mitochondrial cristae. Proc. Natl. Acad. Sci. U.S.A. 105, 13145–13150. doi: 10.1073/pnas.0806192105
Hardingham, G. E., and Bading, H. (2010). Synaptic versus extrasynaptic NMDA receptor signalling: implications for neurodegenerative disorders. Nat. Rev. Neurosci. 11, 682–696. doi: 10.1038/nrn2911
Hardingham, G. E., Fukunaga, Y., and Bading, H. (2002). Extrasynaptic NMDARs oppose synaptic NMDARs by triggering CREB shut-off and cell death pathways. Nat. Neurosci. 5, 405–414. doi: 10.1038/nn835
Hardy, J. (2002). The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science 297, 353–356. doi: 10.1126/science.1072994
Harris, A. Z., and Pettit, D. L. (2007). Extrasynaptic and synaptic NMDA receptors form stable and uniform pools in rat hippocampal slices. J. Physiol. 584, 509–519. doi: 10.1113/jphysiol.2007.137679
Harris, A. Z., and Pettit, D. L. (2008). Recruiting extrasynaptic NMDA receptors augments synaptic signaling. J. Neurophysiol. 99, 524–533. doi: 10.1152/jn.01169.2007
Hartmann, T., Bieger, S. C., Brühl, B., Tienari, P. J., Ida, N., Allsop, D., et al. (1997). Distinct sites of intracellular production for Alzheimer’s disease A beta40/42 amyloid peptides. Nat. Med. 3:1016. doi: 10.1038/nm0997-1016
Hatton, C. J., and Paoletti, P. (2005). Modulation of triheteromeric NMDA receptors by N-terminal domain ligands. Neuron 46, 261–274. doi: 10.1016/j.neuron.2005.03.005
Hell, J. W., Gray, J. A., and Zito, K. (2016). Non-ionotropic signaling by the NMDA receptor: controversy and opportunity. F1000Res. 5:1010. doi: 10.12688/f1000research.8366.1
Henson, M. A., Larsen, R. S., Lawson, S. N., Pérez-Otaño, I., Nakanishi, N., Lipton, S. A., et al. (2012). Genetic deletion of NR3A accelerates glutamatergic synapse maturation. PLoS One 7:e42327. doi: 10.1371/journal.pone.0042327
Henson, M. A., Roberts, A. C., Pérez-Otaño, I., and Philpot, B. D. (2010). Influence of the NR3A subunit on NMDA receptor functions. Prog. Neurobiol. 91, 23–37. doi: 10.1016/j.pneurobio.2010.01.004
Herman, M. A., and Jahr, C. E. (2007). Extracellular glutamate concentration in hippocampal slice. J. Neurosci. 27, 9736–9741. doi: 10.1523/JNEUROSCI.3009-07.2007
Hinoi, E., Fujimori, S., and Yoneda, Y. (2003). Modulation of cellular differentiation by N-methyl-D-aspartate receptors in osteoblasts. FASEB J. 17, 1532–1534. doi: 10.1096/fj.02-0820fje
Hoey, S. E., Williams, R. J., and Perkinton, M. S. (2009). Synaptic NMDA receptor activation stimulates alpha-secretase amyloid precursor protein processing and inhibits amyloid-beta production. J. Neurosci. 29, 4442–4460. doi: 10.1523/JNEUROSCI.6017-08.2009
Hogan-Cann, A. D., and Anderson, C. M. (2016). Physiological roles of non-neuronal NMDA receptors. Trends Pharmacol. Sci. 37, 750–767. doi: 10.1016/j.tips.2016.05.012
Hsieh, H., Boehm, J., Sato, C., Iwatsubo, T., Tomita, T., Sisodia, S., et al. (2006). AMPAR removal underlies Aβ-induced synaptic depression and dendritic spine loss. Neuron 52, 831–843. doi: 10.1016/j.neuron.2006.10.035
Hunter, S., Smailagic, N., and Brayne, C. (2018). Aβ and the dementia syndrome: simple versus complex perspectives. Eur. J. Clin. Invest. 48:e13025. doi: 10.1111/eci.13025
Hynd, M. R., Scott, H. L., and Dodd, P. R. (2001). Glutamate(NMDA) receptor NR1 subunit mRNA expression in Alzheimer’s disease. J. Neurochem. 78:175. doi: 10.1046/j.1471-4159.2001.00409.x
Hynd, M. R., Scott, H. L., and Dodd, P. R. (2004). Differential expression of N-methyl-D-aspartate receptor NR2 isoforms in Alzheimer’s disease. J. Neurochem. 90, 913–919. doi: 10.1111/j.1471-4159.2004.02548.x
Inagaki, N., Kuromi, H., Gonoi, T., Okamoto, Y., Ishida, H., Seino, Y., et al. (1995). Expression and role of ionotropic glutamate receptors in pancreatic islet cells. FASEB J. 9, 686–691. doi: 10.1096/fasebj.9.8.7768362
Ittner, L. M., Ke, Y. D., Delerue, F., Bi, M., Gladbach, A., van Eersel, J., et al. (2010). Dendritic function of tau mediates amyloid-β toxicity in alzheimer’s disease mouse models. Cell 142, 387–397. doi: 10.1016/j.cell.2010.06.036
Jacob, C. P., Koutsilieri, E., Bartl, J., Neuen-Jacob, E., Arzberger, T., Zander, N., et al. (2007). Alterations in expression of glutamatergic transporters and receptors in sporadic Alzheimer’s disease. J. Alzheimers Dis. 11, 97–116. doi: 10.3233/JAD-2007-11113
Jagust, W. (2018). Imaging the evolution and pathophysiology of Alzheimer disease. Nat. Rev. Neurosci. 19, 687–700. doi: 10.1038/s41583-018-0067-3
Jimenez-Blasco, D., Santofimia-Castaño, P., Gonzalez, A., Almeida, A., and Bolaños, J. P. (2015). Astrocyte NMDA receptors’ activity sustains neuronal survival through a Cdk5–Nrf2 pathway. Cell Death Differ. 22, 1877–1889. doi: 10.1038/cdd.2015.49
Jourdain, P., Bergersen, L. H., Bhaukaurally, K., Bezzi, P., Santello, M., Domercq, M., et al. (2007). Glutamate exocytosis from astrocytes controls synaptic strength. Nat. Neurosci. 10, 331–339. doi: 10.1038/nn1849
Kaindl, A. M., Degos, V., Peineau, S., Gouadon, E., Chhor, V., Loron, G., et al. (2012). Activation of microglial N-methyl-D-aspartate receptors triggers inflammation and neuronal cell death in the developing and mature brain. Ann. Neurol. 72, 536–549. doi: 10.1002/ana.23626
Kamenetz, F., Tomita, T., Hsieh, H., Seabrook, G., Borchelt, D., Iwatsubo, T., et al. (2003). APP processing and synaptic function. Neuron 37, 925–937. doi: 10.1016/S0896-6273(03)00124-7
Káradóttir, R., Cavelier, P., Bergersen, L. H., and Attwell, D. (2005). NMDA receptors are expressed in oligodendrocytes and activated in ischaemia. Nature 438, 1162–1166. doi: 10.1038/nature04302
Karakas, E., and Furukawa, H. (2014). Crystal structure of a heterotetrameric NMDA receptor ion channel. Science 344, 992–997. doi: 10.1126/science.1251915
Kato, H., Narita, M., Miyatake, M., Yajima, Y., and Suzuki, T. (2006). Role of neuronal NR2B subunit-containing NMDA receptor-mediated Ca2+ influx and astrocytic activation in cultured mouse cortical neurons and astrocytes. Synapse 59, 10–17. doi: 10.1002/syn.20213
Kavalali, E. T. (2015). The mechanisms and functions of spontaneous neurotransmitter release. Nat. Rev. Neurosci. 16, 5–16. doi: 10.1038/nrn3875
Kessels, H. W., Nabavi, S., and Malinow, R. (2013). Metabotropic NMDA receptor function is required for -amyloid-induced synaptic depression. Proc. Natl. Acad. Sci. U.S.A. 110, 4033–4038. doi: 10.1073/pnas.1219605110
Khoury, R., Patel, K., Gold, J., Hinds, S., and Grossberg, G. T. (2017). Recent progress in the pharmacotherapy of Alzheimer’s disease. Drug Aging 34, 811–820. doi: 10.1007/s40266-017-0499-x
Kim, J., Anwyl, R., Suh, Y., Djamgoz, M. B. A., and Rowan, M. J. (2001). Use-dependent effects of amyloidogenic fragments of {beta}-amyloid precursor protein on synaptic plasticity in rat hippocampus in vivo. J. Neurosci. 21, 1327–1333. doi: 10.1523/JNEUROSCI.21-04-01327.2001
Kim, J. H., Liao, D., Lau, L., and Huganir, R. L. (1998). SynGAP: a synaptic RasGAP that associates with the PSD-95/SAP90 protein family. Neuron 20, 683–691. doi: 10.1016/S0896-6273(00)81008-9
Kimura, T., Whitcomb, D. J., Jo, J., Regan, P., Piers, T., Heo, S., et al. (2014). Microtubule-associated protein tau is essential for long-term depression in the hippocampus. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369:20130144. doi: 10.1098/rstb.2013.0144
Kinoshita, A. (2003). Demonstration by FRET of BACE interaction with the amyloid precursor protein at the cell surface and in early endosomes. J. Cell Sci. 116, 3339–3346. doi: 10.1242/jcs.00643
Kiss, J. P., Szasz, B. K., Fodor, L., Mike, A., Lenkey, N., Kurkó, D., et al. (2012). GluN2B-containing NMDA receptors as possible targets for the neuroprotective and antidepressant effects of fluoxetine. Neurochem. Int. 60, 170–176. doi: 10.1016/j.neuint.2011.12.005
Kodis, E. J., Choi, S., Swanson, E., Ferreira, G., and Bloom, G. S. (2018). N-methyl-D-aspartate receptor-mediated calcium influx connects amyloid-β oligomers to ectopic neuronal cell cycle reentry in Alzheimer’s disease. Alzheimers Dement. 14, 1302–1312. doi: 10.1016/j.jalz.2018.05.017
Köhr, G. (2006). NMDA receptor function: subunit composition versus spatial distribution. Cell Tissue Res. 326, 439–446. doi: 10.1007/s00441-006-0273-6
Köhr, G. (2007). NMDA receptor mobility: cultures versus acute brain slices or neonatalversus mature synapses? J. Physiol. 584, 367–367. doi: 10.1113/jphysiol.2007.144865
Kong, M., Ba, M., Liu, C., Zhang, Y., Zhang, H., and Qiu, H. (2015). NR2B antagonist CP-101,606 inhibits NR2B phosphorylation at tyrosine-1472 and its interactions with Fyn in levodopa-induced dyskinesia rat model. Behav. Brain Res. 282, 46–53. doi: 10.1016/j.bbr.2014.12.059
Krapivinsky, G., Krapivinsky, L., Manasian, Y., Ivanov, A., Tyzio, R., Pellegrino, C., et al. (2003). The NMDA receptor is coupled to the ERK pathway by a direct interaction between NR2B and RasGRF1. Neuron 40, 775–784. doi: 10.1016/S0896-6273(03)00645-7
Kullmann, D. M., and Lamsa, K. P. (2007). Long-term synaptic plasticity in hippocampal interneurons. Nat. Rev. Neurosci. 8, 687–699. doi: 10.1038/nrn2207
Kunz, P. A., Roberts, A. C., and Philpot, B. D. (2013). Presynaptic NMDA receptor mechanisms for enhancing spontaneous neurotransmitter release. J. Neurosci. 33, 7762–7769. doi: 10.1523/JNEUROSCI.2482-12.2013
Kurup, P., Zhang, Y., Xu, J., Venkitaramani, D. V., Haroutunian, V., Greengard, P., et al. (2010). Abeta-mediated NMDA receptor endocytosis in Alzheimer’s disease involves ubiquitination of the tyrosine phosphatase STEP61. J. Neurosci. 30, 5948–5957. doi: 10.1523/JNEUROSCI.0157-10.2010
Kvist, T., Greenwood, J. R., Hansen, K. B., Traynelis, S. F., and Bräuner-Osborne, H. (2013). Structure-based discovery of antagonists for GluN3-containing N-methyl-D-aspartate receptors. Neuropharmacology 75, 324–336. doi: 10.1016/j.neuropharm.2013.08.003
Lacor, P. N., Buniel, M. C., Furlow, P. W., Clemente, A. S., Velasco, P. T., Wood, M., et al. (2007). Abeta oligomer-induced aberrations in synapse composition, shape, and density provide a molecular basis for loss of connectivity in Alzheimer’s disease. J. Neurosci. 27, 796–807. doi: 10.1523/JNEUROSCI.3501-06.2007
LaFerla, F. M., Green, K. N., and Oddo, S. (2007). Intracellular amyloid-β in Alzheimer’s disease. Nat. Rev. Neurosci. 8, 499–509. doi: 10.1038/nrn2168
Lalo, U. (2006). NMDA receptors mediate neuron-to-glia signaling in mouse cortical astrocytes. J. Neurosci. 26, 2673–2683. doi: 10.1523/JNEUROSCI.4689-05.2006
Larsen, R. S., Corlew, R. J., Henson, M. A., Roberts, A. C., Mishina, M., Watanabe, M., et al. (2011). NR3A-containing NMDARs promote neurotransmitter release and spike timing-dependent plasticity. Nat. Neurosci. 14, 338–344. doi: 10.1038/nn.2750
Larsen, R. S., Smith, I. T., Miriyala, J., Han, J. E., Corlew, R. J., Smith, S. L., et al. (2014). Synapse-specific control of experience-dependent plasticity by presynaptic NMDA receptors. Neuron 83, 879–893. doi: 10.1016/j.neuron.2014.07.039
Lau, C. G., Takeuchi, K., Rodenas-Ruano, A., Takayasu, Y., Murphy, J., Bennett, M. V. L., et al. (2009). Regulation of NMDA receptor Ca2+ signalling and synaptic plasticity. Biochem. Soc. Trans. 37, 1369–1374. doi: 10.1042/BST0371369
Lazarov, O., Lee, M., Peterson, D. A., and Sisodia, S. S. (2002). Evidence that synaptically released β-amyloid accumulates as extracellular deposits in the hippocampus of transgenic mice. J. Neurosci. 22, 9785–9793. doi: 10.1523/JNEUROSCI.22-22-09785.2002
Lee, C., Lü, W., Michel, J. C., Goehring, A., Du, J., Song, X., et al. (2014). NMDA receptor structures reveal subunit arrangement and pore architecture. Nature 511, 191–197. doi: 10.1038/nature13548
Lee, G. (2004). Phosphorylation of tau by fyn: implications for Alzheimer’s disease. J. Neurosci. 24, 2304–2312. doi: 10.1523/JNEUROSCI.4162-03.2004
Lee, G., Newman, S. T., Gard, D. L., Band, H., and Panchamoorthy, G. (1998). Tau interacts with src-family non-receptor tyrosine kinases. J. Cell Sci. 111(Pt 21), 3167.
Lee, M., Ting, K. K., Adams, S., Brew, B. J., Chung, R., and Guillemin, G. J. (2010). Characterisation of the expression of NMDA receptors in human astrocytes. PLoS One 5:e14123. doi: 10.1371/journal.pone.0014123
Lesne, S. (2005). NMDA receptor activation inhibits alpha-secretase and promotes neuronal amyloid- production. J. Neurosci. 25, 9367–9377. doi: 10.1523/JNEUROSCI.0849-05.2005
Leveille, F., El, G. F., Gouix, E., Lecocq, M., Lobner, D., Nicole, O., et al. (2008). Neuronal viability is controlled by a functional relation between synaptic and extrasynaptic NMDA receptors. FASEB J. 22, 4258–4271. doi: 10.1096/fj.08-107268
Li, C., Xiao, L., Liu, X., Yang, W., Shen, W., Hu, C., et al. (2013). A functional role of NMDA receptor in regulating the differentiation of oligodendrocyte precursor cells and remyelination. Glia 61, 732–749. doi: 10.1002/glia.22469
Li, J., Zhao, L., Cui, B., Deng, L., Ning, G., and Liu, J. (2011). Multiple signaling pathways involved in stimulation of osteoblast differentiation by N-methyl-D-aspartate receptors activation in vitro. Acta Pharmacol. Sin. 32, 895–903. doi: 10.1038/aps.2011.38
Li, S., Hong, S., Shepardson, N. E., Walsh, D. M., Shankar, G. M., and Selkoe, D. (2009). Soluble oligomers of amyloid β protein facilitate hippocampal long-term depression by disrupting neuronal glutamate uptake. Neuron 62, 788–801. doi: 10.1016/j.neuron.2009.05.012
Li, Y., Chang, L., Song, Y., Gao, X., Roselli, F., Liu, J., et al. (2016). Astrocytic GluN2A and GluN2B oppose the synaptotoxic effects of amyloid-β1-40 in hippocampal cells. J. Alzheimers Dis. 54, 135–148. doi: 10.3233/JAD-160297
Lipton, S. A. (2006). Paradigm shift in neuroprotection by NMDA receptor blockade: memantine and beyond. Nat. Rev. Drug Discov. 5, 160–170. doi: 10.1038/nrd1958
Liu, B., Zhu, Y., Zhou, J., Wei, Y., Long, C., Chen, M., et al. (2014). Endoplasmic reticulum stress promotes amyloid-beta peptides production in RGC-5 cells. Cell Stress Chaperones 19, 827–835. doi: 10.1007/s12192-014-0506-7
Liu, J., Chang, L., Roselli, F., Almeida, O. F. X., Gao, X., Wang, X., et al. (2010). Amyloid-β induces caspase-dependent loss of PSD-95 and synaptophysin through NMDA receptors. J. Alzheimers Dis. 22, 541–556. doi: 10.3233/JAD-2010-100948
Liu, L. (2004). Role of NMDA receptor subtypes in governing the direction of hippocampal synaptic plasticity. Science 304, 1021–1024. doi: 10.1126/science.1096615
Liu, S. B., Zhang, N., Guo, Y. Y., Zhao, R., Shi, T. Y., Feng, S. F., et al. (2012). G-protein-coupled receptor 30 mediates rapid neuroprotective effects of estrogen via depression of NR2B-containing NMDA receptors. J. Neurosci. 32, 4887–4900. doi: 10.1523/JNEUROSCI.5828-11.2012
Liu, Y., Wong, T. P., Aarts, M., Rooyakkers, A., Liu, L., Lai, T. W., et al. (2007). NMDA receptor subunits have differential roles in mediating excitotoxic neuronal death both in vitro and in vivo. J. Neurosci. 27, 2846–2857. doi: 10.1523/JNEUROSCI.0116-07.2007
Low, C. M., and Wee, K. S. L. (2010). New insights into the not-so-new nr3 subunits of n-methyl-D-aspartate receptor: localization, structure, and function. Mol. Pharmacol. 78, 1–11. doi: 10.1124/mol.110.064006
Luscher, C., and Malenka, R. C. (2012). NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Perspect. Biol. 4:a005710. doi: 10.1101/cshperspect.a005710
Lynch, G., Larson, J., Kelso, S., Barrionuevo, G., and Schottler, F. (1983). Intracellular injections of EGTA block induction of hippocampal long-term potentiation. Nature 305, 719–721. doi: 10.1038/305719a0
MacDermott, A. B., Mayer, M. L., Westbrook, G. L., Smith, S. J., and Barker, J. L. (1986). NMDA-receptor activation increases cytoplasmic calcium concentration in cultured spinal cord neurones. Nature 321, 519–522. doi: 10.1038/321519a0
MacDonald, J. F., Jackson, M. F., and Beazely, M. A. (2006). Hippocampal long-term synaptic plasticity and signal amplification of NMDA receptors. Crit. Rev. Neurobiol. 18, 71–84. doi: 10.1615/CritRevNeurobiol.v18.i1-2.80
Maeda, S., Djukic, B., Taneja, P., Yu, G. Q., Lo, I., Davis, A., et al. (2016). Expression of A152T human tau causes age-dependent neuronal dysfunction and loss in transgenic mice. EMBO Rep. 17, 530–551. doi: 10.15252/embr.201541438
Malenka, R. C. (1994). Synaptic plasticity in the hippocampus: LTP and LTD. Cell 78, 535–538. doi: 10.1016/0092-8674(94)90517-7
Malenka, R. C., and Bear, M. F. (2004). LTP and LTD: an embarrassment of riches. Neuron 44, 5–21. doi: 10.1016/j.neuron.2004.09.012
Malinow, R. (2012). New developments on the role of NMDA receptors in Alzheimer’s disease. Curr. Opin. Neurobiol. 22, 559–563. doi: 10.1016/j.conb.2011.09.001
Manczak, M., Anekonda, T. S., Henson, E., Park, B. S., Quinn, J., and Reddy, P. H. (2006). Mitochondria are a direct site of Aβ accumulation in Alzheimer’s disease neurons: implications for free radical generation and oxidative damage in disease progression. Hum. Mol. Genet. 15, 1437–1449. doi: 10.1093/hmg/ddl066
Marquard, J., Otter, S., Welters, A., Stirban, A., Fischer, A., Eglinger, J., et al. (2015). Characterization of pancreatic NMDA receptors as possible drug targets for diabetes treatment. Nat. Med. 21, 363–372. doi: 10.1038/nm.3822
Martel, M. A., Wyllie, D. J. A., and Hardingham, G. E. (2009). In developing hippocampal neurons, NR2B-containing N-methyl-D-aspartate receptors (NMDARs) can mediate signaling to neuronal survival and synaptic potentiation, as well as neuronal death. Neuroscience 158, 334–343. doi: 10.1016/j.neuroscience.2008.01.080
Martínez-Turrillas, R., Puerta, E., Chowdhury, D., Marco, S., Watanabe, M., Aguirre, N., et al. (2012). The NMDA receptor subunit GluN3A protects against 3-nitroproprionic-induced striatal lesions via inhibition of calpain activation. Neurobiol. Dis. 48, 290–298. doi: 10.1016/j.nbd.2012.07.001
Massey, P. V. (2004). Differential roles of NR2A and NR2B-containing NMDA receptors in cortical long-term potentiation and long-term depression. J. Neurosci. 24, 7821–7828. doi: 10.1523/JNEUROSCI.1697-04.2004
Matsuda, K., Fletcher, M., Kamiya, Y., and Yuzaki, M. (2003). Specific assembly with the NMDA receptor 3B subunit controls surface expression and calcium permeability of NMDA receptors. J. Neurosci. 23, 10064–10073. doi: 10.1523/JNEUROSCI.23-31-10064.2003
Mayer, M. L., Westbrook, G. L., and Guthrie, P. B. (1984). Voltage-dependent block by Mg2+ of NMDA responses in spinal cord neurones. Nature 309, 261–263. doi: 10.1038/309261a0
McGuinness, L., Taylor, C., Taylor, R. D. T., Yau, C., Langenhan, T., Hart, M. L., et al. (2010). Presynaptic NMDARs in the hippocampus facilitate transmitter release at theta frequency. Neuron 68, 1109–1127. doi: 10.1016/j.neuron.2010.11.023
Mesic, I., Madry, C., Geider, K., Bernhard, M., Betz, H., and Laube, B. (2016). The N-terminal domain of the GluN3A subunit determines the efficacy of glycine-activated NMDA receptors. Neuropharmacology 105, 133–141. doi: 10.1016/j.neuropharm.2016.01.014
Micu, I., Jiang, Q., Coderre, E., Ridsdale, A., Zhang, L., Woulfe, J., et al. (2006). NMDA receptors mediate calcium accumulation in myelin during chemical ischaemia. Nature 439, 988–992. doi: 10.1038/nature04474
Mishizen-Eberz, A. J., Rissman, R. A., Carter, T. L., Ikonomovic, M. D., Wolfe, B. B., and Armstrong, D. M. (2004). Biochemical and molecular studies of NMDA receptor subunits NR1/2A/2B in hippocampal subregions throughout progression of Alzheimer’s disease pathology. Neurobiol. Dis. 15, 80–92. doi: 10.1016/j.nbd.2003.09.016
Misra, C., Brickley, S. G., Farrant, M., and Cull-Candy, S. G. (2000). Identification of subunits contributing to synaptic and extrasynaptic NMDA receptors in Golgi cells of the rat cerebellum. J. Physiol. 524, 147–162. doi: 10.1111/j.1469-7793.2000.00147.x
Molnar, E., Va Radi, A., McIlhinney, R. A. J., and Ashcroft, S. J. H. (1995). Identification of functional ionotropic glutamate receptor proteins in pancreatic β-cells and in islets of langerhans. FEBS Lett. 371, 253–257. doi: 10.1016/0014-5793(95)00890-L
Momiyama, A., Feldmeyer, D., and Cull-Candy, S. G. (1996). Identification of a native low-conductance NMDA channel with reduced sensitivity to Mg2+ in rat central neurones. J. Physiol. 494, 479–492. doi: 10.1113/jphysiol.1996.sp021507
Montes, D., and Aguilera, P. (2015). A metabotropic-like flux-independent NMDA receptor regulates Ca2+ exit from endoplasmic reticulum and mitochondrial membrane potential in cultured astrocytes. PLoS One 10:e0126314. doi: 10.1371/journal.pone.0126314
Monyer, H., Burnashev, N., Laurie, D. J., Sakmann, B., and Seeburg, P. H. (1994). Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron 12, 529–540. doi: 10.1016/0896-6273(94)90210-0
Morimoto, B. H. (2018). Therapeutic peptides for CNS indications: progress and challenges. Bioorgan Med. Chem. 26, 2859–2862. doi: 10.1016/j.bmc.2017.09.011
Morris, G. P., Clark, I. A., and Vissel, B. (2018). Questions concerning the role of amyloid-β in the definition, aetiology and diagnosis of Alzheimer’s disease. Acta Neuropathol. 136, 663–689. doi: 10.1007/s00401-018-1918-8
Mota, S. I., Ferreira, I. L., Pereira, C., Oliveira, C. R., and Rego, A. C. (2012). Amyloid-beta peptide 1-42 causes microtubule deregulation through N-methyl-D-aspartate receptors in mature hippocampal cultures. Curr. Alzheimer Res. 9, 844–856. doi: 10.2174/156720512802455322
Mota, S. I., Ferreira, I. L., and Rego, A. C. (2014). Dysfunctional synapse in Alzheimer’s disease – A focus on NMDA receptors. Neuropharmacology 76, 16–26. doi: 10.1016/j.neuropharm.2013.08.013
Muller, T., Albrecht, D., and Gebhardt, C. (2009). Both NR2A and NR2B subunits of the NMDA receptor are critical for long-term potentiation and long-term depression in the lateral amygdala of horizontal slices of adult mice. Learn Mem. 16, 395–405. doi: 10.1101/lm.1398709
Murugan, M., Sivakumar, V., Lu, J., Ling, E. A., and Kaur, C. (2011). Expression of N-methyl D-aspartate receptor subunits in amoeboid microglia mediates production of nitric oxide via NF-κB signaling pathway and oligodendrocyte cell death in hypoxic postnatal rats. Glia 59, 521–539. doi: 10.1002/glia.21121
Nabavi, S., Kessels, H. W., Alfonso, S., Aow, J., Fox, R., and Malinow, R. (2013). Metabotropic NMDA receptor function is required for NMDA receptor-dependent long-term depression. Proc. Natl. Acad. Sci. U.S.A. 110, 4027–4032. doi: 10.1073/pnas.1219454110
Nagele, R. G., Andrea, M. R. D., Anderson, W. J., and Wang, H. Y. (2002). Intracellular accumulation of β-amyloid1–42 in neurons is facilitated by the α7 nicotinic acetylcholine receptor in Alzheimer’s disease. Neuroscience 110, 199–211. doi: 10.1016/S0306-4522(01)00460-2
Nagy, J. (2004). The NR2B subtype of NMDA receptor: a potential target for the treatment of alcohol dependence. Curr. Drug Targets CNS Neurol. Disord. 3, 169–179. doi: 10.2174/1568007043337409
Nakanishi, N., Tu, S., Shin, Y., Cui, J., Kurokawa, T., Zhang, D., et al. (2009). Neuroprotection by the NR3A subunit of the NMDA receptor. J. Neurosci. 29, 5260–5265. doi: 10.1523/JNEUROSCI.1067-09.2009
Nakazawa, T., Komai, S., Tezuka, T., Hisatsune, C., Umemori, H., Semba, K., et al. (2001). Characterization of Fyn-mediated tyrosine phosphorylation sites on GluRε2 (NR2B) subunit of the N-Methyl-D-aspartate receptor. J. Biol. Chem. 276, 693–699. doi: 10.1074/jbc.M008085200
Newpher, T. M., and Ehlers, M. D. (2008). Glutamate receptor dynamics in dendritic microdomains. Neuron 58, 472–497. doi: 10.1016/j.neuron.2008.04.030
Nilsson, A., Duan, J., Mo-Boquist, L. L., Benedikz, E., and Sundström, E. (2007). Characterisation of the human NMDA receptor subunit NR3A glycine binding site. Neuropharmacology 52, 1151–1159. doi: 10.1016/j.neuropharm.2006.12.002
Nishi, M., Hinds, H., Lu, H., Kawata, M., and Hayashi, Y. (2001). Motoneuron-specific expression of NR3B, a novel NMDA-type glutamate receptor subunit that works in a dominant-negative manner. J. Neurosci. 21, RC185–RC185. doi: 10.1523/JNEUROSCI.21-23-j0003.2001
Nowak, L., Bregestovski, P., Ascher, P., Herbet, A., and Prochiantz, A. (1984). Magnesium gates glutamate-activated channels in mouse central neurones. Nature 307, 462–465. doi: 10.1038/307462a0
Nyitrai, G., Kékesi, K. A., and Juhász, G. (2006). Extracellular level of GABA and Glu: in vivo microdialysis-HPLC measurements. Curr. Top. Med. Chem. 6, 935–940. doi: 10.2174/156802606777323674
Pachernegg, S., Strutz-Seebohm, N., and Hollmann, M. (2012). GluN3 subunit-containing NMDA receptors: not just one-trick ponies. Trends Neurosci. 35, 240–249. doi: 10.1016/j.tins.2011.11.010
Pagani, L., and Eckert, A. (2011). Amyloid-beta interaction with mitochondria. J. Alzheimers Dis. 2011, 1–12. doi: 10.4061/2011/925050
Palop, J. J., Chin, J., Roberson, E. D., Wang, J., Thwin, M. T., Bien-Ly, N., et al. (2007). Aberrant excitatory neuronal activity and compensatory remodeling of inhibitory hippocampal circuits in mouse models of Alzheimer’s disease. Neuron 55, 697–711. doi: 10.1016/j.neuron.2007.07.025
Palop, J. J., and Mucke, L. (2010). Amyloid-β-induced neuronal dysfunction in Alzheimer’s disease: from synapses toward neural networks. Nat. Neurosci. 13, 812–818. doi: 10.1038/nn.2583
Palygin, O., Lalo, U., and Pankratov, Y. (2011). Distinct pharmacological and functional properties of NMDA receptors in mouse cortical astrocytes. Br. J. Pharmacol. 163, 1755–1766. doi: 10.1111/j.1476-5381.2011.01374.x
Paoletti, P. (2011). Molecular basis of NMDA receptor functional diversity. Eur. J. Neurosci. 33, 1351–1365. doi: 10.1111/j.1460-9568.2011.07628.x
Paoletti, P., Ascher, P., and Neyton, J. (1997). High-affinity zinc inhibition of NMDA NR1–NR2A receptors. J. Neurosci. 17, 5711–5725. doi: 10.1523/JNEUROSCI.17-15-05711.1997
Paoletti, P., Bellone, C., and Zhou, Q. (2013). NMDA receptor subunit diversity: impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 14, 383–400. doi: 10.1038/nrn3504
Papouin, T., Ladépêche, L., Ruel, J., Sacchi, S., Labasque, M., Hanini, M., et al. (2012). Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell 150, 633–646. doi: 10.1016/j.cell.2012.06.029
Parameshwaran, K., Dhanasekaran, M., and Suppiramaniam, V. (2008). Amyloid beta peptides and glutamatergic synaptic dysregulation. Exp. Neurol. 210, 7–13. doi: 10.1016/j.expneurol.2007.10.008
Park, H., Popescu, A., and Poo, M. (2014). Essential role of presynaptic NMDA receptors in activity-dependent BDNF secretion and corticostriatal LTP. Neuron 84, 1009–1022. doi: 10.1016/j.neuron.2014.10.045
Parsons, C. G., Stöffler, A., and Danysz, W. (2007). Memantine: a NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system – Too little activation is bad, too much is even worse. Neuropharmacology 53, 699–723. doi: 10.1016/j.neuropharm.2007.07.013
Patel, L., and Grossberg, G. (2011). Combination therapy for Alzheimer’s disease. Drugs Aging 28, 539–546. doi: 10.2165/11591860-000000000-00000
Paterlini, M., Valerio, A., Baruzzi, F., Memo, M., and Spano, P. (1998). Opposing regulation of tau protein levels by ionotropic and metabotropic glutamate receptors in human NT2 neurons. Neurosci. Lett. 243, 77–80. doi: 10.1016/S0304-3940(98)00087-1
Pérez-Otaño, I., Larsen, R. S., and Wesseling, J. F. (2016). Emerging roles of GluN3-containing NMDA receptors in the CNS. Nat. Rev. Neurosci. 17, 623–635. doi: 10.1038/nrn.2016.92
Perez-Otano, I., Schulteis, C. T., Contractor, A., Lipton, S. A., Trimmer, J. S., Sucher, N. J., et al. (2001). Assembly with the NR1 subunit is required for surface expression of NR3A-containing NMDA receptors. J. Neurosci. 21, 1228–1237. doi: 10.1523/JNEUROSCI.21-04-01228.2001
Perszyk, R. E., DiRaddo, J. O., Strong, K. L., Low, C. M., Ogden, K. K., Khatri, A., et al. (2016). GluN2D-containing N-methyl-D-Aspartate receptors mediate synaptic transmission in hippocampal interneurons and regulate interneuron activity. Mol. Pharmacol. 90, 689–702. doi: 10.1124/mol.116.105130
Petralia, R. S., Al-Hallaq, R. A., and Wenthold, R. J. (2009). “Trafficking and targeting of NMDA receptors,” in Biology of the NMDA Receptor, Chap. 8, ed. A. M. VanDongen (Boca Raton, FL: CRC Press/Taylor & Francis).
Pierson, T. M., Yuan, H., Marsh, E. D., Fuentes-Fajardo, K., Adams, D. R., Markello, T., et al. (2014). GRIN2A mutation and early-onset epileptic encephalopathy: personalized therapy with memantine. Ann. Clin. Trans. Neurol. 1, 190–198. doi: 10.1002/acn3.39
Prybylowski, K., Fu, Z., Losi, G., Hawkins, L. M., Luo, J., Chang, K., et al. (2002). Relationship between availability of NMDA receptor subunits and their expression at the synapse. J. Neurosci. 22, 8902–8910. doi: 10.1523/JNEUROSCI.22-20-08902.2002
Puzzo, D., Privitera, L., Leznik, E., Fa, M., Staniszewski, A., Palmeri, A., et al. (2008). Picomolar amyloid- positively modulates synaptic plasticity and memory in hippocampus. J. Neurosci. 28, 14537–14545. doi: 10.1523/JNEUROSCI.2692-08.2008
Raveendran, R., Devi Suma, Priya, S., Mayadevi, M., Steephan, M., Santhoshkumar, T. R., et al. (2009). Phosphorylation status of the NR2B subunit of NMDA receptor regulates its interaction with calcium/calmodulin-dependent protein kinase II. J. Neurochem. 110, 92–105. doi: 10.1111/j.1471-4159.2009.06108.x
Regan, M. C., Grant, T., McDaniel, M. J., Karakas, E., Zhang, J., Traynelis, S. F., et al. (2018). Structural mechanism of functional modulation by gene splicing in NMDA receptors. Neuron 98, 521.e3–529.e3. doi: 10.1016/j.neuron.2018.03.034
Regan, M. C., Romero-Hernandez, A., and Furukawa, H. (2015). A structural biology perspective on NMDA receptor pharmacology and function. Curr. Opin. Struct. Biol. 33, 68–75. doi: 10.1016/j.sbi.2015.07.012
Reisberg, B., Doody, R., Stöffler, A., Schmitt, F., Ferris, S., and Möbius, H. J. (2003). Memantine in moderate-to-severe Alzheimer’s disease. New. Engl. J. Med. 348, 1333–1341. doi: 10.1056/NEJMoa013128
Reynolds, C. H., Garwood, C. J., Wray, S., Price, C., Kellie, S., Perera, T., et al. (2008). Phosphorylation regulates tau interactions with src homology 3 domains of phosphatidylinositol 3-kinase, phospholipase Cγ1, Grb2, and Src family kinases. J. Biol. Chem. 283, 18177–18186. doi: 10.1074/jbc.M709715200
Rice, L., and Bisdas, S. (2017). The diagnostic value of FDG and amyloid PET in Alzheimer’s disease-a systematic review. Eur. J. Radiol. 94, 16–24. doi: 10.1016/j.ejrad.2017.07.014
Roberson, E. D., Scearce-Levie, K., Palop, J. J., Yan, F., Cheng, I. H., Wu, T., et al. (2007). Reducing endogenous tau ameliorates amyloid-induced deficits in an Alzheimer’s disease mouse model. Science 316, 750–754. doi: 10.1126/science.1141736
Roberts, A. C., Díez-García, J., Rodriguiz, R. M., López, I. P., Luján, R., Martínez-Turrillas, R., et al. (2009). Downregulation of NR3A-containing NMDARs is required for synapse maturation and memory consolidation. Neuron 63, 342–356. doi: 10.1016/j.neuron.2009.06.016
Rogaeva, E., Meng, Y., Lee, J. H., Gu, Y., Kawarai, T., Zou, F., et al. (2007). The neuronal sortilin-related receptor SORL1 is genetically associated with Alzheimer disease. Nat. Genet. 39, 168–177. doi: 10.1038/ng1943
Rong, Y., Lu, X., Bernard, A., Khrestchatisky, M., and Baudry, M. (2001). Tyrosine phosphorylation of ionotropic glutamate receptors by Fyn or Src differentially modulates their susceptibility to calpain and enhances their binding to spectrin and PSD-95. J. Neurochem. 79, 382–390. doi: 10.1046/j.1471-4159.2001.00565.x
Rönicke, R., Mikhaylova, M., Rönicke, S., Meinhardt, J., Schröder, U. H., Fändrich, M., et al. (2011). Early neuronal dysfunction by amyloid β oligomers depends on activation of NR2B-containing NMDA receptors. Neurobiol. Aging 32, 2219–2228. doi: 10.1016/j.neurobiolaging.2010.01.011
Roselli, F., Tirard, M., Lu, J., Hutzler, P., Lamberti, P., Livrea, P., et al. (2005). Soluble β-amyloid1-40 induces NMDA-dependent degradation of postsynaptic density-95 at glutamatergic synapses. J. Neurosci. 25, 11061–11070. doi: 10.1523/JNEUROSCI.3034-05.2005
Rossi, B., Ogden, D., Llano, I., Tan, Y. P., Marty, A., and Collin, T. (2012). Current and calcium responses to local activation of axonal NMDA receptors in developing cerebellar molecular layer interneurons. PLoS One 7:e39983. doi: 10.1371/journal.pone.0039983
Rothman, S. M., and Olney, J. W. (1986). Glutamate and the pathophysiology of hypoxic-ischemic brain damage. Ann. Neurol. 19, 105–111. doi: 10.1002/ana.410190202
Ruthirakuhan, M., Herrmann, N., Suridjan, I., Abraham, E. H., Farber, I., and Lanctot, K. L. (2016). Beyond immunotherapy: new approaches for disease modifying treatments for early Alzheimer’s disease. Expert Opin. Pharmacother. 17, 2417–2429. doi: 10.1080/14656566.2016.1258060
Salomone, S., Caraci, F., Leggio, G. M., Fedotova, J., and Drago, F. (2012). New pharmacological strategies for treatment of Alzheimer’s disease: focus on disease modifying drugs. Br. J. Clin. Pharmacol. 73, 504–517. doi: 10.1111/j.1365-2125.2011.04134.x
Salter, M. G., and Fern, R. (2005). NMDA receptors are expressed in developing oligodendrocyte processes and mediate injury. Nature 438, 1167–1171. doi: 10.1038/nature04301
Salter, M. W., and Kalia, L. V. (2004). Src kinases: a hub for NMDA receptor regulation. Nat. Rev. Neurosci. 5, 317–328. doi: 10.1038/nrn1368
Sanders, E. M., Nyarko-Odoom, A. O., Zhao, K., Nguyen, M., Liao, H. H., Keith, M., et al. (2018). Separate functional properties of NMDARs regulate distinct aspects of spatial cognition. Learn Mem. 25, 264–272. doi: 10.1101/lm.047290.118
Sanderson, J. L., Gorski, J. A., and Dell’Acqua, M. L. (2016). NMDA receptor-dependent LTD requires transient synaptic incorporation of Ca(2)(+)-permeable AMPARs mediated by AKAP150-anchored PKA and calcineurin. Neuron 89, 1000–1015. doi: 10.1016/j.neuron.2016.01.043
Sans, N., Petralia, R. S., Wang, Y., Blahos, J., Hell, J. W., and Wenthold, R. J. (2000). A developmental change in NMDA receptor-associated proteins at hippocampal synapses. J. Neurosci. 20, 1260–1271. doi: 10.1523/JNEUROSCI.20-03-01260.2000
Sanz-Clemente, A., Nicoll, R. A., and Roche, K. W. (2012). Diversity in NMDA receptor composition. Neuroscientist 19, 62–75. doi: 10.1177/1073858411435129
Sattler, R., Xiong, Z., Lu, W., MacDonald, J. F., and Tymianski, M. (2000). Distinct roles of synaptic and extrasynaptic NMDA receptors in excitotoxicity. J. Neurosci. 20, 22–33. doi: 10.1523/JNEUROSCI.20-01-00022.2000
Shankar, G. M., Bloodgood, B. L., Townsend, M., Walsh, D. M., Selkoe, D. J., and Sabatini, B. L. (2007). Natural oligomers of the Alzheimer amyloid- protein induce reversible synapse loss by modulating an NMDA-type glutamate receptor-dependent signaling pathway. J. Neurosci. 27, 2866–2875. doi: 10.1523/JNEUROSCI.4970-06.2007
Sheng, M., and Lee, S. H. (2000). Growth of the NMDA receptor industrial complex. Nat. Neurosci. 3, 633–635. doi: 10.1038/76576
Shi, X., Sun, K., Hu, R., Liu, X., Hu, Q., Sun, X., et al. (2016). Blocking the Interaction between EphB2 and ADDLs by a small peptide rescues impaired synaptic plasticity and memory deficits in a mouse model of Alzheimer’s disease. J. Neurosci. 36, 11959–11973. doi: 10.1523/JNEUROSCI.1327-16.2016
Sindou, P., Couratier, P., Barthe, D., and Hugon, J. (1992). A dose-dependent increase of Tau immunostaining is produced by glutamate toxicity in primary neuronal cultures. Brain Res. 572, 242. doi: 10.1016/0006-8993(92)90476-P
Sindou, P., Lesort, M., Couratier, P., Yardin, C., Esclaire, F., and Hugon, J. (1994). Glutamate increases tau phosphorylation in primary neuronal cultures from fetal rat cerebral cortex. Brain Res. 646, 124. doi: 10.1016/0006-8993(94)90064-7
Sinor, J. D., Du, S., Venneti, S., Blitzblau, R. C., Leszkiewicz, D. N., Rosenberg, P. A., et al. (2000). NMDA and glutamate evoke excitotoxicity at distinct cellular locations in rat cortical neurons in vitro. J. Neurosci. 20, 8831–8837. doi: 10.1523/JNEUROSCI.20-23-08831.2000
Smothers, C. T., and Woodward, J. J. (2003). Effect of the NR3 subunit on ethanol inhibition of recombinant NMDA receptors. Brain Res. 987, 117–121. doi: 10.1016/S0006-8993(03)03315-8
Snyder, E. M., Nong, Y., Almeida, C. G., Paul, S., Moran, T., Choi, E. Y., et al. (2005). Regulation of NMDA receptor trafficking by amyloid-β. Nat. Neurosci. 8, 1051–1058. doi: 10.1038/nn1503
Sofroniew, M. V. (2009). Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 32, 638–647. doi: 10.1016/j.tins.2009.08.002
Song, X., Jensen, M. Ø., Jogini, V., Stein, R. A., Lee, C., Mchaourab, H. S., et al. (2018). Mechanism of NMDA receptor channel block by MK-801 and memantine. Nature 556, 515–519. doi: 10.1038/s41586-018-0039-9
Sproul, A., Steele, S. L., Thai, T. L., Yu, S., Klein, J. D., Sands, J. M., et al. (2011). N-methyl-D-aspartate receptor subunit NR3a expression and function in principal cells of the collecting duct. Am. J. Physiol. Renal. Physiol. 301, F44–F54. doi: 10.1152/ajprenal.00666.2010
Stein, I. S., Gray, J. A., and Zito, K. (2015). Non-ionotropic NMDA receptor signaling drives activity-induced dendritic spine shrinkage. J. Neurosci. 35, 12303–12308. doi: 10.1523/JNEUROSCI.4289-14.2015
Stroebel, D., Carvalho, S., Grand, T., Zhu, S., and Paoletti, P. (2014). Controlling NMDA receptor subunit composition using ectopic retention signals. J. Neurosci. 34, 16630–16636. doi: 10.1523/JNEUROSCI.2736-14.2014
Sucher, N. J., Akbarian, S., Chi, C. L., Leclerc, C. L., Awobuluyi, M., Deitcher, D. L., et al. (1995). Developmental and regional expression pattern of a novel NMDA receptor-like subunit (NMDAR-L) in the rodent brain. J. Neurosci. 15, 6509–6520. doi: 10.1523/JNEUROSCI.15-10-06509.1995
Suga, K., Saito, A., and Akagawa, K. (2015). ER stress response in NG108-15 cells involves upregulation of syntaxin 5 expression and reduced amyloid β peptide secretion. Exp. Cell Res. 332, 11–23. doi: 10.1016/j.yexcr.2015.01.001
Sumi, M., Kiuchi, K., Ishikawa, T., Ishii, A., Hagiwara, M., Nagatsu, T., et al. (1991). The newly synthesized selective Ca2+/calmodulin dependent protein kinase II inhibitor KN-93 reduces dopamine contents in PC12h cells. Biochem. Biophys. Res. Commun. 181, 968–975. doi: 10.1016/0006-291X(91)92031-E
Sun, W., Hansen, K. B., and Jahr, C. E. (2017). Allosteric interactions between NMDA receptor subunits shape the developmental shift in channel properties. Neuron 94, 58.e3–64.e3. doi: 10.1016/j.neuron.2017.03.018
Sun, X., Tuo, Q., Liuyang, Z., Xie, A., Feng, X., Yan, X., et al. (2016). Extrasynaptic NMDA receptor-induced tau overexpression mediates neuronal death through suppressing survival signaling ERK phosphorylation. Cell Death Dis. 7:e2449. doi: 10.1038/cddis.2016.329
Sundström, E., Whittemore, S., Mo, L., and Seiger, Å. (1997). Analysis of NMDA receptors in the human spinal cord. Exp. Neurol. 148, 407–413. doi: 10.1006/exnr.1997.6691
Swanger, S. A., Vance, K. M., Acker, T. M., Zimmerman, S. S., DiRaddo, J. O., Myers, S. J., et al. (2017). A novel negative allosteric modulator selective for GluN2C/2D-containing NMDA receptors inhibits synaptic transmission in hippocampal interneurons. ACS Chem. Neurosci. 9, 306–319. doi: 10.1021/acschemneuro.7b00329
Szasz, B. K., Mike, A., Karoly, R., Gerevich, Z., Illes, P., Vizi, E. S., et al. (2007). Direct inhibitory effect of fluoxetine on N-Methyl-D-Aspartate receptors in the central nervous system. Biol. Psychiatry 62, 1303–1309. doi: 10.1016/j.biopsych.2007.04.014
Sze, C., Bi, H., Kleinschmidt-DeMasters, B. K., Filley, C. M., and Martin, L. J. (2001). N-Methyl- D-aspartate receptor subunit proteins and their phosphorylation status are altered selectively in Alzheimer’s disease. J. Neurol. Sci. 182, 151–159. doi: 10.1016/S0022-510X(00)00467-6
Takai, H., Katayama, K., Uetsuka, K., Nakayama, H., and Doi, K. (2003). Distribution of N-methyl-D-aspartate receptors (NMDARs) in the developing rat brain. Exp. Mol. Pathol. 75, 89–94. doi: 10.1016/S0014-4800(03)00030-3
Tamburri, A., Dudilot, A., Licea, S., Bourgeois, C., and Boehm, J. (2013). NMDA-receptor activation but not ion flux is required for amyloid-beta induced synaptic depression. PLoS One 8:e65350. doi: 10.1371/journal.pone.0065350
Tayeb, H. O., Yang, H. D., Price, B. H., and Tarazi, F. I. (2012). Pharmacotherapies for Alzheimer’s disease: beyond cholinesterase inhibitors. Pharmacol. Therapeut. 134, 8–25. doi: 10.1016/j.pharmthera.2011.12.002
Texidó, L., Martín-Satué, M., Alberdi, E., Solsona, C., and Matute, C. (2011). Amyloid β peptide oligomers directly activate NMDA receptors. Cell Calcium 49, 184–190. doi: 10.1016/j.ceca.2011.02.001
Tezuka, T., Umemori, H., Akiyama, T., Nakanishi, S., and Yamamoto, T. (1999). PSD-95 promotes Fyn-mediated tyrosine phosphorylation of the N-methyl-D-aspartate receptor subunit NR2A. Proc. Natl. Acad. Sci. U.S.A. 96, 435–440. doi: 10.2307/46829
Thiels, E., Xie, X., Yeckel, M. F., Barrionuevo, G., and Berger, T. W. (1996). NMDA receptor-dependent LTD in different subfields of hippocampus in vivo and in vitro. Hippocampus 6, 43–51. doi: 10.1002/(SICI)1098-1063(1996)6:1<43::AID-HIPO8>3.0.CO;2-8
Thomas, C. G., Miller, A. J., and Westbrook, G. L. (2006). Synaptic and extrasynaptic NMDA receptor NR2 subunits in cultured hippocampal neurons. J. Neurophysiol. 95, 1727–1734. doi: 10.1152/jn.00771.2005
Thomas, S. M., and Brugge, J. S. (1997). Cellular functions regulated by Src family kinases. Annu. Rev. Cell Dev. Biol. 13, 513–609. doi: 10.1146/annurev.cellbio.13.1.513
Ting, K., Brew, B. J., and Guillemin, G. J. (2009). Effect of quinolinic acid on human astrocytes morphology and functions: implications in Alzheimer’s disease. J. Neuroinflamm. 6:36. doi: 10.1186/1742-2094-6-36
Tovar, K. R., and Westbrook, G. L. (2017). Modulating synaptic NMDA receptors. Neuropharmacology 112, 29–33. doi: 10.1016/j.neuropharm.2016.08.023
Townsend, M., Yoshii, A., Mishina, M., and Constantine-Paton, M. (2003). Developmental loss of miniature N-methyl-D-aspartate receptor currents in NR2A knockout mice. Proc. Natl. Acad. Sci. U.S.A. 100, 1340–1345. doi: 10.1073/pnas.0335786100
Traynelis, S. F., Wollmuth, L. P., McBain, C. J., Menniti, F. S., Vance, K. M., Ogden, K. K., et al. (2010). Glutamate receptor ion channels: structure, regulation, and function. Pharmacol. Rev. 62, 405–496. doi: 10.1124/pr.109.002451
Tymianski, M., Charlton, M. P., Carlen, P. L., and Tator, C. H. (1993). Source specificity of early calcium neurotoxicity in cultured embryonic spinal neurons. J. Neurosci. 13, 2085–2104. doi: 10.1523/JNEUROSCI.13-05-02085.1993
Ułas, J., and Cotman, C. W. (1997). Decreased expression of N-methyl-D-aspartate receptor 1 messenger RNA in select regions of Alzheimer brain. Neuroscience 79, 973–982. doi: 10.1016/S0306-4522(97)00023-7
Ullian, E. M., Barkis, W. B., Chen, S., Diamond, J. S., and Barres, B. A. (2004). Invulnerability of retinal ganglion cells to NMDA excitotoxicity. Mol. Cell. Neurosci. 26, 544–557. doi: 10.1016/j.mcn.2004.05.002
Um, J. W., Kaufman, A. C., Kostylev, M., Heiss, J. K., Stagi, M., Takahashi, H., et al. (2013). Metabotropic glutamate receptor 5 is a coreceptor for Alzheimer Aβ oligomer bound to cellular prion protein. Neuron 79, 887–902. doi: 10.1016/j.neuron.2013.06.036
Um, J. W., Nygaard, H. B., Heiss, J. K., Kostylev, M. A., Stagi, M., Vortmeyer, A., et al. (2012). Alzheimer amyloid-β oligomer bound to postsynaptic prion protein activates Fyn to impair neurons. Nat. Neurosci. 15, 1227–1235. doi: 10.1038/nn.3178
Usardi, A., Pooler, A. M., Seereeram, A., Reynolds, C. H., Derkinderen, P., Anderton, B., et al. (2011). Tyrosine phosphorylation of tau regulates its interactions with Fyn SH2 domains, but not SH3 domains, altering the cellular localization of tau. FEBS J. 278, 2927–2937. doi: 10.1111/j.1742-4658.2011.08218.x
Verges, D. K., Restivo, J. L., Goebel, W. D., Holtzman, D. M., and Cirrito, J. R. (2011). Opposing synaptic regulation of amyloid-beta metabolism by NMDA receptors in vivo. J. Neurosci. 31, 11328–11337. doi: 10.1523/JNEUROSCI.0607-11.2011
Vicini, S., Wang, J. F., Li, J. H., Zhu, W. J., Wang, Y. H., Luo, J. H., et al. (1998). Functional and pharmacological differences between recombinant N-Methyl-D-aspartate receptors. J. Neurophysiol. 79, 555–566. doi: 10.1152/jn.1998.79.2.555
Villmann, C., and Becker, C. (2007). On the hypes and falls in neuroprotection: targeting the NMDA receptor. Neuroscientist 13, 594–615. doi: 10.1177/1073858406296259
Vizi, E. S. (2000). Role of high-affinity receptors and membrane transporters in nonsynaptic communication and drug action in the central nervous system. Pharmacol. Rev. 52, 63–89.
Vizi, E. S., Kisfali, M., and Lõrincz, T. (2013). Role of nonsynaptic GluN2B-containing NMDA receptors in excitotoxicity: evidence that fluoxetine selectively inhibits these receptors and may have neuroprotective effects. Brain Res. Bull. 93, 32–38. doi: 10.1016/j.brainresbull.2012.10.005
Volianskis, A., France, G., Jensen, M. S., Bortolotto, Z. A., Jane, D. E., and Collingridge, G. L. (2015). Long-term potentiation and the role of N -methyl-D-aspartate receptors. Brain Res. 1621, 5–16. doi: 10.1016/j.brainres.2015.01.016
Wake, H., Lee, P. R., and Fields, R. D. (2011). Control of local protein synthesis and initial events in myelination by action potentials. Science 333, 1647–1651. doi: 10.1126/science.1206998
Walsh, D. M., Klyubin, I., Fadeeva, J. V., Cullen, W. K., Anwyl, R., Wolfe, M. S., et al. (2002). Naturally secreted oligomers of amyloid β protein potently inhibit hippocampal long-term potentiation in vivo. Nature 416, 535–539. doi: 10.1038/416535a
Walsh, D. M., Tseng, B. P., Rydel, R. E., Podlisny, M. B., and Selkoe, D. J. (2000). The oligomerization of amyloid β-protein begins intracellularly in cells derived from human brain†. Biochemistry-US 39, 10831–10839. doi: 10.1021/bi001048s
Wang, C., Pralong, W., Schulz, M., Rougon, G., Aubry, J., Pagliusi, S., et al. (1996). Functional N-Methyl-D-aspartate receptors in O-2A glial precursor cells: a critical role in regulating polysialic acid-neural cell adhesion molecule expression and cell migration. J. Cell Biol. 135, 1565–1581. doi: 10.1083/jcb.135.6.1565
Wang, H. Y., Lee, D. H., D’Andrea, M. R., Peterson, P. A., Shank, R. P., and Reitz, A. B. (2000). beta-Amyloid(1-42) binds to alpha7 nicotinic acetylcholine receptor with high affinity. implications for Alzheimer’s disease pathology. J. Biol. Chem. 275, 5626–5632. doi: 10.1074/jbc.275.8.5626
Wang, R., and Reddy, P. H. (2017). Role of glutamate and NMDA receptors in Alzheimer’s disease. J. Alzheimer’s Dis. 57, 1041–1048. doi: 10.3233/JAD-160763
Wang, Z., Zhao, J., and Li, S. (2013). Dysregulation of synaptic and extrasynaptic N-methyl-D-aspartate receptors induced by amyloid-β. Neurosci. Bull. 29, 752–760. doi: 10.1007/s12264-013-1383-2
Wee, K. S. L., Zhang, Y., Khanna, S., and Low, C. (2008). Immunolocalization of NMDA receptor subunit NR3B in selected structures in the rat forebrain, cerebellum, and lumbar spinal cord. J. Comp. Neurol. 509, 118–135. doi: 10.1002/cne.21747
Wenk, G. L. (2006). Neuropathologic changes in Alzheimer’s disease: potential targets for treatment. J. Clin. Psychiatry 67 (Suppl. 3), 3–7; quiz 23.
Whitesell, J. D., Buckley, A. R., Knox, J. E., Kuan, L., Graddis, N., Pelos, A., et al. (2018). Whole brain imaging reveals distinct spatial patterns of amyloid beta deposition in three mouse models of Alzheimer’s disease. J. Comp. Neurol. doi: 10.1002/cne.24555 [Epub ahead of print].
Wilkinson, D., Fox, N. C., Barkhof, F., Phul, R., Lemming, O., and Scheltens, P. (2012). Memantine and brain atrophy in Alzheimer’s disease: a 1-Year randomized controlled trial. J. Alzheimers Dis. 29, 459–469. doi: 10.3233/JAD-2011-111616
Williams, K. (1993). Ifenprodil discriminates subtypes of the N-methyl-D-aspartate receptor: selectivity and mechanisms at recombinant heteromeric receptors. Mol. Pharmacol. 44:851.
Wong, J. M., and Gray, J. A. (2018). Long-term depression is independent of GluN2 subunit composition. J. Neurosci. 38, 4462–4470. doi: 10.1523/JNEUROSCI.0394-18.2018
Wu, H., Tzeng, N., Qian, L., Wei, S., Hu, X., Chen, S., et al. (2009). Novel neuroprotective mechanisms of memantine: increase in neurotrophic factor release from astroglia and anti-inflammation by preventing microglial activation. Neuropsychopharmacology 34, 2344–2357. doi: 10.1038/npp.2009.64
Wu, H. Y., Hudry, E., Hashimoto, T., Kuchibhotla, K., Rozkalne, A., Fan, Z., et al. (2010). Amyloid induces the morphological neurodegenerative triad of spine loss, dendritic simplification, and neuritic dystrophies through calcineurin activation. J. Neurosci. 30, 2636–2649. doi: 10.1523/JNEUROSCI.4456-09.2010
Yang, J., Sophie, E. L. C., Woodhall, G. L., and Roland, S. G. J. (2008). Mobility of NMDA autoreceptors but not postsynaptic receptors at glutamate synapses in the rat entorhinal cortex. J. Physiol. 586, 4905–4924. doi: 10.1113/jphysiol.2008.157974
Yang, J., Woodhall, G. L., and Jones, R. S. G. (2006). Tonic facilitation of glutamate release by presynaptic NR2B-containing NMDA receptors is increased in the entorhinal cortex of chronically epileptic rats. J. Neurosci. 26, 406–410. doi: 10.1523/JNEUROSCI.4413-05.2006
Yao, Y. (2006). Characterization of a soluble ligand binding domain of the NMDA receptor regulatory subunit NR3A. J. Neurosci. 26, 4559–4566. doi: 10.1523/JNEUROSCI.0560-06.2006
Yao, Y., Harrison, C. B., Freddolino, P. L., Schulten, K., and Mayer, M. L. (2008). Molecular mechanism of ligand recognition by NR3 subtype glutamate receptors. EMBO J. 27, 2158–2170. doi: 10.1038/emboj.2008.140
Yashiro, K., and Philpot, B. D. (2008). Regulation of NMDA receptor subunit expression and its implications for LTD, LTP, and metaplasticity. Neuropharmacology 55, 1081–1094. doi: 10.1016/j.neuropharm.2008.07.046
Yi, F., Mou, T., Dorsett, K. N., Volkmann, R. A., Menniti, F. S., Sprang, S. R., et al. (2016). Structural basis for negative allosteric modulation of GluN2A-containing NMDA receptors. Neuron 91, 1316–1329. doi: 10.1016/j.neuron.2016.08.014
Yu, A., and Lau, A. Y. (2018). Glutamate and glycine binding to the NMDA receptor. Structure 26, 1035.e2–1043.e2. doi: 10.1016/j.str.2018.05.004
Yux, L. X., Foster, T. P., Sarver, R. W., and Moseley, W. M. (1996). Preparation, characterization, and in vivo evaluation of an oil suspension of a bovine growth hormone releasing factor analog. J. Pharm. Sci.-U.S. 85, 396–401. doi: 10.1021/js950110h
Zempel, H., Thies, E., Mandelkow, E., and Mandelkow, E. M. (2010). Abeta oligomers cause localized Ca(2+) elevation, missorting of endogenous tau into dendrites, tau phosphorylation, and destruction of microtubules and spines. J. Neurosci. 30, 11938–11950. doi: 10.1523/JNEUROSCI.2357-10.2010
Zhang, J., and Diamond, J. S. (2006). Distinct perisynaptic and synaptic localization of NMDA and AMPA receptors on ganglion cells in rat retina. J. Comp. Neurol. 498, 810–820. doi: 10.1002/cne.21089
Zhou, X., Ding, Q., Chen, Z., Yun, H., and Wang, H. (2013a). Involvement of the GluN2A and GluN2B subunits in synaptic and extrasynaptic n-methyl-D-aspartate receptor function and neuronal excitotoxicity. J. Biol. Chem. 288, 24151–24159. doi: 10.1074/jbc.M113.482000
Zhou, X., Hollern, D., Liao, J., Andrechek, E., and Wang, H. (2013b). NMDA receptor-mediated excitotoxicity depends on the coactivation of synaptic and extrasynaptic receptors. Cell Death Dis. 4, e560–e560. doi: 10.1038/cddis.2013.82
Zhu, J. J., Qin, Y., Zhao, M., Van Aelst, L., and Malinow, R. (2002). Ras and rap Control AMPA receptor trafficking during synaptic plasticity. Cell 110, 443–455. doi: 10.1016/S0092-8674(02)00897-8
Zhu, S., and Paoletti, P. (2015). Allosteric modulators of NMDA receptors: multiple sites and mechanisms. Curr. Opin. Pharmacol. 20, 14–23. doi: 10.1016/j.coph.2014.10.009
Keywords: NMDA receptor, amyloid beta, memantine, Alzheimer’s disease, synaptic dysfunction
Citation: Liu J, Chang L, Song Y, Li H and Wu Y (2019) The Role of NMDA Receptors in Alzheimer’s Disease. Front. Neurosci. 13:43. doi: 10.3389/fnins.2019.00043
Received: 17 September 2018; Accepted: 16 January 2019;
Published: 08 February 2019.
Edited by:
Tai Wai Yew, The Chinese University of Hong Kong, ChinaReviewed by:
Roland Brandt, University of Osnabrück, GermanyCopyright © 2019 Liu, Chang, Song, Li and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yan Wu, eWFud3VAY2NtdS5lZHUuY24=; MTM5MTEwNjE1OTRAMTYzLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.