 Louisa Dahmani1†
Louisa Dahmani1† Miryam Idriss
Miryam Idriss Greg L. West
Greg L. West Veronique D. Bohbot
Veronique D. Bohbot- 1Department of Psychiatry, Faculty of Medicine, Douglas Mental Health University Institute, McGill University, Verdun, QC, Canada
- 2Department of Psychology, University of Montreal, Montréal, QC, Canada
Sex differences in navigation have been a topic of investigation for decades and has been subjected to various contradictory findings and debates. The aim of this work was to compare the spatial memory of men and women tested in various different types of spatial tasks, while controlling for navigation strategies and aging. It is generally thought that men outperform women in navigation and that women have higher scores on object location tasks. However, many studies fail to control for different factors that may bias one sex or the other. We aggregated the data of 465 participants (349 young adults, 127 older adults) who took part in various studies conducted in our laboratory, which include both published and original unpublished data, in order to investigate sex differences. In these studies, we used a number of different paradigms: virtual radial arm mazes, a virtual wayfinding task, an object location task, a virtual Morris Water Maze, and the invisible sensor task which is a real-life model of the Morris Water Maze. While our results may seem discordant at first glance, they demonstrate that several factors can impact the performance of men and women on spatial tasks, including spontaneous navigation strategies, environmental characteristics, and age. We replicated findings showing that women favor proximal landmarks compared to men who favor distal landmarks, women have better memory than men for the position of objects in the absence of reference frames, but they will have poorer scores when navigation requires specific angles, distances and polar coordinates. Moreover, we found that in aging, women who avoid the use of landmarks when navigating a radial maze show stronger reliance on these non-spatial strategies than men. On the other hand, women who rely on landmarks, do so to the same extent as men. Our findings highlight the need to carefully take into consideration these factors in order to produce a more harmonious understanding of sex differences in navigation. Finally, the interaction between spontaneous navigation strategies, sex, and age is discussed in terms of its implications for risk of Alzheimer’s disease.
Introduction
Spatial ability in men and women has received a lot of interest. It is generally believed that men outperform women in this domain. Spatial ability is a loosely defined function and as such, a wide array of tasks are claimed to measure it, most of them table-top tasks. These table-top tasks are considerably different from real-life navigation. For example, table-top tasks are viewed from a single perspective and all the information is contained within the field of view, while in navigation the information is spread out in a large-scale environment and the environment is experienced from multiple perspectives (Maguire et al., 1999). These types of tests do not predict real-world navigation ability (Habib and Sirigu, 1987; McCarthy et al., 1996; Maguire and Cipolotti, 1998; Maguire et al., 1999).
Navigation has been intensively studied in rodents. This research spurred the development of numerous real-life paradigms meant to test spatial ability. The Morris Water Maze (Morris, 1981) and the radial maze (Olton and Samuelson, 1976) are the most commonly used mazes in the literature and have greatly contributed to the understanding of navigation. Following this profusion in rodent navigation research, efforts were put into creating real-life spatial tasks for humans akin to those used in animal research (Wertlieb and Rose, 1979; Overman et al., 1996; Bohbot et al., 1997; Bohbot et al., 1998). However, these require wide spaces and are subject to environmental variability (weather conditions, noise, etc.), and do not afford a lot of experimental control.
Using virtual reality, many groups were able to recreate mazes typically used in rodent research. Virtual reality furthermore holds other advantages: it allows full experimental control of the physical aspects of the environment, e.g., layout, lighting, object appearance and placement (Gamberini, 2000; Spiers et al., 2001). Although virtual navigation lacks motor and vestibular component (Bohbot et al., 2017) it is ideal for the study of navigation, as it enables participants to experience the same virtual environment in exactly the same physical conditions (Dahmani et al., 2012).
Many rodent tasks were adapted for human research using virtual reality, including the Morris Water Maze task (MWM), the radial arm maze, and the T-maze. A male advantage is usually found in the MWM (Astur et al., 1998; Astur et al., 2004; Newhouse et al., 2007; Mueller et al., 2008; Woolley et al., 2010) and was also found in one study that investigated wayfinding (Iaria et al., 2008). Studies examining performance in radial mazes did not find sex differences (Astur et al., 2004; Levy et al., 2005). A female advantage is typically observed in object location memory tasks age. (see Voyer et al., 2007 for a review).
In this study we focus on the impact of environmental characteristics used in the various navigation paradigms, navigation strategies, and age on spatial memory performance in men and women. The characteristics of the environments in which men and women are tested also have a large impact on navigation ability. For example, individuals tend to overestimate distances where there is a larger geographical gap and increased turns between two landmarks (Lopez and Bosco, 2022). Certain environments may inadvertently favour men, while others may favour women, which can lead to data misinterpretation. For example, a study involving a hometown map completion test based on well-known and familiar landmarks by Lopez et al., 2020b showed a female advantage when encoding the positions of landmarks. Consistent with this idea, the absence of landmarks can disadvantage women as they often rely on landmarks when navigating and are impaired when there are none (Sandstrom et al., 1998; Andersen et al., 2012), while men’s navigation performance will remain unaffected because they use other sources of information such as cardinal directions and Euclidean coordinates (Dabbs Jr et al., 1998; Sandstrom et al., 1998; Saucier et al., 2002; Postma et al., 2004; Ruggiero et al., 2008; Andersen et al., 2012). Additionally, men are better at estimating precise angles and distances (Holding and Holding, 1988; Postma et al., 1998; Lawton and Morrin, 1999; Postma et al., 2004; Iachini et al., 2005), which could explain why they perform better than women in navigation paradigms in which these skills are required, such as the Morris Water Maze (Astur et al., 1998; Astur et al., 2004; Rizk-Jackson et al., 2006; Mueller et al., 2008; Nowak and Moffat, 2011; Dahmani et al., 2012).
Navigation strategies influence how a specific spatial task is perceived, processed, and solved, and therefore can affect navigation ability. Two navigation strategies can be used when people learn to navigate in an environment. One, the spatial strategy, which is supported by the hippocampus involves learning the spatial relationship between objects or landmarks in an environment (Holdstock et al., 2000; O'Keefe and Nadel, 1978). The spontaneous use of this strategy leads to the formation of a cognitive map, or a mental representation, of one’s environment and is associated with more grey matter and functional magnetic resonance imaging (fMRI) in the hippocampus (Tolman, 1948; Iaria et al., 2003; Bohbot et al., 2007; West et al., 2018). On the other hand, the response strategy is supported by the caudate nucleus and involves learning, through repetition, a series of movements in relation to a stimulus set al., 2007; Iaria et al., 2003, Packard et al., 1989; White and McDonald, 2002). The spontaneous use of this strategy is associated with increased fMRI activity and grey matter in the caudate nucleus and reduced activity and grey matter in the hippocampus (Iaria et al., 2003; Bohbot et al., 2007; West et al., 2018). The spatial strategy is flexible in that it allows one to infer shortcuts between landmarks or novel trajectories in the environment, while the response strategy is rigid in its use of well-learned routes. Thus, sex differences may, at least in part, be explained by the possibility that men and women use different navigation strategies, or that the same strategies lead to different navigational behaviour and performance in men and women. However, these strategies are seldom assessed in navigation studies.
Finally, with age, navigation is impaired, which creates larger individual differences within older adults in spatial tasks (Wolbers and Hegarty, 2010). For example, older adults have more difficulty with increased complexity of tasks when encoding the positions of landmarks (Lopez et al., 2020a). Aging can also alter the navigation ability differences between men and women, as aging affects the sexes differently. Moreover, older adults with Mild Cognitive Impairment showed impairment in tasks involving differences in geometry and landmark position (proximal or distal) using a Virtual Reorientation Test (Caffò et al., 2017). Volume reductions in the hippocampus may be early signs of Alzheimer’s disease, and a smaller hippocampus is known to be a risk factor for cognitive decline in normal aging (Reiman et al., 1998; Du et al., 2001). In addition, the hippocampus and entorhinal cortex are also the first brain regions to display Alzheimer’s disease, which is then followed by disruption in the connectivity between the hippocampus and other regions of the brain (Du et al., 2001; Wang et al., 2006). Healthy older adults who maintained spatial navigation strategies displayed increased grey matter and fMRI activity in their hippocampus (Konishi and Bohbot, 2013; Konishi et al., 2013).
In this paper, we combined different studies conducted in our laboratory, which include both published and original unpublished findings, and investigated sex differences in 465 participants (349 young adults, 127 older adults). In these studies, we used a number of different paradigms in a total of 6 experiments: 2 radial arm mazes, a wayfinding task, a virtual MWM, and a real-life navigation task that resembles the MWM and an object location task. We will discuss how our results relate to the literature on sex differences, and how they shed light on the impact of age, navigation strategies, and type of environment on navigation.
In experiment 1 and 2, we aimed to test spontaneous navigation strategies in both men and women, young and old using a 4 on 8 Virtual Maze (4/8 VM) and a Concurrent Spatial Discrimination Learning Task (CSDLT). We consider the 4/8 VM and CSDLT environments to be neutral tasks in that they do not favor men nor women. Considering women are more prone to Alzheimer’s disease than men (Barnes et al., 2005), that a small hippocampus is a predictor of future diagnosis of Alzheimer’s disease (Mielk et al., 2012), that response learners have a smaller hippocampus than spatial learners (Bohbot et al., 2007; Konishi and Bohbot, 2013), we hypothesized that response women would be impaired at hippocampus dependent tasks with aging.
In experiment 3 we aimed to test sex differences using a virtual town wayfinding task and hypothesized that men would outperform women due to their ability to better estimate angles and distances in a large environment (Holding and Holding, 1988; Postma et al., 1998; Lawton and Morrin, 1999; Postma et al., 2004; Iachini et al., 2005). The aim of experiment 4 was to compare spatial and response learners in latencies to find specific targets using a Morris Water Maze, a task that can only be solved using a spatial strategy (Morris, 1987)32. Considering several studies have found men to outperform women on this task, we hypothesized that response women would have higher latencies to find the target compared to other groups (Astur et al., 1998; Astur et al., 2004; Rizk-Jackson et al., 2006; Mueller et al., 2008; Chai and Jacobs, 2009; Nowak and Moffat, 2011). In experiment 5 we aimed to test the use of landmarks using an Invisible Sensor Task (IST), which is a real-life spatial task that was modelled after the Morris Water Maze. The environment is small in scale and therefore does not require individuals to estimate distances and angles over a large environment and contains both proximal and distal landmarks. We therefore hypothesized that women would use proximal landmarks to a greater extent than men. Lastly, we aimed to explore sex differences using the object location memory task which tends to favor women as it involves an in-depth reconstruction process to recall the positions of objects which have to be learned in relation to each other, without necessarily estimating angles, we therefore hypothesized women to outperform men (Saucier et al., 2007).
Experiment 1: The 4 on 8 virtual maze
Methods
Participants
Healthy young adults (N = 248; women: n = 129, M age: = 24.73, SD = 4.61; men: n = 119, M age = 24.95, SD = 4.50) and 99 healthy older adults (women: n = 55, M age = 66.64, SD = 7.32; men: n = 44, M age: 65.77, SD = 5.55) were tested. All participants were right-handed and had no history of neurological or psychiatric disorders. Informed consent was obtained in conformity with the local ethics committee requirements for all experiments.
Procedure
All the virtual environments were created using the Unreal Tournament 2003 game development platform (Unreal Tournament 2003; Epic Games, Raleigh, NC). Participants were seated in front of a standard 17” monitor, with a 1,280 × 1,024 resolution. Participants were placed in a practice virtual environment to familiarize themselves with the keys which allowed them to move around. They were instructed to navigate in the virtual environment using the forward, left, and right arrow keys on a keyboard. In order to best simulate real-world navigation, participants were not allowed to navigate using the backwards key. When both the experimenter and participant agreed that the participant was comfortable enough navigating with the keys, the experimental task was given. This procedure was used in all subsequent virtual navigation experiments.
4 on 8 virtual maze (4/8 VM)
The 4/8 VM is a virtual eight-arm radial maze modelled after the eight-arm maze used in rodents (Olton and Samuelson, 1976). The eight-arm radial maze is surrounded by a rich landscape that contains landmarks (Figure 1). Participants gather four objects from four open pathways in a first phase and they have to remember which are the previously visited pathways in order to avoid them in a second phase and collect the remaining four objects among the eight open pathways. Typically, when a spatial strategy is used to solve the task, participants learn the spatial relationships between features of the background environment (mountains, trees, boulders) and the target objects. On the other hand, people who use a response strategy typically learn a sequence of movements (e.g., skip one arm, enter the next arm, then skip two arms) in order to locate the target objects, while ignoring the relationship between environmental landmarks. To measure the extent to which the landmarks were used to encode the position of the objects, a probe trial is administered in which the environmental landmarks are removed. Participants who previously used landmarks to navigate, i.e., those who used a spatial strategy, will make errors on the probe when trying to find the objects, since there are no landmarks to rely on, while participants who used a response strategy will make fewer errors. Finally, we administered a verbal report, asking participant to describe how they solved the task from the beginning to the end of the experiment. The verbal report served to categorize participants as either spatial learners or response learners. The 4/8 VM is described in more detail in earlier publications (Iaria et al., 2003; Bohbot et al., 2004; Bohbot et al., 2007; Etchamendy and Bohbot, 2007; West et al., 2018). Throughout the entire verbal report, experimenters follow a strict protocol to ensure all questions are read word for word without mentioning words that could potentially bias the participant, such as “landmark” or “sequence” (Iaria et al., 2003). There are a total of 12 questions asked to ensure sufficient information is given regarding how each participant memorized where the specific objects are place throughout the entire task. To be classified a spatial learner, individuals need to specify at least 2 landmarks and no single sequence. In order to classify a verbal report as a response strategy, the participant needs to describe a sequence or pattern to remember where specific objects are. They do not mention any landmarks in their verbal report unless it is used as a reference point such as a starting position for the sequence (e.g., a counting sequence from a tree). To identify a shift, experimenters ask if participants used the method they previously described for all the trials from the beginning to the end, then ask for elaboration). An example of a verbal report scored as a spatial strategy would be “I used the mountains to get the first object, then I knew that next to the tree was another object and the two last objects were next to the pyramid. An example of a verbal report scored as a response strategy would be “I went down the arm directly ahead, then skipped two arms to the right, then skipped one arm…..” Here, the participant used their starting position as the starting point of the counting sequence. Other participants using the response strategy may start the single sequence from a single landmark. For example, “from the tree going counterclockwise: open, closed, closed, open … ” Moreover, when classifying strategies, we typically use the initial spontaneous strategy combining spatial learners with shifters because both groups started with the spatial strategy and combining all response learners who counted from a single point (starting position or single landmark). In this task, we previously reported that the response strategy is more efficient such that response learners make fewer errors and spend less time to complete a trial (Iaria, et al., 2003). This could explain why most of the shifters will shift from the spatial to the response strategy and not the other way around. The verbal report is written word for word by the experimenter and is scored by two independent experimenters, blind to each other’s strategy classification. The inter-rater reliability of strategy assessment showed a 96% overlap between two experimenters that independently assigned the subjects to the different groups (spatial memory, shift, response strategy) (Iaria et al., 2003).

Figure 1. Screenshots of the 4 on 8 Virtual Maze. The task consists of an 8-arm radial maze, in which 4 arms are accessible and 4 are blocked. Participants have to retrieve objects located at the end of the 4 accessible arms. Then, all 8 arms become accessible and participants have to retrieve objects now located in the 4 pathways that were previously blocked. After the task is learned, a probe trial is given in which all landmarks are removed. Created with Unity Editor®. Unity is a trademark or registered trademark of Unity Technologies.
Analysis
We investigated the effect of sex and navigation strategies on the average number of errors made, average time taken to complete a trial, and number of trials to criterion by entering them as dependent variables in a MANOVA with sex and navigation strategies as the independent variables. For probe performance, we conducted a two-way ANOVA with sex and navigation strategies as the independent variables and probe error as the dependent variable. Finally, to compare men and women’s proportion of strategies used, we conducted a chi square analysis.
Results
Task performance
In both the young and older adult samples, men and women did not differ on any of the 4/8 VM measures; they were similar in the number of errors made, in the time taken to complete a trial, and in the number of trials to criterion (p > 0.05) (Figure 2; Table 1): Average errors young: women: n = 125; M = 0.88; SD = 1.17; ε2 = 0.064; men: n = 118; M = 0.74; SD = 1.04; ε2 = 0.064). Trials to criterion young: women: n = 125; M = 1.58; SD = 0.70; ε2 = 0.036; men: n = 118; M 1.52; SD = 0.62, ε2 = 0.036). Average errors old: women: n = 55; M = 1.76; SD = 1.47; ε2 = 0.064; men: n = 44; M = 1.91; SD = 1.57; ε2 = 0.064. Trials to criterion old: women: n = 55; M = 2.40; SD = 1.27; ε2 = 0.064; men: n = 44; M = 2.30; SD = 1.11; ε2 = 0.064). The fact that men and women did not differ in the number of errors across trials and in the number of trials to criterion indicates they learned the task at similar rates.
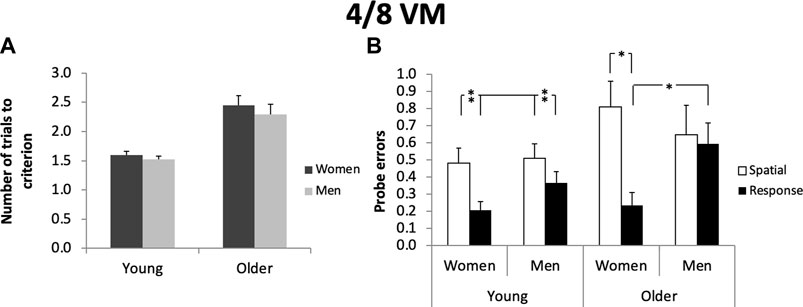
Figure 2. (A): Women and men require the same number of trials to reach the learning criterion in the 4 on 8 Virtual Maze. This was found in both young (p > 0.05) and older adults (p > 0.05). (B): In young adults, response strategy users make fewer probe errors than spatial strategy users (F (1.243) = 8.79, p < 0.01) In older adults, response women make fewer probe errors than spatial women (F (1.95) = 12.05, p < 0.05) and response men (F (1.95) = 5.41, p < 0.05). Error bars represent standard errors of the mean. *p < 0.05; **p < 0.01.
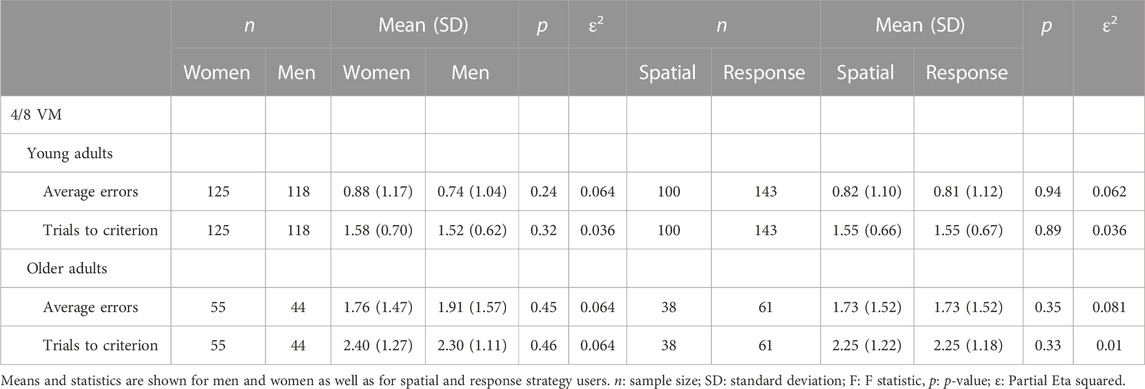
Table 1. 4 on 8 Virtual Maze (4/8 VM): Means and statistics for average errors and number of trials to criterion for young and older adults.
Navigation strategies
In both the young and older adult samples, men and women did not differ in the strategies they used (young women: n = 78 response, n = 49 spatial; young men: n = 76 response, n = 40 spatial; χ2 (1, N = 243) = 0.87, p > 0.05; Cramer’s V = 0.007); older women: n = 40 response and older men: n = 28 response; older women: n = 15 spatial; older men: n = 16 spatial; χ2 (1, N = 99) = 0.57, p > 0.05; Cramer’s V = 0.09) or in the number of probe errors: young women: n = 125; young men: n = 118; older women: n = 55; older men n = 44 (p > 0.05). There was no main effect of navigation strategies or any interaction on average errors, average time taken to complete a trial, or number of trials to criterion in either age group (p > 0.05). As mentioned above, young women: n = 125 average errors (M = 0.88; SD = 1.17); n = 125 trials to criterion (M = 1.58; SD = 0.70). For young men: n = 118 average errors (M = 0.74; SD = 1.04); n = 118 trials to criterion (M = 1.52; SD = 0.62) (Table 1). In young adults, there was a main effect of navigation strategies on probe error: those who used a spatial strategy performed worse on the probe than those who used a response strategy (spatial: M = 0.50, SD = 0.61; response: M = 0.29, SD = 0.50; F (1,243) = 8.79, p < 0.01, ε2 = 0.081), demonstrating that spatial strategy users did use landmarks to a greater extent than response strategy users.
The same was found in older adults (spatial: M = 0.74, SD = 0.69; response: M = 0.39, SD = 0.56; F (1.95) = 6.43, p < 0.05, ε2 = 0.064. However, in the older adults, there was a significant interaction between sex and navigation strategies on probe performance (F (1.95) = 4.40, p < 0.05, ε2 = 0.071). A test of simple effects with Sidak correction showed that there is an effect of navigation strategies in women only (F (1.95) = 12.05, p < 0.05, ε2 = 0.075). Thus, older spatial and response women differ in their probe performance but older spatial and response men do not (spatial women: M = 0.81, SD = 0.68; response women: M = 0.24, SD = 0.43; spatial men: M = 0.65, SD = 0.70; response men: M = 0.59, SD = 0.64). Another test of simple effects revealed that there is an effect of sex within response strategy users only: older women made fewer errors on the probe than older men (F (1.95) = 5.41, p < 0.05, ε2 = 0.074. Within spatial strategy users, older men and women performed equally on the probe (older women M = 0.81; SD = 0.68; older men: M = 0.65; SD = 0.70). To further investigate the difference in probe performance between each of the groups, we conducted a one-way ANOVA with four groups: spatial men, spatial women, response men, and response women. The overall ANOVA was significant (F (3.95) = 4.63, p < 0.01) and post hoc tests showed that older response women made fewer errors on the probe than the other three groups; ε2 = 0.072 (all p’s < 0.05). The same was found in older adults (spatial: M = 0.74, SD = 0.69; response: M = 0.39, SD = 0.56; F (1.95) = 6.43, p < 0.05, ε2 = 0.064). However, in the older adults, there was a significant interaction between sex and navigation strategies on probe performance (F (1.95) = 4.40, p < 0.05, ε2 = 0.071). A test of simple effects with Sidak correction showed that there is an effect of navigation strategies in women only (F (1.95) = 12.05, p < 0.05; ε2 = 0.075). Thus, older spatial and response women differ in their probe performance, but older spatial and response men do not (spatial women: M = 0.81, SD = 0.68; response women: M = 0.24, SD = 0.43; spatial men: M = 0.65, SD = 0.70; response men: M = 0.59, SD = 0.64). Another test of simple effects revealed that there is an effect of sex within response strategy users only: older women made fewer errors on the probe than older men (F (1.95) = 5.41, p < 0.05; ε2 = 0.074; (Figure 2). Within spatial strategy users, older men and women performed equally on the probe F (1.95) = 3.97, p = 0.049; ε2 = 0.061 (women: M = 0.81, SD = 0.68. men: M = 0.65, SD = 0.70; (Figure 2). To further investigate the difference in probe performance between each of the groups, we conducted a one-way ANOVA with four groups: spatial men, spatial women, response men, and response women. The overall ANOVA was significant (F (3.95) = 4.63, p < 0.01) and post hoc tests showed that older response women made fewer errors on the probe than the other three groups (all p’s < 0.05; ε2 = 0.072) (older spatial women: M = 0.81, SD = 0.68; older spatial men M = 0.65, SD = 0.70; older response women: M = 0.24; SD = 0.43; older response men: M = 0.59, SD = 0.64).
Discussion
Characteristics of the 4/8 VM environment
We consider the 4/8 VM environment to be a neutral task in the sense that it does not confer an advantage to either sex. The eight equally spaced pathways with fixed lengths make it so there is no need to measure precise angles or distances to reach a target (which would otherwise constitute a male advantage). The presence of multiple landmarks also allows women to navigate using their preferred method and does not constrain them to use cues that advantage men, such as cardinal directions and Euclidean coordinates. Finally, the target objects are distributed throughout a 3D environment and are not visible within a single field of view, which would otherwise confer women an advantage in object location. The fact that men and women learned the task at similar rates supports the idea that the 4/8 VM environment did not confer an advantage to either sex.
Navigation strategies
Men and women did not differ in the navigation strategies they used. Although it is believed that women rely on landmark information to a greater extent than men (Dabbs Jr et al., 1998; Sandstrom et al., 1998; Andersen et al., 2012), it appears that women are able to use this information in different ways using either a spatial or response strategy. For example, when using a response strategy, it is possible to learn a pattern of movements from a single landmark. It is thus possible to rely on landmarks whether one uses spatial or response strategies.
Aging
Older men and women, like young adults, do not differ in either their navigation performance during learning or in the navigation strategies that they use. However, response women make fewer mistakes on the probe than spatial strategy users and response men, suggesting that they use landmarks to a lesser extent than the other groups. If the use of landmarks is a process that requires the critical contribution of the hippocampus, these results suggest that older adult response women use their hippocampus less than other groups.
Relation of results with current literature
Few studies have used a radial maze paradigm to investigate sex differences in human navigation. Levy et al. (2005) used a 12-arm radial maze with an abundance of proximal cues. They asked young adult participants to learn the location of six objects among the 12 arms. In accordance with our own results, the authors did not find any sex differences in errors or latency during learning. Although Levy and colleagues did not assess navigation strategies in their 12-arm maze, it is clear that both spatial and response strategies could be used to solve the task, as both landmarks and target objects remained stable throughout learning. Thus, one could learn the spatial relationships between the cues and the target arms (spatial strategy) or learn a sequence of movements from a single landmark to locate the target arms (response strategy). Room geometry could also be used to locate the objects. Such a paradigm allowed men and women to use their preferred cues and may explain the lack of sex differences, as was seen in our study. Levy et al. (2005) went on to administer the T-maze, in which participants are placed at the end of an arm in a 4-arm maze and have to learn the location of the target object, which they can find using either a spatial strategy by associating the target arm with a cue or a response strategy by learning to turn left, for example. After learning, a probe trial that dissociates navigation strategies was administered. The authors found that, overall, men and women did not differ in their navigation strategies, which supports our findings.
Astur et al. (2004) used a similar paradigm: their task consisted of an eight-arm maze where proximal cues and room geometry could be used to locate four target objects. They found that women were slower to complete the trials, as we have previously found with the 4/8 VM (Iaria et al., 2003), but that the distance traveled was the same. Therefore, the paths taken by women were just as efficient as men’s. No sex differences were found in terms of errors. Overall, the findings of Levy et al. (2005) and Astur et al. (2004) are in agreement with ours, in that men and women do not differ in their learning of a radial arm maze or in their navigation strategies. On the other hand, our investigation of navigational strategies with age revealed that older adult response women use landmarks to a lesser extent than older adult response men. These results suggest that older adult response women may use their hippocampus less than other groups. This was confirmed by Sodums & Bohbot. (2020)48 which showed that among older adults, response women had the least amount of grey matter in the hippocampus.
Experiment 2: concurrent spatial discrimination learning task (CSDLT)
Methods
Participants
Healthy young adults (N = 78; women: n = 41, M age: 24.51, SD = 4.02; men: n = 37, M age = 24.35, SD = 4.14) and 82 healthy older adults (women: n = 46, M age: = 7.22, SD = 7.49; men: n = 36, M age = 66.00, SD = 6.08) were tested on the CSDLT.
Concurrent spatial discrimination learning task
The CSDLT is a virtual human analogue of the task developed for mice by Marighetto et al. (1999). It consists of a radial arm maze with 12 arms surrounded by a landscape and landmarks (Figure 3). During the learning stage, the arms are presented in isolated pairs of adjacent arms and participants have to learn the location of an object within each of these pairs. Once participants learn the location of the objects to a criterion of 92% correct, they are taken to the probe stage, which serves to assess participants’ navigation strategies. Here, the pairs are recombined into new adjacent pairs in which the object positions remain the same. To do well in this recombined stage, it is necessary to have learned the precise spatial relationships between the pathway containing the object and the landmarks in the landscape. For both the 4/8 virtual maze and CSDLT tasks, individuals who use a spatial learning approach rely on the relationships between landmarks, rather than a rigid pattern based on counting or the sequence of opened and closed pathways. The spatial strategy can rely on both proximal and distal landmarks, rather than solely relying on only distal or proximal landmarks. The landmarks used can be both proximal and distal.

Figure 3. Screenshots of the Concurrent Spatial Discrimination Learning Task. The task is comprised of a 12-arm radial maze surrounded by landmarks and a scenery. Top: In the learning phase, participants have to learn the location of objects within pairs of pathways. Bottom: In the probe phase, the pathways are recombined into new pairs but the object locations remain the same. Screenshots of the Concurrent Spatial Discrimination Learning Task. The task is comprised of a 12-arm radial maze surrounded by landmarks and a scenery. In the learning phase, participants have to learn the location of objects within pairs of arms. In the probe phase, the arms are recombined into new pairs but the object locations remain the same. Created with Unity Editor®. Unity is a trademark or registered trademark of Unity Technologies.
Thus, those who used a spatial strategy during learning will make fewer errors. Those who score at least 88% on the probe trial are considered to have used a spatial strategy during the learning stage, while those who score lower are considered to have used a response strategy (Etchamendy et al., 2012; Dahmani and Bohbot, 2014). The CSDLT is described in more detail in (Banner et al., 2011; Etchamendy et al., 2012; Dahmani and Bohbot, 2014).
The decision to use verbal reports exclusively for the 4/8 virtual maze, which puts participants in an open environment where a high number of variable routes can be used to solve the task, is rooted in the initial research conducted by Bohbot et al. (2004). In this study, it was found that when landmarks are removed during the probe trial, the 15 participants with damage to their medial temporal lobe had trouble remembering the starting direction they were facing relative to the previously displayed landmarks. More specifically, one-third of those who used the spatial strategy took three times longer than the remaining patients to complete the first out of three sections of the 4/8VM. This was also correlated with their increased number of errors during the task (14.9 vs. 8.1 errors) compared to the non-spatial group. Further, all participants, regardless of their strategy, made at least one error during the probe trial. Therefore, the probe trial could not be used to discriminate between the two strategies (Bohbot et al., 2004). Further, because all landmarks were removed during the probe trial, these participants were likely to make errors, behaving similarly to those using a spatial strategy. Consequently, relying solely on probe errors for grouping participants posed a risk of misclassification (Bohbot et al., 2004).
Because of this, the probe trial delivered at the end of the task did not accurately differentiate between spatial and response strategies. In contrast, the verbal report, which asks participants to account how they solved the maze, did allow for the accurate classification of the initial spontaneous navigation strategy used, which are validated by the probe trial in subsequent studies (Iaria et al., 2003; Konishi and Bohbot et al., 2013). Consequently, these studies affirmed the necessity of verbal reports during the 4/8 virtual navigation task.
Conversely, the CSDLT limits the participant to choose one of only two pathways in each trial, and no landmarks are removed from view, and therefore the potential disorientation experienced in the 4/8 virtual maze probe trial is avoided.
Finally, navigation strategies based upon verbal reports have been validated by both functional MRI studies (Iaria et al., 2003) and structural MRI studies (West et al., 2018). These validation studies further support the use of verbal reports for assessing cognitive processes during the 4/8 virtual maze task.
Analysis
We performed two separate ANOVAs with sex and navigation strategies as independent variables: one with trials to criterion as the dependent variable (n = 78) and one with average accuracy across trials as the dependent variable (n = 43). The overall accuracy is referring to the average number of correct entries made during the learning trials before the probe phase. In other words, participants who make few accurate choices of pathways in each trial before reaching criterion have poorer accuracy. In turn, participants who few accurate choices will typically need more trials to learn the task. Therefore, the average number of correct entries made during the learning trials and the number of trials needed to reach the criterion are related. This is because errors made during the learning trials reflect the same underlying cognitive process as the trials needed to reach the criterion. These measures are believed to represent distinct stages (average number of correct entries vs. number of trials to reach criterion where the criterion is dependent on correct entries) of the same learning process. These dependent variables were in separate analyses because accuracy information was missing for a subset of participants. To assess navigation strategy use, we performed a chi-square analysis with the number of spatial (probe score ≥88%) and response (probe score <88%) strategy users within both men and women. To investigate whether spontaneous navigation strategies have an effect on CSDLT performance, we conducted ANOVAs with sex and 4/8 VM strategy as independent variables and trials to criterion, average accuracy across trials, and probe score as dependent variables.
Results
Task performance
In both young and older adults, no sex differences were found on the number of trials to criterion: n = 34 young women (M = 7.15; SD = 0.56); n = 27 young men (M = 6.85; SD = 0.49); ε2 = 0.025; n = 46 older women (M = 9.80; SD = 4.67; n = 36 older men (M = 10.06; SD = 3.49); ε2 = 0.009 (p > 0.05) (Figure 4; Table 2) or on the average accuracy across trials: n = 15 young women (M = 5.03; SD = 0.42); n = 13 young men (M = 4.78; SD = 0.63); ε2 = 0.025; n = 46 older women (M = 3.76; SD = 1.15); n = 36 older men (M = 3.66, SD = 1.07); ε2 = 0.009 (p > 0.05) (Table 2). Thus, men and women learned the task at similar rates.
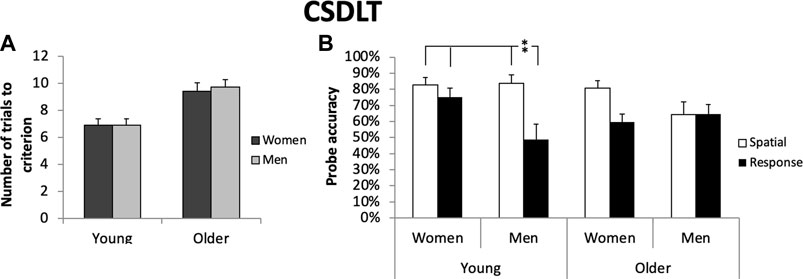
Figure 4. (A): Women and men require the same number of trials to reach the learning criterion in the Concurrent Spatial Discrimination Learning Task, whether they are young (p > 0.05) or older (p > 0.05). (B): In young adults, there is an interaction between sex and 4/8 VM strategies on the CSDLT probe (F (1.57) = 4.72, p < 0.05) whereby there is a larger difference between spatial and response men than between spatial and response women. This effect is driven by response men, who significantly differ from all other groups (all p’s < 0.01). In older adults, there is a tendency towards significance for an interaction effect between sex and 4/8 VM navigation strategies (F (1.78) = 3.35, p = 0.07) whereby there is a larger difference in probe performance between spatial and response women than between spatial and response men. No single group drove this effect (p > 0.05). Error bars represent standard errors of the mean. *p < 0.05; **p < 0.01.
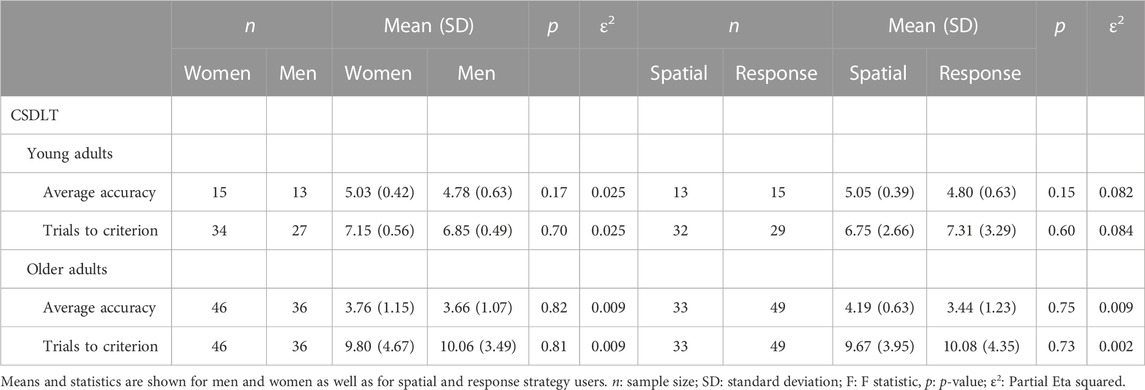
Table 2. Concurrent Spatial Discrimination Learning Task (CSDLT): Means and statistics for average accuracy and number of trials to criterion for young and older adults.
Navigation strategies
Young adult men and women did not differ in their CSDLT navigation strategies (young women: n = 21 response, n = 18 spatial; young men: n = 21 response, n = 16 spatial; χ2 (1, N = 76) = 6e−04, Cramer’s V = 0.0132, p > 0.05). There was a significant main effect of CSDLT strategy on both the number of trials to criterion (F (1.74) = 4.40, p = 0.039; ε2 = 0.082) and average accuracy (F (1.39) = 6.39, p = 0.02; ε2 = 0.084): those who used a spatial strategy had fewer trials to criterion (spatial: M = 6.75, SD = 2.66, response: M = 7.31, SD = 3.29) and higher average accuracy (spatial: M = 5.05, SD = 0.39, response: M = 4.80, SD = 0.63) than those who used a response strategy.
In older adults, there were no sex differences in CSDLT navigation strategy use (spatial men: n = 13, response men: n = 23; spatial women: n = 17, response women: n = 29; χ2 (1, N = 82) = 4e−31, Cramer’s V = 0.008; p > 0.05. As found in young adults, there was a significant main effect of strategy on average accuracy in older adults as well (F (1.81) = 9.66, p < 0.01, ε2 = 0.82). Older adults who used a spatial strategy had higher average accuracy (spatial n = 33, M = 4.19, SD = 0.63) than those who used a response strategy (response n = 49: M = 3.44, SD = 1.23). No difference was observed in the number of trials to criterion in older adults (spatial n = 33, M = 9.67, SD = 3.95; response n = 49, M = 10.08, SD = 4.35 (Table 2). These results indicate that in young and older adults, spatial strategies in the CSDLT are associated with fewer errors per trial. In turn, this lead to spatial learners making fewer trials to criterion, however, this effect was statistically significant in young adults only, because with aging, both spatial and response learners needed the same number of trials to reach criterion.
When we conducted analyses with 4/8 VM strategies to investigate whether spontaneous navigation strategies have an effect on CSDLT performance, there was no effect of 4/8 VM strategies, nor any interaction, on the number of trials to criterion or on the average accuracy in either young or older adults.
In terms of sex differences in probe performance in young adults, there were main effects where women performed better than men: young women probe: n = 34 (M = 78.68, SD = 5.89); young men probe: n = 25 (M = 69.44, SD = 9.45); (F (1.57) = 4.19, p < 0.05, ε2 = 0.067) and of 4/8 VM navigation strategies where spatial learners performed better than response learners: spatial probe: n = 30 (M = 89.2, SD = 5.94); response probe: n = 28 (M = 58.9, SD = 6.85); (F (1.57) = 11.80, p < 0.01, ε2 = 0.079), as well as an interaction between the two factors (F (1.57) = 4.72, p < 0.05, ε2 = 0.079). The significant interaction indicates that there is a larger difference between young spatial and response men on the CSDLT probe than between young spatial and response women.
To find out whether one specific group was driving this effect, we conducted an ANOVA with four groups based on sex and 4/8 VM navigation strategy (spatial women, spatial men, response women, response men). The overall ANOVA was significant (F (3.57) = 5.70, p < 0.01; ε2 = 0.079) and post hoc tests showed that young response men performed worse on the CSDLT probe than all other groups: young response men probe % correct: n = 11 (M = 48.86, SD = 9.45); young response women probe % correct: n = 18 (M = 75.00, SD = 5.89); young spatial men probe % correct: n = 16 (M = 83.58, SD = 5.43); young spatial women probe % correct: n = 4 (M = 82.81, SD = 4.40) (all p’s < 0.01). Thus, young men who spontaneously use response strategies on the 4/8 VM use response strategies on the CSDLT to a greater extent than the other groups.
In older adults, there was a tendency towards significance for the main effect of 4/8 VM navigation strategy (F (1.78) = 3.19, p = 0.08, ε2 = 0.04) as well as a tendency towards significance for the interaction between sex and 4/8 VM navigation strategies (F (1.78) = 3.35, p = 0.07, ε2 = 0.04). To investigate whether a specific group was driving this tendency, we performed an ANOVA with the four groups of participants. However, the overall ANOVA was non-significant (p > 0.05). However, due to our a priori hypotheses that navigation strategies will have an impact on the probe performance of the CSDLT, we still ran paired comparisons. Young men spatial learners made significantly fewer errors than young men response learners: young response men probe % correct: n = 11 (M = 48.86, SD = 9.45); young spatial men probe % correct: n = 16 (M = 83.58, SD = 5.43); (t = 3.41, p < 0.005; Cohen’s d = 4.51). With age, women who use spatial strategies, continue to perform well and are significantly better than response women: older women spatial probe score % correct: n = 16 (M = 80.63, SD = 4.75); older women response probe % correct: n = 30 (M = 59.72, SD = 4.59); (t = 3.11, p < 0.005; Cohen’s d = 4.47). Thus, with aging, we find that there is a tendency for a larger difference between spatial and response women on the probe trial, an effect that is consistent with the 4/8 VM findings described above.
Discussion
Characteristics of the CSDLT environment
As with the 4/8 VM, the CSDLT is comprised of equidistant pathways with fixed lengths, which eliminates any male advantage in estimating distances and angles to reach a target. The presence of landmarks and the distribution of objects in a 3D environment also preclude the male advantage in navigating without landmarks and the female advantage in object location. The task is thus considered to be neutral and, under these conditions, men and women perform equally well, as they learned the task at the same rate.
Navigation strategies
Men and women used spatial and response strategies in similar proportions on the CSDLT. Navigation strategies have an effect on navigation performance: spatial strategy users were more accurate and required fewer trials to learn the object locations. This highlights the importance of accounting for navigation strategies in a navigation study, as strategies can explain some of the variance in performance. This also highlights the importance of making sure that both sexes are balanced in terms of the navigation strategies that they use. Otherwise, navigation differences in performance may emerge that are not due to sex but to navigation strategies.
When looking at spontaneous navigation strategies using the 4/8 VM, we found that young response men performed worse than all other groups of the same age on the probe while older spatial women performed better than all other groups of the same age (see Figure 4). This indicates that young response men use response strategies on the CSDLT to a greater extent, and that older spatial women use spatial strategies on the CSDLT to a greater extent.
Aging
Older men and women did not differ in their accuracy, number of trials to reach criterion, probe performance, or navigation strategies. As in the 4/8 VM, the CSDLT does not advantage one sex or the other. As a result, no sex differences are observed in navigation in either young adults or older adults.
Relation of results with current literature
Our CSDLT findings are in accordance with our 4/8 VM results and with the findings of Levy et al. (2005) and Astur et al. (2004) described above: no sex differences were found in the learning of the CSDLT or in the navigation strategies used to solve the task. This was observed in both young and older adults, which indicates that the navigation performance of one sex relative to the other does not change throughout the lifespan when the paradigm used is not biased towards one sex. Consistent with the results of the 4/8 VM reported above, navigational strategies have an impact on spatial memory performance with age, such that older adult response women tended to have poorer performance than older adult spatial women or older adult men.
Experiment 3: The virtual town wayfinding task
Methods
Participants
Healthy young adults (N = 95; women: n = 52, M age = 23.9, SD = 4.16; men: n = 43,M age: 23.7, SD = 4.1) and 53 healthy older adults (women: n = 26, M age = 63.5, SD = 4.8; men: n = 27, M age: 65.4, SD = 5.0) were tested.
Virtual town wayfinding task
To assess wayfinding, a virtual navigation task involving a virtual town was used, modeled after the task used in Hartley et al. (2003). The virtual town is comprised of streets, buildings, and trees (Figure 5). Participants have to explore the virtual town and learn the location of eight landmarks. They are given a minimum of 20 min to explore the town and the exploration period is ended when participants have visited each street at least once and have passed by each landmark at least twice. This exploration period allows participants to build a cognitive map, which is accomplished by forming relationships between landmarks in the environment (O'Keefe and Nadel, 1978; Tolman, 1948). Then, knowledge of the town is tested by asking participants to find the shortest path between two landmarks, for example, between the cinema and the hospital. The task is described in more detail in Etchamendy and Bohbot (2007) and Dahmani et al. (2012).
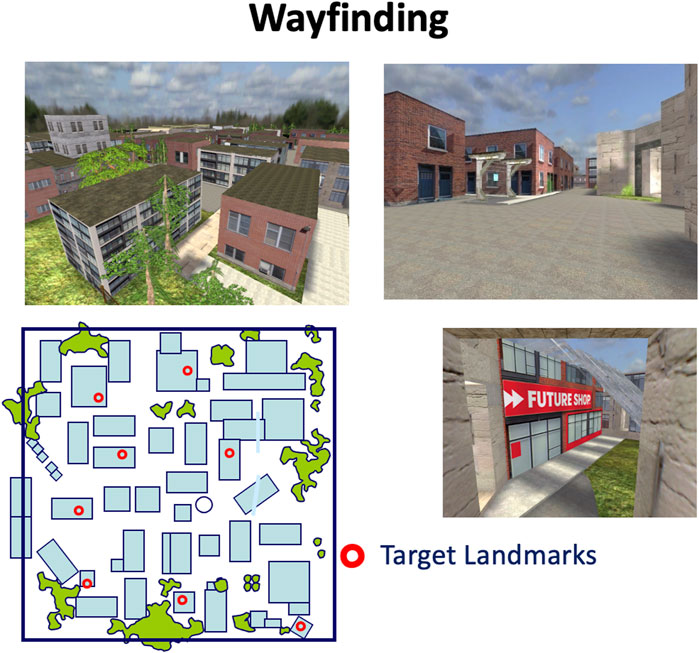
Figure 5. Screenshots and overhead representation of the virtual town wayfinding task. Participants have to explore the town for a minimum of 20 min. After the exploration period, they are given 8 probe trials in which they are placed in front of a landmark and are asked to take the shortest path to another landmark. Created with Unity Editor®. Unity is a trademark or registered trademark of Unity Technologies.
Analysis
In the statistical analyses, we considered only successful trials, defined as trials that were completed within three times the shortest time it takes to reach the target location on a given trial. This was done to minimize the impact on performance of unsuccessful trials (in which the target location was not found within a 5-min time limit) or trials where the target location was likely found by chance (trials where the latency is over three times the shortest possible latency). Unsuccessful trials were taken into account by the variable measuring the percentage of target locations found. The dependent variables thus included mean distance error (percentage of the distance traveled that is beyond the shortest distance possible), mean time error (percentage of the time taken that is beyond the minimum amount of time necessary to reach a target), and mean percentage of target locations found. During navigation, individuals have the option to pause and look around, which introduces a degree of variability. To account for this, we use two metrics: the mean time error and mean distance error. These measures are chosen because the search phase ends after a predetermined duration for each individual. This approach allows us to capture the variability introduced by individual stopping behaviors.
To compare men and women on their wayfinding performance, we conducted a one-way ANOVA with sex as the independent variable and the wayfinding measures as the dependent variables. Because the virtual town task does not allow us to dissociate the navigation strategies used, we conducted a MANOVA with a subset of participants who performed the 4/8 VM, as we have previously done in Etchamendy and Bohbot (2007) (young adults: N = 59; older adults: N = 48), with spontaneous navigation strategies and sex as the independent variables and wayfinding measures as the dependent variables. Finally, we were interested in looking at subsets of strategies within the two main strategy groups: spatial learners, response learners who do not rely on landmarks, and response learners who rely on landmarks. The inclusion of subgroup divisions was limited to a single experiment, the wayfinding task. Due to the restricted number of participants in experiments 4 and 5, specifically the Virtual Water Maze and Invisible Sensor Task, we refrained from further subdividing the spatial and response learners into the three designated categories: response with landmarks, response without landmarks, and spatial. The wayfinding task encompassed a total of 148 participants, comprising 95 older individuals and 53 younger individuals, all of whom were assessed using the 4/8VM to evaluate their navigation strategy. However, experiment 4 only involved testing 51 participants, and experiment 5 further narrowed down the sample to 30 individuals who were tested on the 4/8VM. Consequently, dividing these subcategories would result in insufficient sample sizes for analysis. Additionally, it is important to note that experiment 6, which focused on Object Location, did not include participants who were assessed using the 4/8VM.
We ran a MANOVA with these strategies as part of the navigation strategy independent variable and with wayfinding measures as the dependent variables. Then, we conducted LSD post hoc tests to further look at the differences between groups. Because we have previously found men to outperform women in this task and because response strategy users who did not use landmarks were found to perform poorly compared to the other strategy users (Dahmani et al., 2012), we expected women who spontaneously use response strategies without landmarks to perform more poorly on the wayfinding task. As such, we conducted a set of planned comparisons (independent samples t-tests) and corrected for multiple comparisons using the Bonferroni correction.
Results
Task performance
Young men overall outperformed young women on the wayfinding task. Young men found significantly more target locations than women (young men % targets found n = 43; M = 91.98, SD = 13.27; young women % targets found n = 52; M = 83.60; SD = 22.28). (F (1.93) = 4.70, p < 0.05, ε2 = 0.048) (Figure 6; Table 3), and they did so more quickly (women n = 52: M = 33.47, SD = 13.51; men n = 43: M = 25.54, SD = 14.72) (F (1.93) = 6.47, p < 0.05, ε2 = 0.063). However, in successful trials, the mean distance error did not differ between men and women t (57) = 0.72; Cohen’s d = 0.21 p> 0.05; men n = 27: M = 19.52; SD = 14.63; women n = 32: M = 22.26; SD = 10.21). In older adults, men and women perform equally on all three measures (distance error %: men n = 27; M = 52.57; SD = 13.64; women n = 26: M = 49.80; SD = 11.55; t (51) = 0.78; Cohen’s d = 0.22 p > 0.05; time error %: men n = 27: M = 61.12; SD = 11.12; women n = 26; M = 66.11; SD = 12.12; t (51) = 1.42; Cohen’s d = 0.42 p > 0.05% targets found: men n = 27; M = 49.01; SD = 28.73; women n = 26; M = 42.30; SD = 25.48; t (51) = 0.89; Cohen’s d = 0.24 p > 0.05) (Figure 6; Table 3).
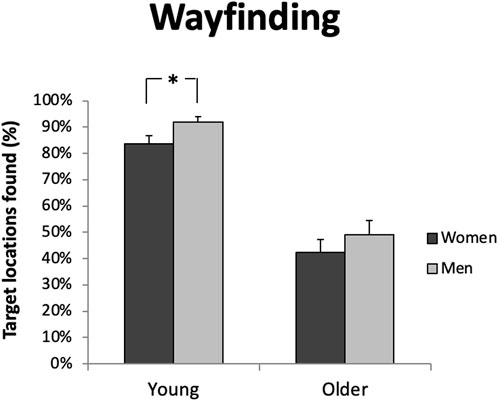
Figure 6. Young men found more target locations than young women in the virtual town (F (1.93) = 4.70, p < 0.05) Older men and women did not differ in this respect (p > 0.05). Error bars represents standard errors of the mean. *p < 0.05.
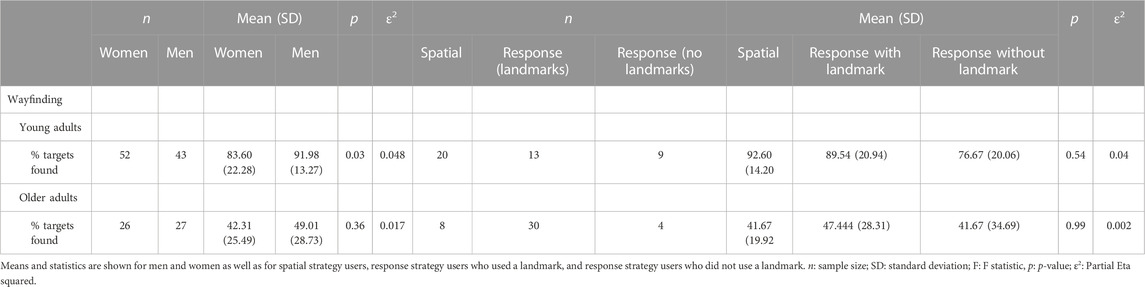
Table 3. Wayfinding virtual town task: Means and statistics for percentage of target locations found for young and older adults.
The presence of a larger standard deviation (SD) in young women compared to young men (22.28 vs. 13.27) suggests that the data for young women exhibit greater variability. Consequently, it is important to exercise caution when interpreting the results. Saying this, since our sample size is relatively large (N = 95), the standard deviations tend to become less sensitive in detecting small differences. This is because larger sample sizes provide more precise estimates of the population parameters. Therefore, even if the difference between the SDs of the means for the two groups is considerable, it still indicates a substantial distinction between the groups.
Navigation strategies
In young adults, navigation strategies did not influence any of the wayfinding measures. However, we divided the spatial and response groups into further strategy groups, as described in the analysis section. In this analysis, navigation strategies had a significant effect on mean distance error (F (3.39) = 3.39, p < 0.05, ε2 = 0.078) and mean time (F (3.39) = 2.80, p < 0.05, ε2 = 0.079), but not target locations found (p > 0.05). Post hoc tests revealed that spatial learners travel shorter distances to target locations (p < 0.01): spatial n = 13: M = 17.59, SD = 7.46; response n = 9 (with no landmarks): M = 29.04, SD = 6.98, reach them more quickly (p < 0.05) spatial n = 20: M = 25.09, SD = 15.38; response n = 9 (no landmarks) M = 38.13, SD = 4.78), and find more target locations (p < 0.05) spatial n = 20 M = 92.60, SD = 14.20; response n = 9 (no landmarks) M = 76.67, SD = 20.06) than response learners who do not use landmarks. Response learners who base their sequence on a landmark also travel shorter distances (p < 0.05) response n = 13 (landmarks): M = 18.33, SD = 7.23) and reach targets more quickly (p < 0.05) response n = 13 (landmarks): M = 24.94, SD = 15.68) than response learners who do not use landmarks. In older adults, navigation strategies did not modulate any of the wayfinding measures (p > 0.05). There was no interaction between sex and navigation strategies on wayfinding measures (p > 0.05).
Planned comparisons, in young adults, between response women who did not use a landmark on the 4/8 VM and the other five groups of participants (response men and women who used a landmark, as well as spatial men and women, and response men who did not use a landmark). They were compared on all three wayfinding variables (mean distance error, mean time error, percentage of target locations found). Response women who did not use a landmark have a greater mean time error than both spatial women (t (11) = 3.72, p < 0.0033; Cohen’s d = 0.91) and response men who used a landmark (t (9) = 7.29, p < 0.0033; Cohen’s d = 1.79); Spatial women time error % n = 6 (M = 26.20, SD = 18.21); spatial men time error % n = 14 (M = 24.61 SD = 17.86); response women time error % (landmark) n = 9 (M = 29.41; SD = 16.57); response men time error % (landmark) n = 4 (M = 14.87, SD = 17.46); response women time error % (without landmark) n = 7 (M = 39.02, SD = 7.74); response men time error % (without landmark) n = 2 (M = 34.99, SD = 8.54). They do not differ from the other groups in terms of mean distance error or percentage of target locations found: spatial women distance error % n = 6 (M = 16.92, SD = 6.99); spatial men distance error % n = 14 (M = 17.88, SD = 7.89); response women (landmark) distance error % n = 9 (M = 20.28, SD = 7.13); response men (landmark) distance error % = n = 4 (M = 13.93, SD = 6.01); response women (without landmarks) distance error % n = 7 (M = 28.85, SD = 7.80); response men (without landmarks) distance error % n = 2 (M = 29.71, SD = 4.86); all p’s > 0.0033. Thus, although response women who did not spontaneously use a landmark in the 4/8 VM are slower than some of the other groups, they do not travel longer distances or find fewer target locations. In older adults, the same planned comparisons yielded no significant results (all p’s > 0.0033).
Discussion
Characteristics of the virtual town environment
Forming a cognitive map of the virtual town and finding the shortest route between two landmarks require estimating precise angles and distances between the various elements in the environment, especially when one needs to keep track of winding roads where a small angular error would lead to a significant deviation from one’s goal. For this reason, the task likely confers men with an advantage. This is supported by the finding that men found more target locations than women and did so more quickly. Despite the absence of a significant difference in mean distance error between young men and women, we did see an overall effect where men outperform women in the wayfinding task: they were able to locate more targets and do so more rapidly. In addition, we had previously found that men traveled shorter distances to target locations than women (Dahmani et al., 2012), which also supports the statement that men are advantaged in tasks that require precise angle and distance estimations (Holding and Holding, 1988; Postma et al., 1998; Lawton and Morrin, 1999; Postma et al., 2004; Iachini et al., 2005). The speed at which participants can navigate during the wayfinding task is constant, therefore if men found targets more quickly it was due to a more efficient, shorter choice of route. Because all participants used the same simple directional button setup on the keyboard and completed a habituation trial that ensured that they were able to navigate in the virtual environments, no differences observed were due to the input interface used.
Navigation strategies
Our current results replicate those of an earlier study (Etchamendy and Bohbot, 2007). People who spontaneously use landmark-based navigation strategies (i.e., spatial learners and response learners who base their sequence or pattern on a landmark), as assessed with the 4/8 VM, are able to find their way better than people who do not spontaneously rely on landmarks (response learners who base their sequence or pattern on their starting position). It is possible that people who spontaneously use landmarks are better able to use landmark information in a wayfinding task. However, there were no interactions between navigation strategies and sex on the task. Therefore, men overall performed better than women regardless of the navigation strategies that they spontaneously use.
Aging
In older adults, men and women performed equally well, regardless of their navigation strategies. Thus, with aging, sex differences in wayfinding disappear. This could be due to a number of factors, such as changes in hormones with age (e.g., menopause) or differential brain volume decline in aging. Indeed, the hippocampus, a brain region associated with spatial learning, undergoes faster volume decline with age in men than in women (Pruessner et al., 2001).
Relation of results with current literature
Few studies have investigated sex differences in a virtual town. Iaria et al. (2008) had participants learn the location of four buildings in a town that contained nine buildings. Participants had to explore the town until they formed a cognitive map of it, which was assessed by asking participants to indicate on a map the location of the target buildings among the nine buildings. Once the cognitive map was formed, participants were given probe trials where they had to travel from one landmark to another using the shortest path possible. The authors found that, during the exploration period, men formed a cognitive map more quickly than women. In the probe trials, men and women performed equally well. At first glance, these results may not seem consistent with ours, as we found that men outperform women in the probe trials while Iaria et al. (2008) found no sex differences. However, in the paradigm used in Iaria et al. (2008), the probe trials were given once each individual participant demonstrated that they had learned the location of the target buildings, i.e., after they had built a cognitive map. In our study, everyone was administered the probe trials after approximately 20 min of exploration during which each individual was required to have visited each landmark at least twice. If men form a cognitive map more quickly than women, then our results are indeed consistent with Iaria et al. (2008): when the exploration period is equated between men and women, men are better able to find the target landmarks. Iaria et al. (2008)’s findings are interesting in that they suggest that, once a cognitive map is formed, men and women do not appear to perform differently. However, it is important to keep in mind that the virtual town used in Iaria et al. (2008) was small in scale and that, in a large-scale environment, men may still outperform women due to their better ability to estimate distances and angles (Holding and Holding, 1988; Postma et al., 1998; Lawton and Morrin, 1999; Postma et al., 2004; Iachini et al., 2005).
Other studies have used virtual labyrinths to study navigation (Castelli et al., 2008; Chen et al., 2009; Head and Isom, 2010), the results of which support our findings. We only consider the ones that allow for active exploration of the environment (instead of passive viewing of an environment). In Castelli et al. (2008), participants explored a labyrinth that contained landmarks. Afterwards, they were placed in a specific place and were asked to angle themselves towards a specific landmark. At the end of the task, participants were asked to place icons depicting each landmark on a map. Men made less angular error than women when asked to turn towards the position of another landmark and had less distance error than women when placing the icons on a map. Chen et al. (2009) asked participants to explore a virtual aquatic exhibition with 28 objects, after which they were asked to find specific objects. The authors found men to take less time to find the objects than women. Head and Isom (2010) tested young and older adults on a task in which they had to explore a series of interconnected hallways containing 28 landmarks. After a 15-min exploration, probes were administered where participants had to find a specified landmark using the shortest path possible. Head and Isom (2010) did not find sex to have an effect on performance, in either young or older adults. The finding in older adults mirrors our own, whereby there were no sex differences between the older men and women in our study. However, the results in young adults are not consistent with the typical male advantage seen in wayfinding tasks. It is possible that the exploration period of 15 min was sufficient to build a detailed cognitive map of the small environment used in the study, and thus would not yield sex differences in probe trials, as argued above with regards to results from Iaria et al. (2008). Additionally, because the virtual task was made up of interconnected pathways and not an open environment, the task alleviated the need to estimate precise distances and angles between landmarks, which may also make this task less sensitive to sex differences.
Overall, previous studies and our results do support the notion of a male advantage in wayfinding. Factors that may modulate sex differences in wayfinding tasks include size and complexity of the environment, having an open environment vs. one that is more restrictive in the movements allowed (e.g., pathways, hallways), task difficulty (e.g., number of landmark positions to memorize), exploration time, and navigation strategies. Navigation strategies in wayfinding tasks are not usually accounted for, as this type of task usually requires a spatial strategy to be used.
Experiment 4: virtual water maze (VWM)
Methods
Participants
Healthy young adults (N = 51; women: n = 31, M = age: 20.83, SD = 2.05; men: n = 20, M age: 22.15, SD = 2.03) were tested on the VWM. They were also tested on the 4/8 VM to assess whether spontaneous navigation strategies have an impact on VWM performance.
Virtual water maze
The VWM is a virtual human analogue of the Morris Water Maze (Morris, 1987). Participants have to learn the location of a hidden target on the floor of an arena surrounded by landmarks (Figure 7). The landmarks for this task were images of objects and we did not distinguish between proximal and distal landmarks. On each trial, participants start from a different position, however the target remains in the same location throughout the learning trials. After 20 trials, the target is removed unbeknownst to the participants (probe trial). The participants’ search pattern provides an assessment of their learning. The dependent variable is percentage of time spent in the correct quadrant, where the platform used to be located. Then, participants are told that the target has been removed, and they have to navigate to where they think the target was located. Once the training is complete, participants are asked whether they believed that the location of the platform was constant or whether it moved from trial to trial.
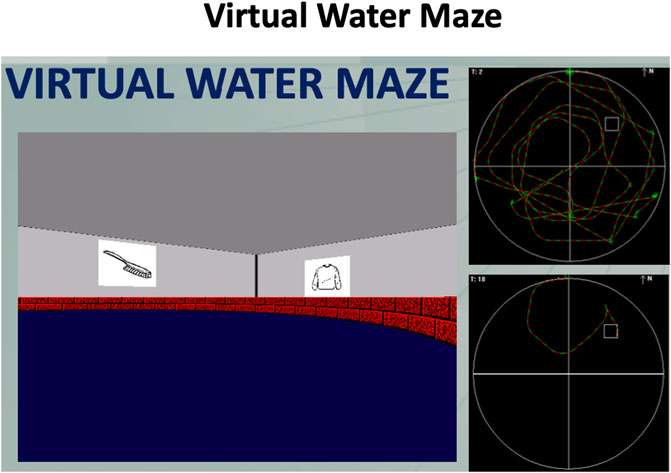
Figure 7. Screenshot of the Virtual Water Maze. Participants have to find and learn the location of a hidden target on the floor of an arena that is surrounded by landmarks (posters on the walls). After the learning phase, a probe trial is given where the target is removed unbeknownst to participants. Created with Unity Editor®. Unity is a trademark or registered trademark of Unity Technologies.
Analysis
Traditionally, the VWM version was not designed to assess and dissociate participants’ spontaneous navigation strategies. Since we previously found that participants’ spontaneous spatial and response strategies, as assessed with the 4/8 VM, correlate with hippocampus and caudate nucleus grey matter and BOLD activity, respectively (Iaria et al., 2003; Bohbot et al., 2007), these were thought to serve as a good measure of the strategies that people naturally use. Consequently, participants tested on the VWM were also tested on the 4/8 VM. Furthermore, the 4/8 VM does not show the sex differences traditionally reported in the VWM, and so assessing participants on both tasks would provide valuable information. We performed a Repeated Measures ANOVA with sex and 4/8 VM strategy as the independent variables and average latency per learning block (five blocks of four trials each) as the dependent variable. The measure of learning per block is only for the Morris Water Maze and not for the other tasks. Because approximately half the sample thought the target platform was not stable from trial to trial, even though it was always in the same location, we also included perception of stability in target location (“place constant” for those who thought the location of the platform was constant and “place changed” for those who believed the platform’s location changed from trial to trial) as an independent variable. We performed ANOVAs with the same independent variables but with average latency during learning and percentage of time spent in the correct quadrant during the probe trials as dependent variables. Examining the latency for learning block allows for the detection of different rates of learning between subgroups as the task progresses. The average learning latency is also used to detect if there were any global differences in learning between subgroups. The percentage of time spent in the correct quadrant allows for an estimation of overall accuracy that can be compared between the subgroups. Considering the trials start randomly within each block, we wanted to compare the progression of learning block to block. We use average overall latency to calculate a stable measure and compare it to the groups. More importantly, looking at learning trajectories block by block allows us to confirm a main effect of learning.
Finally, we conducted ANOVAs looking at the average latency during learning and percentage of time spent in the correct quadrant in the four groups of participants categorized by sex and strategy (spatial women, spatial men, response women, response men), both within the full sample and within the sample who believed the target location to be constant. Because several studies have found women to perform worse than men on the Morris Water Maze, and because we expected response strategy users to perform more poorly on the VMW, which is a task that can only be solved using a spatial strategy, we hypothesized that response women would have higher latencies compared to other groups. We therefore conducted a set of planned comparisons. As such, we use independent samples t-tests to investigate VWM differences (in participants who perceived the target location to be constant) between response women and two other groups: spatial women and response men, as categorized with the 4/8VM. Note that the group of spatial men who perceived the target location as constant were too small for inclusion in these sub-analyses. In the full sample, there were 8 spatial women, 3 spatial men, 23 response women, and 17 response men. In the subsample that thought the target location was constant, there were 4 spatial women, 2 spatial men, 12 response women, and 10 response men.
Results
Task performance
There was no main effect of sex in terms of latency during the five learning blocks or on average latency across all trials (Figure 8, bottom left): men (n = 12) average latency during learning (M = 21.02; SD = 13.62) women (n = 16) average latency during learning (M = 26.34; SD = 13.19). There was a significant main effect of learning blocks, in that latency decreased across blocks (F (4,172) = 5.20, p < 0.01, ε2 = 0.079). There was also a significant main effect of perception of stability in target location, in that those who thought the target location was constant (n = 28) found the target more quickly across learning blocks than those who thought the target location changed (n = 23) (F (1.43) = 14.80, p < 0.001, ε2 = 0.072) (Figure 8, top). The same was found in terms of average latency during learning (place constant: M = 20.30, SD = 10.19, place changed: M = 30.74, SD = 9.96) (F (1.43) = 14.80, p < 0.001, ε2 = 0.082). For the probe trial, there was no main effect of sex in the percentage of time spent searching in the correct quadrant women (n = 16) M = 27.52, SD = 14.84; men (n = 12) M = 20.67, SD = 10.97. However, there was a main effect of perception of stability in the target location (F (1.43) = 17.60, p < 0.001 (p = 0.002, ε2 = 0.083) whereby those who thought the target location was constant spent more time searching in the correct quadrant. There was also a tendency towards an interaction between sex and perception of stability in the target location (F (1.43) = 18.14); p = 0.06; ε2 = 0.067) where the difference in performance (time spent in the correct quadrant) between those who perceived the target location as constant and those who thought it changed is smaller in women than in men. Because the group that believed the target location was not stable may not have learned the location of the target and thus may have performed poorly, we conducted the same analyses as above but only in the group that believed the target location was stable. There was no main effect of sex on any of the variables (p > 0.05) (Table 4).
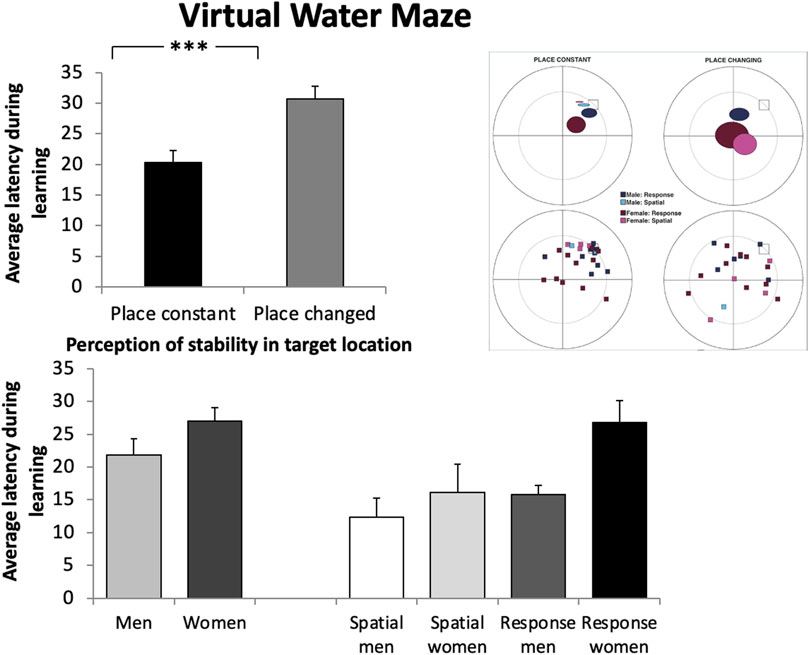
Figure 8. Top: Participants who believed the target location changed from trial to trial (place changed) had a significantly higher average latency than participants who believed it to remain in the same location (place constant) (F (1 (43) = 14.80, p < 0.001). Bottom left: Women and men did not differ in their average latency to find the target (in seconds) in the Virtual Water Maze (p > 0.05). Bottom right: When participants were categorized into four groups based on their sex and navigation strategy, response women had a significantly greater average latency than response men (t = 3.01, p < 0.025). However, an important limitation is that the sample sizes of the groups were low (spatial men: n = 2, spatial women: n = 4, response men: n = 10, response women: n = 12). Error bars represent standard errors of the mean. *p < 0.025; ***p < 0.001.
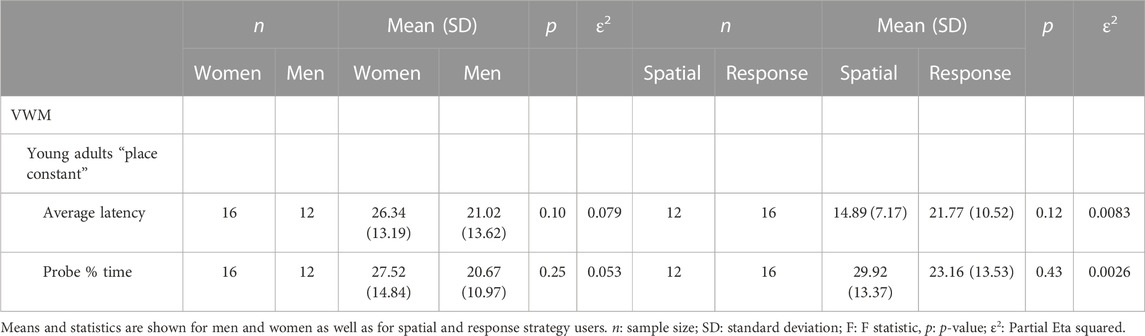
Table 4. Virtual Water Maze (VWM): Means and statistics for latency and percentage of time spent in the correct quadrant means and statistics for young adults who perceived the platform location as being constant.
Navigation strategies
In addition to the VWM, participants were tested on the 4/8 VM. No main effect or interactions with strategies was found: (p > 0.05) (Table 4): Spatial place constant (n = 12) M = 14.89, SD = 7.17; Response place constant (n = 16) M = 21.77 (10.52); ε2 = 0.0083; Spatial latency probe % time (n = 12) M = 29.92, SD = 13.37; Response latency probe % time (n = 16), M = 23.16, SD = 13.53); ε2 = 0.0026. Planned comparisons between the three groups of participants in the sample that perceived the platform as stable (response women vs. spatial women, response men) revealed that response women did not differ from either group on the probe trial (p > 0.05; ε2 = 0.002) in terms of average latency during learning, response women were slower than response men (t (26) = 3.01, p < 0.025; Cohen’s d = 0.45) (Figure 8, bottom right). Although the number of participants is low and statistics cannot be assessed for the spatial groups, it is interesting to note that the average latency to the target is similar for the spatial men and the spatial women (spatial men M = 13.14, SD = 6.12; spatial women M = 16.89, SD = 8.32). Finally, the standard deviations (SDs) observed in this analysis are similar, except for the spatial male group when comparing each of the 5 blocks (7.45 vs. 21.50 vs. 7.75 vs. 12.42 vs. 10.72). Due to the relatively low sample size and the variation in standard deviations, there is a possibility that the means might not accurately represent the characteristics of the larger population. It is also worth noting that a larger sample size may yield a statistically significant effect, or alternatively, the observed difference could be attributed to measurement error. Consequently, when interpreting the results, it is crucial to consider these factors.
Discussion
Characteristics of the VWM environment
The standard Morris Water Maze requires participants to estimate angles and distances in a precise way to reach the target in a direct fashion. We would therefore consider this task to confer an advantage to men. However, there appears to be other factors at play that determine whether sex differences emerge or not, such as spontaneous navigation strategies, number and proximity of landmarks, size of the environment, and navigation interface (joystick vs. keyboard). These factors are discussed in the “Relation of results with current literature” section below.
Navigation strategies
In the current study, response women had a lower average performance (i.e., a higher latency) than response men. No effects were found among spatial strategy users. Although the current sample was small, the identification of the target placement was fairly homogenous and not different from one another in men and women spatial learners. There was no difference in terms of probe performance according to either sex or navigation strategies, indicating that other factors may modulate the emergence of sex differences in locating the platform.
Relation of results with current literature
The Morris Water Maze literature is widely discrepant when it comes to sex differences. Some studies found men to outperform women (Astur et al., 1998; Astur et al., 2004; Rizk-Jackson et al., 2006; Mueller et al., 2008; Chai and Jacobs, 2009; Nowak and Moffat, 2011), while other studies found no sex differences (Driscoll et al., 2005; Levy et al., 2005; Moffat et al., 2007; van Gerven et al., 2012).
Interestingly, two studies by Piccardi et al. (2011) and Roof and Stein. (1999) found a male advantage when the virtual environment was devoid of landmarks or contained landmarks that were not predictive of the platform’s location, and this male advantage disappeared once landmarks were present and stable, consistent with the results from Andersen et al. (2012). These studies may shed light onto what kind of conditions lead to a male advantage and which do not lead to any sex bias. Specifically, landmark proximity may be a factor in sex differences. Some studies refer to proximal and distal landmarks while others use the terms positional and directional landmarks. There is no clear definition for these cue types; for example, it is not always clear whether proximal landmarks are located inside a maze or outside the periphery in close proximity. Similarly, it is not clear whether distal landmarks are considered to be outside the maze or whether they are distant in the background. To avoid confusion, we will not address intra-maze cues as there typically are none in MWM paradigms. We will therefore refer only to landmarks outside the maze, placed either close to the periphery of the virtual pool, which we will refer to as proximal, or further away in the distance, which we will refer to as distal.
Chamizo et al. (2011) used a Morris Water Maze task with four landmarks: two of which were close to the perimeter of the virtual pool (proximal) and two of which were more distant (distal). In a series of probe trials, the authors removed the platform and also manipulated the environment in different ways. When the proximal landmarks were removed and only the two distal landmarks remained, men outperformed women in that they spent more time searching in the correct quadrant where the platform was previously located. When only one distal landmark was present, performance fell to chance for both men and women. When the distal landmarks were removed and the proximal landmarks remained, men and women spent the same amount of time in the correct quadrant. These findings indicate that, while men could rely on both proximal and distal landmarks to locate the platform, women had a preference for the proximal landmarks. Considering males tend to use geometric information whereas females rely more on landmarks, a study by Aguilar-Lattorre et al. (2022) attempt to alter the geometry bias in female rats. The first experiment comprised of 3 groups of female rats that were trained in a triangular-shaped pool to find a hidden platform. There were two main sources of information to find the hidden platform, a landmark outside the pool and a particular corner of the pool. The females with prior experience of two other pool shapes–with a kite-shaped pool and with a rectangular-shaped pool were significantly more accurate than control rats during the trial with a triangular pool and no landmark. Further, males performed significantly better than females when both had no experience and were faced with the triangular pool and no landmark (Aguilar-Lattore et al., 2022) These results can explain the framework of selective attention and suggest that experience with different frameworks can help overcome sex specific biases.
These findings are supported by our Invisible Sensor Task study (discussed below), which used a real-life task modelled after the Morris Water Maze, in which we have shown that women preferentially use proximal landmarks compared to men.
Additionally, Chamizo et al. (2011) found that the presence of only one proximal (extra-maze) landmark yielded worse performance than the presence of two, in both men and women. Chai and Jacobs (2009) found that women’s lower performance compared to men in a MWM-type task was exacerbated when the environment was devoid of proximal (intra-maze) landmarks. This is in agreement with our previous study (Andersen et al., 2012)72, in which we showed that women do poorly when few or no landmarks are available in the 4/8 VM and that women benefit from the presence of multiple landmarks. In our study, there were multiple landmarks and they were located on the walls which themselves were close to the perimeter of the virtual pool. Therefore, the number of landmarks and their proximity may explain why we did not observe sex differences in the average latency to reach the platform or in the ability to locate the platform in the probe trial.
Sandstrom et al. (1998) investigated the role that landmarks, and room geometry play in learning to locate a target platform. Their paradigm included four landmarks around the virtual pool and the irregular shape of the room provided useful geometrical information that could be used to learn the placement of the platform. In probe trials where the landmarks were absent or moved around, men had a smaller latency error (the time taken beyond the minimum time it takes to reach the platform) than women. However, when the landmarks were present in their original locations and the geometry was modified, men and women performed equally well. This study suggests that men are able to use both geometry and landmark information to find the platform, while women lean towards using the landmarks. Moreover, although the authors do not specifically mention this, it is of note that the landmarks are close in proximity to the virtual pool, which might explain why there are no sex differences when these landmarks are made available, as in Chamizo et al. (2011)’s study.
The size of the MWM environment may also modulate sex differences. We would expect large open environments to lead to a male advantage because of men’s ability to better estimate angles and distances, similar to what is seen in a large-scale virtual wayfinding task. Although it is not possible in the current paper to objectively measure the environment size of the MWM paradigms used in previous studies, it is possible that larger environments, as seems to be the case for at least some of the studies that yielded a male advantage (Astur et al., 1998; Astur et al., 2004; Rizk-Jackson et al., 2006; Mueller et al., 2008; Chai and Jacobs, 2009; Nowak and Moffat, 2011), bias performance in favour of men. Accordingly, at least some of the studies in which no sex differences were observed (Driscoll et al., 2005; Levy et al., 2005; Moffat et al., 2007; van Gerven et al., 2012) seem to have a smaller virtual pool and/or surrounding room.
Finally, all the studies that found a male advantage (Astur et al., 1998; Astur et al., 2004; Rizk-Jackson et al., 2006; Mueller et al., 2008; Chai and Jacobs, 2009; Nowak and Moffat, 2011) used joysticks to navigate in the virtual environments. In the other studies, half used such methods as keyboards (Driscoll et al., 2005; Chamizo et al., 2011; van Gerven et al., 2012) and half used joysticks (Sandstrom et al., 1998; Levy et al., 2005; Moffat et al., 2007). Although many studies include joystick training, some do not and this may partly account for the presence of a male advantage as some of the studies have shown that men are more proficient with joysticks (Rizk-Jackson et al., 2006; Mueller et al., 2008; van Gerven et al., 2012).
Thus, although not empirically tested in the current study, we propose that the emergence of sex differences in the MWM may depend on the following factors: number and proximity of landmarks, size of the environment, navigation interface (joystick vs. keyboard) and navigation strategies.
Experiment 5: invisible sensor task (IST)
Methods
Participants
Healthy young adults, aged between 18 and 35 (N = 52; women: n = 26; men: n = 26), were tested on the Invisible Sensor Task (IST) which is a real-life spatial memory test modelled after the Morris Water Maze adapted from Bohbot et al. (1998). A subset of them were tested on the 4/8 VM (women: n = 15; men: n = 15) to investigate whether spontaneous strategies have an impact on the IST.
Invisible sensor task
The IST is a real-life spatial task that was modelled after the Morris Water Maze. The current version of the IST is a modernized cue-controlled environment taking place in a symmetrical square room (5 m × 5 m) containing nothing but a large screen on each of the four black walls. There are two doors on every corner of the room, allowing multiple entries to the room, but also to prevent participants from orienting using the three functional entrance doors from which they could come in. A panoramic view of a valley is projected on the screens, with one landmark that appears closer (hereby referred to as the proximal landmark) and others that appear more distal (hereby referred to as distal landmarks) (Figure 9). As the landmarks are projected on screens around the periphery of the room, they are considered to be extra-maze landmarks. Participants have to learn the location of a virtual target inside the room. The target is detected with a light emitting diode placed at the top of a 1.9 m pole, which is moved around by participants across the floor. The relationship between the projected visual environment and target location is kept constant between trials, but they are rotated between trials such that learning the target location is done independently of the physical environment. In addition, the start position changed between trials and was indicated by arrows pointing to a corner of the environment. The start position changed in relation to the projected visual environment such that the target location could not be learned with an egocentric set of motor actions from the start position. Before each trial, participants were also asked to go to the centre of the room and rotate clockwise and counter-clockwise between trials, in order to allow themselves to be disoriented so as to ignore the physical environment, including the entrance door to the room. Consequently, participants could either learn the target location in relation to the proximal landmark or they could learn the target location in relation to the distal landmarks. Participants reached asymptotic performance when they located the target within 13 s, with a criterion of three times in a row. To determine which type of landmarks participants used to learn the location of the target, the proximal landmark is moved to a different position in the landscape during a probe trial (probe 1). Thus, there are two locations at which participants can find a target: either they learned the position of the target in relation to the distal landmarks which are part of the scenery (mountains) or they learned the position of the target in relation to the proximal landmark (e.g., the barn). The one they choose will reveal whether participants used proximal or distal landmarks to learn the location of the target. Following this, in another trial, the proximal landmark is removed completely and only the distal landmarks remain (probe 2). This last probe is also meant to assess whether participants have acquired information about distal landmarks and can use them to find the target location whether or not they had acquired the task using the proximal landmark, which is now absent. Participants often paused when noticing the change in landscape before engaging in the task as such. For this reason, we found that participants going in a straight path to the target did so under 27 s, which was used as a cut off for a trial to be included as successful.

Figure 9. Picture of the symmetrical room and task. A panoramic view of a landscape is projected on 4 screens located on the 4 walls. The landscape contains proximal (extra-maze) and distal (also extra-maze) landmarks. Participants move a sensor pole that beeps once they find the virtual target. After the learning phase, two probe trials are given. In probe 1, the proximal landmark is moved to a different position in the landscape. In probe 2, the proximal landmark is removed completely and only the distal landmarks remain. Created with Unity Editor®. Unity is a trademark or registered trademark of Unity Technologies.
Analysis
For the learning phase, we conducted an independent samples t-test to assess whether there was a significant difference between men and women in the number of trials to criterion. We performed a chi-square test to test whether men and women use one type of landmark (proximal or distal) more than the other based on probe 1 performance. We conducted another chi-square test to directly compare men and women on their proportion of proximal and distal cue users in probe 1. For this latter test, we used a one-tailed chi-square test because previous studies have shown women to rely more on proximal landmarks than men (Barkley and Gabriel, 2007; Chai and Jacobs, 2009; Chamizo et al., 2011). To examine the latency to reach the target on probe 1, a two-way ANOVA was performed with latency as the dependent variable and with sex and cue type used as the independent variables. For probe 2, an ANOVA was conducted with latency as the dependent variable and sex as the independent variable. To investigate the proportion of men and women who found the target within 27 s in the absence of the proximal landmark (probe 2), a chi-square analysis was conducted with sex as the independent variable and probe 2 success as the dependent variable. To investigate navigation strategies, an ANOVA was performed with sex and strategies as the independent variable and trials to criterion as the dependent variable. Finally, a chi-square analysis was conducted to examine whether the proportion of spatial and response learners differed between proximal and distal cue users, within each sex group.
Results
Task performance
Men and women learned the task at a similar rate; they took the same number of trials to learn the task to criterion: women n = 26 (M = 7.46, SD = 3.46); men n = 26 (M = 6.27, SD = 3.05); ε2 = 0.044; p > 0.05) (Table 5). On probe 1, women used the proximal landmark to a greater extent than the distal landmarks to locate the target (χ2 (1, N = 26) = 7.54, p < 0.01; Cramer’s V = 0.24), while men use both types of landmarks equally often (χ2 (1, N = 26) = 0.62, p > 0.05; Cramer’s V = 0.24) (Figure 10). A greater proportion of women tended to use the proximal landmark compared to men: women proximal n = 20; women distal n = 6; men proximal n = 14; men distal n = 12; χ2 (1, N = 26) = 2.19, p = 0.07; Cramer’s V = 0.24) (Figure 10). In terms of latency to find the target, men and women are equally good at finding the target when both proximal and distal landmarks are present: women probe 1 latency n = 26 (M = 26.78, SD = 41.11); men probe 1 latency n = 26 (M = 20.00, SD = 15.29); p > 0.05; ε2 = 0.002 (probe 1; Table 5) or when only distal landmarks are present (probe 2), irrespective of the type of landmarks used: women probe 2 latency n = 26 (M = 29.52, SD = 26.31); men probe 2 latency n = 26 (M = 23.55, SD = 17.91); p > 0.05 In the absence of a proximal landmark (probe 2), the proportion of women and men who found the target within 27 s was similar, no matter which type of landmarks they used in probe 1: women distal n = 4; women proximal n = 10; men distal n = 10; men proximal n = 7 (p > 0.05).
Table 5. Invisible Sensor Task (IST): Means and statistics for number of trials to criterion and probe 1 latency for young adults.
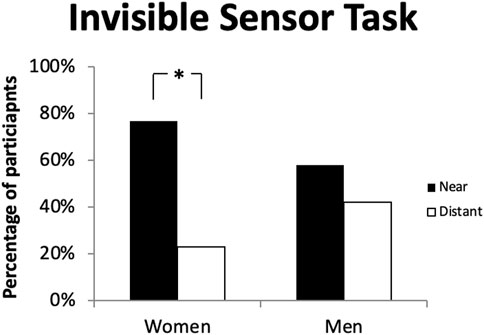
Figure 10. On probe 1, women used the proximal landmark to a greater extent than the distal landmarks to locate the target (χ2 (1, N = 26) = 7.54, p < 0.01; Cramer’s V = 0.24). A greater proportion of women tended to use the proximal landmark compared to men: women proximal n = 20; women distal n = 6; men proximal n = 14; men distal n = 12; χ2 (1, N = 26) = 2.19, p = 0.07; Cramer’s V = 0.24.
Navigation strategies
Spontaneous navigation strategies were assessed with the 4/8 VM for a subset of participants (n = 28: 6 spatial women, 5 spatial men, 7 response women, 10 response men). Navigation strategies modulated learning in the IST: response learners tended to require fewer trials to reach the learning criterion: response n = 17 (M = 5.23, SD = 1.56); spatial n = 11 (M = 6.63, SD = 2.06) (F (1.26) = 4.17, p = 0.05; ε2 = 0.079 (Table 5). There was no interaction between sex and navigation strategies in learning the target location (p > 0.05). There was an equal proportion of spatial and response learners in both the proximal and distal landmark user groups in both men and women: response men distal n = 5; response men proximal n = 5; spatial men distal n = 2, spatial men proximal n = 3; χ2 (1, N = 15) = 2e−31; (p > 0.05; Cramer’s V = 0.09); response women distal n = 4; response women proximal n = 3; spatial women distal n = 3; spatial women proximal n = 3; χ2 (1, N = 13) = 0; (p > 0.05; Cramer’s V = 0.07) (p > 0.05). Thus, there is no relationship between the spontaneous navigation strategy used in the 4/8VM and the type of landmarks used on probe 1. Finally, spontaneous navigation strategies did not influence probe 1 latency: spatial probe 1 latency n = 11 (M = 29.04, SD = 6.47); response probe 1 latency n = 17 (M = 25.37, SD = 11.23); F (1, 26) = 1.28; ε2 = 0.053; p > 0.05) (Table 5). For the probe 1 latency, given the differing standard deviations (6.47 vs. 11.23) and the low sample size, it is possible that the means may not accurately reflect the characteristics of the larger population. A higher sample size might return a significant effect, or this could be simply due to measurement error. it is imperative to consider these factors when interpreting the results.”
Discussion
Characteristics of the IST environment
The IST requires to precisely locate a target inside a room. However, the room size is small in scale and therefore does not require participants to integrate distances and angles over a large environment. Although the target is within the confines of a single room, it is not possible to locate the target within a single viewpoint, as one would need to turn around to see all the components of the virtual environment projected onto the four walls of the room. Additionally, the projected environment contained both proximal and distal landmarks, which accommodates women since they generally rely on proximal landmarks more than on distal landmarks (Barkley and Gabriel, 2007; Chai and Jacobs, 2009; Chamizo et al., 2011).80 For these reasons, we do not consider this task to confer an advantage to either men or women. Accordingly, men and women’s learning rate did not differ in the IST. We replicated the finding that women rely more on proximal than distal landmarks.
Navigation strategies
Spatial learners were slower than response learners in learning the location of the target, regardless of sex. Iaria et al. (2003) found similar results, whereby spatial learners made more errors and took more time to learn the object locations on the 4/8 VM when compared to response learners, suggesting that building relationships between landmarks with the spatial strategy is more cognitively demanding than the response strategy (Nadel and Hardt, 2004).
When the proximal landmark was removed (probe 2), most participants were still able to find the target. This finding suggests that even though most participants used the proximal landmark instead of the distal landmarks when locating the target, as shown in probe 1, they also learned the relationship between the target and distal landmarks. It is possible that this learning was incidental. This finding also demonstrates that participants are flexible in their landmark use.
Relation of results with current literature
The fact that men and women’s performance was the same during learning is supported by a study by Chai and Jacobs (2009). In this study, the authors used a MWM-type task where participants had to locate a hidden target in a virtual environment that contained both proximal (intra-maze) and distal (extra-maze) landmarks. In such an environment, men and women were equally good at locating the target.
Our findings that women rely more on proximal landmarks are consistent with those of Chamizo et al. (2011) and Chai and Jacobs (2009). Chamizo et al. (2011) found that, when proximal (extra-maze) landmarks were removed in a MWM paradigm, women’s ability to locate the platform was impaired. In Chai and Jacobs (2009), using an environment without proximal landmarks that provided positional cues increased the performance difference between men and women compared to when these landmarks were present. The current study shows that, when both proximal and distal landmarks are available, women will use the proximal landmark to learn the location of a target more than men. Therefore, women’s greater reliance on proximal landmarks is replicated in the current study. One difference between Chamizo et al. (2011)’s findings and ours is that we did not find women to be impaired when the proximal landmarks were removed from the virtual environment. In our paradigm, we used photographs of real places in which the position of proximal landmarks overlapped with that of distal landmarks, i.e., distal landmarks (e.g., mountains) could be seen behind the proximal landmark (e.g., barn). Therefore, it is possible that, in the absence of the proximal landmark in probe 2, women associated the nearest distal landmark (a mountain) with the previous position of the proximal landmark and used it to find the target. Such a method would allow women to compensate for the removal of proximal landmarks. In other words, during acquisition of the task, women could have associated the proximal and distal landmarks since they overlap and therefore the removal of the proximal landmark did not impact their performance. We may have observed different results if the landmarks were not overlapping. Alternatively, features of the environment or other experimental design details may encourage the acquisition of distal landmark information in our study but not in the study by Chamizo et al. (2011).
Our results are also supported by a study by Barkley and Gabriel (2007) in which participants viewed a photograph of a scene followed by another photograph that was either an altered version of the original photograph, the exact same photograph, or a different photograph. The authors found that women were slower to identify pairs of photographs in which the altered version had a proximal landmark removed but were not impaired when a distal landmark was removed. Men, on the other hand, were impaired in both cases. This particular study demonstrates that women have a preference for proximal landmarks compared to distal landmarks and that men process distal landmarks to a greater extent than women.
In summary, we have replicated the previous finding that women rely more on proximal than distal landmarks. However, when they are learning the location of a target, women learn as quickly as men. Interestingly, the use of proximal and distal landmarks seems to be unrelated to spontaneous spatial and response strategies. This suggests that women favor proximal landmarks even when they use the spatial strategy in the 4/8 VM, previously shown to be dependent on the hippocampus (Iaria et al., 2003; Sodums and Bohbot, 2020).
Experiment 6: object location memory task
Methods
Participants
Healthy adults between the ages of 32 and 55 (N = 13; women: n = 7, M age = 44.00, SD = 7.81; men: n = 6, M age: 42.50, SD = 7.72) were tested on the object location memory task. A subset of this sample served as a control group in a lesion study (Stepankova et al., 2004).
Object location task
Participants had to learn the location of one to six objects on the floor of a circular arena enclosed by curtains by looking at the objects for 10 s. For each set of objects, participants had to indicate the location of each object on a sheet of paper using photo icons of the objects (Figure 11). Then, the objects were placed together in the center of the arena and participants had to place the objects where they previously had seen them in the arena (see Stepankova et al. (2004) for further details). The main dependent variable was mapping error in placing the icons on the sheet of paper and arena error in placing the objects in the arena. Navigation strategies were not assessed.
Figure 11. Schematic of the object location memory task, shown here with three objects. In this task, participants have to learn the location of up to six objects within an arena enclosed by curtains. The arena spans 3 m in diameter. Objects are shown here disproportionately larger to make them visible.
Analysis
We conducted two Repeated Measures ANOVAs with each of the location error measures (map error and arena error) as the repeated measure for each object set size (1–6) and with sex as the independent variable. We also conducted a MANOVA with sex as the independent variable and the various measures of learning as the dependent variables, including average map error and average arena error.
Results
Task performance
In the Repeated Measures ANOVA, there was a significant main effect of object set size (F (3, 28.98) = 3.13, p < 0.05, ε2 = 0.78) and sex (F (1, 11) = 5.88, p < 0.05, ε2 = 0.067) on map error (Figure 12). As object set size increased, map error increased as well.
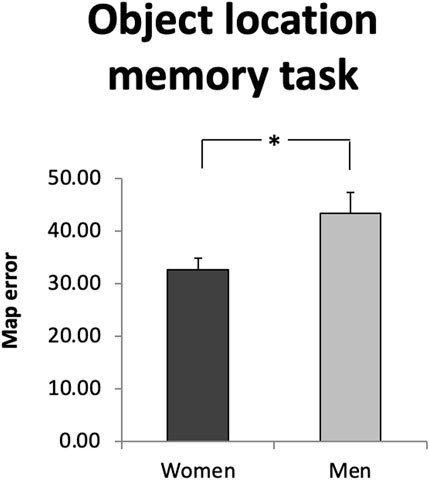
Figure 12. After learning the location of the objects in the arena, had to place icons representing the objects in their correct locations on sheet of paper showing a top view of the environment. Women were more accurate in placing the objects on the map than men (F (1, 11) = 5.88, p < 0.05). Error bars represent standard errors of the mean.
The overall average map error was 36.40 with a range of 37.88 (maximum: 61.83; minimum: 23.95; SD = 12.23). In terms of sex differences, men (M = 43.40, SD = 12.85) had a greater map error than women (M = 32.68, SD = 6.75) across the object sets. Considering our low sample size and difference in SD’s (6.75 vs. 12.85) it is possible the means may not accurately represent the underlying population. This makes it more challenging to draw reliable conclusions between our two groups. In addition, it is important to acknowledge that the observed effects in our study may be underpowered. Therefore, we performed a post hoc power analysis, which revealed a power value of 9.73% indicating that our study may have limited ability to detect significant effects due to small sample size. Further, Men had a range of 34.43 (maximum: 61.84; minimum: 27.41) and women of 18.15 (maximum: 42.10; minimum: 23.95) in map error. For arena error, there was no main effect of object set size or sex (men: M = 16.26, SD = 4.98; women: M = 14.67, SD = 3.14) (p > 0.05) (Table 6). For arena error, men had a range of 12.65 (maximum: 22.99; minimum: 10.34), and the women had a range of 9.27 (maximum: 19.69; minimum: 10.42). The average arena error was 15.92 with a range of 12.23 (maximum: 22.65; minimum: 10.42; SD = 4.04).
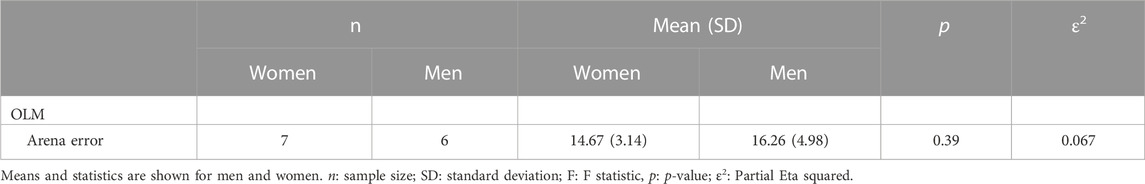
Table 6. Object Location Memory task (OLM): Means and statistics for arena error in adults.
Discussion
Characteristics of the object location memory task environment
To encode the location of the objects, participants had to stand in the entrance of the arena and turn their heads left and right to view all the objects. Although all the objects did not fit within a single field of view, there was no navigation required (participants were standing still) in order to view all the items. In such circumstances, the typical female advantage in object location was observed, but only when the objects had to be placed on a piece of sheet and not in the arena. We suggest that recalling the position of the objects on a sheet of paper requires a more thorough reconstruction process than recalling the position of the objects in the arena. One reason is that in the arena, participants can view the objects from the same perspective, whereas recalling the position of objects on a piece of paper requires a transformation of the information from first-person view to overhead map. Another reason is because the arena environment provides the context of the learning situation, and therefore can serve as a primer and help all participants to perform well. Placing the objects on the sheet of paper is a different context and therefore there is no priming effect, which requires participants to undergo a reconstruction process from memory.
The female advantage observed may have emerged from the fact that women use the relationship between objects but also, because in this study, the arena was composed of a set of curtains shaped in a circle. The fact that the arena was circular and surrounded by a curtain, implies that participants could not use Euclidean coordinates or cardinal directions to learn the location of the objects, thereby disabling the male advantage.
Relation of results with current literature
Object location memory is also an area of contradictory findings (see Voyer et al., 2007 for a review). Voyer et al. (2007) propose that there are several factors that can modulate sex differences in object location memory, including object type and type of measure. Most studies of object location memory used arrays of objects presented on sheets of paper or computer screens (Silverman et al., 1992; Postma et al., 1998; Saucier et al., 2007). Here, we will only consider studies that use real-life objects that did not fit within a single field of view. To our knowledge, only two such studies exist. Iachini et al. (2005) had participants learn the location of seven objects placed inside a large arena. Then, the objects were gathered together along with seven distractor objects. Participants had to choose the correct objects and place them in the arena in their respective locations. They found that men and women did not differ in object location error. This measure is akin to our arena error and the lack of sex differences in this type of measure supports our finding that men and women performed equally well when they had to recall the location of the objects inside the arena.
Montello et al. (1999) asked participants to memorize the identities of 35 objects placed on two table tops, walls, and on the floor underneath the tables. Then, participants had to place a label for each object onto a sheet of paper on which was depicted the tables, walls, and floor. The authors found women to more precisely place the objects on the sheet of paper and they also found them to misplace fewer objects than men. These results are in accordance with our map error results, for which we have observed better scores in women than men. Our findings as well as the studies described above indicate that women have a better memory for object locations when they have to use a thorough reconstruction process to recall the positions of the objects and that the positions of objects have to be learned in relation to each other, without necessarily obtaining Euclidian coordinates and cardinal directions.
General discussion
Young men and women performed similarly on the two virtual radial mazes, as predicted, because these tasks are not sensitive to the male or female advantage. Women performed better than men in the Object Location task, as predicted, because this task is sensitive to the female advantage involving learning the relationships between items without precise distances and Euclidian coordinates. We found a sex difference in the use of distal and proximal landmarks, but this was independent of spatial and response navigation strategies. Moreover, men outperformed women in the Wayfinding and Virtual Water Maze tasks, as predicted, because these tasks are sensitive to the male advantage. Interestingly, while we replicated the sex differences previously reported in the Virtual Water Maze showing an impairment in women, we showed that the impairment was not present in spatial women. Instead, this effect was specific to response women only. A sex differences did emerge on the 4/8 VM and CSDLT radial mazes in aging, when navigation strategies were taken into account. More specifically, older response women made fewer errors during the probe trial of the 4/8 VM, suggesting reduced use of landmarks and reduced recruitment of the hippocampus.
The tasks that did not have environmental characteristics that favoured one sex or the other, such as the 4/8VM and CSDLT, yielded no sex differences. These tasks have restricted navigation: participants could only navigate on defined pathways and were not free to move outside of these pathways. Therefore, the male advantage that is usually seen in open environments due to men’s better ability to estimate distances and angles was not a factor in these tasks. Additionally, all the landmarks in the 4/8 and CSDLT are salient, therefore both men and women were able to navigate using their preferred cue type or both (Barkley and Gabriel, 2007; Chai and Jacobs, 2009; Chamizo et al., 2011). Men and women not only did not differ in their performance, but they also did not differ in the navigation strategies that they used, as they used spatial and response strategies in the same proportion.
For the IST task, women used the proximal landmark to a greater extent than the distal landmarks to locate the target, yet there were no sex differences in the learning rates. Since the IST is comprised of an open environment but it is small in size, it is possible that a male advantage did not emerge for this reason as the estimation of angles is difficult in a small environment.
The wayfinding task in a virtual town is an open environment and is large in scale, in which we hypothesized to find a male advantage based on the literature. Considering women are at a disadvantage regarding estimating angles and distances (Holding and Holding, 1988; Postma et al., 1998; Lawton and Morrin, 1999; Postma et al., 2004; Iachini et al., 2005), it is possible that a small error in estimating angles and distances in a small-scale environment, would get magnified when navigating in a town. In other words, estimation errors would have accumulated as women explored the virtual town, yielding a less accurate cognitive map of the town. Young women found fewer locations and were slower at finding the targets than men.
The emergence of a sex difference in the object location memory task depended on the measure used, even though the same knowledge was being assessed. This indicates that the way performance is measured is important. In this case, when the object locations had to be recalled completely from memory (i.e., by having to draw an overhead map of the arena and its objects), then women’s memory of object locations was more accurate. When the object locations had to be recalled within the same context as the learning phase (e.g., by placing the object in the arena where the object locations were learned), then men and women performed similarly. This finding suggests that women are better when they have to do a complete reconstruction of the objects’ placements. Consistent with the literature, the object location memory task showed that women had a better performance than men at recalling an overhead map of the relationships between objects. Importantly, the environment was devoid of a reference frame, thereby eliminating the male advantage previously seen in navigation task.
For the Virtual Morris Water Maze women spatial learners learned at the same rate as men and that only women response learners showed an impairment on this task. Based on the literature, we hypothesized that this task would confer an advantage to men, as it contains an open environment in which participants can navigate in any direction they want. Other experimental factors are at play in the maze that may influence the sexes differently. Some of these factors can help explain why some studies found sex differences (Astur et al., 1998; Astur et al., 2004; Rizk-Jackson et al., 2006; Mueller et al., 2008; Chai and Jacobs, 2009; Nowak and Moffat, 2011) while others did not (Driscoll et al., 2005; Levy et al., 2005; Moffat et al., 2007; van Gerven et al., 2012). These include the size of the environment, the presence or absence of landmarks, the proximity and number of landmarks, and the navigation interface (e.g., joystick vs. keyboard). More precisely, we suggest that smaller environments, the presence of proximal and multiple landmarks, and using a keyboard as a navigation interface reduce the performance difference between men and women. It is also important to note that many participants misperceived the platform as changing locations from trial to trial, causing noise in the data as these individuals never learned the task. Only those who believed the platform to be stable showed normal learning. Therefore, it is important to take this into account when analyzing data from virtual water maze-type paradigms.
The factors mentioned likely apply to all navigation paradigms, and additionally include the structure of the environment (open vs. restricted, for example, by having hallways lead from one place to another) and type of measure.
Navigation strategies also play a role in performance. In some paradigms, spatial strategy users and individuals who used landmark-based strategies performed better (e.g., CSDLT, virtual town), while in others response strategy users performed better (e.g., IST). Importantly, when it comes to sex differences, some of our paradigms showed interactions between sex and navigation strategies, which became emphasized with age. In the 4/8 VM, older response women made fewer mistakes on the probe than all other groups. In the CSDLT, young response men performed more poorly on the probe and older spatial women performed best on the probe. In the VWM, young response women had longer latencies during learning than response men. This highlights the importance of accounting for navigation strategies, as these effects may have been missed or misinterpreted were it not for navigation strategies. For example, there was a main effect of sex in young adults who performed the CSDLT, whereby men performed more poorly than women on the probe. However, this effect was explained by an interaction, in which response men were driving the effect. If navigation strategies had not been accounted for, we would have mistakenly concluded that women outperform men on the CSDLT. Another factor to keep in mind is the fact that navigational strategies are rarely measured in other studies in the literature. Should the proportion of spatial and response learners vary depending on the sample, it would lead to a bias and misinterpretations of data if one navigation strategy is over-represented. For example, if in one sample women happen to predominantly use response strategies while men use both strategies equally, then we might find that women do more poorly than men if response strategies are associated with worse performance on a given paradigm, as is the case in the MWM. This difference would not be inherent to sex but rather to the greater proportion of response strategy users in that particular sample of women. Therefore, accounting for navigation strategies may help to avoid misinterpreting results.
Finally, age also plays a role in sex differences. Age affects men and women differently: there are changes in hormones that occur with aging (e.g., menopause) but there are also differences in the normal volume decline of the brain. For example, the volume of an area known to be critical to spatial learning, the hippocampus, declines at a faster rate in men than in women (Pruessner et al., 2001). Therefore, if young men perform better than young women in a task that elicits activity in the hippocampus and in which only spatial strategies can be used, such as the wayfinding task (Hartley et al., 2003; Head and Isom, 2010; Ledoux et al., 2013), then this male advantage would decrease and disappear with aging as the volume of the hippocampus declines. For other tasks that do not confer an advantage to one sex or the other and that allow the use of either spatial or response strategies (e.g., 4/8 VM, CSDLT), a sex differences may emerge with age.
The current results also have implications for healthy cognitive aging and Alzheimer’s disease. On the 4/8 VM, older response women made fewer mistakes on the probe trial than the other sex and navigation strategy groups. We have previously found that low probe score on the 4/8 VM negatively correlate with grey matter in the hippocampus (Bohbot et al., 2007). Low hippocampal volume is a strong risk factor for Alzheimer’s disease (den Heijer et al., 2006; Fox et al., 1996; Jagust et al., 2006; Kaye et al., 1997). Therefore, our 4/8 VM results suggest that women who spontaneously use response strategies on the 4/8 VM have lower hippocampal grey matter and as a result may be more at risk for the disease. This hypothesis was confirmed by Sodums and Bohbot (2020). This is important because if we can identify individuals that are most vulnerable, such as older women who spontaneously use response strategies, future clinical implications such as hippocampus stimulation can be considered as potential prevention or as a possible treatment.
Limitations
It is important to explore other factors that could lead to differences in spatial memory performance in men and women. A study by Montello, 2017 suggests that the role of landmarks in spatial cognition is exaggerated and the exact term “landmark” is ambiguous. Further, with object location memory task, the associations built between landmarks also requires learning metric information, which is not independent to the associations learned between landmarks (Chrastil and Warren, 2014). More specifically, Chrastil and Warren, 2014 showed that participants traveled the shortest metric distance to a destination instead of using pure topological knowledge. In our tasks used, we need to consider the possibility that the pure topological information was not enough for women to complete the tasks, or to show their advantage over men. Taking into consideration that men performed better during the wayfinding task, we can also explore the idea that this was due to a more efficient exploration strategy or that they perform at an advantage in a large environment (Brunec et al., 2023).
Another limitation of our study is the relatively low sample size in certain experiments, particularly experiments 5 and 6. As a result, the ability to detect significant effects in these experiments is decreased. This limitation should be taken into consideration when interpreting the findings from these specific experiments.
A significant limitation is our study is that the menstrual cycle phase and contraception were not controlled in young women. This is important as there has been evidence to show that sex steroid hormone levels can have an impact on spatial memory and navigation strategy along with hippocampus size (Hussain et al., 2016 and Pletzer et al., 2010). Furthermore, in older adults, hormone replacement therapy and hysterectomy/ovariectomy was not assessed nor controlled. Both of these have an impact on cognitive functioning and risk of AD (Henderson, 1994). Moreover, in the object location task, the women were aged between 32 and 55 years old but the reproductive age was not assessed such as pre, peri or post menopause which can have an effect on memory, in particular peri-menopause (Unkenstein et al., 2016). Finally, hormones levels were not assessed in this study thereby contributing to the limitations described above.
Conclusion
We found evidence of sex differences at a group level with a female and a male advantage. Importantly, we found that certain tasks that traditionally showed sex differences only apply to response learners. Finally, sex differences interact with spontaneous navigational strategies and get augmented with aging, thereby providing for the first time, a hypothesis for explaining why women are more prone to Alzheimer’s disease than men, more specifically response women. Even in paradigms widely thought to favour one sex, there are still many factors that determine whether sex differences will emerge or not. We hope that describing these factors will help researchers to develop paradigms that will target specific abilities or processes within men and women without confounding factors. We are left with the age-old question: who is better at navigating? Which of our paradigms, some of which seem closer to real-life situations (e.g., virtual town wayfinding task), would best describe navigation ability differences between men and women in standard daily living? We would argue that, as in the experiments we described, the emergence of sex differences depends on the situation. It is true that a lot of environments are open, such as parks and woods, but it is also true that many are restricted: a city whose streets are grid-like restricts navigation and resembles tasks with pathways. Similarly, finding one’s way in a high-rise building implies going along series of corridors that restrict navigation. On the other hand, cities with curving streets do not restrict navigation as much and allow us to take more shortcuts, and therefore they more greatly resemble open environments. For these reasons, we argue that men and women are equally as good at navigating, but that the particular environmental features that they pay attention to differ. Spiers et al. (2021) found that individuals that reported growing up in a city were on average worse at navigating than those who grew up outside cities. Therefore, navigation experience growing up has an impact on spatial performance and may help identify those at greater risk of Alzheimer’s disease Spiers et al. (2021).
Data availability statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Douglas Mental Health McGill University. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
LD: testing participants, statistic calculation, writing, figures, and tables. MI: editing. KK: editing. VB: supervisor and editor. GW: supervisor and editor. All authors contributed to the article and approved the submitted version.
Acknowledgments
Maris Kalnins, Oliver Hardt, Kate Conrad, Rosalind Sham, Kathleen MacDonald, Katarina Stepankova. CFI #9357 CIHR #64381, #82638, #86727, #GSD-121804 NSERC #239896, #CGSM-409063-2011 FRSQ: #2828 McDonnell Foundation #97-34EE, #98-38 CNS-QUA Grant Agency of the Czech Republic #309/02/1218/A.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aguilar-Latorre, A., Romera-Nicolás, V., Gimeno, E., and Chamizo, V. D. (2022). Previous experience seems crucial to eliminate the sex gap in geometry learning when solving a navigation task in rats (rattus norvegicus). Front. Psychol. 13, 838407. doi:10.3389/fpsyg.2022.838407
Andersen, N. E., Dahmani, L., Konishi, K., and Bohbot, V. D. (2012). Eye tracking, strategies, and sex differences in virtual navigation. Neurobiol. Learn Mem. 97 (1), 81–89. doi:10.1016/j.nlm.2011.09.007
Astur, R. S., Ortiz, M. L., and Sutherland, R. J. (1998). A characterization of performance by men and women in a virtual Morris water task. Behav. Brain Res. 93 (1-2), 185–190. doi:10.1016/s0166-4328(98)00019-9
Astur, R. S., Tropp, J., Sava, S., Constable, R. T., and Markus, E. J. (2004). Sex differences and correlations in a virtual Morris water task, a virtual radial arm maze, and mental rotation. Behav. Brain Res. 151 (1-2), 103–115. doi:10.1016/j.bbr.2003.08.024
Banner, H., Bhat, V., Etchamendy, N., Joober, R., and Bohbot, V. D. (2011). The brain-derived neurotrophic factor Val66Met polymorphism is associated with reduced functional magnetic resonance imaging activity in the hippocampus and increased use of caudate nucleus-dependent strategies in a human virtual navigation task. Eur. J. Neurosci. 33 (5), 968–977. doi:10.1111/j.1460-9568.2010.07550.x
Barkley, C. L., and Gabriel, K. I. (2007). Sex differences in cue perception in a visual scene: investigation of cue type. Behav. Neurosci. 121 (2), 291–300. doi:10.1037/0735-7044.121.2.291
Barnes, L. L., Wilson, R. S., Bienias, J. L., Schneider, J. A., Evans, D. A., and Bennett, D. A. (2005). Sex differences in the clinical manifestations of alzheimer disease pathology. Archives General Psychiatry 62 (6), 685. doi:10.1001/archpsyc.62.6.685
Bohbot, V. D., Copara, M. S., Gotman, J., and Ekstrom, A. D. (2017). Low-frequency theta oscillations in the human hippocampus during real-world and Virtual Navigation. Nat. Commun. 8 (1), 14415. doi:10.1038/ncomms14415
Bohbot, V. D., Iaria, G., and Petrides, M. (2004). Hippocampal function and spatial memory: evidence from functional neuroimaging in healthy participants and performance of patients with medial temporal lobe resections. Neuropsychology 18 (3), 418–425. doi:10.1037/0894-4105.18.3.418
Bohbot, V. D., Jech, R., Bures, J., Nadel, L., and Ruzicka, E. (1997). Spatial and nonspatial memory involvement in myasthenia gravis. J. Neurol. 244 (8), 529–532. doi:10.1007/s004150050139
Bohbot, V. D., Kalina, M., Stepankova, K., Spackova, N., Petrides, M., and Nadel, L. (1998). Spatial memory deficits in patients with lesions to the right hippocampus and to the right parahippocampal cortex. Neuropsychologia 36 (11), 1217–1238. doi:10.1016/s0028-3932(97)00161-9
Bohbot, V. D., Lerch, J., Thorndycraft, B., Iaria, G., and Zijdenbos, A. P. (2007). Gray matter differences correlate with spontaneous strategies in a human virtual navigation task. J. Neurosci. 27 (38), 10078–10083. doi:10.1523/jneurosci.1763-07.2007
Brunec, I. K., Nantais, M. M., Sutton, J. E., Epstein, R. A., and Newcombe, N. S. (2023). Exploration patterns shape cognitive map learning. Cognition 233, 105360. doi:10.1016/j.cognition.2022.105360
Caffò, A. O., Lopez, A., Spano, G., Serino, S., Cipresso, P., Stasolla, F., et al. (2017). Spatial reorientation decline in aging: the combination of geometry and landmarks. Aging & Ment. Health 22 (10), 1372–1383. doi:10.1080/13607863.2017.1354973
Castelli, L., Latini Corazzini, L., and Geminiani, G. C. (2008). Spatial navigation in large-scale virtual environments: gender differences in survey tasks. Comput. Hum. Behav. 24 (4), 1643–1667. doi:10.1016/j.chb.2007.06.005
Chai, X. J., and Jacobs, L. F. (2009). Sex differences in directional cue use in a virtual landscape. Behav. Neurosci. 123 (2), 276–283. doi:10.1037/a0014722
Chamizo, V., Artigas, A., Sansa, J., and Banterla, F. (2011). Gender differences in landmark learning for virtual navigation: the role of distance to a goal. Behav. Process. 88 (1), 20–26. doi:10.1016/j.beproc.2011.06.007
Chen, C.-H., Chang, W.-C., and Chang, W.-T. (2009). Gender differences in relation to wayfinding strategies, navigational support design, and wayfinding task difficulty. J. Environ. Psychol. 29 (2), 220–226. doi:10.1016/j.jenvp.2008.07.003
Chrastil, E. R., and Warren, W. H. (2014). From cognitive maps to cognitive graphs. PLoS ONE 9 (11), e112544. doi:10.1371/journal.pone.0112544
Dabbs, J. M., Chang, E.-L., Strong, R. A., and Milun, R. (1998). Spatial ability, navigation strategy, and geographic knowledge among men and women. Evol. Hum. Behav. 19 (2), 89–98. doi:10.1016/s1090-5138(97)00107-4
Dahmani, L., and Bohbot, V. D. (2014). Dissociable contributions of the prefrontal cortex to hippocampus-and caudate nucleus-dependent virtual navigation strategies. Neurobiol. Learn. Mem. 117, 42–50. doi:10.1016/j.nlm.2014.07.002
Dahmani, L., Ledoux, A. A., Boyer, P., and Bohbot, V. D. (2012). Wayfinding: the effects of large displays and 3-D perception. Behav. Res. Methods 44 (2), 447–454. doi:10.3758/s13428-011-0158-9
den Heijer, T., Geerlings, M. I., Hoebeek, F. E., Hofman, A., Koudstaal, P. J., and Breteler, M. M. (2006). Use of hippocampal and amygdalar volumes on magnetic resonance imaging to predict dementia in cognitively intact elderly people. Arch. Gen. Psychiatry 63 (1), 57–62. doi:10.1001/archpsyc.63.1.57
Driscoll, I., Hamilton, D. A., Yeo, R. A., Brooks, W. M., and Sutherland, R. J. (2005). Virtual navigation in humans: the impact of age, sex, and hormones on place learning. Hormones Behav. 47 (3), 326–335. doi:10.1016/j.yhbeh.2004.11.013
Du, A. T., Schuff, N., Amend, D., Laakso, M. P., Hsu, Y. Y., Yagust, W. J., et al. (2001). Magnetic resonanceimaging of the entorhinal cortex and hippocampus in mild cognitive impairment and alzheimer's disease. J. Neurology, Neurosurg. Psychiatry 71 (4), 441–447. doi:10.1136/jnnp.71.4.441
Etchamendy, N., and Bohbot, V. D. (2007). Spontaneous navigational strategies and performance in the virtual town. Hippocampus 17 (8), 595–599. doi:10.1002/hipo.20303
Etchamendy, N., Konishi, K., Pike, G. B., Marighetto, A., and Bohbot, V. D. (2012). Evidence for a virtual human analog of a rodent relational memory task: A study of aging and fMRI in young adults. Hippocampus 22 (4), 869–880. doi:10.1002/hipo.20948
Fox, N. C., Warrington, E. K., Freeborough, P. A., Hartikainen, P., Kennedy, A. M., Stevens, J. M., et al. (1996). Presymptomatic hippocampal atrophy in Alzheimer's disease. A longitudinal MRI study. Brain 119 (Pt 6), 2001–2007. doi:10.1093/brain/119.6.2001
Gamberini, L. (2000). Virtual reality as a new research tool for the study of human memory. CyberPsychology Behav. 3 (3), 337–342. doi:10.1089/10949310050078779
Habib, M., and Sirigu, A. (1987). Pure topographical disorientation: A definition and anatomical basis. Cortex 23 (1), 73–85. doi:10.1016/s0010-9452(87)80020-5
Hartley, T., Maguire, E. A., Spiers, H. J., and Burgess, N. (2003). The well-worn route and the path less traveled: distinct neural bases of route following and wayfinding in humans. Neuron 37 (5), 877–888. doi:10.1016/s0896-6273(03)00095-3
Head, D., and Isom, M. (2010). Age effects on wayfinding and route learning skills. Behav. Brain Res. 209 (1), 49–58. doi:10.1016/j.bbr.2010.01.012
Henderson, V. W. (1994). Estrogen replacement therapy in older women. Archives Neurology 51 (9), 896. doi:10.1001/archneur.1994.00540210068014
Holding, C. S., and Holding, D. H. (1988). Acquisition of route network knowledge by males and females. J. General Psychol. 116, 29–41. doi:10.1080/00221309.1989.9711108
Holdstock, J. S., Mayes, A. R., Cezayirli, E., Isaac, C. L., Aggleton, J. P., and Roberts, N. (2000). A comparison of egocentric and allocentric spatial memory in a patient with selective hippocampal damage. Neuropsychologia 38 (4), 410–425. doi:10.1016/s0028-3932(99)00099-8
Hussain, D., Hanafi, S., Konishi, K., Brake, W. G., and Bohbot, V. D. (2016). Modulation of spatial and response strategies by phase of the menstrual cycle in women tested in a virtual navigation task. Psychoneuroendocrinology 70, 108–117doi. doi:10.1016/j.psyneuen.2016.05.008
Iachini, T., Sergi, I., Ruggiero, G., and Gnisci, A. (2005). Gender differences in object location memory in a real three-dimensional environment. Brain Cognition 59 (1), 52–59. doi:10.1016/j.bandc.2005.04.004
Iaria, G., Lanyon, L. J., Fox, C. J., Giaschi, D., and Barton, J. J. (2008). Navigational skills correlate with hippocampal fractional anisotropy in humans. Hippocampus 18 (4), 335–339. doi:10.1002/hipo.20400
Iaria, G., Petrides, M., Dagher, A., Pike, B., and Bohbot, V. D. (2003). Cognitive strategies dependent on the hippocampus and caudate nucleus in human navigation: variability and change with practice. J. Neurosci. 23 (13), 5945–5952. doi:10.1523/jneurosci.23-13-05945.2003
Jagust, W., Gitcho, A., Sun, F., Kuczynski, B., Mungas, D., and Haan, M. (2006). Brain imaging evidence of preclinical Alzheimer's disease in normal aging. Ann. Neurol. 59 (4), 673–681. doi:10.1002/ana.20799
Kaye, J. A., Swihart, T., Howieson, D., Dame, A., Moore, M. M., Karnos, T., et al. (1997). Volume loss of the hippocampus and temporal lobe in healthy elderly persons destined to develop dementia. Neurology 48 (5), 1297–1304. doi:10.1212/wnl.48.5.1297
Konishi, K., and Bohbot, V. D. (2013). Spatial navigational strategies correlate with gray matter in the hippocampus of healthy older adults tested in a virtual maze. Front. Aging Neurosci. 5, 1. doi:10.3389/fnagi.2013.00001
Konishi, K., Etchamendy, N., Roy, S., Marighetto, A., Rajah, N., and Bohbot, V. D. (2013). Decreased functional magnetic resonance imaging activity in the hippocampus in favor of the caudate nucleus in older adults tested in a virtual navigation task. Hippocampus 23 (11), 1005–1014. doi:10.1002/hipo.22181
Lawton, C. A., and Morrin, K. A. (1999). Gender differences in pointing accuracy in computer-simulated 3D mazes. Sex. roles 40 (1-2), 73–92. doi:10.1023/a:1018830401088
Ledoux, A. A., Phillips, J. L., Labelle, A., Smith, A., Bohbot, V. D., and Boyer, P. (2013). Decreased fMRI activity in the hippocampus of patients with schizophrenia compared to healthy control participants, tested on a wayfinding task in a virtual town. Psychiatry Res. 211 (1), 47–56. doi:10.1016/j.pscychresns.2012.10.005
Levy, L. J., Astur, R. S., and Frick, K. M. (2005). Men and women differ in object memory but not performance of a virtual radial maze. Behav. Neurosci. 119 (4), 853–862. doi:10.1037/0735-7044.119.4.853
Lopez, A., and Bosco, A. (2022). Distortions in spatial mental representation affect sketch maps in Young Adults. Neurol. Int. 14 (4), 771–783. doi:10.3390/neurolint14040064
Lopez, A., Caffò, A. O., Tinella, L., Postma, A., and Bosco, A. (2020a). Studying individual differences in spatial cognition through differential item functioning analysis. Brain Sci. 10 (11), 774. doi:10.3390/brainsci10110774
Lopez, A., Postma, A., and Bosco, A. (2020b). Categorical & coordinate spatial information: can they be disentangled in sketch maps? J. Environ. Psychol. 68, 101392. doi:10.1016/j.jenvp.2020.101392
Maguire, E. A., Burgess, N., and O'Keefe, J. (1999). Human spatial navigation: cognitive maps, sexual dimorphism, and neural substrates. Curr. Opin. Neurobiol. 9 (2), 171–177. doi:10.1016/s0959-4388(99)80023-3
Maguire, E. A., and Cipolotti, L. (1998). Selective sparing of topographical memory. J. Neurol. Neurosurg. Psychiatry 65 (6), 903–909. doi:10.1136/jnnp.65.6.903
Marighetto, A., Etchamendy, N., Touzani, K., Torrea, C. C., Yee, B. K., Rawlins, J. N. P., et al. (1999). Knowing which and knowing what: A potential mouse model for age-related human declarative memory decline. Eur. J. Neurosci. 11 (9), 3312–3322. doi:10.1046/j.1460-9568.1999.00741.x
McCarthy, R. A., Evans, J. J., and Hodges, J. R. (1996). Topographic amnesia: spatial memory disorder, perceptual dysfunction, or category specific semantic memory impairment? J. Neurol. Neurosurg. Psychiatry 60 (3), 318–325. doi:10.1136/jnnp.60.3.318
Mielke, M. M., Okonkwo, O. C., Oishi, K., Mori, S., Tighe, S., Miller, M. I., et al. (2012). Fornix integrity and hippocampal volume predict memory decline and progression to alzheimer's disease. Alzheimer's Dementia 8 (2), 105–113. doi:10.1016/j.jalz.2011.05.2416
Moffat, S. D., Kennedy, K. M., Rodrigue, K. M., and Raz, N. (2007). Extrahippocampal contributions to age differences in human spatial navigation. Cereb. Cortex 17 (6), 1274–1282. doi:10.1093/cercor/bhl036
Montello, D. R. (2017). Landmarks are exaggerated. KI - Künstliche Intell. 31 (2), 193–197. doi:10.1007/s13218-016-0473-5
Montello, D. R., Lovelace, K. L., Golledge, R. G., and Self, C. M. (1999). Sex-related differences and similarities in geographic and environmental spatial abilities. Ann. Assoc. Am. Geogr. 89 (3), 515–534. doi:10.1111/0004-5608.00160
Morris, R. G. (1981). Spatial localization does not require the presence of local cues. Learn. motivation 12 (2), 239–260. doi:10.1016/0023-9690(81)90020-5
Mueller, S. C., Jackson, C., and Skelton, R. W. (2008). Sex differences in a virtual water maze: an eye tracking and pupillometry study. Behav. Brain Res. 193 (2), 209–215. doi:10.1016/j.bbr.2008.05.017
Nadel, L., and Hardt, O. (2004). The spatial brain. Neuropsychology 18 (3), 473–476. doi:10.1037/0894-4105.18.3.473
Newhouse, P., Newhouse, C., and Astur, R. S. (2007). Sex differences in visual-spatial learning using a virtual water maze in pre-pubertal children. Behav. Brain Res. 183 (1), 1–7. doi:10.1016/j.bbr.2007.05.011
Nowak, N. T., and Moffat, S. D. (2011). The relationship between second to fourth digit ratio, spatial cognition, and virtual navigation. Archives Sex. Behav. 40 (3), 575–585. doi:10.1007/s10508-010-9668-2
Olton, D. S., and Samuelson, R. J. (1976). Remembrance of places passed: spatial memory in rats. J. Exp. Psychol. Animal Behav. Process. 2 (2), 97–116. doi:10.1037/0097-7403.2.2.97
Overman, W. H., Pate, B. J., Moore, K., and Peuster, A. (1996). Ontogeny of place learning in children as measured in the radial arm maze, Morris search task, and open field task. Behav. Neurosci. 110 (6), 1205–1228. doi:10.1037/0735-7044.110.6.1205
Packard, M. G., Hirsh, R., and White, N. M. (1989). Differential effects of fornix and caudate nucleus lesions on two radial maze tasks: evidence for multiple memory systems. J. Neurosci. 9 (5), 1465–1472. doi:10.1523/jneurosci.09-05-01465.1989
Piccardi, L., Bianchini, F., Iasevoli, L., Giannone, G., and Guariglia, C. (2011). Sex differences in a landmark environmental re-orientation task only during the learning phase. Neurosci. Lett. 503 (3), 181–185. doi:10.1016/j.neulet.2011.08.031
Postma, A., Izendoorn, R., and De Haan, E. H. F. (1998). Sex differences in object location memory. Brain Cognition 36 (3), 334–345. doi:10.1006/brcg.1997.0974
Postma, A., Jager, G., Kessels, R. P., Koppeschaar, H. P., and van Honk, J. (2004). Sex differences for selective forms of spatial memory. Brain Cogn. 54 (1), 24–34. doi:10.1016/s0278-2626(03)00238-0
Pruessner, J., Collins, D., Pruessner, M., and Evans, A. (2001). Age and gender predict volume decline in the anterior and posterior hippocampus in early adulthood. J. Neurosci. 21 (1), 194–200. doi:10.1523/jneurosci.21-01-00194.2001
Reiman, E. M., Uecker, A., Caselli, R. J., Lewis, S., Bandy, D., De Leon, M. J., et al. (1998). Hippocampal volumes in cognitively normal persons at genetic risk for alzheimer's disease. Ann. Neurology 44 (2), 288–291. doi:10.1002/ana.410440226
Rizk-Jackson, A. M., Acevedo, S. F., Inman, D., Howieson, D., Benice, T. S., and Raber, J. (2006). Effects of sex on object recognition and spatial navigation in humans. Behav. Brain Res. 173 (2), 181–190. doi:10.1016/j.bbr.2006.06.029
Roof, R., and Stein, D. (1999). Gender differences in Morris water maze performance depend on task parameters. Physiology Behav. 68 (1-2), 81–86. doi:10.1016/s0031-9384(99)00162-6
Ruggiero, G., Sergi, I., and Iachini, T. (2008). Gender differences in remembering and inferring spatial distances. Memory 16 (8), 821–835. doi:10.1080/09658210802307695
Sandstrom, N. J., Kaufman, J., and Huettel, S. A. (1998). Males and females use different distal cues in a virtual environment navigation task. Brain Res. Cogn. Brain Res. 6 (4), 351–360. doi:10.1016/s0926-6410(98)00002-0
Saucier, D., Lisoway, A., Green, S., and Elias, L. (2007). Female advantage for object location memory in peripersonal but not extrapersonal space. J. Int. Neuropsychol. Soc. 13 (4), 683–686. doi:10.1017/S1355617707070865
Saucier, D. M., Green, S. M., Leason, J., MacFadden, A., Bell, S., and Elias, L. J. (2002). Are sex differences in navigation caused by sexually dimorphic strategies or by differences in the ability to use the strategies? Behav. Neurosci. 116 (3), 403–410. doi:10.1037/0735-7044.116.3.403
Silverman, I., and Eals, M. (1992). Sex differences in spatial abilities: evolutionary theory and data. Portions of this paper were presented at the meetings of the International Society for Human Ethology in Binghamton, NY, Jun 1990, the Human Behavior and Evolution Society in Los Angeles, CA, Aug 1990, and the European Sociobiological Society in Prague, Czechoslovakia, Aug 1991.
Sodums, D. J., and Bohbot, V. D. (2020). Negative correlation between grey matter in the hippocampus and caudate nucleus in healthy aging. Hippocampus 30 (8), 892–908. doi:10.1002/hipo.23210
Spiers, H. J., Burgess, N., Hartley, T., Vargha-Khadem, F., and O'Keefe, J. (2001). Bilateral hippocampal pathology impairs topographical and episodic memory but not visual pattern matching. Hippocampus 11 (6), 715–725. doi:10.1002/hipo.1087
Spiers, H. J., Coutrot, A., and Hornberger, M. (2021). Explaining world-wide variation in navigation ability from millions of people: citizen science project sea hero quest. Top. Cognitive Sci. 15 (1), 120–138. doi:10.1111/tops.12590
Stepankova, K., Fenton, A. A., Pastalkova, E., Kalina, M., and Bohbot, V. D. (2004). Object–location memory impairment in patients with thermal lesions to the right or left hippocampus. Neuropsychologia 42 (8), 1017–1028. doi:10.1016/j.neuropsychologia.2004.01.002
Tolman, E. C. (1948). Cognitive maps in rats and men. Psychol. Rev. 55 (4), 189–208. doi:10.1037/h0061626
van Gerven, D. J., Schneider, A. N., Wuitchik, D. M., and Skelton, R. W. (2012). Direct measurement of spontaneous strategy selection in a virtual morris water maze shows females choose an allocentric strategy at least as often as males do. Behav. Neurosci. 126 (3), 465–478. doi:10.1037/a0027992
Voyer, D., Postma, A., Brake, B., and Imperato-McGinley, J. (2007). Gender differences in object location memory: A meta-analysis. Psychon. Bull. Rev. 14 (1), 23–38. doi:10.3758/bf03194024
Wertlieb, D., and Rose, D. (1979). Maturation of maze behavior in preschool children. Dev. Psychol. 15 (4), 478–479. doi:10.1037/h0078085
West, G. L., Konishi, K., Diarra, M., Benady-Chorney, J., Drisdelle, B. L., Dahmani, L., et al. (2018). Impact of video games on plasticity of the hippocampus. Mol. Psychiatry 23, 1566–1574. doi:10.1038/mp.2017.155
White, N. M., and McDonald, R. J. (2002). Multiple parallel memory systems in the brain of the rat. Neurobiol. Learn. Mem. 77 (2), 125–184. doi:10.1006/nlme.2001.4008
Wolbers, T., and Hegarty, M. (2010). What determines our navigational abilities? Trends Cogn. Sci. 14 (3), 138–146. doi:10.1016/j.tics.2010.01.001
Woolley, D. G., Vermaercke, B., Op de Beeck, H., Wagemans, J., Gantois, I., D'Hooge, R., et al. (2010). Sex differences in human virtual water maze performance: novel measures reveal the relative contribution of directional responding and spatial knowledge. Behav. Brain Res. 208 (2), 408–414. doi:10.1016/j.bbr.2009.12.019
Keywords: hippocampus, aging, navigation, sex, navigation strategies
Citation: Dahmani L, Idriss M, Konishi K, West GL and Bohbot VD (2023) Sex differences in spatial tasks: Considering environmental factors, navigation strategies, and age. Front. Virtual Real. 4:1166364. doi: 10.3389/frvir.2023.1166364
Received: 15 February 2023; Accepted: 21 August 2023;
Published: 04 September 2023.
Edited by:
Pierre Bourdin, Open University of Catalonia, SpainReviewed by:
Chuanxiuyue He, Rutgers, The State University of New Jersey, United StatesAndrea Bosco, University of Bari Aldo Moro, Italy
Copyright © 2023 Dahmani, Idriss, Konishi, West and Bohbot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Veronique D. Bohbot, dmVyb25pcXVlLmJvaGJvdEBtY2dpbGwuY2E=
†Present address: Louisa Dahmani, Department of Psychiatry, Massachusetts General Hospital and Harvard Medical School, Cambridge, MA, United States; Kyoko Konish, Department of Psychiatry, Massachusetts General Hospital and Harvard Medical School, Cambridge, MA, United States