
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Virol. , 02 December 2024
Sec. Emerging and Reemerging Viruses
Volume 4 - 2024 | https://doi.org/10.3389/fviro.2024.1448192
 Sara Thabet1,2*
Sara Thabet1,2* Rim Lajnef3
Rim Lajnef3Bluetongue is a non-zoonotic arboviral disease transmitted by biting midges of the genus Culicoides. Among 1,347 Culicoides species, 30 are known to be vectors of the disease. Bluetongue affects domestic and wild ruminants across all continents, except the Antarctica, causing growing economic losses in livestock production and international trade. In this work, we focused on potential mechanisms underlying the bluetongue virus emergence and spread. Of these mechanisms, the emergence of novel serotypes makes vaccination controversial despite its effectiveness, along with the vector’s ability to acquire the virus and successfully transmit it, spatio-temporal adaptation, and identification of new transmission routes. We also discussed the predisposition of the vertebrate and invertebrate hosts to contract the virus, as well as factors hijacking vector population activity and abundance. Indeed, the ecological dynamics of biting midges strongly influence the ability of the virus to be transmitted and to cause epizootic into parts of the world never before affected, helped by interactions between the pathogen and its host in addition to multiple routes deployed to escape defense barriers in the target infected cells. The points raised in this review can help inform aspects of interest for future studies that may elucidate factors to consider in monitoring and controlling the expansion of Culicoides-transmitted arboviruses.
Bluetongue virus (BTV) is an arthropod-vectored emerging pathogen of wild ruminants and livestock endemic in many parts of the world. BTV causes notifiable severe hemorrhagic disease mainly in sheep, some species of deer, and pronghorn antelope (1–3) as reported by the zoo-sanitary code of the World Organisation for Animal Health (WOAH) (4–7). In fact, bluetongue (BT) is considered potentially debilitating, infectious but a noncontagious disease in ruminants (8) with clinical signs that differ widely between species. Infected cattle rarely exhibit clinical manifestations but constitute an important reservoir for epidemiological studies because they have longer viremia than observed in sheep (9). In goat and camelid, the infection usually goes unnoticed with subclinical manifestation. Recovery is spontaneous and rapid in most cases. Whereas, in sheep, the disease produces severe physical and cellular damage ranging from fever, edema, excessive salivation, anorexia, nasal discharge, lameness, coronitis, and swelling of the head to death in extreme cases. Since the 2000s, high morbidity has been recorded with the incursion of several virulent viral strains in sheep and cattle leading to growing economic impact in livestock production (10). Mortality in sheep ranges from 5% to 30% and sometimes reaches 70%, which can cause serious socio-economic and trade restrictions at local and global levels (11–13).
BTV is the prototype member of genus Orbivirus within the family Sedoreoviridae (5, 10, 14, 15) transmitted by competent adult female Culicoides midges. The virus particle is icosahedral and is made of an outer capsid (consisting of VP2 and VP5 proteins) and a double-layered core (VP3 forms the inner layer of the core and VP7 forms the outer layer of the core). Five additional non-structural proteins, NS1, 2, 3/3a, 4, and 5 (16–18) are produced in virus-infected cells (19) and are involved in BTV release from mammalian and insect cells via interactions between viral and host cell proteins (20).
BTV genome is known to undergo high frequency of genetic polymorphism and reassortment events responsible for the emergence of new serotypes or antigenic variants of the same serotype (20). BTV as a species is identified by the group-specific antigen, that is the VP7 protein, while the serotype is determined by the VP2 protein (19). Reassortment involving genome segment 2 will cause an antigenic shift thus changing the serotype of the particle. However, vaccination remains the most adequate response to protect the animals and control virus spread, yet a vaccine-matching strategy should be implemented. BT continues to be debilitating with wide virus propagation in both ruminant and insect cells inducing considerable socioeconomic concern. In addition, incursion of novel serotypes in enzootic regions as well as combination of genetic drift and genomic reassortment offer increasing pathways for the virus to bypass and evade the defense barriers of the host (19). Hence, a broad-spectrum vaccination strategy based on the use of cross-protective vaccine subunits such as the NS1 would confer a cross-serotype protection.
The aim of this paper is to discuss potential mechanisms underlying bluetongue virus emergence and epidemiology with an overview of disease outbreaks prediction criteria.
Sanitary protection measures (21) are based on two axes:
● The control of animal movements from infected countries by isolating or even slaughtering sick and infected animals and destroying cadavers in endemic areas.
● Vector control through regular insecticide treatments.
These measures can be reinforced by serological surveys in sentinel farms to monitor the possible spread of the disease. Trapping of Culicoides helps define areas of vector activity.
As a medical approach, vaccination remains the best way to protect against bluetongue, although controversial due to the emergence of new serotypes, given the difficulty of protecting oneself from insect bites. Different types of vaccines are available, live attenuated vaccines, inactivated virus vaccines, and new generation vaccines. In non-African breeds of sheep, live attenuated vaccines cause severe clinical signs and hence, are not licensed for use in several countries.
BT was considered for a long time as an exotic disease limited to tropical and temperate sub-tropical regions (5, 12). It is believed to be that until the 1950s. The disease was first recognized in South Africa, the most probable origin of BT, in Merino sheep in the late 18th century (22). Alkhamis et al. (23) proposed that goats are the ancestral hosts for BTV. It is even thought to be endemic in wild ruminants in sub-Saharan Africa since antiquity. In 1906, Thelier et al. (24) demonstrated that BT was caused by a filterable pathogen. Serotype 4 is the first BTV serotype identified in South Africa. They also introduced the first attenuated vaccine against BTV-4 that has been used in farming sheep for over 40 years and that provides cross-protection to other serotypes (25). Outside of Africa, infectious outbreaks were reported in Cyprus in 1943 (26, 27). After that, BTV has spread subsequently to regions between the latitudes of 35°S and 40°N (1, 3, 25, 28–31). In addition to the African continent, these areas encompass Asia including the Indian subcontinent and China, Central and South America, Middle East, Israel, and Australia where BTV was isolated for the first time from Culicoides in the Northern Territory in 1975 (29–32). In the autumn of 1977, serotype 4 was isolated from female Culicoides obsoletus during a second epizootic of BT in Cyprus (27, 29). The European continent was considered safe from the disease with just a few sporadic incursions in the Iberian Peninsula (5, 33) until 1998. However, since the virus incursion into several Mediterranean basin countries in 1998 with the arrival of BTV-9 in Greece, the emergence of new serotypes in disease-free areas as naïve populations, as well as in endemic regions has modified the epizootic situation generating massive losses for the livestock industry through reduced productivity and death affecting herds particularly in farmed sheep. Since then and for a few years, at least six BTV serotypes (BTV-1, BTV-2, BTV-4, BTV-8, BTV-9, and BTV-16) have been continuously present in parts of the Mediterranean Basin (34).
Explosive unprecedented outbreak of BT disease has been reported in 2006 in Northern and Central Europe with the arrival of BTV-8 in the Netherlands and the disease spread to neighboring countries. During the hot seasons of 2006 and 2007, the situation was alarming with the outbreak of a devastating epidemic (5), which triggered Europe-wide monitoring projects on the dynamics of biting midges of the genus Culicoides (35) as well as intensive vaccination campaigns against BTV-8. The propagation of BTV-8 involved two previously unknown to be effective BTV vectors, Culicoides obsoletus and C. pulicaris (12, 36).
The first occurrence of BT in Tunisia dates to 1999. BTV-2, BTV-1, and BTV-4 have been reported in 2000, 2006, and 2009 respectively (37). Since then, active circulation of Mediterranean BTV strains has been noted.
Since the onset of the disease until 2008, 24 classical serotypes of BTV (BTV-1, BTV-24), replicating in vertebrates and Culicoides, were officially recognized according to cross neutralization assays (38, 39). Live attenuated vaccine strains, originally developed in South Africa were the origin of BTV-6 and BTV-11 strain outbreaks in Northern Europe between late 2008 and early 2009 (40). Other serotypes have been proven to come from reassortments; a field reassortant between BTV-1 and BTV-8 named FRA2008/24 had been described in France (41). Reassortment was also detected in Culicoides sonorensis being co-infected with BTV-10 and BTV17 (42).
Due to the movement of infected ruminants, the abundance and wind dispersal of infected midges and the changes in climate, novel BTV strains have appeared with the possibility of reassortment between different serotypes (32). Genome comparison, conventional serotyping completed by evolutionary divergence data, allows determining the source of the infection and its spread in different areas. At present, the disease is present all over the inhabited world except Antarctica (43). BT is a notifiable disease and while the WOAH manual cites BTV-1 to BTV-27, at least 12 ‘atypical’ serotypes have been recognized so far, bringing the total number of published BTV-serotypes to 36 (44, 45). The serotype identification relies on the sequence of Seg-2/VP2. (3, 19, 46–48). Unlike classical serotypes, atypical ones appear to be unable to replicate in the arthropod vector suggesting direct transmission between vertebrate hosts (between infected and susceptible animals). Contamination occurs possibly via aerosols expelled into the air by hosts carrying the virus (49). To date, this mode of transmission is suspected for certain serotypes such as BTV-25 (50, 51), BTV-26 (12, 52, 53), BTV-27 (47), and BTV-28 (54). Although the recognized atypical serotypes have not caused severe bluetongue epidemics or economic losses to date, some of them have been shown to cause mild to moderate clinical signs in small ruminants exclusively (by experimentation or direct observation in the field) (55).
Ongoing epidemiological studies based on sequencing of complete or partial nucleotide genomic regions complemented by several phylogenetic analyses allow statistical estimation of genetic variability generated by RNA segments reassortment, recombination events, genetic drift, and mutation within serotypes. BTV genome replication is highly error-prone due to the lack of proof-reading mechanisms (56, 57). These biological events could lead to the emergence of more or less virulent strains by driving phenotypic change resulting in a fitness advantage for the virus (19, 41, 58). Intrasegment recombination is likely to occur between BTV strains either within vertebrate host (ruminant) or insect host (Culicoides) (19). Such information helps in the appreciation of the geographical spread, and subsequently, the emergence of novel putative BTV serotypes into enzootic or non-enzootic regions (59).
Apart from the presence of different serotypes, two major geographic groups of BTVs are described and designated as eastern (e) or western (w) topotypes even within the same serotype. The eastern topotype includes viruses from Australia and the Middle/Far East; the western viruses originate from Africa and the Americas (38, 39).
An interesting aspect of BTV infection is the variability of the host susceptibility. This is attributed to several factors including species of the infected subject, its age, and its immune system, as well as the infecting strain (30, 60). Indeed, different degrees of virulence have been described depending on the infecting serotype and resulting in different clinical manifestations. Variable severity of BTV infection comes from phenotypic and genotypic polymorphism related to high mutation rates in viral genome inducing the expression of more or less pathogenic character. For the same serotype, different pathogenicity levels would be observed according to its geographical origin. For example, the serotype 1 is reputed to be very aggressive in Spain and France displaying intense clinical signs and severe pathological lesions in sheep and goat flocks (61–63). Whereas, in Africa, Asia, and Indian subcontinent countries, the same serotype 1 is commonly associated with high pathogenic potential in sheep but no evidence for severity in goat has been reported. Neither clinical signs nor mortality were observed in cattle herds. It has also been reported that serotypes 1 and 3 from South Africa are more virulent than those from Australia. In Tunisia, infection with BTV-3 has resulted in high mortality rates in sheep (64). The European serotype 8 that triggered BT outbreak in 2006 has shown to be highly virulent with high mortality rates in naive sheep and many cases of severe clinical manifestations in cattle (25, 29, 63, 65). Other BTV strains (BTV-6, BTV-11, BTV-14) detected in Europe exhibited very little pathogenicity in farm animals. In enzootic areas, BTV serotypes 1, 2, 3, 4, 6, 8, 10, 11, and 24 are known to have a high epizootic potential mainly in sheep (22, 66) unlike serotypes 20 and 21 that would be clinically the most moderate forms. BTV-25 was initially identified in goats following cases of still births and BTV-26 was isolated from sheep showing BT clinical signs. BTV-28 caused clinical signs in experimentally infected sheep. Care should be taken not to bestow the non-pathogenic character onto all atypical BTV serotypes. Co-infection with two or more serotypes was reported in fields as well in experimental context implying biological consequences during viral replication within co-infected host cells (61). Moreover, Westrich et al. (45) established a model of BTV-17 infection in sheep including a longitudinal vertebrate host’s immunological response and viral infection progression using a combination of conventional methods and cutting-edge technology.
Upon feeding, the virus contained in the blood meals from infected vertebrate hosts is deposited in the mid-gut diverticulum. Viral particles attach to the surface of the gut cells and enter them by direct penetration or endocytosis (2). Culicoides cells are readily infected by core particles (usually non-infectious to mammalian cells). The core particle of an orbivirus is composed of an outer layer (VP7) and an inner layer (VP3) and this structure encloses the transcriptase complex (VP1, VP4, and VP6) together with the dsRNA genome (10). This core particle can bind to Culicoides cell surface and this is believed to be mediated by the interaction of VP7 with the RGD tripeptide of integrins (67, 68). Following replication within the midgut cells, virions are released from the basolateral membrane of the gut cells and migrate to sites of secondary replication. These include the salivary glands in which the virus replicates and is subsequently released into the salivary ducts. During subsequent blood meals, the infected female Culicoides injects the virus into the vertebrate host, by depositing it in the mammal's skin (to avoid recurrence). The viral particles travel through lymph to draining lymph nodes where the first replication in the ruminant takes place. The virus is then disseminated, via efferent lymphatic circulation, to secondary replication sites including lymphoid organs (lymph nodes and spleen) and lungs. Replication in these sites is followed by the release of BTV into the bloodstream. The VP2 is the cell attachment protein which mediates binding to the cell surface. Following endocytosis, acidification of the early endosome leads to the stepwise release of the VP2 and conformational changes of the VP5 (69). The VP5 then perforates the endosome, releasing the transcriptionally active core into the cell cytoplasm. Viral inclusion bodies (VIBs) are formed in the infected cell cytoplasm. The NS2 is the major component of VIBs and is the first viral protein that is detected in infected cells. It is a phosphoprotein able to bind ssRNA. These properties suggest that NS2 plays a significant role in genome assembly and packaging. BTV replication is accompanied by the synthesis of structural and non-structural viral proteins which are involved in the replication cycle, packaging, and assembly of novel viral particles but also play important roles in attenuating cellular responses (18, 70–72). In mammalian cells, the virus is released by budding during the early stages of infection (73), although virus egress progresses toward lytic release during the later stages of infection. In insect cells, the virus is released by budding, involving the NS3 viral protein (6, 10, 31). Studies suggested that NS3 forms viroporins, thus facilitating virus release (74).
The main source of BTV transmission is infected blood carried by Culicoides species. Only adult and sexually mature females, 28– 48 h post-hatching, feed on blood. Their longevity is estimated to be at 10–20 days with a maximum of 50. They become infected during a blood meal on a viremic animal and transmit the virus to other susceptible hosts during subsequent blood meals using their piercing mouthparts (75). The virus is not shed, so it is not found in saliva, waste, oral lesions. The level of exposure to infectious bites is the most influential parameter in BTV transmission depending on the period of infection. However, other modes of transmission are evoked such as via seminal fluid, oral, and iatrogenic routes, and vertically across the placenta in both cattle and sheep (7, 40) (Figure 1).
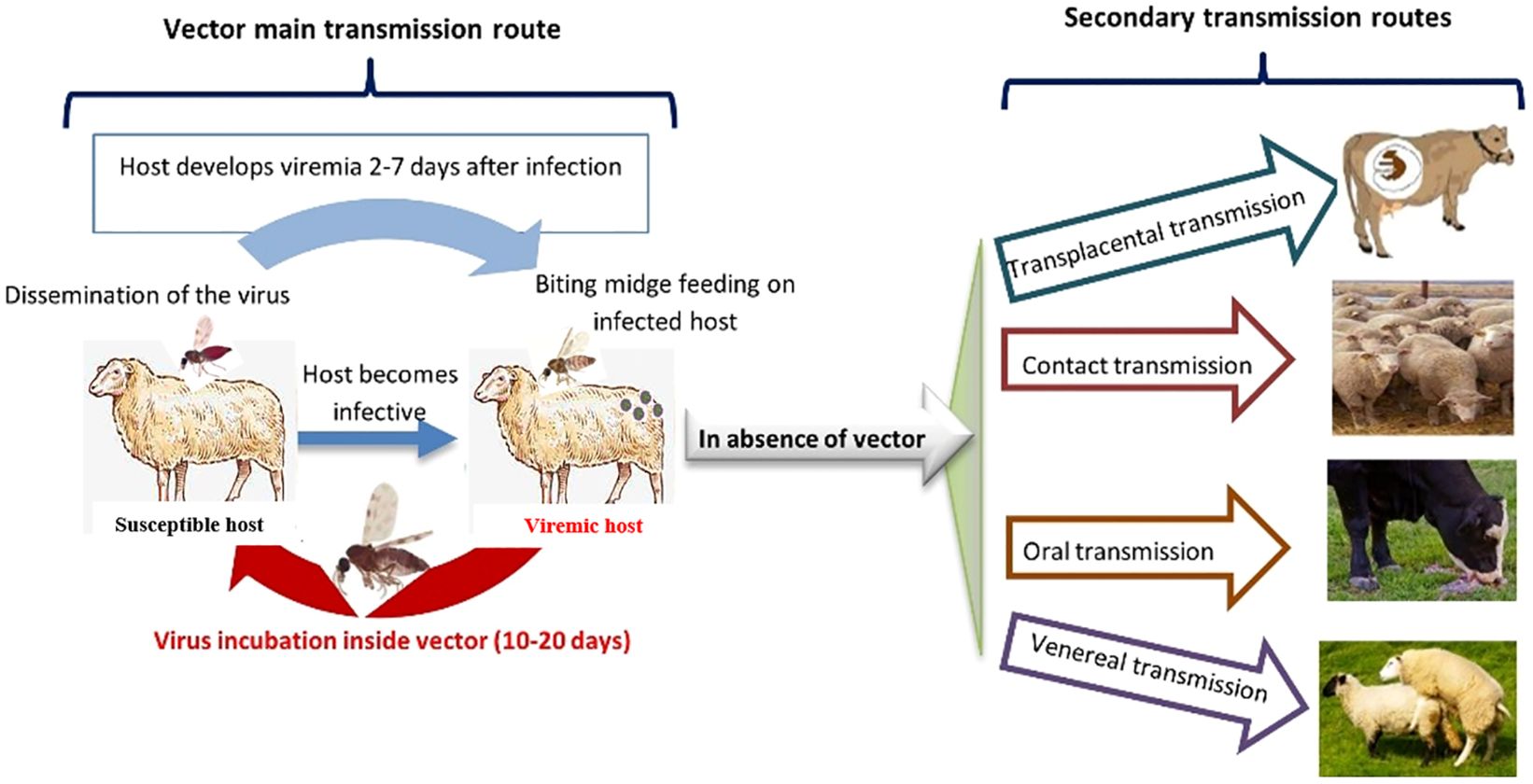
Figure 1. BTV transmission routes.
BTV transplacental transmission has been described firstly in sheep and later in cattle mainly with live attenuated BTV, European BTV-8, and other vaccine strains (3). Live attenuated vaccines used in the USA in the early 1950s caused severe degenerative central nervous system lesions, an increase in stillbirths, blind live lambs, and other congenital malformations in the areas where vaccines were used. Such notifications suggested causative relationship between BTV and signs seen in newborn lambs resulting from vertical transmission from vaccinated pregnant females to fetuses at different gestational stages (12, 19, 76). Vertical transmission of BTV from infected gestating heifers to their calves had been reported in various studies for several BTV serotypes (BTV-1, BTV‐2, BTV‐4, BTV‐8, BTV‐9, BTV‐10, BTV‐11, BTV‐13, BTV‐16, and BTV‐23) in cattle, sheep, goat, and elk (77). The ability of the BTV-8 strain to cross the placental barrier in cattle, sheep, and goats had been proven and vertical transmission could be quantified. Moreover, infection in early stage of fetal development may result in abortion or severe brain defects, while fetal infection in late stage of gestation could more likely result in the birth of viable RT-PCR positive calves (7). The BTV-8 epizootic in Europe was followed by an outbreak of hydranencephaly in aborted calves; spleen tissues of several of them were positive for BTV RNA (12). Nevertheless, estimates of vertical transmission may vary depending on the epidemiological context. Although vertical transmission has been proven, there is no evidence yet that infected calves could transmit BTV to Culicoides vectors nor how long they can keep antibodies. Moreover, vertical transmission is still considered a secondary route for BTV infection compared to transmission through vector bites. In the absence of an active vector transmission during the overwintering of the infection, BTV could be maintained in the host population through vertical transmission from an infected adult to their offspring resulting in the birth of healthy but BT-positive progeny. The virus could survive despite the absence of the vector due to adverse weather conditions (78).
Unusual features of BTV infection have been reported in the literature via horizontal transmission in the absence of the insect vector. Infection in a vector-free environment has been reported in cattle, goats, and sheep in experimental context as well as in field conditions for several serotypes such as BTV-1, BTV-2, BTV-8, and BTV-26 provided there is close contact between animals. However, it has been discussed that vector-independent transmission of BTV-1, BTV-2, or BTV-8 was only observed in gestating animals. Therefore, this observation is likely linked to the stage of gestation because no vector-independent transmission has been observed in non-gestating animals at least for BTV-8 or even BTV-4 (79).
Newly emerged atypical serotypes (BTV-25 - BTV-36) may be transmitted horizontally by direct contact between vertebrate hosts without the involvement of biting midges (19). In fact, over the past decade, atypical BTV strains have been shown to replicate efficiently only in mammalian cells, whereas their replication is severely restricted in Culicoides cells. BTV transmission is suggested to occur via aerosols expelled into the air by hosts carrying the virus (49), which needs to be explored more. To date, horizontal transmission is experimentally confirmed for some serotypes such as serotype 26 (52, 53), serotype 27 (47) and serotype 28 (54). For other serotypes, such as serotype 25, direct transmission is only suspected (50, 51). Although being an infrequent event, vector-independent transmission can induce an increase in morbidity rates, especially within farms (12, 80).
Transmission of BTV-8 by artificial insemination of contaminated frozen semen has been described (81). The infecting semen came from naturally infected bulls and was received by BT negative heifers causing their seroconversion with detectable BTV viremia, in most cases, displaying mild clinical signs of BT. The viral load transmitted may have significant impact on the infection rate in inseminated females and abortion may occur at early stages of gestation although other infected heifers gave birth at term to healthy calves.
BTV-8 infection of breeding females previously negative for BT was supported by the hypothesis of Pascall et al. (82) assuming that the re-emergence of BTV-8 in France in 2015 may be the result of the use of contaminated BTV bovine semen collected during the 2006-2010 epizootic (82). This feature of BTV-8 should not be extrapolated and the potential for other BTV serotypes to occur by artificial insemination needs to be more investigated.
Oral BTV transmission has been described in many cases in both domestic and wild ruminants. Infectious colostrum has been shown to be a source of infection for calves (19). Transmission of BTV-8 by ingestion of infected placenta has also been reported (76).
In the veterinary sector, sharing blood-infected needles between animals could result in mechanical transmission. In 2010, a study showed that some tick species experimentally fed with BTV-spiked blood became infected, suggesting that they may have ‘vectoring capacities’. However, the capacity of ticks to transmit the virus should be more deeply studied to clearly classify their role in the transmission cycle. Furthermore, BTV transmission by some mosquitoes and ticks has been discussed by Saminathan et al. (3).
Culicoides are among hematophagous insects that occur throughout most of the inhabited world (29) (Table 1). Species of the Culicoides genus, which act as vectors for BT are recognizable based on their morphological features (83). They are small diptera belonging to the order Diptera of the family Ceratopogonidae. The genus Culicoides includes 1,347 species of which 30 are known to be potential vectors of bluetongue. Adult Culicoides (imago) are 2–3-mm long. They have slender, elongated bodies with antennae consisting of 14 to 16 articles and short, often spotted, sparsely hairy wings (Figure 2) (83). As mentioned above, only adult females are hematophagous (Figure 3) (84), and therefore, able to transmit the virus to domestic and wild ruminants. Cyclically, viral particles adsorbed to the erythrocyte membrane are infectious and can thus infect any competent vector feeding on viremic animals.
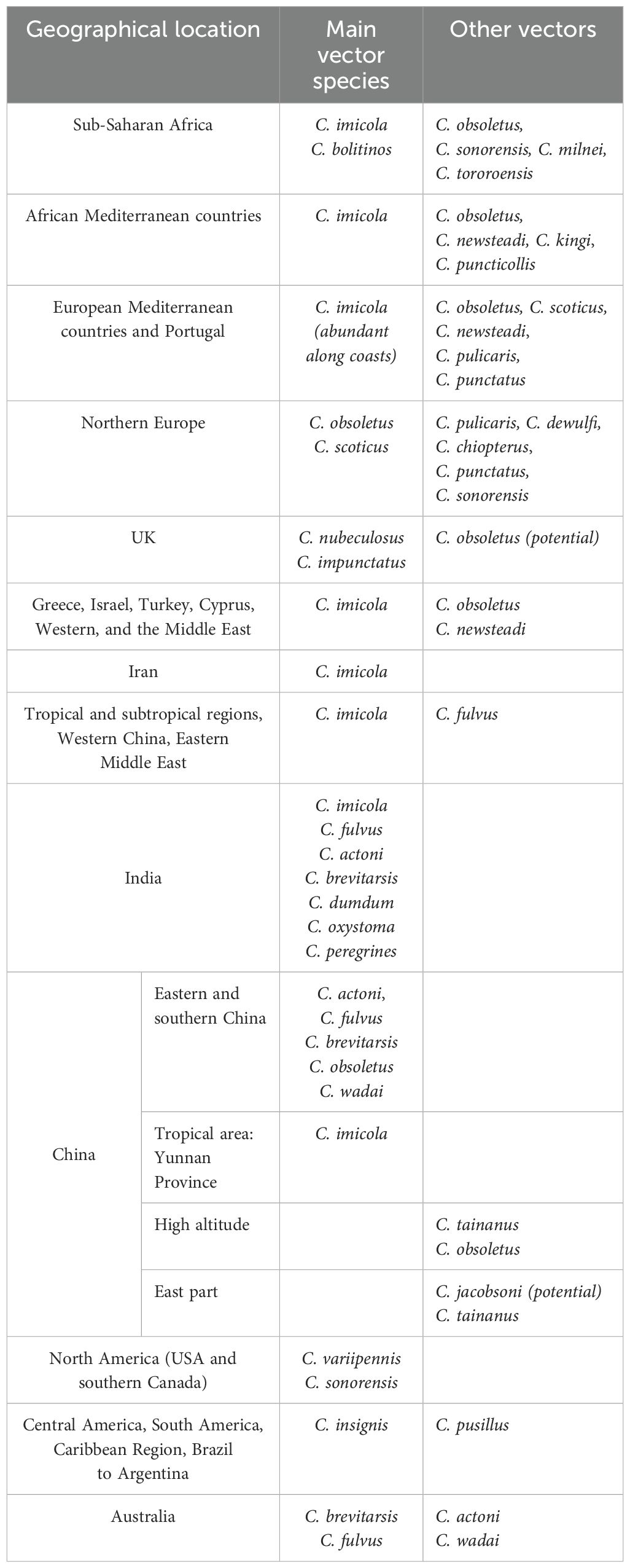
Table 1. Most predominant efficient vectors of the bluetongue virus in the inhabited world.

Figure 2. Female Culicoides sp imicola.

Figure 3. Engorged Culicoides sp obsoletus.
In Africa, most countries have reported the presence of C. imicola, as the main vector for BTV transmission (85). An Afro-Asiatic species, C. imicola, is recognizable by its size and patterns on its wings. The most severe BT outbreaks are recorded when vector activity is optimal during the late summer and autumn. The virus has been isolated several times in South Africa, Tunisia, Kenya, and in other countries (2, 40, 83, 86). Populations of C. Imicola have also been detected in areas close to the African continent such as Greece, Israel, and Turkey (2), coinciding with BTV occurrence. In addition to C. Imicola, other species have been proven to be BTV invertebrate host vectors, C. tororoensis, C. milnei, and C. obsoletus in Kenya and Cyprus, respectively, without significant effect on the epidemiology of BT. C. bolitinos, an African blood-sucking insect closely related to C. imicola, is considered a possible vector for African horse sickness, a viral disease closely related to BT, that breeds in the dung of the African buffalo and cattle.
Toward the northern part of the Mediterranean basin, incursions of the virus into Europe, for a long time free from the infection, are due to the circulation of C. imicola, as predominant vector in the Mediterranean region. BTV outbreaks have also been reported in the European part of Turkey and Greece where C. obsoletus of the sub-genus Avaritia was common, and C. imicola was absent (30). In Northern and Western Europe, the main Culicoides species involved in BTV transmission are C. obsoletus and C. scoticus (87).
In North America, C. variipennis is the major vector with the implication of at least three subspecies: C. v. occidentalis, C. v. sonorensis, and C. v. variipennis, which are genetically different with variable susceptibility to viral infection (2). Works on genetic control and polymorphism provided evidence for the involvement of gene expression in controlling the susceptibility of C. variipennis species to viral transmission. In Central and South America and the Caribbean Region, C. insignis is potentially the most predominant vector of BTV. It has been shown capable of transmitting the virus to susceptible sheep (88).
In Asia, the presence of several species of Culicoides has been confirmed. Nevertheless, C. imicola seems to be the most documented vector in Middle East countries, India, China, and other tropical and subtropical regions. Works carried out in the southern and eastern parts of the Asian continent have described other species of Culicoides (C. schultzei, C. gemellus, C. peregrinus, C. actoni, C. fulvus, and others) as being able to harbor and transmit BTV. Along with these species, C. wadai and C.brevitarsis are considered efficient vectors in Australia (89).
Vector competence is measured by its ability to contract the virus, amplify it, and successfully transmit it to a susceptible host species. It depends on several factors. Firstly, the number of inoculations per host is based on insect population dynamics, abundance, trophic preferences, and survival rates. Secondly, vector capacity depends largely on extrinsic factors such as temperature, barometric pressure, and wind speed. Culicoides survive at temperatures ranging from 15°C to 40°C. At temperatures below 17°C, their ability to survive and replicate the virus is diminished (19). When there is little or no wind, midges can be carried over distances of up to hundreds of kilometers (up to 700 km). However, strong winds prevent vector from flying, which hinders virus dissemination (90).
The interaction between climatic changes, the midges’ population dynamic, and ecology has been reported, and BTV transmission seems to be closely correlated to Culicoides vector’s seasonal behavior. The assessment of the role of Culicoides species as a vector requires in-depth knowledge of their spatio-temporal occurrence. In fact, understanding their distribution is crucial to enable the identification of high-risk transmission areas (91). They are specifically present in agricultural areas with varying abundance. Most species are characterized by twilight and nocturnal activities with peaks around sunset and sunrise (92). Response to climatic conditions raises real challenge and has been used as a topographical predictive factor in modeling conception aided by the seasonal abundance of Culicoides.
Worldwide trade of animals can spread the disease to livestock herds across vast distances by introducing infectious animals. Such importations involve transport of midges, which by feeding on infected animals, the real BTV reservoir, will increase transmission rates (93). This is the reason why livestock movement restrictions are applied during an outbreak. Otherwise, BTV may persist in the ruminant host during the winter (5), through prolonged viremia, which can last up to 45 days in sheep and 60 days in cattle, the lifespan of infected red blood cells (75). Under these conditions, the virus can overwinter for months without infecting new hosts in the absence of the vector (5).
In temperate to subtropical enzootic areas, the insect vector, the virus, and the mammalian host have the ability to coexist in climatic conditions suitable for BTV replication and transmission. Most infections occur between late summer and late autumn, which partly reverts to the seasonal nature of the lifestyle of Culicoides populations. The population density of midges is extremely reduced in over-wintering periods. Although infected adult midges can survive for a long time during cold periods, a study has shown that eggs are the most tolerant life stage of these insects in the cold season (94). Monitoring of Culicoides biting midges has been conducted at 12 locations in Switzerland over 3 years, using overnight trapping once a week (35). Results showed a statistically significant annual variation in Culicoides abundance and activity. High activity was observed between April and November and strongly reduced in winter. More than 80% of the annual midge collection was counted from June to September.
Among a panoply of environmental factors influencing the propagation and activity of Culicoides species, temperature remains one of the most influential variables by affecting the survival time and seasonal dynamic of adult biting midges. These insects are carried by hot and humid winds. Their survival would require temperatures above 12.5°C. A four-degree increase in temperature (13°C –17°C) was associated with a 5-day decrease in egg development time. In addition, fertility in females maintained at 13°C was significantly lower than in females maintained at temperatures of 17°C or above (19). Low vector competence for BTV-4 has been shown compared to other serotypes (such as BTV-1) after incubation at 23°C–25°C (85). A temperature of 37°C has been described as the optimal temperature for infectious blood during the experimental feeding of Culicoides.
Little is known regarding how or whether environmental factors such as temperature can affect the frequency of genetic reassortment within segmented arboviruses (95, 96). Therefore, the recent study conducted by Carpenter et al. (42) represents one of the first attempts to better characterize the impact of temperature on rates of reassortment in arthropod vectors. They suggested that reassortment frequency increased with higher temperature in Culicoides sonorensis coinfected with two BTV serotypes, BTV- 10 and BTV-17.
In the winter, low Culicoides biting midges activity is observed depending on unfavorable weather conditions and low temperatures. Populations are strongly reduced (35).
In northern and central Europe, Culicoides activity appears to reach its lowest level during the winter. Their endophagic behavior allows them to shelter from cold conditions inside farm buildings. Thus, the vector’s life cycle can be maintained during the cold season, which may impact both the virus survival and the probability of infection. Unlike in Afro-tropical and Mediterranean basin countries, a complete cessation of the vector activity of C. imicola as an exophagic species, is observed (5).
In cool conditions, the persistence of orbiviruses in vectors for a long time have been reported. Replication of these latent viruses resumes when subsequent exposure to warm temperatures occurs, resulting in pathogen transmission.
Some groups of biting midges are dominant in altitudes up to 1,200 m. Others are more frequent above 1,500 m (35). Small populations of Culicoides species remain active during winter, which could explain the persistence of the transmission cycle. At northern latitudes, most of them survive the winter as larvae (5).
Northward expansion of C. imicola to Europe is suggested to be aided by climate change given that 1°C rise in temperature, corresponding to 90 km of latitude and 150 m of altitude (2).
Interactions between climatic changes, midges’ population dynamic, and ecology are found to be likely risk factors for BTV infection and virogenesis rates. Culicoides species are hematophagous insects that occur throughout most of the inhabited world, and understanding their distribution is crucial to enable the identification of high-risk transmission areas. Moreover, there is evidence now of BTV infection through routes other than vector transmission, although still rarely considered as factors of the spread of the disease. Vector-independent transmission is considered limited in terms of epizootiology. However, it may be associated with the spread of novel serotypes and can have a substantial economic impact due to the birth of malformed calves (congenital malformations) or the loss of progeny.
Further investigation into predictive factors is required to better evaluate the risks associated with BT spread in endemic regions and outbreaks of new BTV serotypes to make appropriate assumptions for global control strategies and disease monitoring.
ST: Writing – original draft, Writing – review & editing. RL: Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The author thanks the members of research team of the virology laboratory, veterinary research institute of Tunisia.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Mellor PS, Boorman J. The transmission and geographical spread of African horse sickness and bluetongue viruses. Ann Trop Med Parasitol. (1995) 89:1–15. doi: 10.1080/00034983.1995.11812923
2. Mellor PS, Boorman J, Baylis M. Culicoides biting midges: their role as arbovirus vectors. Annu Rev Entomol. (2000) 45:307–40. doi: 10.1146/annurev.ento.45.1.307
3. Saminathan M, Pal Singh K, Khorajiya JH, Dinesh M, Vineetha S, Maity M, et al. An updated review on bluetongue virus: epidemiology, pathobiology, and advances in diagnosis and control with special reference to India. Vet Q. (2020) 40:258–321. doi: 10.1080/01652176.2020.1831708
4. Schwartz-Cornil I, Mertens PPC, Contreras V, Hemati B, Pascale F, Bréard E, et al. Bluetongue virus: virology, pathogenesis and immunity. Veterinary Res. (2008) 39:1. doi: 10.1051/vetres:2008023
5. Napp S, Gubbins S, Calistri P, Allepuz A, Alba A, García-Bocanegra I, et al. Quantitative assessment of the probability of bluetongue virus overwintering by horizontal transmission: application to Germany. Veterinary Res. (2011) 42:45. doi: 10.1186/1297-9716-42-4
6. Van Gennip R,GP, Van de Water SGP, Van Rijn PA. Bluetongue virus nonstructural protein NS3/NS3a is not essential for virus replication. PloS One. (2014) 9:e85788. doi: 10.1371/journal.pone.0085788
7. Courtejoie N, Bournez L, Zanella G, Durand B. Quantifying bluetongue vertical transmission in French cattle from surveillance data. Veterinary Res. (2019) 50:34. doi: 10.1186/s13567-019-0651-1
8. White JR, Williams DT, Wang J, Chen H, Melville LF, Davis SS, et al. Identification and genomic characterization of the first isolate of bluetongue virus serotype 5 detected in Australia. Veterinary Med science. (2019) 5:129–45. doi: 10.1002/vms3.156
9. Van den Bergh C, Coetzee P, Venter EH. Reassortment of bluetongue virus vaccine serotypes in cattle. J S Afr Vet Assoc. (2018) 89:e1–7. doi: 10.4102/jsava.v89i0.1649
10. Mohl BP, Kerviel A, Labadie T, Matsuo E, Roy P. Differential localization of structural and non-structural proteins during the bluetongue virus replication cycle. Viruses. (2020) 12:343. doi: 10.3390/v12030343
11. MacLachlan NJ, Bennie IO. Impact of bluetongue virus infection on the international movement and trade of ruminants. J Am Veterinary Med Assoc. (2006) 228:1346−49. doi: 10.2460/javma.228.9.1346
12. Van der Sluijs MTW, De Smit AJ, Moormann RJM. Vector independent transmission of the vector-borne bluetongue virus. Crit Rev Microbiol. (2014) 42:57–64. doi: 10.3109/1040841X.2013.879850
13. White SM, Sanders CJ, Shortall CR, Purse BV. Mechanistic model for predicting the seasonal abundance of Culicoides biting midges and the impacts of insecticide control. Parasites Vectors. (2017) 10:162. doi: 10.1186/s13071-017-2097-5
14. MacLachlan NJ, Drew CP, Darpel KE, Worwa G. The pathology and pathogenesis of bluetongue. J Com. Pathol. (2009) 141:1–16. doi: 10.1016/j.jcpa.2009.04.003
15. MacLachlan NJ, Mayo CE, Daniels PW, Savini G, Zientara S, Gibbs EP. Bluetongue. Rev Sci Tech. (2015) 34:329–40. doi: 10.20506/rst.34.2.2360
16. Stewart M, Hardy A, Barry G, Pinto RM, Caporale M, Melzi E, et al. Characterization of a second open reading frame in genome segment 10 of bluetongue virus. J Gen Virol. (2015) 96:3280–93. doi: 10.1099/jgv.0.000267
17. Roy P. Bluetongue virus proteins and particles and their role in virus entry, assembly, and release. Adv Virus Res. (2005) 64:69−123. doi: 10.1016/S0065-3527(05)64004-3
18. Mohd Jaafar F, Monsion B, Mertens PPC, Attoui H. Identification of orbivirus non-structural protein 5 (NS5), its role and interaction with RNA/DNA in infected cells. Int J Mol Sci. (2023) 24:6845. doi: 10.3390/ijms24076845
19. Mayo C, McDermott E, Kopanke J, Stenglein M, Lee J, Mathiason C, et al. Ecological dynamics impacting bluetongue virus transmission in North America. Front Veterinary Science. (2020) 7:186. doi: 10.3389/fvets.2020.00186
20. Chacko N, Nihar N, Mohanty NN, Biswas SK, Chand K, Yogisharadhya R, et al. A coiled-coil motif in non-structural protein 3 (NS3) of bluetongue virus forms an oligomer. Virus Genes. (2015) 51:244–51. doi: 10.1007/s11262-015-1230-9
21. OIE. Code sanitaire pour les animaux terrestres. Paris: Organisation Mondiale de la Santé Animale. Vol. 11. (2014).
22. Gerdes GH. A South African overview of the virus, vector, surveillance, and unique features of bluetongue. Vet Ital. (2004) 40:39–42. https://www.izs.it/vet_italiana/2004/40_3/39.
23. Alkhamis MA, Aguilar-Vega C, Fountain-Jones NM, Lin K, Perez AM, Sanchez-Vizcaıno JM. Global emergence and evolutionary dynamics of bluetongue virus. Sci Rep. (2020) 10:21677. doi: 10.1038/s41598-020-78673-9
24. Thelier A. The inoculation of sheep and the results in practice. Vet J. (1908) 64:600–7. doi: 10.1016/S0372-5545(17)68234-8
25. Coetzee P, Stokstad M, Venter EH, Myrmel M, Van Vuuren M. Bluetongue: A historical and epidemiological perspective with the emphasis on South Africa. Virol J. (2012) 9:1985. doi: 10.1186/1743-422X-9-198
26. MacLachlan N. J. Bluetongue: pathogenesis and duration of viraemia. Veterinaria italiana, (2004). 40(4):462–7.
27. Mellor PS, Pitzolis G. Observations on breeding sites and light-trap collections of Culicoides during an outbreak of bluetongue in Cyprus. Bull Entomol Res. (1979) 69:229–34. doi: 10.1017/S0007485300017697
28. Baldet T, Mathieu B, Delécolle JC, Gerbier G, Roger F. Emergence de la fièvre catarrhale ovine dans le Bassin méditerranéen et surveillance entomologique en France [in french]. Rev Élev. Méd. vét. Pays Trop. (2005) 58:125–32. doi: 10.19182/remvt.9923
29. Wilson AJ, Mellor PS. Bluetongue in Europe: past, present and future. Philos Trans R Soc B: Biol Sci. (2009) 364:2669−81. doi: 10.1098/rstb.2009.0091
30. Sperlova A, Zendulkova D. Bluetongue: a review. Veterinarni Medicina. (2011) 56:430–52. doi: 10.17221/3206-VETMED
31. Labadie T, Edward S, Polly R. Multiple routes of bluetongue virus egress. Microorganisms. (2020) 8:965. doi: 10.3390/microorganisms8070965
32. Koltsov A, Sodnom T, Andrey G, Denis K, et Galina K. Identification and Characterization of Bluetongue Virus Serotype 14 in Russia. Frontiers in Veterinary Sci. (2020) 7(février):26. doi: 10.3389/fvets.2020.00026
33. Saegerman C, Berkvens D, Mellor PS. Bluetongue epidemiology in the European union. Emerg Infect Dis. (2008) 14:4. doi: 10.3201/eid1404.071441
34. Sana K, Soufien S, Thameur BH, Liana T, Massimo S, Kaouther G, et al. Risk-based serological survey of bluetongue and the first evidence of bluetongue virus serotype 26 circulation in Tunisia. Veterinary Med science. (2022) 8:1671–82. doi: 10.1002/vms3.818
35. Kaufmann C, Steinmann IC, Hegglin D, Schaffner F, Mathis A. Spatio-temporal occurrence of Culicoides biting midges in the climatic regions of Switzerland, along with large scale species identification by MALDI-TOF mass spectrometry. Parasites Vectors. (2012) 5:246. doi: 10.1186/1756-3305-5-246
36. Wilson A, Mellor P. Bluetongue in Europe: vectors, epidemiology and climate change. Parasitol Res. (2008) 103:69−77. doi: 10.1007/s00436-008-1053-x
37. Ben Hassine T, Amdouni J, Monaco F, Savini G, Sghaier S, Ben Selimen I, et al. Emerging vector-borne diseases in dromedaries in Tunisia: West Nile, bluetongue, epizootic haemorrhagic disease and Rift Valley fever. Onderstepoort J Vet Res. (2017) 84:1316. doi: 10.4102/ojvr.v84i1.1316
38. Maan S, Maan NS, Ross-smith N, Batten CA, Shaw AE, Anthony SJ, et al. Sequence analysis of bluetongue virus serotype 8 from the Netherlands 2006 and comparison to other European strains. Virology. (2008) 377:308−18. doi: 10.1016/j.virol.2008.04.028
39. Cappai S, Rolesu S, Loi F, Liciardi M, Leone A, Marcacci M, et al. Western Bluetongue virus serotype 3 in Sardinia, diagnosis and characterization. Transbound Emerg Dis. (2019) 66:1426–31. doi: 10.1111/tbed.13156
40. Sossah CC, Madani H, Sailleau C, Nomikou K, Sadaoui H, Zientara S, et al. Molecular epidemiology of bluetongue virus serotype 1 isolated in 2006 from Algeria. Res Veterinary Science. (2011) 91:486–97. doi: 10.1016/j.rvsc.2010.10.002
41. Shaw AE, Ratinier M, Nunes SF, Nomikou K, Caporale M, Golder M, et al. Reassortment between two serologically unrelated bluetongue virus strains is flexible and can involve any genome segment. J Virol. (2013) 87:543–57. doi: 10.1128/JVI.02266-12
42. Carpenter M, Kopanke J, Lee J, Rodgers C, Reed K, Sherman TJ, et al. Assessing Reassortment between Bluetongue Virus Serotypes 10 and 17 at Different Coinfection Ratios in Culicoides sonorenesis. Viruses. (2024) 16:240. doi: 10.3390/v16020240
43. Goud AR, Pritchard LI. Relationships amongst bluetongue viruses revealed by comparisons of capsids and outer coat protein nucleotide sequences. Virus Res. (1990) 17:31–52. doi: 10.1016/0168-1702(90)90078-p
44. Ries C, Vögtlin A, Hüssy D, Jandt T, Gobet H, Hilbe M, et al. Putative novel atypical BTV serotype ‘36’identified in small ruminants in Switzerland. Viruses. (2021) 13:721. doi: 10.3390/v13050721
45. Westrich JA, McNulty EE, Carpenter M, Burton M, Reed K, Nalls A, et al. Monitoring longitudinal immunological responses to bluetongue virus 17 in experimentally infected sheep. Virus Res. (2023) 338:199246. doi: 10.1016/j.virusres.2023.199246
46. Chaignat V, Worwa G, Scherrer N, Hilbe M, Ehrensperger FC, Cortyen M, et al. Toggenburg Orbivirus, a new bluetongue virus: initial detection, first observations in field and experimental infection of goats and sheep. Vet Microbiol. (2009) 138:11–9. doi: 10.1016/j.vetmic.2009.02.003
47. Bréard E, Schulz C, Sailleau C, Bernelin-Cottet C, Viarouge C, Vitour D, et al. Bluetongue virus serotype 27: experimental infection of goats, sheep and cattle with three BTV-27 variants reveal atypical characteristics and likely direct contact transmission BTV-27 between goats. Transboundary Emerging Dis. (2018) 65:e251−63. doi: 10.1111/tbed.12780
48. Golender N, Bumbarov V, Eldar A, Lorusso A, Kenigswald G, Varsano JS, et al. Bluetongue serotype 3 in Israel 2013–2018: clinical manifestations of the disease and molecular characterization of Israeli strains. Front Veterinary Science. (2020) 7:112. doi: 10.3389/fvets.2020.00112
49. Belbis G, Zientara S, Bréard E, Sailleau C, Caignard G, Vitour D, et al. Chapter seven - bluetongue virus: from BTV-1 to BTV-27. Adv Virus Res. (2017) 99:161–97. doi: 10.1016/bs.aivir.2017.08.003
50. Reber A, Kreienbrock L, Casati S, Chaignat V, Schwermer H. Putative risk factors for infections with toggenburg orbivirus in goat herds in southern Switzerland (Canton of ticino). Veterinary Microbiol. (2012) 160:29−34. doi: 10.1016/j.vetmic.2012.05.027
51. Vögtlin A, Hofmann MA, Nenniger C, Renzullo S, Steinrigl A, Loitsch A, et al. Long-term infection of goats with bluetongue virus serotype 25. Veterinary Microbiol. (2013) 166:165−73. doi: 10.1016/j.vetmic.2013.06.001
52. Batten CA, Henstock MR, Steedman HM, Waddington S, Edwards L, Oura CAL. Bluetongue virus serotype 26: infection kinetics, pathogenesis and possible contact transmission in goats. Veterinary Microbiol. (2013) 162:62−67. doi: 10.1016/j.vetmic.2012.08.014
53. Batten CA, Darpel K, Henstock MR, Fay P, Veronesi E, Gubbins S, et al. [amp]]laquo; Evidence for Transmission of Bluetongue Virus Serotype 26 through Direct Contact ». Édité par Tetsuro Ikegami. PloS One. (2014) 9:e96049. doi: 10.1371/journal.pone.0096049
54. Bumbarov V, Golender N, Jenckel M, Wernike K, Beer M, Khinich E, et al. Characterization of bluetongue virus serotype 28. Transboundary Emerging Dis. (2020) 67:171−82. doi: 10.1111/tbed.13338
55. Batten CA, Henstock MR, Bin-Tarif A, Steedman HM, Waddington S, Edwards L, et al. Bluetongue virus serotype 26: infection kinetics and pathogenesis in Dorset Poll sheep. Veterinary Microbiol. (2012) 157:119–24. doi: 10.1016/j.vetmic.2011.11.033
56. Bonneau K, Mullens BA, MacLachlan NJ. Occurrence of Genetic Drift and Founder Effect during Quasi species Evolution of the VP2 and NS3/NS3A Genes of Bluetongue Virus upon Passage between Sheep, Cattle, and Culicoides sonorensis. J Virol. (2001) 75:8298–305. doi: 10.1128/jvi.75.17.8298-8305.2001
57. Kowalik TF, Li JK. Bluetongue virus evolution: sequence analyses of the genomic sl segments and major core protein VP7. Virology. (1991) 181:749–55. doi: 10.1016/0042-6822(91)90911-t
58. Nomikou K, Hughes J, Wash R, Kellam P, Breard E, Zientara S, et al. Widespread reassortment shapes the evolution and epidemiology of Bluetongue virus following European invasion. PloS Pathog. (2015) 11:e1005056. doi: 10.1371/journal.ppat.1005056
59. Gaudrealt NN, Mayo CE, Jasperson DC, Crossley BM, Breitmeyer RE, Johnson DJ, et al. Whole genome sequencing and phylogenetic analysis of Bluetongue virus serotype 2 strains isolated in the Americas including a novel strain from the western United States. J Vet Diagn. Invest. (2014) 26:553–7. doi: 10.1177/1040638714536902
60. Caporale M, Di Gialleonorado L, Janowicz A, Wilkie G, Shaw A, Savini G, et al. Virus and host factors affecting the clinical outcome of bluetongue virus infection. J Virology. (2014) 88:10399 –10411. doi: 10.1128/JVI.01641-14
61. Dal Pozzo F, Martinelle L, Thys C, Sarradin P. Experimental co-infections of calves with bluetongue virus serotypes 1 and 8. Vet Microbiol. (2013) 26:165(1–2):167-72. doi: 10.1016/j.vetmic.2013.01.016
62. Allepu A, Garcia-Bocanegra I, Napp S, Casal J, Arenas A, Saez M, et al. Monitoring bluetongue disease (BTV-1) epidemic in southern Spain during 2007. Prev Veterinary Med. (2007) 96:263–71. doi: 10.1016/j.prevetmed.2010.06.005
63. Sanchez-Cordon PJ, Pedrera M, Risalde MA, Molina V, Rodriguez-Sanchez B, Nunez A, et al. Potential role of proinflammatory cytokines in the pathogenetic mechanisms of vascular lesions in goats naturally infected with bluetongue virus serotype 1. Transboundary Emerging Diseases. (2013) 60:252–62. doi: 10.1111/j.1865-1682.2012.01343.x
64. Sghaier S, Hammami S, Goffredo M, Hammami M, Portanti O, Lorusso A, et al. New species of the genus Culicoides (Diptera Ceratopogonidae) for Tunisia, with detection of Bluetongue viruses in vectors. Veterinaria Ital. (2017) 53:357–66. doi: 10.12834/VetIt.986.5216.2
65. Celma CC, Bhattacharya B, Eschbaumer M, Wernike K, Beer M, Roy P. Pathogenecity study in sheep using reverse-genetics-based reassortant bluetongue viruses. Vet Micrpbiol. (2014) 174:139–47. doi: 10.1016/j.vetmic.2014.09.012
66. Dungu B, Gerdes T, Smit T. The use of vaccination in the control of bluetongue in southern Africa. Vet Ital. (2004) 40:616–22. https://www.izs.it/vet_italiana/2004/40_4/616.
67. Basak AK, Gouet P, Grimes J, Roy P, Stuart D. Crystal structure of the top domain of African horse sickness virus VP7: comparisons with bluetongue virus VP7. J Virol. (1996) 70:3797–806. doi: 10.1128/jvi.70.6.3797-3806.1996
68. Tan BH, Nason E, Staeuber N, Jiang W, Monastryrskaya K, Roy P. RGD tripeptide of bluetongue virus VP7 protein is responsible for core attachment to Culicoides cells. J Virol. (2001) 75:3937–47. doi: 10.1128/JVI.75.8.3937-3947.2001
69. Xia X, Wu W, Cui Y, Roy P, Zhou ZH. Bluetongue virus capsid protein VP5 perforates membranes at low endosomal pH during viral entry. Nat Microbiol. (2021) 6:1424–32. doi: 10.1038/s41564-021-00988-8
70. Ratinier M, Shaw AE, Barry G, Gu Q, Di Gialleonardo L, Janowicz A, et al. Bluetongue virus NS4 protein is an interferon antagonist and a determinant of virus virulence. J Virol. (2016) 90:5427–39. doi: 10.1128/JVI.00422-16
71. Fablet A, Kundlacz C, Dupré J, Hirchaud E, Postic L, Sailleau C, et al. Comparative virus-host protein interactions of the bluetongue virus NS4 virulence factor. Viruses. (2022) 14:182. doi: 10.3390/v14020182
72. Mohd Jaafar F, Belhouchet M, Monsion B, Bell-Sakyi L, Mertens PPC, Attoui H. Orbivirus NS4 proteins play multiple roles to dampen cellular responses. Viruses. (2023) 15:1908. doi: 10.3390/v15091908
73. Haiyan Fu. Mechanisms controlling the infection of Culicoides biting midges with bluetongue virus. University of Hertfordshire in United Kingdom: University of Hertfordshire Research Archive (1995). doi: 10.18745/th.14153
74. Han Z, Harty RN. The NS3 Protein of Bluetongue Virus Exhibits Viroporin-like Properties. J Biol Chem. (2004) 279:43092–7. doi: 10.1074/jbc.M403663200
75. Busquets G, Gillian M, Darpel D, Cooke L, Armstrong S, Simpson J, et al. An early block in the replication of the atypical bluetongue virus serotype 26 in culicoides cells is determined by its capsid proteins. Viruses. (2021) 13:919. doi: 10.3390/v13050919
76. Menzies FD, McCullough SJ, Mckeown IM, Forster JL, Jess S, Batten C, et al. Evidence for transplacental and contact transmission of bluetongue virus in cattle. Vet Res. (2008) 163:203–9. doi: 10.1136/vr.163.7.203
77. Rasmussen LD, Savini G, Lorusso A, Bellacicco A, Palmarini M, Caporale M, et al. Transplacental transmission of field and rescued strains of BTV-2 and BTV-8 in experimentally infected sheep. Vet Res. (2013) 44:75. http://www.veterinaryresearch.org/content/44/1/75.
78. EFSA Panel on Animal Health and Welfare. Bluetongue: control, surveillance and safe movement of animals. EFSA J. (2017) 15(3):e04698. doi: 10.2903/j.efsa.2017.4698
79. Attoui H, Monsion B, Klonjkowski B, Zientara S, Mertens PPC, Mohd Jaafar F, et al. Identification of the Genome Segments of Bluetongue Virus Type 26/Type 1 Reassortants Influencing Horizontal Transmission in a Mouse Model. Viruses. (2021) 13:2208. doi: 10.3390/v13112208
80. Zientara S, MacLachlan NJ, Calistri P, Sanchez-Vizcaino JM, Giovani S. Bluetongue vaccination in Europe. Expert Rev Vaccines. (2010) 9:989–91. doi: 10.1016/j.cimid.2007.07.006
81. De Clercq K, Vandaele L, Vanbinst T, Riou M, Deblauwe I, Wesselingh W, et al. Transmission of bluetongue virus serotype 8 by artificial insemination with frozen–thawed semen from naturally infected bulls. Viruses. (2021) 13:652. doi: 10.3390/v13040652
82. Pascall DJ, Nomikou K, Bréard E, Zientara S, Filipe A, Hoffmann B, et al. Frozen evolution” of an RNA virus suggests accidental release as a potential cause of arbovirus re-emergence. PloS Biol. (2020) 18:e3000673. doi: 10.1371/journal.pbio.3000673
83. Slama D, Babba H, Chaker E. Culicoides spp. (Diptera: ceratopogonidae) in Tunisia. In: Shields VDC, editor. Biological Control of Pest and Vector Insects. Biological Control of Pest and Vector Insects. USA: InTech (2017). doi: 10.5772/66944
84. Bartsch S, Bauer B, Wiemann A, Clausen PH, Steuber S. Feeding patterns of biting midges of the Culicoides obsoletus and Culicoides pulicaris groups on selected farms in Brandenburg, Germany. Parasitol Res. (2009) 105:373–80. doi: 10.1007/s00436-009-1408-y
85. Paslaru AL, Mathis A, Torgerson P, Veronesi E. Vector competence of pre-alpine Culicoides (Diptera: Ceratopogonidae) for bluetongue virus serotypes 1, 4 and 8. Parasites Vectors. (2018) 11:466. doi: 10.1186/s13071-018-3050-y
86. Walker AR, Davies FG. A preliminary survey of the epidemiology of bluetongue in Kenya. J Hyg. (1971) 69:47–60. doi: 10.1017/s0022172400021239
87. Balenghien T, Garros C, Mathieu B, Setier-Rio ML, Allene X, Gardes L, et al. La surveillance des Culicoïdes en France. Bull Epidémiologique. (2010) 35:8–9. https://be.anses.fr/fr/issue/35%20Bulletin%20%C3%A9pid%C3%A9miologique.
88. Tanya VN, Greiner EC, Gibbs EPJ. Evaluation of Culicoides insignis (Diptera: Ceratopogonidae) as a vector of bluetongue virus. Veterinary Microbiol. (1992) 32:1–14. doi: 10.1016/0378-1135(92)90002-b
89. Cybinski DH, Muller MJ. Isolation of arboviruses from cattle and insects at two sentinel sites in Queensland, Australia 1979-1985. Aust J Zool. (1990) 38:25–32. https://www.publish.csiro.au/zo/zo9900025.
90. Hendrickx G, Gilbert M, Staubach C, Elbers A, Mintiens K, Gerbier G, et al. A wind density model to quantify the airborne spread of Culicoides species during north-western Europe bluetongue epidemi. Prev veterinary Med. (2008) 87:162–81. doi: 10.1016/j.prevetmed.2008.06.009
91. Ba Gahn MC, Niakh F, Ciss M, Seck I, Lo MM, Fall AG, et al. Assessing the risk of occurrence of bluetongue in Senegal. Microorganisms. (2020) 8:1766. doi: 10.3390/microorganisms8111766
92. Braverman YA, Friger CGM, Mumcuoglu KY. Nocturnal activity of Culicoides imicola Kieffer (Diptera: Ceratopogonidae) in Israel. Russian Entomol J. (2008) 17:37–9.
93. Brand SPC, Keeling MJ. The impact of temperature changes on vector-borne disease transmission: Culicoides midges and bluetongue virus. J R Soc interface. (2017) 14:20160481. doi: 10.1098/rsif.2016.0481
94. McDermott EG, Mayo CE, Mullens BA. Low temperature tolerance of Culicoides sonorensis (Diptera: Ceratopogonidae) eggs, larvae, and pupae from temperate and subtropical climates. J Med Entomol. (2016) 54:264–74. doi: 10.1093/jme/tjw190
95. Carpenter S, Wilson A, Barber J, Veronesi E, Mellor P, Venter G, et al. Temperature dependence of the extrinsic incubation period of orbiviruses in Culicoides biting midges. PloS One. (2011) 6:e27987. doi: 10.1371/journal.pone.0027987
Keywords: bluetongue virus, serotypes, transmission routes, Culicoides, ecological dynamics, bluetongue monitoring
Citation: Thabet S and Lajnef R (2024) Potential mechanisms underlying bluetongue virus emergence and spread. Front. Virol. 4:1448192. doi: 10.3389/fviro.2024.1448192
Received: 12 June 2024; Accepted: 07 October 2024;
Published: 02 December 2024.
Edited by:
Glenn Andrew Marsh, Commonwealth Scientific and Industrial Research Organisation (CSIRO), AustraliaReviewed by:
Murat Şevik, Necmettin Erbakan University, TürkiyeCopyright © 2024 Thabet and Lajnef. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sara Thabet, c2FyYWh0aGFiZXRiZW5hYmRlbGplbGlsQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.