 Toscane Fourié1,2*
Toscane Fourié1,2* Guillaume André Durand1,2
Guillaume André Durand1,2 Franck Touret2
Franck Touret2 Géraldine Piorkowski2Audrey Dubot-Pérès2Xavier de Lamballerie2Isabelle Leparc-Goffart1,2Gilda Grard1,2
Géraldine Piorkowski2Audrey Dubot-Pérès2Xavier de Lamballerie2Isabelle Leparc-Goffart1,2Gilda Grard1,2- 1Centre National de Référence des Arbovirus, Institut de Recherche Biomédicale des Armées, Marseille, France
- 2Unité des Virus Émergents (UVE), Aix Marseille Université, IRD 190, INSERM 1207, Marseille, France
Laboratory-confirmed dengue virus (DENV) infections in Africa are rarely reported. In this study, we report 18 DENV serotype 1 (DENV-1) infections, diagnosed by the French National Reference Center for Arboviruses, in patients who had histories of recent travel in Africa. Our analyses revealed two cases, one from Niger in 2018 and one from the Republic of the Congo in 2016, where dengue fever had not been previously reported, and one case from Mauritania in 2015, where DENV-1 had not been previously reported. These cases support the reported spread of DENV outside its well-established tropical and subtropical environment toward the arid deserts of the Sahel. Phylogenetic analyses suggest that a single monophyletic DENV-1 lineage is currently in circulation in West Africa, having spread from East Africa after its original importation from Asia. Our study provides an improved understanding of DENV dynamics in Africa and underlines the importance of surveillance of travel-acquired infections.
1 Introduction
Dengue fever is a high-burden mosquito-borne disease endemic in tropical and subtropical regions of the world. It is caused by four antigenically distinct dengue viruses (DENV-1 to DENV-4) that are transmitted to humans by bites from mosquitoes belonging to the genus Aedes. Dengue virus (DENV) infections can be asymptomatic or manifest as flu-like symptoms, but occasionally develop into a severe, hemorrhagic form that can lead to severe shock syndrome. DENV is an 11-kb, positive-sense, single-stranded RNA virus belonging to the Flavivirus genus. The four distinct serotypes share around 65% nucleotide identity and do not induce cross-protective immunity (1). Within each serotype, multiple genotype lineages exist, with differing global distributions, as classified by Weaver and Vasilakis in 2009 (2).
Laboratory-confirmed DENV outbreaks have been reported in Africa since 1968 (3), predominantly caused by DENV-1 and DENV-2. Despite major improvements in the diagnosis and surveillance of infectious diseases in Africa during the last decade, dengue fever is widely considered to be underdiagnosed (4, 5), and laboratory-confirmed cases remain rare.
DENV-1 was first isolated in Africa from a febrile patient in Nigeria in 1968 (3). Of the five recognized DENV-1 genotypes (2), two have been reported to circulate in Africa, with distinct geographic distributions. The African lineage of genotype I (GI) has been reported solely in East Africa: in Djibouti in 1998, in Eritrea in 2010, and in Somalia in 2011 (6–8). The African lineage of genotype V (GV) was originally identified in West Africa, specifically in Nigeria in 1968 and in Côte d’Ivoire in 1985 and 1998 (3, 9, 10). However, recent observations from Gabon in 2012 and Angola in 2013 indicate that the GV African lineage is also present in Central Africa (11, 12). In addition to these DENV-1 lineages endemic in Africa, strains phylogenetically related to the Asian lineage of GV have recently been detected in travel-acquired infections from East and West Africa (13–16) and in outbreaks in West Africa (17, 18).
In the present study, we report laboratory-confirmed human DENV-1 infections thought to have been acquired by French patients during a stay in West and Central Africa between 2013 and 2019. We provide molecular characterization of the detected strains in aid of an improved understanding of DENV-1 distribution and circulation in Africa.
2 Materials and methods
2.1 DENV-1 infections associated with recent travel history to Africa
This retrospective study was conducted on samples collected by the French National Reference Center for Arboviruses (NRC). The NRC’s missions include surveillance, delivery of expertise, and confirmatory diagnosis of arbovirus infections. Various French facilities (e.g., hospitals, private laboratories, and medical teams temporarily based in Africa) may request confirmatory diagnosis for suspected arboviral cases from the NRC by submitting blood samples in the form of sera, plasma, or dried whole blood spots (DBS). An analysis request form, completed by the medical examiner, provides detailed clinical and travel information, as well as patient consent for medical research. Data from patient records collected and anonymously presented in this report were data used in routine diagnosis by the NRC.
Criteria for selection of DENV-1 cases from the NRC records were as follows: (i) symptoms developed during a stay in Africa or during the week following return to mainland France, according to the travel data provided by medical staff in a mandatory information form supplied with each sample, and (ii) a laboratory-confirmed DENV-1 diagnosis by reverse transcription quantitative polymerase chain reaction (RT-qPCR). All cases were tested with pan-DENV RT-qPCR and serotype was confirmed via DENV-1-specific RT-qPCR in accordance with previously described protocols (19). Exclusion criteria were as follows: (i) serotyping diagnosis based on high qPCR titers (Ct value > 36), (ii) insufficient sample volume, (iii) incomplete travel details, (iv) multi-country travel, and (v) cases submitted by facilities in French overseas territories where DENV is endemic. Samples were selected from the period 2013 to 2019. Sera and plasma were stored at –20°C and DBS samples at 4°C after initial diagnostic analysis by the NRC.
2.2 Virus isolation
DENV PCR-positive plasma and sera samples were inoculated on Vero (ATCC® CCL-81™) and C6/36 (ATCC CRL-1660™) standard cell lines for virus isolation and amplification. Viral amplification was detected by RT-qPCR (19).
2.3 Dengue virus genome sequencing
Viral RNA from cell culture supernatant or primary sample was extracted using the EZ1 Advanced XL instrument (Qiagen, Hilden, Germany) and the EZ1 Virus mini kit, in accordance with the manufacturer’s recommendations. Two sets of primers were used for full-genome amplification (20, 21). Next-generation sequencing was attempted as previously described (20). All complete coding sequences (CDSs) obtained in this study were deposited in GenBank under accession numbers listed in Table 1.
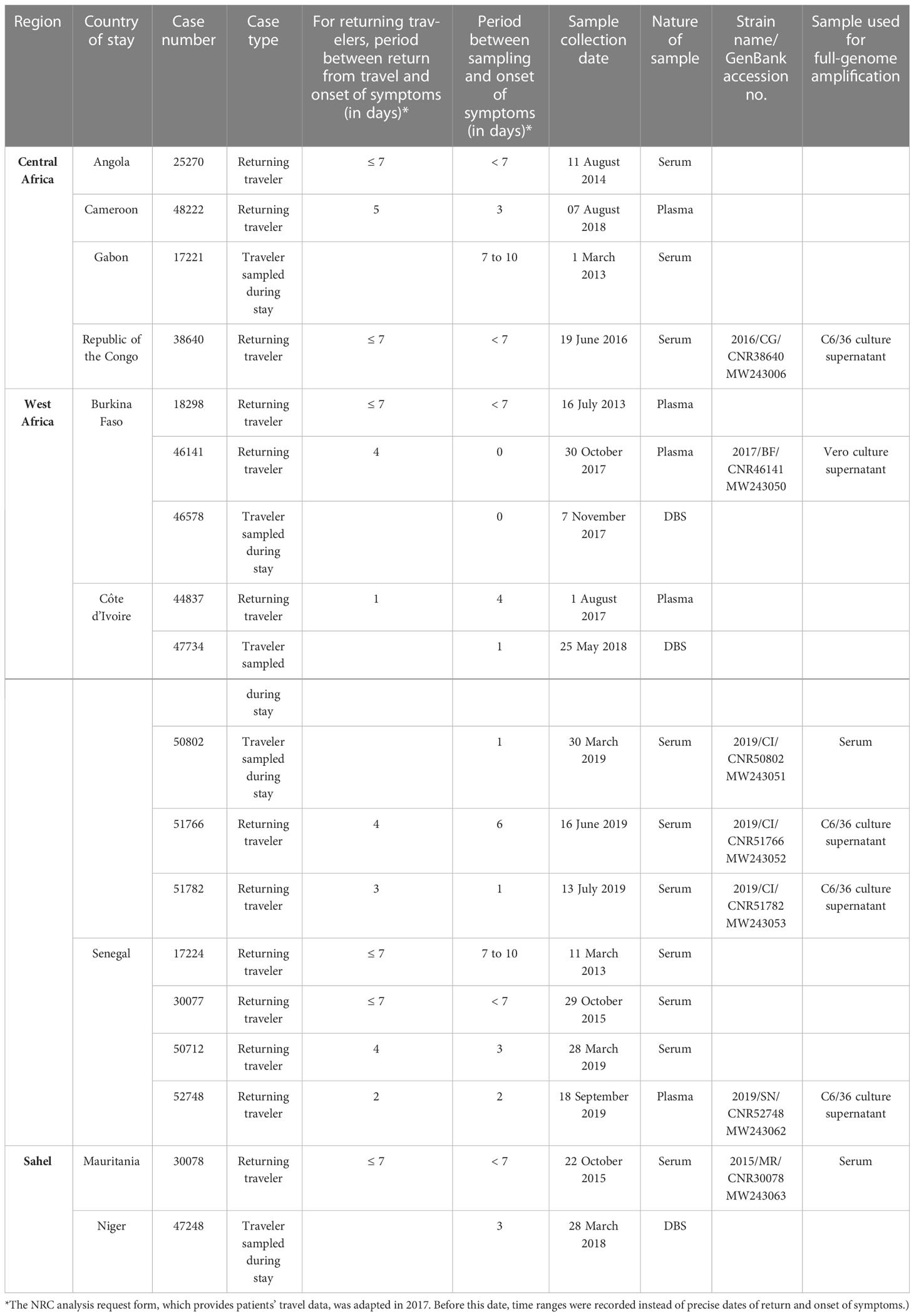
Table 1 Characteristics of DENV-1 cases analyzed in this study.
2.4 Phylogenetic analysis
2.4.1 Complete CDS dataset
A total of 105 strains representing the major DENV-1 clades from various locations and years were selected and complete CDSs with dates and locations of isolation were retrieved from GenBank. In addition, all sequences available in GenBank (containing data on isolation year and origin) that met one of the following criteria were selected: (i) DENV-1 sequence from an African country; or (ii) DENV-1 sequence sharing over 95% nucleotide identity with sequences obtained in this study. All partial (length < 10,100 bases), ambiguous (presence of continuous N; >0.2% single ambiguous sites), and redundant sequences (sharing >99.5% nucleotide identity, with identical epidemiological characteristics, i.e., identical country and year of isolation) were excluded. The final complete CDS dataset contained 110 sequences (Supplementary Table 1).
2.4.2 Envelope gene dataset
Approximately 40% of DENV-1 strains from Africa retrieved from GenBank were excluded from the complete CDS dataset due to insufficient length. Consequently, a partial genome dataset focusing on the Envelope (E) gene was constructed. All sequences from the complete CDS dataset were included, and additional E-gene sequences were retrieved from GenBank using identical selection criteria. All partial (length < 1,460 bases), ambiguous (presence of continuous N; >0.2% single ambiguous sites), and redundant sequences (sharing >99.5% nucleotide identity, with identical epidemiological characteristics) were excluded. The final E-gene dataset contained 123 sequences (including 13 additional E-gene sequences) (Supplementary Table 1).
2.4.3 Bioinformatics analysis
Sequences obtained in this study were added to both the complete CDS and the E-gene datasets and were aligned using the online version of MAFFT (v7.182) (22). All regions containing free-end gaps were trimmed manually from the nucleotide MAFFT output, leaving a final complete CDS alignment length of 10,102 bases and a final E-gene alignment length of 1,464 bases. Using the Galaxy web platform (usegalaxy.org) (23), maximum-likelihood phylogenetic trees were constructed with the online version of IQ-Tree (v2.1.2) (24). In terms of model parameters, ModelFinder indicated that substitution model GTR+F+I+G4 (i.e., a general time reversible model with empirical base frequencies and a rate heterogeneity model allowing discrete γ distribution, with four categories and evolutionarily invariant sites) was the best fit for both the E-gene and complete CDS alignments based on the Bayesian information criterion (–merit BIC) (25). One thousand bootstrap replicates were performed by UFBoot using the recommended optimized parameters (–ufboot 1,000; –bnni) (26). Final trees were midpoint-rooted and annotated with the online version of iTol (version 6.6; itol.embl.de) (Figures 1, 2).
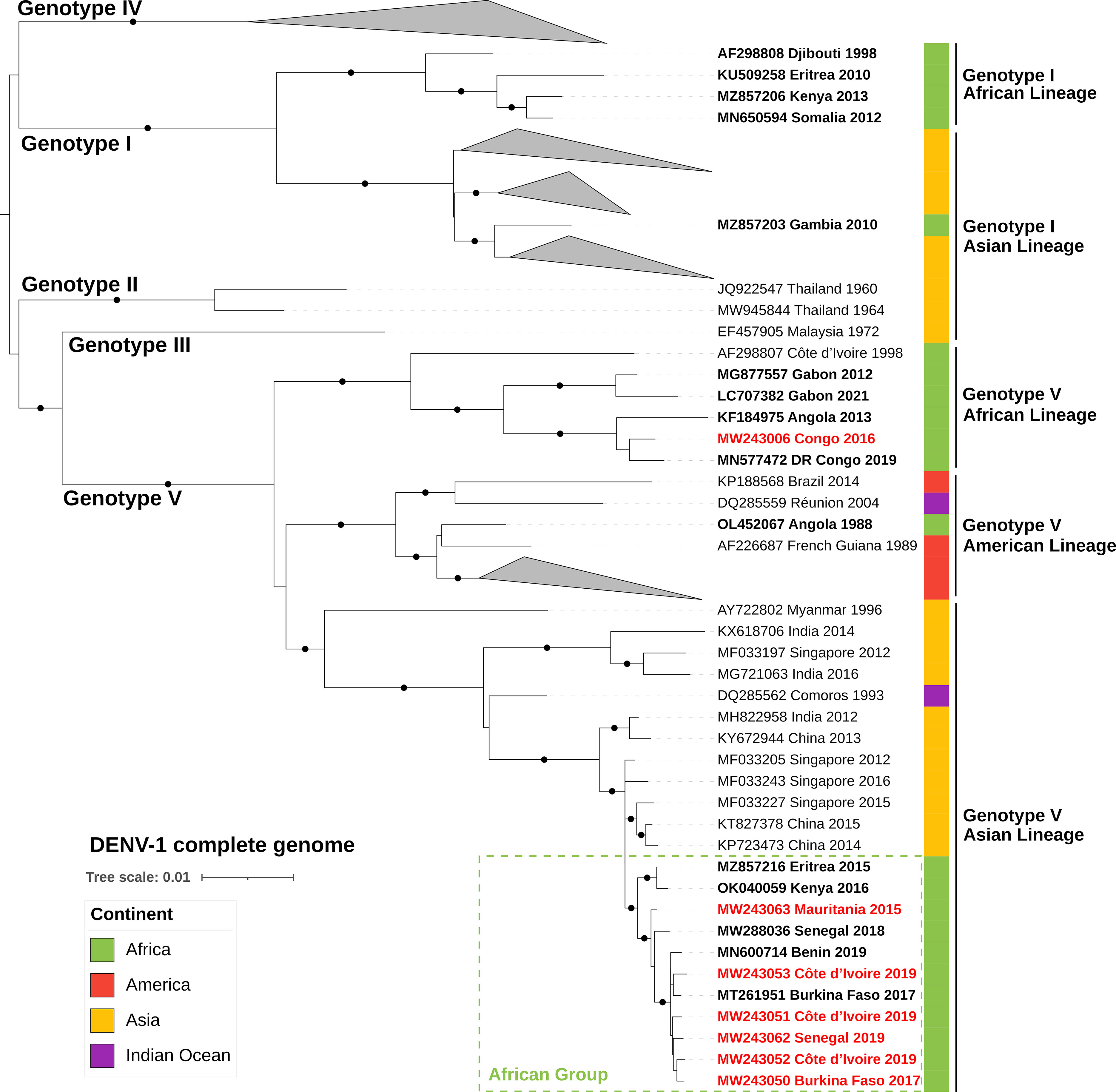
Figure 1 ML phylogenetic tree based on 117 complete CDSs of DENV-1 strains (10,102-nucleotide-long sequences after trimming). DENV-1 strains from African countries are indicated in bold and strains from our dataset are in red text. Each strain is identified by its GenBank accession number, country of origin, and year of sample collection. The tree was rooted at the mid-point and bootstrap values equal to 100% are indicated by a black circle on the branch. The scale bar indicates genetic distance (nucleotide substitutions per site). The color strip indicates location of infection. DENV, dengue virus; DR Congo, Democratic Republic of the Congo.
3 Results
3.1 DENV-1 infections associated with recent travel history to Africa
A total of 18 PCR-confirmed DENV-1 infections met the inclusion criteria for this study, as detailed in Table 1 and Figure 3. Of these 18 cases, 13 had been imported to mainland France and the patients had developed symptoms within 1 week of return from their stay in to Africa (Table 1). In the other five cases, onset of symptoms had occurred during the patient’s stay in Africa: medical consultation and sample collection were performed by French medical teams based locally (Table 1). Overall, 12 infections were acquired in West Africa: in Burkina Faso (n=3), Côte d’Ivoire (n=5), and Senegal (n=4). Four were acquired in Central Africa: in Angola (n=1), Cameroon (n=1), Gabon (n=1), and the Republic of the Congo (n=1). Two were acquired in the Sahel: in Niger (n=1) and Mauritania (n=1).
Interestingly, we have found two DENV-1 infections acquired in areas where dengue fever had not been previously reported, one in Niger in 2018 and one in the Republic of the Congo in 2016, and one infection acquired in Mauritania in 2015, where DENV-1 had not previously been reported (to our knowledge). Burkina Faso, Senegal, and Côte d’Ivoire accounted for the majority (12/18) of cases of DENV-1 infection in our study.
3.2 Virus isolation and genome sequencing
Five DENV-1 isolates were successfully cultured (Table 1). Three were isolated from patients who had visited Senegal (n=1; isolate 2019/SN/CNR52748) or Côte d’Ivoire (n=2; isolates 2019/CI/CNR51766 and 2019/CI/CNR51782) in 2019. Isolates 2016/CG/CNR38640 and 2017/BF/CNR46147 were obtained from individuals who had visited the Republic of the Congo in 2016 and Burkina Faso in 2017, respectively.
Full-genome sequences were obtained from all five isolates. In addition, two full-genome sequences were obtained from primary blood samples: one from Mauritania (2015/MR/CNR30078) and one from Côte d’Ivoire (2019/CI/CNR50802). Overall, seven full-genome sequences of DENV from West Africa (n=5), Central Africa (n=1), and the Sahel (n=1) were obtained (Table 1).
3.3 Phylogenetic analysis
Phylogenetic tree reconstruction of the complete CDSs showed that the DENV-1 strains of our sample set belonged to genotype V (Figure 1). Strain 2016/CG/CNR38640 (MW243006) from the Republic of the Congo (Central Africa) belonged to the previously described endemic African lineage and clustered with other Central African strains. Phylogenetic analysis of available E-gene sequences from the GV African lineage showed that strains currently circulating in Central Africa (i.e., detected from 2012 to 2021) were phylogenetically distinct from an older West African cluster from that same lineage (Figure 2). In addition, E-gene sequence KX270806 identified in a traveler returning to China in 2013 (without further travel information available) belonged to this Central African cluster (identical E-gene sequence with the Angolan 2013 KM277610 and DRC 2015 LC360648 strains) (Figure 2).
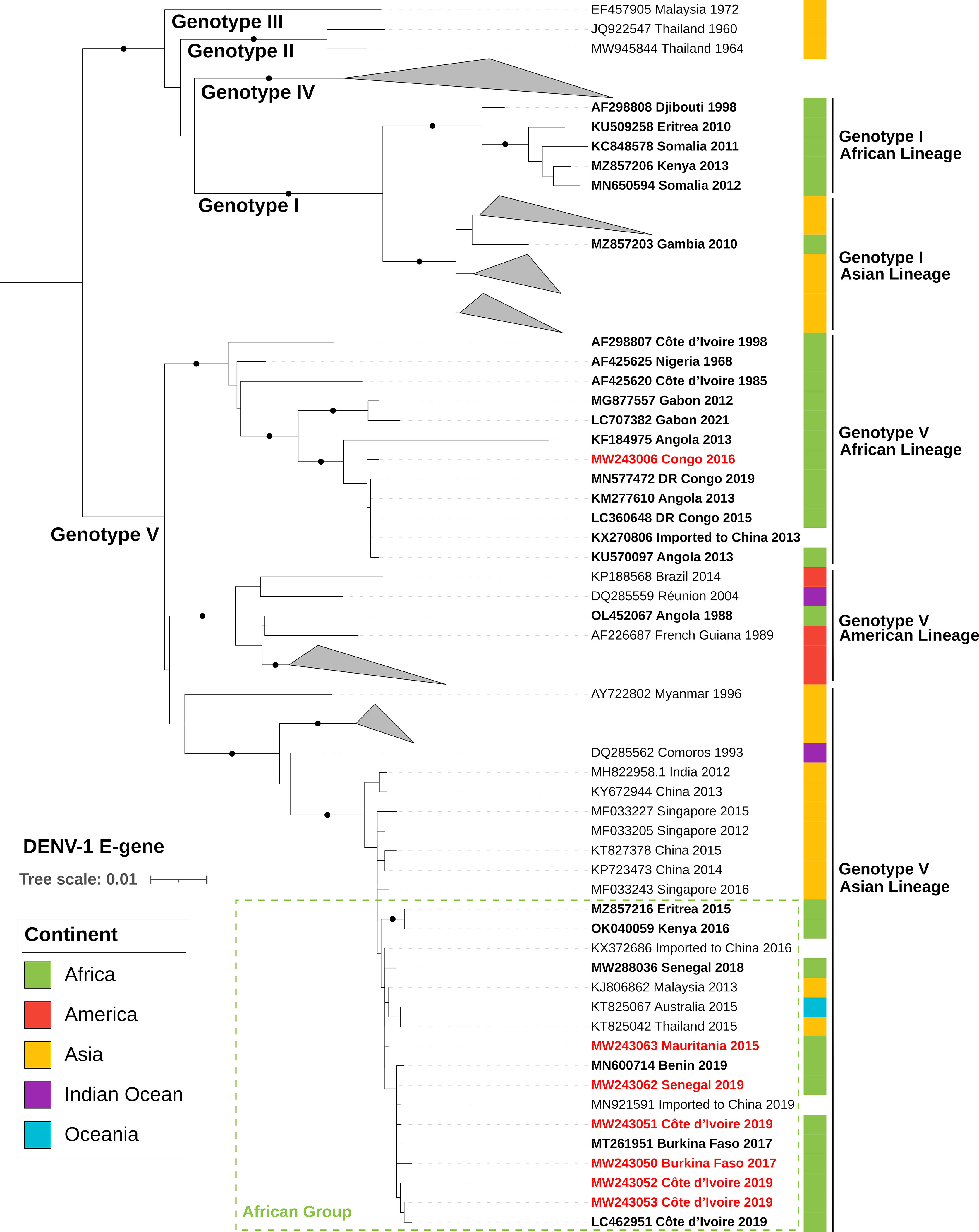
Figure 2 ML phylogenetic tree based on 130 E-gene sequences of DENV-1 strains (1,464-nucleotide-long sequences after trimming). DENV-1 strains from African countries are indicated in bold and strains from our dataset are in red text. Each strain is identified by its GenBank accession number, country of origin, and year of sample collection. The tree was rooted at the mid-point and bootstrap values equal to 100% are indicated by a black circle on the branch. The scale bar indicates genetic distance (nucleotide substitutions per site). The color strip indicates location of infection. DENV, dengue virus; DR Congo, Democratic Republic of the Congo.
All West African and Sahelian strains (i.e., those from Burkina Faso, Côte d’Ivoire, Senegal, and Mauritania) from our sample set formed a distinct cluster within the GV Asian lineage, in both complete CDS and E-gene phylogenies (Figures 1, 2). They clustered with recently described strains from East Africa (i.e., Kenya OK040059 and Eritrea MZ857216) and West Africa (i.e., Côte d’Ivoire LC462951, Burkina Faso MT261951, Senegal MW288036, and Benin MN600714). The most closely related sequences were from Singaporean and Chinese strains sampled from 2012 to 2016 (i.e., MF033205, MF033227, KT827378, and MF033243), which shared up to 99.2% complete CDS nucleotide pairwise identity with this African cluster within the GV Asian lineage. In addition, the E-gene phylogeny revealed the presence of strains reported in Oceania and Asia that clustered with this African lineage (Figure 2). Several strains were detected during an outbreak in Malaysia in 2013 (e.g., KJ806860–62) without any reported travel in Africa. Two strains were imported to China by travelers in 2016 and 2019, with no travel details available.
4 Discussion
Cases from our dataset of travelers have revealed the presence of DENV-1 in countries where the serotype and/or virus had yet to be reported (Figure 3).
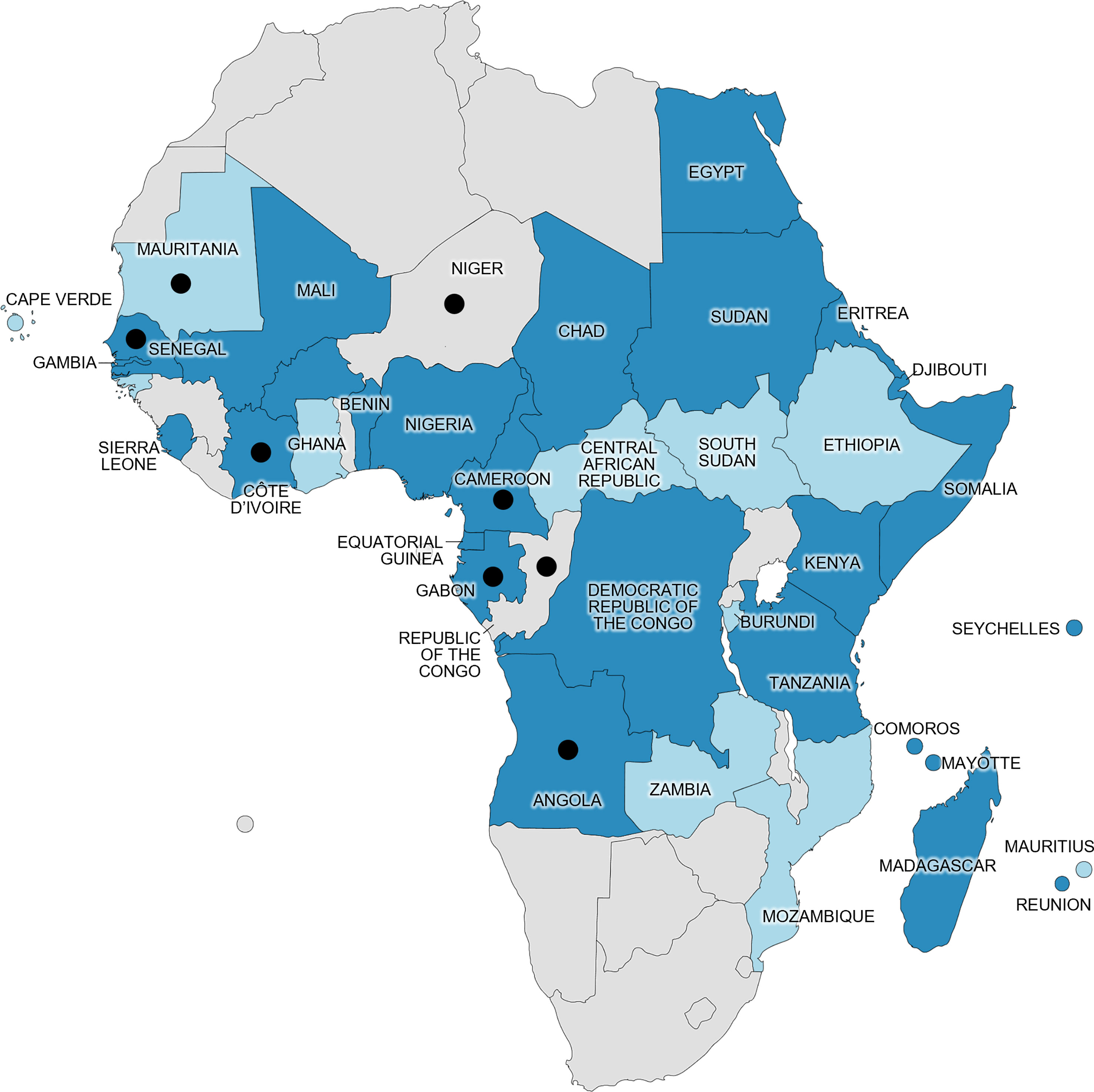
Figure 3 Distribution of confirmed human DENV cases in Africa prior to our study (at least one PCR-diagnosed case acquired in the country mentioned in a published report, as of December 2022) and location of DENV-1 cases in our study (black dots). Dark blue represents DENV-1 presence specifically, and light blue represents DENV presence in general (other serotype or unserotyped). Gray represents absence of reported confirmed DENV human cases, as defined by the World Health Organization (27). White represents bodies of water. Map generated using mapchart.net©.
We report two cases of DENV-1 acquired in the Sahel, a region where DENV circulation is rarely documented. The Sahelian climate is characterized by an absence of rainfall for several months of the year, with no natural water surfaces for mosquito breeding. However, Aedes species have been reported across the entire Sahel region (28–31). Indeed, anthropophilic mosquitoes such as Aedes species can thrive in anthropogenic water sources, especially during the moderate rainy season, from July to September. Interestingly, some laboratory-confirmed DENV infections have been detected in Sahelian countries, such as Chad, Sudan, Mali, and Mauritania, since 1986 (6, 32–37) (Figure 3). In Niger, Aedes vectors are established in dwellings from the south to the central regions (28). The first reported case of dengue fever was an imported case from Cuba in August 2022 of unknown serotype (38); however, our observation in case number 47248 suggests that DENV was present in the country prior to this, at least as early as 2018. Whether it resulted from a recent viral introduction or had been in undetected circulation for years remains unknown. The infection occurred in March during the 8-month-long dry season (i.e., the period of no rainfall from October to April), suggesting the presence of competent vectors outside the rainy season. Among other Sahelian countries, DENV-1 had previously been found in travelers returning from Eritrea (7, 15), Chad (35), and Sudan (39), and it caused an outbreak in Bamako, the capital city of Mali, in 2006 (32) (Figure 3). The DENV-1 infection of case number 30078 also places this serotype in Mauritania, 1 year after the description of the establishment of Ae. aegypti in the capital city, Nouakchott (40).
In West and Central Africa, substantial rainfall all year round, the tropical climate, and forests provide a suitable environment for annual vector proliferation, with sylvatic and urban Aedes species, including Ae. aegypti and Ae. albopictus, being broadly established (40). Consequently, DENV (and DENV-1 in particular) has been reported in most countries of the region since 1968, mostly in travelers returning from the region and in the form of localized outbreaks (3, 6, 9, 10, 13–15, 18, 33, 39, 41–54). However, no official reports of DENV in the Republic of the Congo have been published, despite the presence of abundant competent vectors and known recurrent arboviral outbreaks (55, 56). The DENV-1 infection of case number 38640 was acquired during a stay to the Republic of the Congo, indicating that DENV was present in the country at least as early as 2016, and its occurrence at several time points in all neighboring countries (51, 53, 54, 57) suggests that it has probably been in undetected circulation for years.
Available genomic data from DENV-1 in Africa, supplemented with our sequences, reveal the current circulation of several distinct lineages.
All known Central African DENV-1 strains cluster within the GV African lineage. The oldest strains of this lineage were isolated in Nigeria in 1968 (AF425625) and in Côte d’Ivoire in 1985 (AF425620), indicating endemic circulation in sub-Saharan Africa (Figure 2). The GV African lineage was implicated in a major urban outbreak in Angola in 2013 (11, 33, 58–60), thus demonstrating its epidemic potential on the continent. Viruses from this cluster have already been exported from Africa by travelers returning to Asia (53, 61), Europe (54, 59), and Israel (57).
DENV-1 strains currently detected in West Africa (including Mauritania) cluster in the GV Asian lineage, in a monophyletic group composed of strains detected in Africa since 2015. An introduction from Southeast Asia to Africa, probably through East African countries, could have occurred around 2012, as suggested by a recent Swedish study (15). This lineage was found to be responsible for sporadic cases in Senegal and Mali since 2015 and for significant outbreaks in the following years in Senegal (43). Genomic data from this study were not available at the time of writing and are therefore not included in our analysis. This Senegalese study suggested that DENV-1 was introduced to Senegal via Mali around 2014 (43), which is consistent with the timeline reported by Alfsnes et al. (15). Since its introduction to Africa, the GV Asian lineage has been detected in multiple countries throughout the continent, revealing rapid geographical spread: Eritrea in 2015 (15), Mauritania in 2015, Senegal from 2015 to 2019 (43), Kenya in 2016 (15), Mali in 2017 (43), Burkina Faso in 2017 (17), Benin in 2019 (13), and Côte d’Ivoire in 2019 (16). It is important to note that during this period no other DENV genotype was reported in West Africa. Based on current knowledge, it could be hypothesized that the GV Asian lineage of DENV-1 has replaced the historical African lineage that was last detected in West Africa in 1998 in Côte d’Ivoire (AF298807). However, the disparity in publicly available data limits our understanding of the DENV-1 dynamic within the continent. Outside Africa, viruses belonging to this monophyletic group according to the E-gene phylogenetic tree have been identified in travelers in China in 2016 and 2019 (KX372686 and MN921591, respectively), in Australia and Thailand in 2015, and in an outbreak in Malaysia in 2013 (Figure 2). Due to the high nucleotide identity of the E-gene sequences within this group and the lack of full-genome sequences, care should be taken in the interpretation of phylogenetic trajectories. Nevertheless, these observations reveal the exchange of highly similar viruses on multiple continents over the last 10 years.
Increasing economic exchanges between Asia and Africa over the past decades have resulted in an influx of Asian workers and freight shipping through African ports, thus creating putative importation routes for infectious diseases from Asia to Africa and vice versa (62). Considering the presence of local competent vectors in sub-Saharan Africa and the endemicity of DENV in Asia, more introduction events are to be expected in the future.
5 Conclusion
Our findings from molecular analyses indicated active circulation of two distinct DENV-1 genetic lineages in Africa: (i) the African group of GV Asian lineage, imported from Asia probably around 2012, which is currently spreading in the Sahel and West Africa; and (ii) the historical GV African lineage detected in Central Africa. Our cases found in the Sahel suggest that DENV, alongside its anthropophilic vectors, is spreading outside its well-established tropical and subtropical environments into the arid deserts of the Sahel. Moreover, this study underlines the importance of surveillance of travel-acquired infections, as publicly shared epidemiological surveillance data can only improve our overall understanding of disease circulation, in this instance in remote locations with limited access to molecular analysis.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession numbers can be found in the article/Supplementary Material.
Ethics statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Author contributions
IL-G, XL, GG, and GD contributed to conception and design of the study. TF and GP performed laboratory analysis. GP and AD-P contributed to primer design. GP, FT, and TF performed bioinformatic treatment and analysis. TF wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by (i) the French Defense Health Service, (ii) the European Virus Archive goes global project (EVAg; European Union—Horizon 2020 program under grant agreement no. 871029; http://www.european-virus-archive.com/), and (iii) Santé Publique France. The funders had no role in study design, data collection, analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
The authors thank Manon Geulen, Laurent Bosio, Thomas Canivez, Vincent Palomo, and Camille Placidi for their support in laboratory investigations, and Dr David Wilkinson for his useful insights on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fviro.2023.1124848/full#supplementary-material
References
1. Guzman MG, Halstead SB, Artsob H, Buchy P, Farrar J, Gubler DJ, et al. Dengue: a continuing global threat. Nat Rev Microbiol (2010) 8(12):S7–16. doi: 10.1038/nrmicro2460
2. Weaver SC, Vasilakis N. Molecular evolution of dengue viruses: Contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infect Genet Evol (2009) 9(4):523–40. doi: 10.1016/j.meegid.2009.02.003
3. Carey DE, Causey OR, Reddy S, Cooke AR. Dengue viruses from febrile patients in Nigeria, 1964-68. Lancet (1971) 297(7690):105–6. doi: 10.1016/S0140-6736(71)90840-3
4. Amarasinghe A, Kuritsk JN, Letson GW, Margolis HS. Dengue virus infection in Africa. Emerg Infect Dis (2011) 17(8):1349–54. doi: 10.3201/eid1708.101515
5. Jaenisch T, Junghanss T, Wills B, Brady OJ, Eckerle I, Farlow A, et al. Dengue expansion in Africa-not recognized or not happening? Emerg Infect Dis (2014) 20(10):e140487. doi: 10.3201/eid2010.140487
6. de Laval F, Plumet S, Simon F, Deparis X, Leparc-Goffart I. Dengue surveillance among French military in Africa. Emerg Infect Dis (2012) 18(2):342–3. doi: 10.3201/eid1802.111333
7. Vainio K, Noraas S, Holmberg M, Fremstad H, Wahlstrøm M, Ånestad G, et al. Fatal and mild primary dengue virus infections imported to Norway from Africa and south-east Asia, 2008-2010. Eurosurveillance (2010) 15(38):19666. doi: 10.2807/ese.15.38.19666-en
8. Bosa HK, Montgomery JM, Kimuli I, Lutwama JJ. Dengue fever outbreak in Mogadishu, Somalia 2011: Co-circulation of three dengue virus serotypes. Int J Infect Dis (2014) 21:3. doi: 10.1016/j.ijid.2014.03.412
9. Goncalvez AP, Escalante AA, Pujol FH, Ludert JE, Tovar D, Salas RA, et al. Diversity and evolution of the envelope gene of dengue virus type 1. Virology (2002) 303(1):110–9. doi: 10.1006/viro.2002.1686
10. Durand J, Vallée L, de Pina J, Tolou H. Isolation of a dengue type 1 virus from a soldier in West Africa (Côte d’Ivoire). Emerg Infect Dis J (2000) 6(1):83–4. doi: 10.3201/eid0601.000116
11. Parreira R, Conceição C, Centeno-Lima S, Marques N, da CJS, Abreu C, et al. Angola’s 2013 dengue outbreak: clinical, laboratory and molecular analyses of cases from four Portuguese institutions. J Infect Dev Ctries. (2014) 8(09):1210–5. doi: 10.3855/jidc.4910
12. Fontaine A, Lequime S, Moltini-Conclois I, Jiolle D, Leparc-Goffart I, Reiner RC, et al. Epidemiological significance of dengue virus genetic variation in mosquito infection dynamics. PloS Pathog (2018) 14(7):e1007187. doi: 10.1371/journal.ppat.1007187
13. Fourié T, Luciani L, Amrane S, Zandotti C, Leparc-Goffart I, Ninove L, et al. Dengue virus type 1 infection in traveler returning from Benin to France, 2019. Emerg Infect Dis (2020) 26(8):1946–9. doi: 10.3201/eid2608.200055
14. Ayolabi CI, Olusola BA, Ibemgbo SA, Okonkwo GO. Detection of dengue viruses among febrile patients in Lagos, Nigeria and phylogenetics of circulating dengue serotypes in Africa. Infect Genet Evol (2019) 75:103947. doi: 10.1016/j.meegid.2019.103947
15. Alfsnes K, Eldholm V, Gaunt MW, de Lamballerie X, Gould EA, Pettersson JHO. Tracing and tracking the emergence, epidemiology and dispersal of dengue virus to Africa during the 20th century. One Health (2021) 13:100337. doi: 10.1016/j.onehlt.2021.100337
16. Suzuki T, Kutsuna S, Nakamoto T, Ota M, Ishikane M, Yamamoto K, et al. Dengue virus serotype 1 exported to Japan from côte d’Ivoire, 2019. Jpn J Infect Dis (2021) 74(2):148–50. doi: 10.7883/yoken.JJID.2019.303
17. Letizia AG, Pratt CB, Wiley MR, Fox AT, Mosore M, Agbodzi B, et al. Retrospective genomic characterization of a 2017 dengue virus outbreak, Burkina Faso. Emerg Infect Dis (2022) 28(6):1198–210. doi: 10.3201/eid2806.212491
18. Gaye A, Ndiaye T, Sy M, Deme AB, Thiaw AB, Sene A, et al. Genomic investigation of a dengue virus outbreak in thiès, Senegal, in 2018. Sci Rep (2021) 11(1):10321. doi: 10.1038/s41598-021-89070-1
19. Leparc-Goffart I, Baragatti M, Temmam S, Tuiskunen A, Moureau G, Charrel R, et al. Development and validation of real-time one-step reverse transcription-PCR for the detection and typing of dengue viruses. J Clin Virol (2009) 45(1):61–6. doi: 10.1016/j.jcv.2009.02.010
20. Baronti C, Piorkowski G, Leparc-Goffart I, de Lamballerie X, Dubot-Pérès A. Rapid next-generation sequencing of dengue, EV-A71 and RSV-a viruses. J Virol Methods (2015) 226:7–14. doi: 10.1016/j.jviromet.2015.09.004
21. Dubot-Pérès A, Vongphrachanh P, Denny J, Phetsouvanh R, Linthavong S, Sengkeopraseuth B, et al. An epidemic of dengue-1 in a remote village in rural Laos. PloS Negl Trop Dis (2013) 7(8):e2360. doi: 10.1371/journal.pntd.0002360
22. Katoh K, Rozewicki J, Yamada KD. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings Bioinf (2019) 20(4):1160–6. doi: 10.1093/bib/bbx108
23. Galaxy Community. The galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res (2022) 50(W1):W345–351. doi: 10.1093/nar/gkac247
24. Nguyen LT, Schmidt HA, von Haeseler A, Minh BQ. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol (2015) 32(1):268–74. doi: 10.1093/molbev/msu300
25. Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods (2017) 14(6):587–9. doi: 10.1038/nmeth.4285
26. Hoang DT, Chernomor O, von Haeseler A, Minh BQ, Vinh LS. UFBoot2: Improving the ultrafast bootstrap approximation. Mol Biol Evol (2018) 35(2):518–22. doi: 10.1093/molbev/msx281
27. Control of Neglected Tropical Diseases, Emergencies Preparedness, Guidelines Review Committee. Laboratory testing for zika virus and dengue virus infections: interim guidance. World Health Organization (2022) p. 1–14. Available at: https://www.who.int/publications-detail-redirect/WHO-ZIKV_DENV-LAB-2022.1.
28. Labbo R, Doumma A, Mahamadou I, Arzika I, Soumana A, Kadri S, et al. Distribution and relative densities of aedes aegypti in Niger. Med Sante Trop (2019) 29(1):47–54. doi: 10.1684/mst.2019.0882
29. Tandina F, Doumbo O, Yaro AS, Traoré SF, Parola P, Robert V. Mosquitoes (Diptera: Culicidae) and mosquito-borne diseases in Mali, West Africa. Parasit Vectors (2018) 11:467. doi: 10.1186/s13071-018-3045-8
30. Mint Mohamed Lemine A, Ould Lemrabott MA, Hasni Ebou M, Mint Lekweiry K, Ould Ahmedou Salem MS, Ould Brahim K, et al. Mosquitoes (Diptera: Culicidae) in Mauritania: a review of their biodiversity, distribution and medical importance. Parasit Vectors (2017) 10(1):35. doi: 10.1186/s13071-017-1978-y
31. Müller GC, Tsabari O, Traore MM, Traore SF, Doumbia S, Kravchenko VD, et al. First record of aedes albopictus in inland Africa along the river Niger in Bamako and mopti, Mali. Acta Trop (2016) 162:245–7. doi: 10.1016/j.actatropica.2016.07.008
32. Phoutrides EK, Coulibaly MB, George CM, Sacko A, Traore S, Bessoff K, et al. Dengue virus seroprevalence among febrile patients in Bamako, Mali: results of a 2006 surveillance study. Vector Borne Zoonotic Dis Larchmt N (2011) 11(11):1479–85. doi: 10.1089/vbz.2011.0622
33. Toro C, Trevisi P, López-Quintana B, Amor A, Iglesias N, Subirats M, et al. Imported dengue infection in a Spanish hospital with a high proportion of travelers from Africa: A 9-year retrospective study. Am J Trop Med Hyg (2017) 96(3):701–7. doi: 10.4269/ajtmh.16-0335
34. Msimang V, Weyer J, le Roux C, Leman P, Kemp A, Paweska J. Dengue fever in south Africa: an imported disease. Commun Dis Surveill Bull (2013) 11(3):58–62.
35. Biernat B, Stańczak J, Szostakowska B, Wroczyńska A, Kuna A, Nahorski WL, et al. Different serotypes of dengue virus (DENV) imported by polish travellers from dengue endemic areas to Poland. Int Marit Health (2015) 66(2):72–6. doi: 10.5603/IMH.2015.0018
36. Fourié T, El Bara A, Dubot-Pérès A, Grard G, Briolant S, Basco LK, et al. Emergence of dengue virus serotype 2 in Mauritania and molecular characterization of its circulation in West Africa. PloS Negl Trop Dis (2021) 15(10):e0009829. doi: 10.1371/journal.pntd.0009829
37. Hyams KC, Oldfield EC, Scott RM, Bourgeois AL, Gardiner H, Pazzaglia G, et al. Evaluation of febrile patients in port Sudan, Sudan: isolation of dengue virus. Am J Trop Med Hyg (1986) 35(4):860–5. doi: 10.4269/ajtmh.1986.35.860
38. World Health Organization (WHO). Outbreaks and emergencies bulletin, week 36: 29 august - 4 September 2022. WHO | Regional Office for Africa (2022). Available at: https://apps.who.int/iris/bitstream/handle/10665/362389/OEW36-290804092022.pdf.
39. Domingo C, Niedrig M, Gascón J, Palacios G, Reyes N, Malo MJ, et al. Molecular surveillance of circulating dengue genotypes through European travelers. J Travel Med (2011) 18(3):183–90. doi: 10.1111/j.1708-8305.2011.00501.x
40. Mint Lekweiry K, Ould Ahmedou Salem MS, Ould Brahim K, Ould Lemrabott MA, Brengues C, Faye O, et al. Aedes aegypti (Diptera: Culicidae) in Mauritania: First report on the presence of the arbovirus mosquito vector in Nouakchott. J Med Entomol (2015) 52(4):730–3. doi: 10.1093/jme/tjv051
41. La Ruche G, Renaudat C, Tarantola A, Caro V, Ledrans M, Dejour-Salamanca D, et al. Increase in dengue fever imported from côte d’Ivoire and West Africa to France (2010). Available at: http://www.who.int/iris/handle/10665/170965.
42. Im J, Balasubramanian R, Ouedraogo M, Wandji Nana LR, Mogeni OD, Jeon HJ, et al. The epidemiology of dengue outbreaks in 2016 and 2017 in Ouagadougou, Burkina Faso. Heliyon (2020) 6(7):e04389. doi: 10.1016/j.heliyon.2020.e04389
43. Dieng I, Cunha M, Diagne MM, Sembène PM, Zanotto PM de A, Faye O, et al. Origin and spread of the dengue virus type 1, genotype V in Senegal, 2015-2019. Viruses (2021) 13(1):57. doi: 10.3390/v13010057
44. Rico-Hesse R. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology (1990) 174(2):479–93. doi: 10.1016/0042-6822(90)90102-W
45. Fagbami A. Epidemiological investigations on arbovirus infections at igbo-ora, Nigeria. Trop Geogr Med (1977) 29(2):187–91.
46. de Araújo Lobo JM, Mores CN, Bausch DG, Christofferson RC. Short report: Serological evidence of under-reported dengue circulation in Sierra Leone. PloS Negl Trop Dis (2016) 10(4):e0004613. doi: 10.1371/journal.pntd.0004613
47. Wichmann O, Lauschke A, Frank C, Shu P, Niedrig M, Huang J, et al. Dengue antibody prevalence in German travelers - volume 11, number 5–may 2005 - emerging infectious disease journal - CDC. Emerg Infect Dis (2005) 11(5):762–5. doi: 10.3201/eid1105.050097
48. Yousseu FBS, Nemg FBS, Ngouanet SA, Mekanda FMO, Demanou M. Detection and serotyping of dengue viruses in febrile patients consulting at the new-bell district hospital in douala, Cameroon. PloS One (2018) 13(10):e0204143. doi: 10.1371/journal.pone.0204143
49. Nemg Simo FB, Sado Yousseu FB, Evouna Mbarga A, Bigna JJ, Melong A, Ntoude A, et al. Investigation of an outbreak of dengue virus serotype 1 in a rural area of kribi, south Cameroon: A cross-sectional study. Intervirology (2018) 61(6):265–71. doi: 10.1159/000499465
50. Demanou M, Pouillot R, Grandadam M, Boisier P, Kamgang B, Hervé JP, et al. Evidence of dengue virus transmission and factors associated with the presence of anti-dengue virus antibodies in humans in three major towns in Cameroon. PloS Negl Trop Dis (2014) 8(7):e2950. doi: 10.1371/journal.pntd.0002950
51. Ushijima Y, Abe H, Mbadinga MJVM, Ondo GN, Bikangui R, Agnandji ST, et al. Re-emergence of dengue, chikungunya, and zika viruses in 2021 after a 10-year gap in Gabon. IJID Reg. (2022) 5:68–71. doi: 10.1016/j.ijregi.2022.08.013
52. Makiala-Mandanda S, Ahuka-Mundeke S, Abbate JL, Pukuta-Simbu E, Nsio-Mbeta J, Berthet N, et al. Identification of dengue and chikungunya cases among suspected cases of yellow fever in the democratic republic of the Congo. Vector-Borne Zoonotic Dis (2018) 18(7):364–70. doi: 10.1089/vbz.2017.2176
53. Yamamoto SP, Kasamatsu Y, Kanbayashi D, Kaida A, Shirano M, Kubo H, et al. Dengue virus in traveler returning to Japan from the democratic republic of the Congo, 2015. Jpn J Infect Dis (2019) 72(6):426–8. doi: 10.7883/yoken.JJID.2018.554
54. Colavita F, Vairo F, Carletti F, Boccardo C, Ferraro F, Iaiani G, et al. Full-length genome sequence of a dengue serotype 1 virus isolate from a traveler returning from democratic republic of Congo to Italy, July 2019. Int J Infect Dis (2020) 92:46–8. doi: 10.1016/j.ijid.2019.12.023
55. Mombouli JV, Bitsindou P, Elion DOA, Grolla A, Feldmann H, Niama FR, et al. Chikungunya virus infection, Brazzaville, republic of Congo, 2011. Emerg Infect Dis (2013) 19(9):1542–3. doi: 10.3201/eid1909.130451
56. Fritz M, Taty Taty R, Portella C, Guimbi C, Mankou M, Leroy EM, et al. Re-emergence of chikungunya in the republic of the Congo in 2019 associated with a possible vector-host switch. Int J Infect Dis (2019) 84:99–101. doi: 10.1016/j.ijid.2019.05.013
57. Sessions OM, Khan K, Hou Y, Meltzer E, Quam M, Schwartz E, et al. Exploring the origin and potential for spread of the 2013 dengue outbreak in Luanda, Angola. Glob Health Action (2013) 6:21822. doi: 10.3402/gha.v6i0.21822
58. Abreu C, Silva-Pinto A, Lazzara D, Sobrinho-Simões J, Guimarães JT, Sarmento A. Imported dengue from 2013 Angola outbreak: Not just serotype 1 was detected. J Clin Virol (2016) 79(Supplement C):77–9. doi: 10.1016/j.jcv.2016.04.011
59. Schwartz E, Meltzer E, Mendelson M, Tooke A, Steiner F, Gautret P, et al. Detection on four continents of dengue fever cases related to an ongoing outbreak in Luanda, Angola, march to may 2013. Euro Surveill Bull Eur Sur Mal Transm Eur Commun Dis Bull (2013) 18(21):20488. doi: 10.2807/ese.18.21.20488-en
60. Hunsperger EA, Sharp TM, Lalita P, Tikomaidraubuta K, Cardoso YR, Naivalu T, et al. Use of a rapid test for diagnosis of dengue during suspected dengue outbreaks in resource-limited regions. J Clin Microbiol (2016) 54(8):2090–5. doi: 10.1128/JCM.00521-16
61. Shi Y, Li S, Li X, Zheng K, Yuan S, Huang J. Epidemiological and molecular characterization of dengue viruses imported into guangzhou during 2009–2013. SpringerPlus (2016) 5(1):1635. doi: 10.1186/s40064-016-3257-3
Keywords: dengue virus serotype 1, traveler, phylogeny, dengue fever, Africa
Citation: Fourié T, Durand GA, Touret F, Piorkowski G, Dubot-Pérès A, de Lamballerie X, Leparc-Goffart I and Grard G (2023) Molecular characterization of dengue virus serotype 1 infections in French travelers from Africa between 2013 and 2019. Front. Virol. 3:1124848. doi: 10.3389/fviro.2023.1124848
Received: 15 December 2022; Accepted: 27 March 2023;
Published: 20 April 2023.
Edited by:
Gerald Misinzo, Sokoine University of Agriculture, TanzaniaReviewed by:
Edgar Simulundu, University of Zambia, ZambiaJacky Flipse, Rijnstate Hospital, Netherlands
Copyright © 2023 Fourié, Durand, Touret, Piorkowski, Dubot-Pérès, de Lamballerie, Leparc-Goffart and Grard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Toscane Fourié, dG9zY2FuZS5mb3VyaWVAZ21haWwuY29t