 Angelica A. Palm1*
Angelica A. Palm1* Srinivas Veerla2
Srinivas Veerla2 Jacob Lindman2Per-Erik Isberg3
Jacob Lindman2Per-Erik Isberg3 Emil Johansson1,4Antonio Biague5
Emil Johansson1,4Antonio Biague5 Fredrik Månsson4Hans Norrgren2
Fredrik Månsson4Hans Norrgren2 Joakim Esbjörnsson4
Joakim Esbjörnsson4 Patrik Medstrand4
Patrik Medstrand4 Marianne Jansson1 on behalf of the SWEGUB CORE group
Marianne Jansson1 on behalf of the SWEGUB CORE group- 1Department of Laboratory Medicine, Lund University, Lund, Sweden
- 2Department of Clinical Sciences Lund, Lund University, Lund, Sweden
- 3Department of Statistics, School of Economics and Management, Lund University, Lund, Sweden
- 4Department of Translational Medicine, Lund University, Lund, Sweden
- 5The National Laboratory of Public Health (LNSP), Bissau, Guinea-Bissau
Disease progression is slower in HIV-2, as compared with HIV-1 infection, in accordance with low or undetectable plasma viremia at viral setpoint. However, it is unclear why most HIV-2 infected individuals are still at risk of developing AIDS. To explore if specific host responses are linked to HIV disease severity, we have compared blood gene expression profiles between HIV seronegative and HIV-1, HIV-2 or dually HIV-1/HIV-2 infected individuals. In this study the gene encoding Interferon alpha-inducible protein 27 (IFI27) was found to be the most differentially expressed. Detailed expression analysis revealed significantly higher IFI27 expression in HIV infected individuals compared with seronegative individuals, irrespectively of HIV type. Moreover, IFI27 expression was higher in HIV-1 than in HIV-2 infected individuals. Multiple linear regression analysis, adjusting for age and sex, showed also that plasma viral load was the strongest predictor of IFI27 expression, followed by CD4% and HIV type. In line with this, IFI27 expression was found to be higher in HIV-2 viremic, compared with HIV-2 aviremic individuals. Still, HIV-2 aviremic individuals displayed elevated IFI27 expression compared with seronegative individuals. Furthermore, in HIV-2 infected individuals, IFI27 expression was also correlated with plasma markers previously linked to inflammation and disease progression in HIV infection. Taken together, our findings suggest that sustained elevation of type I interferon signaling, here reflected by elevated IFI27 expression in the chronic infection phase, is a key pathogenic feature of both HIV-1 and HIV-2.
Introduction
Both HIV-1 and HIV-2 are causative agents of AIDS and are characterized by a progressive decline in CD4+ T-cells (1). However, the CD4+ T-cell decline and the rate of disease progression in untreated individuals is faster in HIV-1 than in HIV-2 infection (2, 3). In individuals that are dually infected with both HIV-1 and HIV-2 (HIV-D), it has also been shown that a preceeding HIV-2 infection delays disease progression (4, 5). The plasma viral load (pVL) at set point, and when adjusted for CD4+ T-cell level, is higher in HIV-1 infected than in HIV-2 infected individuals (6–8). Accordingly, a large proportion of HIV-2 infected individuals will remain aviremic, i.e. with low or undetectable viremia, for extended periods of time (9). Still, disease progression occurs also in aviremic HIV-2 infected individuals (10–12), although the mechanisms behind this phenomenon are not clear.
HIV-1 disease progression is characterized by chronic immune activation and exhaustion (13). HIV-1 elite controllers, a group that in some aspects resembles HIV-2 aviremic individuals, presents with reduced immune activation and exhaustion compared to HIV-1 viremic individuals (6, 13). Less is known about the HIV-2 pathogenesis, but disease progression has been coupled to T-cell activation and exhaustion, and to the level of plasma inflammation markers (10–12, 14–19). Interestingly, also aviremic HIV-2 infected individuals have been suggested to harbor increased frequencies of activated and exhausted CD4+ and CD8+ T-cells compared to HIV seronegative individuals (10, 12).
Upon virus encounter, the innate immune response provides the first line of defence, including production of type I interferons (IFN-I) (20). This in turn results in the transcription of interferon-stimulated genes (ISGs), which, for example, inhibits viral transcription, translation and replication, thereby contributing to viral control during acute infections (20, 21). However, persistent up-regulation of ISGs during chronic infections, is associated with chronic and generalized immune activation, which is a key feature of CD4+ T-cell depletion and disease progression in pathogenic HIV-1 and SIV infection (22–25). By contrast, nonpathogenic chronic SIV infection is associated with lower levels of ISG expression and immune activation during the chronic infection phase, compared with pathogenic SIV and HIV-1 infection (22–24). Although associations between IFN-I and ISG expression and disease severity have been investigated for HIV-1 infection and both pathogenic and nonpathogenic SIV infection (22–25), such studies have not included HIV-2, a virus strain originating from nonpathogenic SIVsm (26).
In this study, gene expression in whole blood was compared between individuals infected with HIV-1, HIV-2 or HIV-D, and HIV seronegative individuals. Results showed that the ISG encoding the Interferon alpha-inducible protein 27 (IFI27), was significantly linked to disease severity in both HIV-1 and HIV-2 infection.
Materials and Methods
Study Participants
The individuals included in this study were part of a well-defined occupational cohort of police officers in Guinea-Bissau (27, 28). In this study, 39 HIV-1, 43 HIV-2, and 14 HIV-D infected individuals and 35 HIV seronegative individuals were enrolled (Table 1). All individuals were treatment naïve or unsuccessfully treated [defined by a pVL above 1000 RNA copies/ml plasma (29)]. We also included nine HIV-1 infected individuals that were successfully treated, i.e. with a pVL < 1000 copies/ml. The HIV-2 infected individuals were further subdivided according to level of pVL, where 13 had a pVL ≥ 75 copies/ml, and were referred to as viremic, and 30 had a pVL < 75 copies/ml and were referred to as aviremic (Table 1). Among the aviremic HIV-2 infected individuals 16 had pVL that was detected below the technical quantification threshold (above 0 but below 75 copies/ml). Informed consent was obtained from all the study participants, and the study was approved by the National Ethical Committee, Ministry of Public Health in Guinea-Bissau, and the Ethical Committee at Lund University.

Table 1 Characteristics of study participants included in the qPCR analysis.
CD4+ T-Cell Level and Plasma Viral Load Determination
HIV status, CD4+ T-cell levels and pVL were determined as previously described (10). We used CD4% instead of absolute CD4+ T-cell counts for analysis, as CD4% has been shown to be a more stable marker in settings with high pathogenic burden (30). The technical pVL quantification threshold was 75 copies/ml plasma for HIV-1 or HIV-2 single-infected, and 135 copies/ml plasma for HIV-D infected individuals (31).
Blood Collection and RNA Isolation
Whole blood (2.5 ml) was collected from study participants in PAXgene tubes (PreAnalytiX). Sample tubes were stored frozen until use. The tubes were thawed at room temperature overnight, before proceeding with total RNA extraction using the PAXgene Blood RNA kit (Qiagen) according to the manufacturer’s instructions.
Microarray Analysis
An exploratory microarray gene expression analysis was performed on samples from HIV seronegative (n = 8), HIV-1 (n = 8), HIV-2 (n = 7) and HIV-D (n = 8) infected individuals to identify candidate genes for further analyses. HIV infected individuals were matched on CD4% to allow identification of virus specific differences in gene expression. Total RNA was depleted of globins using GLOBINclear kit (ThermoFisher Scientific) according to manufacturer’s instructions. RNA quantity and integrity were analyzed using NanoDrop Spectrophotometry and the Agilent 2100 Bioanalyzer (Agilent). Gene expression data were generated using the Illumina HT12 v4 BeadChip (Illumina) which provides genome-wide transcriptional coverage of well-characterized genes, gene candidates, and splice variants. It targets more than 25,000 annoted genes with more than 48,000 probes. Of note, IFI27 is targeted by a single probe. Expression data were initially pre-processed and normalized using Quantile Normalization method (32). These analyses were performed using GenomeStudio software V2011.1. Non-annotated probe sets and probe sets with signal intensities below the median of the negative control intensities in 80% of the samples that did not belong to one condition were excluded. Following this the remaining probe set comprised of 20595, 20819, 20562 and 20689 probes for the comparisons of gene expression between HIV seronegative individuals and HIV infection groups combined (i.e. HIV-1, HIV-2 and HIV-D), HIV-1, HIV-2 and HIV-D infected individuals, respectively. Raw and processed data have been deposited in the Gene Expression Omnibus (GSE200606).
qPCR Analysis
Total RNA was quantified using a NanoDrop 2000 Spectrophotometer (ThermoFisher Scientific). cDNA synthesis was performed on 500 ng total RNA using Superscript IV (ThermoFisher Scientific) with random hexamers (Invitrogen) according to manufacturer’s instructions, in a total reaction volume of 20 µl. IFI27 and GAPDH (used as internal control) were amplified using the Hs00271467_m1 and Hs03929097_g1 TaqMan gene expression assays, respectively, with TaqMan Fast Advanced Master Mix (all from ThermoFisher Scientific). Reactions were set up according to manufacturer’s instructions, using 2 µl of cDNA as input, and analyzed on a StepOnePlus instrument (Applied Biosystems). The fold change in gene expression was calculated using the 2^-ΔΔCt method (33) and geometric means with confidence intervals are presented.
Plasma Inflammation Markers
For HIV-2 infected individuals, plasma concentrations of interferon gamma-induced protein 10 (IP-10, also known as CXCL10), soluble CD14 (sCD14), and beta-2-microglobulin (b2m) were analyzed using the Magnetic Luminex assay (R&D Systems Inc.) on the Bio-Plex 200 platform (Bio-Rad Laboratories Inc.) according to manufacturer’s instructions. Plasma was diluted 1:2, 1:600 and 1:4000 for quantification of IP-10, sCD14 and b2m, respectively.
Statistical Analysis
Statistical analyses of differentially expressed genes were performed in R by student’s t-test with Benjamini-Hochberg post-hoc correction for multiple comparisons. Differentially expressed genes were considered when the fold change was above 1.5 and the adjusted p-value was below 0.05. All other statistical analyses were performed using GraphPad Prism 9.2.0 (GraphPad software, La Jolla, California). To achieve normal distribution, the following variables were log10-transformed prior to statistical analysis: IFI27 fold change, CD4%, pVL, b2m, sCD14 and IP-10. Differences between study participants were analyzed using t-test, comparing two groups, and One-way ANOVA with Tukey’s post-hoc test, comparing more than two groups. Pearson correlation analysis was used to assess correlations between IFI27 and CD4%, pVL and plasma inflammation markers. Multiple linear regression analysis was used to identify significant predictors of IFI27 expression.
Results
IFI27 Differentiates HIV Infected From HIV Seronegative Individuals
We performed an exploratory array expression analysis to identify genes that were differentially expressed in HIV infected [HIV-1 (n = 8), HIV-2 (n = 7) or HIV-D (n = 8)] and HIV seronegative (n = 8) individuals (Table S1), where the HIV infected individuals were matched on CD4%. IFI27 was found to be the most strongly upregulated gene, with a 16.7-fold higher expression in HIV infected individuals compared with seronegative controls (adjusted p-value = 0.004, Figure S1).
To validate the array results, we determined IFI27 gene expression using qPCR in 39 HIV-1, 43 HIV-2, and 14 HIV-D infected individuals, and 35 HIV seronegative controls (Table 1). Among these study participants HIV-2 infected individuals had lower pVL levels compared to HIV-1 (p < 0.001) and HIV-D infected individuals (p < 0.001). Moreover, the CD4% was higher in HIV-2 infected compared with HIV-1 (p < 0.001) and HIV-D (p = 0.040) infected individuals. Seronegative individuals also had higher CD4% compared with HIV-1 (p < 0.001), HIV-2 (p < 0.001) and HIV-D (p < 0.001) infected individuals. HIV-2 infected individuals were older, as compared to HIV-1 and HIV-D infected individuals (p = 0.001 and p = 0.009, respectively).
IFI27 expression was found to be 42.2 (p < 0.001), 5.5 (p < 0.001) and 24.5 (p < 0.001) fold higher in HIV-1, HIV-2 and HIV-D infected as compared with seronegative individuals, respectively (Figure 1). Furthermore, the IFI27 expression was 7.7 fold higher in HIV-1 compared with HIV-2 infected individuals (p < 0.001, Figure 1).
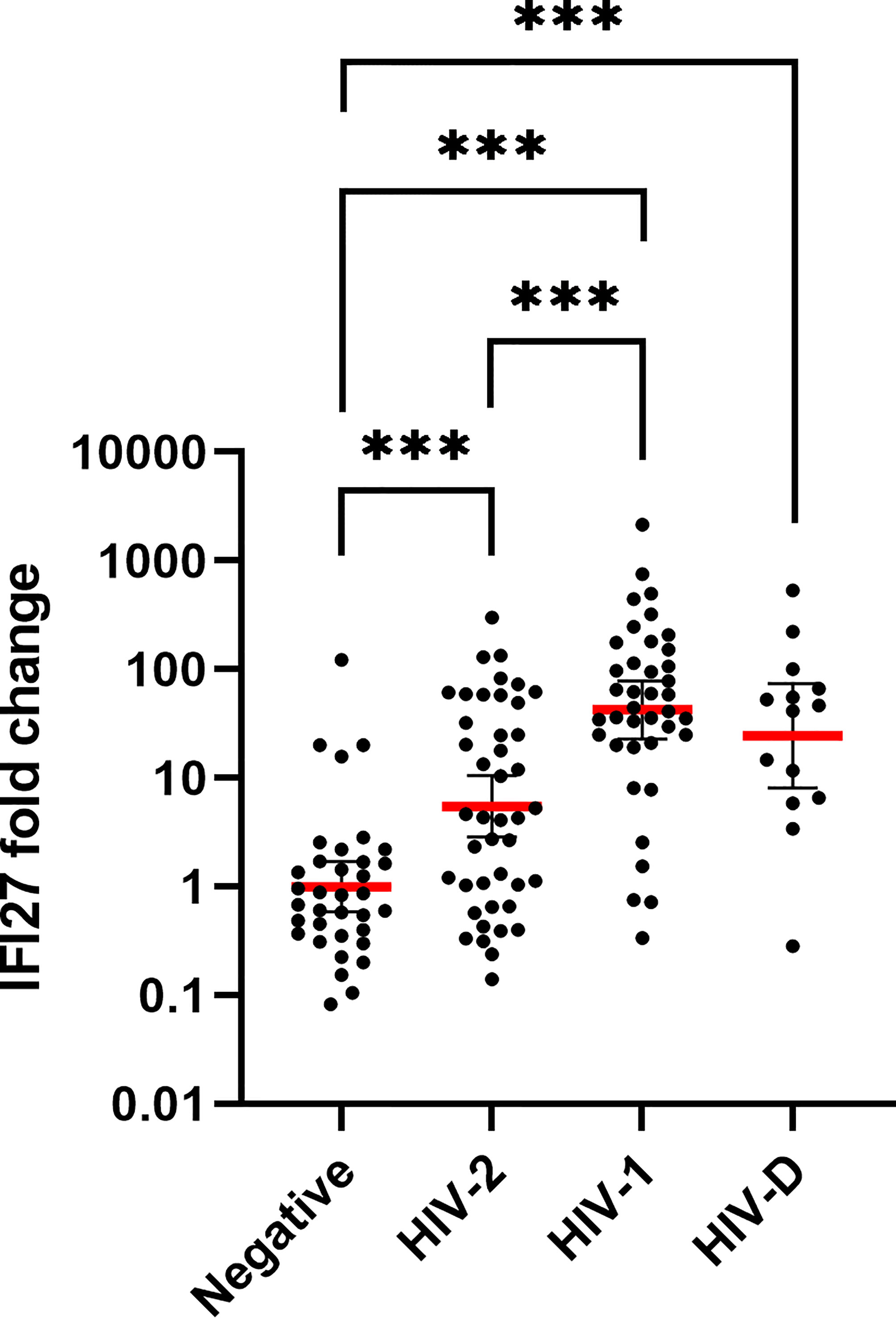
Figure 1 IFI27 gene expression by HIV infection status. The relative IFI27 fold change gene expression, compared to HIV seronegative controls, was quantified and calculated by qPCR and the 2^-ΔΔCT method, in whole blood of HIV seronegative (n = 35), HIV-2 (n = 43), HIV-1 (n = 39) and HIV-D (n = 14) infected individuals. The red line illustrates the geometric mean with confidence interval as error bars. IFI27 expression was log10-transformed prior to statistical comparison between groups using One-way ANOVA with Tukey’s post-hoc test. ***p < 0.001.
IFI27 Is Linked to Disease Severity in Both HIV-1 and HIV-2 Infections
Next, we investigated if IFI27 expression correlated with CD4% and pVL in HIV infected individuals (Table S2). IFI27 was negatively correlated with CD4% in both HIV-1 (r = -0.559, p < 0.001) and HIV-2 infection (r = -0.592, p < 0.001), and positively correlated with pVL in both HIV-1 (r = 0.717, p < 0.001) and HIV-2 (r = 0.676 and p < 0.001) infection. In HIV-D infection, we did not observe any correlations between IFI27 expression and pVL or CD4% (Table S2) and further analyses within this group were not considered.
To determine the relative contribution of HIV type (HIV-1 or HIV-2), sex, age, pVL, and CD4% on IFI27 gene expression, we performed multiple linear regression. The overall regression was significant (p < 0.001, r2 = 0.621) and IFI27 was found to be significantly predicted by HIV type (p = 0.019), CD4% (p = 0.008) and pVL (p < 0.001) when adjusting for age and sex (Table S3). Next, we investigated the interaction between HIV type and CD4%, and pVL, by multiple linear regression. At undetectable pVL levels, the level of IFI27 expression was found to be significantly higher in HIV-2 infected individuals compared with expression levels extrapolated for the HIV-1 infected individuals, both in the crude analysis (p = 0.036, Figure 2A and Table S3), and when adjusting for age and sex (p = 0.015, Table S3). No significant interaction was observed between HIV type and pVL (i.e. the slope of the regression line did not differ between the groups) in either crude (p = 0.148) or adjusted analysis (p = 0.108). For CD4%, there was no significant difference in level of IFI27 expression between the groups in either crude analysis (p = 0.126, Figure 2B and Table S3) or when adjusting for age and sex (p = 0.170, Table S3). Instead, the rate of IFI27 increase with CD4% decline was higher for HIV-2 as compared with HIV-1 infected individuals in crude analysis (p = 0.048, Figure 2B and Table S3), while this difference was lost when adjusting for age and sex (p = 0.094).
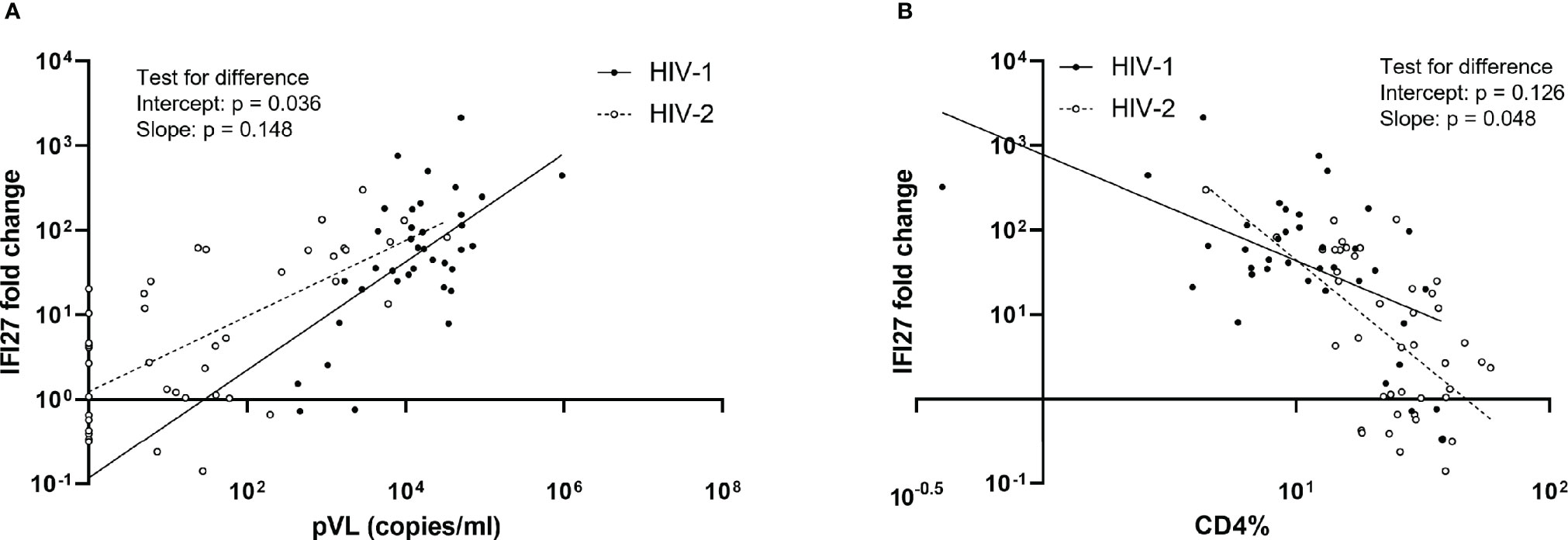
Figure 2 Associations between IFI27 expression and plasma viral load (pVL) (A) and CD4% (B). The relative IFI27 fold change gene expression, compared to HIV seronegative controls, was quantified and calculated by qPCR and the 2^-ΔΔCT method. IFI27 fold change, pVL and CD4% were log10-transformed prior to analysis. P-values for difference between HIV-1 (n = 39 for pVL and 38 for CD4%) and HIV-2 (n = 43 for both pVL and CD4%) in intercept and slope (i.e. the interaction between IFI27 and pVL or CD4%) were calculated by crude multiple linear regression analysis, without adjustment for sex and age. Additional results from these analyses are provided in Table S3.
IFI27 Expression Is Increased in Both Viremic and Aviremic HIV-2 Infected Individuals
To further dissect the relationship between IFI27 expression and disease severity in HIV-2 infected individuals, we stratified this group by level of pVL. We compared the IFI27 expression in HIV-2 aviremic (pVL < 75 copies/ml, n = 30) and HIV-2 viremic individuals (pVL ≥ 75 copies/ml, n = 13) and found that aviremic individuals had 19.8 fold lower levels of IFI27 (p < 0.001, Figure 3A). However, even in the absence of quantifiable pVL, the IFI27 expression was still 2.2 fold higher in aviremic individuals compared to HIV seronegative individuals (n = 35, p = 0.049, Figure 3B). Next, we investigated if the IFI27 expression differed in HIV-2 aviremic individuals, i.e. individuals that naturally controlled their infection, and HIV-1 successfully treated individuals, i.e. individuals with a treatment-induced control of viremia. Here, we did not observe any significant difference in IFI27 expression between natural HIV-2 and treatment-induced HIV-1 control of viremia (p = 0.245, Figure 3C).
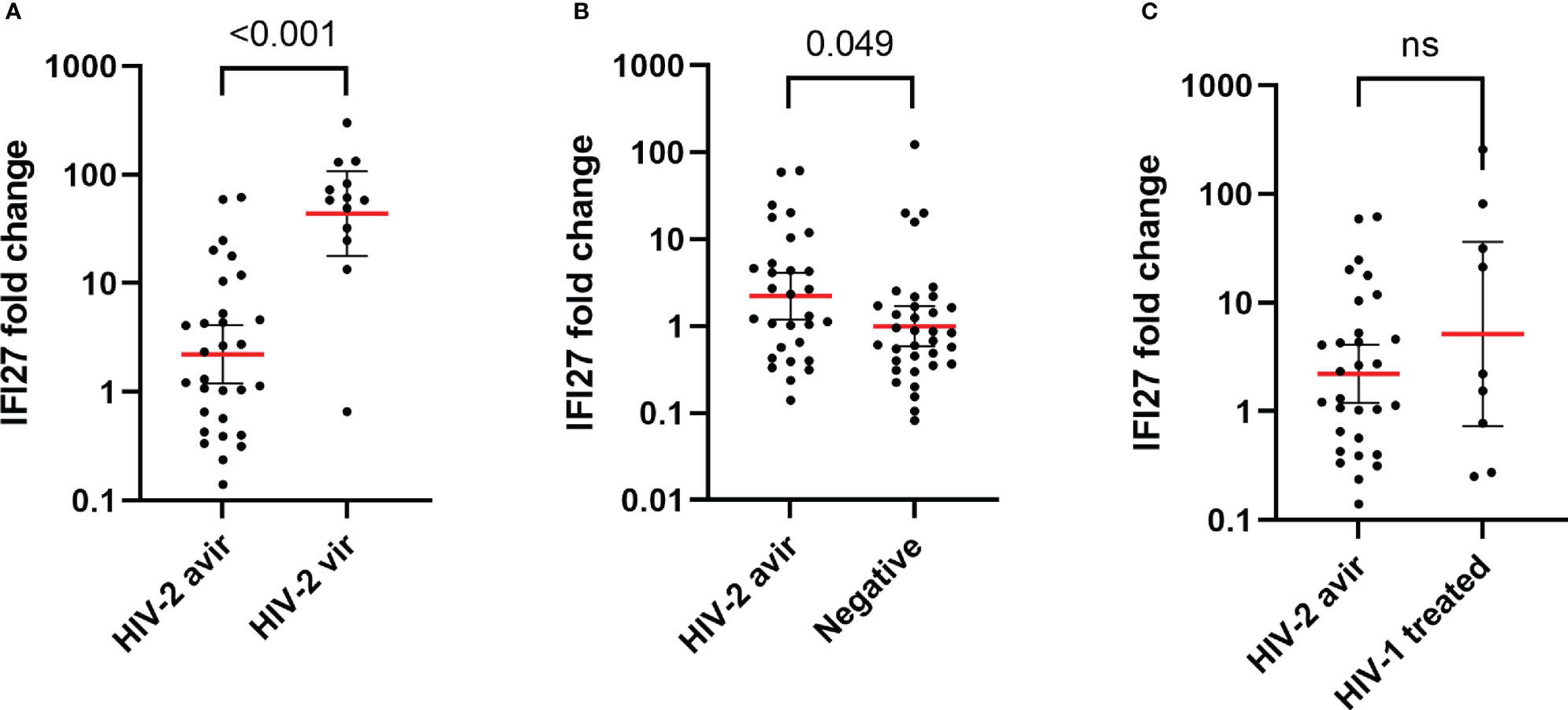
Figure 3 IFI27 fold change expression in HIV-2 aviremic (n = 30) vs HIV-2 viremic (n = 13) (A), HIV seronegative (n = 35) (B), and HIV-1 successfully treated (C) (n = 9) individuals. The relative IFI27 fold change gene expression, compared to HIV seronegative controls, was quantified and calculated by qPCR and the 2^-ΔΔCT method. The red line illustrates the geometric mean with confidence interval as error bars. IFI27 expression was log10-transformed prior to statistical comparison between groups using t-test. ns = non significant.
Finally, we set out to determine if IFI27 expression was correlated with plasma markers previously linked to immune activation and inflammation, viremia and disease progression, i.e. beta-2-microglobulin (b2m), soluble CD14 (sCD14), and IP-10 (also known as CXCL10) (17–19). Results showed that IFI27 was positively correlated with b2m, sCD14 and IP-10, (r = 0.724, p < 0.001; r = 0.710, p < 0.001; and r = 0.663, p < 0.001, respectively) in the studied HIV-2 infected individuals (Figure S2 and Table S2).
Discussion
In this study, comparing gene expression profiles of whole blood sampled from HIV seronegative and HIV-1, HIV-2 or dually HIV-1/HIV-2 infected individuals, IFI27, an IFN-I induced gene, was found to be the most differentially expressed. IFI27 was significantly upregulated in HIV-2, and even more so in HIV-1, chronically infected individuals as compared with HIV seronegative controls. Our exploration of variables associated with IFI27 expression levels, revealed that the IFI27 expression was associated with both pVL and CD4%, with the strongest association to pVL, when adjusting for HIV infection type, sex and age.
Importantly, individuals with aviremic HIV-2 infection also displayed elevated IFI27 expression levels, as compared with HIV seronegative individuals. Thus, despite the lack of viremia above 75 copies/ml, this group of aviremic HIV-2 infected individuals also presented with chronic innate immune activation, in our study reflected by elevated IFI27 expression. This observation is in line with our previous reports on elevated frequencies of CD4+ and CD8+ T-cells expressing markers of immune activation and exhaustion in aviremic HIV-2 infected individuals (10, 12). Moreover, the strong associations between IFI27 expression and plasma inflammation markers, b2m, sCD14 and IP-10, suggest that IFI27 is part of a larger inflammatory network linked to disease in HIV-2 infection. Accordingly, this further supports that although level of viremia contributes significantly to the expression of IFI27, other factors, such as infection duration, may be involved. Indeed, our multivariate analysis showed that, at undetectable pVL levels, the IFI27 expression level was higher in HIV-2 than extrapolated for HIV-1 infected individuals. Furthermore, in aviremic HIV-2 infected individuals we did not observe a difference in IFI27 expression between individuals supgrouped according to undetectable or unquantifiable pVL (data not shown). It has also been reported that HIV-1 viremic nonprogressors have lower levels of ISGs compared with rapid progressors (34). Further, it was recently demonstrated that ART treated HIV-1 infected immunological non-responders, (i.e. individuals under treatment with successfully suppressed pVL but whose CD4+ T-cell count do not recover) had higher IFI27 levels as compared with immunological responders (35).
It has been suggested that the CD4+ T-cell recovery upon treatment is lower in HIV-2 infected individuals compared to HIV-1 infected individuals (36). Clinical follow-up of treatment outcome based on CD4+ T-cell count may therefore not be sufficient in HIV-2 infection. Since the HIV-2 aviremic individuals cannot be monitored by viral load measurements it would be valuable to identify other potential markers of disease progression and treatment outcome. Our data suggests that IFI27 could be such potential suitable marker to further explore in larger cohorts, both during disease progression and treatment.
In a study based on single-cell RNA sequencing, it was recently shown that IFI27, and other ISGs, were upregulated in the acute phase of HIV-1 infection (37). Of note, during SARS-CoV-2 infection, the ISG levels have been shown to be important determinants for the outcome in Covid-19, such that mild acute infections are characterized by a strong and transient expression of ISGs, whereas severe Covid-19 is characterized by a lower and sustained ISG expression (38). Such detailed description of how acute IFI27 levels translates to subsequent HIV-1 or HIV-2 disease progression is yet to be presented. Previous studies have shown that the upregulated expression of IFN-I is maintained during chronic HIV-1 infection (25). However, despite an expected antiviral effect of IFN-I and ISGs, chronic IFN-I expression has a seemingly limited impact on viral load and instead contributes to the systemic immune activation (25). Data from animal studies have shown that expression of various ISGs is upregulated in the chronic phase of nonpathogenic SIV infection, but to a lower level as compared to in pathogenic SIV infections (22–24). Our data demonstrates that HIV-2, an infection that presents as an intermediate between pathogenic and nonpathogenic HIV-1/SIV infections (6, 23), is also associated with upregulated IFI27 expression, but to a lower level than in HIV-1 infection. Thus, we speculate that the inability of the adaptive immune responses to durably control virus replication in HIV infection permits ongoing innate immune activation, in turn driving the disease progression in infected individuals. We have previously shown that HIV-2 delays HIV-1 disease progression in dually infected individuals and that this effect appears to be established during early infection (4, 5). Thus, it would have been interesting to study if IFN-I signaling is linked to this observation. However, this merits a separate investigation and could not be addressed in the current study comprised of a limited number of HIV-D infected individuals sampled during the chronic infection phase.
Both HIV-1 and HIV-2 infection results in the loss and dysregulation of CD4+ T cell (1). However, other target cells, including CD4+ plasmacytoid dendritic cells (pDCs), have also been shown to be dysregulated (39–41). Moreover, in both SIV and HIV infections, pDCs have been shown to be the major source of IFN-alpha (a type I IFN) (23). Still, it has been reported that HIV-2, in contrast to HIV-1, favors pDC to develop into a antigen presenting phenotype, rather than becoming excessive IFN-alpha producers (42). It has also been suggested that HIV-2, via the expression of vpx may contribute to productive infection of DCs via blockade of the SAMHD1 (43, 44). Thus, it is tempting to speculate that the early and productive infection of pDCs in acute HIV-2 infection results in a balanced IFN-alpha response, combined with efficient antigen presentation, subsequently contributing to a less pathogenic chronic infection. Furthermore, gene expression analysis of monocytes from chronically HIV-1 infected individuals identified IFN-alpha as the main driver of the monocyte gene expression profile (45). Interestingly, IFI27 was found to be the most upregulated gene, especially in the HIV-1 infected individuals with high pVL (45). Monocytes in chronic HIV-1 and HIV-2 infection have also been shown to display an activated phenotype (45, 46) and to be desensitized to TLR-stimulation (45). These findings are in line with data previously reported by us showing a defective TLR responsiveness of cells in whole blood from both HIV-1 and HIV-2 infected individuals (47). Taken together these studies point to an important role of myeloid cells in the regulatory network between HIV and IFN-I. However, this network is complex, and the beneficial or detrimental effects of IFN-I signaling is likely determined by numerous factors, including timing, i.e. acute vs chronic infection phase. While both administration of IFN-I and inhibition of its signaling pathways has been therapeutically pursued, the clinical impact has so far been limited (48).
In conclusion, our current findings suggest a scenario in which the level of IFN-I signaling, here reflected by IFI27 expression, is a key feature in determining the pathogenesis of both HIV-1 and HIV-2 infections. We believe that a better understanding of the factors involved in shaping the IFN-I signaling in response to both HIV-1 and HIV-2 infections could unravel new therapeutics and treatment monitoring approaches.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/geo/, GSE200606.
Ethics Statement
The studies involving human participants were reviewed and approved by The National Ethics Committee, Ministry of Public Health in Guinea-Bissau and the Ethical Committee at Lund university. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
AP designed and conducted the RNA extractions and qPCR analyses, interpreted the data, and drafted the manuscript. SV designed, conducted, and summarized the bioinformatics analyses. EJ conducted the Luminex analyses. P-EI contributed substantially to the statistical analyses. JL, FM, and HN coordinated the study cohort. JL, FM, JE, HN, PM, and MJ contributed to the study design and results interpretation, and critically reviewed the manuscript. AB was medically responsible for the study participants. MJ contributed to the study design, interpreted data, revised and critically reviewed the manuscript. AP, PM, and MJ conceptualized the study. All authors read and approved the submitted version.
Funding
This research was funded by grants given to MJ from the Swedish Research Council (grant number 2016-02285, 2019-01439) and The Swedish Fund for Research without Animal Experiments (N2019-0009, F2021-0010). PM was supported by the Swedish Research Council (grant numbers 2019-05235 and 2020-02344) and Österlund Foundation. JE received funding from the Swedish Research Council (grant number 2020-06262).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank SCIBLU Genomics Center at Lund University for technical support concerning the microarray. The listed authors and the members of the Sweden Guinea-Bissau Cohort Research (SWEGUB CORE) group, including Babetida N’Buna, Antonio Biague, Ansu Biai, Cidia Camara, Zacarias Jose da Silva, Joakim Esbjörnsson, Marianne Jansson, Emil Johansson, Sara Karlson, Jacob Lopatko Lindman, Patrik Medstrand, Fredrik Månsson, Hans Norrgren, Angelica A. Palm, Gülsen Özkaya Sahin and Sten Wilhelmson are indebted to the staff of the Police Clinics and the National Public Health Laboratory (LNSP) in Bissau, Guinea-Bissau.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fviro.2022.929053/full#supplementary-material
References
1. Nyamweya S, Hegedus A, Jaye A, Rowland-Jones S, Flanagan KL, Macallan DC. Comparing HIV-1 and HIV-2 Infection: Lessons for Viral Immunopathogenesis. Rev Med Virol (2013) 23(4):221–40. doi: 10.1002/rmv.1739
2. Esbjornsson J, Mansson F, Kvist A, da Silva ZJ, Andersson S, Fenyo EM, et al. Long-Term Follow-Up of HIV-2-Related AIDS and Mortality in Guinea-Bissau: A Prospective Open Cohort Study. Lancet HIV (2019) 6(1):e25–31. doi: 10.1016/S2352-3018(18)30254-6
3. Jaffar S, Wilkins A, Ngom PT, Sabally S, Corrah T, Bangali JE, et al. Rate of Decline of Percentage CD4+ Cells is Faster in HIV-1 Than in HIV-2 Infection. J Acquir Immune Defic Syndr Hum Retrovirol (1997) 16(5):327–32. doi: 10.1097/00042560-199712150-00003
4. Esbjornsson J, Mansson F, Kvist A, Isberg PE, Biague AJ, da Silva ZJ, et al. Increased Survival Among HIV-1 and HIV-2 Dual-Infected Individuals Compared to HIV-1 Single-Infected Individuals. AIDS (2014) 28(7):949–57. doi: 10.1097/QAD.0000000000000101
5. Esbjornsson J, Mansson F, Kvist A, Isberg PE, Nowroozalizadeh S, Biague AJ, et al. Inhibition of HIV-1 Disease Progression by Contemporaneous HIV-2 Infection. N Engl J Med (2012) 367(3):224–32. doi: 10.1056/NEJMoa1113244
6. Esbjornsson J, Jansson M, Jespersen S, Mansson F, Honge BL, Lindman J, et al. HIV-2 as a Model to Identify a Functional HIV Cure. AIDS Res Ther (2019) 16(1):24. doi: 10.1186/s12981-019-0239-x
7. Popper SJ, Sarr AD, Travers KU, Gueye-Ndiaye A, Mboup S, Essex ME, et al. Lower Human Immunodeficiency Virus (HIV) Type 2 Viral Load Reflects the Difference in Pathogenicity of HIV-1 and HIV-2. J Infect Dis (1999) 180(4):1116–21. doi: 10.1086/315010
8. Andersson S, Norrgren H, da Silva Z, Biague A, Bamba S, Kwok S, et al. Plasma Viral Load in HIV-1 and HIV-2 Singly and Dually Infected Individuals in Guinea-Bissau, West Africa: Significantly Lower Plasma Virus Set Point in HIV-2 Infection Than in HIV-1 Infection. Arch Intern Med (2000) 160(21):3286–93. doi: 10.1001/archinte.160.21.3286
9. van der Loeff MF, Larke N, Kaye S, Berry N, Ariyoshi K, Alabi A, et al. Undetectable Plasma Viral Load Predicts Normal Survival in HIV-2-Infected People in a West African Village. Retrovirology (2010) 7:46. doi: 10.1186/1742-4690-7-46
10. Buggert M, Frederiksen J, Lund O, Betts MR, Biague A, Nielsen M, et al. CD4+ T Cells With an Activated and Exhausted Phenotype Distinguish Immunodeficiency During Aviremic HIV-2 Infection. AIDS (2016) 30(16):2415–26. doi: 10.1097/QAD.0000000000001223
11. Sousa AE, Carneiro J, Meier-Schellersheim M, Grossman Z, Victorino RM. CD4 T Cell Depletion is Linked Directly to Immune Activation in the Pathogenesis of HIV-1 and HIV-2 But Only Indirectly to the Viral Load. J Immunol (2002) 169(6):3400–6. doi: 10.4049/jimmunol.169.6.3400
12. Scharf L, Pedersen CB, Johansson E, Lindman J, Olsen LR, Buggert M, et al. Inverted CD8 T-Cell Exhaustion and Co-Stimulation Marker Balance Differentiate Aviremic HIV-2-Infected From Seronegative Individuals. Front Immunol (2021) 12:744530. doi: 10.3389/fimmu.2021.744530
13. Klatt NR, Chomont N, Douek DC, Deeks SG. Immune Activation and HIV Persistence: Implications for Curative Approaches to HIV Infection. Immunol Rev (2013) 254(1):326–42. doi: 10.1111/imr.12065
14. Nowroozalizadeh S, Mansson F, da Silva Z, Repits J, Dabo B, Pereira C, et al. Microbial Translocation Correlates With the Severity of Both HIV-1 and HIV-2 Infections. J Infect Dis (2010) 201(8):1150–4. doi: 10.1086/651430
15. Bachle SM, Malone DF, Buggert M, Karlsson AC, Isberg PE, Biague AJ, et al. Elevated Levels of Invariant Natural Killer T-Cell and Natural Killer Cell Activation Correlate With Disease Progression in HIV-1 and HIV-2 Infections. AIDS (2016) 30(11):1713–22. doi: 10.1097/QAD.0000000000001147
16. Leligdowicz A, Feldmann J, Jaye A, Cotten M, Dong T, McMichael A, et al. Direct Relationship Between Virus Load and Systemic Immune Activation in HIV-2 Infection. J Infect Dis (2010) 201(1):114–22. doi: 10.1086/648733
17. Nyamweya S, Townend J, Zaman A, Steele SJ, Jeffries D, Rowland-Jones S, et al. Are Plasma Biomarkers of Immune Activation Predictive of HIV Progression: A Longitudinal Comparison and Analyses in HIV-1 and HIV-2 Infections? PloS One (2012) 7(9):e44411. doi: 10.1371/journal.pone.0044411
18. Thiebaut R, Charpentier C, Damond F, Taieb A, Antoine R, Capeau J, et al. Association of Soluble CD14 and Inflammatory Biomarkers With HIV-2 Disease Progression. Clin Infect Dis (2012) 55(10):1417–25. doi: 10.1093/cid/cis708
19. Lei J, Yin X, Shang H, Jiang Y. IP-10 is Highly Involved in HIV Infection. Cytokine (2019) 115:97–103. doi: 10.1016/j.cyto.2018.11.018
20. Ivashkiv LB, Donlin LT. Regulation of Type I Interferon Responses. Nat Rev Immunol (2014) 14(1):36–49. doi: 10.1038/nri3581
21. McNab F, Mayer-Barber K, Sher A, Wack A, O'Garra A. Type I Interferons in Infectious Disease. Nat Rev Immunol (2015) 15(2):87–103. doi: 10.1038/nri3787
22. Bosinger SE, Li Q, Gordon SN, Klatt NR, Duan L, Xu L, et al. Global Genomic Analysis Reveals Rapid Control of a Robust Innate Response in SIV-Infected Sooty Mangabeys. J Clin Invest (2009) 119(12):3556–72. doi: 10.1172/JCI40115
23. Klatt NR, Silvestri G, Hirsch V. Nonpathogenic Simian Immunodeficiency Virus Infections. Cold Spring Harbor Perspect Med (2012) 2(1):a007153. doi: 10.1101/cshperspect.a007153
24. Jacquelin B, Mayau V, Targat B, Liovat AS, Kunkel D, Petitjean G, et al. Nonpathogenic SIV Infection of African Green Monkeys Induces a Strong But Rapidly Controlled Type I IFN Response. J Clin Invest (2009) 119(12):3544–55. doi: 10.1172/JCI40093
25. Bosinger SE, Utay NS. Type I Interferon: Understanding Its Role in HIV Pathogenesis and Therapy. Curr HIV/AIDS Rep (2015) 12(1):41–53. doi: 10.1007/s11904-014-0244-6
26. Hirsch VM, Olmsted RA, Murphey-Corb M, Purcell RH, Johnson PR. An African Primate Lentivirus (SIVsm) Closely Related to HIV-2. Nature (1989) 339(6223):389–92. doi: 10.1038/339389a0
27. Norrgren H, Andersson S, Naucler A, Dias F, Johansson I, Biberfeld G. HIV-1, HIV-2, HTLV-I/II and Treponema Pallidum Infections: Incidence, Prevalence, and HIV-2-Associated Mortality in an Occupational Cohort in Guinea-Bissau. J Acquir Immune Defic Syndr Hum Retrovirol (1995) 9(4):422–8. doi: 10.1097/00042560-199508000-00014
28. Mansson F, Biague A, da Silva ZJ, Dias F, Nilsson LA, Andersson S, et al. Prevalence and Incidence of HIV-1 and HIV-2 Before, During and After a Civil War in an Occupational Cohort in Guinea-Bissau, West Africa. AIDS (2009) 23(12):1575–82. doi: 10.1097/QAD.0b013e32832cedfb
29. Organization WH. Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection (2016) (2016). Available at: https://apps.who.int/iris/handle/10665/208825.
30. Anglaret X, Diagbouga S, Mortier E, Meda N, Verge-Valette V, Sylla-Koko F, et al. CD4+ T-Lymphocyte Counts in HIV Infection: Are European Standards Applicable to African Patients? J Acquir Immune Defic Syndr Hum Retrovirol (1997) 14(4):361–7. doi: 10.1097/00042560-199704010-00009
31. Ozkaya Sahin G, Mansson F, Palm AA, Vincic E, da Silva Z, Medstrand P, et al. Frequent Intratype Neutralization by Plasma Immunoglobulin a Identified in HIV Type 2 Infection. AIDS Res Hum Retroviruses (2013) 29(3):470–8. doi: 10.1089/aid.2012.0219
32. Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U, et al. Exploration, Normalization, and Summaries of High Density Oligonucleotide Array Probe Level Data. Biostatistics (2003) 4(2):249–64. doi: 10.1093/biostatistics/4.2.249
33. Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (2001) 25(4):402–8. doi: 10.1006/meth.2001.1262
34. Rotger M, Dalmau J, Rauch A, McLaren P, Bosinger SE, Martinez R, et al. Comparative Transcriptomics of Extreme Phenotypes of Human HIV-1 Infection and SIV Infection in Sooty Mangabey and Rhesus Macaque. J Clin Invest (2011) 121(6):2391–400. doi: 10.1172/JCI45235
35. Liu X, Lin L, Lu L, Li X, Han Y, Qiu Z, et al. Comparative Transcriptional Analysis Identified Characteristic Genes and Patterns in HIV-Infected Immunological Non-Responders. Front Immunol (2022) 13:807890. doi: 10.3389/fimmu.2022.807890
36. Matheron S, Damond F, Benard A, Taieb A, Campa P, Peytavin G, et al. CD4 Cell Recovery in Treated HIV-2-Infected Adults is Lower Than Expected: Results From the French ANRS CO5 HIV-2 Cohort. AIDS (2006) 20(3):459–62. doi: 10.1097/01.aids.0000199829.57112.2f
37. Kazer SW, Aicher TP, Muema DM, Carroll SL, Ordovas-Montanes J, Miao VN, et al. Integrated Single-Cell Analysis of Multicellular Immune Dynamics During Hyperacute HIV-1 Infection. Nat Med (2020) 26(4):511–8. doi: 10.1038/s41591-020-0799-2
38. Ziegler CGK, Miao VN, Owings AH, Navia AW, Tang Y, Bromley JD, et al. Impaired Local Intrinsic Immunity to SARS-CoV-2 Infection in Severe COVID-19. Cell (2021) 184(18):4713–33 e22. doi: 10.1016/j.cell.2021.07.023
39. Cavaleiro R, Baptista AP, Soares RS, Tendeiro R, Foxall RB, Gomes P, et al. Major Depletion of Plasmacytoid Dendritic Cells in HIV-2 Infection, an Attenuated Form of HIV Disease. PloS pathogens (2009) 5(11):e1000667. doi: 10.1371/journal.ppat.1000667
40. Feldman S, Stein D, Amrute S, Denny T, Garcia Z, Kloser P, et al. Decreased Interferon-Alpha Production in HIV-Infected Patients Correlates With Numerical and Functional Deficiencies in Circulating Type 2 Dendritic Cell Precursors. Clin Immunol (2001) 101(2):201–10. doi: 10.1006/clim.2001.5111
41. Pacanowski J, Kahi S, Baillet M, Lebon P, Deveau C, Goujard C, et al. Reduced Blood CD123+ (Lymphoid) and CD11c+ (Myeloid) Dendritic Cell Numbers in Primary HIV-1 Infection. Blood (2001) 98(10):3016–21. doi: 10.1182/blood.V98.10.3016
42. Royle CM, Graham DR, Sharma S, Fuchs D, Boasso A. HIV-1 and HIV-2 Differentially Mature Plasmacytoid Dendritic Cells Into IFN-Producing Cells or APCs. J Immunol (2014) 193(7):3538–48. doi: 10.4049/jimmunol.1400860
43. Mohamed A, Bakir T, Al-Hawel H, Al-Sharif I, Bakheet R, Kouser L, et al. HIV-2 Vpx Neutralizes Host Restriction Factor SAMHD1 to Promote Viral Pathogenesis. Sci Rep (2021) 11(1):20984. doi: 10.1038/s41598-021-00415-2
44. Goujon C, Arfi V, Pertel T, Luban J, Lienard J, Rigal D, et al. Characterization of Simian Immunodeficiency Virus SIVSM/human Immunodeficiency Virus Type 2 Vpx Function in Human Myeloid Cells. J Virol (2008) 82(24):12335–45. doi: 10.1128/JVI.01181-08
45. Rempel H, Sun B, Calosing C, Pillai SK, Pulliam L. Interferon-Alpha Drives Monocyte Gene Expression in Chronic Unsuppressed HIV-1 Infection. AIDS (2010) 24(10):1415–23. doi: 10.1097/QAD.0b013e32833ac623
46. Cavaleiro R, Tendeiro R, Foxall RB, Soares RS, Baptista AP, Gomes P, et al. Monocyte and Myeloid Dendritic Cell Activation Occurs Throughout HIV Type 2 Infection, an Attenuated Form of HIV Disease. J Infect Dis (2013) 207(11):1730–42. doi: 10.1093/infdis/jit085
47. Nowroozalizadeh S, Mansson F, da Silva Z, Repits J, Dabo B, Pereira C, et al. Studies on Toll-Like Receptor Stimuli Responsiveness in HIV-1 and HIV-2 Infections. Cytokine (2009) 46(3):325–31. doi: 10.1016/j.cyto.2009.03.003
Keywords: HIV-1, HIV-2, interferon-stimulated gene (ISG), viremia, chronic infection, disease severity, interferon alpha-inducible protein 27 (IFI27)
Citation: Palm AA, Veerla S, Lindman J, Isberg P-E, Johansson E, Biague A, Månsson F, Norrgren H, Esbjörnsson J, Medstrand P and Jansson M (2022) Interferon Alpha-Inducible Protein 27 Expression Is Linked to Disease Severity in Chronic Infection of Both HIV-1 and HIV-2. Front.Virol. 2:929053. doi: 10.3389/fviro.2022.929053
Received: 26 April 2022; Accepted: 19 May 2022;
Published: 13 June 2022.
Edited by:
Nuno Taveira, Instituto Universitário Egas Moniz (IUEM), PortugalCopyright © 2022 Palm, Veerla, Lindman, Isberg, Johansson, Biague, Månsson, Norrgren, Esbjörnsson, Medstrand and Jansson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angelica A. Palm, YW5nZWxpY2EucGFsbUBtZWQubHUuc2U=