
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 31 March 2025
Sec. Parasitology
Volume 12 - 2025 | https://doi.org/10.3389/fvets.2025.1568298
This article is part of the Research Topic Epidemiology, Diagnosis, and Treatment of Toxoplasma gondii Infection in a One Health Context View all articles
 Mubashra Salim1†
Mubashra Salim1† Asmat Ullah Khan2†Alaudin Khan Niazi1Amna Aqdas3
Asmat Ullah Khan2†Alaudin Khan Niazi1Amna Aqdas3 Turki M. Dawoud4Muhammad Usman5Hira Muqaddas6Shakir Ullah7Hanène Belkahia8Adil Khan9*
Turki M. Dawoud4Muhammad Usman5Hira Muqaddas6Shakir Ullah7Hanène Belkahia8Adil Khan9* Mourad Ben Said8,10*
Mourad Ben Said8,10* Furhan Iqbal1*
Furhan Iqbal1*Introduction: Despite Pakistan’s rich amphibian fauna, frog and toad species remain largely unexplored regarding blood-borne parasites.
Methods: This study aims to investigate potential infections by toxoplasma gondii and Hepatozoon spp. in four amphibian species (n = 223) collected from various regions in Punjab and Khyber Pakhtunkhwa provinces.
Results: Molecular analyses revealed that 17 out of 223 amphibians (7.6%) were infected with Hepatozoon spp., with the highest infection rates found in Bufo olivaceous (20.0%), followed by Bufo stomaticus (9.3%) and Hoplobatrachus tigerinus (5.05%). DNA sequencing and BLAST analysis confirmed the presence of Hepatozoon ixoxo and Lankesterella sp. phylogenetic analysis of both pathogens demonstrated genetic diversity among the Pakistani isolates, clustering with isolates from birds, amphibians, and reptiles worldwide. To the best of our knowledge, this is first ever report globally where we are documenting that 4.5% of the screened Pakistani anurans, including frogs (H. tigerinus, 8.1%) and toads (B. stomaticus, 1.9%), were infected with toxoplasma gondii-like strains. Parasite prevalence varied between sampling sites and amphibian species. This study represents the first report from Pakistan documenting the prevalence and genetic characterization of Hepatozoon sp., Lankesterella sp., and T. Gondii-like strains among amphibians. We recommend conducting similar large-scale studies across various geo-climatic regions of Pakistan to further explore the epidemiology, genetic diversity, host–parasite interactions, and effective control of these pathogens among local frog and toad species. Identifying genetically related T. Gondii strains in unexpected host animals, such as amphibians, has been crucial for contributing to the elucidation of the parasite’s evolutionary history.
Amphibians, a diverse group of vertebrates, require water or moist environments for survival (1). Globally, there are currently 7,947 amphibian species, with 7,013 species being frogs and toads (Anura) found in various aquatic and terrestrial habitats, excluding estuarine and marine environments (2). Anurans are notable for their feeding habits, medicinal value, and their economic, esthetic and cultural significance (3). They are also consumed as food internationally (4).
In Pakistan, information on anurans distribution is limited due to the lack of attention from the scientific community (5). The country’s amphibian fauna is solely represented by the order Anura, comprising 21 species, 12 genera, and 4 families: Ranidae, Microhylidae, Bufonidae, and Megophryidae (3). Amphibians in Pakistan are reported from the Indus valley in the west bank of the river, runnel of Himalayan North region, water channel and sub-mountainous regions of western Baluchistan (6). Despite their diversity, anurans are experiencing the highest rate of population decline among vertebrates (7). Factors contributing to this decline include anthropogenic activities such as deforestation, industrialization, urbanization, mechanized agriculture, pesticide use, road causalities and vector borne diseases (3).
Parasites can negatively affect their hosts, leading to disturbed physiology and decreased reproductive success (8). Hepatozoon species, belonging to the family Hepatozoidae, are apicomplexan protozoans that target the red and white blood cells of various hosts, including reptiles, amphibians, and mammals (9, 10). These species have also been reported in several invertebrate species that primarily act as vectors, transmitting the parasites from invertebrates to vertebrates and among different vertebrate species (11, 12). A common mode of transmission for Hepatozoon spp. is through the ingestion of an infected invertebrate or intermediate prey, leading to the release of developmental stages in endothelial cells, hepatocytes, and other visceral organs of a wide variety of vertebrate hosts (13). Toxoplasma gondii is an intracellular protozoan that infects nearly all warm-blooded animals, both domestic and wild (14, 15). Its prevalence in wildlife is closely linked to the presence of felids, the definitive hosts, as the parasite’s oocysts are excreted in feces and subsequently ingested by new hosts (16, 17). Common symptoms of toxoplasmosis in animals include fever, loss of appetite, and lethargy, which can vary depending on whether the infection is acute or chronic (18).
In Pakistan, there are limited reports documenting amphibian diversity, and information regarding the endo-parasites infecting frogs and toads is scarce (3, 19, 20). To fill this knowledge gap, blood samples were collected from anuran species across six districts in Punjab and Khyber Pakhtunkhwa provinces and screened for the DNA of Haemogregarines and T. gondii and related strains complex using PCR and sequencing methods. Additionally, risk factors associated with infection were evaluated. The findings offer valuable insights into the epidemiology of these parasites in Pakistan, with significant implications for wildlife conservation and public health.
Samples were collected from three regions in Punjab [Layyah, Multan and Sargodha (Bhera)] and three regions in Khyber Pakhtunkhwa (KPK) [Upper Dir (Sheringal), Mardan and Buner] in Pakistan. The survey took place during the summer and rainy months (July to October) over two consecutive years (2022–2023). The study areas in the two provinces were geographically and climatically distinct, allowing for the observation of geo-climatic differences on the prevalence of studied parasites (Figure 1).
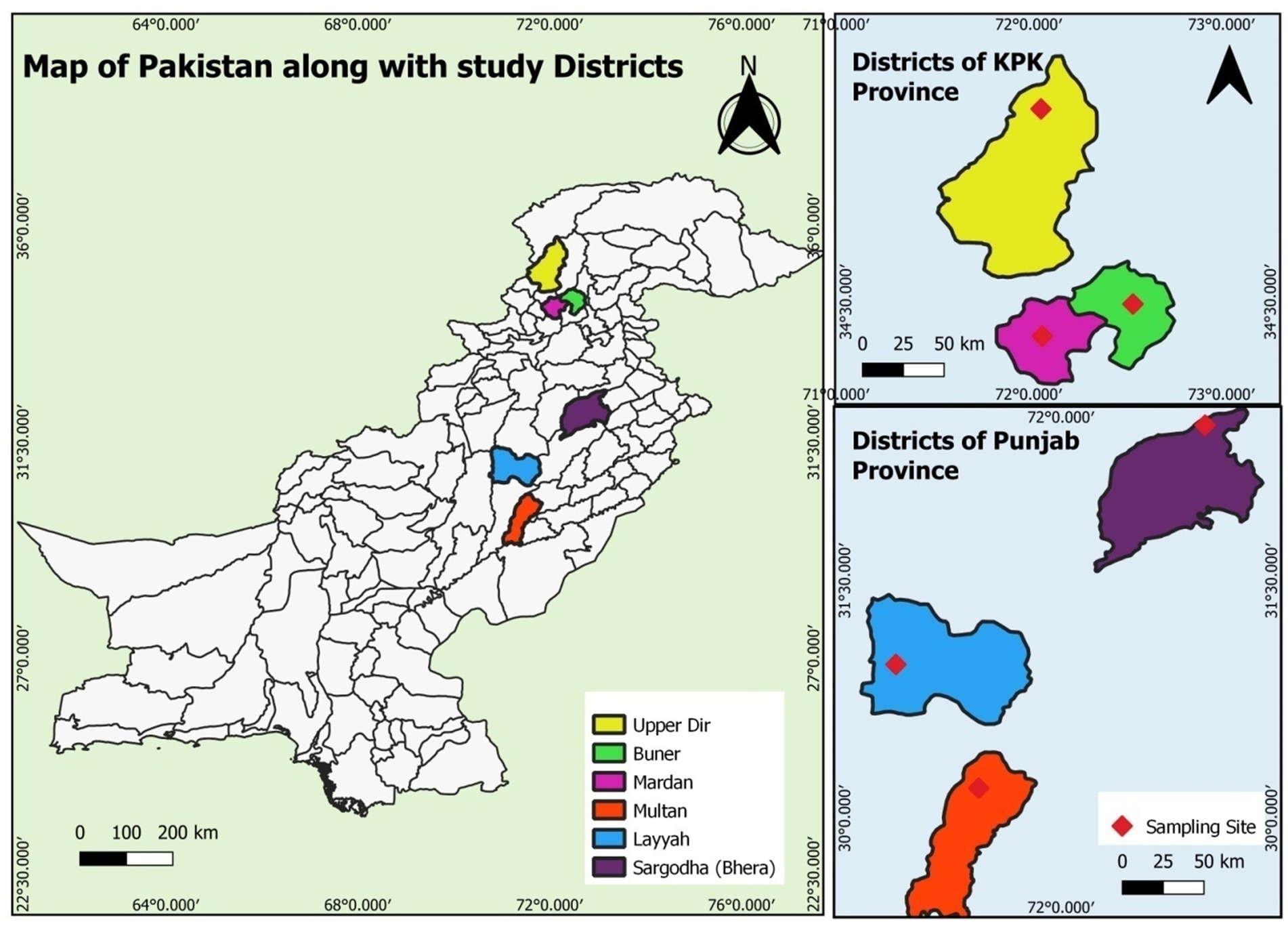
Figure 1. Magnified map of Punjab and KPK showing different sampling sites (Multan, Layyah, Sargodha, Upper Dir, Buner, and Mardan).
A total of 223 amphibians, representing four different species of toads and frogs, were collected from three regions each in Punjab (n = 126) and KPK (n = 97). Samples were collected during the summer and rainy months as during autumn and winters these animals hibernate being cold blooded animals. Frogs and toads were caught by hand or using nets and transported in clean, ventilated plastic containers to the Molecular Parasitology Laboratory at the Institute of Zoology, Bahauddin Zakariya University, Multan. The specimens were identified following the methodologies of Ingerm and Stuebing (21) and Frost et al. (1). Briefly, frogs can be differentiated as they have long legs, longer than their head and body, used for hopping. Toads, on the other hand, have much shorter legs and prefer to crawl. Frogs have smooth, somewhat slimy skin while toads have dry, warty skin (6).
A questionnaire was completed for each animal to gather information about each animal in order to calculate the prevalence of studied parasites according to each risk factor. The captured frogs and toads were anesthetized and blood samples (> 0.1 mL) were collected from each animal via cardiac puncture with a disposable syringe. The blood was transferred into screw capped tubes containing EDTA for DNA extraction.
Genomic DNA was extracted from the blood of each frog and toad using the blood genomic DNA extraction kit (Solarbio, China) following the manufacturer’s instructions.
The extracted DNA samples were analyzed for the presence of Hepatozoon spp., by targeting their 18S rRNA gene, using the forward primer HepF300 5′-GTT TCT GAC CTA TCA GCT TTC GAC G-3′ and reverse primer Hep900 5′-C AAA TCT AAG AAT TTC ACC TCT GAC-3′, following the PCR protocol reported by Ujvari et al. (22). The primers amplified a 600 base pair fragment from 18S rRNA gene that is shared by variety of species in addition to Hepatozoon spp. (23). For the detection of T. gondii and strains genetically related, the ITS-1 gene was targeted using only the internal primers of the nested PCR protocol (ITS_fw 5′-GAT TTG CAT TCA AGA AGC GTG ATA GTA T − 3′ and ITS_rv 5′-AGT TTA GGA AGC AAT CTG AAA GCA CAT C -3′) as reported by Zintl et al. (24).
DNA amplification was carried out in a DNA thermal cycler (Gene Amp® PCR system 2,700 Applied Biosystems Inc., United Kingdom). During each reaction, distilled water was used as a negative control, while DNA from positive animals (for screened parasites available at our laboratory from previous studies) was used as a positive control.
Amplified PCR products of all parasites were sequenced by a commercial company (First base, Malaysia), and the resultant partial gene sequences were submitted to NCBI GenBank. The newly generated sequences targeting the 18S SSU gene (600 bp) were compared with other isolates from Lankesterella and Hepatozoon spp. available on GenBank. Similarly, the ITS-1 partial sequences (300 bp) of T. gondii-like strains were compared with those isolated from T. gondii species previously deposited in GenBank. The sequences were aligned using Geneious version 7.1.3 (25) with the MUSCLE algorithm1 and default settings, including related sequences identified through a Blastn search.
For phylogenetic analyses, Bayesian inference (BI) and Maximum Likelihood (ML) methods were performed. The program JModeltest v.2.1.10 (26) was used for the ML method to identify the best evolutionary model. Based on the Akaike Information Criterion (AIC), the transitional model with a discrete Gamma distribution (TVM + G) was chosen (55). The analysis was inferred using PhyML (27) with 1,000 bootstrap replicates (>50%). For BI analysis, MrBayes was implemented using the computational resource CIPRES (28). The best BIC score indicated the general time reversible model (GTR + I + Γ) (29). In addition, the Markov chain Monte Carlo (MCMC) algorithm was run for 10,000,000 generations, sampling one tree every 1,000 generations. For burn-in, the first 25% of generations were discarded, and the consensus tree was estimated using the remaining trees. Bayesian posterior probabilities (BPP) cut-off considered was >50%. The BI and ML constructed trees were edited in FigTree v1.4 (30). Adelina dimidiata Schneider, 1875, Adelina grylli (DQ096835 and DQ096836) and Klossia helicina Schneider, 1875 (HQ224955) from the suborder Adeleorina were used as outgroups during the phylogenetic analysis of Hepatozoon. Dactylosoma piperis (MW264134), Dactylosoma ranarum (HQ224958), and Dactylosoma kermiti (MN839798) (Apicomplexa, Dactylosomatidae) were used as outgroups for the Lankesterella spp. genetic diversity analysis. While Hammondia (AF159240 and KJ394594) and Sarcocystis species (KF601312, KM657771, EF079887, MG493471, AY082645, AY082647 and AF098245) were used as outgroups for T. gondii and related strains’ analysis.
The statistical analysis of data was performed using Minitab (Minitab, Pennsylvania, United States). A p-value of less than 0.05 was considered statistically significant. The PCR-based pathogen prevalence between various sampling sites and anuran species was compared using one way ANOVA. The association between the presence of each pathogen and the studied epidemiological factors were assessed using contingency table analysis with Fisher’s exact test (for 2 × 2 tables).
The captured amphibians included one frog species [Hoplobatrachus tigerinus (N = 99)] and three toad species [Bufo stomaticus n = 108), Bufo olivaceous (n = 10) and Bufo melanostictus n = 6)].
A total of four anuran species were captured and identified during this investigation including one frog (H. tigerinus) and three toad (B. stomaticus, B. olivaceous, and B. melanostictus) species. Polymerase chain reaction amplified a 600 base pairs fragment from the 18S rRNA gene of Hepatozoon spp. in 17 out of 223 (7.6%) frog and toad blood samples collected from two provinces in Pakistan (Table 1).

Table 1. Haemogregarines prevalence among frog and toad species in overall and according to the six sampling districts.
BLAST analysis of the amplified parasite sequences confirmed that our Pakistani anurans were infected with both Hepatozoon spp. and Lankesterella spp., as anticipated based on the generalized primers capable of amplifying the 18S rRNA gene from various organisms. The analysis revealed that our Hepatozoon sp. sequence (PP481405) is genetically closest to Hepatozoon ixoxo, showing 99.77% identity with an isolated sequence (KP119772). Similarly, our Lankesterella sp. sequence (F11, PP476386) is genetically similar to Lankesterella sp. (KX453649) with 96.22% identity. To further explore genetic diversity, we analyzed Hepatozoon spp. and Lankesterella spp. separately. The haplotype PP481405 for Hepatozoon spp. clustered with sequences from amphibians and reptiles in South Africa (MG519501, MG519504, KP119772, KP119773, MG041591, MG041598, and MG041600), Canada (JN181157, HQ224960, and HQ224962), Brazil (MW591556), and the United States (AF176837) (Figure 2). For Lankesterella spp., the two haplotypes identified (PP544154 and PP541577) were genetically similar and formed a monophyletic cluster with 100% bootstrap support. Our third haplotype (PP476386) was clustered separately. These haplotypes also displayed similarities with global Lankesterella spp. sequences from birds, amphibians, and reptiles (Figure 3). Notably, Pakistani haplotypes PP544154 and PP541577 clustered with Lankesterella sp. from Spain (KJ131417) and Yemen (MW076442), while isolate PP476386 clustered with Lankesterella sp. from Oman (KX453649) (Figure 3).
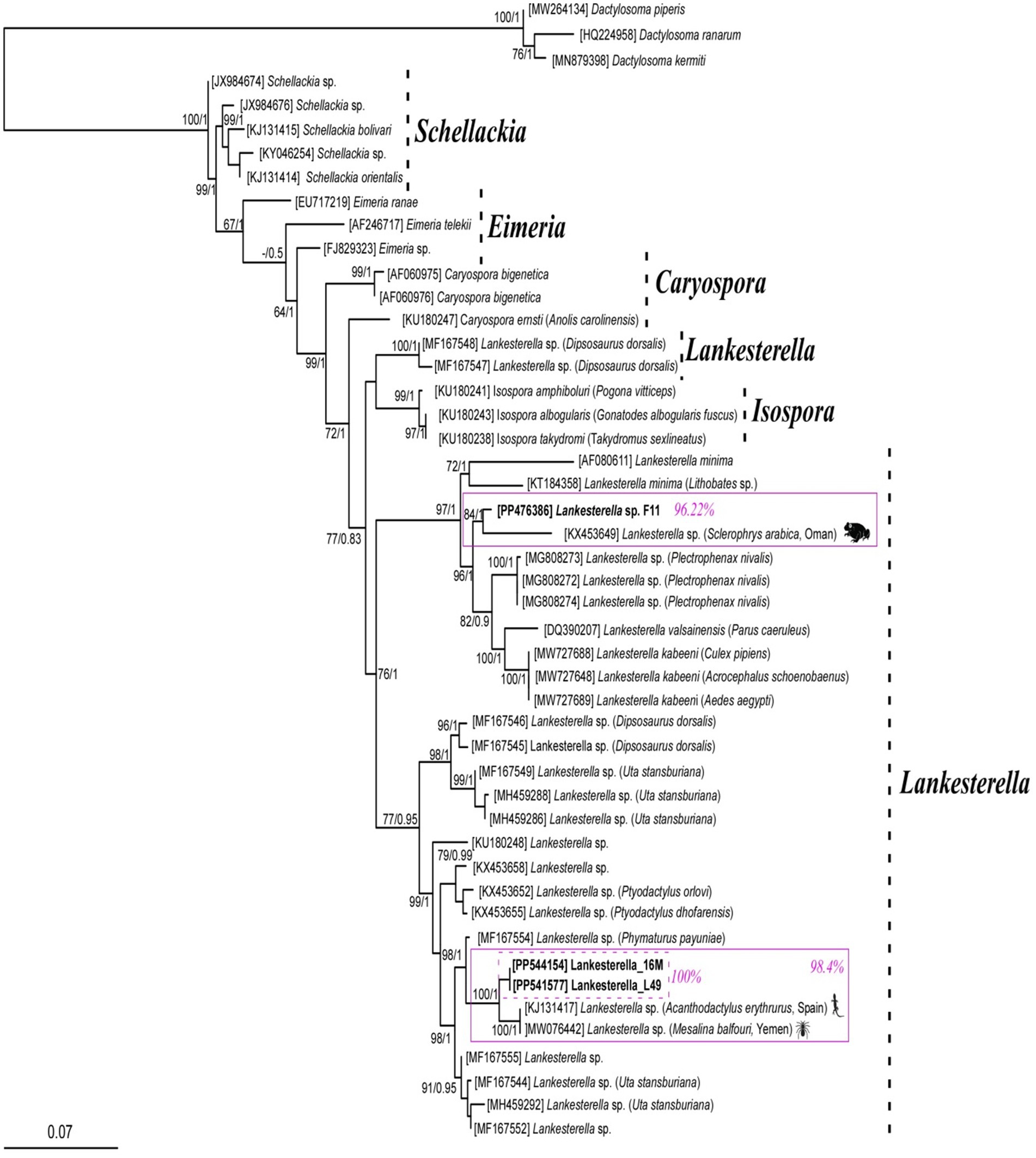
Figure 2. Phylogenetic tree of Hepatozoon spp. based on the partial 18S rRNA gene sequences. New sequence of Hepatozoon sp. obtained in this study (PP481405) is highlighted in bold. Scale bar represents 0.2 substitutions per nucleotide positions. Bootstrap values are shown as numbers on the nodes.
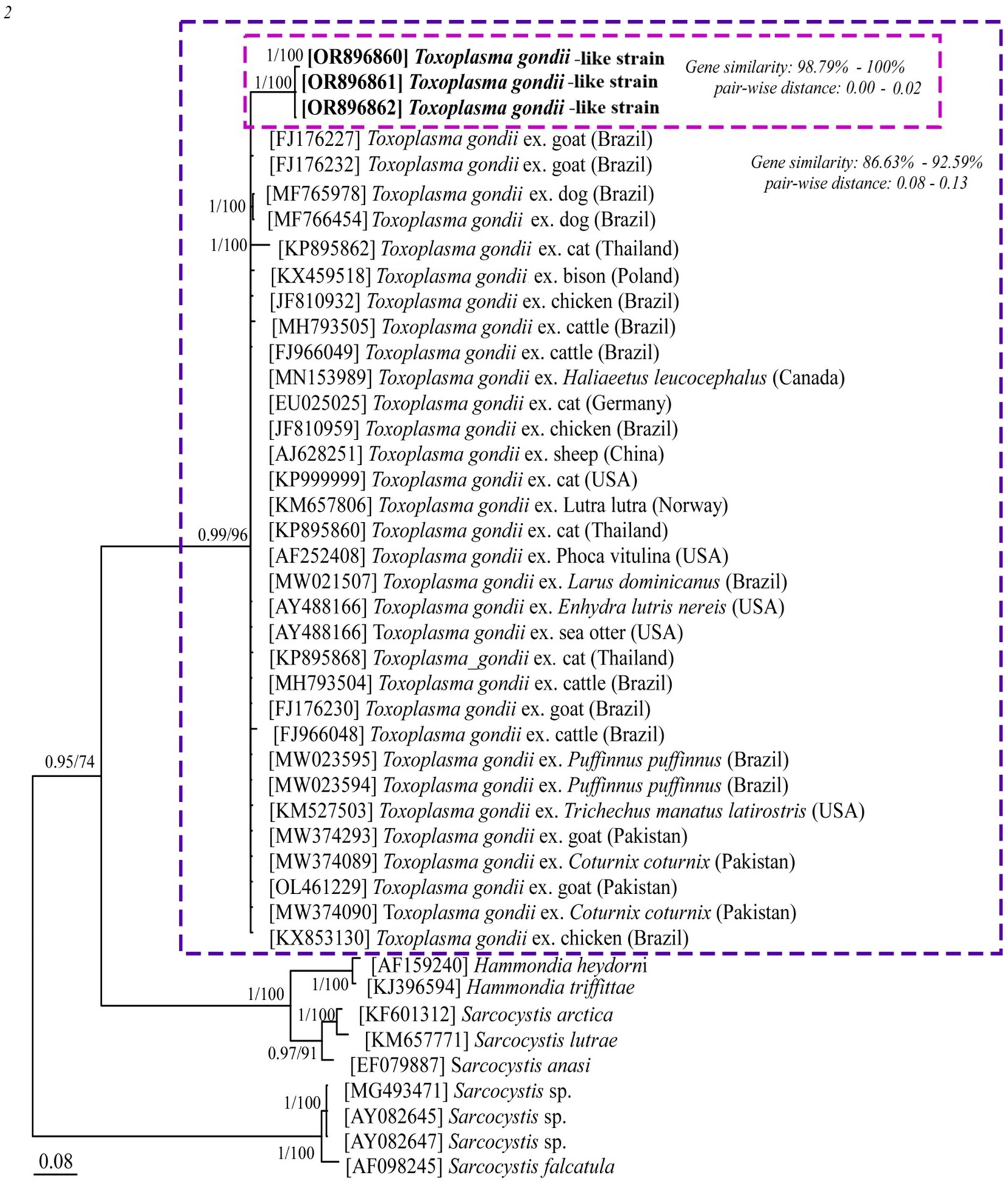
Figure 3. Phylogenetic tree of Lankesterella sp. based on the partial 18S rRNA gene sequences. The three new sequences of Lankesterella sp. obtained in this study (PP476386, PP544154 and PP541577) are highlighted in bold and presented in box. Scale bar represents 0.07 substitutions per nucleotide position. Bootstrap values are shown as numbers on the nodes.
For Lankesterella spp., the two haplotypes revealed in this study (PP544154 and PP541577) were genetically similar and clustered together with a monophyletic branch with same number of substitutions at 100% bootstrap support values. Our third haplotype (PP476386) was clustered separately from the other two haplotypes. These haplotypes also showed similarities with Lankesterella spp. sequences reported from birds, amphibian, and reptiles globally (Figure 3). Pakistani haplotypes PP544154 and PP541577 clustered with Lankesterella sp. reported from Spain (KJ131417) and Yemen (MW076442). The remaining Pakistani isolate PP476386 clustered with Lankesterella sp. from Oman (KX453649) (Figure 3).
When comparing the prevalence of Hepatozoon spp. among the anurans (frogs and toads) captured from various sampling sites, one-way ANOVA results indicated that prevalence of this parasite was not restricted to a particular sampling site (p = 0.260) (Table 1). Fisher’s exact test showed that Hepatozoon spp. infection was not associated with the sex of H. tigerinus captured from six districts in Pakistan (p = 0.631) (Table 2). A similar trend was observed for the three toad species included in this study, with Fisher’s exact test indicating that Hepatozoon spp. infection was not related to the sex of B. stomaticus (p = 0.589) and B. olivaceous (p = 0.617). This analysis was not possible for B. melanostictus as none of the toad belonging to this species was found infected with Hepatozoon spp. (Table 2).
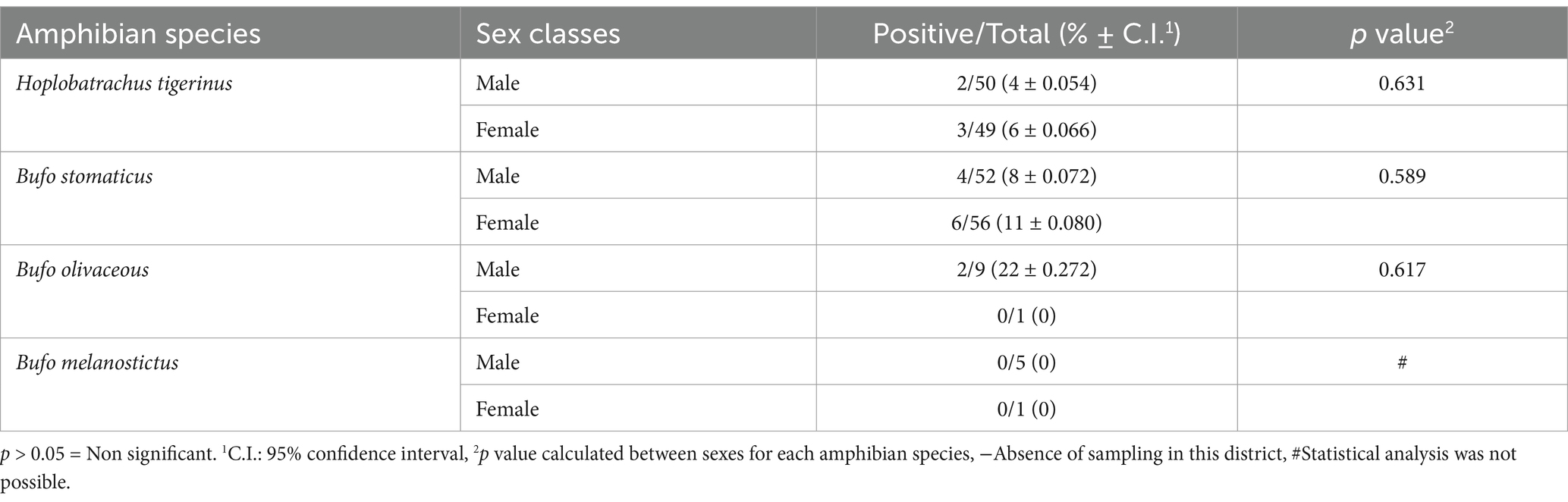
Table 2. Prevalence rates of haemogregarines in amphibian species by sex.
Polymerase chain reaction amplified a 300 base pair fragment specific to the ITS-1 marker of T. gondii-like strains in 10 out of 223 (4.5%) frog and toad blood samples collected from three sampling districts (Layyah, Sargodha and Multan) in Punjab and three sampling districts (Upper Dir, Buner and Mardan) in KPK during present study (Table 3).
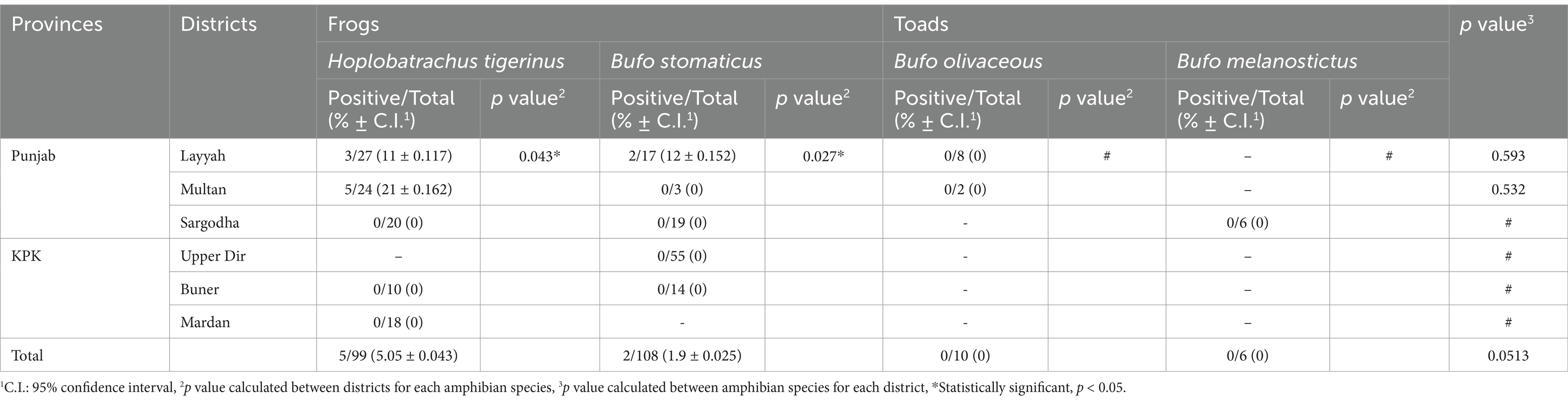
Table 3. Toxoplasma gondii-like strains’ prevalence rates among different species of frogs and toads in overall and according to the six sampling districts.
BLAST analysis conducted on haplotypes of our T. gondii-like strains revealed a sequence identity of 90.57% with T. gondii isolates reported from various mammals and birds across different countries (FJ176227, MH793500, KP895860, and MW374089). Sequence comparisons among our haplotypes showed a high identity of 98.87%. Phylogenetic analysis of the ITS-1 region of T. gondii and related strains indicated that the three Pakistani haplotypes generated in this study (OR896860, OR896861, and OR896862) clustered together but were genetically distinct from the ITS-1 sequences of T. gondii reported in various mammals and birds in Brazil (FJ176227, FJ176230, FJ176232, FJ966048, MF765978, MF766454, JF810932, JF810959, MH793504, MH793505, FJ966049, MW021507, MW023594, MW023595, and KX853130), as well as in Thailand (KP895860, KP895862, and KP895868), Poland (KX459518), Canada (MN153989), Germany (EU025025), China (AJ628251), the United States (KP999999, AF252408, AY488166, and KM525503), Norway (KM657806), and Pakistan (MW374089, MW374090, MW374293, and OL461229) (Figure 4).
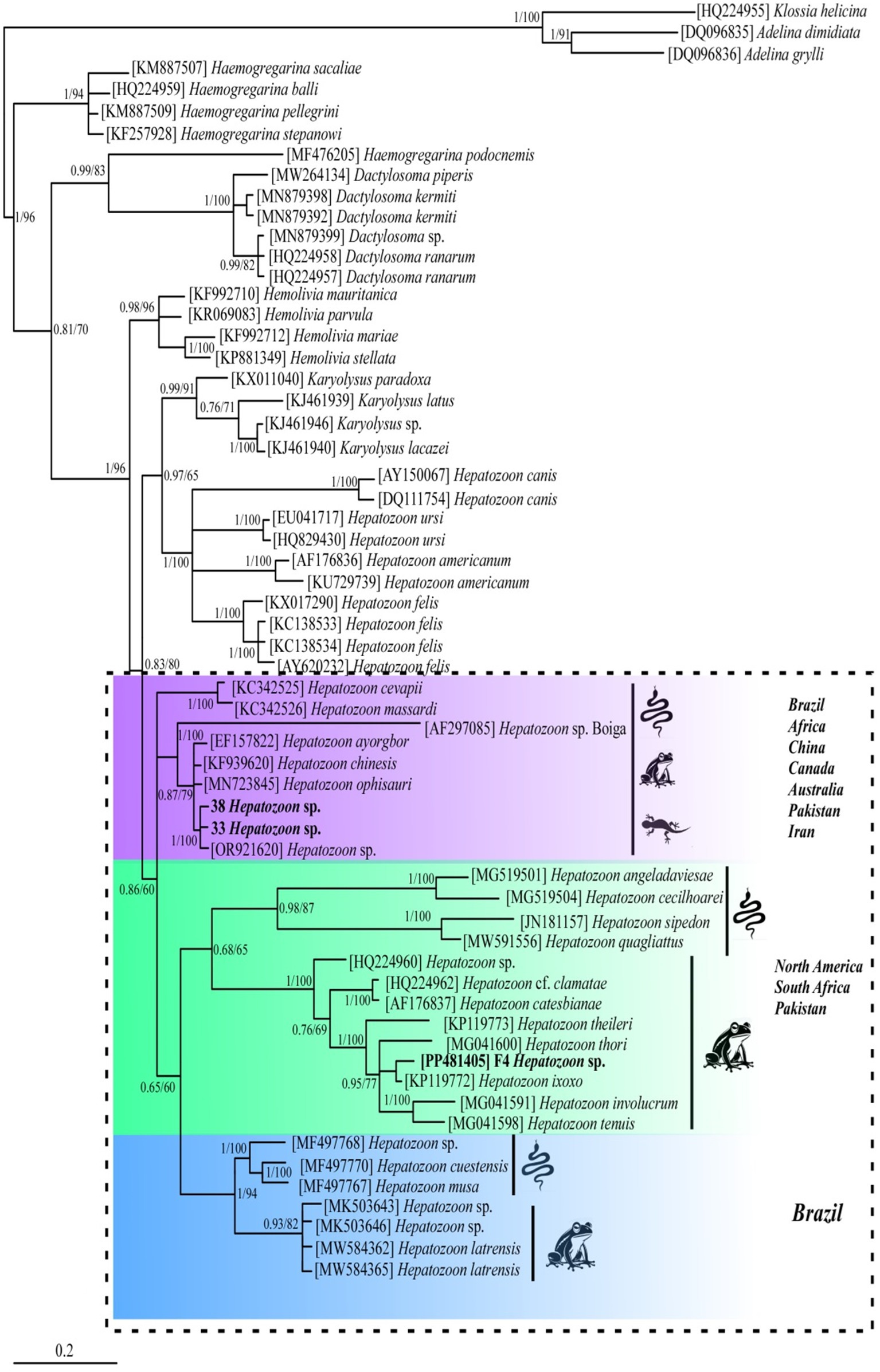
Figure 4. Phylogenetic tree of Toxoplasma gondii and related strains based on ITS-1 partial sequences. The three new sequences of T. gondii-like strains obtained in this study (OR896860, OR896861, and OR896862) are highlighted in bold and presented in box. Scale bar represents 0.08 substitutions per nucleotide position. Bootstrap values are shown as numbers on the nodes.
When comparing the prevalence of T. gondii-like strains among anurans (frogs and toads) captured from various sampling sites, One-way ANOVA results indicated a significant variation in parasite prevalence between sampling sites. The infected anurans included one frog species (H. tigerinus; p = 0.043) and one toad species (B. stomaticus; p = 0.027), all of which were captured from Punjab province. No infections were found in animals captured from KPK during present investigation (Table 3). Fisher’s exact test analysis revealed that T. gondii-like strains’ infection was not associated with the sex of H. tigerinus captured from five districts in Pakistan (p = 0.481; Table 4). A similar trend was observed for B. stomaticus (p = 0.958). This analysis could not be performed for B. olivaceous and B. melanostictus as no infections were detected in these toad species (Table 4).
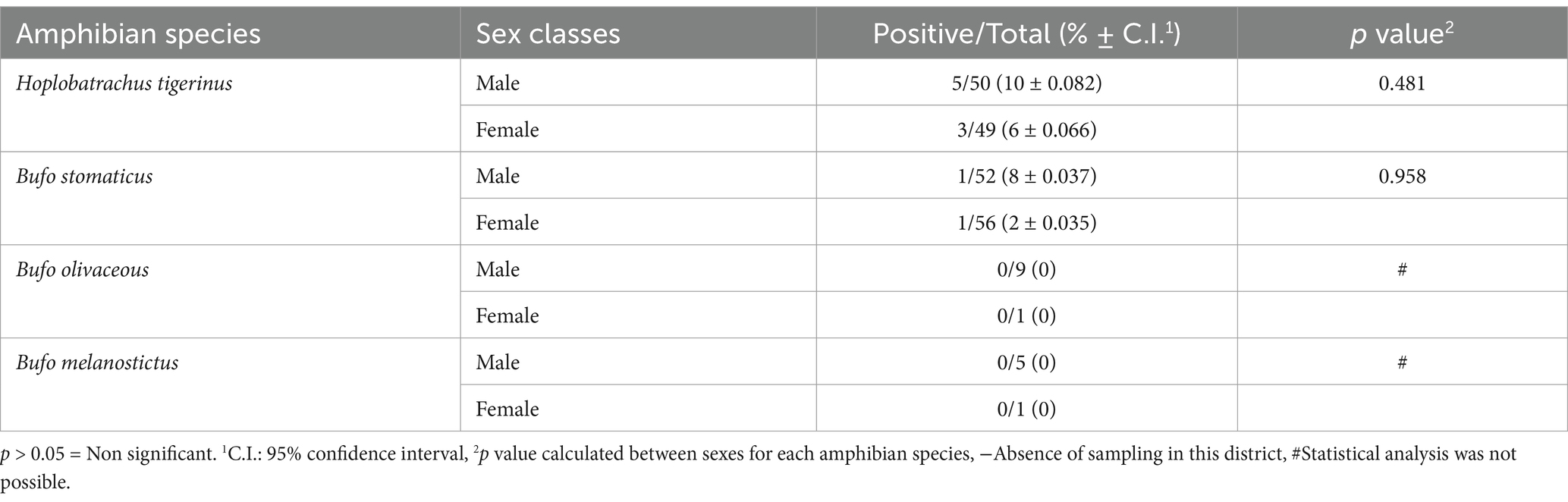
Table 4. Prevalence rates of Toxoplasma gondii-like strains in amphibian species by sex.
Amphibians are a transitional group of tetrapods that remain closely tied to water, particularly for reproduction, and they are among the most threatened vertebrate groups in the world. The main causes include climate change, habitat destruction and emerging diseases (23). As ectotherms with permeable skin, amphibians are extremely vulnerable to habitat alterations as well as for parasitic infections (31). They are known to host a wide variety of hemoparasites, including Haemogregarina, Hepatozoon, Lankesterella and Schellackia (32, 33). The presence of Toxoplasma gondii has never been reported in cold blooded animals before, especially in anurans. In light of these facts, the present study was designed to report the molecular prevalence and phylogeny of Hepatozoon spp. and T. gondii-like strains among various frog and toad species captured from three districts each in Punjab and KPK (Pakistan).
In the past decade, the understanding of apicomplexan parasite diversity in amphibians has significantly advanced due to the use of molecular techniques (34, 35). Despite this progress, the molecular prevalence of Hepatozoon spp. has not been previously reported in any animal species in Pakistan, particularly in anurans. Our study provides the first data on this unexplored research area in Pakistan. We found that 7.6% of anurans from the family Ranidae and Bufonidae were infected with Hepatozoon spp., including one frog species (H. tigerinus) and three toad species (B. stomaticus, B. olivaceous and B. melanostictus) (Table 1). DNA sequence analysis indicated that our primers amplified two parasite species: Hepatozoon spp. and Lankesterella sp. consistent with previous findings that these primers can detect multiple species (23).
Few reports exist on the presence of Hepatozoon spp. and/or Lankesterella sp. in anurans worldwide. Our findings are in line with those of Úngari et al. (36), who reported a 6.06% prevalence of Hepatozoon spp. in anurans from Mato Grosso State, Brazil. In another Brazilian study, de Abreu Reis Ferreira et al. (23) reported a 100% prevalence rate of haemogregarines parasites in Leptodactylus latrans and the first molecular report of Lankesterella sp. in amphibians from Brazil. Similarly, Isaak Delgado et al. (37) reported an 85% infection rate of Hepatozoon sp. and a 57% prevalence of Lankesterella sp. in Lithobates vaillanti from La Florida Biological Station, Tabasco, Mexico. Netherlands et al. (32) also reported a 73% infection rate of Hepatozoon spp. among African anurans of the family Bufonidae in Northern, Southern and Central Africa. Similarly, Leal et al. (38) found a 28.9% prevalence of Hepatozoon spp. in Leptodactylus podicipinus and Leptodactylus chaquensis from two regions of the Pantanal, state of Mato Grosso do Sul, Brazil (38). Readel and Goldberg (39) documented a 17% infection rate in frogs screened in Uganda, with infections from Hepatozoon, Trypanosoma, or microfilariae. In addition, Al-Khamesi et al. (40) observed a 13.33% prevalence of Hepatozoon spp. in 70 adult common frogs collected from different locations in Baghdad, Iraq (40). Netherlands et al. (2) reported a 8.9% prevalence in hyperoliid frogs from northern KwaZulu-Natal, South Africa. In contrast, Parejo-Pulido et al. (33) did not detect haemosporidian and haemogregarine parasites in 86 amphibian blood samples from southwestern Iberia. Similarly, Seabra-Babo et al. (56) failed to amplify DNA of haemogregarines in amphibian blood samples from Europe and North Africa, despite clear visual identification in blood smears. The absence of detected parasites in these studies could be attributed to a lack of appropriate aquatic vectors in their study areas or limited exposure to terrestrial vectors [(41)]. These findings highlight the limited information available on the prevalence of Lankesterella sp. and Hepatozoon spp., especially in Pakistani amphibians and globally. This underscores the importance of exploring these under-researched areas and screening wild hosts from various geo-climatic regions to enhance our understanding of their pathogenicity and diversity, which could inform effective control measures in amphibians.
The primers used in this study were originally designed by Ujvari et al. (22) to target Hepatozoon parasites. However, these primers are non-specific and also amplify other genera such as Eimeria, Sarcocystis and Isospora, as well as various other genetic groups (35). Out of 17 amplified PCR products, only 6 were successfully sequenced, with the remaining products failing due to insufficient blood volume or poor quantity or quality of extracted DNA. Of the six successfully sequenced samples, one Hepatozoon spp. was identified from a frog (H. tigerinus), while two Lankesterella spp. was identified from H. tigerinus and one from the toad B. stomaticus. Consequently, phylogenetic analyses were performed separately for the two detected parasite genera.
No prior attempts have been made to assess the genetic diversity of Hepatozoon spp. among Pakistani anurans. Thus, the 18S rRNA gene sequences from the amplified PCR products were utilized for phylogenetic analysis. The 18S rRNA gene is commonly used in molecular studies to reconstruct the evolutionary histories due to its slow rate of evolution, making it suitable for tracing ancient divergences (23). The DNA sequence analysis indicated that the Hepatozoon species infecting Pakistani anuran was 99.77% similar to H. ixoxo. The haplotype (GenBank aaccession number PP481405) obtained in this study showed similarity to 18S rRNA sequences of Hepatozoon spp. isolated from amphibians and reptiles from South Africa by Cook et al. (42) (GenBank accession numbers MG519501 and MG519504), Netherlands et al. (32), (GenBank accession numbers KP119772 and KP119773), and Netherlands et al. (2) (GenBank accession numbers MG041591, MG041598, and MG041600), adeleorinid coccidians in Canada by Barta et al. (43) (GenBank accession numbers JN181157, HQ224960, and HQ224962), snakes in Brazil by Úngari et al. (36) (GenBank accession number MW591556), and bullfrogs in the United States by Mathew et al. (44) (GenBank accession number AF176837) (Figure 2).
Three amplified PCR products from the 18S rRNA gene were used for the phylogenetic analysis of Lankesterella spp. The haplotypes identified in this study resided in distinct clades indicating sequence variations within the parasite (Figure 3). Lankesterella lineage PP476386 was most similar to sequences isolated from bird species unlike the other lineage (PP544154 and PP541577). Specifically, the Pakistani haplotypes PP544154 and PP541577 clustered with Lankesterella sp. reported in lizards in Spain [GenBank accession number KJ131417 (45)] and reptiles in Yemen [GenBank accession number MW076442 (46)]. In contrast, the Pakistani isolate PP476386 clustered with Lankesterella sp. from reptiles in Oman [GenBank accession number KX453649 (47)] (Figure 3). This data suggests that while the Pakistani haplotypes exhibit distinct phylogenetic relationships, they are closely related to known Lankesterella species found in various reptiles. This may indicate that the local haplotypes could represent new taxonomic entities, reflecting differences in host–parasite compatibility and potentially revealing undescribed endemic species. Our results underscore the influence of host ecology and relatedness on Lankesterella species distributions. More broadly, they highlight the necessity of screening wild hosts from remote and underexplored regions of Pakistan to gain deeper insights into parasite diversity.
Both Hepatozoon spp. and Lankesterella spp. belong to the order Eucoccidiorida (subclass Coccidia, class Conoidasida, Apicomplexa) and collectively considered as Haemogregarines (48). We have mentioned above that our primers were generalized and they are capable of amplifying both Hepatozoon spp. and Lankesterella spp. and as we were not successful in DNA sequencing all the amplified partial 18S rRNA gene sequences that were amplified in this study, so we were unable to analyze the data separately for Hepatozoon spp. and Lankesterella spp. Hence, we have analyzed all of our data for Haemogregarines prevalence. In this study, Haemogregarines infection among Pakistani anurans did not vary significantly between different sampling sites (Table 1). This finding aligns with the results of Leal et al. (38) who reported similar non-significant variations in Hepatozoon sp. prevalence among Leptodactylus podicipinus and Leptodactylus chaquensis frog species across different sampling sites. However, our results contrast with Readel and Goldberg (39), who observed significant differences in Hepatozoon spp. prevalence among different frog species in Uganda. This disparity may be attributed to varying habitat preferences and associated differences in vector abundance and contact rates. Hepatozoon spp. are typically transmitted through the ingestion of infected mosquitoes, and frogs living in more aquatic environments might experience higher blood parasite burdens due to increased exposure to insect vectors (39). Regarding the association between parasite infection and the sex of the amphibians, our study found no significant difference in Hepatozoon spp. prevalence between males and females across the anuran species (H. tigerinus, B. stomaticus, B. olivaceous, and B. melanostictus) captured from different districts (Table 4). This contrasts with the findings of Mohamed and Osman (49), who reported higher Hepatozoon sp. prevalence in males compared to females of A. regularis toads. The reasons for the observed differences in infection rates between sexes in their study remain unclear, and the authors recommended further screening of toad samples to validate these findings.
Among protozoan diseases, toxoplasmosis is particularly significant due to its zoonotic nature, with T. gondii, the causative agent, known to infect nearly all warm-blooded animals (16). Although T. gondii is not typically associated with cold-blooded animals, its presence has been documented in marine mammals such as cetaceans, pinnipeds, sirenians, and sea otters (50–52). This raises concerns about the potential role of cold-blooded animals, such as frogs, toads, turtles, crocodiles, snakes, fish, and shellfish, as reservoirs for T. gondii (53).
Our study reports a 4.5% prevalence of T. gondii-like infections in Pakistani anurans for the first time, detecting the parasite in both frog (H. tigerinus) and toad (B. stomaticus) species (Table 3). These findings highlight the importance of advanced diagnostic techniques, such as PCR, in understanding the prevalence and transmission dynamics of T. gondii and genetically related strains among frog and toad populations, with implications for both animal and public health.
This study also represents the first report on the genetic diversity of T. gondii-like strains among Pakistani amphibians. Phylogenetic analysis was performed using PCR-amplified products from the ITS-1 partial sequence of the pathogen, a commonly targeted marker in molecular phylogenetic studies due to its ease of amplification and relatively high variability (54). The identified haplotypes clustered together, forming a distinct clade separate from all published T. gondii sequences. The isolates included in this analysis originated from a variety of hosts across different geographical regions, underscoring the widespread distribution of T. gondii-like strains and their potential transmission among diverse host species.
Importantly, while the strains identified in this study are genetically related to T. gondii, they do not belong to the reference pathogenic strain, and their pathogenicity remains uncertain. However, these strains found in aquatic animals may represent ancestral species of those currently present in terrestrial animals, which could pose a significant threat due to their pathogenic potential. This highlights the need for further research to evaluate the health risks associated with these genetically related strains. Our findings illustrate the evolution of this parasite across various hosts, shedding light on its adaptability and potential reservoirs within amphibian populations. Understanding the evolution and diversity of T. gondii-like strains is essential for informing future control measures and public health strategies.
Our risk factor analysis indicated that the prevalence of T. gondii-like strains varied between sampling sites and among different anuran species. However, the prevalence was not associated with the sex of the amphibians screened (Tables 4). As this is the first report regarding the presence of T. gondii-like strains among amphibians, direct comparisons with previous studies are not possible. Nonetheless, these findings pave the way for further research into this widely distributed parasite across diverse amphibian species globally. Expanding this research will enhance our understanding of T. gondii and related strains in various animal hosts and elucidate host–parasite interactions, potentially leading to more effective control measures for this common pathogen. We recommend employing serological assays for the detection of T. gondii-like strains in frog and toad blood and suggest future studies attempt to isolate these strains from the muscles and brain of anurans for definitive confirmation.
This study provides the first documented evidence of T. gondii-like strains, Hepatozoon sp., and Lankesterella sp. infections in amphibians from Pakistan, revealing significant insights into parasite diversity and prevalence. The detection of T. gondii-like strains in both frogs and toads, along with the identification of H. ixoxo and Lankesterella spp., highlights the complex interactions between amphibians and their parasitic pathogens. Molecular analyses indicated notable genetic diversity among these parasites, suggesting the presence of potential new taxonomic entities and underscoring the necessity for further research. The variation in parasite prevalence across different sampling sites and amphibian species, combined with the lack of sex-specific infection patterns, reflects the dynamics of parasite transmission within amphibian populations. Importantly, the genetically related strains identified in aquatic animals may represent ancestral species of those currently found in terrestrial animals, posing a significant threat due to their pathogenic potential. Our findings contribute to a broader understanding of the evolution of T. gondii-like strains and Hepatozoon spp. in wildlife, emphasizing the potential implications of our studied hosts in this evolutionary context. This study enhances awareness of animal health and potential zoonotic risks, highlighting the importance of amphibians as reservoirs for these parasites. Future research should explore these relationships further to improve management strategies and reduce the impact of these parasites on amphibian health and ecosystems.
The datasets generated and/or analyzed during the current study are available in the GenBank repository, with accession numbers PP481405 (Hepatozoon sp.), PP544154, PP541577, and PP476386 (Lankesterella sp.) and OR896860, OR896861, and OR896862 (T. gondii-like strains).
The animal study was approved by Ethical Research Committee of the Bahauddin Zakariya University Multan (Pakistan). The study was conducted in accordance with the local legislation and institutional requirements.
MS: Formal analysis, Investigation, Resources, Writing – original draft. AUK: Conceptualization, Investigation, Resources, Writing – review & editing. AN: Formal analysis, Resources, Writing – review & editing. AA: Software, Writing – review & editing. TA: Funding acquisition, Software, Writing – review & editing. MU: Resources, Writing – review & editing. HM: Software, Validation, Writing – review & editing. SU: Resources, Writing – review & editing. HB: Software, Writing – review & editing. AK: Conceptualization, Resources, Software, Writing – review & editing. MB: Methodology, Software, Validation, Writing – original draft, Writing – review & editing. FI: Conceptualization, Formal analysis, Investigation, Project administration, Resources, Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This work is financially supported by the Researchers Supporting Project number (RSP2025R197), King Saud University, Riyadh, Saudi Arabia.
Authors are grateful to Letícia Pereira Úngari from Universidade de São Paulo – USP, Departamento de Parasitologia, Instituto de Ciências Biomédicas, São Paulo, SP, Brasil for technical support and her help in phylogenetic analysis of the pathogens.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. ^Biomatters, www.geneious.com
1. Frost, RD, Grant, T, Faivovich, JN, Bain, RH, Haas, A, Haddad, CFB, et al. The amphibian tree of life. Bull Amr Museum Nat Hist. (2006) 297:1–291. doi: 10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2
2. Netherlands, EC, Cook, CA, Du Preez, LH, Vanhove, MPM, Brendonck, L, and Smit, NJ. Monophyly of the species of Hepatozoon (Adeleorina: Hepatozoidae) parasitizing (African) anurans, with the description of three new species from hyperoliid frogs in South Africa. Parasitology. (2018) 145:1039–50. doi: 10.1017/S003118201700213X
3. Sarwar, MK, Malik, MF, Hussain, M, Azam, I, Iqbal, W, and Ashiq, U. Distribution and current status of amphibian fauna of Pakistan: a review. Electron J Biol. (2016) 12:243–6.
4. Crittini, AGF, Casiraghi, M, and Padoa-Schioppa, E. New data on amphibian and reptiles of the northern area of Pakistan: distribution, genetic variability, and conservation issue. North West J Zool. (2010) 6:1–12.
5. Kumar, R, Mahar, MA, Jumani, S, Bhanbro, R, Qazi, F, Ibupoto, M, et al. Biodiversity of amphibians in Pakistan, causes of their decline and their conservation. Ind J Sci Technol. (2020) 13:1243–7. doi: 10.17485/IJST/v13i11.149583_2020
6. Khan, MS. Key for identification of amphibians and reptiles of Pakistan. Pak J Zool. (1994) 26:249–55.
7. Ali, W, Javid, A, Hussain, SM, Azmat, H, and Jabeen, G. The amphibians and reptiles collected from different habitat types in district Kasur, Punjab, Pakistan. Pak J Zool. (2016) 48:1201–4.
8. Quillfeldt, P, Romeike, T, Masello, JF, Reiner, G, Willems, H, and Bedolla-Guzmán, Y. Molecular survey of coccidian infections of the side-blotched lizard Uta stansburiana on San Benito Oeste Island, Mexico. Étude moléculaire des infections à coccidies du lézard Uta stansburiana sur l’île de San Benito Oeste, Mexique. Parasit. (2018) 25:43. doi: 10.1051/parasite/2018043
9. Aktas, M, Ozübek, S, and Ipek, DN. Molecular investigations of Hepatozoon species in dogs and developmental stages of Rhipicephalus sanguineus. Parasitol Res. (2013) 112:2381–5. doi: 10.1007/s00436-013-3403-6
10. Han, H, Wu, Y, Dong, H, Zhu, S, Li, L, Zhao, Q, et al. First report of Hepatozoon (Apicomplexa: Adeleorina) from king ratsnakes (Elaphe carinata) in Shanghai, with description of a new species. Acta Parasitol. (2015) 60:266–74. doi: 10.1515/ap-2015-0038
11. Aktas, M, Özübek, S, Altay, K, Balkaya, İ, Utuk, AE, Kırbas, A, et al. A molecular and parasitological survey of Hepatozoon canis in domestic dogs in Turkey. Vet Parasitol. (2015) 209:264–7. doi: 10.1016/j.vetpar.2015.02.015
12. O’Dwyer, LH, Moço, TC, Paduan, KS, dos Santos Paduan, K, Spenassatto, C, da Silva, RJ, et al. Description of three new species of Hepatozoon (Apicomplexa, Hepatozoidae) from rattlesnakes (Crotalus durissus terrificus) based on molecular, morphometric and morphologic characters. Exp Parasitol. (2013) 135:200–7. doi: 10.1016/j.exppara.2013.06.019
13. Maia, JP, Harris, DJ, and Perera, A. Molecular survey of Hepatozoon species in lizards from North Africa. J Parasitol. (2011) 97:513–7. doi: 10.1645/GE-2666.1
14. Almuzaini, AM. Flow of zoonotic toxoplasmosis in food chain. Pak Vet J. (2023) 43:1–16. doi: 10.29261/pakvetj/2023.010
15. Elfadaly, HA, Shaapan, RM, Barakat, AM, Hassanain, NA, and Maher, A. The accuracy of developed peroxidase Toxoplasma gondii IgG ELISA plates for evaluating toxoplasmosis in sheep. Int J Vet Sci. (2023) 12:236–41. doi: 10.47278/journal.ijvs/2022.174
16. Ahmad, G, Masud, A, Naeem, M, Ghafar, A, Muqaddas, H, Qamar, MF, et al. Molecular prevalence, phylogeny and hematological impact of Toxoplasma gondii and Plasmodium spp. in common quails from Punjab, Pakistan. PLoS One. (2024) 19:e0304179. doi: 10.1371/journal.pone.0304179
17. Irshad, A, Noreen, S, Khan, W, and Rasool, A. Sero-epidemiology, spatial distribution and phylogenetic analysis of Toxoplasma gondii in goats of Malakand division of Pakistan. Pak Vet J. (2024) 44:442–8. doi: 10.29261/pakvetj/2024.152
18. Fredebaugh, SL, Mateus, PNE, McAllister, M, Warner, RE, and Weng, HY. Prevalence of antibody to Toxoplasma gondii terrestrial wildlife in a natural area. J Wildlif Dis. (2011) 47:381–92. doi: 10.7589/0090-3558-47.2.381
19. Khan, MS. Conservation biology of amphibians of Asia In: H Heatwole and I Das, editors. Amphibians of Pakistan and their conservation status. Kota Kinabalu: Natural History Publications Borneo (2014). 1–35.
20. Mansoor, R. An annotated checklist of amphibians and reptiles of Margalla Hill National Park. Pakistan. Pak J Zool. (2011) 43:20–522.
21. Ingerm, RF, and Stuebing, RB. A field guide to the frogs of Borneo. 2nd ed. Borneo: Natural History Publication, USA (2005).
22. Ujvari, B, Madsen, T, and Olsson, M. High prevalence of Hepatozoon spp. (Apicomplexa, Hepatozoidae) infection in water pythons (Liasis fuscus) from tropical Australia. J Parasitol. (2004) 90:670–2. doi: 10.1645/GE-204R
23. De Abreu Reis Ferreira, D, Perles, L, Machado, RZ, Prado, CPA, and André, MR. Molecular detection of apicomplexan hemoparasites in anurans from Brazil. Parasitol Res. (2020) 119:3469–79. doi: 10.1007/s00436-020-06835-9
24. Zintl, A, Halova, D, Mulcahy, G, O’Donovan, J, Markey, B, and DeWaal, T. In vitro culture combined with quantitative TaqMan PCR for the assessment of Toxoplasma gondii tissue cyst viability. Vet Parasitol. (2009) 164:167–72. doi: 10.1016/j.vetpar.2009.05.024
25. Kearse, M, Moir, R, Wilson, A, Sones-Havas, S, Cheung, M, Sturrock, S, et al. Geneious basic: an integrated a extendable desktop software platform for the organization and analysis of sequence data. Bioinformat. (2012) 28:1647–9. doi: 10.1093/bioinformatics/bts199
26. Darriba, D, Taboada, GL, Doallo, R, and Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods. (2012) 9:772.
27. Guindon, S, and Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. (2003) 52:696–704. doi: 10.1080/10635150390235520
28. Miller, MA, Pfeiffer, W, and Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In: Proceedings of the 2010 gateway computing environments workshop (GCE), New Orleans, LA, USA, pp. 1–8. (2010).
29. Tavaré, S, and Miura, RM. Some probabilistic and statistical problems in the analysis of DNA sequences, lectures on mathematics in the life sciences. Lect Math Lif Sci. (1986) 17:57–86.
30. Rambaut, A. FigTree v1.4.2. (2012). Available online at: http://tree.bio.ed.ac.uk/software/figtree/ (Accessed on December 01, 2023).
31. Blaustein, AR, Walls, SC, Bancroft, BA, Lawler, JJ, Searle, CL, and Gervasi, SS. Direct and indirect effects of climate change on amphibian populations. Diversity. (2010) 2:281–313. doi: 10.3390/d2020281
32. Netherlands, EC, Cook, CA, and Smit, NJ. Hepatozoon species (Adeleorina: Hepatozoidae) of African bufonids, with morphological description and molecular diagnosis of Hepatozoon ixoxo sp. nov. parasitising three Amietophrynus species (Anura: Bufonidae). Parasit Vect. (2014) 7:552. doi: 10.1186/s13071-014-0552-0
33. Parejo-Pulido, D, Mora-Rubio, C, Marzal, A, and Magallanes, S. Molecular characterization of haemosporidian and haemogregarine diversity in southwestern Iberian amphibians and reptiles. Parasitol Res. (2023) 122:1139–49. doi: 10.1007/s00436-023-07814-6
34. Baneth, G. Perspectives on canine and feline hepatozoonosis. Vet Parasitol. (2011) 181:3–11. doi: 10.1016/j.vetpar.2011.04.015
35. Harris, DJ, Spigonardi, MP, Maia, JP, and Cunha, RT. Molecular survey of parasites in introduced Pelophylax perezi (Ranidae) water frogs in the Azores. Acta Parasitol. (2013) 58:607–11. doi: 10.2478/s11686-013-0176-0
36. Úngari, LP, Netherlands, EC, de Alcantara, EP, Emmerich, E, Silva, RJ, and O’Dwyer, LH. Description of a new species Hepatozoon quagliattus sp. nov. (Apicomplexa: Adeleorina: Hepatozoidae), infecting the sleep Snake, Dipsas mikanii (Squamata: Colubridae: Dipsadinae) from Goiás state, Brazil. Acta Parasitol. (2021) 66:871880. doi: 10.1007/s11686-021-00355-x
37. Isaak-Delgado, AB, López-Díaz, O, Romero-Callejas, E, Martínez-Hernández, F, Muñoz-García, CI, Villalobos, G, et al. Morphological and molecular characteristics of hemoparasites in Vaillant's frogs (Lithobates vaillanti). Parasitol Res. (2020) 119:1891–901. doi: 10.1007/s00436-020-06689-1
38. Leal, DD, Dreyer, CS, da Silva, RJ, Ribolla, PE, Paduan, K, Dos, S, et al. Characterization of Hepatozoon spp. in Leptodactylus chaquensis and Leptodactylus podicipinus from two regions of the Pantanal, state of Mato Grosso do Sul, Brazil. Parasitol Res. (2015) 114:1541–9. doi: 10.1007/s00436-015-4338-x
39. Readel, AM, and Goldberg, TL. Blood parasites of frogs from an equatorial African montane forest in western Uganda. J Parasitol. (2010) 96:448–50. doi: 10.1645/GE-2284.1
40. Al-Khamesi, MB, Salman, IS, Abid, SA, and Ibrahim, SM. Study prevalence and effect of internal parasites and histopathological changes on common frogs at Baghdad city. Iraq. J Agricul Sci. (2018) 49:445–52.
41. Apperson, CS, Hassan, HK, Harrison, BA, Savage, HM, Aspen, SE, Farajollahi, A, et al. Host feeding patterns of established and potential mosquito vectors of West Nile virus in the eastern United States. Vect Born Zoonot Dis. (2004) 4:71–82. doi: 10.1089/153036604773083013
42. Cook, CA, Netherlands, EC, As, JV, and Jacobus Smit, N. Two new species of Hepatozoon (Apicomplexa: Hepatozoidae) parasitising species of Philothamnus (Ophidia: Colubridae) from South Africa. Folia Parasitol. (2018) 65:004. doi: 10.14411/fp.2018.004
43. Barta, JR, Ogedengbe, JD, Martin, DS, and Smith, TG. Phylogenetic position of the Adeleorinid coccidia (Myzozoa, Apicomplexa, Coccidia, Eucoccidiorida, Adeleorina) inferred using 18S rDNA sequences. J Eukaryot Microbiol. (2012) 59:171–80. doi: 10.1111/j.1550-7408.2011.00607.x
44. Mathew, JS, Van Den Bussche, RA, Ewing, SA, Malayer, JR, Latha, BR, and Panciera, RJ. Phylogenetic relationships of Hepatozoon (Apicomplexa: Adeleorina) based on molecular, morphologic, and life-cycle characters. J Parasitol. (2000) 86:366–72. doi: 10.1645/0022-3395(2000)086[0366:PROHAA]2.0.CO;2
45. Megía-Palma, R, Martínez, J, and Merino, S. Molecular characterization of haemococcidia genus Schellackia (Apicomplexa) reveals the polyphyletic origin of the family Lankesterellidae. Zool Script. (2014) 43:304–12. doi: 10.1111/ZSC.12050
46. Tomé, B, Maia, J, Perera, A, Carranza, S, and Vasconcelos, R. Parasites in a hotspot: diversity and specificity patterns of apicomplexans infecting reptiles from the Socotra archipelago. Parasitology. (2021) 148:42–52. doi: 10.1017/S0031182020002000
47. Maia, JP, Harris, DJ, Carranza, S, and Goméz-Díaz, E. Assessing the diversity, host-specificity and infection patterns of apicomplexan parasites in reptiles from Oman, Arabia. Parasitology. (2016) 143:1730–47. doi: 10.1017/S0031182016001372
48. Al-Quraishy, S, Abdel-Ghaffar, F, Dkhil, MA, and Abdel-Gaber, R. Haemogregarines and criteria for identification. Anim. (2021) 11:170. doi: 10.3390/ani11010170
49. Mohamed, OAEH, and Osman, A. Blood parasites of toads (Amietophrynus regularis) in Al-Sorojia and Jebel Aulia, Khartoum state, Sudan. J Mar Sci Environ Technol. (2022) 8:16–24.
50. Cole, RA, Lindsay, DS, Howe, DK, Roderick, CL, Dubey, JP, Thomas, NJ, et al. Biological and molecular characterizations of Toxoplasma gondii strains obtained from southern sea otters (Enhydra lutris nereis). J Parasitol. (2000) 86:526–30. doi: 10.1645/0022-3395(2000)086[0526:BAMCOT]2.0.CO;2
51. Mathews, PD, Da Silva, VM, Rosas, FC, Neto, JA, Lazzarini, SM, and Ribeiro, DC. Occurrence of antibodies to Toxoplasma gondii and Lepstospira spp. in manatees (Trichechus inunguis) of the Brazilian Amazon. J Zool Wildlif Med. (2012) 43:85–8. doi: 10.1638/2011-0178.1
52. Ubey, JP, Murata, FH, Cerqueira-Cézar, CK, Kwok, OC, and Grigg, ME. Recent epidemiologic and clinical importance of Toxoplasma gondii infections in marine mammals: 2009–2020. Vet Parasitol. (2020) 288:109296. doi: 10.1016/j.vetpar.2020.109296
53. Nayeri, T, Sarvi, S, and Daryani, A. Toxoplasma gondii in mollusks and cold-blooded animals: a systematic review. Parasitology. (2021) 148:895–903. doi: 10.1017/S0031182021000433
54. Tippery, NP, and Les, DH. Phylogenetic analysis of the internal transcribed spacer (ITS) region in Menyanthaceae using predicted secondary structure. Mol Phylogenet Evol. (2008) 49:526–37. doi: 10.1016/j.ympev.2008.07.019
55. Posada, D., and Buckley, T. R. (2004). Model selection and model averaging in phylogenetics: advantages of akaike information criterion and bayesian approaches over likelihood ratio tests. Systemat. Biol. 53, 793–808. doi: 10.1080/10635150490522304
Keywords: Bufo olivaceous, Bufo stomaticus, Hoplobatrachus tigerinus, prevalence and phylogeny, amphibians, Pakistan
Citation: Salim M, Khan AU, Niazi AK, Aqdas A, Dawoud TM, Usman M, Muqaddas H, Ullah S, Belkahia H, Khan A, Ben Said M and Iqbal F (2025) Molecular detection and phylogenetic identification of Toxoplasma gondii-like strains, Hepatozoon ixoxo and Lankesterella sp. in frogs and toads. Front. Vet. Sci. 12:1568298. doi: 10.3389/fvets.2025.1568298
Edited by:
Rao Zahid Abbas, University of Agriculture, Faisalabad, PakistanReviewed by:
Münir Aktaş, Firat University, TürkiyeCopyright © 2025 Salim, Khan, Niazi, Aqdas, Dawoud, Usman, Muqaddas, Ullah, Belkahia, Khan, Ben Said and Iqbal. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Adil Khan, ZHJhZGlsa2hhbkBia3VjLmVkdS5waw==; Mourad Ben Said, bW91cmFkLmJlbnNhaWRAaXNic3QudW1hLnRu; Furhan Iqbal, ZnVyaGFuLmlxYmFsQGJ6dS5lZHUucGs=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.