
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Vet. Sci. , 06 March 2025
Sec. Veterinary Epidemiology and Economics
Volume 12 - 2025 | https://doi.org/10.3389/fvets.2025.1553152
This article is part of the Research Topic Sentinels of Health: Advancements in Monitoring and Surveillance of Vector-Borne Diseases in Domestic and Wild Animals and Vectors View all 13 articles
 Xiaoshuang Han1,2†
Xiaoshuang Han1,2† Ziheng Liu1,2†Zhixian Jiang3†Shanshan Zhao1,2†
Ziheng Liu1,2†Zhixian Jiang3†Shanshan Zhao1,2† Sándor Hornok4,5Meihua Yang3Gang Liu1,2*
Sándor Hornok4,5Meihua Yang3Gang Liu1,2* Yuanzhi Wang1,2*
Yuanzhi Wang1,2*Long-tailed ground squirrels (LTGRs, Spermophilus undulatus) are known as reservoirs of multiple arthropod-borne pathogens, such as Yersinia pestis and Bartonella rochalimae. However, data on the prevalence of spotted fever group rickettsiae (SFGR) and Coxiella burnetii in LTGRs and its ectoparasites are limited. In two alpine regions of Xinjiang Uygur Autonomous Region (XUAR, northwestern China), a total of 346 samples were collected from 142 LTGRs, including 142 livers and 204 pooled ectoparasites (Citellophilus tesquorum dzetysuensis: 120 pools of 484 fleas; Frontopsylla elatoides elatoides: 19 pools of 71 fleas; Neopsylla mana: 1 pool of 4 fleas; and Linognathoides urocitelli: 64 pools of 865 lice). From these samples, the DNA was extracted, followed by PCR amplification of different genetic markers. Particularly, genes encoding the outer membrane protein A and B (ompA, ompB), citrate synthase (gltA), and surface cell antigen 1 (sca1) were used to identify the SFGR. Additionly, the capsular outer membrane protein (Com1) gene and insertion sequence (IS1111) genes were used to detect Coxiella. Rickettsia sibirica subsp. sibirica, Rickettsia felis, and C. burnetii were detected in LTGRs, as well as in flea and louse pools. Rickettsia raoultii was found in LTGRs and flea pools. Furthermore, Rickettsia slovaca was also identified in the flea pools. This study provides molecular evidence for the occurrence of SFGR and C. burnetii in LTGRs and their ectoparasites. These findings suggest that R. sibirica, R. slovaca, R. raoultii, R. felis and C. burnetii are transmitted between LTGRs (as potential reservoirs) and their fleas and lice (as potential vectors).
Species of the genus Rickettsia belong to four distinct phylogenetic clades: spotted fever group, typhus group, ancestral group, and transitional group (1). The spotted fever group rickettsiae (SFGR) include over 30 distinct species that may cause severe infections in humans, domestic animals and wildlife (2). The majority of SFGR are tick-borne, while R. felis is typically transmitted by fleas. Notably, some SFGR species can be transmitted by lice and mosquitoes, albeit rarely (3–5).
Coxiella burnetii, recognized as the causative agent of Q fever, stands out as the most notorious member of its genus (6). As a globally distributed pathogen, it can infect a diverse range of mammals, from rodents to bats (7, 8). The main vectors of C. burnetii are hard ticks (7), but it is also potentially transmitted by fleas and lice (9, 10).
Long-tailed ground squirrels (LTGRs, Spermophilus undulatus) are medium-sized ground-dwelling rodents, inhabiting distinct alpine habitats in Central Asia, including Kazakhstan, Mongolia, the southern region of the Russian Federation and northwestern China (11). Previously, LTGRs and arthropods infesting them were shown to act as reservoirs or potential vectors for some pathogens, such as Yersinia pestis, Borrelia burgdoferi sense lato, Anaplasma phagocytophilum, Trypanosoma otospermophili, Blechomonas luni, tick-borne encephalitis virus and Hepacivirus C (12–16). However, the evidence of SFGR and C. burnetii in LTGRs and their ectoparasites, especially in fleas and lice, remains still unknown. The aim of this study was to screen these pathogens in liver samples of LTGRs and their associated arthropods.
In total, 142 LTGRs were captured in July, 2024 in Wenquan County and Jinghe County (1200–2,500 m above sea level, both adjacent to Kazakhstan), Bortala Mongolian Autonomous Prefecture, northwestern China (Supplementary Figure 1). To achieve this goal, Sherman traps (H.B. Sherman Traps, Tallahassee, FL, United States) were deployed at the entry points of occupied burrows. The survey encompassed a total of 150 traps per site, which were inspected once an hour (17).
All captured rodents were identified by experienced zoologists based on morphological characteristics, such as body length, fur color, tail length and other features (17). Subsequently, the rodent were euthanized and killed via cervical dislocation by certified personnel at the enhanced biosafety level 2 laboratory, Shihezi University (18). Each sampled rodent was then put in individual ziplock bags stored at −80°C. The liver was removed from individual LTGR and placed into each labeled tubes. The species was confirmed from four liver samples of representative rodents by sequencing the cytochrome b (cytb) gene (19). All procedures involving wild rodents adhered to the ethical guidelines of Animal Ethics Committee of Shihezi University (Approval No. A2022-029-01).
Arthropod ectoparasites (559 fleas, 865 lice and 1,136 ticks) were collected from individual rodents through gentle brushing of their fur, and then preserved in 70% ethanol. All sampled ticks were used for virus research by another team. For morphological identification, the fleas and lice were treated with 10% NaOH for 1–3 days and put onto slides for microscopical examination (11, 20). Meanwhile, the cytochrome c oxidase subunit II (COII) gene for fleas, and the 18S ribosomal RNA (18S rRNA) gene for lice were amplified and sequenced, in order to confirm their taxonomy (21–23). Subsequently, the ectoparasite samples were grouped into pools according to individual host from where they were collected, flea or louse species, number and sampling sites. Flea pools contained 2 to 5 individuals, while lice pools contained 8 to 15 individuals. Finally, fleas were allocated into a total of 140 pools and lice into 64 pools which were used to screen pathogens as described below.
Each arthropod pool and liver sample were extracted with TIANamp Genomic DNA Kit (TIANGEN, Beijing, China) according to the manufacturer’s instructions. Four genetic markers, including outer membrane proteins A and B (ompA and ompB), citrate synthase (gltA) and surface cell antigen 1 (sca1), were used to detect SFGR (17). In addition, the capsular outer membrane protein (Com1) gene and the insertion sequence (IS1111) gene were targeted to investigate the presence of C. burnetii (24, 25). The primer sequences and PCR conditions are shown in Supplementary Tables 1, 2. Negative controls consisted of double-distilled water, which consistently showed no detectable PCR product in all tests. Positive controls were DNA samples of Rickettsia lusitaniae from common pipistrelles and Coxiella-like symbiont from ticks, both preserved in our laboratory (26, 27). The PCR products were purified using the TIANgel Midi Purification Kit (TIANGEN, Beijing, China), and sequenced with Sanger and 454-pyrosequenced PCR amplicons (28, 29). The above sequencing was conducted three times to check the reproducibility. Obtained sequences were compared to reference sequences found in GenBank using BLAST.1 Phylogenetic analysis was conducted using MEGA 7.0 software, employing the neighbor-joining method and 1,000 replicates for bootstrap support.
All sampled rodents were identified as long-tailed ground squirrels (S. undulatus) based on their morphology and a 98.86% sequence identity (1,124/1137 bp) to the cytb gene of this species found in Russia (OQ695583). A total of 559 fleas were collected. The average flea index was 3.94 (559/142). Subsequently, three species were identified after microscopical examination (Figure 1), including Citellophilus tesquorum dzetysuensis (484), Frontopsylla elatoides elatoides (71) and Neopsylla mana (4). The former flea species was morphologically indientified by genal comb absent, reduced frontal bristles (1 in males, absent in females), pronotal ctenidium with 18–22 vertical spines, labial palps extending to or beyond forecoxa. The middle was by prominent frontal tubercle, 6–7 frontal bristles, 3 ocular bristles above eye, pronotal ctenidium with 22 spines, labial palps reaching forecoxa apex. The latter was by two genal combs (outer comb with short, broad spines; inner comb with narrow, posteriorly inclined spines), pronotal ctenidium with 17–20 spines, labial palps reaching two-thirds of forecoxa. The COII gene sequences of these species exhibited 100, 99.86% (713/714 bp), and 100% sequence identities to those of conspecific fleas reported from China, respectively (PP475165, MF000677 and MF000670). Furthermore, 865 lice were also collected. The average louse index was 6.09 (865/142). All sampled lice in this study were identified as Linognathoides urocitelli by molecular detection and morphological key features, which included dorsoventrally flattened body, reduced eyes, 5-segmented antennae with sexually dimorphic spines on third segment (males), thoracic sternal plate with posterior medial lobe, and paratergal plates on abdominal segments III-VII bearing spiracular openings. The 18S rRNA gene sequence of this species exhibited 99.81% (522/523 bp) sequence identity to the same louse species from LTGR in Mongolia (MK478719).
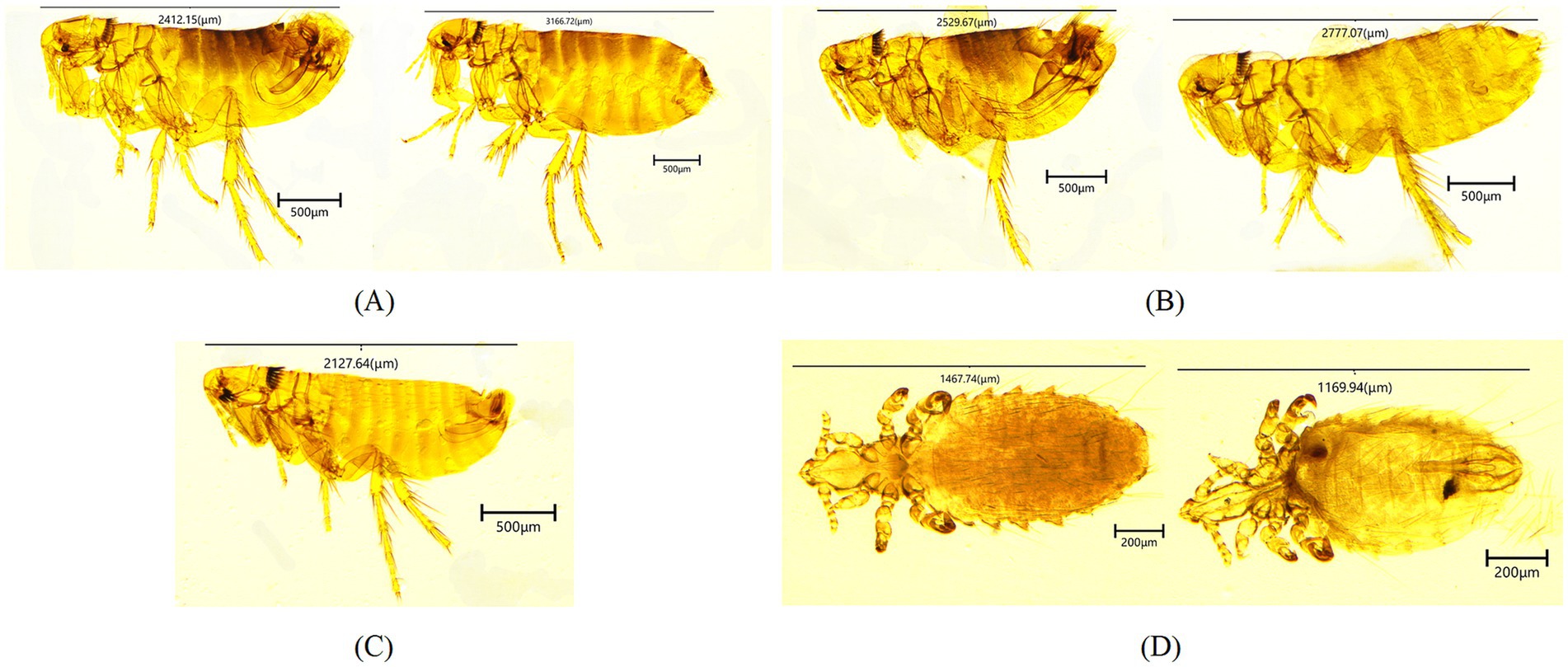
Figure 1. Photomicrographs of morphologically identified fleas and lice [Citellophilus tesquorum dzetysuensis (A), Frontopsylla elatoides elatoides (B), Neopsylla mana (C), Linognathoides urocitelli (D)].
Eleven LTGRs (7.75%, 11/142), 12 flea pools (8.57%, 12/140) and 15 louse pools (23.44%, 15/64) tested positive for SFGR. Among the 11 SFGR-positive LTGRs, 10 liver samples were also SFGR-positive in their associated flea and louse pools. BLAST and phylogenetic analyses revealed that R. sibirica and R. felis were present in LTGRs, flea and louse pools. Additionally, R. raoultii was identified in LTGRs and flea pools. Moreover, R. slovaca was also detected in the flea pools (Table 1).
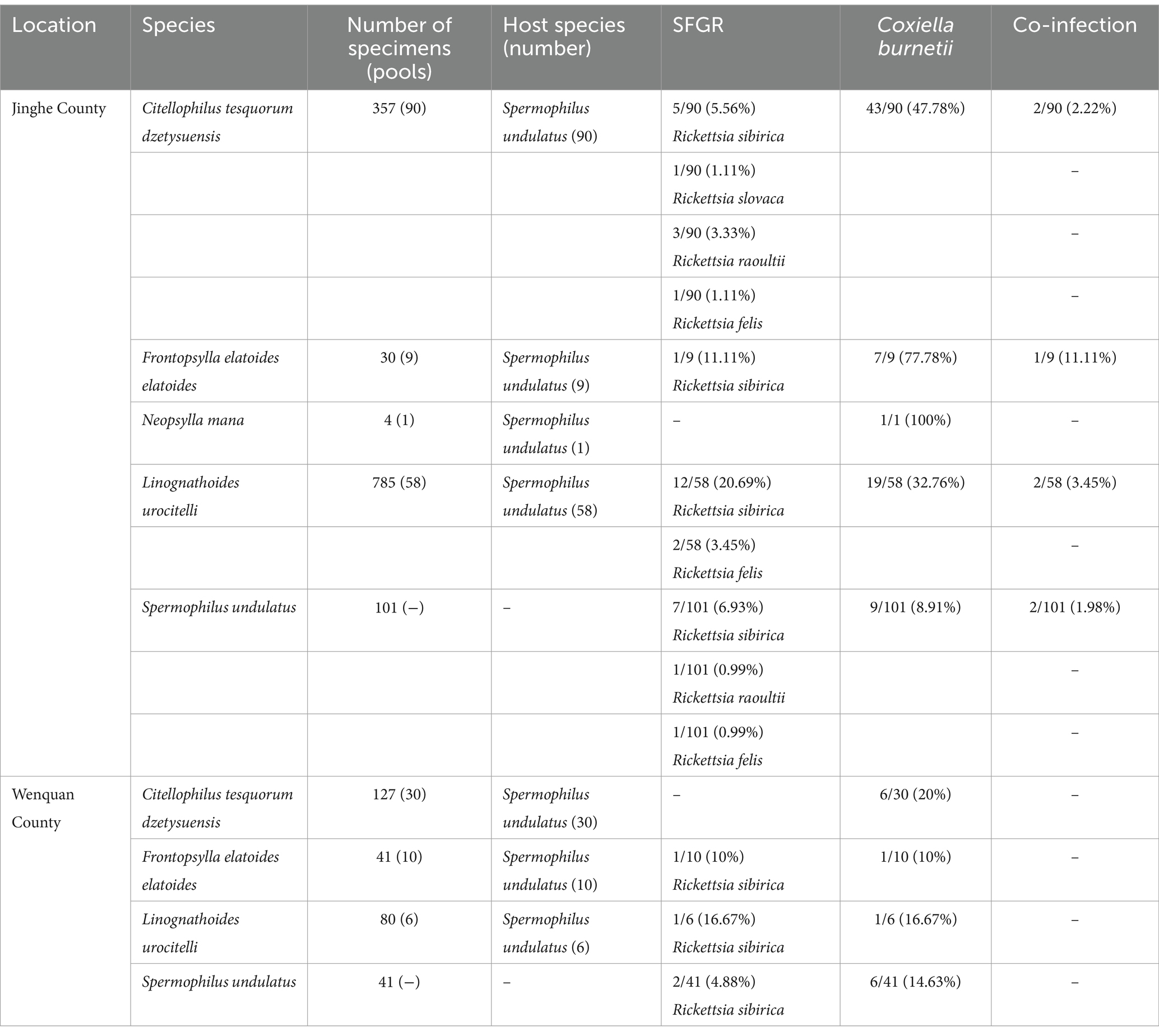
Table 1. Information on specimens used in this study, including geographical location of collection site, total number, pool number, hosts, and prevalence of pathogens.
Regarding sequence comparisons based on the four genetic markers, Rickettsia sibirica subsp. sibirica showed 100% identity compared with the sequence of conspecific bacteria from a tick-bitten patient in Russia (KT006594); R. slovaca showed 100% identity to R. slovaca from Dermacentor marginatus ticks in Kazakhstan (MW922580); R. raoultii showed 99.47–99.50% identity to R. raoultii from striped field mouse (Apodemus agrarius) in China (MZ297809); and R. felis showed 99.76–100% identity compared with the sequence of R. felis from cat fleas (Ctenocephalides felis) in Indonesia (MT499365; Figure 2). The detailed similarities and divergences of the sequences in this study are shown in Supplementary Table 3.
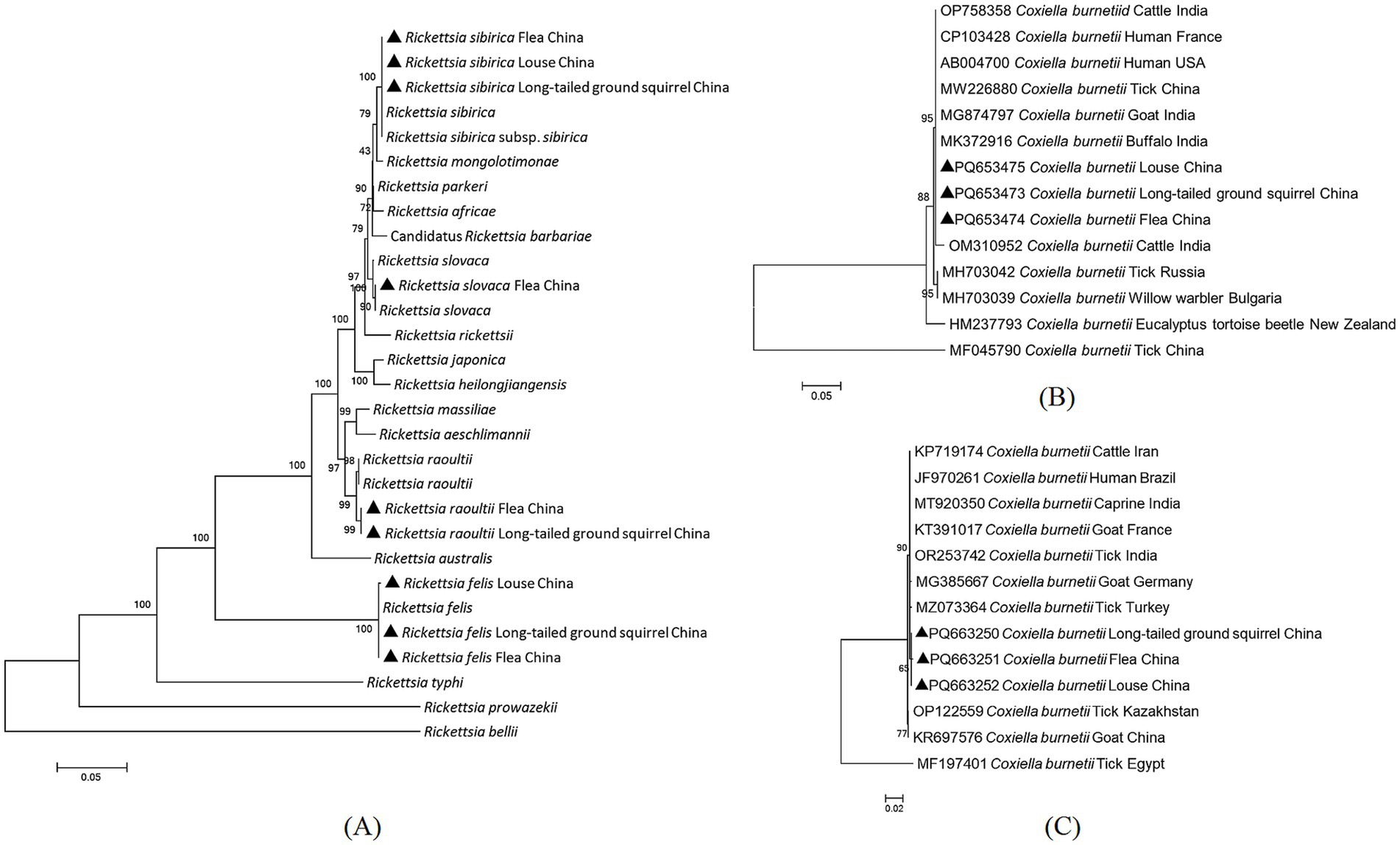
Figure 2. Phylogenic tree of spotted fever group rickettsiae (SFGR) and Coxiella burnetii from long-tailed ground squirrels and their ectoparasites [the ompA-ompB-gltA-sca1 concatenated sequences for SFGR (A), the Com1 concatenated sequences for C. burnetii (B), the IS1111 concatenated sequences for C. burnetii (C)]. The new sequences provided in the present study are indicated by a black triangle (containing the accession number).
C. burnetii was detected in 15 LTGRs (10.56%, 15/142), 58 flea pools (41.43%, 58/140) and 20 louse pools (31.25%, 20/64). The prevalence of C. burnetii in both LTGRs and their ectoparasites was significantly higher than that of SFGR (X2 = 35.09, df = 2, p < 0.05), suggesting a more prominent circulation of C. burnetii in these region. Based on BLASTn analysis, the IS1111 sequences (PQ663250-PQ663252) in this study were closest related to a C. burnetii isolate from Rhipicephalus sanguineus ticks in Turkey (MZ073364), showing 99.54–99.69% identities (Figure 2).
Notably, two LTGRs (1.41%, 2/142) were co-infected with R. sibirica and C. burnetii. More interestingly, their associated three flea pools (2.14%, 3/140) and two louse pools (3.13%, 2/64) were also co-infected with R. sibirica and C. burnetii.
In this study, 142 LTGRs and their ectoparasites, 559 fleas and 865 lice were collected in Central Asia. In these samples, four SFGR species were detected, including R. sibirica, R. slovaca, R. raoultii, and R. felis, as well as C. burnetii. To our best knowledge, (i) R. sibirica, R. felis and C. burnetii were identified here for the first time in LTGRs and their parasites (fleas and lice); (ii) in this study, first molecular evidence is also provided for R. raoultii in LTGRs and their ectoparasitic fleas; (iii) additionally, R. slovaca is newly recognized in LTGR fleas.
SFGR have worldwide distribution and affect a wide range of wild and domestic vertebrates. Although most reports focus on tick-borne transmission of SFGR, growing evidence suggests that other arthropods-including fleas (notably for R. felis), lice, keds, and bugs-may serve as potential vectors (2). R. felis is primarily transmitted by fleas, particularly by the cat flea (C. felis) (30). Similarly, molecular evidence was provided for R. slovaca in Haematopinus suis lice from boars in Algeria and later in Laelaps agilis, Laelaps jettmari and Eulaelaps stabularis mites from rodents in Slovakia (31, 32). With regard to R. raoultii, its occurrence was detected in fleas from yellow necked mouse (Apodemus favicollis) and bank vole (Myodes glareolus) in Germany (33). In this study, we reported novel findings of R. sibirica and R. felis in C. tesquorum dzetysuensis, F. elatoides elatoides and L. urocitelli, as well as R. slovaca and R. raoultii in C. tesquorum dzetysuensis and F. elatoides elatoides. In addition, SFGR were previously detected in wild boars, birds, dogs and cats, which act as reservoirs (34–37). Rodents are also notably recognized as reservoirs including yellow necked mouse (Apodemus flavicollis), common vole (Microtus arvalis) and European water vole (Arvicola terrestris) (38). Interestingly, we found that R. sibirica, R. raoultii and R. felis were present not only in the aforementioned arthropods but also in LTGRs. Notably, among 11 SFGR-positive LTGRs, 10 liver samples were also SFGR-positive in their associated flea and louse pools. This high concordance suggests that ectoparasites may play a critical role in either acquiring pathogens from infected hosts or transmitting SFGR within rodent populations. Although we have detected these SFGR in LTGRs and their ectoparasites, further experiments are still needed to confirm the role of these arthropods as vectors.
C. burnetii, listed as category B bioterrorism agent in United States, has diverse infection routes including air-borne and venereal transmission (39). Furthermore, it is transmitted by infected arthropods, such as ticks, fleas, lice and mites (10, 40–42). Previously, C. burnetii was found in Haematopinus eurysternus lice from a cow in Egypt (10). More recently, a report revealed the presence of C. burnetii in fleas collected from Norway rat (Rattus norvegicus), black rat (Rattus rattus) and Cyprus red fox (Vulpes vulpes indutus) in Cyprus (43). In this study, we extend these findings with the novel detection of C. burnetii in C. tesquorum dzetysuensis, F. elatoides elatoides, N. mana and L. urocitelli. Additionally, C. burnetii was previously identified in deer, sheep, cattle, goats, birds and rabbits, which serve as reservoirs (7). Rodents also have an important role as reservoirs, as exemplified by the flying squirrel (Pteromys volans), red squirrel (Sciurus vulgaris), and red-backed vole (Myodes gapperi) (44). In our research, LTGRs have been found to harbor C. burnetii as well. Crucially, all C. burnetii-positive LTGRs in this study concurrently carried infected fleas or lice, suggesting a potential bi-directional transmission between hosts and ectoparasites.
Rodents serve as hosts for several species of blood-sucking arthropods, and infected ectoparasites may transmit pathogens to these mammals through their bites. Consequently, naïve ectoparasites themselves can become infected after biting these rodent reservoirs, leading to new opportunities for pathogens to spread by horizontal transmission (Supplementary Figure 2). Examples of such pathogens include R. slovaca, R. raoultii, R. felis and R. monacensis (33, 45). Therefore, the concurrent detection of pathogens in hosts, fleas, and lice is of significant importance, underscoring the complexity of pathogen transmission and the potential roles of these vectors in pathogen spillover and host shift.
In this study, we present the first molecular evidence of SFGR and C. burnetii in LTGRs and their ectoparasites. Our findings suggested that R. sibirica, R. slovaca, R. raoultii, R. felis and C. burnetii are transmitted between LTGRs (as potential reservoirs) and fleas and lice (as potential vectors) infesting them.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, Rodent species cytb: PQ653469; flea species COII: PQ653470-PQ653472; louse species 18S rRNA: PQ640277; SFGR ompA: PQ677906-PQ677914; SFGR ompB: PQ677915-PQ677923; SFGR gltA: PQ677924-PQ677932; SFGR sca1: PQ677933-PQ677941; Coxiella burnetii Com1: PQ653473-PQ653475; Coxiella burnetii IS1111: PQ663250-PQ663252.
This study was approved by the Animal Ethics Committee of Shihezi University (approval no. A2022-029-01). The study was conducted in accordance with the local legislation and institutional requirements.
XH: Conceptualization, Investigation, Software, Writing – original draft. ZL: Conceptualization, Investigation, Writing – original draft. ZJ: Conceptualization, Investigation, Writing – original draft. SZ: Conceptualization, Investigation, Writing – original draft. SH: Conceptualization, Writing – review & editing. MY: Investigation, Resources, Software, Writing – original draft. GL: Conceptualization, Investigation, Resources, Software, Writing – original draft, Writing – review & editing. YW: Conceptualization, Investigation, Resources, Software, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (82260399, 82260414, and 82260410), the Natural Science Key Project of Xinjiang Uygur Autonomous Region (2022B03014), the Science & Technology Innovation Team Project of TIANSHAN Elite (2023TSYCTD0020), and the High-Level Talent Initiative Foundation of Shihezi University (RCZK202369).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1553152/full#supplementary-material
SUPPLEMENTARY FIGURE 1 | The locations for capturing rodents and associated ectoparasitic arthropods in the current study.
SUPPLEMENTARY FIGURE 2 | Circulation of pathogens between ectoparasites and rodents (LTGRs): (A) Ectoparasites (e.g., fleas and lice) acquire pathogens by biting infected rodents (LTGRs); (B) Pathogen-carrying ectoparasites bite uninfected rodents (LTGRs); (C) Uninfected rodents (LTGRs) become infected by bites from pathogen-carrying ectoparasites.
SUPPLEMENTARY TABLE 1 | Target genes and sequences of primers used in this study for identifying Long-tailed ground squirrels (LTGRs), fleas, lice, spotted fever group rickettsiae (SFGR) and Coxiella burnetii.
SUPPLEMENTARY TABLE 2 | Cycling conditions of PCR assays.
SUPPLEMENTARY TABLE 3 | The BLAST analysis of Long-tailed ground squirrels (LTGRs), fleas, lice, spotted fever group rickettsiae (SFGR) and Coxiella burnetii.
LTGR, Long-tailed ground squirrel; SFGR, spotted fever group rickettsiae; XUAR, Xinjiang Uygur Autonomous Region; ompA, outer membrane protein A; ompB, outer membrane protein B; gltA, citrate synthase; sca1, surface cell antigen 1; com1, capsular outer membrane protein; IS1111, insertion sequence.
1. De Vito, A, Geremia, N, Mameli, SM, Fiore, V, Serra, PA, Rocchitta, G, et al. Epidemiology, clinical aspects, laboratory diagnosis and treatment of Rickettsial diseases in the Mediterranean area during COVID-19 pandemic: a review of the literature. Mediterr J Hematol Infect Dis. (2020) 12:e2020056. doi: 10.4084/MJHID.2020.056
2. Zhang, YY, Sun, YQ, Chen, JJ, Teng, AY, Wang, T, Li, H, et al. Mapping the global distribution of spotted fever group rickettsiae: a systematic review with modelling analysis. Lancet Digit Health. (2023) 5:e5–e15. doi: 10.1016/S2589-7500(22)00212-6
3. Parola, P, Paddock, CD, Socolovschi, C, Labruna, MB, Mediannikov, O, Kernif, T, et al. Update on tick-borne rickettsioses around the world: a geographic approach. Clin Microbiol Rev. (2013) 26:657–702. doi: 10.1128/CMR.00032-13
4. Horta, MC, Scott, FB, Correia, TR, Fernandes, JI, Richtzenhain, LJ, and Labruna, MB. Rickettsia felis infection in cat fleas ctenocephalides felis felis. Braz J Microbiol. (2010) 41:813–8. doi: 10.1590/S1517-83822010000300035
5. Blanda, V, D’Agostino, R, Giudice, E, Randazzo, K, La Russa, F, Villari, S, et al. New real-time PCRs to differentiate Rickettsia spp. and Rickettsia conorii. Molecules. (2020) 25:4431. doi: 10.3390/molecules25194431
6. Kazar, J. Coxiella burnetii infection. Ann N Y Acad Sci. (2005) 1063:105–14. doi: 10.1196/annals.1355.018
7. Celina, SS, and Cerný, J. Coxiella burnetii in ticks, livestock, pets and wildlife: a mini-review. Front Vet Sci. (2022) 9:1068129. doi: 10.3389/fvets.2022.1068129
8. Silva-Ramos, CR, Faccini-Martínez, ÁA, Pérez-Torres, J, Hidalgo, M, and Cuervo, C. First molecular evidence of Coxiella burnetii in bats from Colombia. Res Vet Sci. (2022) 150:33–5. doi: 10.1016/j.rvsc.2022.05.009
9. Wu, YL, Hu, SF, Zhang, XL, Wang, HM, Pan, HY, Liu, GH, et al. Complete bacterial profile and potential pathogens of cat fleas Ctenocephalides felis. Acta Trop. (2023) 243:106923. doi: 10.1016/j.actatropica.2023.106923
10. Reeves, WK, Szumlas, DE, Moriarity, JR, Loftis, AD, Abbassy, MM, Helmy, IM, et al. Louse-borne bacterial pathogens in lice (Phthiraptera) of rodents and cattle from Egypt. J Parasitol. (2006) 92:313–8. doi: 10.1645/GE-717R.1
11. Durden, LA, Robinson, C, Cook, JA, McLean, BS, Nyamsuren, B, and Greiman, SE. A new species of sucking louse from the long-tailed ground squirrel, Urocitellus undulatus, from Mongolia, with a key to species, and a review of host associations and geographical distributions of members of the genus Linognathoides (Psocodea: Anoplura: Polyplacidae). J Parasitol. (2019) 105:469–79. doi: 10.1645/18-198
12. Bazanova, LP, and Innokent’eva, TI. Reservation forms of plague infectious agent in Tuva natural focus. Zh Mikrobiol Epidemiol Immunobiol. (2012) 5:115–9.
13. Li, Bo, Shan, Lu, Luo, Tao, and Abulikemu, Abudurexiti·, Hao, Qin, Wang, Xinhui, et al. Infection status of pathogens in Citellus undulatus in Guertu, Xinjiang. Disease Surveillance (2020) 35:623–626. doi: 10.3784/j.issn.1003-9961.2020.07.015
14. Han, X, Zhao, S, Liu, Z, Zhang, Y, Zhao, G, Zhang, C, et al. Bartonella, Blechomonas and Trypanosoma in fleas from the long-tailed ground squirrel (Spermophilus undulatus) in northwestern China. Int J Parasitol Parasites Wildl. (2024) 24:100958. doi: 10.1016/j.ijppaw.2024.100958
15. Demina, TV, Tkachev, SE, Kozlova, IV, Doroshchenko, EK, Lisak, OV, Suntsova, OV, et al. Comparative analysis of complete genome sequences of European subtype tick-borne encephalitis virus strains isolated from Ixodes persulcatus ticks, long-tailed ground squirrel (Spermophilus undulatus), and human blood in the Asian part of Russia. Ticks Tick Borne Dis. (2017) 8:547–53. doi: 10.1016/j.ttbdis.2017.03.002
16. Li, LL, Liu, MM, Shen, S, Zhang, YJ, Xu, YL, Deng, HY, et al. Detection and characterization of a novel hepacivirus in long-tailed ground squirrels (Spermophilus undulatus) in China. Arch Virol. (2019) 164:2401–10. doi: 10.1007/s00705-019-04303-z
17. Zhao, S, Yang, M, Jiang, M, Yan, B, Zhao, S, Yuan, W, et al. Rickettsia raoultii and Rickettsia sibirica in ticks from the long-tailed ground squirrel near the China-Kazakhstan border. Exp Appl Acarol. (2019) 77:425–33. doi: 10.1007/s10493-019-00349-5
18. Elangbam, CS, Qualls, CW Jr, Lochmiller, RL, and Boggs, JF. Strongyloidiasis in cotton rats (Sigmodon hispidus) from Central Oklahoma. J Wildl Dis. (1990) 26:398–402. doi: 10.7589/0090-3558-26.3.398
19. Nicolas, V, Quérouil, S, Verheyen, E, Verheyen, W, Mboumba, JF, Dillen, M, et al. Mitochondrial phylogeny of African wood mice, genus Hylomyscus (Rodentia, Muridae): implications for their taxonomy and biogeography. Mol Phylogenet Evol. (2006) 38:779–93. doi: 10.1016/j.ympev.2005.11.024
20. Liu, Q. General account: Chapter on morphology and anatomy In: H Wu, editor. Fauna Sinica: Insecta. Siphonaptera. China: Science Press (2007). 359–61.
21. Maekawa, K, Kitade, O, and Matsumoto, T. Molecular phylogeny of orthopteroid insects based on the mitochondrial cytochrome oxidase II gene. Zoologica (Stuttg). (1999) 16:175–84. doi: 10.2108/zsj.16.175
22. Johnson, KP, Cruickshank, RH, Adams, RJ, Smith, VS, Page, RD, and Clayton, DH. Dramatically elevated rate of mitochondrial substitution in lice (Insecta: Phthiraptera). Mol Phylogenet Evol. (2003) 26:231–42. doi: 10.1016/s1055-7903(02)00342-1
23. Yoshizawa, K, and Johnson, KP. How stable is the “Polyphyly of lice” hypothesis (Insecta: Psocodea)?: a comparison of phylogenetic signal in multiple genes. Mol Phylogenet Evol. (2010) 55:939–51. doi: 10.1016/j.ympev.2010.02.026
24. Han, HJ, Liu, JW, Wen, HL, Qin, XR, Zhao, M, Wang, LJ, et al. Babesia vesperuginis in insectivorous bats from China. Parasit Vectors. (2018) 11:317. doi: 10.1186/s13071-018-2902-9
25. Fard, SN, and Khalili, M. PCR-detection of Coxiella burnetii in ticks collected from sheep and goats in Southeast Iran. Iran J Arthropod Borne Dis. (2011) 5:1–6.
26. Zhao, S, Yang, M, Liu, G, Hornok, S, Zhao, S, Sang, C, et al. Rickettsiae in the common pipistrelle Pipistrellus pipistrellus (Chiroptera: Vespertilionidae) and the bat soft tick Argas vespertilionis (Ixodida: Argasidae). Parasit Vectors. (2020) 13:10. doi: 10.1186/s13071-020-3885-x
27. Xu, B, Gu, M, Wu, Q, Shu, C, Tan, W, Wang, S, et al. The bacterial patterns suggesting the dynamic features of tick-associated microorganisms in hard ticks. BMC Microbiol. (2024) 24:179. doi: 10.1186/s12866-024-03323-3
28. Li, E, Wu, X, Tang, L, Yang, M, Hornok, S, Zhang, C, et al. Molecular-phylogenetic analyses of Babesia and Theileria species from small mammals and their ticks in northern China suggest new reservoirs of bovine and equine piroplasms. Vet Parasitol. (2024) 332:110304. doi: 10.1016/j.vetpar.2024.110304
29. Chen, P, Xu, H, Tang, H, Zhao, F, Yang, C, Kwok, LY, et al. Modulation of gut mucosal microbiota as a mechanism of probiotics-based adjunctive therapy for ulcerative colitis. Microb Biotechnol. (2020) 13:2032–43. doi: 10.1111/1751-7915.13661
30. Brown, LD, and Macaluso, KR. Rickettsia felis, an emerging flea-borne rickettsiosis. Curr Trop Med Rep. (2016) 3:27–39. doi: 10.1007/s40475-016-0070-6
31. Zeroual, F, Leulmi, H, Benakhla, A, Raoult, D, Parola, P, and Bitam, I. Molecular evidence of Rickettsia slovaca in wild boar lice, in northeastern Algeria. Vector Borne Zoonotic Dis. (2018) 18:114–6. doi: 10.1089/vbz.2017.2165
32. Špitalská, E, Kraljik, J, Miklisová, D, Boldišová, E, Sparagano, OAE, and Stanko, M. Circulation of Rickettsia species and rickettsial endosymbionts among small mammals and their ectoparasites in eastern Slovakia. Parasitol Res. (2020) 119:2047–57. doi: 10.1007/s00436-020-06701-8
33. Obiegala, A, Oltersdorf, C, Silaghi, C, Kiefer, D, Kiefer, M, Woll, D, et al. Rickettsia spp. in small mammals and their parasitizing ectoparasites from Saxony, Germany. Vet Parasitol Reg Stud Reports. (2016) 5:19–24. doi: 10.1016/j.vprsr.2016.08.008
34. Orkun, Ö, and Çakmak, A. Molecular identification of tick-borne bacteria in wild animals and their ticks in Central Anatolia. Turkey Comp Immunol Microbiol Infect Dis. (2019) 63:58–65. doi: 10.1016/j.cimid.2018.12.007
35. Borşan, SD, Ionică, AM, Galon, C, Toma-Naic, A, Peştean, C, Sándor, AD, et al. High diversity, prevalence, and co-infection rates of tick-borne pathogens in ticks and wildlife hosts in an urban area in Romania. Front Microbiol. (2021) 12:645002. doi: 10.3389/fmicb.2021.645002
36. Liesner, JM, Krücken, J, Schaper, R, Pachnicke, S, Kohn, B, Müller, E, et al. Vector-borne pathogens in dogs and red foxes from the federal state of Brandenburg. Germany Vet Parasitol. (2016) 224:44–51. doi: 10.1016/j.vetpar.2016.05.012
37. Muz, MN, Erat, S, and Mumcuoglu, KY. Protozoan and microbial pathogens of house cats in the province of Tekirdag in Western Turkey. Pathogens. (2021) 10:1114. doi: 10.3390/pathogens10091114
38. Fischer, S, Spierling, NG, Heuser, E, Kling, C, Schmidt, S, Rosenfeld, UM, et al. High prevalence of Rickettsia helvetica in wild small mammal populations in Germany. Ticks Tick Borne Dis. (2018) 9:500–5. doi: 10.1016/j.ttbdis.2018.01.009
39. Arricau Bouvery, N, Souriau, A, Lechopier, P, and Rodolakis, A. Experimental Coxiella burnetii infection in pregnant goats: excretion routes. Vet Res. (2003) 34:423–33. doi: 10.1051/vetres:2003017
40. Yessinou, RE, Katja, MS, Heinrich, N, and Farougou, S. Prevalence of Coxiella-infections in ticks - review and meta-analysis. Ticks Tick Borne Dis. (2022) 13:101926. doi: 10.1016/j.ttbdis.2022.101926
41. Huang, HHH, Power, RI, Mathews, KO, Ma, GC, Bosward, KL, and Šlapeta, J. Cat fleas (Ctenocephalides felis clade ‘Sydney’) are dominant fleas on dogs and cats in New South Wales, Australia: presence of flea-borne Rickettsia felis, Bartonella spp but absence of Coxiella burnetii DNA. Curr Res Parasitol Vector Borne Dis. (2021) 1:100045. doi: 10.1016/j.crpvbd.2021.100045
42. Reeves, WK, Loftis, AD, Szumlas, DE, Abbassy, MM, Helmy, IM, Hanafi, HA, et al. Rickettsial pathogens in the tropical rat mite Ornithonyssus bacoti (Acari: Macronyssidae) from Egyptian rats (Rattus spp.). Exp Appl Acarol. (2007) 41:101–7. doi: 10.1007/s10493-006-9040-3
43. Psaroulaki, A, Chochlakis, D, Ioannou, I, Angelakis, E, and Tselentis, Y. Presence of Coxiella burnetii in fleas in Cyprus. Vector Borne Zoonotic Dis. (2014) 14:685–7. doi: 10.1089/vbz.2013.1399
44. Thompson, M, Mykytczuk, N, Gooderham, K, and Schulte-Hostedde, A. Prevalence of the bacterium Coxiella burnetii in wild rodents from a Canadian natural environment park. Zoonoses Public Health. (2012) 59:553–60. doi: 10.1111/j.1863-2378.2012.01493.x
Keywords: Spermophilus undulatus, flea, louse, spotted fever group rickettsiae (SFGR), Coxiella burnetii
Citation: Han X, Liu Z, Jiang Z, Zhao S, Hornok S, Yang M, Liu G and Wang Y (2025) Detection of spotted fever group rickettsiae and Coxiella burnetii in long-tailed ground squirrels (Spermophilus undulatus) and their ectoparasites. Front. Vet. Sci. 12:1553152. doi: 10.3389/fvets.2025.1553152
Edited by:
Vesna Milicevic, University of Belgrade, SerbiaReviewed by:
Danica Bogunović, University of Belgrade, SerbiaCopyright © 2025 Han, Liu, Jiang, Zhao, Hornok, Yang, Liu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gang Liu, MTUxOTgxNjYxMkBxcS5jb20=; Yuanzhi Wang, d2FuZ3l1YW56aGk2MjFAMTI2LmNvbQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.