 Junna Wang
Junna Wang Jin Cui
Jin Cui Guoxin Li
Guoxin Li Lingxue Yu
Lingxue Yu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci. , 03 March 2025
Sec. Veterinary Infectious Diseases
Volume 12 - 2025 | https://doi.org/10.3389/fvets.2025.1535328
This article is part of the Research Topic The application of new technologies such as new vaccines, therapeutic cytokines and antibodies, and antiviral drugs in the prevention and treatment of animal infectious diseases View all 15 articles
In recent years, replication-deficient viral vector vaccines have attracted much attention in the field of vaccine research and development due to their high safety and immunogenicity. These vaccines use genetic modifications to engineer viral vectors that make them unable to replicate but effective in expressing recombinant proteins and induce immune responses. Currently, replication-deficient adenovirus vectors and poxvirus vectors are widely used in vaccine R&D for a variety of infectious diseases in humans and animals, including AIDS, hepatitis B, pseudorabies, avian influenza, infectious bronchitis in poultry, and foot-and-mouth disease. Replication-deficient viral vaccines have been shown to effectively induce neutralizing antibodies and cellular immune responses, thereby providing effective immune protection. Future development of genetic engineering technology and continuous in-depth research on viral vectors should lead to replication-deficient viral vector platforms that have an essential role in preventing and controlling existing and emerging infectious diseases.
Viral vector vaccines utilize modified viruses, including recombinant attenuated and non-host viruses to deliver antigens, thereby eliciting an immune response (1). These vaccines have demonstrated efficacy across various applications, with notable examples including adenovirus [Adv; (2)], modified vaccinia virus (pox) (3), and turkey herpesvirus [HVT; (4, 5)]. However, traditional live attenuated viral vector vaccines face challenges mainly safety concerns, such as potential reversion to virulence, and uncontrolled shedding into the environment (6, 7). Replication-defective vectors address these issues by enhancing safety, making them an ideal choice for the next generation of vaccines.
Replication-deficient viruses are genetically modified viruses lacking functions essential for the replication of one or more viral genomes or virion synthesis and assembly in which the critical replication gene regions in the genomes are dysfunctional (8). Such viruses cannot complete the replication process independently and must rely on exogenous proteins provided by the host cell for assembly into mature viral particles. Once infecting host cells, they can only undergo one round of infection without continuing to replicate themselves, and this dependence explains why replication-deficient viruses exhibit a high level of biosecurity. In normal cells, one or more steps of viral replication are blocked, resulting in viral gene expression within infected cells without producing progeny virus (9). The tight blockade of these mutant viruses in replication makes their high level of safety one of the major advantages of using replication-deficient mutant viruses as vaccine vectors.
Current research on replication-deficient viruses also includes the study of single-cycle mutant viruses. By contrast, a single-cycle infectious virus is somewhat similar to a viral vaccine but is technically different. It is defective in the virion proteins that function after virus assembly and multiplies in complementary cells that express the deletion gene product. First infection, the replication cycle proceeds normally and produces progeny virions. However, since these virions are non-infectious, the infection cannot spread to the second round of cells, but the released non-infectious virions may provide inert antigens that can spread beyond the infected cells, thereby potentially leading to incomplete blocking of viral transmission (10).
Replication-deficient recombinant adenoviruses are currently the most widely investigated and applied replication-deficient vaccine vectors. Adenoviruses are non-enveloped double-stranded DNA viruses with a genome length of ~36 kb in which the first generation adenoviral vectors become replication-deficient by deleting the E1 and E3 genomic proteins necessary for viral replication and gene expression while inserting transgene expression sites, resulting in high immunogenicity (11, 12). Second generation adenoviral vectors further delete the E2 and E4 genomic regions, increasing their carrying capacity while maintaining high immunogenicity (13, 14), and third generation adenoviral vectors delete the entire adenoviral genome while retaining only the essential components to achieve high levels of gene expression but requiring helper adenoviruses for packaging (15). In this regard, the main drawback of adenoviral vectors is that the first generation adenoviral vectors, which lack the E1 protein and are the most widely used, can only insert up to 9 kb of exogenous gene sequences (16). Despite the ability of second and third generation vectors to accommodate larger exogenous gene sequences because more viral genes are deleted, the viral titer and expression levels of the exogenous genes are significantly lower than those of the first generation adenoviral vectors [(17, 18); Table 1].
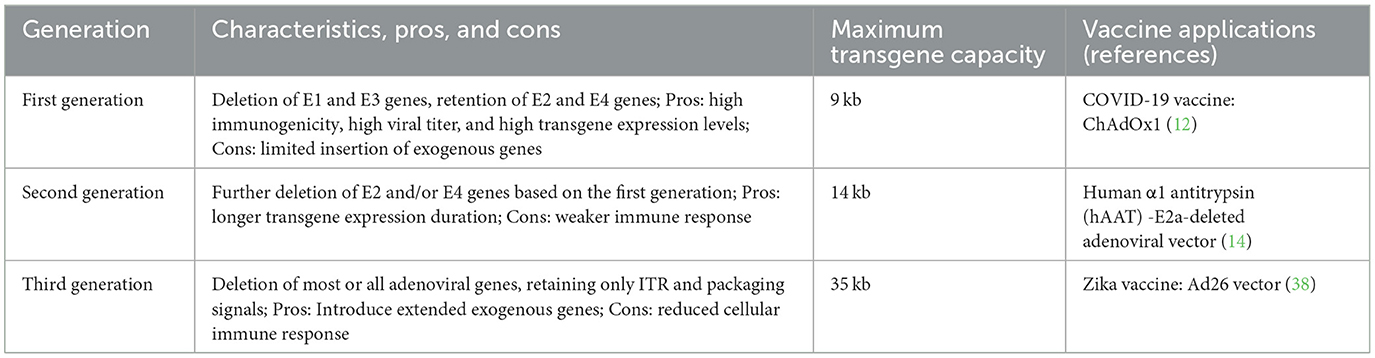
Table 1. The replication-deficient adenoviral vectors.
Replication-deficient recombinant adenovirus vaccines inhibit the progression of chronic Chagas's disease and reverse cardiac injury by reprogramming immune responses (19). A vaccine for blocking the transmission of malaria (AdPvs25) was developed by use of replication-deficient human adenovirus type 5 (rAd5) as a vector to express the surface protein (Pvs25) of the malignant Plasmodium parasite, and immunizing mice with this vaccine effectively induced antibodies against Pvs25. Compared with recombinant Pvs25 protein vaccine mixed with aluminum adjuvant, AdPvs25 had higher efficacy (20). Numerous studies have reported the application of Ad5 vector vaccines in human immunodeficiency virus type 1 (HIV-1), and Ad5 vaccines can induce potent and long-lasting CD8+ T-cell responses and specific cytotoxic T-cell responses (21). Additionally, other researchers found that a combination of DNA vaccine immunization followed by immunization with a replication-deficient adenoviral vector HIV-1 vaccine (Ad-Bal) induced higher humoral and cell-mediated immune responses (22).
The coronavirus disease 2019 (COVID-19) pandemic caused by the SARS-CoV-2 coronavirus in 2019 posed a significant challenge to healthcare systems worldwide, and investigators constructed a vaccine (Ad5-nCoV) that used the Ad5 adenoviral vector to express the S protein of SARS-CoV-2, assessed the vaccine's immunogenicity and protective efficacy in BALB/c mice and ferrets as animal models, and compared two different routes of administration: intramuscular injection and mucosal inoculation. The results indicated that a single dose of mucosal vaccination protects mice and ferrets from infection and inhibits virus replication in the upper respiratory tract. In comparison, intramuscular immunizations is insufficient to provide complete protection for the upper respiratory tract of mice against viral infections [(23); Table 2].
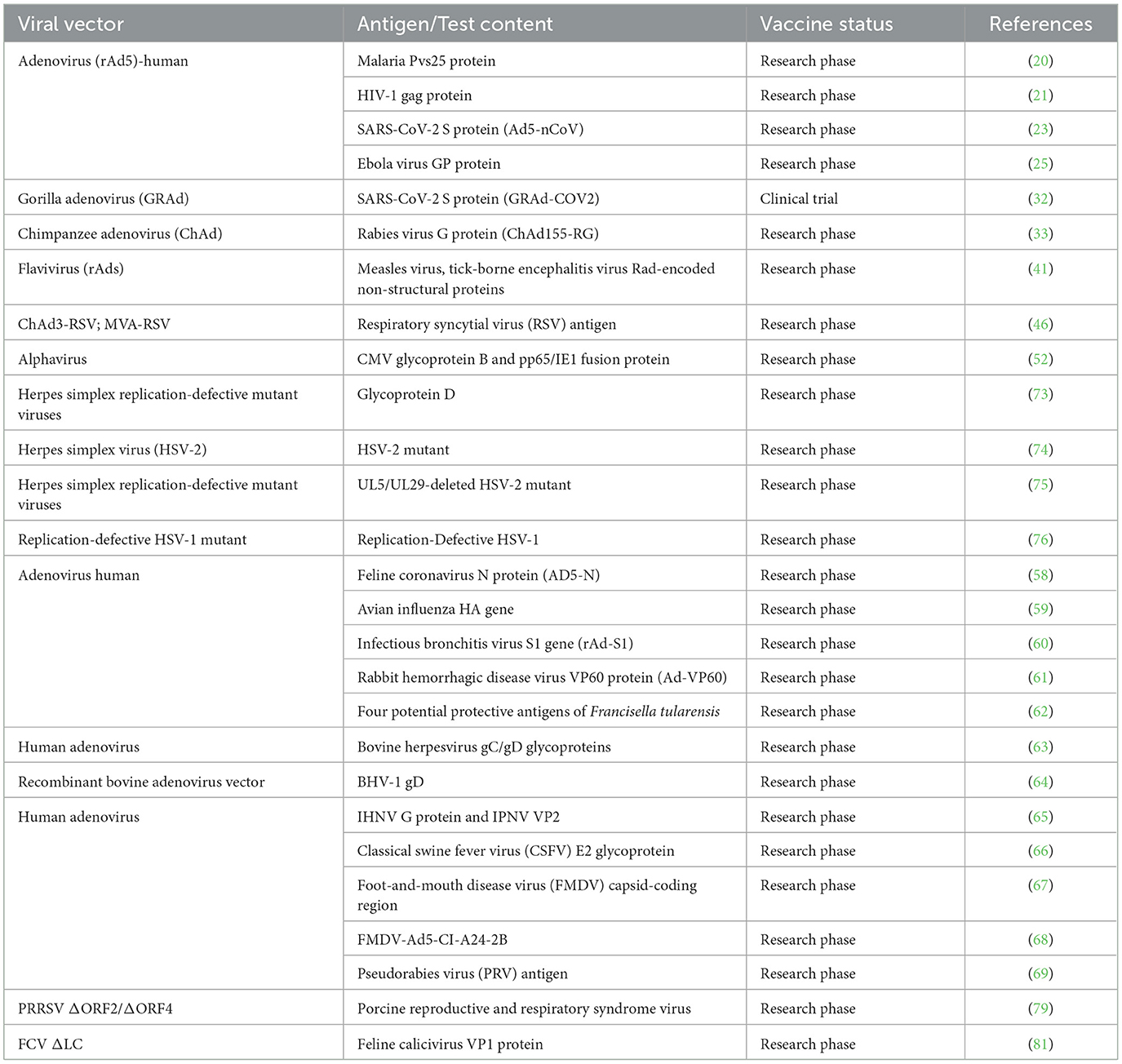
Table 2. A summary of examples of defective virus vaccines.
Adenoviruses derived from non-human primates (NHP) share a similar genomic structure with human adenoviruses, albeit with some differences, such as sequence diversity in the major capsid proteins and E3 region (24). Compared with those of human adenoviruses, the immune responses induced by NHP adenoviruses may be more versatile and generate more memory T cells, which could help enhance the persistence of antitumor immunity. In this regard, NHP adenoviruses can serve as a valuable resource for developing novel oncolytic adenoviral vectors (24).
Researchers have developed vaccines based on human adenovirus type 5 (rAd5) that can provide protection against acute lethal Ebola virus (EBOV) challenges in macaques (25). However, such vaccines fail to protect macaques with preexisting Ad5 antibodies, indicating that prior infection with wild-type Ad5 may limit the vaccine's immunogenic efficacy in humans. Although adenoviruses from non-human primates (including chimpanzees and apes) may overcome this limitation and have shown promise for protection against the EBOV, the effect is limited to modified viruses in mice (25).
Adenoviruses have been isolated from chimpanzees, bonobos, gorillas, and macaques (26). Replication-deficient vector vaccine have been developed on the basis of the adenovirus backbone from gorillas and chimpanzees (27–31). One candidate vaccine, GRAd-COV2, is based on a replication-deficient gorilla adenoviral vector that stably expresses the spike protein of the SARS-CoV-2 virus. The results from postvaccination clinical trials showed that GRAd-COV2 was well-tolerated at all doses, with most participants in young and elderly groups producing sustained specific antibodies that neutralized variant strains. Additionally, they also produced potent T-cell responses targeting the spike protein, with mild and short-lasting side effects (32). A chimpanzee-derived replication-deficient adenovirus (ChAd) was utilized to express the glycoprotein (G protein) of the rabies virus, followed by the development of a novel rabies vaccine, ChAd155-RG, for serotype C, the administration method for which is simpler than existing methods. In addition, the vaccine exhibited a robust and enduring neutralizing antibody response in non-human primates, demonstrating superior protective efficacy (33).
The broad applicability of adenoviral vectors is further exemplified in the development of flavivirus vaccines (34, 35). Members of the Flavivirus family are responsible for various diseases in humans, including dengue fever, yellow fever, Japanese encephalitis, tick-borne encephalitis, Zika virus, and St. Louis encephalitis, etc. (36). Replication-deficient recombinant adenoviruses (rAds) have potential as flavivirus vaccines (37–40). For instance, animal model experiments using rAd vaccines for the measles virus and tick-borne encephalitis virus have induced good humoral and cellular immune responses, ultimately protecting animals from viral infections. rAds can express non-structural proteins of flaviviruses, effectively stimulating strong cellular immune responses chronic viral infections (41).
Poxvirus is a large, enveloped double-stranded DNA virus with a genome size of 130–230 kb, and the prototype virus is the causative agent of smallpox (42). The Modified Vaccinia virus Ankara (MVA) strain, derived from an attenuated vaccinia virus strain, is a highly effective and safe replication-deficient viral vector platform (3). Previous studies on recombinant MVA vaccines have confirmed their protective efficacy, indicating the promising potential of this platform for vaccine development (43–45). Research has involved the vaccination of healthy adults with two different replication-deficient viral vector vaccines: the African green monkey adenovirus, ChAd3-RSV, and the Ankara vaccinia virus, MVA-RSV, both targeting respiratory syncytial virus (RSV; Table 2). Experimental data suggest that the MVA-RSV vaccine has immunogenicity and safety comparable to those of the ChAd3-RSV vaccine in healthy adults and can effectively induce humoral and cellular immune responses (46). The avian poxvirus is unable to produce progeny viruses when infecting mammalian cells (47).
Alphavirus is a member of the Togaviridae family (48), characterized as an enveloped virus with a single-stranded positive-sense RNA genome. The non-structural genes are translated from the genomic RNA once it is released into the cytoplasm, forming replication complexes that drive the production of negative-sense antigenomic RNA. This process also involves the transcription of full-length positive-sense genomic RNA and shorter subgenomic RNA. As genome-wide RNAs are replicated, more templates are transcribed via subgenomic promoters, leading to the exponential amplification of subgenomic RNAs and rapid proliferation of subgenomic mRNAs (49). The use of alphavirus vaccine vectors is increasing because of their broad range of host cells, tropism for dendritic cells and monocytes, high levels of gene expression capability (50), and immunity to very few individuals, which makes them very promising candidates for replication-deficient vectors (51).
Investigators have used a replication-deficient, single-cycle, single-capsid alphavirus replicon vector system (52) to construct viral-like replicon particles expressing the extracellular domains of cytomegalovirus glycoprotein B and pp65/IE1 fusion proteins, which induced high-titer antibody responses and strong cellular immune responses in mice and rabbits while demonstrating satisfactory immunogenicity and safety [(53); Table 2].
In recent years, various viral vaccines for animal diseases have been continuously developed to create safer and more effective novel vaccines (54). Replication-deficient adenovirus vaccines have demonstrated higher safety and more potent immunogenicity than traditional vaccines in clinical prevention and protection against many diseases, with several adenovirus vector vaccines from different species exhibiting superior immunological effects in immunization experiments conducted in mice and other animals (55). The main viral vectors include human adenovirus, chimpanzee adenovirus, and avian adenovirus, and there have been numerous applications in animal diseases beyond humans, such as swine influenza virus, porcine epidemic diarrhea (56), small ruminant viral diseases (57), and foot-and-mouth disease.
Previous laboratory studies on feline infectious peritonitis (FCoV) found limited efficacy in the current vaccines against FCoV, primarily due to antibody-dependent enhancement in affected cats, and the nucleocapsid (N) protein of FCoV was constructed as a novel adenovirus vector (AD5-N) to develop a new vaccine to address this issue (Table 2). Compared with the control group, the AD5-N immunized animals group exhibited a strong cellular immune response and specifically induced high levels of IgG and SIgA antibodies, as well as showed evidently reduced viral loads in their feces and intestinal tissues after challenge assays. The experimental results indicated that the AD5-N vaccine provided effective protection for cats against FCoV infection (58).
Regarding avian diseases research, investigators have inserted the HA genes of different subtypes of avian influenza virus (AIV) into the HAdV5 vector, constructing replication-deficient adenoviruses that express various subtype genes (Table 2). In mouse models, polyvalent adenovirus vaccines can induce broad-spectrum immune responses and provide cross-protection against multiple subtypes of avian influenza virus (59). Aside from avian influenza viruses, the currently used infectious bronchitis virus (IBV) vaccines for poultry encounter issues, such as low cost-effectiveness and insufficient protective efficacy, particularly against nephropathogenic IBV (Table 2). In view of this, a recombinant adenovirus vaccine (rAd-S1) expressing the S1 gene of nephropathogenic IBV that effectively induces humoral and cellular immune responses in vaccinated chickens while producing antibodies and cytokines against IBV has been developed. Compared with the control group, chickens in the rAd-S1 group exhibited significantly relieved renal lesions and clinical symptoms after challenge assays, with evident decreased mortality (60).
Regarding infectious diseases in rabbits, some researchers have constructed an adenovirus vector vaccine (Ad-VP60) that efficiently expresses the VP60 protein of rabbit hemorrhagic disease virus (RHDV; Table 2). This Ad-VP60 vaccine exhibits stronger immunogenicity and induces high levels of anti-RHDV antibodies in immunized mice and rabbits. Additionally, rabbits immunized via the nasal route showed saliva containing IgA antibodies comparable to those found in sera from rabbits vaccinated with inactivated RHDV vaccines (61). Rabbit fever, also known as rabbit plague or Tularemia, is a naturally occurring infectious disease caused by Francisella tularensis, which is a highly contagious Gram-negative pathogen capable of infecting various mammals, including rodents, wild rabbits, domestic rabbits, and humans. Researchers have developed a subunit vaccine and an adenovirus vector vaccine expressing four potential protective antigens from F. tularensis (Table 2). In mouse experiments, the adenovirus vector vaccine was able to induce protective immunity after a single immunization and stimulated an immune response characterized by Th1 cell cytokines, which had significantly higher protective efficacy than the subunit vaccine (62).
Investigators have constructed a vaccine by inserting glycoprotein C (gC) or glycoprotein D (gD) of bovine herpesvirus type 1 (BHV-1) into the HAd5 vector (Table 2), followed by assessing its protective efficacy in rabbits and cattle through intranasal and intramuscular administration, with intranasal injection in rabbits showing higher BHV-1 neutralizing antibody levels than those in the intramuscular group and the intranasal injection in cattle showing higher levels than those in the commercial live vaccine group (63). Additionally, there are some reports of modifying bovine adenovirus to express BHV-1 gD, constructing replication-capable and -deficient recombinant bovine adenoviruses (Table 2). Animal experiments found that the replication-capable virus provided partial protection and reduced shedding without completely preventing BHV-1 infection. By contrast, the replication-deficient virus could stimulate an immune memory response without providing protection (64).
For aquatic animals, prevention of infectious hematopoietic necrosis virus (IHNV) and infectious pancreatic necrosis virus (IPNV) in rainbow trout is critically important. Investigators successfully coexpressed the G protein of IHNV and the VP2 protein of IPNV by inserting them into a recombinant adenovirus vector (Table 2). After immersion immunization of juvenile fish, the immune levels were substantially elevated, producing high levels of neutralizing antibodies against IHNV and IPNV. Challenge tests indicated that the recombinant adenovirus vaccine exhibited high protection rates against IHNV and IPNV, with relative survival rates of 81.25% and 78.95%, respectively (65).
Immunogenic results from an adenovirus vaccine expressing the E2 glycoprotein of classical swine fever virus (CSFV) in pigs supported the construction of a recombinant adenovirus rAd-E2-InvC by coexpressing the C-terminal domain of the invasion protein (InvC) from Yersinia enterocolitica with the E2 glycoprotein (Table 2). Pigs immunized with rAd-E2-InvC did not show detectable virus or pathological changes upon necropsy after infection with CSFV, demonstrating complete resistance to virulent infection. By contrast, pigs in the rAd-E2 group exhibited clinical symptoms and viral nucleic acids, indicating that the coexpression group can enhance the efficacy of HAdV-based CSFV vaccines (66).
Investigators also have inserted the capsid-coding region of foot-and-mouth disease virus (FMDV) into an adenovirus vector and added an Arg-Gly-Asp sequence to fibronectin, and the fiber-modified adenovirus vector vaccine was able to express FMDV capsid proteins more effectively in vitro, particularly in cells lacking the Coxackie and adenovirus receptor (Table 2). However, no significant differences were observed in protective efficacy between the 2 vaccine types in bovine in vivo experiments (67). Additionally, Poly ICLC was used as an adjuvant in combination with a replication-deficient human adenovirus vector FMD vaccine (Ad5-CI-A24-2B). After immunization and challenge, the animals showing no detectable FMDV-specific neutralizing antibodies were still protected because Poly ICLC enhanced the specific T-cell response. Given this, Poly ICLC can serve as an adjuvant for the Ad5-FMD vaccine, reducing the vaccine dosage, improving immunogenicity, and lowering production costs (68).
Other studies have also found that piglets immunized via intramuscular injection with a recombinant adenovirus (rAd) vaccine for pseudorabies virus (PRV) could effectively resist PRV infection (Table 2), regardless of whether or not the dam was immune. However, piglets vaccinated intranasally were poorly protected. Particularly, the intramuscular administration of the rAd vaccine effectively induced anti-PRV immune responses in newborn piglets, even in the presence of maternal antibodies (69).
Herpes simplex virus (HSV) is a common human disease pathogen that causes mucosal infections, spreads to sensory neurons, and establishes latent infections, which lead to recurrent diseases (70). In particular, HSV infections occur throughout the carrier's lifetime and manifest as various clinical syndromes, including herpes labialis, herpetic whitlow, eczema herpeticum, keratitis, encephalitis, and genital diseases (71). In individuals with compromised immune systems, such as patients with AIDS, HSV infections can lead to severe illnesses (72).
Herpes simplex virus type 2 (HSV-2), a sexually transmitted virus, is the most common cause of genital diseases worldwide. There is a current lack of effective treatment options, making vaccine development crucial. Various forms of replication-deficient mutant HSV-2 viruses have been tested as vaccine strains. In mouse models of HSV-2 infection, the protective effect of live vaccines was proportional to the vaccination dose. With increasing doses, viral replication in the reproductive tract, disease severity and mortality rates were remarkably reduced (73). Another group of investigators compared the protective effects of a replication-deficient vaccine and a subunit vaccine via different immunization routes, and the replication-deficient vaccine had significant advantages in protecting vaginal mucosa and dorsal root ganglia from infection. Additionally, intramuscular injection produced immunogenicity and protective effects higher than those of subcutaneous injection in genital HSV-2 infection models (74). Moreover, an assessment of the protective efficacy of replication-deficient vaccines lacking viral replication genes (UL5 and UL29) and subunit vaccines in guinea pigs and rabbits indicated good safety and tolerability (75). Subcutaneous immunization with replication-deficient herpes simplex virus type 1 (HSV-1) mutants has also been found to reduce viral infections in the cornea and latent infections in the trigeminal ganglia while inducing persistent immunity. They exhibited good safety and avoided the risks associated with replication-competent viruses (76).
Currently, PRRSV vaccines (77) have issues with safety (live attenuated vaccines may regain virulence) and efficacy [unsatisfactory efficacy of inactivated vaccines; (78)]. Since the deletion of ORF2 and ORF4 genes results in the virus being unable to replicate in MARC-145 cells and porcine alveolar macrophages, investigators successfully constructed two full-length infectious cDNA clones of gene-deleted viruses: ΔORF2-PRRSV and ΔORF4-PRRSV. At the same time, two complementary cell lines were constructed to stably express GP2 and GP4 proteins to rescue the viruses lacking these genes. Additionally, although pigs immunized with ΔORF2-PRRSV and ΔORF4-PRRSV showed reduced viral loads after challenge assays, there was no significant improvement in their clinical symptoms (79).
Notably, African Swine Fever (ASF) is a highly contagious viral disease that causes substantial economic losses in the global pig industry. The ASF virus has multiple genotypes, and the high variability of its genes presents great challenges for vaccine protection. Despite the ability to induce immune responses and provide long-term protection against homologous ASFV infections, live attenuated ASFV vaccines fail to protect against heterologous strains, resulting in very limited protection. Additionally, attenuated strains developed by deleting virulence-related genes, such as ASFV-G-ΔI177L/ΔLVR and BA71ΔCD2, which can stably replicate in COS-1 cell lines and provide cross-protection are already known (80).
Reports have indicated the successful rescue of a virus containing the genome-wide feline calicivirus (FCV), named rBAC-FCV, with deletions made to the LC gene associated with FCV virulence, resulting in the infectious clone pBAC-FCV-ΔLC. Virus rescue using the feline kidney cell line F81-VP1, which expresses the FCV-VP1 protein, was conducted, followed by obtaining the LC gene-deleted strain rBAC-FCV-ΔLC. Additionally, cell experiments confirmed that deletion of the LC gene resulted in reduced viral virulence, and the virus lacking the LC gene could only replicate in cell lines that provide the VP1 protein, thereby significantly enhancing its safety. In conclusion, the successful rescue of this replication-deficient virus supports the use of alternative vaccine strains for developing novel FCV vaccines based on reverse genetics technology (81).
Vaccines are one of the most critical tool-kit to prevent infectious diseases. There are various types of existing vaccines available commercially (82); traditional inactivated vaccines are considered safe, but often fail to elicit strong T-cell responses (83), whereas live attenuated vaccines can produce potent antibody and cellular immune responses, but carry potential risks of causing disease (84). By contrast, viral vector vaccines deliver genes from pathogens via viral vectors, to stimulate immune responses (85).
In recent years, replication-deficient viral vaccines have gained increasing attention. In the field of animal health. For example, there are currently no therapeutic drugs for feline infectious peritonitis, which makes the application of replication-deficient adenoviral vector vaccines that have demonstrated high levels of protection in animal experiments a promising research area. Moreover, aside from common replication-deficient adenoviral vector vaccines, single-cycle adenoviral vector vaccines induce higher antibody levels and more potent immune responses in animal experiments for EBOV (86). If these findings are confirmed, single-cycle adenoviral vector vaccines might have significant potential for infectious disease vaccines.
Currently, the commercially available PRRS (87) vaccines are mainly inactivated vaccines and live attenuated vaccines, neither of which achieves a balance between protective efficacy and safety (88). However, a significant portion of the strains in pigs are recombinant viruses derived from vaccine strains (89), and safety is the greatest advantage of developing replication-deficient vaccines (90) that can also stimulate humoral and cellular immunity in immunized animals, thereby compensating for the shortcomings of existing vaccines.
In conclusion, replication-deficient vaccines represent a transformative advancement within the rapidly evolving field of vaccinology. Through targeted genetic engineering, these platforms are being refined to achieve an optimal balance between immunogenicity and biosafety. As our understanding of pathogen biology and host immunity expands-complemented by breakthroughs in gene-editing technologies such as CRISPR and prime editing-replication-deficient vaccines are poised to assume a central role in mitigating both emerging and re-emerging infectious diseases.
Looking forward, the convergence of precision gene editing, structural virology, and computational immunology will propel the development and application of replication-deficient viral vectors and vaccines to address existing and emerging infectious disease.
JW: Writing – original draft, Writing – review & editing. JC: Writing – original draft, Writing – review & editing. GL: Funding acquisition, Project administration, Resources, Writing – original draft, Writing – review & editing. LY: Funding acquisition, Project administration, Resources, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. We gratefully acknowledge support from Shanghai Municipal Agriculture Science and Technology Project (Grant No. 2022-02-08-00-12-F01115), National Key Research and Development Program of China (2022YFD1800800), Natural Science Foundation of Shanghai (24ZR1479400), The Key Laboratory of Veterinary Biotechnology (No. BB1500010), Shanghai, P.R. China, and Agricultural-Science and Technology Innovation Program (CAAS-ASTIP-2021-SHVRI).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Small JC, Ertl HC. Viruses – from pathogens to vaccine carriers. Curr Opin Virol. (2011) 1:241–5. doi: 10.1016/j.coviro.2011.07.009
2. Feng L, Wang Q, Shan C, Yang C, Feng Y, Wu J, et al. An adenovirus-vectored COVID-19 vaccine confers protection from SARS-CoV-2 challenge in rhesus macaques. Nat Commun. (2020) 11:4207. doi: 10.1038/s41467-020-18077-5
3. Volz A, Sutter G. Modified Vaccinia virus Ankara. Adv Virus Res. (2017) 97:187–243. doi: 10.1016/bs.aivir.2016.07.001
4. Sadigh Y, Powers C, Spiro S, Pedrera M, Broadbent A, Nair V. Gallid herpesvirus 3 SB-1 strain as a recombinant viral vector for poultry vaccination. NPJ Vaccines. (2018) 3:21. doi: 10.1038/s41541-018-0056-6
5. Reemers S, Verstegen I, Basten S, Hubers W, van de Zande S. A broad spectrum HVT-H5 avian influenza vector vaccine which induces a rapid onset of immunity. Vaccine. (2021) 39:1072–9. doi: 10.1016/j.vaccine.2021.01.018
6. Brisse M, Vrba SM, Kirk N, Liang Y, Ly H. Emerging concepts and technologies in vaccine development. Front Immunol. (2020) 11:583077. doi: 10.3389/fimmu.2020.583077
7. Poria R, Kala D, Nagraik R, Dhir Y, Dhir S, Singh B, et al. Vaccine development: current trends and technologies. Life Sci. (2024) 336:122331. doi: 10.1016/j.lfs.2023.122331
8. Thomas CE, Ehrhardt A, Kay MA. Progress and problems with the use of viral vectors for gene therapy. Nat Rev Genet. (2003) 4:346–58. doi: 10.1038/nrg1066
9. Dudek T, Knipe DM. Replication-defective viruses as vaccines and vaccine vectors. Virology. (2006) 344:230–9. doi: 10.1016/j.virol.2005.09.020
10. Nogales A, Baker SF, Domm W, Martínez-Sobrido L. Development and applications of single-cycle infectious Influenza A Virus (sciIAV). Virus Res. (2016) 216:26–40. doi: 10.1016/j.virusres.2015.07.013
11. Tatsis N, Ertl HCJ. Adenoviruses as vaccine vectors. Mol Ther. (2004) 10:616–29. doi: 10.1016/j.ymthe.2004.07.013
12. Ramasamy MN, Minassian AM, Ewer KJ, Flaxman AL, Folegatti PM, Owens DR, et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): a single-blind, randomised, controlled, phase 2/3 trial. Lancet (2020) 396:1979–93. doi: 10.1016/S0140-6736(20)32466-1
13. Janssen JM, Liu J, Skokan J, Gonçalves MAFV, de Vries AAF. Development of an AdEasy-based system to produce first- and second-generation adenoviral vectors with tropism for CAR- or CD46-positive cells. J Gene Med. (2013) 15:1–11. doi: 10.1002/jgm.2687
14. O'Neal WK, Zhou H, Morral N, Aguilar-Cordova E, Pestaner J, Langston C, et al. Toxicological comparison of E2a-deleted and first-generation adenoviral vectors expressing α1-antitrypsin after systemic delivery. Hum Gene Ther. (1998) 9:1587–98. doi: 10.1089/hum.1998.9.11-1587
15. Gorziglia MI, Lapcevich C, Roy S, Kang Q, Kadan M, Wu V, et al. Generation of an adenovirus vector lacking E1, E2a, E3, and all of E4 except open reading frame 3. J Virol. (1999) 73:6048–55. doi: 10.1128/JVI.73.7.6048-6055.1999
16. Kay MA, Glorioso JC, Naldini L. Viral vectors for gene therapy: the art of turning infectious agents into vehicles of therapeutics. Nat Med. (2001) 7:33–40. doi: 10.1038/83324
17. Trivedi PD, Byrne BJ, Corti M. Evolving horizons: adenovirus vectors' timeless influence on cancer, gene therapy and vaccines. Viruses. (2023) 15:2378. doi: 10.3390/v15122378
18. Dormond E, Perrier M, Kamen A. From the first to the third generation adenoviral vector: what parameters are governing the production yield? Biotechnol Adv. (2009) 27:133–44. doi: 10.1016/j.biotechadv.2008.10.003
19. Pereira IR, Vilar-Pereira G, Marques V, da Silva AA, Caetano B, Moreira OC, et al. A human type 5 adenovirus-based Trypanosoma cruzi therapeutic vaccine re-programs immune response and reverses chronic cardiomyopathy. PLoS Pathog. (2015) 11:e1004594. doi: 10.1371/journal.ppat.1004594
20. Miyata T, Harakuni T, Sugawa H, Sattabongkot J, Kato A, Tachibana M, et al. Adenovirus-vectored Plasmodium vivax ookinete surface protein, Pvs25, as a potential transmission-blocking vaccine. Vaccine. (2011) 29:2720–26. doi: 10.1016/j.vaccine.2011.01.083
21. Casimiro DR, Chen L, Fu T-M, Evans RK, Caulfield MJ, Davies M-E, et al. Comparative immunogenicity in rhesus monkeys of DNA plasmid, recombinant vaccinia virus, and replication-defective adenovirus vectors expressing a human immunodeficiency virus type 1 gag gene. J Virol. (2003) 77:6305–13. doi: 10.1128/JVI.77.11.6305-6313.2003
22. Yoshida T, Okuda K, Xin K-Q, Tadokoro K, Fukushima J, Toda S, et al. Activation of HIV-1-specific immune responses to an HIV-1 vaccine constructed from a replication-defective adenovirus vector using various combinations of immunization protocols. Clin Exp Immunol. (2001) 124:445–52. doi: 10.1046/j.1365-2249.2001.01540.x
23. Wu S, Zhong G, Zhang J, Shuai L, Zhang Z, Wen Z, et al. A single dose of an adenovirus-vectored vaccine provides protection against SARS-CoV-2 challenge. Nat Commun. (2020) 11:4081. doi: 10.1038/s41467-020-17972-1
24. Bots STF, Hoeben RC. Non-human primate-derived adenoviruses for future use as oncolytic agents? Int J Mol Sci. (2020) 21:4821. doi: 10.3390/ijms21144821
25. Stanley DA, Honko AN, Asiedu C, Trefry JC, Lau-Kilby AW, Johnson JC, et al. Chimpanzee adenovirus vaccine generates acute and durable protective immunity against ebolavirus challenge. Nat Med. (2014) 20:1126–9. doi: 10.1038/nm.3702
26. Duncan M, Cranfield MR, Torano H, Kuete HM, Lee GP, Glenn A, et al. Adenoviruses isolated from wild gorillas are closely related to human species C viruses. Virology. (2013) 444:119–23. doi: 10.1016/j.virol.2013.05.041
27. Collignon C, Bol V, Chalon A, Surendran N, Morel S, van den Berg RA, et al. Innate immune responses to chimpanzee adenovirus vector 155 vaccination in mice and monkeys. Front Immunol. (2020) 11:579872. doi: 10.3389/fimmu.2020.579872
28. de Jong R, Stockhofe-Zurwieden N, Bonsing J, Wang K-F, Vandepaer S, Bouzya B, et al. ChAd155-RSV vaccine is immunogenic and efficacious against bovine RSV infection-induced disease in young calves. Nat Commun. (2022) 13:6142. doi: 10.1038/s41467-022-33649-3
29. Lu ZH, Dmitriev IP, Brough DE, Kashentseva EA, Li J, Curiel DT. A new gorilla adenoviral vector with natural lung tropism avoids liver toxicity and is amenable to capsid engineering and vector retargeting. J Virol. (2020) 94:e00265–20. doi: 10.1128/JVI.00265-20
30. Limbach K, Stefaniak M, Chen P, Patterson NB, Liao G, Weng S, et al. New gorilla adenovirus vaccine vectors induce potent immune responses and protection in a mouse malaria model. Malar J. (2017) 16:263. doi: 10.1186/s12936-017-1911-z
31. Dicks MDJ, Spencer AJ, Edwards NJ, Wadell G, Bojang K, Gilbert SC, et al. A novel chimpanzee adenovirus vector with low human seroprevalence: improved systems for vector derivation and comparative immunogenicity. PLoS ONE. (2012) 7:e40385. doi: 10.1371/journal.pone.0040385
32. Lanini S, Capone S, Antinori A, Milleri S, Nicastri E, Camerini R, et al. GRAd-COV2, a gorilla adenovirus-based candidate vaccine against COVID-19, is safe and immunogenic in younger and older adults. Sci Transl Med. (2022) 14:eabj1996. doi: 10.1126/scitranslmed.abj1996
33. Napolitano F, Merone R, Abbate A, Ammendola V, Horncastle E, Lanzaro F, et al. A next generation vaccine against human rabies based on a single dose of a chimpanzee adenovirus vector serotype C. PLoS Negl Trop Dis. (2020) 14:e0008459. doi: 10.1371/journal.pntd.0008459
34. Shoushtari M, Roohvand F, Salehi-Vaziri M, Arashkia A, Bakhshi H, Azadmanesh K. Adenovirus vector-based vaccines as forefront approaches in fighting the battle against flaviviruses. Hum Vaccin Immunother. (2022) 18:2079323. doi: 10.1080/21645515.2022.2079323
35. Bullard BL, Corder BN, Gordon DN, Pierson TC, Weaver EA. Characterization of a species E adenovirus vector as a Zika virus vaccine. Sci Rep. (2020) 10:3613. doi: 10.1038/s41598-020-60238-5
36. Luria-Pérez R, Sánchez-Vargas LA, Muñoz-López P, Mellado-Sánchez G. Mucosal vaccination: a promising alternative against flaviviruses. Front Cell Infect Microbiol. (2022) 12:887729. doi: 10.3389/fcimb.2022.887729
37. Jacobs SC, Stephenson JR, Wilkinson GW. High-level expression of the tick-borne encephalitis virus NS1 protein by using an adenovirus-based vector: protection elicited in a murine model. J Virol. (1992) 66:2086–95. doi: 10.1128/jvi.66.4.2086-2095.1992
38. Cox F, van der Fits L, Abbink P, Larocca RA, van Huizen E, Saeland E, et al. Adenoviral vector type 26 encoding Zika virus (ZIKV) M-Env antigen induces humoral and cellular immune responses and protects mice and nonhuman primates against ZIKV challenge. PLoS ONE. (2018) 13:e0202820. doi: 10.1371/journal.pone.0202820
39. Jaiswal S, Khanna N, Swaminathan S. Replication-defective adenoviral vaccine vector for the induction of immune responses to dengue virus type 2. J Virol. (2003) 77:12907–13. doi: 10.1128/JVI.77.23.12907-12913.2003
40. Appaiahgari MB, Saini M, Rauthan M, Jyoti, Vrati S. Immunization with recombinant adenovirus synthesizing the secretory form of Japanese encephalitis virus envelope protein protects adenovirus-exposed mice against lethal encephalitis. Microbes Infect. (2006) 8:92–104. doi: 10.1016/j.micinf.2005.05.023
41. Stephenson J. Defective adenoviruses as novel vaccines for the Flaviviridae. Clin Diagn Virol. (1998) 10:187–94. doi: 10.1016/S0928-0197(98)00038-5
42. Sasso E, D'Alise AM, Zambrano N, Scarselli E, Folgori A, Nicosia A. New viral vectors for infectious diseases and cancer. Semin Immunol. (2020) 50:101430. doi: 10.1016/j.smim.2020.101430
43. Breathnach CC, Clark HJ, Clark RC, Olsen CW, Townsend HGG, Lunn DP. Immunization with recombinant modified vaccinia Ankara (rMVA) constructs encoding the HA or NP gene protects ponies from equine influenza virus challenge. Vaccine. (2006) 24:1180–90. doi: 10.1016/j.vaccine.2005.08.091
44. Veits J, Römer-Oberdörfer A, Helferich D, Durban M, Suezer Y, Sutter G, et al. Protective efficacy of several vaccines against highly pathogenic H5N1 avian influenza virus under experimental conditions. Vaccine. (2008) 26:1688–96. doi: 10.1016/j.vaccine.2008.01.016
45. Orlova OV, Glazkova DV, Bogoslovskaya EV, Shipulin GA, Yudin SM. Development of modified vaccinia virus ankara-based vaccines: advantages and applications. Vaccines. (2022) 10:1516. doi: 10.3390/vaccines10091516
46. Green CA, Scarselli E, Sande CJ, Thompson AJ, De Lara CM, Taylor KS, et al. Chimpanzee adenovirus– and MVA-vectored respiratory syncytial virus vaccine is safe and immunogenic in adults. Sci Transl Med. (2015) 7:300. doi: 10.1126/scitranslmed.aac5745
47. Weli SC, Tryland M. Avipoxviruses: infection biology and their use as vaccine vectors. Virol J. (2011) 8:49. doi: 10.1186/1743-422X-8-49
48. Strauss JH, Strauss EG. The alphaviruses: gene expression, replication, and evolution. Microbiol Rev. (1994) 58:491–562. doi: 10.1128/mr.58.3.491-562.1994
49. Wang KS, Kuhn RJ, Strauss EG, Ou S, Strauss JH. High-affinity laminin receptor is a receptor for Sindbis virus in mammalian cells. J Virol. (1992) 66:4992–5001. doi: 10.1128/jvi.66.8.4992-5001.1992
50. Guerrero-Arguero I, Tellez-Freitas CM, Weber KS, Berges BK, Robison RA, Pickett BE. Alphaviruses: host pathogenesis, immune response, and vaccine & treatment updates. J Gen Virol. (2021) 102:001644. doi: 10.1099/jgv.0.001644
52. Reap EA, Dryga SA, Morris J, Rivers B, Norberg PK, Olmsted RA, et al. Cellular and humoral immune responses to alphavirus replicon vaccines expressing cytomegalovirus pp65, IE1, and gB proteins. Clin Vaccine Immunol. (2007) 14:748–55. doi: 10.1128/CVI.00037-07
53. Reap EA, Morris J, Dryga SA, Maughan M, Talarico T, Esch RE, et al. Development and preclinical evaluation of an alphavirus replicon particle vaccine for cytomegalovirus. Vaccine. (2007) 25:7441–9. doi: 10.1016/j.vaccine.2007.08.016
54. Ferreira TB, Alves PM, Aunins JG, Carrondo MJT. Use of adenoviral vectors as veterinary vaccines. Gene Ther. (2005) 12:S73–83. doi: 10.1038/sj.gt.3302618
55. Miao X, Zhang L, Zhou P, Zhang Z, Yu R, Liu X, et al. Recombinant human adenovirus type 5 based vaccine candidates against GIIa- and GIIb-genotype porcine epidemic diarrhea virus induce robust humoral and cellular response in mice. Virology. (2023) 584:9–23. doi: 10.1016/j.virol.2023.05.001
56. Liu X, Zhao D, Zhou P, Zhang Y, Wang Y. Evaluation of the efficacy of a recombinant adenovirus expressing the spike protein of porcine epidemic diarrhea virus in pigs. Biomed Res Int. (2019) 2019:8530273. doi: 10.1155/2019/8530273
57. Rojas JM, Sevilla N, Martín V. A new look at vaccine strategies against PPRV focused on adenoviral candidates. Front Vet Sci. (2021) 8:729879. doi: 10.3389/fvets.2021.729879
58. Wang Y, Liu Y, Wang J, Zhang M, Deng X, Song J, et al. An adenovirus-vectored vaccine based on the N protein of feline coronavirus elicit robust protective immune responses. Antiviral Res. (2024) 223:105825. doi: 10.1016/j.antiviral.2024.105825
59. Vemula SV, Ahi YS, Swaim A-M, Katz JM, Donis R, Sambhara S, et al. Broadly protective adenovirus-based multivalent vaccines against highly pathogenic avian influenza viruses for pandemic preparedness. PLoS ONE. (2013) 8:e62496. doi: 10.1371/journal.pone.0062496
60. Zeshan B, Zhang L, Bai J, Wang X, Xu J, Jiang P. Immunogenicity and protective efficacy of a replication-defective infectious bronchitis virus vaccine using an adenovirus vector and administered in ovo. J Virol Methods. (2010) 166:54–9. doi: 10.1016/j.jviromet.2010.02.019
61. Fernández E, Toledo JR, Chiong M, Parra F, Rodríguez E, Montero C, et al. Single dose adenovirus vectored vaccine induces a potent and long-lasting immune response against rabbit hemorrhagic disease virus after parenteral or mucosal administration. Vet. Immunol. Immunopathol. (2011) 142:179–88. doi: 10.1016/j.vetimm.2011.05.007
62. Zhao M, Zhai Y, Zai X, Mao Y, Hu E, Wei Z, et al. Comparative evaluation of protective immunity against Francisella tularensis induced by subunit or adenovirus-vectored vaccines. Front Cell Infect Microbiol. (2023) 13:1195314. doi: 10.3389/fcimb.2023.1195314
63. Gogev S, Vanderheijden N, Lemaire M, Schynts F, D'Offay J, Deprez I, et al. Induction of protective immunity to bovine herpesvirus type 1 in cattle by intranasal administration of replication-defective human adenovirus type 5 expressing glycoprotein gC or gD. Vaccine. (2002) 20:1451–65. doi: 10.1016/S0264-410X(01)00458-3
64. Reddy PS, Idamakanti N, Pyne C, Zakhartchouk AN, Godson DL, Papp Z, et al. The immunogenicity and efficacy of replication-defective and replication-competent bovine adenovirus-3 expressing bovine herpesvirus-1 glycoprotein gD in cattle. Vet Immunol Immunopathol. (2000) 76:257–68. doi: 10.1016/S0165-2427(00)00217-8
65. Li S, Li X, Yuan R, Chen X, Chen S, Qiu Y, et al. Development of a recombinant adenovirus-vectored vaccine against both infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. (2023) 132:108457. doi: 10.1016/j.fsi.2022.108457
66. Li H, Ning P, Lin Z, Liang W, Kang K, He L, et al. Co-expression of the C-terminal domain of Yersinia enterocolitica invasin enhances the efficacy of classical swine-fever-vectored vaccine based on human adenovirus. J Biosci. (2015) 40:79–90. doi: 10.1007/s12038-014-9495-z
67. Medina GN, Montiel N, Diaz-San Segundo F, Sturza D, Ramirez-Medina E, Grubman MJ, et al. Evaluation of a fiber-modified adenovirus vector vaccine against foot-and-mouth disease in cattle. Clin Vaccine Immunol. (2016) 23:125–36. doi: 10.1128/CVI.00426-15
68. Diaz-San Segundo F, Dias CC, Moraes MP, Weiss M, Perez-Martin E, Salazar AM, et al. Poly ICLC increases the potency of a replication-defective human adenovirus vectored foot-and-mouth disease vaccine. Virology. (2014) 468–70:283–92. doi: 10.1016/j.virol.2014.08.012
69. Monteil M, Le Pottier MF, Ambriović Ristov A, Cariolet R, L'Hospitalier R, Klonjkowski B, et al. Single inoculation of replication-defective adenovirus-vectored vaccines at birth in piglets with maternal antibodies induces high level of antibodies and protection against pseudorabies. Vaccine. (2000) 18:1738–42. doi: 10.1016/S0264-410X(99)00545-9
70. Piret J, Boivin G. Immunomodulatory strategies in herpes simplex virus encephalitis. Clin Microbiol Rev. (2020) 33:e00105–19. doi: 10.1128/CMR.00105-19
71. Kukhanova MK, Korovina AN, Kochetkov SN. Human herpes simplex virus: life cycle and development of inhibitors. Biochemistry. (2014) 79:1635–52. doi: 10.1134/S0006297914130124
72. Cohen JI. Therapeutic vaccines for herpesviruses. J Clin Invest. (2024) 134:e179483. doi: 10.1172/JCI179483
73. Da Costa XJ, Morrison LA, Knipe DM. Comparison of different forms of herpes simplex replication-defective mutant viruses as vaccines in a mouse model of HSV-2 genital infection. Virology (2001) 288:256–63. doi: 10.1006/viro.2001.1094
74. Delagrave S, Hernandez H, Zhou C, Hamberger JF, Mundle ST, Catalan J, et al. Immunogenicity and efficacy of intramuscular replication-defective and subunit vaccines against herpes simplex virus type 2 in the mouse genital model. PLoS ONE. (2012) 7:e46714. doi: 10.1371/journal.pone.0046714
75. Piras F, Plitnick LM, Berglund P, Bernard M, Desert P. Nonclinical safety evaluation of two vaccine candidates for herpes simplex virus type 2 to support combined administration in humans. J Appl Toxicol. (2023) 43:534–56. doi: 10.1002/jat.4404
76. Morrison LA, Knipe DM. Mechanisms of immunization with a replication-defective mutant of herpes simplex virus 1. Virology. (1996) 220:402–13. doi: 10.1006/viro.1996.0328
77. Renukaradhya GJ, Meng X-J, Calvert JG, Roof M, Lager KM. Inactivated and subunit vaccines against porcine reproductive and respiratory syndrome: current status and future direction. Vaccine. (2015) 33:3065–72. doi: 10.1016/j.vaccine.2015.04.102
78. Bai X, Wang Y, Xu X, Sun Z, Xiao Y, Ji G, et al. Commercial vaccines provide limited protection to NADC30-like PRRSV infection. Vaccine. (2016) 34:5540–5. doi: 10.1016/j.vaccine.2016.09.048
79. Welch S-KW, Jolie R, Pearce DS, Koertje WD, Fuog E, Shields SL, et al. Construction and evaluation of genetically engineered replication-defective porcine reproductive and respiratory syndrome virus vaccine candidates. Vet Immunol Immunopathol. (2004) 102:277–90. doi: 10.1016/j.vetimm.2004.09.022
80. Urbano AC, Ferreira F. African swine fever control and prevention: an update on vaccine development. Emerg Microbes Infect. (2022) 11:2021–33. doi: 10.1080/22221751.2022.2108342
81. Yu W, Sun H, Fang C, Song J, Bao E. Establishment of a reverse genetics system for feline calicivirus and construction of an attenuated strain. Anim Husband Vet Med. (2024) 56:107–12.
82. Pollard AJ, Bijker EM. A guide to vaccinology: from basic principles to new developments. Nat Rev Immunol. (2021) 21:83–100. doi: 10.1038/s41577-020-00479-7
83. Lv Z, Li Q, Feng Z, Zheng X, NaYin, Yang H, et al. Inactivated SARS-CoV-2 vaccines elicit immunogenicity and T-cell responses in people living with HIV. Int Immunopharmacol. (2022) 102:108383. doi: 10.1016/j.intimp.2021.108383
84. Niu X, Wang Q. Prevention and control of porcine epidemic diarrhea: the development of recombination-resistant live attenuated vaccines. Viruses. (2022) 14:1317. doi: 10.3390/v14061317
85. Sheerin D, Dold C, O'Connor D, Pollard AJ, Rollier CS. Distinct patterns of whole blood transcriptional responses are induced in mice following immunisation with adenoviral and poxviral vector vaccines encoding the same antigen. BMC Genomics. (2021) 22:777. doi: 10.1186/s12864-021-08061-8
86. Anguiano-Zarate SS, Matchett WE, Nehete PN, Sastry JK, Marzi A, Barry MA. A Replicating single-cycle adenovirus vaccine against Ebola virus. J Infect Dis. (2018) 218:1883–89. doi: 10.1093/infdis/jiy411
87. Kappes MA, Faaberg KS. PRRSV structure, replication and recombination: origin of phenotype and genotype diversity. Virology. (2015) 479:475–86. doi: 10.1016/j.virol.2015.02.012
88. Montaner-Tarbes S, del Portillo HA, Montoya M, Fraile L. Key gaps in the knowledge of the porcine respiratory reproductive syndrome virus (PRRSV). Front Vet Sci. (2019) 6:38. doi: 10.3389/fvets.2019.00038
89. Zhou L, Ge X, Yang H. Porcine reproductive and respiratory syndrome modified live virus vaccine: a “leaky” vaccine with debatable efficacy and safety. Vaccines. (2021) 9:362. doi: 10.3390/vaccines9040362
90. Jiang W, Jiang P, Li Y, Tang J, Wang X, Ma S. Recombinant adenovirus expressing GP5 and M fusion proteins of porcine reproductive and respiratory syndrome virus induce both humoral and cell-mediated immune responses in mice. Vet Immunol Immunopathol. (2006) 113:169–80. doi: 10.1016/j.vetimm.2006.05.001
Keywords: replication-deficient virus, replication-deficient vaccine, single-cycle replication, safety, vector, vaccine
Citation: Wang J, Cui J, Li G and Yu L (2025) Research advances in replication-deficient viral vector vaccines. Front. Vet. Sci. 12:1535328. doi: 10.3389/fvets.2025.1535328
Received: 27 November 2024; Accepted: 11 February 2025;
Published: 03 March 2025.
Edited by:
Dirk Werling, Royal Veterinary College (RVC), United KingdomReviewed by:
Michael Rahe, North Carolina State University, United StatesCopyright © 2025 Wang, Cui, Li and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lingxue Yu, eXVsaW5neHVlQHNodnJpLmFjLmNu; Guoxin Li, Z3VveGlubGlAc2h2cmkuYWMuY24=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.