
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
CASE REPORT article
Front. Vet. Sci., 19 March 2025
Sec. Parasitology
Volume 12 - 2025 | https://doi.org/10.3389/fvets.2025.1534378
 Rubén L. Rodríguez-Expósito1,2,3
Rubén L. Rodríguez-Expósito1,2,3 Loles Carbonell4
Loles Carbonell4 Jesús Recuero-Gil5Javier Martinez4Rosa Martinez-Valverde5
Jesús Recuero-Gil5Javier Martinez4Rosa Martinez-Valverde5 Carmen Martinez-Fernandez6
Carmen Martinez-Fernandez6 Joaquín Ortega-Porcel7Agustín Barragán Hernández7Juan M. Corpa7
Joaquín Ortega-Porcel7Agustín Barragán Hernández7Juan M. Corpa7 Estefanía Montero Cortijo7Jesús Sánchez-Nicolás8Sergio Moya9
Estefanía Montero Cortijo7Jesús Sánchez-Nicolás8Sergio Moya9 Patricia Pérez-Pérez1,2
Patricia Pérez-Pérez1,2 María Reyes-Batlle1,2,3Angélica Domíngez-de-Barros1,3Omar García-Pérez1,3
María Reyes-Batlle1,2,3Angélica Domíngez-de-Barros1,3Omar García-Pérez1,3 Angela Magnet6
Angela Magnet6 Fernando Izquierdo6
Fernando Izquierdo6 Soledad Fenoy6
Soledad Fenoy6 Carmen del Águila6
Carmen del Águila6 Elizabeth Córdoba-Lanús1,3*Francisco de Asis García-González5Miguel Casares4*
Elizabeth Córdoba-Lanús1,3*Francisco de Asis García-González5Miguel Casares4* José E. Piñero1,2,3*
José E. Piñero1,2,3* Jacob Lorenzo-Morales1,2,3*
Jacob Lorenzo-Morales1,2,3*Balamuthia mandrillaris is an amoeba that can cause granulomatous amoebic encephalitis (GAE) as well as lung and skin infections in both humans and animals. Studies on B. mandrillaris-related GAE cases have increased in recent years. This amoeba has been identified as a cause of encephalitis and death in several non-human primates. In this study, we report a case of a 4-year-old female Bornean orangutan (Pongo pygmaeus) in a zoological center that exhibited neurological symptoms for several days. After unsuccessful treatments and a worsening in her condition, euthanasia was deemed necessary. Additionally, we describe the case of a 4-year-old male chimpanzee (Pan troglodytes verus) who died suddenly in a different zoo. Postmortem analysis revealed brain lesions with multiple hemorrhages, oedema, and inflammation in various organs in both cases. Histology showed the presence of B. mandrillaris trophozoites in necrotic and inflamed brain tissues, consistent with granulomatous amoebic meningoencephalitis. The diagnosis was confirmed using a multiplex qPCR assay on brain tissue samples from both animals water and soil samples from the chimpanzee’s and orangutan’s enclosure tested positive for B. mandrillaris DNA by qPCR, confirming environmental exposure. An immunofluorescent antibody (IFA) assay detected B. mandrillaris in chimpanzee brain slices. According to the authors’ knowledge, this report documents the first known cases of Balamuthia amoebic encephalitis in non-human primates in Spain and the first case in Pan troglodytes verus.
Among free-living amoebae (FLA), which are ubiquitous and widely distributed in the environment, only four genera or species are known agents of human and animal infection of the central nervous system (CNS): Acanthamoeba spp., Naegleria fowleri, Balamuthia mandrillaris, and Sappinia pedata (1). Amoebic brain infections are an important emerging parasitic disease, mainly in immunodeficient patients. B. mandrillaris and several Acanthamoeba species can cause granulomatous amoebic encephalitis (GAE), and lung and skin infections (2–5). In contrast, N. fowleri causes acute and fulminant primary amoebic meningoencephalitis (PAM), leading in most cases to the death of the patient within a few days (6).
Hitherto, B. mandrillaris is an amoeba and the only known species of Balamuthia (1). Its life cycle, like Acanthamoeba, presents only two stages: a vegetative trophozoite 12–60 μm in size, and a dormant spherical cyst 12–30 μm (1, 3, 7). These protozoans can induce infection directly through contaminated skin lesions, by inhalation via the olfactory nerve, lung and gastrointestinal tract, or organ transplants. This is followed by CNS invasion via haematogenous spread, leading to severe neurological disease. Most patients present with subacute onset of neurological symptoms and rapid progression to death within weeks to years, showing a fatality rate of over 95.0% in most of cases. The high fatality rate may be explained by the delayed diagnosis and ineffective treatment approaches (3, 4, 8).
The first reported case of Balamuthia mandrillaris encephalitis occurred in 1986, isolated from a 3-year-old pregnant mandrill monkey (Mandrillus sphinx) at the San Diego Wild Animal Park after showing signs of severe paralysis (9). In this regard, the number of studies about B. mandrillaris related to GAE cases has gradually increased in recent years. This amoeba has been identified in several non-human primates, such as in western lowland gorilla (Gorilla gorilla gorilla) (10–12), Northwest Bornean orangutan (Pongo pygmaeus pygmaeus) (13, 14), Kikuyu Colobus monkey (Colobus guereza kikuyuensis), or in white-cheeked gibbon (Nomascus leucogenys) (12). In addition, cases have been reported in other animals on postmortem examination, including a sheep (15), horses (16), an Indian flying fox (17), a tiger (18) and dogs (19–22).
Herein, we report a case of a 4-year-old female Bornean orangutan whose initial sign was hemiparesis. During the evolution of the symptoms the animal was treated with several different drugs without clinical improvement. A computed tomography (CT) scan revealed multiple masses in the brain. Finally, the orangutan’s condition rapidly progressed to worsening neurological symptoms, and euthanasia was conducted.
On the other hand, we present another case of a 4-year-old male chimpanzee (Pan troglodytes verus) without relevant neurological symptoms, who died suddenly in the facilities of the center.
This study summarized the clinical features and detailed the postmortem anatomopathological findings for both cases. The amoebic trophozoites presented in the brain tissue and the multiplex qPCR performed confirmed the final diagnosis of amoebic meningoencephalitis caused by Balamuthia mandrillaris. To the authors’ knowledge, this is the first report of Balamuthia amoebic encephalitis described in non-human primates in Spain and the first time it has been reported in Pan troglodytes verus.
In May 2020, a 4-year-old female Bornean orangutan (Pongo pygmaeus) from Bioparc Fuengirola developed acute neurological symptoms: hemiparesis of the left arm and leg and subtle paresis of the left side of the mouth. Head injury was initially suspected due to the acute presentation of symptoms, and treatment with dexamethasone was initiated orally to avoid separating the infant from her mother, but without clinical improvement. On the fifth day after the onset of symptoms, the patient was anaesthetized for a computed tomography (CT) scan, which revealed multiple masses in the brain, predominantly on the right side of the brain (Figure 1A). Blood test values were normal, except for a marked increase in alkaline phosphatase. In addition, thoracic and abdominal CT scans, X-rays and ultrasound scans showed no abnormalities. A broad-spectrum serological test for pathogens causing multifocal encephalitis was performed at the Biomedical Primate Research Centre (BPRC, NL), without any positive results for the main agents causing neurological diseases.
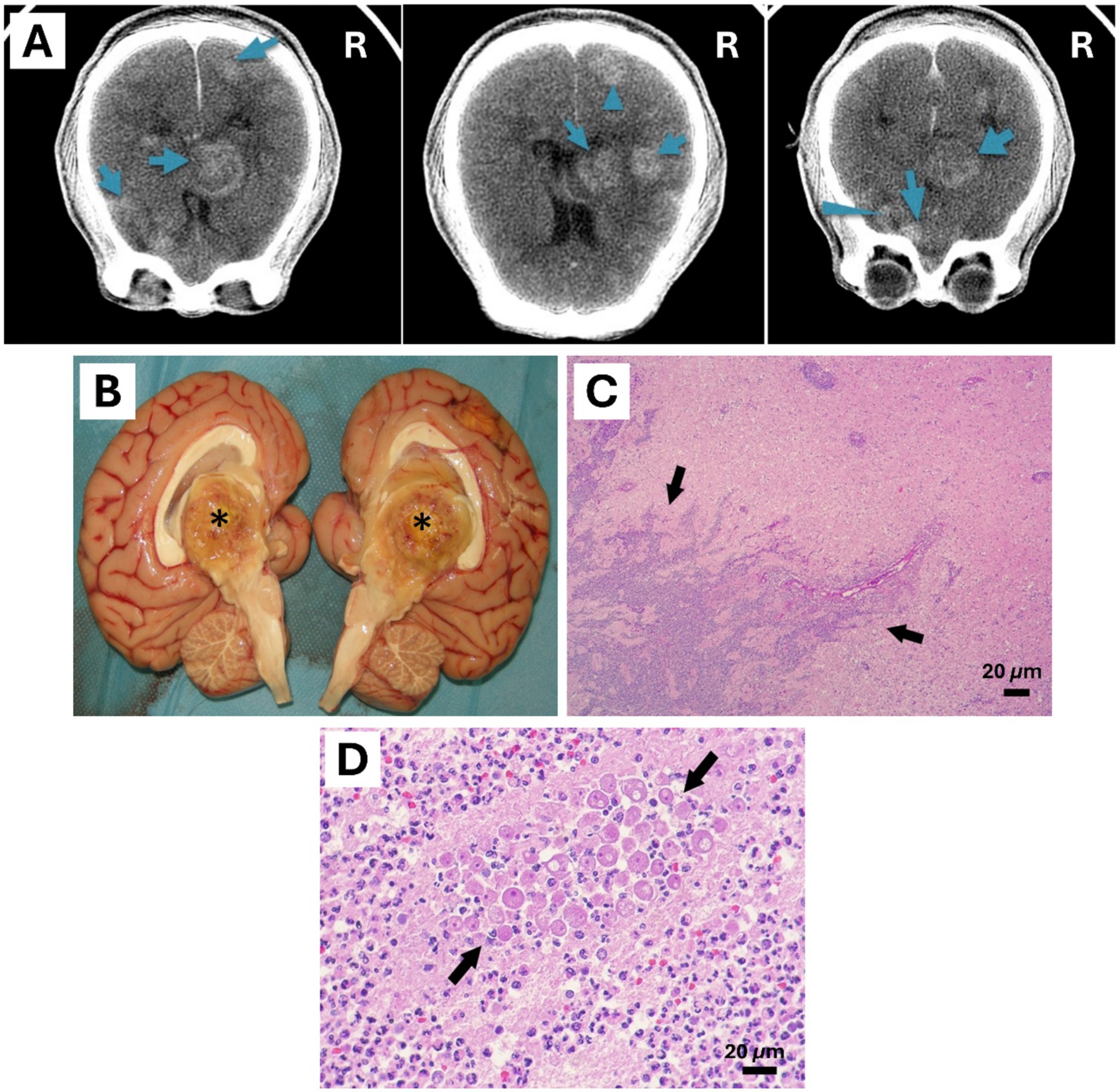
Figure 1. Central nervous system of a 4-year-old female Bornean orangutan (Pongo pygmaeus). (A) Antemortem CT scan of the brain showed multiple masses mainly located on the right side of the encephalon (masses were marked with blue arrows). (B) Mass with central caseous appearance found in the diencephalon and third ventricle, 4 cm in diameter granuloma (*). (C,D) Histological findings of the necropsied encephalon. Rounded amoebic trophozoites with granular cytoplasm and prominent 1 to multiple nuclei were observed inside the area of inflammation in the brain (Black arrows). Hematoxylin & Eosin stain (Scale bar represents 20 μm).
Without a definitive diagnosis, treatment was started for the most common agents that can cause multifocal encephalitis: nematodes, cestodes, protozoa, fungi, viruses and bacteria. Antibiotics (cefixime, ceftazidime, rifampicin), antifungals (itraconazole), anti-toxoplasma (pyrimethamine, clindamycin, folinic acid), antiparasitics (praziquantel and albendazole), and antiprotozoals (metronidazole) were administered. Moreover, she was supplemented with corticosteroids from the first day to control inflammation and potential brain oedema. Levetiracetam and diuretics (furosemide and mannitol) were also administered to control status epilepticus, which appeared within a week of the onset of symptoms. However, neurological signs worsened over the days, with seizures, progressive paresis, strabismus, blindness and a comatose state 16 days after the onset of symptoms. Finally, euthanasia was conducted.
Postmortem study was performed at Bioparc fuengirola. Main lesions were found in the brain. Meninges were slightly congestive, multiple yellowish masses with a caseous center, some visible on the brain’s surface. The largest mass of 4 cm in diameter was located in the right side of the third ventricle and diencephalon (Figure 1B). Other smaller ones: 1.5 cm mass in the right frontal lobe, 1.5 cm mass in the right caudal lobe, 1.5 cm mass in the left anterior frontal lobe, 2 cm mass in the left frontal parietal lobe, and a 1.1 cm mass in the right posterior occipital lobe. The remaining organs showed no macroscopic lesions. Likewise, no skin lesions were found. Formalin fixed tissues were sent to Noah’s Path for histopathology study, frozen tissue samples were preserved to allow for further diagnostics.
The macroscopically observed brain lesions corresponded with large areas of necrosis (malacia), infiltrated by vacuolized macrophages (gitter cells), and other areas of intense neutrophilic infiltration. These foci showed peripheral areas of perivascular and neuropil infiltration by numerous lymphocytes, plasma cells and macrophages, and sparse multinucleated giant cells. At the perivascular level, giant cells often formed multiple concentric layers. The suppurative and lymphoplasmocellular-histiocytic inflammation involved the tunica adventitia and even extended to the media of the blood vessels. In addition, fibrin deposits as well as foci of acute hemorrhage could be distinguished in these areas. Numerous blood vessels, particularly in areas of advancing necrosis, contained fibrin thrombi. On the other hand, the cerebellum was unaffected, except for occasional mild perivascular lymphoplasmacytic infiltrates in the meninges.
Regarding to histological findings, gliosis was seen with numerous prominent gemistocytic astrocytes. Round and oval amoebic trophozoites with one or rarely 2 to multiple small, eccentric basophilic or eosinophilic nuclei and abundant cytoplasm containing one or more clear perinuclear vacuoles were distinguished intralesional. These trophozoites presented a size approximately of a 14 to 30 μm in greatest diameter and were concentrated around the adventitia of blood vessels (Figures 1C,D). In larger areas of lesion, particularly in areas of malacia, trophozoites were scarcely visible, while in other areas, especially foci of suppurative inflammation, they were abundant.
Different other lesions, all considered incidental, were also observed in the rest of the organs. In the colon, foci of acute hemorrhage were found scattered in the lamina propria of the mucosa. Furthermore, in the small intestine, moderate hemosiderin deposits were observed. A diffuse pattern was observed in the liver with mild hepatocellular lipidosis. A diffuse pattern was also observed in the adrenal glands, and the cells in the fasciculate area showed high vacuolation in their cytoplasm. In the thyroid gland, the interstice was infiltrated by numerous hypertrophic adipocytes. In the lymph nodes, diffuse depletion of lymphoid tissue was observed, and a dilatation of the medullary sinuses. Finally, numerous cardiomyocytes with mild to moderate hypertrophy of their nuclei were observed in the heart.
In November 2022 a 4-year-old male chimpanzee (Pan troglodytes verus) was referred already dead to the Clinical Veterinary Hospital of the CEU Cardenal Herrera University by the veterinarians of Bioparc Valencia. On 20 October, the first symptoms appeared, presenting apathy, anorexia and fever. He was administered ibuprofen 7 mg/kg every 12 h for only 3 days, due to the improvement of the symptoms and the fact that his body condition was adequate. On 24 October, he presented a right unilateral strabismus without any other complication. Twelve days after the onset of symptoms, the animal died suddenly in the facilities. No tests or diagnostic procedures were performed during the development of the symptoms, only a postmortem CT scan was performed, but no pathology was observed.
Postmortem examination revealed that the occipital bone on the right side showed several reddish areas, 0.5–2 cm in diameter, compatible with hemorrhages (Figure 2A). The most severe lesions in this chimpanzee were observed in the brain. The meninges were congestive, and the surface of the brain showed diffuse yellowish areas, slightly elevated in the frontal lobe of the left hemisphere (Figure 2B). In addition, a hemorrhage was observed on the inner side of the caudal part of the right mandible during the necropsy of the animal (Figure 2C).
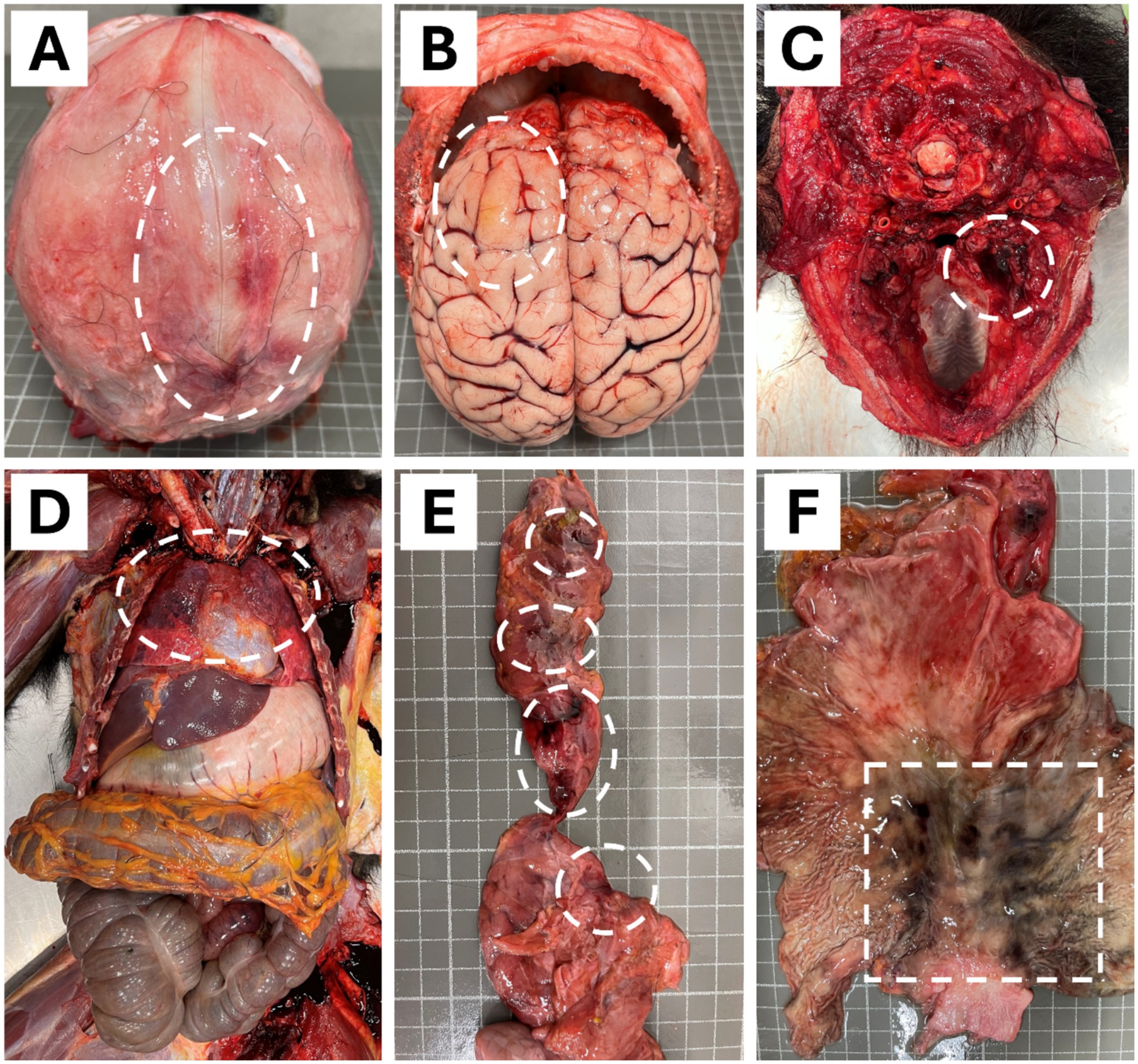
Figure 2. Macroscopic findings in the necropsy of a 4-years-old male chimpanzee (Pan troglodytes verus). (A) Occipital bone presented hemorrhage lesions of 0.5 to 2 cm on the right side. (B) Encephalon showed a delimited congestive and diffuse yellowish areas in the frontal lobe of the left hemisphere. (C) Hemorrhage on the inner side of the caudal part of the right mandible. (D) Obscured, slightly enlarged, diffuse areas with moderate congestion and oedema in the dorsal portion of both lungs. (E) Reddish areas of 0.5 to 1 cm in diameter with multifocal distribution on the pancreas. (F) Blackish areas of 1 to 2 cm in diameter with multifocal distribution and poorly defined contours on the gastric mucosa. Circles and a square in the images depicted all lesions.
In the thoracic cavity, both lungs were partially obscured in their dorsal portion, slightly enlarged, diffusely, with moderate congestion and oedema (Figure 2D). Besides this, the pancreas showed reddish areas, 0.5 to 1 cm in diameter, with multifocal distribution (Figure 2E). The gastric mucosa showed blackish areas, 1–2 cm in diameter, with multifocal distribution and poorly defined contours (Figure 2F). The pathology and findings in both organs are compatible with hemorrhages (ecchymosis). The kidneys showed congestion and mild interstitial nephritis; the liver also showed mild congestion.
Examination of the encephalon revealed multifocal areas of necrosis characterized by yellowish color and granular or cavitated appearance (Figure 3A). Histologically, the brain showed large areas of necrosis with the presence of gitter cells, perivascular cuffs, and inflammation of the meninges. Within the affected areas, parasitic structures were observed with size and morphology consistent with amoebae. Pleomorphic trophozoites with one or multiple nuclei and 12–35 μm in size were observed (Figures 3B–D).
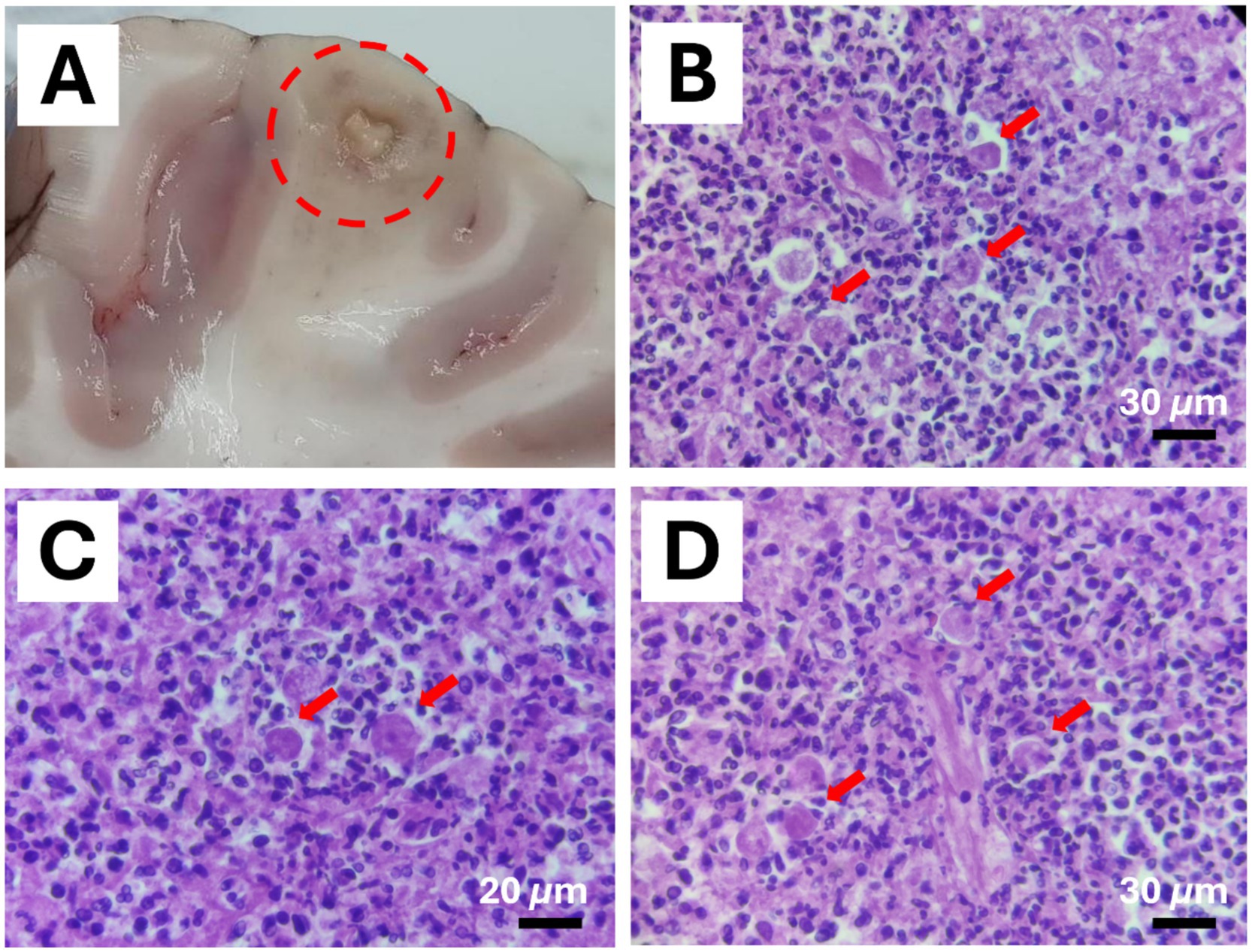
Figure 3. Balamuthia mandrillaris brain infection in a 4-years-old male chimpanzee (Pan troglodytes verus). (A) Granulomatous inflammation in the encephalon (depicted by a circle). (B–D) Intralesional polygonal trophozoites with 1 to multiple nuclei and vacuolated cytoplasm were observed (Red arrows). Hematoxylin & Eosin stain (Scale bar represents 30 μm in B and D; 20 μm in C).
Balamuthia mandrillaris DNA was detected postmortem in brain tissue samples of the orangutan and chimpanzee cases using a multiplex qPCR assay. In addition, the water and soil samples from the facilities where the chimpanzee was housed, and water and soil samples from the orangutan’s facilities were analysed for B. mandrillaris DNA detection. The multiplex qPCR assay was performed for simultaneous molecular detection of Acanthamoeba spp., N. fowleri, and B. mandrillaris specific to the region of the 18 s ribosomal RNA gene using the assay conditions and TaqMan probes reported in our previous study (23), with primer sequences first described by Qvarnstrom et al. (24). Cycle threshold (Ct) values of B. mandrillaris were 30.4 and 28.7 cycles for orangutan and chimpanzee brain tissue, respectively. Soil and water samples from the chimpanzee facilities were positive to: Acanthamoeba spp. (Ct = 31) and B. mandrillaris (Ct = 33.9) for the soil, and Acanthamoeba spp. (Ct = 31.5) and B. mandrillaris (Ct = 33.7) for the water samples. Regarding the soil and water samples from orangutan facilities, multiplex qPCR detected B. mandrillaris (Ct = 32.6) in the water samples, and Acanthamoeba spp. (Ct = 32.5) and B. mandrillaris (Ct = 31.7) in the soil samples.
An immunofluorescent antibody (IFA) assay for detecting B. mandrillaris was performed using 5 μm brain slices from chimpanzee. Slides containing the paraffin-embedded tissue section were deparaffinized and incubated with a rabbit polyclonal antibody against B. mandrillaris. Amoebae were detected with a fluorochrome-conjugated polyclonal goat anti-rabbit antibody (Sigma F9887; Sigma-Aldrich, Darmstadt, Germany) according to a previously described method (25, 26). IFA assay revealed intense positive green staining of amoebic trophozoites in the chimpanzee brain (Figures 4C,D).
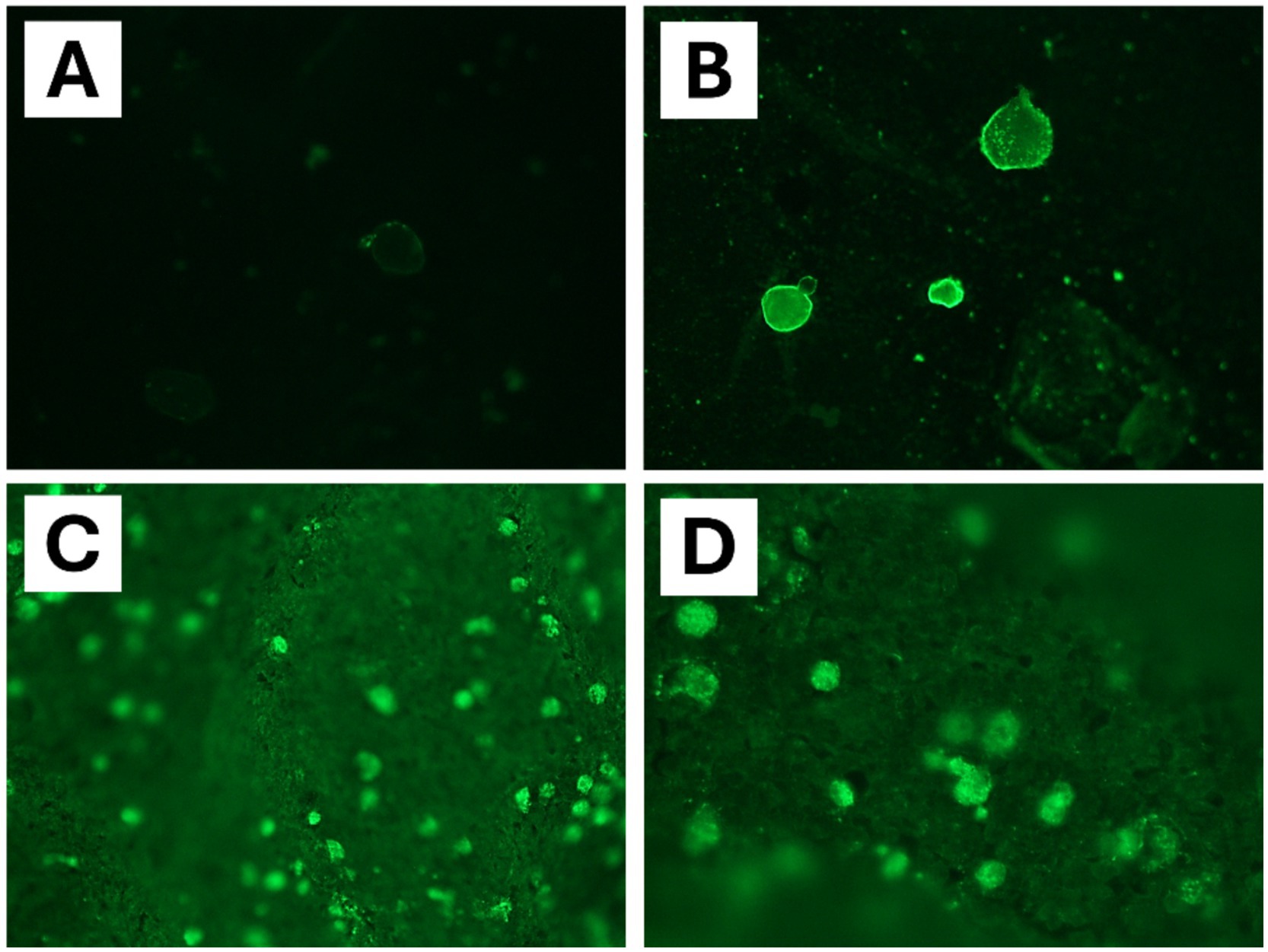
Figure 4. Immunofluorescent antibody (IFA) assay specific for Balamuthia mandrillaris detection in brain tissue samples of chimpanzee (Pan troglodytes verus). (A) Negative control. (B) Positive control with cultured amoebae. (C,D) IFA assay highlighted the presence of B. mandrillaris with bright solid green fluorescence in chimpanzee brain tissues (polyclonal rabbit anti-Balamuthia serum).
Finally, multiplex qPCR assay combined with clinical features and histological findings supports our final diagnosis of Balamuthia amoebic encephalitis in the two cases reported. Furthermore, an IFA assay performed on the chimpanzee brain tissues slides revealed the presence of amoebae, confirming the diagnosis in this case.
Infection produced by Balamuthia mandrillaris has been recognized as an emerging disease. Only about 300 cases have been reported worldwide in humans and animals since 1990, mainly in warmer climate regions (3, 7, 27). Most cases have been reported in the US and Peru (27–30), but several cases have also been reported in the Asia-Pacific region (31–35). In this regard, non-human primate cases have been reported in different species. First description of B. mandrillaris infection was in a mandrill monkey in the US in 1986 (9). Subsequently, in 1997 a retrospective study revealed this infection in two western lowland gorillas, in a white-cheeked gibbon and in a Kikuyu Colobus monkey in the same zoo as the mandrill monkey (12). In the same year, Canfield et al. (14) reported a case in a Bornean orangutan in New South Wales, Australia. However, it was not until 2011 when Mätz-Rensing et al. (11) reported an infection in Germany in a gorilla. Then, Gjeltema et al. (10) described a new case involving a gorilla in North Carolina in the US. In addition, a second case in an orangutan was reported by Hawkins et al. (13) in the US. In this case report, we describe the first documented case of an infection caused by B. mandrillaris in Spain in an orangutan and the first case report describing this infection in a chimpanzee (Pan troglodytes verus).
Infection requires environmental exposure to cysts or trophozoites of B. mandrillaris and can occur in both immunocompromised and immunocompetent hosts. Certain cases report a history of exposure to soil or water contaminated with this amoeba at animal facilities. In this context, cysts are highly resistant to freezing, temperatures up to 70°C, detergents, ultraviolet radiation, chlorine, and disinfectants (3, 5, 36–39). Therefore, contact with this amoeba through contaminated soil or water in treated or disinfected facilities is still possible. In a previous report, water samples collected in the orangutan facility from the B. mandrillaris case infection reported by Hawkins et al. (13) tested real-time PCR positive for this amoeba. In another retrospective study, direct examination of water and soil samples collected in gorilla and mandril enclosures showed several different FLA, but none were identified to species level (12). In the present case, the water and soil samples obtained from the chimpanzee and orangutan facilities revealed the presence of B. mandrillaris by multiplex qPCR, suggesting that this could be the source of infection, based on previously reported cases and the biology of amoebae. In this sense, the route of B. mandrillaris infection is generally considered to be through skin trauma (3, 40). Nevertheless, the route of exposure in our cases was unknown. No external skin lesions were observed in either the chimpanzee or the orangutan during the examinations or at necropsy. Other reported potential routes of infection include inhalation or even gastrointestinal absorption (41), suggesting these as possible routes of infection in our cases, through the ingestion of water or direct contact with the water source and soil in the facilities.
B. mandrillaris infections can have an onset of symptoms that develops over weeks to years as the amoeba replicates in host tissue, eventually causing clinical disease. The infection typically leads to rapid deterioration of the host’s condition, with symptoms such as headache, dysphagia, focal paralysis, seizures, and coma (1–3, 27). In our case, the orangutan primarily exhibited neurological symptoms, including left-sided hemiparesis of the arm and leg, slight facial paresis, seizures, strabismus and confusion that progressed to a comatose state within 2 weeks of symptoms onset. In a previously reported case, an infected orangutan showed symptoms of depression, lethargy, inappetence and head pain (14). More recently, other case reported neurological symptoms, including moderate ataxia, thoracic limb muscle fasciculations, depressed mentation, and reduced fine motor skills, with a progression of years since symptoms onset (13). All these clinical signs align with those observed in our orangutan case, which are typical of B. mandrillaris infection. Nonetheless, the chimpanzee case only presented apathy, fever and mild unilateral strabismus, leading to death 12 days after symptoms onset.
Common pathological findings at necropsy include a necrotizing granulomatous encephalitis with infiltration of inflammatory cells and focal hemorrhages. Additionally, histological preparations typically reveal lesions consistent with suppurative necrotizing meningoencephalitis, along with fibrinoid necrosis of small blood vessels, hemorrhage or oedema (3, 4, 10, 13, 14, 28, 42). Both current case reports align with these usual findings. The orangutan and the chimpanzee presented meningoencephalitis with extensive necrotic areas and suppurative inflammation in the affected regions, macroscopically and histologically. Extracranial lesions in hosts infected with B. mandrillaris are often rare, but disseminated lesions affecting other organs have been described (2, 19, 20, 28, 30, 31, 41). In our cases, necropsy findings in the chimpanzee included moderate congestion, oedema and hemorrhage in the gastric system, pancreas and lungs. The orangutan showed focal acute hemorrhage in the colon, intestines, liver and adrenal glands, and several infiltrative inflammation and lesions in other regions. However, according to the pathologist, these were mild findings and unrelated to the cause of death. In this context, no secondary cause for these findings was identified in either case. Previously reported cases in orangutans have not documented extracranial lesions associated with this amoebic infection (13, 14).
In most cases, the diagnosis of B. mandrillaris infection in non-human primates is usually reported at postmortem. Furthermore, since postmortem analysis of these animals is uncommon in many countries, it suggests that the number of deaths due to B. mandrillaris could be significantly higher than reported. A further complication is that diagnosing this amoebic infection usually requires a brain tissue biopsy for confirmatory analysis. Balamuthia is rarely detectable in the cerebrospinal fluid of infected hosts, either through microscopy or PCR (4, 8, 13, 18, 26, 43). In our report, a postmortem multiplex qPCR assay was performed on brain tissue samples to confirm the final diagnosis of B. mandrillaris infection in both orangutan and chimpanzee cases. Recent advances in molecular biology allow for highly sensitivity and specificity detection of this amoeba in water and soils samples (23, 24). Likewise, we conducted an IFA assay, which revealed stained trophozoites in the chimpanzee brain tissue. These rapid and highly sensitive/specific assays could facilitate earlier antemortem treatment. Concerning this, Ferris et al. (25) studied the presence of B. mandrillaris antibodies in stored serum samples from five orangutans using a human IFA assay, suggesting that this may enable novel antemortem diagnosis in individuals with symptoms consistent with an amoebic encephalitis, which is critical for timely treatment.
Optimal treatment for B. mandrillaris encephalitis remains a challenge. Delay in diagnosis, misdiagnosis, lack of effective drugs, and the severity of infection, all contributing to the low survival rate (4, 8, 27, 39, 43, 44). Current treatment strategies involve combination therapy with miltefosine, fluconazole, miconazole, ketoconazole, 5-fluorocytosine, macrolides or pentamidine. The Centers for Disease Control and Prevention (CDC) recommends a regimen including pentamidine, sulfadiazine, flucytosine, albendazole, fluconazole, azithromycin and miltefosine based on cases of surviving patients (28, 30, 45). However, there is no specific protocol for treating this parasitic disease. In the present case, the chimpanzee died suddenly, and no therapy was administered. The orangutan, in absence of diagnosis, was treated with combination therapy targeting the most common causes of encephalitis in non-human primates, but this was unsuccessful, leading to euthanasia decision. In this regard, fluoroquinolone nitroxoline has recently been used to successfully treat B. mandrillaris encephalitis in a human case (46, 47). Additionally, in previously studies we demonstrate that nitroxoline could inhibit other amoebas that cause encephalitis in humans and animals, Acanthamoeba spp. and Naegleria fowleri specifically, inducing a programmed cell death in these parasites (48, 49). Therefore, nitroxoline may represent a promising new therapeutic option for developing more effective therapies against B. mandrillaris.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because this is a report on zoo animals. Written informed consent was obtained from the participants for the publication of this case report.
RR-E: Conceptualization, Data curation, Writing – original draft, Writing – review & editing. LC: Data curation, Supervision, Writing – review & editing. JR-G: Writing – review & editing, Data curation. JM: Writing – review & editing, Data curation. RM-V: Writing – review & editing, Data curation. CM-F: Conceptualization, Data curation, Writing – review & editing. JO-P: Conceptualization, Data curation, Supervision, Writing – review & editing. AH: Writing – review & editing, Data curation. JC: Conceptualization, Data curation, Supervision, Writing – review & editing. EC: Writing – review & editing, Data curation. JS-N: Writing – review & editing, Data curation. SM: Writing – review & editing, Data curation. PP-P: Writing – review & editing, Data curation. MR-B: Writing – review & editing, Data curation. AD-d-B: Writing – review & editing, Data curation. OG-P: Writing – review & editing, Data curation. AM: Writing – review & editing, Data curation, Conceptualization, Supervision. FI: Data curation, Writing – review & editing. SF: Writing – review & editing, Data curation. CÁ: Writing – review & editing, Data curation, Conceptualization, Supervision. EC-L: Conceptualization, Data curation, Supervision, Writing – review & editing. FG-G: Funding acquisition, Project administration, Resources, Writing – review & editing. MC: Writing – review & editing, Funding acquisition, Project administration, Resources. JP: Writing – review & editing, Funding acquisition, Project administration, Conceptualization, Data curation, Supervision. JL-M: Supervision, Writing – review & editing, Conceptualization, Data curation, Funding acquisition, Project administration.
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the Consorcio Centro de Investigación Biomédica (CIBER) de Enfermedades Infecciosas (CIBERINFEC); Instituto de Salud Carlos III, 28006 Madrid, Spain (CB21/13/00100); and Ministerio de Sanidad, Spain. RR-E and OG-P were funded by the Cabildo Insular de Tenerife 2023–2028 PROYECT CC20230222, CABILDO.23. PP-P was funded by a grant (TESIS2021010070) from the Agencia Canaria de Investigación, Innovación y Sociedad de la Información, 85% cofunded by Fondo Social Europeo (FSE).
The authors acknowledge Dr. Consuelo Rubio Guerri and Dr. María Magdalena Garijo Toledo from the University Cardenal Herrera, CEU Universities (UCH-CEU). Carles Juan-Sallés, Noah’s Path for providing histopathological diagnosis and pictures. Dr. Henk Niphuis, BPRC, Netherlands for primate pathogens full molecular and serological screening.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
GAE, granulomatous amoebic encephalitis; IFA, immunofluorescent antibody assay; FLA, free-living amoebae; CNS, central nervous system; PAM, primary amoebic meningoencephalitis; CT, computed tomography.
1. Visvesvara, GS, Moura, H, and Schuster, FL. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol Med Microbiol. (2007) 50:1–26. doi: 10.1111/j.1574-695X.2007.00232.x
2. Lee, DC, Fiester, SE, Madeline, LA, Fulcher, JW, Ward, ME, Schammel, CM-G, et al. Acanthamoeba spp. and Balamuthia mandrillaris leading to fatal granulomatous amebic encephalitis. Forensic Sci Med Pathol. (2020) 16:171–6. doi: 10.1007/s12024-019-00202-6
3. Bhosale, NK, and Parija, SC. Balamuthia mandrillaris: an opportunistic, free-living ameba–an updated review. Trop Parasitol. (2021) 11:78–88. doi: 10.4103/tp.tp_36_21
4. Berger, JR. Amebic infections of the central nervous system. J Neurovirol. (2022) 28:467–72. doi: 10.1007/s13365-022-01096-x
5. Magnet, A, Fenoy, S, Galván, AL, Izquierdo, F, Rueda, C, Fernandez Vadillo, C, et al. A year long study of the presence of free living amoeba in Spain. Water Res. (2013) 47:6966–72. doi: 10.1016/j.watres.2013.09.065
6. Piñero, JE, Chávez-Munguía, B, Omaña-Molina, M, and Lorenzo-Morales, J. Naegleria fowleri. Trends Parasitol. (2019) 35:848–9. doi: 10.1016/j.pt.2019.06.011
7. Zhang, Z, Liang, J, Wei, R, Feng, X, Wang, L, Wang, L, et al. Facial Balamuthia mandrillaris infection with neurological involvement in an immunocompetent child. Lancet Infect Dis. (2022) 22:e93–e100. doi: 10.1016/S1473-3099(21)00334-0
8. Haston, JC, and Cope, JR. Amebic encephalitis and meningoencephalitis: an update on epidemiology, diagnostic methods, and treatment. Curr Opin Infect Dis. (2023) 36:186–91. doi: 10.1097/QCO.0000000000000923
9. Visvesvara, GS, Martinez, AJ, Schuster, FL, Leitch, GJ, Wallace, SV, Sawyer, TK, et al. Leptomyxid ameba, a new agent of amebic meningoencephalitis in humans and animals. J Clin Microbiol. (1990) 28:2750–6. doi: 10.1128/jcm.28.12.2750-2756.1990
10. Gjeltema, JL, Troan, B, Muehlenbachs, A, Liu, L, Da Silva, AJ, Qvarnstrom, Y, et al. Amoebic meningoencephalitis and disseminated infection caused by Balamuthia mandrillaris in a Western lowland gorilla (Gorilla gorilla gorilla). J Am Vet Med Assoc. (2016) 248:315–21. doi: 10.2460/javma.248.3.315
11. Mätz-Rensing, K, Kunze, M, Zöller, M, Roos, C, Kiderlen, AF, Ludwig, C, et al. Fatal Balamuthia mandrillaris infection in a gorilla–first case of balamuthiasis in Germany. J Med Primatol. (2011) 40:437–40. doi: 10.1111/j.1600-0684.2011.00479.x
12. Rideout, BA, Gardiner, CH, Stalis, IH, Zuba, JR, Hadfield, T, and Visvesvara, GS. Fatal infections with Balamuthia mandrillaris (a free-living amoeba) in gorillas and other old world primates. Vet Pathol. (1997) 34:15–22. doi: 10.1177/030098589703400103
13. Hawkins, SJ, Struthers, JD, Phair, K, Ali, IKM, Roy, S, Mull, B, et al. Diagnostic evaluation of fatal Balamuthia mandrillaris meningoencephalitis in a captive Bornean orangutan (Pongo pygmaeus) with identification of potential environmental source and evidence of chronic exposure. Primates. (2021) 62:51–61. doi: 10.1007/s10329-020-00860-z
14. Canfield, PJ, Vogelnest, L, Cunningham, ML, and Visvesvara, GS. Amoebic meningoencephalitis caused by Balamuthia mandrillaris in an orangutan. Aust Vet J. (1997) 75:97–100. doi: 10.1111/j.1751-0813.1997.tb14165.x
15. Fuentealba, IC, Wikse, SE, Read, WK, Edwards, JF, and Visvesvara, GS. Amebic meningoencephalitis in a sheep. J Am Vet Med Assoc. (1992) 200:363–5. doi: 10.2460/javma.1992.200.03.363
16. Kinde, H, Read, DH, Daft, BM, Manzer, M, Nordhausen, RW, Kelly, DJ, et al. Infections caused by pathogenic free-living amebas (Balamuthia Mandrillaris and Acanthamoeba sp.) in horses. J Vet Diagn Invest. (2007) 19:317–22. doi: 10.1177/104063870701900318
17. Crossland, NA, Ali, I, Higbie, C, Jackson, J, Pirie, G, and Bauer, R. Neurologic amebiasis caused by Balamuthia mandrillaris in an Indian flying fox (Pteropus giganteus). J Vet Diagn Invest. (2016) 28:54–8. doi: 10.1177/1040638715614346
18. Niedringhaus, KD, Gordon, M, Yabsley, MJ, Gai, J, Uzal, FA, and Woolard, KD. Fatal balamuthosis in a Siberian tiger and a literature review of detection options for free-living amoebic infections in animals. J Vet Diagn Invest. (2023) 35:311–6. doi: 10.1177/10406387231160771
19. Foreman, O, Sykes, J, Ball, L, Yang, N, and De Cock, H. Disseminated infection with Balamuthia mandrillaris in a dog. Vet Pathol. (2004) 41:506–10. doi: 10.1354/vp.41-5-506
20. Finnin, PJ, Visvesvara, GS, Campbell, BE, Fry, DR, and Gasser, RB. Multifocal Balamuthia mandrillaris infection in a dog in Australia. Parasitol Res. (2007) 100:423–6. doi: 10.1007/s00436-006-0302-0
21. Chien, RC-C, Telford, CR, Roy, S, Ali, IKM, Shieh, W-J, and Confer, AW. Canine amoebic meningoencephalitis due to Balamuthia mandrillaris. Vet Parasitol Reg Stud Rep. (2018) 13:156–9. doi: 10.1016/j.vprsr.2018.06.003
22. Hodge, PJ, Kelers, K, Gasser, RB, Visvesvara, GS, Martig, S, and Long, SN. Another case of canine amoebic meningoencephalitis—the challenges of reaching a rapid diagnosis. Parasitol Res. (2011) 108:1069–73. doi: 10.1007/s00436-010-2197-z
23. Córdoba-Lanús, E, Reyes-Batlle, M, Domínguez-de-Barros, A, Pérez-Pérez, P, Rodríguez-Expósito, RL, García-Ramos, A, et al. Multiplex real-time polymerase chain reaction assay to detect Acanthamoeba spp., Vermamoeba vermiformis, Naegleria fowleri, and Balamuthia mandrillaris in different water sources. Am J Trop Med Hyg. (2024) 111:785–90. doi: 10.4269/ajtmh.24-0028
24. Qvarnstrom, Y, Visvesvara, GS, Sriram, R, and da Silva, AJ. Multiplex real-time PCR assay for simultaneous detection of Acanthamoeba spp., Balamuthia mandrillaris, and Naegleria fowleri. J Clin Microbiol. (2006) 44:3589–95. doi: 10.1128/JCM.00875-06
25. Ferris, RL, Ali, IK, and West, G. Use of a human indirect immunofluorescence antibody assay for Balamuthia mandrillaris in a group of captive Northwest Bornean orangutans (Pongo pygmaeus pygmaeus). J Zoo Wildl Med. (2021) 52:310–4. doi: 10.1638/2019-0018
26. Schuster, FL, Honarmand, S, Visvesvara, GS, and Glaser, CA. Detection of antibodies against free-living amoebae Balamuthia mandrillaris and Acanthamoeba species in a population of patients with encephalitis. Clin Infect Dis. (2006) 42:1260–5. doi: 10.1086/503037
27. Bravo, FG, and Seas, C. Balamuthia Mandrillaris amoebic encephalitis: an emerging parasitic infection. Curr Infect Dis Rep. (2012) 14:391–6. doi: 10.1007/s11908-012-0266-4
28. Cope, JR, Landa, J, Nethercut, H, Collier, SA, Glaser, C, Moser, M, et al. The epidemiology and clinical features of Balamuthia mandrillaris disease in the United States, 1974–2016. Clin Infect Dis. (2019) 68:1815–22. doi: 10.1093/cid/ciy813
29. Cabello-Vílchez, AM, Rodríguez-Zaragoza, S, Piñero, J, Valladares, B, and Lorenzo-Morales, J. Balamuthia mandrillaris in South America: an emerging potential hidden pathogen in Perú. Exp Parasitol. (2014) 145:S10–9. doi: 10.1016/j.exppara.2014.05.007
30. Centers for Disease Control and Prevention (CDC). Balamuthia amebic encephalitis--California, 1999-2007. MMWR Morb Mortal Wkly Rep. (2008) 57:768–71.
31. Wang, L, Cheng, W, Li, B, Jian, Z, Qi, X, Sun, D, et al. Balamuthia mandrillaris infection in China: a retrospective report of 28 cases. Emerg Microbes Infect. (2020) 9:2348–57. doi: 10.1080/22221751.2020.1835447
32. Xu, H, Wang, D, Cui, K, Wan, R, Chi, Q, and Wu, T. 18F-FDG PET/CT findings in fatal Balamuthia mandrillaris encephalitis in brain stem: a case report. Radiol Case Rep. (2024) 19:1851–4. doi: 10.1016/j.radcr.2024.02.021
33. Yamanouchi, K, Arima, H, Sakamoto, Y, Kanto, K, Kasai, K, Ito, K, et al. First report of the isolation of Balamuthia mandrillaris in the northern region of Japan. Parasitol Res. (2018) 117:2895–900. doi: 10.1007/s00436-018-5980-x
34. Lee, JY, Yu, IK, Kim, SM, Kim, JH, and Kim, HY. Fulminant disseminating fatal granulomatous amebic encephalitis: the first case report in an immunocompetent patient in South Korea. Yonsei Med J. (2021) 62:563–7. doi: 10.3349/ymj.2021.62.6.563
35. Intalapaporn, P, Suankratay, C, Shuangshoti, S, Phantumchinda, K, Keelawat, S, and Wilde, H. Balamuthia mandrillaris meningoencephalitis: the first case in Southeast Asia. Am J Trop Med Hyg. (2004) 70:666–9. doi: 10.4269/ajtmh.2004.70.666
36. Cabello-Vílchez, AM, Reyes-Batlle, M, Montalbán-Sandoval, E, Martín-Navarro, CM, López-Arencibia, A, Elias-Letts, R, et al. The isolation of Balamuthia mandrillaris from environmental sources from Peru. Parasitol Res. (2014) 113:2509–13. doi: 10.1007/s00436-014-3900-2
37. Yamanouchi, K, Arima, H, Sakamoto, Y, Kanto, K, Itoh, K, and Tsujiguchi, T. Isolation and habitat analysis of Balamuthia mandrillaris from soil. Parasitol Res. (2024) 123:163. doi: 10.1007/s00436-024-08182-5
38. Schuster, FL, Dunnebacke, TH, Booton, GC, Yagi, S, Kohlmeier, CK, Glaser, C, et al. Environmental isolation of Balamuthia mandrillaris associated with a case of amebic encephalitis. J Clin Microbiol. (2003) 41:3175–80. doi: 10.1128/JCM.41.7.3175-3180.2003
39. Siddiqui, R, Ortega-Rivas, A, and Khan, NA. Balamuthia mandrillaris resistance to hostile conditions. J Med Microbiol. (2008) 57:428–31. doi: 10.1099/jmm.0.47694-0
40. Alvarez, P, Torres-Cabala, C, Gotuzzo, E, and Bravo, F. Cutaneous balamuthiasis: a clinicopathological study. JAAD Int. (2022) 6:51–8. doi: 10.1016/j.jdin.2021.11.005
41. Kiderlen, AF, Laube, U, Radam, E, and Tata, PS. Oral infection of immunocompetent and immunodeficient mice with Balamuthia mandrillaris amebae. Parasitol Res. (2007) 100:775–82. doi: 10.1007/s00436-006-0334-5
42. Hara, T, Yagita, K, and Sugita, Y. Pathogenic free-living amoebic encephalitis in Japan. Neuropathology. (2019) 39:251–8. doi: 10.1111/neup.12582
43. Parija, SC, Dinoop, K, and Venugopal, H. Management of granulomatous amebic encephalitis: laboratory diagnosis and treatment. Trop Parasitol. (2015) 5:23–8. doi: 10.4103/2229-5070.149889
44. Spottiswoode, N, Haston, JC, Hanners, NW, Gruenberg, K, Kim, A, DeRisi, JL, et al. Challenges and advances in the medical treatment of granulomatous amebic encephalitis. Ther Adv Infect Dis. (2024) 11:20499361241228340. doi: 10.1177/20499361241228340
45. Centers for Disease Control and Prevention (CDC) (2024) Clinical care of balamuthia infection. In: Treatment recommendations. Available online at: https://www.cdc.gov/balamuthia/hcp/clinical-care/index.html (accessed November 13, 2024).
46. Spottiswoode, N, Pet, D, Kim, A, Gruenberg, K, Shah, M, Ramachandran, A, et al. Successful treatment of Balamuthia mandrillaris granulomatous amebic encephalitis with nitroxoline. Emerg Infect Dis. (2023) 29:197–201. doi: 10.3201/eid2901.221531
47. Laurie, MT, White, CV, Retallack, H, Wu, W, Moser, MS, Sakanari, JA, et al. Functional assessment of 2,177 U.S. and international drugs identifies the quinoline nitroxoline as a potent amoebicidal agent against the pathogen Balamuthia mandrillaris. mBio. (2018) 9:e02051-18. doi: 10.1128/mBio.02051-18
48. Rodríguez-Expósito, RL, Sifaoui, I, Reyes-Batlle, M, Fuchs, F, Scheid, PL, Piñero, JE, et al. Induction of programmed cell death in Acanthamoeba culbertsoni by the repurposed compound nitroxoline. Antioxidants. (2023) 12:2081. doi: 10.3390/antiox12122081
Keywords: Balamuthia mandrillaris, meningoencephalitis, non-human primates, Pongo pygmaeus, Pan troglodytes verus, multiplex qPCR, IFA assay, Spain
Citation: Rodríguez-Expósito RL, Carbonell L, Recuero-Gil J, Martinez J, Martinez-Valverde R, Martinez-Fernandez C, Ortega-Porcel J, Hernández AB, Corpa JM, Cortijo EM, Sánchez-Nicolás J, Moya S, Pérez-Pérez P, Reyes-Batlle M, Domíngez-de-Barros A, García-Pérez O, Magnet A, Izquierdo F, Fenoy S, del Águila C, Córdoba-Lanús E, García-González FdA, Casares M, Piñero JE and Lorenzo-Morales J (2025) Fatal amoebic meningoencephalitis caused by Balamuthia mandrillaris in Pongo pygmaeus and first case report in Pan troglodytes verus. Front. Vet. Sci. 12:1534378. doi: 10.3389/fvets.2025.1534378
Edited by:
Binod Rayamajhee, University of New South Wales, AustraliaReviewed by:
Zorica D. Dakić, University of Belgrade, SerbiaCopyright © 2025 Rodríguez-Expósito, Carbonell, Recuero-Gil, Martinez, Martinez-Valverde, Martinez-Fernandez, Ortega-Porcel, Hernández, Corpa, Cortijo, Sánchez-Nicolás, Moya, Pérez-Pérez, Reyes-Batlle, Domíngez-de-Barros, García-Pérez, Magnet, Izquierdo, Fenoy, del Águila, Córdoba-Lanús, García-González, Casares, Piñero and Lorenzo-Morales. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elizabeth Córdoba-Lanús, YWNvcmRvYmFAdWxsLmVkdS5lcw==; Miguel Casares, bWlndWVsLmNhc2FyZXNAYmlvcGFyY3ZhbGVuY2lhLmVz; José E. Piñero, anBpbmVyb0B1bGwuZWR1LmVz; Jacob Lorenzo-Morales, am1sb3JlbnpAdWxsLmVkdS5lcw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.