 Raúl David Guevara
Raúl David Guevara Sergi López-Vergé
Sergi López-Vergé Jose J. Pastor
Jose J. Pastor Xavier Manteca
Xavier Manteca Gemma Tedo
Gemma Tedo Pol Llonch
Pol Llonch- 1AWEC Advisors S.L., Animal Welfare Education Centre (AWEC), Cerdanyola del Vallès, Spain
- 2Department of Animal and Food Science, Universitat Autònoma de Barcelona, Barcelona, Spain
- 3Animal Science Innovation Division, Lucta, Cerdanyola del Vallès, Spain
Regrouping practices are frequent in pig production, altering hierarchy and triggering aggressive behaviors. The present study aimed to investigate the physiological responses of piglets to an experimental model designed to induce stress through systematic social mixing in two trials. In Trial A, a total of 144 crossbred piglets (25 days postweaning) housed in one room within 36 pens (four piglets/pen) were used and randomly assigned to either a control group (piglets maintained in their pen, Ctrl-A) or a social challenge group (piglets mixed, SC-A). In Trial B, the same number of animals (33 days postweaning) and crossbreed line was used, and each piglet was assigned either to a control group (Ctrl-B) or a social challenge group (SC-B) in two independent rooms (rooms Ctrl and SC, 12 pens/ room, six piglets/pen). The social challenge consisted of daily moves of three out of four pen mates and five out of six pen mates, for Trials A and B, respectively. In the Ctrl groups, all piglets stayed in their original pen. Before the 1st mixing day and at the end of the 3rd mixing day, saliva (cortisol concentration) and blood (cortisol concentration changes, hemogram, and immunologic activation) samples were collected from two random piglets per pen. Skin lesion scores of all piglets were also recorded on the front, middle, and rear body regions. In Trial A, the total skin lesions score was higher in the SC-A group compared to the Ctrl-A group after the social challenge (0.53 vs. 0.17; p < 0.05), but an unexpected increase between sampling days in the Ctrl-A piglets (0.06 vs. 0.17; p < 0.05) was also recorded, suggesting that Ctrl-A pigs showed similar aggressivity levels to the SC-A group. Hematological parameters hemoglobin, red blood cell counts, and leukocyte counts present similar changes in both treatment groups after the social challenge. Contrarily, in Trial B, the lesion score only increased in the piglets in room SC (0.08 vs. 0.34; p < 0.05). Results suggest that stable groups may show aggressive behaviors if they are in the same room with socially challenged pigs. Thus, the physical separation of treatment groups in social stress studies is recommended.
1 Introduction
Social stress in pigs is often caused by common management practices (cross-fostering during lactation, litters’ mixing at weaning, pens’ mixtures at nursery and fattening, and transportation, among other events) that involve regrouping with unfamiliar individuals (1–3). During these mixing events, the established social order among pigs may be disrupted, leading to aggressive behaviors like fights, bites, intimidation, and competition for resources (4, 5). Pigs exhibit various behavioral responses when dealing with violent interactions during social mixing. These actions can be offensive, such as head-to-head clashes, head knocks, biting, chasing, and intimidating through grunts and threatening postures (6–8). Alternatively, responsive actions can be defensive, involving freezing in place or attempting to escape from confrontations (4). However, constant high levels of aggression can compromise the welfare of pigs due to social stress. Social stress can increase cortisol levels (8–10), acute phase protein concentrations (11, 12), immune activation (13, 14), and other hematological parameters (9, 13). On top of that, social stress can negatively affect animal performance and the physiological condition of pigs (13, 15).
Pig production stakeholders have been interested in the mitigation of agonistic behaviors (e.g., tail biting, fighting, intimidating, among others) as it affects the profitability of the production system (16) and raises social concerns about the production practices (17). Plenty of literature has already reported the negative impacts of agonistic behaviors on pigs but the methods to predict and control aggressive outbreaks remain unclear (5, 18, 19). Thus, pig production stakeholders are interested in understanding how negative behaviors disseminate among a group of animals, to develop and apply strategies (e.g., nutritional, husbandry, or management) to reduce the incidence of such negative behaviors.
The present study aimed to investigate the impact of social stress during mixing events and the occurrence of non-socially mixed piglets. A better understanding of negative behaviors dissemination might help to develop approaches to mitigate social stress in pigs.
2 Materials and methods
2.1 Animals, housing, and diets
All procedures were approved by the Laboratory Animal Care Advisory Committee of the Faculty of Veterinary Sciences of the Autonomous University of Barcelona (CEEAH-5754-5755-CEEA-UAB). Two independent trials were performed: (i) Trial A took place at the Institute of Agrifood Research and Technology (IRTA) experimental farm, Mas Bové (Constantí, Tarragona, Spain); and (ii) Trial B was carried out at the LUCTA® Swine Experimental Unit transition farm, “El Castell” (Sant Aniol de Finestres, Girona, Spain). In both trials, piglets from 20 sows were selected (piglets over 4 kg body weight) to be moved to a nursery site at weaning. In trial A, this occurred approximately on day 26, and in trial B, it happened approximately on day 29. At the nursery site, the piglets were distributed according to sex and body weight, and all animals were identified with ear tags.
For Trial A, a total of 144 25-days postweaning (p.w.) crossbred [(Largewhite × Landrace) × Pietrain] piglets (BW 11.90 kg ± 0.79 kg) were housed in one room and distributed into 36 pens according to sex (2 female and 2 male/pen). Each individual pen (4.42 m2; 2 × 2.21 m) was equipped with a hopper feeder featuring four eating spaces, and a drinker.
The same number of animals (BW 14.42 kg ± 0.37 kg, 33 days p.w.) and crossbreed line were used in Trial B. However, pigs were housed in two separate rooms, each containing 12 pens (2.7 m2; 2.2 × 1.3 m), allocating six piglets per pen, maintaining the same proportion of males and females in each pen (3 female and 3 male/pen). Each pen had two drinkers covered with a mobile metal lid to prevent external contamination (feces and urine) and minimize water waste. Pens had one feeder per pen, with 4 separations per feeder, and were also be covered with a metal lid.
In both trials, all pens were equipped with completely slatted plastic floors; each pen had plastic chewable toys (plastic balls with chains attached to the wall); and feed and water were offered ad libitum throughout the experimental period (see Table 1). All diets were formulated to meet or exceed (20) nutrient requirements.
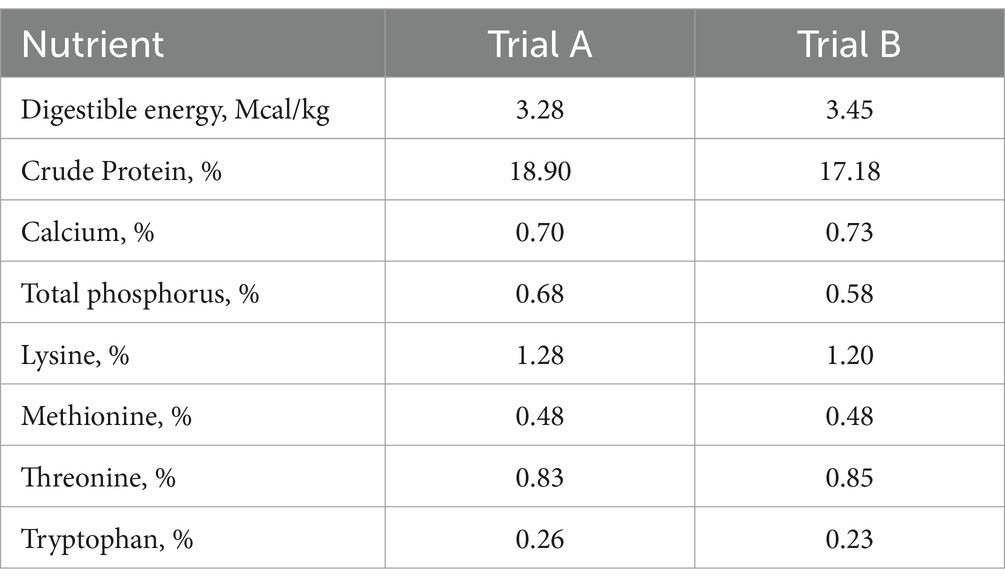
Table 1. Nutrient composition of the diets supplied during the starter phases of the trials (from day 21 to day 41 p.w.).
2.2 Study design–social challenge
In both trials piglets were randomized to one of the two treatment groups: (i) control (Ctrl); and (ii) socially challenged (SC). The social challenge consisted of the daily relocation of three out of four pen mates and five out of six pen mates for Trials A and B, respectively. Relocated piglets were moved to pens with other piglets of similar body weight to maintain groups with similar average body weights. Meanwhile, in the Ctrl pens, all piglets remained in their original pens. The process of social mixing was conducted for three consecutive days, starting at 8:00 a.m. During the 3 days of social challenge, systematic regrouping events were done at 8:00 am before food was placed on the feeders. On the fourth day of the social challenge period, all animals returned to their original pens at 8:00 a.m. for sampling (saliva and blood) and skin lesion score measurements.
In Trail A, both Ctrl and SC pens were housed in the same experimental room, whereas in Trial B, Ctrl, and SC pens were housed in two separate rooms (i.e., one room of Ctrl pens and one room of SC pens).
2.3 Sampling and measurements
2.3.1 Lesion score
Lesion score recording was meant to measure piglets’ aggressiveness during the social mixing events.
On trial A, lesion scores were recorded twice, on the first day and on the last day of the social challenge (days 25, 28 p.w.). Whereas in trial B, the lesion score was recorded thrice, each morning of the social challenge period before pen mixtures (days 33, 34, and 35 p.w.). More skin lesion observations were done in trial B to refine aggressions detection during the challenge period, which was an improvement of the observation protocol from trial A. The lesion score assessment protocol was based on Turner et al. (21), counting the number of lesions on each of the three areas of the body: front, middle and rear, to assign a score from 0 (<6 lesions), 1 (6–15 lesions), or 2 (>15 lesions) (Figure 1). The lesion score aimed to measure the aggressivity of the pigs during the social challenge period.
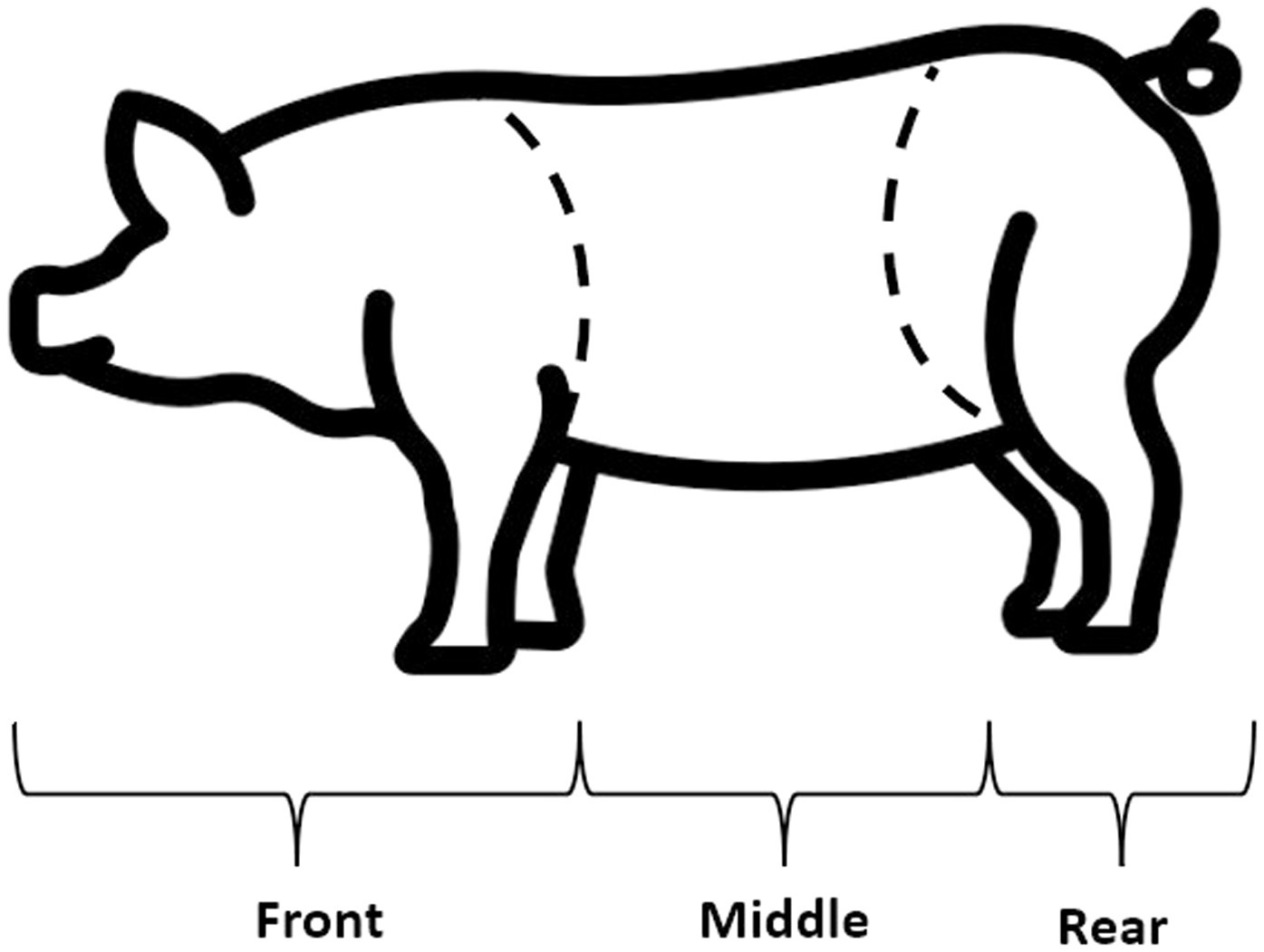
Figure 1. Pig body regions assessed through the skin lesion score (21).
2.3.2 Blood, and saliva samples collection
Blood samples were collected to detect hematological changes triggered by the social mixing. Also, plasmatic and salivary cortisol concentration was measured to monitor piglets stress response.
Piglets were fasted overnight before the sampling to reduce variability in the markers (hematological parameters). Blood samples were collected from the vena cava cranialis using a vacutainer into pre-labeled tubes with EDTA anticoagulant. The blood tubes were immediately centrifuged (1200 g × 10 min). After centrifugation, supernatant was collected, frozen immediately in dry ice and stored (−80°C) until analysis (22).
Saliva samples were collected using saliva collection tubes containing a sponge (Sarstedt, Aktiengesellschaft & Co., Nümbrecht, Germany) each sampling day at the same time (±30 min) to avoid variations due to physiological circadian rhythm. A sponge clipped to a hemostatic clamp was playfully presented to the pigs to encourage them to chew it. Pigs were allowed to chew for approximately 1 min, and subsequently, the samples underwent a centrifugation cycle at 3000 g for 10 min at room temperature. Immediately after centrifugation, the samples were rapidly frozen using dry ice and stored at −80°C until analysis (23).
2.4 Sample analyses
Samples were analyzed for stress activation: cortisol concentration, hematology, hemoglobin, red blood cell counts (RBC), medium corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), and mean corpuscular hemoglobin concentration (MCHC). Cortisol concentration measurements were done aiming to detect a stress response in the animals after the social challenge period. Platelet counts, leukocyte counts, eosinophil counts, basophil counts, lymphocyte counts, monocyte counts, and segmented neutrophil counts were used to evaluate the activation of the immune system before and after the social challenge.
Blood samples were analyzed in a commercial lab (Echevarne, Barcelona, Spain).
Saliva samples were analyzed for cortisol concentration through an automated chemiluminescence immunoassay (Immulite 1000, Siemens Medical Solutions Diagnostics), previously validated for its use in pigs by Escribano et al. (24).
2.5 Statistical analyses
All statistical analyses were performed with the statistical package SAS (version 9.4, SAS Institute Inc., Cary, NC, USA). PROC GLIMMIX was used to assess the lesion score changes between pre pre-social challenge record and post-challenge record in both trials. PROC MIXED with piglet as a random effect and sampling day and treatment as a fixed effect, and adjustment by Tuckey was used to analyze the hematology parameters in the trials. Salivary stress markers activity was performed by PROC GLIMMIX without any adjustment. The main effects and the interaction between them were compared by LSMEANS. Significant differences were declared at p ≤ 0.05 whereas near-significant trends were considered at 0.05 < p ≤ 0.10. Results are presented as mean values with their standard error from the mean (Mean ± SEM).
3 Results
3.1 Lesion score
Figure 2 illustrates the lesion scores observed in both trials. No differences were found between Ctrl-A and SC-A (day 25; p = 0.4645) or between Ctrl-B and SC-B (day 33; p = 0.6257) on their initial lesion scores. After the social mixing, SC-A had a significant increase in the lesion score, relative to the pre-social mixing measurement (p < 0.0001). Ctrl-A piglets also displayed an increase in the lesion score, despite not being directly exposed to social mixing (p = 0.0127), although it was reduced in lower magnitude compared with the SC-A. Regarding the distribution of body lesions, both Ctrl-A (p = 0.0101) and SC-A (p < 0.0001) treatments show a higher concentration of lesions in the front and middle areas relative to the rear body area.
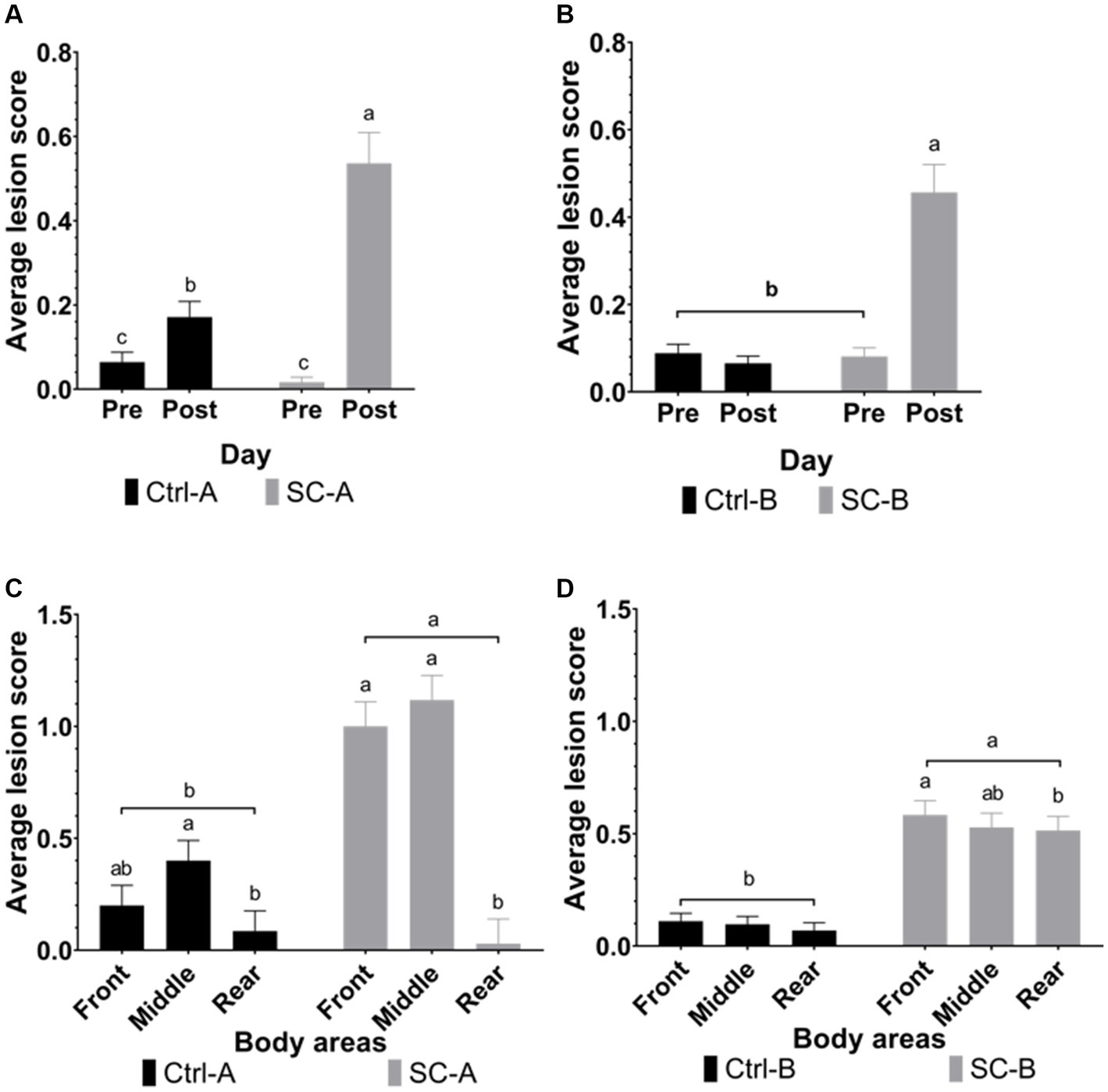
Figure 2. Lesion scores were grouped by treatment groups on days pre- and post-social mixing. (A) Trial A: all animals in the same experimental room at 25 days after weaning (pre-social challenge) and at 28 days after weaning (post-social challenge). (B) Trial B: animals with each treatment group allocated in separate rooms at 33 days after weaning (pre-social challenge) and 36 days after weaning (post-social challenge). (C) Body lesions distribution in trial A at 28 days after weaning (post-social challenge). (D) Body lesions distribution in trial B at 33 days after weaning (post-social challenge). Different lowercase letters indicate significant differences (p < 0.05) whereas near-significant trends were considered at 0.05 < p ≤ 0.10 and represented with capital letters.
SC-B pigs presented a higher lesion score after the social mixing than the measurement previously to the mixing (p < 0.0001), but Ctrl-B pigs did not increase their lesion score after the social mixing period (p = 0.3379). Concerning the distribution of the lesions in the body areas, SC-B piglets presented a higher concentration of body lesions in the front (p = 0.0158) relative to the rear body area but not different from the middle body area (p = 0.0677). SC-B middle area lesion score was similar to the rear body area (p = 0.8412). On the other hand, Ctrl-B pigs maintained their body lesion score in the different body areas after the social mixing period (p = 0.0140).
3.2 Blood markers
3.2.1 Hematological parameters
Hematological parameters summary for trials A and B are summarized in Tables 2, 3, respectively.
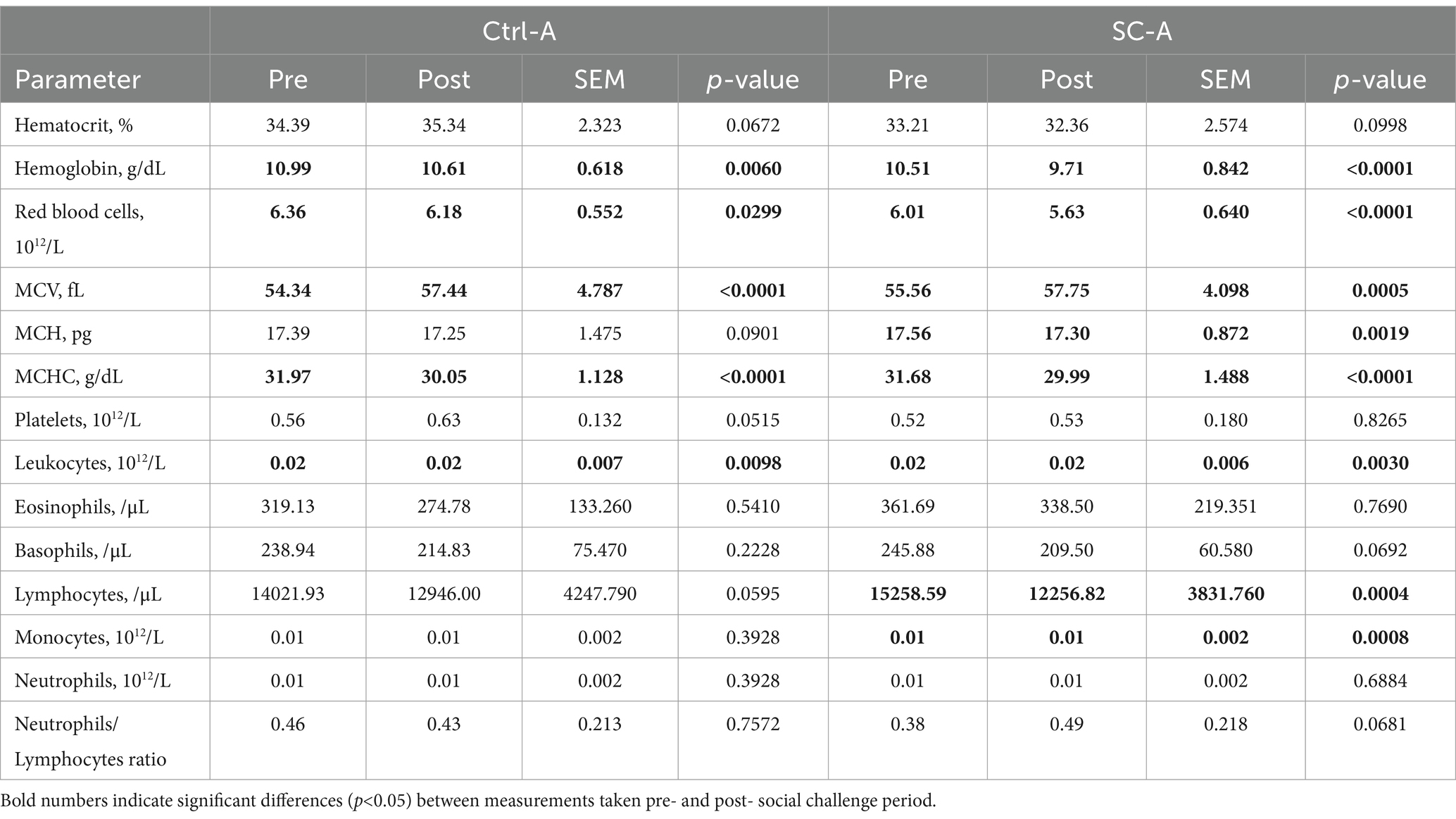
Table 2. Hematological parameters from treatment groups Ctrl-A and SC-A before (Pre) and after (Post) social challenge period (from day 25 post-weaning until day 28) on trial A.
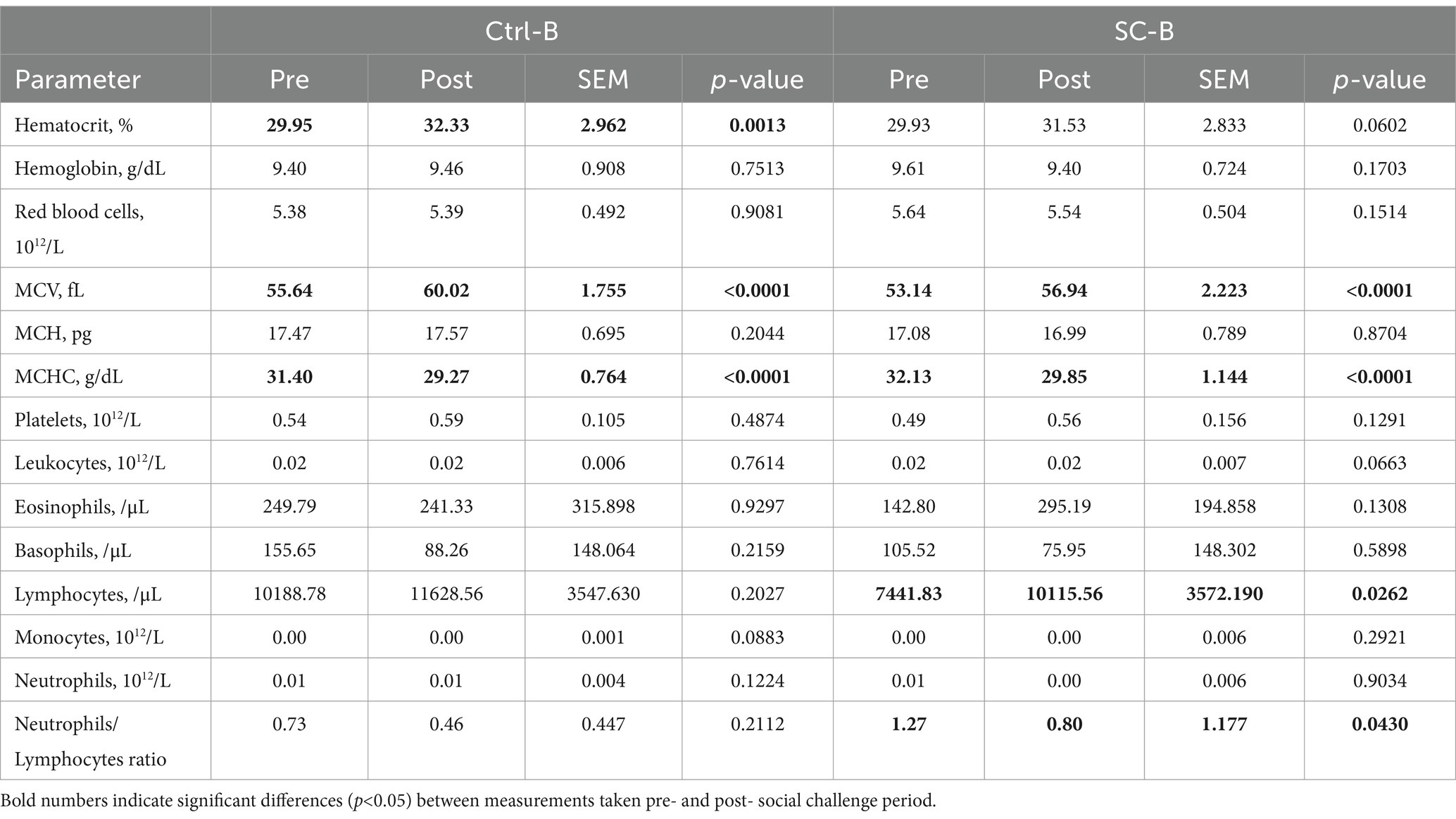
Table 3. Hematological parameters from treatment groups Ctrl-A and SC-A before (Pre) and after (Post) social challenge period (from day 33 post-weaning until day 36) on trial B.
In trial A, Ctrl-A eosinophils, basophils, and neutrophils counts did not present differences after the social mixing. SC-A platelet, eosinophils, and neutrophils counts were not different between pre- and post-social challenge. The Ctrl-A ratio of neutrophils/lymphocytes was not different pre- and post-challenge, but the SC-A neutrophils/lymphocytes ratio presented a trend to increase after the challenge period (29.79%). On the other hand, hemoglobin concentration (3.48%), RBC count (2.79%), MCHC (6.00%), leukocytes (13.61%), and monocytes (29.37%) counts were reduced after the social challenge period in the Ctrl-A piglets. Similarly, SC-A piglets had significant reductions in hemoglobin concentration (7.66%), RBC count (6.36%), MCH concentration (1.48%), MCHC (5.33%), leukocytes (17.14%), lymphocytes (19.67%), and monocytes (21.39%) counts after the social mixing. MCV increased after the social mixing in both Ctrl-A (5.71%) and SC-A (3.94%) groups. Ctrl-A MCH concentration (0.77%) and lymphocyte counts (7.67%) tended to decrease, and hematocrit percentage (2.76%) and platelet counts (12.59%) tended to increase after the social challenge in the Ctrl-A group. The SC-A hematocrit percentage (2.55%) and basophil count (14.79%) tended to be reduced after the social challenge period. When comparing treatment groups and the interaction of the treatment*day factors in terms of the different hematological parameters, no significant differences were found (p > 0.05).
On trial B, no significant differences between the pre- and post-social challenge were detected on Ctrl-B piglets’ hemoglobin and MCH concentrations, RBC, platelets, leukocytes, eosinophils, basophils, lymphocytes, and neutrophils counts, and on SC-B piglets’ hemoglobin and MCH concentrations, RCB, platelets, eosinophils, basophils, monocytes and neutrophils counts. The Ctrl-B- neutrophils/lymphocyte ratio did not differ between pre- and post-challenge period measurements. Contrarily, Ctrl-B piglets’ hematocrit percentage (7.92%) and MCV (7.87%) were increased after the social challenge period, but Ctrl-B MCHC decreased after the challenge period (6.78%). SC-B pigs’ MCV concentration (7.17%) and lymphocyte count (35.92%) increased after the social mixing, although SC-B MCHC was reduced after the social challenge (7.06%). Monocyte count tended to increase on Ctrl-B piglets after the challenge period (3.84%), and hematocrit concentration (5.36%) and leukocyte count (17.75%) tended to increase after the challenge on the SC-B piglets. The SC-B neutrophil/lymphocyte ratio decreased significantly (37.04%) after the social challenge. Similarly to trial A, there were no significant differences observed between treatment groups or the interaction treatment*day in the various hematological parameters (p > 0.05).
3.2.2 Plasmatic cortisol
Plasmatic cortisol in trials A & B are depicted in Figure 3. In trial A, no significant differences in plasmatic cortisol were detected after the social challenge (between day 25 and day 28 after weaning) in any treatment group (Ctr-A p = 0.7202; SC-A p = 0.3623). Plasmatic cortisol concentration in the Ctrl-B had a declining trend (p = 0.0711) after the social challenge period (between day 33 and day 36 after weaning). On the other hand, the plasmatic cortisol concentration of SC-B piglets was not different after the social challenge (p = 0.7187).
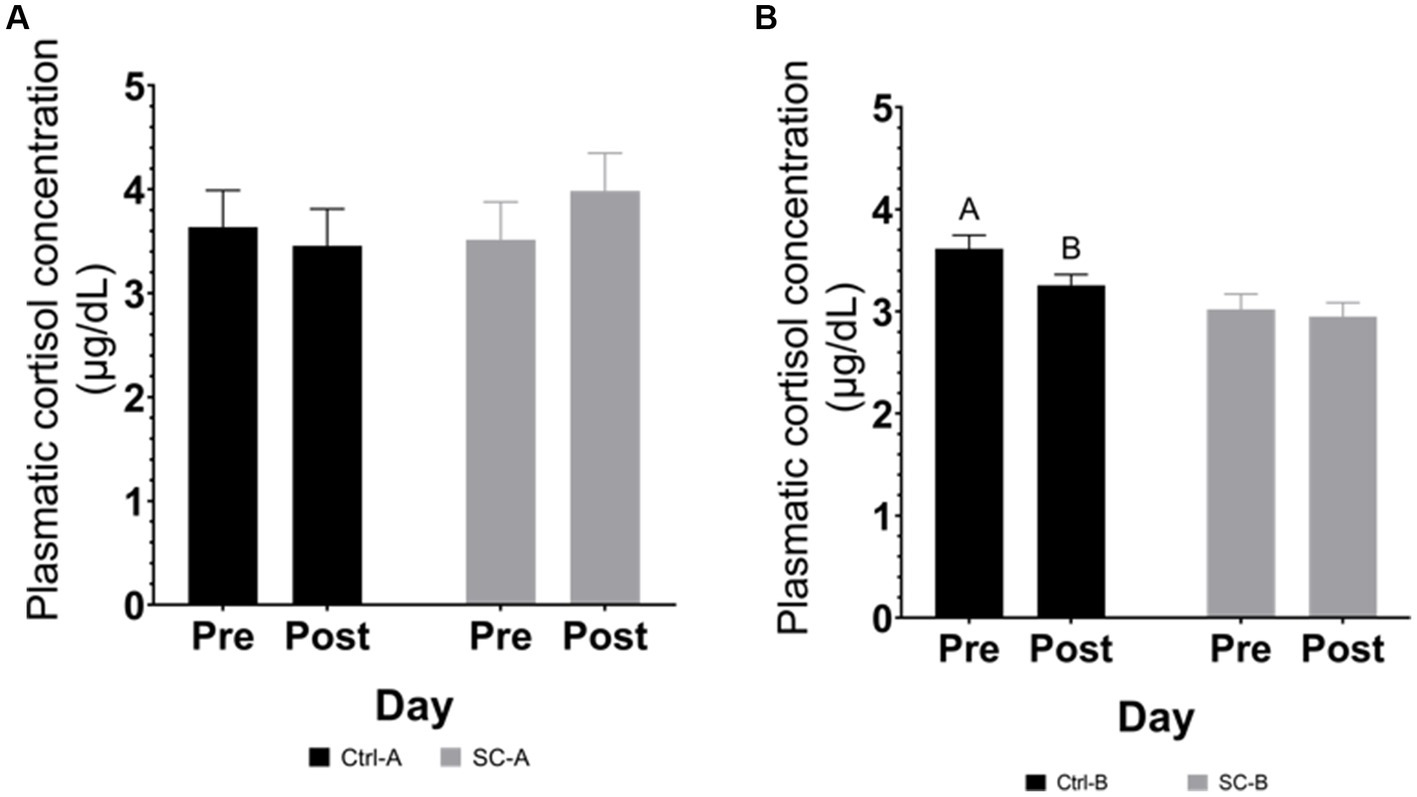
Figure 3. Plasmatic cortisol concentration by treatment groups on days pre- and post-social mixing. (A) Trial A: all animals in the same experimental room at 25 days after weaning (pre-social challenge) and at 28 days after weaning (post-social challenge). (B) Trial B: animals with different treatments in different rooms at 33 days after weaning (pre-social challenge) and 36 days after weaning (post-social challenge). Different lowercase letters indicate significant differences (p < 0.05) whereas near-significant trends were considered at 0.05 < p ≤ 0.10 and represented with capital letters.
3.2.3 Salivary cortisol
Cortisol concentration in saliva is represented in Figure 4. In Trial A no significant differences were detected in any of the tested conditions, neither treatment nor date (between day 25 and day 28 after weaning). In Trial B, Ctrl-B piglets displayed higher cortisol concentration than SC-B piglets (p = 0.0187) before the social stress (day 33 after weaning). After the stress period (day 36 after weaning), Ctrl-B showed a reduction in salivary cortisol (p = 0.0394) while no salivary cortisol changes were detected in the SC-B group (p = 0.9480).

Figure 4. Salivary cortisol concentration by treatment groups on days pre- and post-social mixing. (A) Trial A: all animals in the same experimental room at 25 days after weaning (pre-social challenge) and at 28 days after weaning (post-social challenge). (B) Trial B: animals with different treatments in different rooms at 33 days after weaning (pre-social challenge) and 36 days after weaning (post-social challenge). Different lowercase letters indicate significant differences (p < 0.05) whereas near-significant trends were considered at 0.05 < p ≤ 0.10 and represented with capital letters.
4 Discussion
The spread of agonistic behaviors among pigs may trigger social stress (21). Social stress in pig production is considered a major animal welfare issue (14). Therefore, a deep understanding of the factors that can trigger and spread aggression among pigs is important for production stakeholders to minimize stress and improve animal welfare. The goal of the current research was to study the appearance and effects of agonistic behaviors triggered by systematic social mixing procedures in piglets. This knowledge will help pig stakeholders develop strategies to mitigate the impacts of social stress on pigs.
In Trial A, both Ctrl-A and SC-A treatment groups were randomly placed in the same experimental room. In contrast in Trial B, Ctrl-B and SC-B piglets were placed in separate experimental rooms. As anticipated, piglets exposed to social mixing in both trials (SC-A and SC-B) exhibited significant increases in body lesion scores. This outcome aligns with findings from other studies that also investigated the impacts of social mixing (21, 25, 26). Surprisingly, Ctrl-A piglets also displayed a notable increase in body lesion scores, although to a lesser degree compared to the pronounced increase observed in SC-A piglets following social mixing. It is worth noting that this increase occurred even in the absence of deliberate exposure to the social challenge. Furthermore, the observed body lesions in both Ctrl-A and SC-A groups were primarily concentrated in the front and middle areas of their bodies. Several authors (21, 27–29) reported that lesions accumulation in the front and middle areas are related to fights and confrontations, while body lesions in the rear part of the body of the pig might be related with insufficient access to the feeder or the drinker, respectively. Additionally, the change in hematological parameters was comparable in both Ctrl-A and SC-A pigs after social mixing. Therefore, it is possible to assume that Ctrl-A piglets engage in agonistic interactions and confrontations without experiencing a social challenge. Contrarily, in trial B, where the treatment groups were physically separated, Ctrl-B lesion scores and hematological parameters were unaffected during the social mixture period, while the SC-B piglets had a significantly higher lesion score.
Regarding the hematological parameters measured, pre- and post-social challenge results fell within the reference intervals (30). Hematological parameters are particularly sensitive to the impacts of stress, influencing fundamental functions like growth, maintenance, and immune activities (31–33). Hematocrit percentage, hemoglobin concentration, and red blood cell counts have been shown to have connections with pig growth and voluntary feed intake (34). In both Ctrl-A and SC-A groups, there was a similar pattern in hemoglobin concentration and red blood cell counts after the social challenge. These indicators displayed decreased levels, likely influenced by the stress experienced by the piglets (35, 36). Parameters linked to the morphology of red blood cells, such as MCV and MCHC, were similarly influenced in both Ctrl-A and SC-A piglets. This similarity suggests that social stress was induced in SC-A pigs, whereas Ctrl-A pigs might experience a stress response triggered by the higher activity caused by the pen neighbor’s noise or the auditory cues of aggression from other pigs in the room.
Concerning immune activity parameters, the process of social mixing led to a decrease in the activity of leukocytes, lymphocytes, and monocytes in both Ctrl-A and SC-A piglets. Acute stress can disrupt the typical concentration of these indicators, as cortisol affects the organism’s response (31, 33). Meanwhile, in Trial B, where both Ctrl-B and SC-B pigs were physically separated, the immune activity of Ctrl-B pigs remained unaffected following the social challenge period. The only change observed was an increase in hematocrit percentage among Ctrl-B pigs during that timeframe, which could be attributed to their natural growth pattern as the piglets are in the growing stage (34). On the other hand, lymphocyte activity increased after the social mixing in the SC-B piglets. Thus, Ctrl-B piglets were not affected at the same level as Ctrl-A during the social mixing period, and SC-B piglets did show a milder response to social mixing compared to SC-A. Even if aggression levels were expected to be more severe in SC-B piglets, compared to SC-A, as older or heavier animals are expected to engage in more intense agonistic interactions (7, 37, 38), skin lesion score and hematological parameters were less severe relative to SC-A piglets. This milder physiological response of the SC-B piglets might be related to the fact that the piglets on trial B were older and heavier relative to trial A piglets, they might have been able to cope better with the physiological imbalance, which could have made them more resilient to the social challenge (39). The reductions of MCHC in both Ctrl and SC in both trials A and B might be related to the acute stress of the sampling protocol performed to obtain the blood samples (31).
In both trials, measurements of cortisol concentration in both blood and saliva were unable to identify changes following the social mixing period. This result might be attributed to the sampling schedule, potentially missing the peak of cortisol activity. Cortisol levels are expected to increase as a physiological response to the aggression arising from social mixing, including fights, confrontations, and intimidations (9, 40, 41). This reaction has been observed to occur within the initial hours (ranging from 40 min to 5 h) following the stress event (4, 42, 43). Moreover, pig confrontations resulting from social mixing have been documented to take place within the initial 90 min after mixing (4). In this study, saliva and blood sampling were conducted 24 h after the final social mixing, once the pigs had returned to their original pen. Post social mixing sampling point might have overlooked the peak cortisol concentration as cortisol concentration might be dropped after 24 h of stress peak (44). The initial intention of cortisol (blood and saliva) pre and post-social mixing was to minimize constant manipulation for the piglets that might add stress due to handling or operator presence, affecting the physiological responses of the pigs (45). Additionally, the purpose of the repeated social mixing protocol (spanning 3 consecutive days) was to escalate social stress. Nonetheless, the repetition of the social mixing event could lead to a decrease in the intensity of fights and confrontations (4), the intensity observed after the mixing period could potentially be the least among the 3 days of the social challenge. Cortisol or any other biomarker as a stress indicator is fragile and easily disrupted by several factors (44), some of which are the handling (invasive) procedures and the appropriate time to collect the measurement sample. Moreover, these traditional measurement methodologies are time-spot and might lose information (18), as occurred in the current study. As an alternative to traditional measurements, behavioral assessments (remote method) are emerging as a solution to detect abnormalities or a high incidence of agonistic behaviors (18, 46). Computer vision and machine learning have boosted the evolution of technology capable of detecting situations that can trigger stress and deteriorate the welfare of animals in production conditions. Gómez et al. (47), present computer vision technologies (i.e., cameras, accelerometers, and other PLF devices to record animal activity and aggressive interactions) externally or internally validated capable of detecting and measuring antagonistic activities among pigs.
Therefore, two hypotheses might be proposed based on the results obtained and the interpretation of the information collected (i.e., biomarkers and behavioral information). When pigs are exposed to loud noises, it can change their behavior, such as increasing their activity level and curiosity toward the source of the sound or causing them to freeze. Also, it can raise their heart rate (48). Therefore, during the social mixing process on a farm, the loud sounds (i.e., vocalizations, grunts, stepping sounds, sounds of the lids closing sounds, and sounds from piglets’ collisions against the pen walls) from the staff and the activity in the mixed pens could cause anxiety and fear in the Ctrl-A piglets, who were not part of the mixing process. This could increase the physical activity of the Ctrl-A piglets and the chance to invade other individuals’ spaces, making them more likely to engage in confrontations and display aggressive behaviors. According to a study by Talling et al. (48), loud noises can cause small behavioral responses such as freezing, curiosity, or higher physical activity. However, Talling et al. did not describe if such higher activity triggered by the noise could provoke intense behavioral responses like fighting or escaping.
Another more complex hypothesis to explain the unexpected display of aggressive behaviors in Ctrl-A piglets, even in the absence of direct exposure to social mixing, might be the phenomenon of emotional contagion. Emotional contagion has been described as “The emotional state matching of a subject with an object” (i.e., the adoption of emotions of others) (49). Emotional contagion is provoked when an individual perceives another individual’s emotions and mimics the behavioral and physiological states of the transmitter (50). Emotional contagion has been theorized as a communication strategy to share environmental information with members of the community (51, 52). This information and cues can affect the physiological status, and the behavior of the observer animal (49, 50). Emotional contagion can disseminate emotional valence (positive or negative), and arousal level (high and low) (53). In line with this, a potential explanation is that the fighting grunts and fear/pain vocalizations (auditive cues) of piglets engaged in confrontations during the social challenge could potentially make Ctrl-A piglets more anxious and predisposed to engaging in fights with their pen mates (54, 55). Emotional contagion seems conceivable when the behavior and physiological responses of the observer align with those of the stressed individual (50). This phenomenon can be conveyed through visual means (54) or auditive cues (56–59). Only a limited number of studies have focused on comprehending the factors that govern this behavioral mimicry mechanism. Nonetheless, research in other species (i.e., rodents, nonhuman primates, ruminants, dogs, birds, and fishes, among others), as mentioned earlier, has extensively explored this phenomenon (52). However, the current experimental design does not allow for the confirmation of the hypothesis of emotional contagion.
5 Conclusion
In this study, the social challenge used resulted in aggressive behaviors among piglets, provoking mild to acute social stress in both trials. The social stress response was observed in the mixed piglets due to changes in the physiological condition of the animals. This was evidenced by an increase in body lesions and alterations in hematological parameters. Conversely, piglets without social mixing events also engaged in confrontations with their pen mates, as indicated by an increased skin lesion score and similar patterns in hematological markers compared to mixed piglets when they were in the same experimental room. This might suggest a phenomenon of contagion among the experimental pens. When socially mixed piglets and non-mixed piglets were physically located in different rooms, no changes were detected in the non-socially mixed pigs as a result of aggressive behaviors from socially mixed piglets. Consequently, future research endeavors must acknowledge that treatment groups causing behavioral responses, such as general noise or pain-related or stress vocalizations, could potentially alter the behavior of the animals in different treatment groups.
Moreover, invasive and time spot monitoring methods did not completely detect the physiological responses triggered by the social challenge. It’s important to note that spot-time measurements and scan samples may miss critical information that holds key relevance for concluding studies and research projects. The development of remote, minimally invasive, and continuous measurement methods is of great importance for animal research. These methods enhance the accuracy of monitoring animal welfare without causing disruptions to the animals. Furthermore, to collect the most information triggered by social stress, measurements for aggressive behaviors and stress biomarkers are suggested to be collected within the following hours after the regrouping event.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the Laboratory Animal Care Advisory Committee of the Faculty of Veterinary Sciences of the Autonomous University of Barcelona (CEEAH-5754-5755-CEEA-UAB). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
RG: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. SL-V: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing. JP: Data curation, Formal analysis, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing – review & editing. XM: Investigation, Supervision, Validation, Writing – review & editing. GT: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing. PL: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. PL received funding from Ramon y Cajal program of the Spanish Ministry of Science (Spain; Grant Ref. RYC2020-029067-I). RG received funding from the program Industrial Doctorate of the regional government of Catalonia (Spain; Grant Ref. 2021 DI 00071).
Acknowledgments
We would like to thank the farm staff, animal caretakers, and veterinarians who performed the study at both experimental farms. Also, to acknowledge and reward Lucta’s laboratory staff, Almudena Martínez and Marc Vujadinovic, oversaw running all the laboratory analyses required to obtain the results of both studies presented in the current manuscript.
Conflict of interest
RG was employed by AWEC Advisors S.L. The authors declare that this study received funding from Lucta SA. The funder had the following involvement in the study: covering all expenses coming from the use of Lucta’s facilities, personnel, materials and equipments required for running the experiments, and analysis.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Büttner, K, Czycholl, I, Mees, K, and Krieter, J. Agonistic interactions in pigs-comparison of dominance indices with parameters derived from social network analysis in three age groups. Animals. (2019) 9:929. doi: 10.3390/ani9110929
2. Hou, YH, Chen, J, Tong, GZ, Tian, ZJ, Zhou, YJ, Li, GX, et al. A recombinant plasmid co-expressing swine ubiquitin and the GP5 encoding-gene of porcine reproductive and respiratory syndrome virus induces protective immunity in piglets. Vaccine. (2008) 26:1438–49. doi: 10.1016/j.vaccine.2007.12.057
3. Salazar, LC, Ko, HL, Yang, CH, Llonch, L, Manteca, X, Camerlink, I, et al. Early socialisation as a strategy to increase piglets’ social skills in intensive farming conditions. Appl Anim Behav Sci. (2018) 206:25–31. doi: 10.1016/j.applanim.2018.05.033
4. Coutellier, L, Arnould, C, Boissy, A, Orgeur, P, Prunier, A, Veissier, I, et al. Pig’s responses to repeated social regrouping and relocation during the growing-finishing period. Appl Anim Behav Sci. (2007) 105:102–14. doi: 10.1016/j.applanim.2006.05.007
5. Gimsa, U, Tuchscherer, M, and Kanitz, E. Psychosocial stress and immunity—what can we learn from pig studies? Front Behav Neurosci. (2018) 12:64. doi: 10.3389/fnbeh.2018.00064
6. Camerlink, I, Arnott, G, Farish, M, and Turner, SP. Complex contests and the influence of aggressiveness in pigs. Anim Behav. (2016) 121:71–8. doi: 10.1016/j.anbehav.2016.08.021
7. D’Eath, RB. Individual aggressiveness measured in a resident-intruder test predicts the persistence of aggressive behaviour and weight gain of young pigs after mixing. Appl Anim Behav Sci. (2002) 77:267–83. doi: 10.1016/S0168-1591(02)00077-1
8. Turner, SP, Nevison, IM, Desire, S, Camerlink, I, Roehe, R, Ison, SH, et al. Aggressive behaviour at regrouping is a poor predictor of chronic aggression in stable social groups. Appl Anim Behav Sci. (2017) 191:98–106. doi: 10.1016/j.applanim.2017.02.002
9. Escribano, D, Ko, HL, Chong, Q, Llonch, L, Manteca, X, and Llonch, P. Salivary biomarkers to monitor stress due to aggression after weaning in piglets. Res Vet Sci. (2019) 123:178–83. doi: 10.1016/j.rvsc.2019.01.014
10. Teixeira, DL, and Boyle, LA. A comparison of the impact of behaviours performed by entire male and female pigs prior to slaughter on skin lesion scores of the carcass. Livestock Sci. (2014) 170:142–149. doi: 10.1016/j.livsci.2014.09.026
11. Niekamp, SR, Sutherland, MA, Dahl, GE, and Salak-Johnson, JL. Immune responses of piglets to weaning stress: impacts of photoperiod. J Animal Sci. (2007) 85:93–100. doi: 10.2527/jas.2006-153
12. Oster, M, Muráni, E, Ponsuksili, S, D’Eath, RB, Turner, SP, Evans, G, et al. Hepatic expression patterns in psychosocially high-stressed pigs suggest mechanisms following allostatic principles. Physiol Behav. (2014) 128:159–65. doi: 10.1016/j.physbeh.2014.02.014
13. Couret, D, Otten, W, Puppe, B, Prunier, A, and Merlot, E. Behavioural, endocrine and immune responses to repeated social stress in pregnant gilts. Animal. (2009) 3:118–27. doi: 10.1017/S1751731108003236
14. De Groot, J, Ruis, M, Scholten, JW, Koolhaas, JM, and Boersma, W. Long-term effects of social stress on anti-viral immunity in pigs. Physiol Behav. (2001) 73:145–58. doi: 10.1525/california/9780520233256.003.0009
15. Heo, J, Kattesh, HG, Roberts, MP, Morrow, JL, Dailey, JW, and Saxton, AM. Hepatic corticosteroid-binding globulin (CBG) messenger RNA expression and plasma CBG concentrations in young pigs in response to heat and social stress. J Animal Sci. (2005) 83:208–15. doi: 10.2527/2005.831208x
16. Wellock, IJ, Emmans, GC, and Kyriazakis, I. Predicting the consequences of social stressors on pig food intake and performance. J Anim Sci. (2003) 81:2995–3007. doi: 10.2527/2003.81122995x
17. Boogaard, BK, Boekhorst, LJS, Oosting, SJ, and Sørensen, JT. Socio-cultural sustainability of pig production: citizen perceptions in the Netherlands and Denmark. Livest Sci. (2011) 140:189–200. doi: 10.1016/j.livsci.2011.03.028
18. Guevara, RD, Pastor, JJ, Manteca, X, Tedo, G, and Llonch, P. Systematic review of animal-based indicators to measure thermal, social, and immune-related stress in pigs. PLoS One. (2022) 17:e0266524. doi: 10.1371/journal.pone.0266524
19. Prunier, A, Averos, X, Dimitrov, I, Edwards, SA, Hillmann, E, Holinger, M, et al. Review: early life predisposing factors for biting in pigs. Animal. (2020) 14:570–87. doi: 10.1017/S1751731119001940
20. National Research Council. Nutrient requirements of swine. Washington, DC: National Academies Press (2012).
21. Turner, SP, Farnworth, MJ, White, IMS, Brotherstone, S, Mendl, M, Knap, P, et al. The accumulation of skin lesions and their use as a predictor of individual aggressiveness in pigs. Appl Anim Behav Sci. (2006) 96:245–59. doi: 10.1016/j.applanim.2005.06.009
22. Goering, MJ, Mumm, JM, Coffin, MJ, Bortoluzzi, EM, Ruiz, LA, and Hulbert, LE. 502 low-stress sampling and cortisol concentrations in periparturient sows. J Anim Sci. (2018) 96:268–8. doi: 10.1093/jas/sky073.499
23. Escribano, D, Gutiérrez, AM, Tecles, F, and Cerón, JJ. Changes in saliva biomarkers of stress and immunity in domestic pigs exposed to a psychosocial stressor. Res Vet Sci. (2015) 102:38–44. doi: 10.1016/j.rvsc.2015.07.013
24. Escribano, D, Fuentes-Rubio, M, and Cerón, JJ. Validation of an automated chemiluminescent immunoassay for salivary cortisol measurements in pigs. J Vet Diagnostic Investig. (2012) 24:918–23. doi: 10.1177/1040638712455171
25. Poletto, R, Kretzer, FC, and Hötzel, MJ. Minimizing aggression during mixing of gestating sows with supplementation of a tryptophan-enriched diet. Physiol Behav. (2014) 132:36–43. doi: 10.1016/j.physbeh.2014.04.043
26. Tönepöhl, B, Appel, AK, Voß, B, König von Borstel, U, and Gauly, M. Interaction between sows’ aggressiveness post mixing and skin lesions recorded several weeks later. Appl Anim Behav Sci. (2013) 144:108–15. doi: 10.1016/j.applanim.2013.01.004
27. Ko, HL, Chong, Q, Escribano, D, Camerlink, I, Manteca, X, and Llonch, P. Pre-weaning socialization and environmental enrichment affect life-long response to regrouping in commercially-reared pigs. Appl Anim Behav Sci. (2020) 229:105044. doi: 10.1016/j.applanim.2020.105044
28. Stukenborg, A, Traulsen, I, Puppe, B, Presuhn, U, and Krieter, J. Agonistic behaviour after mixing in pigs under commercial farm conditions. Appl Anim Behav Sci. (2011) 129:28–35. doi: 10.1016/j.applanim.2010.10.004
29. Stukenborg, A, Traulsen, I, Stamer, E, Puppe, B, and Krieter, J. The use of a lesion score as an indicator for agonistic behaviour in pigs. Arch Anim Breed. (2012) 55:163–70. doi: 10.5194/aab-55-163-2012
30. Carr, J, and Marín, H In: A Gráficas, editor. Guía práctica para el manejo del ganado porcino. SERVET (2014)
31. Bacou, E, Haurogné, K, Mignot, G, Allard, M, De Beaurepaire, L, Marchand, J, et al. Acute social stress-induced immunomodulation in pigs high and low responders to ACTH. Physiol Behav. (2017) 169:1–8. doi: 10.1016/j.physbeh.2016.11.012
32. Barnett, JL, Hemsworth, PH, and Hand, AM. Effects of chronic stress on some blood parameters in the pig. Appl Anim Ethol. (1983) 9:273–7. doi: 10.1016/0304-3762(83)90007-X
33. Elenkov, IJ, and Chrousos, GP. Stress hormones, proinflammatory and antiinflammatory cytokines, and autoimmunity. Ann N Y Acad Sci. (2002) 966:290–303. doi: 10.1111/j.1749-6632.2002.tb04229.x
34. Lindholm-Perry, AK, Kuehn, LA, Wells, JE, Rempel, LA, Chitko-Mckown, CG, Keel, BN, et al. Hematology parameters as potential indicators of feed efficiency in pigs. Transl Anim Sci. (2021) 5:txab219. doi: 10.1093/tas/txab219
35. Dubreuil, P, Couture, Y, Farmer, C, and Petitclerc, D. Hematological and biochemical changes following an acute stress in control and somatostatin-immunized pigs. Can J Anim Sci. (1993) 73:241–52. doi: 10.4141/cjas93-026
36. Hicks, TA, McGlone, JJ, Whisnant, CS, Kattesh, HG, and Norman, RL. Behavioral, endocrine, immune, and performance measures for pigs exposed to acute stress. J Anim Sci. (1998) 76:474–83. doi: 10.2527/1998.762474x
37. D’Eath, RB. Consistency of aggressive temperament in domestic pigs: the effects of social experience and social disruption. Aggress Behav. (2004) 30:435–48. doi: 10.1002/ab.20077
38. D’Eath, RB. Socialising piglets before weaning improves social hierarchy formation when pigs are mixed post-weaning. Appl Anim Behav Sci. (2005) 93:199–211. doi: 10.1016/j.applanim.2004.11.019
39. Islam, MA, Uddin, MJ, Tholen, E, Tesfaye, D, Looft, C, Schellander, K, et al. Age-associated differential production of IFN-γ, IL-10 and GM-CSF by porcine alveolar macrophages in response to lipopolysaccharide. Vet J. (2013) 198:245–51. doi: 10.1016/j.tvjl.2013.07.026
40. Camerlink, I, Peijnenburg, M, Wemelsfelder, F, and Turner, SP. Emotions after victory or defeat assessed through qualitative behavioural assessment, skin lesions and blood parameters in pigs. Appl Anim Behav Sci. (2016) 183:28–34. doi: 10.1016/j.applanim.2016.07.007
41. Oldham, L, Camerlink, I, Arnott, G, Doeschl-Wilson, A, Farish, M, and Turner, SP. Winner–loser effects overrule aggressiveness during the early stages of contests between pigs. Sci Rep. (2020) 10:13338–13. doi: 10.1038/s41598-020-69664-x
42. Němečková, M, Popelková, T, and Chloupek, P. Sampling methods for determination of cortisol in pig saliva and their use in the assessment of pig welfare. Acta Vet Brno. (2022) 91:261–6. doi: 10.2754/avb202291030261
43. Shen, C, Tong, X, Chen, R, Gao, S, Liu, X, Schinckel, AP, et al. Identifying blood-based biomarkers associated with aggression in weaned pigs after mixing. Appl Anim Behav Sci. (2020) 224:104927. doi: 10.1016/j.applanim.2019.104927
44. Ruis, MAW, Te Brake, JHA, Engel, B, Ekkel, ED, Buist, WG, Blokhuis, HJ, et al. The circadian rhythm of salivary cortisol in growing pigs: effects of age, gender, and stress. Physiol Behav. (1997) 62:623–30. doi: 10.1016/S0031-9384(97)00177-7
45. Baldi, A, Verga, M, Maffii, M, Canali, E, Chiaraviglio, D, and Ferrari, C. Effects of blood sampling procedures, grouping and adrenal stimulation on stress responses in the growing pig. Reprod Nutr Dév. (1989) 29:95–103. doi: 10.1051/rnd:19890108
46. Dybkjær, L. The identification of behavioural indicators of “stress” in early weaned piglets. Appl Anim Behav Sci. (1992) 35:135–47. doi: 10.1016/0168-1591(92)90004-U
47. Gómez, Y, Stygar, AH, Boumans, IJMM, Bokkers, EAM, Galán, E, Llonch, P, et al. A systematic review on validated precision livestock farming technologies for pig production and its potential to assess animal welfare. Front Vet Sci. (2021) 8:660565. doi: 10.3389/fvets.2021.660565
48. Talling, JC, Waran, NK, Wathes, CM, and Lines, JA. Behavioural and physiological responses of pigs to sound. Appl Anim Behav Sci. (1996) 48:187–201. doi: 10.1016/0168-1591(96)01029-5
49. de Waal, FBM. Putting the altruism Back into altruism: the evolution of empathy. Annu Rev Psychol. (2008) 59:279–300. doi: 10.1146/annurev.psych.59.103006.093625
50. Düpjan, S, Krause, A, Moscovice, LR, and Nawroth, C. Emotional contagion and its implications for animal welfare. CAB Rev Perspect Agric Vet Sci Nutr Nat Resour. (2020) 15:1–6. doi: 10.1079/PAVSNNR202015046
51. Paul, ES, and Mendl, MT. Animal emotion: descriptive and prescriptive definitions and their implications for a comparative perspective. Appl Anim Behav Sci. (2018) 205:202–9. doi: 10.1016/j.applanim.2018.01.008
52. Pérez-Manrique, A, and Gomila, A. Emotional contagion in nonhuman animals: a review. WIREs Cogn Sci. (2022) 13:e1560. doi: 10.1002/wcs.1560
53. Mendl, M, Burman, OHP, and Paul, ES. An integrative and functional framework for the study of animal emotion and mood. Proc R Soc B Biol Sci. (2010) 277:2895–904. doi: 10.1098/rspb.2010.0303
54. Goumon, S, and Špinka, M. Emotional contagion of distress in young pigs is potentiated by previous exposure to the same stressor. Anim Cogn. (2016) 19:501–11. doi: 10.1007/s10071-015-0950-5
55. Reimert, I, Bolhuis, JE, Kemp, B, and Rodenburg, TB. Social support in pigs with different coping styles. Physiol Behav. (2014) 129:221–9. doi: 10.1016/j.physbeh.2014.02.059
56. Düpjan, S, Tuchscherer, A, Langbein, J, Schön, P-C, Manteuffel, G, and Puppe, B. Behavioural and cardiac responses towards conspecific distress calls in domestic pigs (Sus scrofa). Physiol Behav. (2011) 103:445–52. doi: 10.1016/j.physbeh.2011.03.017
57. Reimert, I, Bolhuis, JE, Kemp, B, and Rodenburg, TB. Indicators of positive and negative emotions and emotional contagion in pigs. Physiol Behav. (2013) 109:42–50. doi: 10.1016/j.physbeh.2012.11.002
58. Reimert, I, Bolhuis, JE, Kemp, B, and Rodenburg, TB. Emotions on the loose: emotional contagion and the role of oxytocin in pigs. Anim Cogn. (2015) 18:517–32. doi: 10.1007/s10071-014-0820-6
Keywords: social stress, pig welfare, pig behavior, piglets, emotional contagion, aggression
Citation: Guevara RD, López-Vergé S, Pastor JJ, Manteca X, Tedo G and Llonch P (2024) When the neighbors are noisy: effect of social challenge in collateral pens of stressed animals. Front. Vet. Sci. 11:1433628. doi: 10.3389/fvets.2024.1433628
Edited by:
Leonie Jacobs, Virginia Tech, United StatesReviewed by:
Oceane Schmitt, University of Bristol, United KingdomTimothy John Mahony, The University of Queensland, Australia
Copyright © 2024 Guevara, López-Vergé, Pastor, Manteca, Tedo and Llonch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raúl David Guevara, cmF1bC5ndWV2YXJhQGF3ZWMuZXM=