 Benjamín Bennett-Laso1,2†
Benjamín Bennett-Laso1,2† Bárbara Berazay1,3†Gabriela Muñoz1,2
Bárbara Berazay1,3†Gabriela Muñoz1,2 Naomi Ariyama1,2Nikita Enciso1
Naomi Ariyama1,2Nikita Enciso1 Christina Braun4
Christina Braun4 Lucas Krüger5,6
Lucas Krüger5,6 Miloš Barták7
Miloš Barták7 Marcelo González-Aravena5
Marcelo González-Aravena5 Victor Neira1*
Victor Neira1*- 1Departamento de Medicina Preventiva Animal, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile, Santiago, Chile
- 2Programa de Doctorado en Ciencias Silvoagropecuarias y Veterinarias, Universidad de Chile, Santiago, Chile
- 3Programa de Magister en Ciencias Animales y Veterinarias, Universidad de Chile, Santiago, Chile
- 4Polar & Bird Ecology Group, Institute of Ecology and Evolution, Friedrich Schiller University Jena, Jena, Germany
- 5Instituto Antártico Chileno, Punta Arenas, Chile
- 6Millennium Institute of Biodiversity of Antarctic and Subantarctic Ecosystems (BASE), Santiago, Chile
- 7Department of Experimental Biology, Masaryk University, Faculty of Science, Brno, Czechia
From December 2023 to March 2024, a surveillance program aiming to detect Highly Pathogenic Avian Influenza (HPAI) H5N1 was conducted on Antarctica territories, specifically at Fildes Peninsula (King George Island, Maritime Antarctic), and James Ross Island. At Fildes Peninsula, samples from marine birds and mammals were collected from four accessible sampling locations with significant animal colonies: Ardley Island, hosting a large concentration of Gentoo penguins (Pygoscelis papua); Ardley Cove, where small groups of likely non-breeding Chinstrap penguins (Pygoscelis antarcticus) were present; seal haul-out sites of Southern elephant (Mirounga leonina) and Weddell (Leptonycotes wedellii); and, a nesting site of Southern giant petrels (Macronectes giganteus). Additionally, six samples were collected from five dead skuas near the Lachman lakes on James Ross Island (63.7989S, 57.8105W) on March 3, 2024. Despite collecting a total of 943 samples from Fildes Peninsula, all results tested negative for HPAI, and no animals displayed clinical signs or behaviors consistent with HPAI infection. However, all skua samples from James Ross Island tested positive for HPAI H5N1 clade 2.3.4.4 by specific real-time RT-PCR reactions, confirming the first recorded HPAI-related mortality event in Antarctica (south of 60°S), specifically in skuas. Further research is necessary to genetically characterize the virus and better understand the role of skuas in viral dissemination in Antarctica.
1 Introduction
Highly pathogenic avian influenza (HPAI) subtype H5NX, clade 2.3.4.4b has been responsible for numerous global outbreaks, leading to multiple mass mortality events in avian wildlife and marine mammal populations. Human cases of this virus have also been reported, underscoring its zoonotic potential and public health risk (1–3). Additionally, HPAI infections may significantly stress breeding seabirds, potentially resulting in breeding failure or death due to increased energy demands (4, 5). The rapid and widespread dissemination of this virus is primarily driven by infected migratory species (6–8), particularly those with long-range dispersal capabilities (9, 10). The current HPAI H5N1 strain, clade 2.3.4.4b, emerged in 2020 after extensive viral evolution and was first confirmed in South America in October 2022 (11). Prior to this, the virus had not been detected in Oceania (Australia and New Zealand) or Antarctica.
Given the close proximity of South America to Antarctica, coupled with the migratory patterns of seabirds between the two regions, concerns have risen about the virus’s potential introduction to Antarctica via migratory birds. Previous evidence suggests that influenza viruses detected in Antarctica are genetically related to strains found outside the continent (12, 13). Migratory species such as Arctic terns (Sterna paradisaea) (14) and South polar skuas (Stercorarius maccormicki) (15) are known to undertake trans-equatorial migrations, with both species demonstrating the ability to shed the virus. With the arrival of the virus in the southern hemisphere, other species with shorter migration patterns and wide foraging areas, such as giant petrels (Macronectes spp.), and Brown skuas (Stercorarius antarcticus lonnbergi) may play a more significant role in viral spread (16), potentially introducing the virus to Antarctic wildlife before developing visible clinical signs. Although some positive cases of HPAI H5NX 2.3.4.4b have been confirmed on South Georgia island, a Sub-Antarctic territory (17), no published cases have been reported in more southern latitudes. This suggests that Antarctic species may be highly susceptible to infection, making them vulnerable to mass mortality events if the virus spreads to Antarctic colonies (18). To investigate this risk, we conducted surveillance for HPAI in seabirds and marine mammals at key site on King George Island (South Shetland Islands) and the Maritime Antarctic Peninsula, areas close to previously confirmed cases and where wildlife-human interaction is increasing (19, 20). We also coordinated with researchers across the Antarctic Peninsula to receive reports and samples from unexpected mortality events. In response, a Czech Antarctic team alerted us to dead seabirds near Lachman Lakes on James Ross Island (63.7989 S, 57.8105 W), 4 km east of Mendel Base. This report summarizes findings from December 2023 and March 2024, confirming the presence of HPAI H5N1.
2 Materials and methods
2.1 Locations at Fildes Peninsula, King George Island
The surveillance team was based at Professor Julio Escudero Base, located on King George Island, in the South Shetland Islands, Maritime Antarctic (33). From December 16, 2023, to March 23, 2024, regular surveillance expeditions were carried out around the base (see Supplementary Figure S1), covering key wildlife areas with high concentrations of penguins, flying seabirds, and marine mammals. During these expeditions, clinical observation and sample collections were performed. The selected locations were based on known animal aggregations and colony sites, as detailed by environmental monitoring studies (21). Ten specific sites were visited at least once (see Supplementary Figure S1), including Ardley Cove, Ardley Island (ASPA 150), Hydrographers Cove, Diomedea Island, Flat-top Peninsula, Biologists Cove, Basalt Creek, Gradzinski Cove, and Green Point. These locations were selected based on accessibility and the presence of notable species, such as Gentoo penguins (Pygoscelis papua), Chinstrap penguins (Pygoscelis antarcticus), Weddell seals (Leptonychotes weddellii), Southern giant petrels (Macronectes giganteus), Southern elephant seals (Mirounga leonina) and others (Supplementary Table S1). The sampling effort was recorded by tracking researchers’ movements using a Garmin InReach Explorer and a Garmin Enduro Watch device, which recorded geographic positions at 60- and 30-s intervals, respectively. The tracking data was processed using the “trip” R-package (22) to quantify time spent at different sites, while ArcMap was used to visualize the distribution of effort. Additionally, veterinarians conducted clinical observations, monitoring for signs of respiratory, neurologic, or digestive syndromes associated with HPAI H5N1. Sampling was performed on live and dead animals in compliance with the guidelines of the University of Chile’s Institutional Ethics Committee (approval no. 22603-VET-UCH).
2.2 Sampling
Sampling activities were conducted under permission numbers 669–2023, 670–2023, and 201–2024 from the Chilean Antarctic Institute INACH. Field researchers used personal protective equipment (PPE), including coveralls, gloves, face shields, and masks, which were disinfected and safely disposed of at the sampling site. Afterward, the sealed waste was transported to Professor Julio Escudero Base for proper disposal, minimizing the risk of viral spread. Researchers focused on collecting fresh fecal samples to reduce animal handling and increase the number of samples obtained. In some cases, direct samples were collected from live Southern giant petrels, skuas, and opportunistically from freshly deceased animals. Sampling tools included rayon swabs, which were placed into Viral Transport Media (Inactivated Type) (ALLTEST catalog number ITM-001). For environmental samples, pooled groups of five swabs were placed in a single tube of viral transport media. Direct samples from cloacal swabs were collected by capturing and restraining individuals according to pre-approved protocols (069/CEC/2018 and 3/CBSCUA/2022). All procedures adhered to Antarctic Treaty guidelines, and the field team consisted of a biologist and four veterinarians who also monitored the animals for clinical signs of disease. No blood samples were collected, as there was no prior evidence of HPAIV in King George Island populations.
2.3 Unexpected mortality case at James Ross Island
On February 28, 2024, a report of three dead skuas (unidentified species) was received from a field crew working with the Czech Antarctic Research Program near the Base Johann Gregor Mendel (63.7989S, 57.8105W) on James Ross Island. The site, a known nesting area for skuas, is adjacent to the freshwater Lachman lakes. The researchers, who had over 20 years of experience working in the region, indicated that finding multiple dead skuas in one location was unprecedented. Our surveillance team traveled from King George Island to James Ross Island in the Chilean vessel Janequeo (ATF-65), arriving on March 3, 2024. At the site, five dead skuas were found, and samples were collected from all individuals. Swabs from tissues were pooled from different organs, following established protocols. One skua was positively identified as a brown skua (Figure 1), though the state of decomposition and developmental stages made species identification of the other individuals more challenging.

Figure 1. Deceased skua (brown skua) at James Ross Island.
2.4 Diagnostic testing
The collected samples were processed within 24 h after collection at the molecular biology laboratory at Professor Julio Escudero Base. RNA extraction was performed using the acid guanidinium thiocyanate-phenol-chloroform method with Chomczynski phenol solution (Winkler Ltda.), following the manufacturer’s instructions. A real-time reverse transcriptase polymerase chain reaction (RT PCR) assay performed targeting the conserved M gene of the influenza virus was conducted using the primers infA F (5’-GACCRATCCTGTCACCTCTGAC-3′), infA R (5’-AGGGCATTYTGGACAAAKCGTCTA-3′), infA Probe1 (5’ FAM-TGC AGT CCT CGC TCA CTG GGC ACG-BHQ1-3′) (11). Samples with cycle threshold (CT) values greater than 35 were considered inconclusive, while those with CT values greater than 40 were deemed negative for influenza virus. Positive samples were further subtyped and pathotyped to confirm HPAI H5N1 clade 2.3.4.4 using specific protocols from the NVSL (protocols 1732.02, 1767.01, and 1768.01) (11, 23).
3 Results
Our observation and sampling efforts on Fildes Peninsula totaled 141 h and 30 min, covering 209.5 km of transects (Supplementary Figure S1). The highest efforts were at Diomedea Island (25.4 h), Ardley Island (22.7 h), Biologists Cove (10.8 h), Gemel Peaks (9.2 h), and Gradzinski Cove (7.4 h). We observed various species, including Gentoo, Chinstrap, and Adélie penguins, Southern giant petrels, Antarctic shags, Antarctic terns, Brown and South polar skuas, Kelp gulls, Antarctic fur seals, and non-breeding Southern elephant, Weddell, and Leopard seals. None showed clinical signs of HPAI, and no unexpected mortality was detected. However, skua populations, particularly South polar skuas, declined compared to previous years. One skua carcass was found near Ardley Island, along with a few Gentoo penguin chick carcasses, consistent with typical mortality. In late February, 15 Chinstrap penguin carcasses were observed at Elephant’s Beach, all testing negative for HPAI. In total, 943 samples were collected, 857 environmental fecal samples and 88 direct samples, all testing negative for Influenza A by real-time RT-PCR.
The real-time RT-PCR testing results for skuas from James Ross Island are detailed in Table 1. All deceased skuas tested positive for HPAI H5N1 clade 2.3.4.4, with CT values ranging from 13 to 20.7, indicating high viral loads. There was no evidence of predation or scavenging on the dead animals. Although we lacked permission to histopathological analysis, evident macroscopic lesions were not observed. Additionally, two healthy skuas were noted in proximity to the deceased individuals.
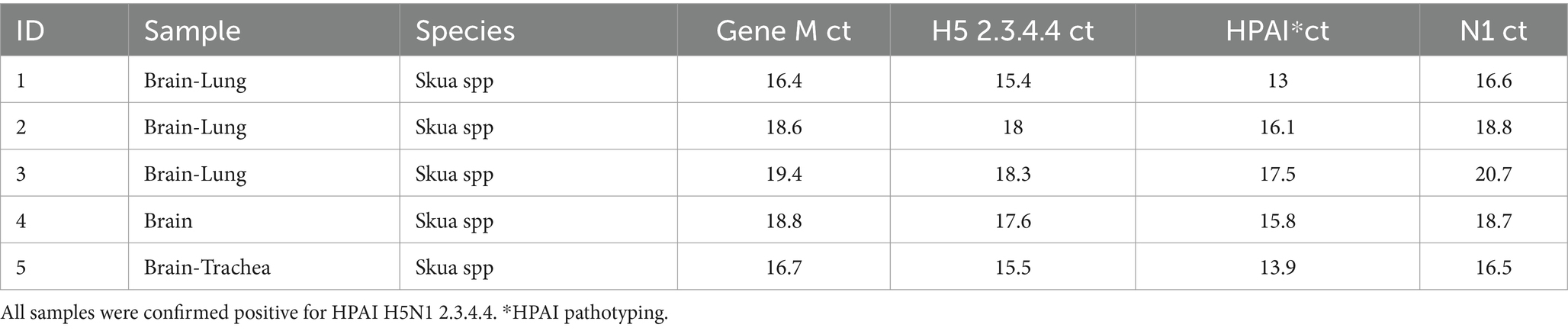
Table 1. Real-time RT-PCR cycle threshold (ct) values from suspected skuas at James Ross Island.
4 Discussion
During the 2022–2023 Southern Hemisphere spring and summer, HPAI outbreaks were confirmed across several South American countries, bringing the virus closer to Antarctica (4). Before this report, HPAI had been detected in sub-Antarctic regions, including South Georgia and the Falkland Islands (17). Then, this study aimed to monitor HPAI in marine mammals and seabirds at key sites like Fildes Peninsula on King George Island, where human activity increases the risk of viral spread (24). Additionally, our network with Antarctic researchers facilitated rapid reporting of unexpected mortality events.
Our surveillance confirmed the presence of HPAI H5N1 in a mortality event involving skuas at James Ross Island. Specific TaqMan PCR assays confirmed the H5 subtype, clade 2.3.4.4, as well as the N1 subtype. Although further genetic analysis and sequencing are necessary, these results provide timely and crucial evidence of the virus’s presence in Antarctica. The reliability of multiple TaqMan PCR reactions enhances confidence in these findings.
The mortality event on James Ross Island was significant, as dead skuas are rarely observed in large numbers. The low CT values from the samples suggest that HPAI H5N1 was the likely cause of death. This finding aligns with recent reports of the virus in skuas within Antarctic territories. On February 25, 2024, just days before the James Ross Island case, two skuas at the Primavera Antarctic Base on Cape Primavera (64°09′00 ″S 60°57′50″W) were confirmed positive for HPAI H5N1, though the report has yet to be published. Additionally, (34) reported suspected cases of HPAI affecting Adélie penguins and Antarctic shags at Red Rock Ridge, though those animals exhibited no clinical signs. These suspected cases remain tentative due to reliance on conventional PCR assays and also raised questions about their accuracy (25).
In contrast, no signs of HPAI were observed in the wildlife of Fildes Peninsula. The extensive sampling and negative RT-PCR results suggest that HPAI was not present at this site during our surveillance period. The decline in the skua population, particularly among South polar skuas, raises questions about the virus’s impact. It is possible that infected birds died during migration, never reaching their breeding grounds in Antarctica. The absence of mass mortality events among other species in the region could indicate a slower viral spread or lower transmission rates in Antarctic ecosystems.
The confirmation of HPAI H5N1 in skuas highlights their potential susceptibility to the virus, which could explain the population decline at Fildes Peninsula. Previous studies have documented the susceptibility of Great Skuas to HPAI H5N1, with a massive mortality event occurring on Foula Island, United Kingdom (26, 27). However, as top predators and scavengers, skuas may have a lower risk of rapid virus dissemination compared to densely populated species like penguins. The study of skua species, including serological analysis, is essential for understanding their role in HPAI dynamics in Antarctica.
Other migratory species, such as Southern giant petrels and Kelp gulls, may also contribute to HPAI dissemination due to their movements between temperate, sub-Antarctic, and Antarctic waters. These species, known for scavenging behaviors, have been implicated in the spread of low-pathogenicity avian influenza viruses (LPAIV) in previous studies (12, 13, 28, 29). While Kelp gulls and Snowy sheathbills have been suggested as potential vectors, tracking studies to confirm their migratory movements are lacking. The role of migratory marine mammals, particularly male Southern elephant seals, in HPAI transmission, also cannot be excluded.
The arrival of HPAI H5N1 in Antarctica presents a significant risk to its wildlife. Outbreaks in South Africa, Chile, and Argentina have demonstrated the high susceptibility of Spheniscus penguins (30, 31). Although no mass mortality events involving penguins have been reported in Antarctica, a recent massive mortality event in Argentina resulted in the deaths of hundreds of Southern elephant seal pups due to HPAI H5N1 (32).
Although sequencing data is pending, our findings are highly reliable, supported by consistent clinical signs and multiple TaqMan PCR detections. This data holds particular value, especially as no other peer-reviewed reports have confirmed the virus in Antarctica to date. The detection of HPAI H5N1 in skuas at James Ross Island, despite the absence of clinical signs in Fildes Peninsula wildlife, indicates that viral transmission in Antarctic species may be slower and less widespread than in other regions. Future studies focus on key species like skuas to fully understand the virus’s evolution and transmission in this fragile ecosystem.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
The animal study was approved by the Institutional Ethics Committee of the University of Chile number 22603-VET-UCH. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
BB-L: Investigation, Methodology, Writing – original draft. BB: Investigation, Writing – original draft. GM: Formal analysis, Investigation, Methodology, Writing – original draft. NA: Conceptualization, Investigation, Methodology, Writing – original draft. NE: Methodology, Writing – original draft. CB: Investigation, Writing – original draft. LK: Conceptualization, Investigation, Methodology, Writing – original draft, Writing – review & editing. MB: Investigation, Methodology, Writing – original draft. MG-A: Conceptualization, Funding acquisition, Investigation, Resources, Writing – original draft. VN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The funders had no role in study design, sample collection, data collection and analysis, decision to publish, or preparation of the article. This work was supported by INACH, Grant N° RT_08_21; ANID Fondecyt Regular N° 1211517 to VN. GM was supported by ANID Programa Beca Doctorado Nacional Grant N° 21220065/2022. LK was supported by the Instituto Antártico Chileno Programa Áreas Marinas Protegidas (AMP 24 03 052) and ANID – Programa Iniciativa Milenio ICN2021_002 (BASE).
Acknowledgments
We thank the INACH personnel for supporting the sample collection and logistics. Thanks also go to the Czech Antarctic Infrastructure (CzechPolar2) at James Ross Island for the support provided during sample collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2024.1423404/full#supplementary-material
References
1. Venkatesan, P. Human cases of avian influenza a(H5) in the USA. The Lancet Microbe. (2024):100978. doi: 10.1016/j.lanmic.2024.100978
2. Zhang, J, Geng, X, Ma, Y, Ruan, S, Xu, S, Liu, L, et al. Fatal avian influenza (H5N1) infection in human, China. Emerg Infect Dis. (2010) 16:1799–801. doi: 10.3201/eid1611.090212
3. Potdar, V, Brijwal, M, Lodha, R, Yadav, P, Jadhav, S, Choudhary, ML, et al. Identification of human case of avian influenza a(H5N1) infection, India. Emerg Infect Dis. (2022) 28:1269–73. doi: 10.3201/eid2806.212246
4. Charostad, J, Rezaei Zadeh Rukerd, M, Mahmoudvand, S, Bashash, D, SMA, H, Nakhaie, M, et al. A comprehensive review of highly pathogenic avian influenza (HPAI) H5N1: an imminent threat at doorstep. Travel Med Infect Dis. (2023) 55:102638. doi: 10.1016/j.tmaid.2023.102638
5. Mills, WF, Ibañez, AE, Carneiro, APB, Morales, LM, Mariano-Jelicich, R, McGill, RAR, et al. Migration strategies of skuas in the Southwest Atlantic Ocean revealed by stable isotopes. Mar Biol. (2023) 171:27. doi: 10.1007/s00227-023-04347-5
6. Caliendo, V, Lewis, NS, Pohlmann, A, Baillie, SR, Banyard, AC, Beer, M, et al. Transatlantic spread of highly pathogenic avian influenza H5N1 by wild birds from Europe to North America in 2021. Sci Rep. (2022) 12:11729. doi: 10.1038/s41598-022-13447-z
7. Prosser, DJ, Chen, J, Ahlstrom, CA, Reeves, AB, Poulson, RL, Sullivan, JD, et al. Maintenance and dissemination of avian-origin influenza a virus within the northern Atlantic flyway of North America. PLoS Pathog. (2022) 18:e1010605. doi: 10.1371/journal.ppat.1010605
8. Gass, JD, Dusek, RJ, Hall, JS, Hallgrimsson, GT, Halldórsson, HP, Vignisson, SR, et al. Global dissemination of influenza a virus is driven by wild bird migration through arctic and subarctic zones. Mol Ecol. (2023) 32:198–213. doi: 10.1111/mec.16738
9. Boulinier, T. Avian influenza spread and seabird movements between colonies. Trends Ecol Evol. (2023) 38:391–5. doi: 10.1016/j.tree.2023.02.002
10. Lane, JV, We, JJ, Avery-gomm, S, Ballstaedt, E, Banyard, AC, Barychka, T, et al. High pathogenicity avian influenza (H5N1) in northern gannets: global spread, clinical signs, and demographic consequences. IBIS Int J Avian Sci. (2024) 166:633–50. doi: 10.1111/ibi.13275
11. Ariyama, N, Pardo-Roa, C, Muñoz, G, Aguayo, C, Ávila, C, Mathieu, C, et al. Highly pathogenic avian influenza a(H5N1) clade 2.3.4.4b virus in wild birds, Chile. Emerg Infect Dis. (2023) 29:1842–5. doi: 10.3201/eid2909.230067
12. Barriga, GP, Boric-Bargetto, D, San Martin, MC, Neira, V, Van Bakel, H, Thompsom, M, et al. Avian influenza virus H5 strain with north American and Eurasian lineage genes in an Antarctic penguin. Emerg Infect Dis. (2016) 22:2221–3. doi: 10.3201/eid2212.161076
13. de Seixas, MMM, de Araújo, J, Krauss, S, Fabrizio, T, Walker, D, Ometto, T, et al. H6N8 avian influenza virus in Antarctic seabirds demonstrates connectivity between South America and Antarctica. Transbound Emerg Dis. (2022) 69:e3436–46. doi: 10.1111/tbed.14728
14. Egevang, C, Stenhouse, IJ, Phillips, RA, Petersen, A, Fox, JW, and Silk, JRD. Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proc Natl Acad Sci USA. (2010) 107:2078–81. doi: 10.1073/pnas.0909493107
15. Kopp, M, Peter, H-U, Mustafa, O, Lisovski, S, Ritz, M, Phillips, R, et al. South polar Skuas from a single breeding population overwinter in different oceans though show similar migration patterns. Mar Ecol Prog Ser. (2011) 435:263–7. doi: 10.3354/meps09229
16. Krietsch, J, Hahn, S, Kopp, M, Phillips, RA, Peter, H-U, and Lisovski, S. Consistent variation in individual migration strategies of brown skuas. Mar Ecol Prog Ser. (2017) 578:213–25. doi: 10.3354/meps11932
17. Banyard, AC, Bennison, A, Byrne, AMP, Reid, SM, Lynton-Jenkins, JG, Mollett, B, et al. Detection and spread of high pathogenicity avian influenza virus H5N1 in the Antarctic region. Nat Commun. (2024) 15:7433. doi: 10.1038/s41467-024-51490-8
18. Dewar, M, Wille, M, Gamble, A, Vanstreels, RET, Bouliner, T, Smith, A, et al. The risk of highly pathogenic avian influenza in the Southern Ocean: a practical guide for operators and scientists interacting with wildlife. Antarct Sci. (2023) 35:407–14. doi: 10.1017/S0954102023000342
19. Pertierra, LR, Hughes, KA, Vega, GC, and Olalla-Tárraga, MÁ. High resolution spatial mapping of human footprint across Antarctica and its implications for the strategic conservation of avifauna. PLoS One. (2017) 12:e0168280. doi: 10.1371/journal.pone.0168280
20. McCarthy, AH, Peck, LS, David, C, and Aldridge, D. Ship traffic connects Antarctica’s fragile coasts to worldwide ecosystems. Proc Natl Acad Sci. (2022) 119:e2110303118. doi: 10.1073/pnas.2110303118
21. Environmental monitoring and management proposals for the Fildes Region, King George Island, Antarctica | Semantic scholar. Available at: https://www.semanticscholar.org/paper/Environmental-monitoring-and-management-proposals-Braun-Mustafa/810f2a556661ed974cd85b541851eeefbdfc3558 (Accessed July 12, 2024)
22. Sumner, MD, Wotherspoon, SJ, and Hindell, MA. Bayesian estimation of animal movement from archival and satellite tags. PLoS One. (2009) 4:e7324. doi: 10.1371/journal.pone.0007324
23. Ulloa, M, Fernández, A, Ariyama, N, Colom-Rivero, A, Rivera, C, Nuñez, P, et al. Mass mortality event in south American Sea lions (Otaria flavescens) correlated to highly pathogenic avian influenza (HPAI) H5N1 outbreak in Chile. Vet Q. (2023):1–13. doi: 10.1080/01652176.2023.2265173
24. Convey, P, and Peck, LS. Antarctic environmental change and biological responses. Sci Adv. (2019) 5:eaaz0888. doi: 10.1126/sciadv.aaz0888
25. Wille, M, Vanstreels, RET, Uhart, M, Reid, S, Dewar, M, and Banyard, AC. (2024): Interpretation and use of in-field diagnostics for high pathogenicity avian influenza (HPAI) in Antarctica– a cautionary tale. doi: 10.20944/preprints202404.1498.v1
26. Furness, RW, Gear, SC, Camphuysen, KCJ, Tyler, G, de Silva, D, Warren, CJ, et al. Environmental samples test negative for avian influenza virus H5N1 four months after mass mortality at a seabird Colony. Pathogens. (2023) 12:584. doi: 10.3390/pathogens12040584
27. Lean, FZX, Falchieri, M, Furman, N, Tyler, G, Robinson, C, Holmes, P, et al. Highly pathogenic avian influenza virus H5N1 infection in skua and gulls in the United Kingdom, 2022. Vet Pathol. (2024) 61:421–31. doi: 10.1177/03009858231217224
28. Petersen, E, Araujo, J, Krüger, L, Seixas, MM, Ometto, T, Thomazelli, L, et al. First detection of avian influenza virus (H4N7) in Giant petrel monitored by geolocators in the Antarctic region. Mar Biol. (2017) 164:62. doi: 10.1007/s00227-017-3086-0
29. Finger, JVG, Krüger, L, Corá, DH, and Petry, MV. Habitat selection of southern giant petrels: potential environmental monitors of the Antarctic peninsula. Antarct Sci. (2023) 35:256–69. doi: 10.1017/S0954102023000147
30. Pardo-Roa, C, Nelson, MI, Ariyama, N, Aguayo, C, Almonacid, LI, Munoz, G, et al. Cross-species transmission and PB2 mammalian adaptations of highly pathogenic avian influenza a/H5N1 viruses in Chile. bioRxiv. (2023). 547205 doi: 10.1101/2023.06.30.547205
31. Molini, U, Aikukutu, G, Roux, J-P, Kemper, J, Ntahonshikira, C, Marruchella, G, et al. Avian influenza H5N8 outbreak in African penguins (Spheniscus demersus), Namibia, 2019. J Wildl Dis. (2020) 56:214. doi: 10.7589/2019-03-067
32. Campagna, C, Uhart, M, Falabella, V, Campagna, J, Zavattieri, V, Vanstreels, RET, et al. Catastrophic mortality of southern elephant seals caused by H5N1 avian influenza. Mar Mamm Sci. (2024) 40:322–5. doi: 10.1111/mms.13101
33. Pavel W, Predator–prey interactions between the South Polar skua Catharacta maccormicki and Antarctic tern Sterna vittata. Catastrophic mortality of southern elephant seals caused by H5N1 avian influenza. J Avian Biol. (2023) 44:89–95. doi: 10.1111/j.1600-048X.2012.05731.x
Keywords: highly pathogenic avian influenza, avian influenza, Antarctica, Antarctic wildlife, skuas, H5N1 2.3.4.4b
Citation: Bennett-Laso B, Berazay B, Muñoz G, Ariyama N, Enciso N, Braun C, Krüger L, Barták M, González-Aravena M and Neira V (2024) Confirmation of highly pathogenic avian influenza H5N1 in skuas, Antarctica 2024. Front. Vet. Sci. 11:1423404. doi: 10.3389/fvets.2024.1423404
Edited by:
Kate Worthing, The University of Sydney, AustraliaReviewed by:
Aruna Pal, West Bengal University of Animal and Fishery Sciences, IndiaBiljana Djurdjević, Scientific Veterinary Institute Novi Sad, Serbia
Copyright © 2024 Bennett-Laso, Berazay, Muñoz, Ariyama, Enciso, Braun, Krüger, Barták, González-Aravena and Neira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Victor Neira, dm5laXJhcmFtQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work