
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Vet. Sci., 20 May 2024
Sec. Comparative and Clinical Medicine
Volume 11 - 2024 | https://doi.org/10.3389/fvets.2024.1395423
This article is part of the Research TopicComparative Coagulation Profiles and Clot Function in Veterinary Medicine: Unravelling Species-Specific Hemostatic Mechanisms and Fibrin Meshwork StructuresView all 3 articles
 Arianna Miglio1*
Arianna Miglio1* Emanuela Falcinelli2Anna Maria Mezzasoma2
Emanuela Falcinelli2Anna Maria Mezzasoma2 Sara Busechian1Fabrizio Rueca1
Sara Busechian1Fabrizio Rueca1 Paolo Gresele2†
Paolo Gresele2† Maria Teresa Antognoni1†
Maria Teresa Antognoni1†Physical exercise has an activating effect on platelet function that differs between trained and untrained subjects, depending on the type of exercise and training status. In humans, soluble P-selectin (sP-sel) and platelet-derived extracellular vesicles (PEVs) are considered reliable markers of in vivo platelet activation during exercise. In untrained humans, they increase after transient physical exercise, whereas long-term training induces a decrease in their resting levels due to an improved ability to adapt to hemodynamic changes. The aim of this study was to assess whether circulating levels of sP-sel and PEVs may be useful markers to explore in vivo platelet function in never-trained Thoroughbreds during their first 4 months of incremental training. A total of 29 clinically healthy, untrained Thoroughbreds (17 males and 12 females) were enrolled. All horses were trained with the same training schedule (90 days). Blood samples were collected on the day the training program began (T0), 30 days (T30), and 90 days (T90) after its incremental increase to quantify platelet count, sP-sel (horse enzyme-linked immunosorbent assay) and PEVs (flow cytometry). Statistical analysis was performed using RM one-way analysis of variance with the Geisser–Greenhouse correction. Soluble P-selectin tended to increase at T30 compared with T0, while T90 levels returned to baseline values. Significantly higher circulating levels of PEVs CD61+/AnnV+ were observed at T30 and T90 compared to baseline confirming platelet hyperactivity. The detection and quantification of sP-sel and PEVs in equine racehorses during the training period appears to be a promising tool to study exercise-induced primary hemostatic changes and may provide an important marker for exercise selection.
Physical exercise has an activating effect on the coagulation system and platelet function in healthy individuals, both in humans and racehorses. However, it is not clear whether this is beneficial or not. Interestingly, acute exercise induces effects that are different from those induced by long-term exercise training, and these effects are different in trained and untrained subjects, demonstrating that the hemostatic response depends on the type of exercise and training status (1–5).
Changes in hemostatic markers induced by long-term training in Thoroughbred horses have been reported (3, 6–10) showing contrasting results of hyper-or hypocoagulable response, probably due to differences in training schedules, breeds of horses, performance status, and methods used for analysis.
Recently, for the first time, we studied the effect of the first period of long-term incremental exercise training on the hemostatic system in untrained Thoroughbreds showing changes in platelet count, primary hemostasis (in vitro evaluation of platelet function assessed by platelet aggregation and PFA100®) and secondary hemostasis (Prothrombin Time, Partial Thromboplastin Time, Thrombin Clotting Time, Fibrinogen, thrombin-antithrombin complex-TAT). In particular, we observed thrombocytosis, an initial hypercoagulable state and platelet hyperreactivity at the initiation of training followed by downregulation of the hemostatic potential at the end of the training period (3). It is worth mentioning that alterations in platelet function appear to be implicated in exercise-induced pulmonary hemorrhage and hemorrhagic syndromes (EIPH) in equine athletes (11, 12). Platelets from horses affected by EIPH tend to be predisposed to thrombus formation and subsequent microvascular obstruction (11) and appear to be significantly less responsive to platelet agonists (12). These few data points suggest that further research on this topic, focusing on the effect of exercise on in vivo platelet function, is highly warranted.
In human studies, soluble P-selectin (sP-sel) and platelet-derived extracellular vesicles (PEVs) have gained increasing importance in recent years as reliable markers of in vivo platelet activation both in health and disease (13, 14).
P-selectin is a cell adhesion molecule that is stored in the alpha granules of platelets and in the Weibel-Palade bodies of endothelial cells and is expressed on their surface upon activation. A soluble form of P-selectin, derived from both platelet and endothelial sources, can be found in plasma as a circulating protein and it is considered not only a marker of platelet activation but also as direct inducer of pro-coagulant activity associated with vascular and thrombotic diseases in humans (15, 16).
PEVs are 0.1–1 μm vesicles shed from the plasma membrane of platelets upon activation with physiological agonists such as thrombin or collagen or in response to high-shear stress (17). In addition to their procoagulant function, several studies suggest a role for PEVs in inflammatory processes (18).
Both of these biomarkers are elevated in humans with cardiac and pulmonary diseases, cardiovascular disorders, sepsis, and diabetes mellitus (1, 19–21), and their dynamic changes and possible function induced by exercise have received much attention. It is well established that in untrained humans they increase after transient physical exercise, driven by vessels’ exposure to high-share stress, whereas long-term training induces a decrease in their resting levels due to an improved ability to adapt to changes in hemodynamics and cellular function during exercise (1, 22, 23).
To date, PEVs have been assessed in horses in a single study of resting animals (24). Information about the measurement of platelet-specific molecules released from platelet granules in equine samples is also limited (25).
The present study was designed to assess whether circulating levels of sP-sel and PEVs may be useful markers to explore in vivo platelet function in never-trained Thoroughbreds evaluating their dynamic changes during the first 4 months of incremental long-term training with the aim of improving knowledge on the fitness and health status of horses and possibly guiding the adjustment of training schedules.
Animals included in the study were those reported in our previous studies (3, 10). A total of 29 clinically healthy (no history of hemostatic abnormalities; heart examination, thorax auscultation, rectal temperature, routine hematology, serum biochemistry, and hemostatic analyses were within reference limits) Thoroughbred racehorses (17 males and 12 females, 2 years of age) were enrolled. No pharmacological treatment was administered before or during the study.
Horses were housed in an AALAC-approved facility in accordance with protocols prescribed by the Institutional Animal Care and Use Committee. All horses were managed with individual housing, a natural photoperiod and natural indoor temperature, and the same feeding schedule. These animals were never trained for flat racing (canter and gallop) prior to entering this study. Each horse followed an individualized feeding program designed by a nutritionist and training team to maximize health conditions, performance, and well-being. Horses were fed three times a day (8:00 a.m., 12:00 noon, and 5:00 p.m.) with hay supplemented with green grass and mixed cereal concentrate. Water was available ad libitum.
All horses were trained according to the same training schedule (Table 1). Training was performed at the same time for each horse from Monday to Saturday with 1 day of rest (Sunday), for a 90-day period. Clinical evaluations and blood samples were collected at rest, before training, on the day of initiation of the training program, which included canter and gallop (T0), 30 days (T30), and 90 days (T90) after the incremental increase in the training program to assess whether or not the animals had adapted to the exercises. Each animal competed in one or more races after the end of the experimental period. None of the horses exhibited poor performance during the study period or during the races.
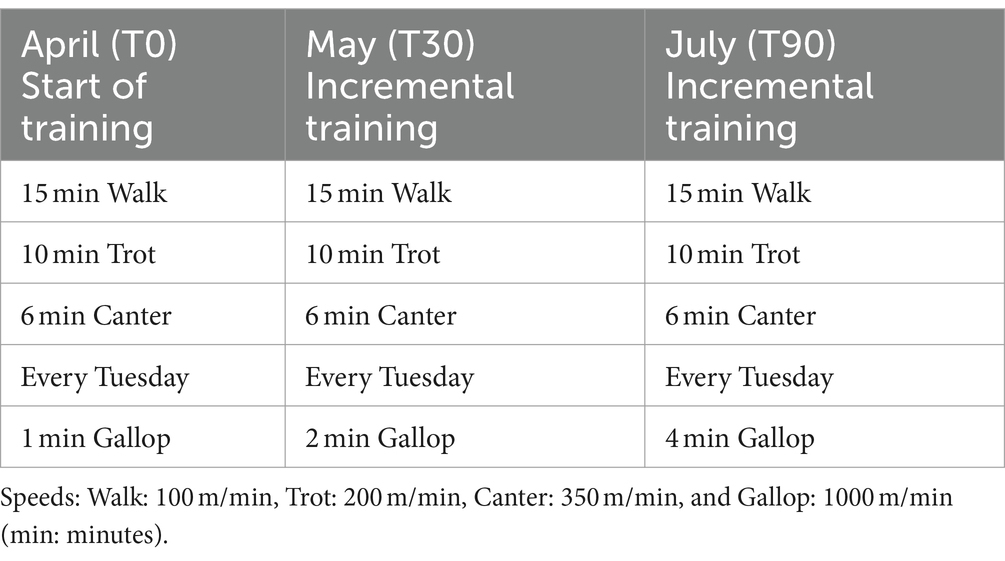
Table 1. Standard daily training program completed by each horse involved in the study.
Blood sampling was performed over a 4-month period from April to July, at 6:30 a.m., at rest, before training and feeding. At T90, the sample collection was done before the race. April (T0) was the sample before the first month of training simulating competition (gallop). From April to July, training was incremental. Blood samples were collected by venipuncture of the jugular vein, paying attention to avoid unnecessary manipulation of the sampling site, which could result in the activation of coagulation. Blood samples were collected in Vacutainer tubes (Terumo Corporation, BD brand; Tokyo, Japan) containing K3-ethylenediamine tetra acetic acid-EDTA and 3.2% sodium citrate. The first few milliliters were discarded to avoid the potential artifacts generated by the contact phase activation. Moreover, to avoid cross-contamination of additives between tubes, blood was drawn and collected in tubes in the following order: first sodium citrate and then K3-EDTA tubes. All the tubes were immediately transported to the local hospital at room temperature to limit cold-induced activation of platelets and were processed within 2 h of collection. K3-EDTA tubes were used to evaluate the platelet counts using an automated hematology analyzer (Sysmex-XT1800iV; Sysmex, Kobe, Japan) validated for horses and equipped with multispecies software. Sodium citrate tubes were centrifuged at 4,000 g for 10 min to obtain platelet-poor plasma (PPP). PPP was then divided into small aliquots in pre-labeled tubes to minimize freeze–thaw cycles and frozen at −80°C until assayed.
For the detection and quantification of PEVs, 1 mL of PPP was further centrifuged for 2 min at 13,000 g and the resulting supernatant was stored at −80°C (24, 26, 27).
Soluble P-Selectin was measured in PPP from citrated blood using a horse enzyme-linked immunosorbent assay (ELISA) (MyBioSource). The sensitivity was 1 ng/mL while the intra- and inter-assay CV (%) was less than 15%.
In total, 50 μL of PFP were incubated with 2.5 μL anti-GPIIb/IIIa antibody (CD61FITC, MyBioSource, clone JM2E5) plus 2 μL AnnexinV-PE (PEVs CD61+/AnnV+) for 30 min at room temperature in the dark (24). The reaction was stopped with 500 μL of calcium-rich binding buffer and a known volume (identical to that of the sample) and the concentration of Flow Count Fluorospheres (Beckman Coulter) was added prior to analysis (26, 27). The morphological gate for extracellular vesicles was set using Megamix (BioCytex, Marseille, France), a mix of fluorescent beads of various diameters covering the size range of microvesicles (0.5 and 0.9 μm) and platelets (0.9 and 3 μm), according to the recommendations of international guidelines (28). After analysis on the flow cytometer, the absolute count was automatically calculated by the instrument software using the Assayed Concentration (in our experiments, 998/μl), using the following formula: (total number of cells counted/total number of flow counts) × flow-count assayed concentration. More details are provided in the Supplementary material.
All results are expressed as mean ± standard error (SEM). Differences between time points were analyzed by the RM one-way analysis of variance with Geisser–Greenhouse correction, and the mixed-effects model was applied and adjusted by Tukey’s multiple comparison test, with individual variances computed for each comparison; initial values were based on GLM. p < 0.05 was considered statistically significant.
No significant effect of training on platelet counts was observed at 30 and 90 days compared with basal (T0). The mean values were within the reference ranges (Supplementary Table S1).
Soluble P-selectin increased after 30 days (T30) of the training program compared to baseline (T0), whereas after 90 days (T90) of the training program, levels returned to baseline (Figure 1A; Supplementary Table S1).

Figure 1. Levels of sP-sel (A) and PEVs CD61+/AnnV+ (B) in plasma collected on the day of the beginning of the training program, which included canter and gallop (T0) and 30 days (T30) and 90 days (T90) after the incremental increase of the training program. *p = 0.0143; **p = 0.0020; ***p = 0.0006; # = 0.0011.
Significantly higher circulating levels of PEVs CD61+/AnnV+ were observed at T30 compared to baseline confirming platelet hyperreactivity. This increase was maintained also after 90 days of the training program compared to the baseline (Figure 1B; Supplementary Table S1).
The present study reports for the first time the effect of long-term training on circulating levels of in vivo platelet activation markers, sP-sel, and PEVs CD61+/AnnV+, in healthy, untrained racehorses. Our results show that sP-sel and PEVs CD61+/AnnV+ levels increased 1 month after the start of training, although at the border of significance for sP-Sel, confirming a condition of platelet hyperreactivity and in vivo platelet activation demonstrating the exercise-induced changes in hemostasis as supported by previous results in humans (29).
It is well known that exercise intensity, duration, type, and individual training status are important factors involved in exercise-induced changes in platelet in vivo and platelet reactivity function; in fact, in vivo, platelet activation appears to be suppressed in the course of moderate exercise, whereas it is increased during strenuous exercise (1, 30, 31).
P-selectin is an adhesion receptor molecule found in platelet α-granules and in the Weibel-Palade bodies of endothelial cells. A soluble form of P-selectin can be found in plasma as a circulating protein that is shed from endothelial cells and the platelet surface membrane after activation, although platelets are currently considered to be the major source of circulating soluble P-selectin in healthy individuals (32). Its biological role is unknown, but its levels are commonly used as a reliable marker of in vivo platelet activation, capable of predicting a pro-thrombotic state in both humans and various animal species (15, 33). It appears to be an agonist of the coagulation process mediating platelet–platelet and platelet-leukocyte interactions and favoring irreversible platelet aggregation upon thrombin stimulation (34).
A significant increase in sP-sel has been found following strenuous exercise in humans, although its release mechanism has not been well characterized (2).
PEVs are a heterogeneous population of small circular platelet membrane fragments generated during platelet activation by various stimuli, senescence, and apoptosis. They retain membrane antigens and cytoplasmic components of their cellular origin and play an essential role in coagulation. Elevated concentrations of circulating PEVs have been reported in thrombotic conditions in humans and are considered an independent predictor of poor outcomes.
Moreover, in humans, the level of PEVs offers insight into the state of circulating platelets during exercise and is considered a useful marker for monitoring physical training activities. The level of PEVs appears to be related to training intensity, with little change during moderate exercise and a high increase during high-intensity exercise (29, 35). Blood levels of EVs change immediately and remain high for up to 3 h after a single bout of exercise (1).
Interestingly, in our study, circulating EV levels increased significantly 1 month after the beginning of training and remained high until the end of the training period while sP-sel increased after 30 days of training and tended to return to basal levels at the end of the training period.
The circulating levels of soluble adhesion molecules are likely to be determined not only by their synthesis and shedding but also by their clearance. Little is known about the clearance of soluble selectins such as P-selectin and we can assume that incremental training could increase the clearance of these soluble molecules. Similarly, our finding that PEV levels are persistently elevated after long-term training may reflect that, although the detailed clearance mechanism of EVs in circulation is not clear, the release and clearance of EVs may coexist during exercise.
Flow cytometry is considered the gold standard method for the study of EVs, allowing the accurate identification and enumeration of EV subpopulations based on the use of distinct monoclonal antibodies that detect cell-specific surface proteins in both humans and horses (1, 36, 37). In this study, we applied a standardized flow cytometry protocol for the identification and enumeration of EVs in equine plasma using an anti-CD61 antibody, as previously reported (24). Moreover, we adapted our protocol from those standardized for humans, which are able to minimize preanalytical influences on PEV counts (38) with the advantage of good accuracy and ease of application.
Platelets are affected by a multitude of physical and chemical factors including sampling collection, handling, processing, and artificial in vitro activation, which may limit the reliability of results. In our study, precautions were taken to minimize ex vivo and in vitro platelet activation according to international guidelines (28).
The present results confirm our previous observations showing an initial hypercoagulability state and increased in vitro platelet reactivity induced by incremental training in thoroughbred horses, which reached maximum levels at 30 days and then tended to return to baseline at the end of the exercise period (3, 10).
The initial hypercoagulable state places racehorses at risk for thrombosis with its clinical sequelae of microthrombotic events, ischemic tissue injury, and organ dysfunction (12, 39–42). Since platelets play a crucial role in the process of thrombus formation and have a pro-inflammatory effect, this vicious cycle between inflammation and coagulation may contribute to the pathogenesis of different common pathologies affecting sport horses such as laminitis, acute renal failure, and EIPH. For this reason, early identification of subclinical alterations in hemostasis in training horses may be of help in the management of equine athletes (11, 43).
This study had some limitations, such as the limited number of animals and the lack of a control group that did not enter training; however, we sampled a group of horses that had never been trained before to have control over untrained subjects. Also, to reduce environmental effects, we waited 30 days before housing the animals in the same training center, which standardized management and diet. Moreover, the lack of a CD62P marker for the identification of MVs does not allow us to discriminate between circulating microparticles derived from platelets following activation and those continuously derived from megakaryocytes although we used a commonly used double staining approach (CD61+/AnnV+) for the identification of MVs (38).
Larger prospective studies testing the effect of regular exercise training on platelet function in healthy horses during training are warranted.
The detection and quantification of soluble P-selectin and PEVs in equine racehorses during the training period appears to be a promising tool to study exercise-induced changes in primary hemostasis and further studies are warranted to better establish reference ranges. The changes in sP-sel and MVs in healthy Thoroughbreds during their first long-term training may provide an important surrogate marker for exercise selection in racehorses. Future studies with larger numbers of racehorses are needed to reassess the possible relationship between in vivo platelet activation, intensity of training, and training status.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethical approval was not required for the studies involving animals in accordance with the local legislation and institutional requirements because The study was performed on material collected during clinical procedures. Blood sampling was approved by the Italian Horse Racing Board and performed by the authorized veterinary practitioner during routine controls to assess the health of the animals in the course of the training season. Prior to sample collection, written owner consent was obtained for all animals included in the study. Animal care procedures were compliant with the European recommendations (Directive 2010/63/EU) for protecting animals used for scientific purposes. Written informed consent was obtained from the owners for the participation of their animals in this study.
AMi: Writing – review & editing, Writing – original draft, Visualization, Validation, Supervision, Software, Resources, Project administration, Methodology, Investigation, Funding acquisition, Formal Analysis, Data curation, Conceptualization. EF: Writing – original draft, Visualization, Validation, Software, Methodology, Formal Analysis, Data curation, Conceptualization. AMe: Writing – review & editing, Visualization, Validation, Formal Analysis, Data curation. SB: Writing – review & editing, Validation. FR: Writing – review & editing, Validation, Funding acquisition. PG: Writing – review & editing, Validation, Supervision, Methodology, Funding acquisition, Conceptualization. MA: Writing – review & editing, Visualization.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2024.1395423/full#supplementary-material
1. Han, X, Li, T, Li, Y, Yang, J, Chen, S, Zhu, X, et al. Exercise and circulating microparticles in healthy subjects. J Cardiovasc Transl Res. (2021) 14:841–56. doi: 10.1007/s12265-021-10100-4
2. Kristiansen, J, Grove, EL, Pedersen, OB, Kristensen, SD, and Hvas, A-M. Effect of regular exercise training on platelet function in patients with cardiovascular disease and healthy individuals: a systematic review. Thromb Res. (2022) 231:223–35. doi: 10.1016/j.thromres.2022.12.017
3. Miglio, A, Falcinelli, E, Mezzasoma, AM, Cappelli, K, Mecocci, S, Gresele, P, et al. Effect of first long-term training on whole blood count and blood clotting parameters in thoroughbreds. Animals (Basel). (2021) 11:447. doi: 10.3390/ani11020447
4. Piccione, G, Grasso, F, Fazio, F, and Giudice, E. The effect of physical exercise on the daily rhythm of platelet aggregation and body temperature in horses. Vet J. (2008) 176:216–20. doi: 10.1016/j.tvjl.2007.01.026
5. Wang, J-S, Li, Y-S, Chen, J-C, and Chen, Y-W. Effects of exercise training and deconditioning on platelet aggregation induced by alternating shear stress in men. Arterioscler Thromb Vasc Biol. (2005) 25:454–60. doi: 10.1161/01.ATV.0000151987.04607.24
6. Assenza, A, Tosto, F, Casella, S, Fazio, F, Giannetto, C, and Piccione, G. Changes in blood coagulation induced by exercise training in young athletic horses. Res Vet Sci. (2013) 95:1151–4. doi: 10.1016/j.rvsc.2013.07.005
7. Fazio, F, Assenza, A, Tosto, F, Casella, S, Piccione, G, and Caola, G. Modifications of some acute phase proteins and the white blood cell count in thoroughbreds during training. Vet Rec. (2010) 167:370–2. doi: 10.1136/vr.c3761
8. Giannetto, C, Arfuso, F, Fazio, F, Giudice, E, Pietro, SD, Bruschetta, D, et al. Different training schedules influence platelet aggregation in show jumping horses. Pol J Vet Sci. (2017) 20:149–54. doi: 10.1515/pjvs-2017-0019
9. Piccione, G, Bazzano, M, Giannetto, C, Marafioti, S, and Fazio, F. Training-induced changes in clotting parameters of athletic horses. J Vet Sci. (2014) 15:45–9. doi: 10.4142/jvs.2014.15.1.45
10. Miglio, A, Falcinelli, E, Cappelli, K, Mecocci, S, Mezzasoma, AM, Antognoni, MT, et al. Effect of regular training on platelet function in untrained thoroughbreds. Animals (Basel). (2024) 14:414. doi: 10.3390/ani14030414
11. Weiss, DJ, McClay, CB, Smith, CM, Rao, GHR, and White, JG. Platelet function in the racing thoroughbred: implication for exercise-induced pulmonary hemorrhage. Vet Clin Pathol. (1990) 19:35–9. doi: 10.1111/j.1939-165X.1990.tb00540.x
12. Johnstone, IB, Viel, L, Crane, S, and Whiting, T. Hemostatic studies in racing standardbred horses with exercise-induced pulmonary hemorrhage. Hemostatic parameters at rest and after moderate exercise. Can J Vet Res. (1991) 55:101–6.
13. Gresele, P, Migliacci, R, Vedovati, MC, Ruffatti, A, Becattini, C, Facco, M, et al. Patients with primary antiphospholipid antibody syndrome and without associated vascular risk factors present a normal endothelial function. Thromb Res. (2009) 123:444–51. doi: 10.1016/j.thromres.2008.05.015
14. Petito, E, Colonna, E, Falcinelli, E, Mezzasoma, AM, Cesari, E, Giglio, E, et al. Anti-severe acute respiratory syndrome coronavirus-2 adenoviral-vector vaccines trigger subclinical antiplatelet autoimmunity and increase of soluble platelet activation markers. Br J Haematol. (2022) 198:257–66. doi: 10.1111/bjh.18245
15. André, P . P-selectin in haemostasis. Br J Haematol. (2004) 126:298–306. doi: 10.1111/j.1365-2141.2004.05032.x
16. Ridker, PM, Buring, JE, and Rifai, N. Soluble P-selectin and the risk of future cardiovascular events. Circulation. (2001) 103:491–5. doi: 10.1161/01.cir.103.4.491
17. Melki, I, Tessandier, N, Zufferey, A, and Boilard, E. Platelet microvesicles in health and disease. Platelets. (2017) 28:214–21. doi: 10.1080/09537104.2016.1265924
18. Provost, P . The clinical significance of platelet microparticle-associated microRNAs. Clin Chem Lab Med. (2017) 55:657–66. doi: 10.1515/cclm-2016-0895
19. Fijnheer, R, Frijns, CJ, Korteweg, J, Rommes, H, Peters, JH, Sixma, JJ, et al. The origin of P-selectin as a circulating plasma protein. Thromb Haemost. (1997) 77:1081–5. doi: 10.1055/s-0038-1656116
20. Semenov, AV, Kogan-Ponomarev, MI, Ruda, MI, Komarov, AL, Panchenko, EP, Chazova, IE, et al. Soluble P-selectin - a marker of platelet activation and vessel wall injury: increase of soluble P-selectin in plasma of patients with myocardial infarction, massive atherosclerosis and primary pulmonary hypertension. Ter Arkh. (2000) 72:15–20.
21. Siljander, PRM . Platelet-derived microparticles - an updated perspective. Thromb Res. (2011) 127:S30–3. doi: 10.1016/S0049-3848(10)70152-3
22. Chaar, V, Romana, M, Tripette, J, Broquere, C, Huisse, M-G, Hue, O, et al. Effect of strenuous physical exercise on circulating cell-derived microparticles. Clin Hemorheol Microcirc. (2011) 47:15–25. doi: 10.3233/CH-2010-1361
23. Maruyama, K, Kadono, T, and Morishita, E. Plasma levels of platelet-derived microparticles are increased after anaerobic exercise in healthy subjects. J Atheroscler Thromb. (2012) 19:585–7. doi: 10.5551/jat.11791
24. Springer, NL, Smith, E, Brooks, MB, and Stokol, T. Flow cytometric detection of circulating platelet-derived microparticles in healthy adult horses. Am J Vet Res. (2014) 75:879–85. doi: 10.2460/ajvr.75.10.879
25. López, C, Carmona, JU, Giraldo, CE, and Alvarez, ME. Bacteriostatic effect of equine pure platelet-rich plasma and other blood products against methicillin-sensitive Staphylococcus aureus. An in vitro study. Vet Comp Orthop Traumatol. (2014) 27:372–8. doi: 10.3415/VCOT-14-04-0054
26. Petito, E, Franco, L, Falcinelli, E, Guglielmini, G, Conti, C, Vaudo, G, et al. COVID-19 infection-associated platelet and neutrophil activation is blunted by previous anti-SARS-CoV-2 vaccination. Br J Haematol. (2023) 201:851–6. doi: 10.1111/bjh.18726
27. Petito, E, Falcinelli, E, Paliani, U, Cesari, E, Vaudo, G, Sebastiano, M, et al. Neutrophil more than platelet activation associates with thrombotic complications in COVID-19 patients. J Infect Dis. (2020) 223:933–44. doi: 10.1093/infdis/jiaa756
28. Lacroix, R, Judicone, C, Poncelet, P, Robert, S, Arnaud, L, Sampol, J, et al. Impact of pre-analytical parameters on the measurement of circulating microparticles: towards standardization of protocol. J Thromb Haemost. (2012) 10:437–46. doi: 10.1111/j.1538-7836.2011.04610.x
29. Sossdorf, M, Otto, GP, Claus, RA, Gabriel, HHW, and Lösche, W. Cell-derived microparticles promote coagulation after moderate exercise. Med Sci Sports Exerc. (2011) 43:1169–76. doi: 10.1249/MSS.0b013e3182068645
30. El-Sayed, MS, Ali, N, and El-Sayed, AZ. Aggregation and activation of blood platelets in exercise and training. Sports Med. (2005) 35:11–22. doi: 10.2165/00007256-200535010-00002
31. Kestin, AS, Ellis, PA, Barnard, MR, Errichetti, A, Rosner, BA, and Michelson, AD. Effect of strenuous exercise on platelet activation state and reactivity. Circulation. (1993) 88:1502–11. doi: 10.1161/01.cir.88.4.1502
32. Ferroni, P, Martini, F, Riondino, S, La Farina, F, Magnapera, A, Ciatti, F, et al. Soluble P-selectin as a marker of in vivo platelet activation. Clin Chim Acta. (2009) 399:88–91. doi: 10.1016/j.cca.2008.09.018
33. Kumar, A, Villani, MP, Patel, UK, Keith, JC, and Schaub, RG. Recombinant soluble form of PSGL-1 accelerates thrombolysis and prevents reocclusion in a porcine model. Circulation. (1999) 99:1363–9. doi: 10.1161/01.cir.99.10.1363
34. André, P, Hartwell, D, Hrachovinová, I, Saffaripour, S, and Wagner, DD. Pro-coagulant state resulting from high levels of soluble P-selectin in blood. Proc Natl Acad Sci USA. (2000) 97:13835–40. doi: 10.1073/pnas.250475997
35. Wilhelm, EN, Mourot, L, and Rakobowchuk, M. Exercise-derived microvesicles: a review of the literature. Sports Med. (2018) 48:2025–39. doi: 10.1007/s40279-018-0943-z
36. Theuerkauf, K, Obach-Schröck, C, Staszyk, C, Moritz, A, and Roscher, KA. Activated platelets and platelet-leukocyte aggregates in the equine systemic inflammatory response syndrome. J Vet Diagn Invest. (2022) 34:448. doi: 10.1177/10406387221077969
37. Weiss, DJ, and Evanson, OA. Detection of activated platelets and platelet-leukocyte aggregates in horses. Am J Vet Res. (1997) 58:823–7. doi: 10.2460/ajvr.1997.58.08.823
38. Cointe, S, Judicone, C, Robert, S, Mooberry, MJ, Poncelet, P, Wauben, M, et al. Standardization of microparticle enumeration across different flow cytometry platforms: results of a multicenter collaborative workshop. J Thromb Haemost. (2017) 15:187–93. doi: 10.1111/jth.13514
39. Lo Feudo, CM, Stucchi, L, Conturba, B, Stancari, G, Zucca, E, and Ferrucci, F. Medical causes of poor performance and their associations with fitness in Standardbred racehorses. J Vet Int Med. (2023) 37:1514–27. doi: 10.1111/jvim.16734
40. Morley, PS, Bromberek, JL, Saulez, MN, Hinchcliff, KW, and Guthrie, AJ. Exercise-induced pulmonary haemorrhage impairs racing performance in thoroughbred racehorses. Equine Vet J. (2015) 47:358–65. doi: 10.1111/evj.12368
41. Poole, DC, and Erickson, HH. Exercise-induced pulmonary hemorrhage: where are we now? Vet Med (Auckl). (2016) 7:133–48. doi: 10.2147/VMRR.S120421
42. Sullivan, S, and Hinchcliff, K. Update on exercise-induced pulmonary hemorrhage. Vet Clin N Am Equine Pract. (2015) 31:187–98. doi: 10.1016/j.cveq.2014.11.011
Keywords: soluble P-selectin, platelet-derived extracellular vesicles, equine, exercise, platelet
Citation: Miglio A, Falcinelli E, Mezzasoma AM, Busechian S, Rueca F, Gresele P and Antognoni MT (2024) Biomarkers of in vivo platelet activation in thoroughbreds during their first long-term training. Front. Vet. Sci. 11:1395423. doi: 10.3389/fvets.2024.1395423
Edited by:
Nanette Oberholzer, University of Pretoria, South AfricaReviewed by:
Magdalena Żmigrodzka, Warsaw University of Life Sciences, PolandCopyright © 2024 Miglio, Falcinelli, Mezzasoma, Busechian, Rueca, Gresele and Antognoni. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Arianna Miglio, YXJpYW5uYS5taWdsaW9AdW5pcGcuaXQ=
†These authors have contributed equally to this work and share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.