 Annalisa Viani1*
Annalisa Viani1* Tommaso Orusa2
Tommaso Orusa2 Sara Divari3
Sara Divari3 Stella Lovisolo3
Stella Lovisolo3 Stefania Zanet3
Stefania Zanet3 Riccardo Orusa4
Riccardo Orusa4 Enrico Borgogno-Mondino2
Enrico Borgogno-Mondino2 Enrico Bollo3
Enrico Bollo3- 1Azienda USL della Valle d'Aosta, S.C. Sanità Animale, Quart, Italy
- 2GEO4Agri DISAFA Laboratory, Department of Agricultural, Forest and Food Sciences (DISAFA), University of Turin, Grugliasco, Italy
- 3Department of Veterinary Sciences, University of Turin, Grugliasco, Italy
- 4Experimental Zooprophilactic Institute of Piedmont, Liguria and Aosta Valley (IZS PLV) S.C Valle d'Aosta—CeRMAS (National Reference Center for Wildlife Diseases), Quart, Italy
Bartonella is a genus of bacteria known to cause various rare but potentially dangerous diseases in humans and wildlife. The objective of this study was to investigate the presence of Bartonella spp. in red foxes (Vulpes vulpes) from Piedmont and Aosta Valley (NW Italy) and to explore the potential association between environmental humidity and Bartonella infection using remote sensing data. A total of 114 spleen samples were collected from hunted red foxes and screened for Bartonella spp. DNA using a qPCR assay targeting the ssrA locus. Samples that tested positive were further analyzed using end-point PCR to detect the ssrA, gltA, and rpoB loci. The overall prevalence of Bartonella spp. infection was found to be 7.9% (9/114), with a prevalence of 6.9% (7/101) in foxes from Piedmont and 15.4% (2/13) in foxes from Aosta Valley. Sequencing results identified Bartonella schoenbuchensis R1 as the most commonly isolated species (5/9, 62.5%), followed by Candidatus “Bartonella gerbillinarum” in two samples (2/9, 28.6%). To investigate the relationship between environmental factors and Bartonella infection, data from NASA USGS Landsat missions (TOA collection) from 2011 to 2022 were processed using Google Earth Engine. The Tasseled Cap Wetness Index (TCW), an indicator of landscape moisture, was calculated for each meteorological season. The study found that Bartonella spp. infections in foxes were positively associated with higher TCW values (>0.7). Canonical Correspondence Analysis further demonstrated a strong link between pathogen prevalence and municipal-level TCW, suggesting that TCW could be used as a parameter to facilitate disease management and control. This study provides a starting point for a more comprehensive epidemiological assessment of Bartonella spp. on a national scale and highlights the potential role of environmental humidity in influencing pathogen distribution.
1 Introduction
The widespread distribution of the fox (Vulpes vulpes Linnaeus) on the Italian territory has resulted in the conduct of multiple studies on the diseases of this species. Over time, some of these diseases have attracted great interest from the scientific world (1, 2), while less attention has been paid to others, such as bartollenosis. Few studies on the presence and spread of this infection in foxes have been published worldwide (3–5); moreover, among the wide range of hosts of this parasite, the epidemiology of bartonellosis in foxes is one of the least investigated. It is well-known that the knowledge and control of animal diseases contributes significantly to the protection of public health, and the health of free-ranging wildlife can act as sensitive sentinel or bio-indicator for broader health issues (6).
Bartonella infection in foxes has been described in different part of the world (3–5, 7–13), and it has been reported in wolves and foxes in Spain and other European countries such as Italy (14), Lithuania (15), and Slovakia (16). Since both the wolf and the red fox can come in contact with domesticated animals, especially dogs, they may play a crucial role in the ecology and spread of Bartonella (17). In Italy, Bartonella bovis and Bartonella chomelii are the only species reported in wildlife, mostly in deer ticks (18). In particular, there is a lack of data on infection in wild canids, although Bartonella vinsonii subsp. berkhoffii and Candidatus Bartonella merieuxii have been reported in hunting dogs in southern Italy (19). It is worth to note that B. vinsonii subsp. berkhofii genotype III can be highly pathogenic to dogs (20).
Bartonella spp. is widespread worldwide, but in some areas the prevalence is higher in relation to the climatic conditions that favor the spread of arthropod vectors (11). Moreover, the distribution of Bartonella is partly linked to the variety of blood-sucking vectors (12). Transmission occurs indirectly, by means of arthropod vectors, mainly fleas, lice, ticks, and blood-sucking flies, which ingest erythrocytes containing bacterial cells through a blood meal and then transmit them to a reservoir or to an accidental host (13). In the study conducted by de Paiva Diniz et al. (68) in 2018, Bartonella strains were identified from environmental and arthropod samples. Furthermore, investigations on the gut microbiome of a variety of insects have revealed the presence of the microorganism among arthropods, i.e., cockroaches, butterflies, bees, various ant species, and a wide variety of ectoparasitic species (15, 16). Other researchers have hypothesized that Bartonella may also have a commensal role in arthropods (17), and therefore how this microorganism originated as an environmental bacterium and evolved and diversified through contact with arthropod vectors. Bartonella represents an ideal pathogen to focus on to evaluate the impact of anthropogenic activity on the distribution models of the main infectious diseases in animals, within the same species and between different species.
The recent exponential development of science and technologies applied to GIS has contributed to the enhancement of epidemiological data analysis capabilities and has provided new and powerful tools for animal disease surveillance (11). GIS, spatial analysis techniques and the use of satellites provide useful methods for collecting and managing the information necessary for epidemiological studies. Still little explored in the veterinary field is the use and development of applications and methodologies based on “Earth Observation Data”. Within the European space program “Copernicus” and other historical programs such as the NASA Landsat missions, geospatial data with medium-high geometric and temporal resolution allow to exploit and expand risk analysis techniques, translating them into a transfer technology for veterinary studies. To date, in fact, only a few applications have been explored with the use of medium-low resolution data in the context of the NASA “Terra” and “Aqua” missions with the “Modis” sensors (21). Although promising, it should be considered that for eco-epidemiological modeling studies, especially in the Alpine area, these data have strong limitations (22–25). Therefore, to avoid bias related to image processing or to a lack of knowledge of passive satellite remote sensing sensors, it is necessary to have high skills in the field of geomatics and remote sensing (26–28).
The aim of this study was to investigate the prevalence and distribution of Bartonella spp., a genus of bacteria responsible for rare but serious diseases, within red fox populations in the provinces of Cuneo and Biella (Piedmont) and Aosta Valley in northwestern Italy. Specifically, the research sought to understand the geographical and environmental factors that might influence the presence of Bartonella spp. at the municipal level. To achieve this, the study employed a combination of molecular diagnostic techniques to detect Bartonella DNA in fox spleen samples and advanced satellite remote sensing to analyze environmental conditions. By integrating these approaches, the research aimed to explore possible correlations between the prevalence of Bartonella infections and environmental factors, particularly focusing on landscape moisture as indicated by the Tasseled Cap Wetness Index (TCW). The ultimate objective was to determine whether specific environmental conditions, such as higher humidity levels, could be associated with increased Bartonella infection rates in fox populations. This interdisciplinary approach aimed to provide a more comprehensive understanding of the epidemiology of Bartonella spp. in wildlife and contribute valuable insights for disease management and control efforts on a broader scale. The study period involved data from 2011 to 2022, during which satellite imagery was processed to create composite TCW images aggregated for each meteorological season of every year.
2 Materials
The foxes examined in this study belong to the species V. vulpes. A total of 114 spleen samples were collected from foxes either hunted during the hunting seasons or found dead across different locations in the Piedmont districts of the provinces of Cuneo and Biella, as well as the Aosta Valley in northwestern Italy. These samples were collected over a period from 2011 to 2022, with the majority coming from Biella (57 samples), followed by Cuneo (44 samples), and Aosta Valley (13 samples). The samples were stored at −20°C until DNA extraction. Detailed information on the location, sex (59 males and 55 females), age (87 adults, 13 subadults, 14 juveniles), and season of collection (88 in autumn-winter, 26 in spring-summer) is summarized in Supplementary Tables S1a–c.
2.1 Data collection
2.1.1 Lab data
For the detection of Bartonella spp. DNA, ~30 mg of spleen tissue was thawed and subjected to DNA extraction using the “E.Z.N.A. Tissue DNA Kit” (Omega Bio-Tek, Norcross, USA), following the manufacturer's instructions. The CFX Connect Real-Time System (BioRad, Hercules, USA) was used to perform qPCR targeting the ssrA locus, which codes for transfer-messenger RNA. Samples with a Cq value lower than 32 were considered positive for Bartonella spp. Positive samples underwent further analysis using end-point PCR to detect the gltA, rpoB, and ssrA loci. Detailed protocols for each PCR assay are provided, including the conditions for amplification and subsequent sequencing of the PCR products. The end-point PCR products were separated, purified, and sent for bidirectional DNA Sanger sequencing.
2.1.2 GIS data
The study utilized multispectral optical remote sensing data from the NASA USGS Landsat 5 and Landsat 8 missions. The data was processed using the cloud-based platform Google Earth Engine (GEE), which allows for large-scale processing of satellite images. The study specifically focused on the Tasseled Cap Wetness (TCW) index, a multispectral index related to soil and canopy moisture interactions, computed from Landsat data. Composite images were created for each meteorological season from 2011 to 2022, aggregating data to capture seasonal variations in environmental conditions. The TCW index was calculated for each scene to assess the relationship between landscape moisture and the prevalence of Bartonella spp. in foxes. The Moran's I index was used to test for spatial autocorrelation.
3 Methods
The approach reported below in each sub-chapter was based on the following materials.
3.1 DNA extraction and qPCR
The CFX Connect Real-Time System (BioRad, Hercules, USA) was used for qPCR and the locus taken into consideration was ssrA codifying for transfer-messenger RNA (29). The protocol includes an initial denaturation at 95°C for 3 min, followed by 45 cycles of denaturation (at 95°C for 15 s), hybridization and extension (at 60°C for 1 min). qPCR was performed using IQ Multiplex Powermix (BioRad, Hercules, USA).
The genomic DNA of Bartonella spp. FG4-1 was used as a positive control (30). Samples with a Cq value lower than 32 were considered positive and were subjected to subsequent analysis.
Primers and probe related to the loci considered in this study are reported in Supplementary Table S2.
3.2 PCR end-point of gltA, rpoB and ssrA loci
The citrate synthase (gltA), RNA polymerase (rpoB), and ssrA loci were detected in qPCR positive samples using end-point PCR. For the gltA locus, two PCR end-point assays were applied, generating 380 bp (31) and 700 bp (32) amplicons. Amplification was performed using a T100 Thermal Cycler (BioRad, Hercules, USA). The protocols applied in the end-point PCR were the following:
- gltA (380 bp): an initial denaturation at 95°C for 15 min followed by 35 cycles at 95°C for 20 s, at 51°C for 30 s and at 72°C for 2 min;
- gltA (700 bp): an initial denaturation at 95°C for 15 min followed by 40 cycles at 95°C for 30 s, at 48°C for 1 min, at 72°C for 1 min; finally, an extension at 72°C for 7 min;
- rpoB: an initial denaturation at 95°C for 15 min followed by 35 cycles at 95°C for 30 s, at 53°C 78 for 30 s, at 72°C for 1 min; finally, an extension at 72°C for 2 min;
- ssrA: an initial denaturation at 95°C for 30 s followed by 40 cycles at 95°C for 15 s, at 60°C for 1 min, at 72°C for 30 s; finally, an extension at 72°C for 3 min.
Reactions were performed using HotStartTaq matrer mix (Qiagen) using primers described in Supplementary Table S2.
End-point PCR products were separated on 1.5% agarose gel in Tris-Acetate-EDTA (TAE) buffer using MIDORI Green Advance DNA stain (Nippon Genetics, Düren, Germany).
3.3 Sequencing
The amplicons obtained were purified using MinElute PCR Purification Kit (Qiagen, Hilden, Germany) and they were sent to BMR Genomics (Padua, Italy) for bidirectional sequencing (DNA Sanger sequencing) (BMR Genomics, Padua, Italy). Sequences obtained were manually corrected using the Geneious Prime 2020.1. The corrected sequences were aligned with the sequences deposited in GenBank, by BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi, last access on 19 April 2023).
3.4 Statistic analysis
The results obtained were analyzed using the GraphPad Prism 8.4.2 software (GraphPad Software, California, USA). Fisher's test was used to evaluate the association between the presence of Bartonella spp. and gender, age, season, and period of collection of samples. The Chi square test was used to evaluate the association between the presence of Bartonella spp. and the place of recovery of animals. A 95% confidence interval was set, and results with p-value < 0.05 were considered statistically significant.
3.5 GIS and remote sensing
The GIS analyzes were carried out using the software QGIS version 3.16.4 Hannover (23). For the geospatial analysis, we used multispectral optical remote sensing data from the NASA USGS Landsat missions with 16-day time resolution starting from 1972 for the whole globe and spatial resolution as a function of the considered spectral band of 30 m for the most part of the spectral bands used. For this work, data from the USGS Landsat 5 and USGS Landsat 8 missions were used, respectively from the Multispectral Scanner (MSS) and Operational Land Imager (OLI) sensors. In particular, the cloud-based platform Google Earth Engine (hereinafter called as GEE) was adopted for the processing of remote sensing data, which allows large-scale processing of satellite images to detect changes, map trends and quantify differences on the Earth's surface (33–40) by accessing petabytes of various satellite missions from different global space agencies.
In particular, the following GEE collections were used:
- LANDSAT/LT05/C02/T1_TOA: Landsat 5 MSS TM with reflectance calibrated at the level of the upper part of the atmosphere (TOA). Calibration coefficients are extracted from each scene's metadata for TOA calculation based on (41).
- LANDSAT/LC08/C02/T1_TOA: Landsat 8 OLI TIRS with reflectance calibrated at the level of the upper part of the atmosphere (TOA). Calibration coefficients are extracted from each scene's metadata for TOA calculation based on (41, 42).
Starting from the sample collection data of the entire population of foxes considered, aggregated by meteorological seasons (43) and related year using a GEO4Agri DISAFA Lab script, average composite images were created for each meteorological season (Supplementary Table S5) starting from the year 2011 until summer 2022 (Supplementary Tables S3, S4).
From the empirical evidence reported in the literature (44, 45), in which the pathogen appears to be influenced by humidity, for each scene from which the composites were obtained the multispectral TCW index was computed.
The Tasseled Cap (46) was generated based on spectral information from the Landsat satellite. The Tasseled Cap coefficients used in the linear equation of the Tasseled Cap transformation are sensor specific and therefore have been derived for each sensor system. The index is related to principal component analysis and vegetation indices. In this case reference was made only to the TCW, a multispectral index of humidity. The Tasseled Cap Wetness provides a measured value for soil and canopy moisture interactions, highlighting how moisture is distributed between the soil and the plant canopy.
where:
ρ = TOA reflectance corresponding to the regions of the electromagnetic spectrum of interest.
α = coefficient for the specific TCW transformation for each Landsat band 5 and 8, respectively. Reference was made to (47, 48).
To demonstrate a possible relationship between the environmental components TCW mappable by remote sensing and municipal-based positivity, Moran's index I was calculated to test the existence of a spatial problem, which in all cases was higher than Moran I > 0.90, consequently significant:
where:
- N = is the number of geographical units;
- xi = is the variable that describes the phenomenon under study in region i;
- μ = represents the sample mean and therefore (xi-μ) is the deviation from the mean of the variable of interest;
- Wij = is the weight matrix which in many cases is equivalent to a binary matrix i, j in which weights are used that are inversely proportional to the distance between point i and point j (where “i” is different from “j”);
- I = varies between −1.0 and +1.0 and its numerator is interpreted as the covariance between contiguous units.
4 Results
4.1 qPCR and PCR end-point
All 114 selected animals were analyzed by qPCR for the presence of the ssrA locus. Nine out of 114 (7.9 %) samples were positive by qPCR analysis and, in particular, 7/101 (6.9%) and 2/13 (15.4%) in red foxes collected in Piedmont and Aosta Valley, respectively. The positive samples further analyzed with end-point PCR yielded 7/9 samples positive for the ssrA locus, of which 3/9 positive for the gltA locus (380 and 700 bp) and none for the rpoB locus. Two sequences were not evaluable due to low quality.
In Table 1 sequencing results of Bartonella spp. positive foxes are reported. The closest relative sequence and identity results for ssrA and gltA are described.
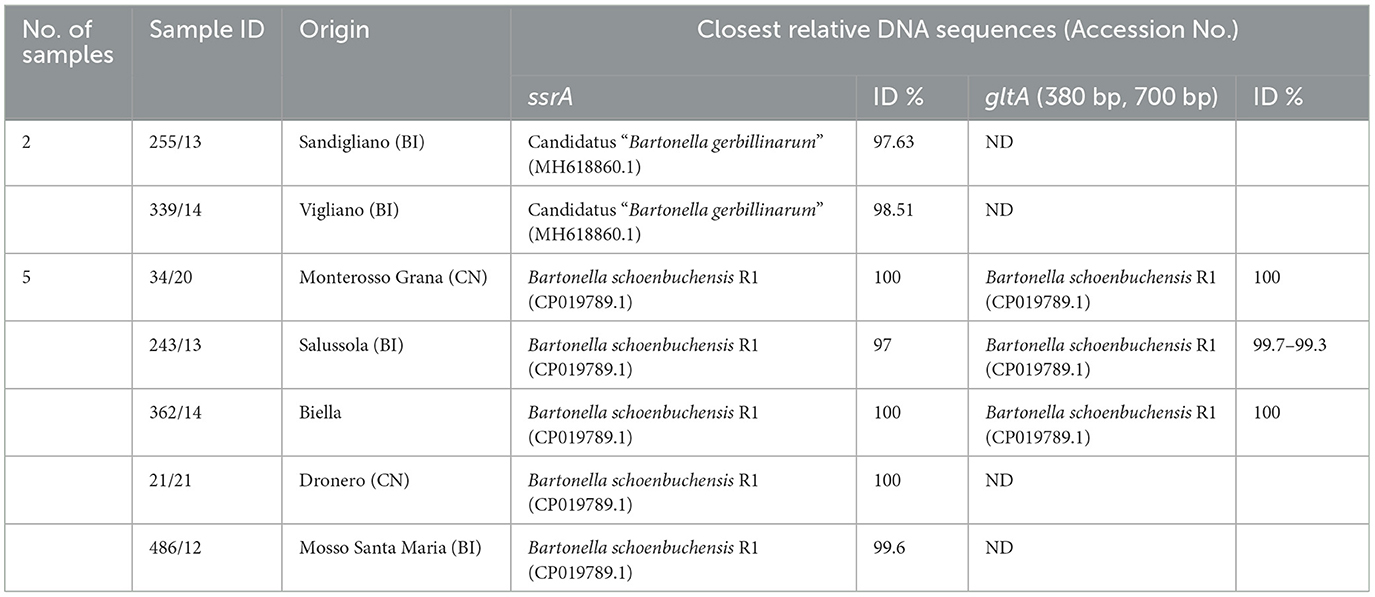
Table 1. Sequencing results of Bartonella spp. positive foxes, including the closest relative sequence (query cover 97–100%) and identity results (ID %) for ssrA and gltA loci, number (n.), and origin of finding.
Based on the sequencing of the amplicons related to the ssrA locus, the most represented Bartonella species was B. schoenbuchensis R1 (5/9, 55.6%); Candidatus “Bartonella gerbillinarum” was identified in two animals (2/9, 22.2%). B. schoenbuchensis R1 was also confirmed by sequencing the gltA locus in 3/9 animals.
4.2 Data distribution
No statistically significant associations were observed between the presence of Bartonella spp. and the sex, age of the foxes, animal area (province), years (period) and seasons. Further information regarding statistical tests specifically: F-test, t-test, Fligner-Kileen test for equal coefficients of variation and Chi-Square can be found in Supplementary material.
4.3 Geostatistical analysis
In order to analyze a possible relationship between TCW and the presence of Bartonella spp. for the composite timeseries on a seasonal basis for the years of interest, zonal statics were created: in this case Spatial Join on a municipal basis for the Aosta Valley and for the provinces of Biella and Cuneo, respectively. The municipal-based and non-punctual analysis was dictated by the fact that the samples were not punctually geo-referenced and starting from the assumption that the place where the animal was found (the municipality and neighboring area) is its core home range (moreover since we are dealing with foxes, the assumption finds correspondence in the ecology and ethology of this species).
In order to map the risk of distribution based on geostatistical results obtained, LISA maps (Local Spatial Autocorrelation Indicators) were created on a municipal scale for the periods of interest [Figure 1 reports LISA map for winter season in (Piedmont province of Cuneo) during the year 2020]. The other LISA maps are attached in Supplementary material. In particular, figure A reports LISA map for spring season in Piedmont (province of Biella) during the year 2013, figure B the spring season in Piedmont (province of Biella) during the year 2014, figure C the winter season in Piedmont (province of Biella) during the year 2014, figure D the winter season in Piedmont (province of Cuneo) during the year 2021, and finally figure E the winter season in Aosta Valley during the year 2022. Specifically, in all the maps (Figures 1A–E), warm colors (dark red, red, orange) pointed out a strong correlation between samples and TCW, while cold colors (dark blue, blue, green) a low correlation. It's worth to note that in areas where low correlations occur there were no positive samples. Due to the strong spatial correlation observed between TCW and the presence of Bartonella spp., a threshold of 0.7 TCW was identified as a potential minimum limit range to detect the pathogen, as determined by the Moran's I index.
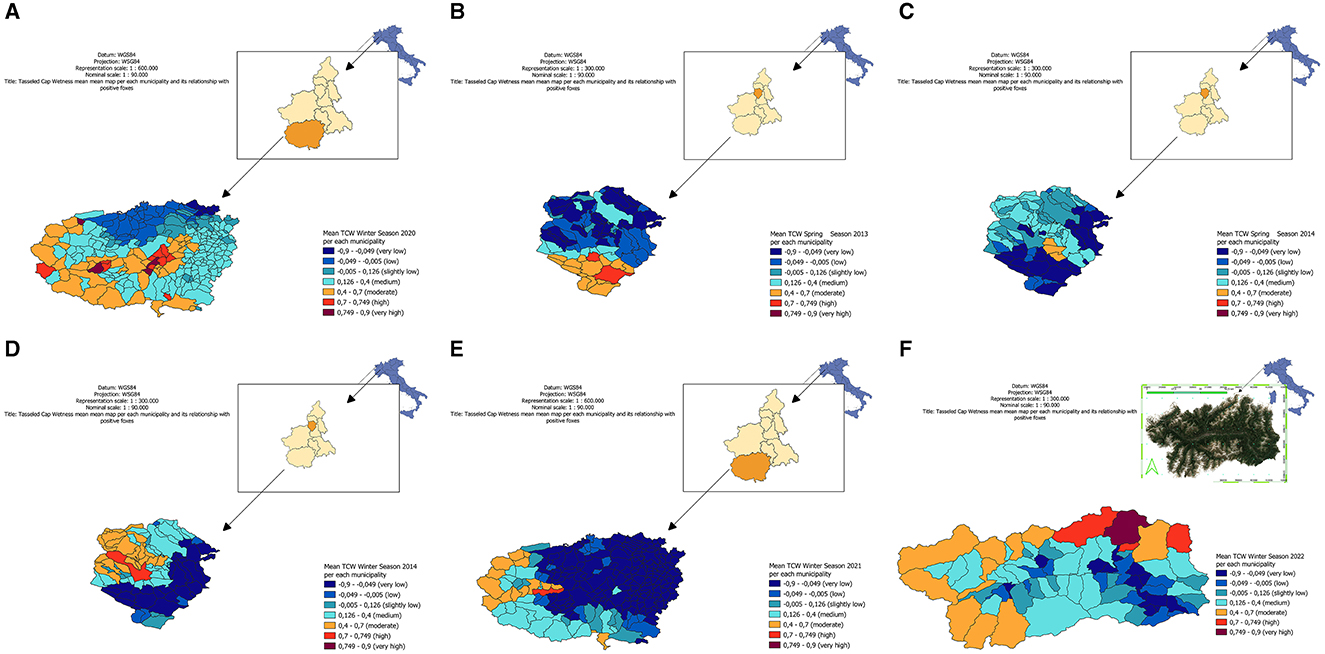
Figure 1. Local spatial autocorrelation indicator (LISA) map for Tasseled Cap Wetness (TCW) for each municipality, in the different periods analyzed. (A, E) in Piedmont, province of Cuneo. (B–D) in Piedmont, province of Biella. (F) in Aosta Valley region.
5 Discussion
The activity carried out in this study provides an interesting contribution to the knowledge of bartonellosis in the fox populations of Piedmont and Aosta Valley, starting from the year 2011 until today. First, the results show how this pathology is present in the area with rather low prevalence values, equal to 7.9% (nine positive samples out of a total of 114). Molecular investigations have highlighted the presence of Bartonella DNA belonging to the species B. schoenbuchensis R1 (sequence identity of 99.6–100%) in five foxes, two from the province of Cuneo and three from the province of Biella. This species of Bartonella causes bacteremia in ruminants and has been isolated from bloodsucking lice such as Lipoptena cervi which affect deer, roe deer, moose and sika in Europe, Siberia, and northern China, as well as red deer, moose, horses, and white-tailed cattle in North America. Accidental human infestation with Lipoptena cervi is well-documented.
Candidatus “Bartonella gerbillinarum” DNA was identified in two animals with a sequence identity of 97.6–98.5%. Moreover, for this species, several insect vectors seem to play an important role in its spread, the microbiological evaluation and epidemiological significance of which require further studies (49, 50).
Statistical analysis of the data did not reveal a statistically significant association in the distribution of data between season, year, age, sex, and municipality of origin of the samples. This result can be attributed to the scarcity of positive animals and to a rather homogeneous distribution of positivity in relation to the above factors mentioned. In fact, previous studies conducted in other species (51) have showed a different prevalence of Bartonella in summer instead of in autumn. Moreover, the prevalence was high in juvenile and subadults and declined considerably in largest-oldest individuals.
The weather and environment affect rat infections, including the ecology of zoonotic pathogens in urban ecosystems (45): high minimum temperatures were positively correlated with Bartonella tribocorum infection in rats for several time periods before the rats were captured. This result implies that B. tribocorum transmission among rats and flea vector survival may require a baseline minimum temperature.
The trend of the infection during the year is certainly also linked to the presence of the vector, which strictly depends on the environmental conditions, which can vary greatly in different seasons. Regarding fleas, it must be considered that the immature stages complete their life outside the host and consequently depend more than adults on environmental conditions (52). Humidity seems to play a fundamental role in the survival of fleas: larvae of Xenopsylla conformis and X. ramesis are more sensitive to low humidity conditions, as they are unable to close their spiracles and consequently prevent water loss through the respiratory tract (52). It's worth to note that some environmental factors are necessary for arthropod-borne infections in order to promote the development of suitable vectors, and infectious agents within these vectors (53). An increasing number of arthropods could act as potential Bartonella species vectors and, for this reason, they should be taken into consideration on clinical and serological data from human and canine cases. In particular, arthropod vectors that spend a large part of their life cycle away from their hosts, like ticks and mosquitoes, are predicted to have strict habitat requirements, whereas lice and fleas, which are always on their hosts or in their shelters, typically have more flexible requirements (54, 55).
As regards the geostatistical analysis, the TCW seems to be a good environmental indicator for identifying the presence of conditions favorable to the presence of Bartonella spp. It is interesting to note that based on the analysis conducted in the years of positivity of the tested samples, the TCW is >0.7 in the considered municipalities. Therefore, it is possible to establish a relationship between a threshold TCW and Bartonella spp. presence. In fact, the seasonal TCW was positively associated with Bartonella spp. infection in foxes as infection was always associated to TCW > 0.7. However, this is a preliminary analysis that deserves further insights. In fact, the population analyzed was small and the geographic area was limited. Despite this, the obtained results seem to corroborate the thesis of a close link between Bartonella spp. and humidity, demonstrating how ordinary epidemiological monitoring techniques can be associated and supported by remote sensing, for a territorial scale understanding of health phenomena in a perspective of “One Health” and technology transfer (56–61). While humidity may influence vector abundance and survival, its direct correlation with Bartonella infection rates may be influenced by a myriad of other factors. Conducting epidemiological studies and vector surveillance in specific geographic regions can provide valuable insights into the relationship between humidity and Bartonella transmission dynamics. In particular, due to the fact that Tasseled Cap Wetness is a spectral transformation technique commonly used in remote sensing to capture and quantify surface moisture content, by leveraging satellite imagery and spectral bands sensitive to moisture, this factor can provide valuable insights into environmental conditions conducive to vector proliferation and subsequent pathogen transmission, such as Bartonella. In regions where humidity plays a critical role in vector abundance and activity, Tasseled Cap Wetness can serve as a proxy for moisture levels in the environment. Areas exhibiting higher Tasseled Cap Wetness values may indicate regions with elevated humidity, potentially creating favorable conditions for vector breeding and survival. By integrating Tasseled Cap Wetness data with epidemiological records of Bartonella infection rates, researchers can explore spatial patterns and temporal trends in Bartonella transmission dynamics. Areas characterized by high Tasseled Cap Wetness values could be prioritized for targeted vector surveillance and control efforts, helping to mitigate the risk of Bartonella transmission to human and animal populations. Furthermore, the incorporation of Tasseled Cap Wetness into predictive model frameworks can enhance the accuracy and reliability of Bartonella risk assessments. By incorporating environmental variables such as humidity, temperature, land cover, and vector habitat suitability, researchers can develop more robust models for forecasting Bartonella outbreaks and identifying high-risk areas. For these reasons, the integration of Tasseled Cap Wetness data into studies examining the relationship between humidity and Bartonella infection holds great potential for advancing our understanding of vector-borne disease dynamics. In this way, researchers can gain valuable insights into the complex interactions between environmental factors and infectious disease transmission, ultimately informing more effective public health interventions and control strategies (62–65).
Future studies could play a crucial role in advancing our understanding of Bartonella spp. on both national and international levels. Expanding research to include a broader range of geographic regions and wildlife hosts would provide a more comprehensive epidemiological map of Bartonella distribution. Such efforts could lead to the development of more effective monitoring and control strategies for Bartonella-related diseases, which are increasingly recognized as emerging threats to both animal and human health.
Moreover, these studies would not only enhance the field of veterinary medicine by fostering new levels of expertise but also encourage greater collaboration across disciplines, including ecology, epidemiology, molecular biology, and public health. By promoting an interdisciplinary approach, researchers can better address the complexities of pathogen-host-environment interactions. This is especially important in the context of the “One Health” framework, which emphasizes the interconnectedness of human, animal, and environmental health.
The integration of advanced technologies, such as remote sensing, geographic information systems (GIS), and bioinformatics, could facilitate the identification of environmental factors that influence Bartonella transmission dynamics. High-resolution data from newer satellite missions, such as the Sentinel-2 or WorldView series or the newer space Italo-European space program IRIDE (66, 67), could be particularly valuable in this context. These missions offer greater spatial and temporal resolution, allowing for more detailed mapping of environmental conditions and finer-scale analysis of habitat characteristics. Such precise data could enhance our ability to detect subtle changes in landscape features that may affect the spread of Bartonella spp., enabling more targeted and effective interventions.
In addition, these technological advancements could support the development of predictive models, enabling more proactive disease management and potentially reducing the risk of zoonotic transmission to humans. Ultimately, this research could contribute to the establishment of a global surveillance network for Bartonella and other vector-borne pathogens, ensuring a more coordinated and effective response to emerging health threats.
6 Conclusions
This study provided a better understanding of the prevalence and genetic diversity of Bartonella species in the fox populations of Piedmont and Aosta Valley (NW Italy). Two species of Bartonella have been detected in foxes, including B. schoenbuchensis R1, responsible of infection in humans. The results raise the potential threats to public health from Bartonella: for these reasons, its surveillance in animals and investigations on suspected clinical cases in humans must be strengthened not only in the considered provinces, but also to the rest of the Italian territory, as already demonstrated by the recent studies (14).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author/s.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the study was conducted onto dead animals from incidents or hunted by hunters.
Author contributions
AV: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. TO: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing – original draft, Writing – review & editing. SD: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Validation, Visualization, Writing – review & editing. SL: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. SZ: Data curation, Investigation, Methodology, Writing – review & editing. RO: Project administration, Supervision, Validation, Visualization, Writing – review & editing. EB-M: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Supervision, Validation, Visualization, Writing – review & editing. EB: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Regione Piemonte—rinnovo atto d'intesa raccolta e smaltimento animali morti—Prof. Bollo. Codice Progetto: U-GOV: BOLE_RIC_N_COMP_17_01.
Acknowledgments
The study represent a collaboration between scientific institutions (Reference Center for Wild Animal Diseases of Aosta-CeRMAS, Department of Veterinary Sciences of the University of Turin, Department of Agricultural and Forestry Sciences of the University of Turin), and among these and the pool of research collaborators, such as hunters and public agencies responsible for wildlife management (Forestry Corps and Agriculture Department, Natural Resources of the Autonomous Region of Valle d'Aosta). A remarkable thanks to GEO4Agri DISAFA Lab.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer FS declared a shared parent affiliation with the authors to the handling editor at the time of review.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2024.1388440/full#supplementary-material
References
1. Holmala K, Kauhala K. Ecology of Wildlife Rabies in Europe. Mamm Rev. (2006) 36:17–36. doi: 10.1111/j.1365-2907.2006.00078.x
2. Nouvellet P, Donnelly CA, De Nardi M, Rhodes CJ, De Benedictis P, Citterio C, et al. Rabies and canine distemper virus epidemics in the red fox population of Northern Italy (2006–2010). PLoS ONE. (2013) 8:e61588. doi: 10.1371/journal.pone.0061588
3. Henn JB, Chomel BB, Boulouis H-J, Kasten RW, Murray WJ, et al. Bartonella Rochalimae in Raccoons, Coyotes, and Red Foxes. Emerg Infect Dis. (2009) 15:1984. doi: 10.3201/eid1512.081692
4. Schaefer JD, Kasten RW, Coonan TJ, Clifford DL, Chomel BB. Isolation or detection of Bartonella vinsonii subspecies Berkhoffii and Bartonella Rochalimae in the Endangered Island Foxes (Urocyon Littoralis). Vet Microbiol. (2011) 154:135–9. doi: 10.1016/j.vetmic.2011.06.031
5. Chomel BB, McMillan-Cole AC, Kasten RW, Stuckey MJ, Sato S, Maruyama S, et al. Candidatus Bartonella merieuxii, a potential new zoonotic Bartonella Species in Canids from Iraq. PLoS ONE. (2012) 6:e1843. doi: 10.1371/journal.pntd.0001843
6. Mccallum, H. Models for managing wildlife disease. Parasitology. (2016) 143:805–20. doi: 10.1017/S0031182015000980
7. Gerrikagoitia X, Gil H, García-Esteban C, Anda P, Juste R, Barral M. Presence of Bartonella species in wild carnivores of Northern Spain. Appl Environ Microbiol. (2012) 78:885–8. doi: 10.1128/AEM.05938-11
8. Fleischman D, Chomel BB, Kasten R, André M, Gonçalves L, Machado R. Bartonella Clarridgeiae and Bartonella vinsonii Subsp. Berkhoffii exposure in captive wild Canids in Brazil. Epidemiol Infect. (2015) 143:573–7. doi: 10.1017/S0950268814001277
9. Bai Y, Gilbert A, Fox K, Osikowicz L, Kosoy M. Bartonella Rochalimae and B. vinsonii Subsp Berkhoffii in Wild Carnivores from Colorado, USA. J Wildlife Dis. (2016) 52:844–9. doi: 10.7589/2016-01-015
10. Marciano O, Gutierrez R, Morick D, King R, Nachum-Biala Y, Baneth G, et al. Detection of Bartonella Spp. in wild carnivores, hyraxes, hedgehog and rodents from Israel. Parasitology. (2016) 143:1232–42. doi: 10.1017/S0031182016000603
11. Millán J, Proboste T, de Mera IGF, Chirife AD, de la Fuente J, Altet L. Molecular detection of vector-borne pathogens in wild and domestic carnivores and their ticks at the human–wildlife interface ticks and tick-borne. Diseases. (2016) 7:284–90. doi: 10.1016/j.ttbdis.2015.11.003
12. HodŽić A, Mrowietz N, Cezanne R, Bruckschwaiger P, Punz S, Habler VE, et al. Occurrence and diversity of arthropod-transmitted pathogens in Red Foxes (Vulpes vulpes) in Western Austria, and possible vertical (transplacental) transmission of hepatozoon Canis. Parasitology. (2018) 145:335–44. doi: 10.1017/S0031182017001536
13. Kosoy M, Goodrich I. Comparative ecology of Bartonella and Brucella infections in wild carnivores. Front Vet Sci. (2019) 5:322. doi: 10.3389/fvets.2018.00322
14. Greco G, Zarea AAK, Sgroi G, Tempesta M, D'Alessio N, Lanave G, et al. Zoonotic Bartonella species in Eurasian wolves and other free-ranging wild mammals from Italy. Zoonoses Public Health. (2021) 68:316–26. doi: 10.1111/zph.12827
15. Sakalauskas P, Lipatova I, Radzijevskaja J, Paulauskas A. pathogen screening in the Red Fox (Vulpes vulpes) from Lithuania. Biologija. (2019) 65:4122. doi: 10.6001/biologija.v65i4.4122
16. Víchová B, Bona M, Miterpáková M, Kraljik J, Cabanová V, Nemčíková G, et al. Fleas and ticks of red foxes as vectors of canine bacterial and parasitic pathogens, in Slovakia, Central Europe. Vector Borne Zoonot Dis. (2018) 18:611–9. doi: 10.1089/vbz.2018.2314
17. Bateman PW, Fleming PA. Big city life: carnivores in urban environments. J Zool. (2012) 287:1–23. doi: 10.1111/j.1469-7998.2011.00887.x
18. Ebani VV, Bertelloni F, Turchi B, Filogari D, Cerri D. Molecular survey of tick-borne pathogens in Ixodid Ticks collected from hunted wild animals in Tuscany, Italy. Asian Pac J Trop Med. (2015) 8:714–7. doi: 10.1016/j.apjtm.2015.07.033
19. de Paiva Diniz PPV, Wood M, Maggi RG, Sontakke S, Stepnik M, Breitschwerdt EB. Co-Isolation of Bartonella Henselae and Bartonella vinsonii Subsp. Berkhoffii from blood, joint and subcutaneous seroma fluids from two naturally infected dogs. Vet Microbiol. (2009) 138:368–72. doi: 10.1016/j.vetmic.2009.01.038
20. Shelnutt LM, Balakrishnan N, DeVanna J, Batey KL, Breitschwerdt EB. Death of military working dogs due to Bartonella vinsonii Subspecies Berkhoffii Genotype III endocarditis and myocarditis. Mil Med. (2017) 182:e1864–9. doi: 10.7205/MILMED-D-16-00125
21. Viani A, Orusa T, Mandola ML, Robetto S, Belvedere M, Renna G, et al. R07. 1 Tick's suitability habitat maps and tick-host relationships in wildlife. A One Health approach based on multitemporal remote sensed data, entropy and Meta® Population Dataset in Aosta Valley, NW Italy. In: Proceedings of the GeoVet 2023 International Conference. Veterinaria Italiana (2023).
22. Ottinger M, Clauss K, Kuenzer C. Aquaculture: relevance, distribution, impacts and spatial assessments–a review. Ocean Coast Manag. (2016) 119:244–66. doi: 10.1016/j.ocecoaman.2015.10.015
23. Orusa T, Orusa R, Viani A, Carella E, Borgogno Mondino E. Geomatics and EO data to support wildlife diseases assessment at landscape level: a pilot experience to map infectious keratoconjunctivitis in chamois and phenological trends in Aosta Valley (NW Italy). Remote Sensing. (2020) 12:3542. doi: 10.3390/rs12213542
24. Malone JB, Bergquist R, Martins M, Luvall, J.C. Use of geospatial surveillance and response systems for vector-borne diseases in the elimination phase. Trop Med Infect Dis. (2019) 4:15. doi: 10.3390/tropicalmed4010015
25. Carella E, Orusa T, Viani A, Meloni D, Borgogno-Mondino E, Orusa R. An integrated, tentative remote-sensing approach based on NDVI entropy to model canine distemper virus in wildlife and to prompt science-based management policies. Animals. (2022) 12:1049. doi: 10.3390/ani12081049
26. Kuenzer C, Ottinger M, Wegmann M, Guo H, Wang C, Zhang J, et al. Earth observation satellite sensors for biodiversity monitoring: potentials and bottlenecks. Int J Remote Sens. (2014) 35:6599–647. doi: 10.1080/01431161.2014.964349
27. Viani A, Orusa T, Borgogno-Mondino E, Orusa R. Snow metrics as proxy to assess sarcoptic mange in wild boar: preliminary results in Aosta Valley (Italy). Life. (2023) 13:987. doi: 10.3390/life13040987
28. Dell'Acqua F, De Vecchi D. Potentials of active and passive geospatial crowdsourcing in complementing sentinel data and supporting copernicus service portfolio. Proc IEEE. (2017) 105:1913–25. doi: 10.1109/JPROC.2017.2727284
29. Diaz MH, Bai Y, Malania L, Winchell JM, Kosoy MY. Development of a novel genus-specific real-time PCR assay for detection and differentiation of Bartonella species and genotypes. J Clin Microbiol. (2012) 50:1645–9. doi: 10.1128/JCM.06621-11
30. Divari S, Danelli M, Pregel P, Ghielmetti G, Borel N, Bollo E. Biomolecular investigation of Bartonella spp. in wild rodents of two swiss regions. Pathogens. (2021) 10:1331. doi: 10.3390/pathogens10101331
31. Norman A, Regnery R, Jameson P, Greene C, Krause D. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J Clin Microbiol. (1995) 33:1797–803. doi: 10.1128/jcm.33.7.1797-1803.1995
32. Birtles R, Laycock G, Day M, Kenny M, Shaw S. Prevalence of Bartonella species causing bacteraemia in domesticated and companion animals in the United Kingdom. Ve Record. (2002) 151:225–9. doi: 10.1136/vr.151.8.225
33. Khan S, Mohiuddin K. Evaluating the parameters of ArcGIS and QGIS for GIS applications. Int J Adv Res Sci Eng. (2018) 7:582–94.
34. Orusa T, Borgogno Mondino E. Exploring short-term climate change effects on rangelands and broad-leaved forests by free satellite data in Aosta Valley (Northwest Italy). Climate. (2021) 9:47. doi: 10.3390/cli9030047
35. Jamali AA, Kalkhajeh RG, Randhir TO, He S. Modeling relationship between land surface temperature anomaly and environmental factors using GEE and giovanni. J Environ Manage. (2022) 302:113970. doi: 10.1016/j.jenvman.2021.113970
36. Orusa T, Mondino EB. Landsat 8 thermal data to support urban management and planning in the climate change era: a case study in Torino Area, NW Italy. Proc Remote Sensing Technol Appl Urban Environ. (2019) 11157:133–49. doi: 10.1117/12.2533110
37. Velastegui-Montoya A, Montalván-Burbano N, Carrión-Mero P, Rivera-Torres H, Sadeck L, Adami M. Google Earth Engine: a global analysis and future trends. Remote Sensing. (2023) 15:3675. doi: 10.3390/rs15143675
38. Orusa T, Viani A, Moyo B, Cammareri D, Borgogno-Mondino E. Risk Assessment of rising temperatures using Landsat 4–9 LST time series and Meta® population dataset: an application in Aosta Valley, NW Italy. Remote Sensing. (2023) 15:2348. doi: 10.3390/rs15092348
39. Viani A, Orusa T, Borgogno-Mondino E, Orusa R. A One Health Google Earth Engine Web-GIS Application to evaluate and monitor water quality worldwide. Euromediterr J Environ Integra. (2024). doi: 10.1007/s41207-024-00528-w
40. Orusa T, Cammareri D, Borgogno Mondino E. A scalable earth observation service to map land cover in geomorphological complex areas beyond the dynamic world: an application in Aosta Valley (NW Italy). Appl Sci. (2022) 13:390. doi: 10.3390/app13010390
41. Chander G, Markham BL, Helder DL. Summary of current radiometric calibration coefficients for Landsat MSS, TM, ETM+, and EO-1 ALI sensors. Remote Sens Environ. (2009) 113:893–903. doi: 10.1016/j.rse.2009.01.007
42. Mishra N, Haque MO, Leigh L, Aaron D, Helder D, Markham B. Radiometric cross calibration of Landsat 8 Operational Land Imager (OLI) and Landsat 7 Enhanced Thematic Mapper plus (ETM+). Remote Sensing. (2014) 6:12619–38. doi: 10.3390/rs61212619
43. Orusa T, Viani A, Cammareri D, Borgogno Mondino E. A Google Earth Engine Algorithm to map phenological metrics in mountain areas worldwide with Landsat collection and Sentinel-2. Geomatics. (2023) 3:221–38. doi: 10.3390/geomatics3010012
44. Jameson P, Greene C, Regnery R, Dryden M, Marks A, Brown J, et al. Prevalence of Bartonella henselae antibodies in pet cats throughout regions of North America. J Infect Dis. (1995) 172:1145–9. doi: 10.1093/infdis/172.4.1145
45. Rothenburger JL, Himsworth CG, Nemeth NM, Pearl DL, Jardine CM. Beyond abundance: how microenvironmental features and weather influence Bartonella tribocorum Infection in Wild Norway Rats (Rattus norvegicus). Zoonoses Public Health. (2018) 65:339–51. doi: 10.1111/zph.12440
46. Kauth RJ, Thomas G. The Tasselled Cap–a Graphic description of the spectral-temporal development of agricultural crops as seen by Landsat. In: Proceedings of the LARS Symposia. Purdue University (1976). p. 159. Available at: https://docs.lib.purdue.edu/cgi/viewcontent.cgi?article=1160&context=lars_symp
47. DeVries B, Pratihast AK, Verbesselt J, Kooistra L, Herold M. Characterizing forest change using community-based monitoring data and Landsat time series. PLoS ONE. (2016) 11:e0147121. doi: 10.1371/journal.pone.0147121
48. Baig MHA, Zhang L, Shuai T, Tong Q. Derivation of a tasselled cap transformation based on Landsat 8 at-satellite reflectance. Remote Sensing Lett. (2014) 5:423–31. doi: 10.1080/2150704X.2014.915434
49. Weber JM, Aubr S, Lachat N, Meia JS, Mermod C, Paratte A. Fluctuations and behaviour of foxes determined by nightlighting. Preliminary results. Acta Theriologica. (1991) 36:285–91. doi: 10.4098/AT.arch.91-29
50. Alsarraf M, Mohallal EM, Mierzejewska EJ, Behnke-Borowczyk J, Welc-Faleciak R, Bednarska M, et al. Description of Candidatus Bartonella fadhilae n. sp and Candidatus Bartonella sanaae n Sp(Bartonellaceae) from Dipodillus dasyurus and Sekeetamys calurus (Gerbillinae) from the Sinai massif (Egypt). Vector Borne Zoonot Dis. (2017) 17:483–94. doi: 10.1089/vbz.2016.2093
51. Kosoy M, Mandel E, Green D, Marston E, Childs J. Prospective studies of Bartonella of rodents. Part I demographic and temporal patterns in population dynamics. Vector Borne Zoonot Dis. (2004) 4:285–95. doi: 10.1089/vbz.2004.4.285
52. Krasnov B, Khokhlova I, Fielden L, Burdelova N. Development rates of two xenopsylla flea species in relation to air temperature and humidity. Med Vet Entomol. (2001) 15:249–58. doi: 10.1046/j.0269-283x.2001.00295.x
53. Service M. Agricultural development and arthropod-borne diseases: a review. Rev Saúde Pública. (1991) 25:165–78. doi: 10.1590/S0034-89101991000300002
54. Beldomenico P, Chomel B, Foley J, Sacks B, Baldi C, Kasten R, et al. Environmental factors associated with Bartonella vinsonii Subsp. Berkhoffii seropositivity in free-ranging coyotes from Northern California. Vect Borne Zoonot Dis. (2005) 5:110–9. doi: 10.1089/vbz.2005.5.110
55. Billeter S, Levy M, Chomel B, Breitschwerdt E. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med Vet Entomol. (2008) 22:1–15. doi: 10.1111/j.1365-2915.2008.00713.x
56. Orusa T, Viani A, Borgogno-Mondino E. IRIDE the Euro-Italian Earth observation program: overview, current progress, global expectations and recommendations. Perspective. (2023) 2:10.
57. Ippoliti C, Candeloro L, Gilbert M, Goffredo M, Mancini G, Curci G, et al. Defining ecological regions in Italy based on a multivariate clustering approach: a first step towards a targeted vector borne disease surveillance. PLoS ONE. (2019) 14:e0219072. doi: 10.1371/journal.pone.0219072
58. Conte A, Candeloro L, Ippoliti C, Monaco F, De Massis F, Bruno R, et al. Spatio-temporal identification of areas suitable for West Nile Disease in the Mediterranean Basin and Central Europe. PLoS ONE. (2015) 10:e0146024. doi: 10.1371/journal.pone.0146024
59. Candeloro L, Ippoliti C, Iapaolo F, Monaco F, Morelli D, Cuccu R, et al. Predicting WNV circulation in Italy using Earth Observation Data and Extreme Gradient Boosting Model. Remote Sensing. (2020) 12:3064. doi: 10.3390/rs12183064
60. De Marinis P, De Petris S, Sarvia F, Manfron G, Momo EJ, Orusa T, et al. Supporting pro-poor reforms of agricultural systems in Eastern DRC (Africa) with remotely sensed data: a possible contribution of spatial entropy to interpret land management practices. Land. (2021) 10:1368. doi: 10.3390/land10121368
61. Latini G, Bagliani M, Orusa T. Lessico e Nuvole: Le Parole Del Cambiamento Climatico. Università degli Studi di Torino: Youcanprint (2021).
62. Orusa T, Viani A, Borgogno-Mondino E. Earth observation data and geospatial deep learning AI to assign contributions to european municipalities Sen4MUN: an empirical application in Aosta Valley (NW Italy). Land. (2024) 13:80. doi: 10.3390/land13010080
63. Napp S, Petrić D, Busquets N. West Nile virus and Other Mosquito-Borne viruses present in Eastern Europe. Pathog Glob Health. (2018) 112:233–48. doi: 10.1080/20477724.2018.1483567
64. Orusa T, Cammareri D, Borgogno Mondino E, A. Possible land cover EAGLE approach to overcome remote sensing limitations in the alps based on Sentinel-1 and Sentinel-2: the Case of Aosta Valley (NW Italy). Remote Sensing. (2022) 15:178. doi: 10.3390/rs15010178
65. Caminade C, McIntyre KM, Jones AE. Impact of recent and future climate change on vector-borne diseases. Ann N Y Acad Sci. (2019) 1436:157–73. doi: 10.1111/nyas.13950
66. Orusa T, Viani A, Borgogno-Mondino E. IRIDE, the Euro-Italian Earth Observation Program: overview, current progress, global expectations, and recommendations. Environ Sci Proc. (2024) 29:74. doi: 10.3390/ECRS2023-16839
67. Mastracci F, Geraldini S. IRIDE: dai Servizi definiti dall'Utente alle Costellazioni di Satelliti, il primo Sistema Italiano end-to-end di Osservazione della Terra da Satellite. GEOmedia. (2023) 27. Available at: https://www.mediageo.it/ojs/index.php/GEOmedia/article/view/1977
68. Vissotto de Paiva Diniz PP, Maggi RG, Schwartz DS, Cadenas MB, Bradley JM, Hegarty B, et al. Canine bartonellosis: serological and molecular prevalence in Brazil and evidence of co-infection with Bartonella henselae and Bartonella vinsonii subsp berkhoffii. Vet Res. (2007) 38:697–710. doi: 10.1051/vetres:2007023
Keywords: Bartonella spp., fox, public health, epidemiology, GIS and remote sensing
Citation: Viani A, Orusa T, Divari S, Lovisolo S, Zanet S, Orusa R, Borgogno-Mondino E and Bollo E (2025) Detection of Bartonella spp. in foxes' populations in Piedmont and Aosta Valley (NW Italy) coupling geospatially-based techniques. Front. Vet. Sci. 11:1388440. doi: 10.3389/fvets.2024.1388440
Received: 19 February 2024; Accepted: 25 October 2024;
Published: 21 January 2025.
Edited by:
Bedaso Edao, Addis Ababa University, EthiopiaReviewed by:
Mariana Alejandra Rivero, Universidad Nacional del Centro de Buenos Aires, ArgentinaYokomi N. Lozano-Sardaneta, National Autonomous University of Mexico, Mexico
Filippo Sarvia, University of Turin, Italy
Copyright © 2025 Viani, Orusa, Divari, Lovisolo, Zanet, Orusa, Borgogno-Mondino and Bollo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annalisa Viani, YXZpYW5pQGF1c2wudmRhLml0