 Ondřej Máca1,2†
Ondřej Máca1,2† Marek Kouba3,4†
Marek Kouba3,4† Iva Langrová2†
Iva Langrová2† Lucie Panská2,5†
Lucie Panská2,5† Erkki Korpimäki6†
Erkki Korpimäki6† David González-Solís7*†
David González-Solís7*†- 1Department of Pathology and Parasitology, State Veterinary Institute Prague, Prague, Czechia
- 2Department of Zoology and Fisheries, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 3Department of Ethology and Companion Animal Science, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 4Department of Game Management and Wildlife Biology, Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Prague, Czechia
- 5Department of Ecology, Faculty of Environmental Sciences, Czech University of Life Sciences Prague, Prague, Czechia
- 6Section of Ecology, Department of Biology, University of Turku, Turku, Finland
- 7Department of Systematics and Aquatic Ecology, El Colegio de la Frontera Sur, Chetumal, Quintana Roo, Mexico
Background: Owls have been reported as definitive hosts, whereas wild small mammals (naturally and experimentally) as intermediate hosts of several species of Sarcocystis. Recently, dead fledglings were found infected by an unnamed species of Sarcocystis since its intermediate host was unknown. After collecting additional samples of owls and wild small mammals, the present study focused on elucidating the identity, potential intermediate host, and complete life cycle of the found Sarcocystis through experimentally infected rodents. The developmental stages’ morphological and molecular characterizations (28S rRNA gene, ITS1 region) are presented herein.
Methods: In total, 21 Tengmalm’s owl carcasses (15 nestlings, 5 fledglings, and 1 adult male) were collected in Kauhava (west-central Finland) and parasitologically examined by wet mounts. Intestinal mucosa scrapings were used to isolate oocysts/sporocysts and employed for experimental infections in dexamethasone-immunosuppressed BALB/cOlaHsd mice. Additionally, sarcocysts were searched in the skeletal muscle of 95 samples from seven wild small mammal species. All these developmental stages were molecularly characterized by the 28S rRNA gene and ITS1 region. Experimental infections were carried out by using immunosuppressed female 8-week-old BALB/cOlaHsd mice, divided into three groups: (1) water with 15 μg/mL of dexamethasone, (2) water with 30 μg/mL of dexamethasone, (3) no dexamethasone treatment. Each group consisted of four individuals. In each group, two mice were infected with 1,000 sporocysts each, and the remaining two with 10,000 sporocysts each. All mice were euthanized on specific days post-infection.
Results: The intestinal mucosa of 11 nestlings and 5 fledglings of the Tengmalm’s owl were positive for Sarcocystis funereus sp. nov. The adult male owl and all owls’ breast and heart muscles were negative for Sarcocystis. Two dexamethasone-immunosuppressed BALB/cOlaHsd mice (group 2) were positive to S. funereus sp. nov. in diaphragm and leg muscles after 22- and 24-day post-infection. Some sarcocysts were found in the wild small mammals. Molecular identification at 28S rRNA revealed sequences from naturally infected Tengmalm’s owls, as well as sarcocysts of dexamethasone-immunosuppressed BALB/cOlaHsd mice were 99.87–100% similar to Sarcocystis sp. isolate Af1 previously found in the Tengmalm’s owl. At the ITS1 region, the S. funereus sp. nov. isolates Af2 haplotype B and Af3 haplotype A were 98.77–100% identical to Sarcocystis sp. isolate Af1. The sequences from sarcocysts of naturally infected wild small mammals were 75.23–90.30% similar at ITS1 region to those of S. funereus sp. nov.
Conclusion: The morphological and molecular characterizations and phylogenetic placement of S. funereus sp. nov. are presented here for the first time and support the erection of the new species.
Introduction
Species of Sarcocystis (Apicomplexa) have an indirect, two-host life cycle in which mammals, birds, reptiles, and humans are involved. Due to the wide variety of hosts acting as intermediate or definitive hosts, it is not easy to know the natural life cycle of many species. Birds represent a group of vertebrates commonly utilized as either intermediate and definitive hosts by various members of the genus Sarcocystis. Particularly, several species of owls have been reported as definitive hosts, while wild small mammals (rodents and shrews) serve as (natural and experimental) intermediate hosts of Sarcocystis. Such a relationship was, for instance, described in the case of the barn owl Tyto alba and the house mouse Mus musculus (1, 2) and the masked owl Tyto novaehollandiae (3) with Sarcocystis dispersa, the northern saw-whet owl Aegolius acadicus and the eastern deer mouse Peromyscus maniculatus with Sarcocystis espinosai (4), the tawny owl Strix aluco and M. musculus with Sarcocystis scotti [this species is considered invalid by Dubey et al. (5)] (3, 6, 7), St. aluco and the wood mouse Apodemus sylvaticus with Sarcocystis sebeki (8, 9), as well as the snowy owl Bubo scandiacus (reported as Nyctea scandica) and Richardson’s collared lemming Dicrostonyx richardsoni with Sarcocystis rauschorum (10, 11). Wiesner (12) further described an unnamed Sarcocystis species in the Tengmalm’s owl Aegolius funereus and experimentally found the bank vole Clethrionomys (=Myodes) glareolus to be its intermediate host. Thus, the involvement of wild small mammals in the life cycles of Sarcocystis parasitizing owls appears to be mandatory and obligate.
A recent analysis of a 45-year breeding data set on Tengmalm’s owl population in the Kauhava study area (Finland) revealed a decreasing trend in fledgling production corresponding with the long-term decline of the whole population (13). Additionally, we have documented long-term decline in the body condition of both male and females parent owls (14). Though the primary reasons for the observed trends regarding the local population are inappropriate forest management, we know practically nothing about the possible detrimental effects of different internal or external parasites on the individual Tengmalm’s owl’s long-term survival. The recent findings, indicating that Sarcocystis sp. isolate Af1 [(see 15)] infected 100% of dead fledglings, have raised doubts about the presumed harmlessness of the mentioned parasite. Thus, the main aim of the present study was to determine the identity, potential intermediate host/s, and complete life cycle of the Sarcocystis sp. isolate Af1, through experimentally infecting rodents with newly collected oocysts and sporocysts isolated from the Tengmalm’s owls inhabiting the same study area. The morphological and molecular characterizations (28S rRNA gene, ITS1 region) of the developmental stages are presented herein.
Methods
The Tengmalm’s owl carcasses were collected in the Kauhava study area (west-central Finland) throughout the breeding season 2021 during regular nest box visits starting in early April and later during radio-tracking of fledged young [see details in Kouba et al. (16)]. A total of 21 specimens (15 nestlings, 5 fledglings, and 1 adult male) were sent frozen to the State Veterinary Institute (SVI) Prague, Czech Republic, where parasitological examinations of intestine and muscles (breast, legs, and heart) were carried out by wet mounts. Intestinal mucosa scrapings were used to isolate oocysts/sporocysts under light microscopy with an optical microscope (Leica DM2500 LED), a digital camera (Leica DMC5400), and Leica Application Suite X microscope software (both Leica Microsystems, Wetzlar, Germany). Prior to experimental infections, 25 whole dead small mammals and 70 hind leg samples of seven wild species [i.e., the short-tailed field vole Microtus agrestis (2 whole bodies, 2 legs), the sibling vole Microtus rossiaemeridionalis (5 bodies), the European water vole Arvicola amphibius (1 body), the Eurasian harvest mouse Micromys minutus (5 bodies, 23 legs), the bank vole (4 bodies, 30 legs), the common shrew Sorex araneus (5 bodies, 15 legs), and the Eurasian pygmy shrew Sorex minutus (3 bodies)] were collected from owl nests and examined by wet smear of skeletal muscle. The prey items/bodies were exchanged for frozen newly hatched chickens not to deprive the owls of food.
Oocysts/sporocysts isolated from 2 birds were used to experimentally infect dexamethasone-immunosuppressed BALB/cOlaHsd mice (see below). At the same time, other parasite developmental stages were stored in Eppendorf tubes for DNA extraction under −20°C until further use. All measurements are given in micrometers unless otherwise specified. The molecular analysis of the oocysts/sporocysts isolates (n = 16) from the intestinal mucosa of owls and sarcocyst isolates from wild small mammals (n = 9) was carried out following that of Máca et al. (15), with minor changes. Genomic DNA of oocysts/sporocysts was extracted by the QIAamp® Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany), while that of sarcocysts by using NucleoSpin tissue XS kit (Macherey-Nagel, Düren, Germany).
All isolates were characterized by the 28S rRNA gene and ITS1 region by using the following primers: KL_P1F/KL_P2R, KL_P2F/P1R, and ITSR/ITSF, respectively (17). PCR procedures were performed in reaction mixtures consisting of 12.50 μL of GoTaq® G2 Hot Start Green Master Mix (Promega, Madison, WI, United States), 0.4 μM of each primer, and 5 μL DNA template. RNase/Dnase-free water was used to top up the reaction mixture to a final volume of 25 μL. PCR amplification of negative controls was also conducted simultaneously. PCR conditions were as follows: 95°C for 3 min, 5 cycles of 94°C for 45 s, 64°C for 60 s, 72°C for 90 s; followed by 30 cycles of 95°C for 30 s, 58°C for 30 s, 72°C for 1 min, with a final elongation step of 72°C for 10 min. Amplified products were checked on 1% agarose gel electrophoresis and visualized on a UV transilluminator. Positive PCR products were purified with the ExoSAP-IT™ Express PCR Product Cleanup Reagent Kit (Thermo Fisher Scientific) and sent for sequencing on both strands (using the same forward and reverse primers as for the PCR) to the commercial company Eurofins Genomics (Ebersberg, Germany).
The reference nucleotide sequences used were selected based on similarities using the Basic Local Alignment Search Tool (BLAST) for sequence analysis1. The most similar sequences were downloaded and compared with the newly obtained sequences, aligned using the MAFFT software version 7 online server2 (18) for phylogenetic analysis using the MEGA 11 software version 11.0.13 (19). The phylogenetic trees were inferred by using the Maximum Likelihood (ML) with evolutionary distances calculated by the best-fitting model based on the lowest Bayesian Information Criterion (BIC) scores and resulted as Hasegawa-Kishino-Yano model (20) for the 28S rRNA gene (analysis involved 38 nucleotide sequences with a total of 1,409 positions). The Hasegawa-Kishino-Yano model (20) was also the best model based on BIC scores for the ITS1 region (involving 25 nucleotide sequences with a total of 1,610 positions in the final dataset), both modeled with a Gamma distribution and invariant sites, with 1,000 bootstrap replications.
Immunosuppression of 8-week-old BALB/cOlaHsd female mice (ENVIGO) was required to establish a successful infection with Sarcocystis sporocysts. The mice were divided into three groups, each consisting of four individuals. Two of these groups received water-soluble dexamethasone sodium phosphate (Sigma-Aldrich) dissolved in their drinking water. The first group (n = 4) received water with a concentration of 15 μg/mL of dexamethasone continuously starting 2 weeks before the infection and continuing until the end of the experiment. The second group (n = 4) received water with a concentration of 30 μg/mL of dexamethasone 1 day before the infection until the end of the experiment. The third group (n = 4) did not receive any dexamethasone treatment. All mice were orally inoculated with oocysts/sporocysts delivered with food. In each group, two mice were infected with 1,000 sporocysts each, and the other two were infected with 10,000 sporocysts each. The data for parasitological examination was collected as follows: all mice were euthanized by intraperitoneal injection of ketamine (Narkamon 5%, Bioveta; 1.2 mL/kg) in combination with xylazine (Rometar 2%, Bioveta; 0.6 mL/kg) on specific days (22, 24, 52, 58, 69, and 77 days) after the infection with oocysts/sporocysts (Table 1). Subsequently, muscles containing sarcocysts were fixed in 10% formalin, embedded in paraffin, and sectioned. The histological sections were stained with hematoxylin and eosin and examined under the microscope (Leica DM2500 LED).
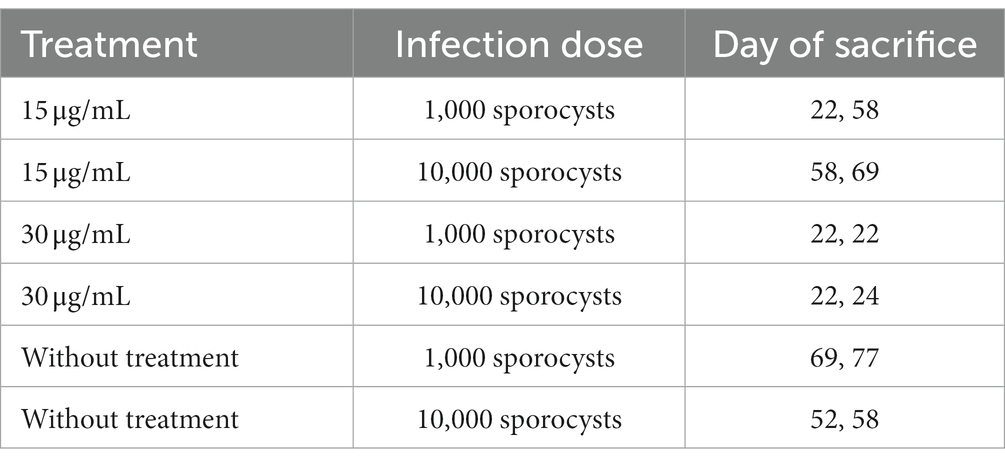
Table 1. Experimental design for establishing the Sarcocystis infection in 12 dexamethasone-immunosuppressed BALB/cOlaHsd mice (2 mice per treatment).
Owls were tagged and radio-tracked, and the carcasses were transported to Czechia under the approval of the Centre for Economic Development, Transport, and the Environment (Varsinais-Suomen Elinkeino-, Liikenne-ja Ympäristökeskus: permit numbers VARELY/1389/2018 and VARELY/5933/2019, respectively). All experiments and the maintenance of experimental animals were consistent with current animal welfare laws of the Czech Republic and were approved by the Animal Welfare Committee of the Czech University of Life Sciences Prague (permit number: MSMT-15824/2023-4).
Results
The intestinal mucosa of 11 out of 15 (73%) Tengmalm’s owl nestlings and 5 out of 5 (100%) fledglings were positive for Sarcocystis. The only adult male owl available was negative to the presence of the parasite. The breast and heart muscles of all 21 examined owls were negative for Sarcocystis. No macroscopic lesions were observed in the organs of infected birds. No other protozoan gastrointestinal parasites were found. Sarcocysts from wild small mammals were found in 9 (1 M. rossiaemeridionalis, 7 S. araneus, 1 S. minutus) out of 95 samples (9.5% prevalence). All samples from A. amphibius, C. glareolus, M. agrestis, and Mi. minutus were negative for sarcocysts.
Developmental stages were described as follows:
Family Sarcocystidae Poche, 1913.
Sarcocystis funereus sp. nov. (Figure 1).
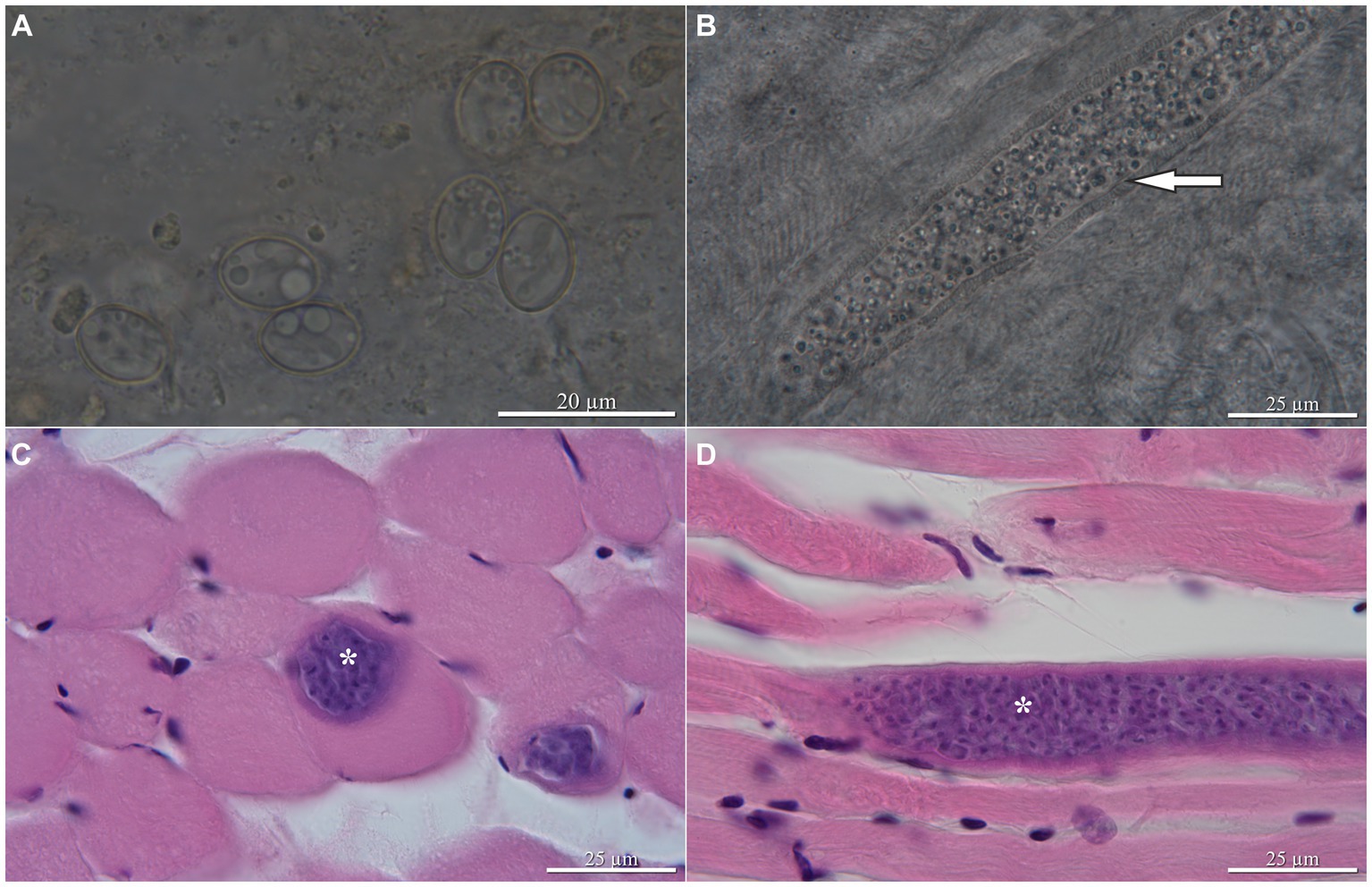
Figure 1. Sarcocystis funereus sp. nov., light micrographs. Oocysts/sporocysts from the intestinal mucosa of the Tengmalm’s owl Aegolius funereus from Finland; arrow indicates cyst wall with finger-like protrusions (A), sarcocyst in wet mount (B) and hematoxylin and eosin staining (C,D) from the skeletal muscles of experimentally infected dexamethasone-immunosuppressed BALB/cOlaHsd mice 24 days postinfection, longitudinal and transversal sections, respectively. Asterisks indicate sarcocysts.
Description: Thin-walled sporulated oocysts, 16.7–17.0 × 11.5–12.2 (n = 5), and sporocysts (Figure 1A) were 11.3–12.9 × 8.1–9.3 (n = 50). Sarcocysts were microscopic, the largest 219.5 long and 30.7 wide (found at 24 days post-infection), elongate, ribbon-shaped, with both ends rounded (Figures 1B–D). Wall was 3.1 thick, characterized by dense finger-shaped villar protrusions, 2.6 long (Figure 1B).
Taxonomic summary
Definitive host: Tengmalm’s owl Aegolius funereus Linnaeus, 1758 (Strigiformes: Strigidae).
Natural Intermediate host: Unknown.
Experimental intermediate host: Dexamethasone-immunosuppressed BALB/cOlaHsd mouse.
Distribution: Kauhava region, west-central Finland (~63° N, 23° E).
Site of infection: Small intestine (definitive host), skeletal muscle (experimental intermediate host).
Deposited material: Symbiotype (oocysts/sporocysts in 2.5% potassium dichromate) and genomic DNA in an Eppendorf tube were stored at SVI Prague. GenBank sequences OR725602 and OR726006 (28S rRNA gene), OR726007 and OR726008 (ITS1 region). Positive mice were frozen at −20°C, and histology slides were stored in lab SVI Prague.
Sequences obtained from the experimental study: OR725602 (28S rRNA gene), and OR726007 (ITS1 region).
ZooBank registration: To comply with the regulations in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (21), details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) for Sarcocystis funereus sp. nov. is urn:lsid:zoobank.org:pub:536F1351-1157-4C09-999A-D41078EE3CBC.
Etymology: The specific epithet is derived from the species name of its definitive host, i.e., funereus.
Molecular identification at 28S rRNA revealed that 12 sequences [isolate Af3 haplotype A (OR726006) and isolate Af2 haplotype B (OR725602), both 1,509 bp] obtained from the 16 oocyst/sporocyst isolates (4 failed sequencing) from the naturally infected Tengmalm’s owls, as well as the 6 sequences (haplotype B) of the 6 sarcocyst isolates in the skeletal muscles of dexamethasone-immunosuppressed BALB/cOlaHsd mouse were 99.87–100% similar to Sarcocystis sp. isolate Af1 (MW349707), 97.59% similar to Sarcocystis strixi (MF162316) and 97.46–97.53% to Sarcocystis lari (MF946611), in the white-tailed sea eagle Haliaeetus albicilla from Norway; and 97.42–97.49% similar to Sarcocystis lutrae (KM657771) in the Eurasian otter Lutra lutra from Norway. Haplotypes A and B were 99.87% similar each other and showed single cases of double peaks at nucleotide positions 666 and 667 (TT/CC), especially in those samples from owls and experimental mice that resulted in TT (haplotype A) or CC (haplotype B) peaks or double peaks at these positions. Isolates used for experimental infections and those of all sarcocysts resulted in CC dominant peak at this position and represent S. funereus sp. nov. isolate Af2 haplotype B (OR725602). Sarcocystis cf. strixi isolate LTAfl120 (OQ557459) in the tawny mouse Apodemus flavicollis from Lithuania and Sarcocystis sp. isolate No. 5 (AF513497) in S. araneus from Czech Republic were 95.37 (47% query cover; 733 bp) and 96.22% (35% query cover; 554 bp), respectively, similar to haplotypes A and B of S. funereus sp. nov., although both have short sequences and were not used in the phylogenetic analysis.
All isolates were successfully sequenced at the ITS1 region. Like at the 28S rRNA gene, the isolates S. funereus sp. nov. isolate Af2 (OR726007, 1,300 bp) and S. funereus sp. nov. isolate Af3 (OR726008, 1,297 bp) were used for infection and all sequences obtained from sarcocysts (OR726007) showed 10 SNPs and 3 nucleotide insertions of GTG in position 1,011–1,013. Those insertions were not found in other ITS1 region sequences that Máca et al. (15) obtained. Similarly, there was only one SNP (T/C) at nucleotide position 466 in newly obtained sequences. Moreover, S. funereus sp. nov. isolate Af2 (OR726007) and S. funereus sp. nov. isolate Af3 (OR726008) were 98.77–100% identical to Sarcocystis sp. isolate Af1 (MW373964), and 88.43–89.57% (26–45% query cover) to S. lutrae (MG372108) in the European badger Meles meles from the Czech Republic and Sarcocystis halieti (MF946596) in the white-tailed sea eagle from Norway.
The phylogenetic tree showed that isolates Af3 haplotype A and Af2 haplotype B of S. funereus sp. nov. at the 28S rRNA gene grouped with Sarcocystis sp. Af1, which was previously reported in A. funereus. Moreover, S. strixi appears as a sister species in a close clade (Figure 2A). At the ITS1 region, sequences of haplotypes A and B are also grouped in a single clade with Sarcocystis sp. Af1 (Figure 2B).
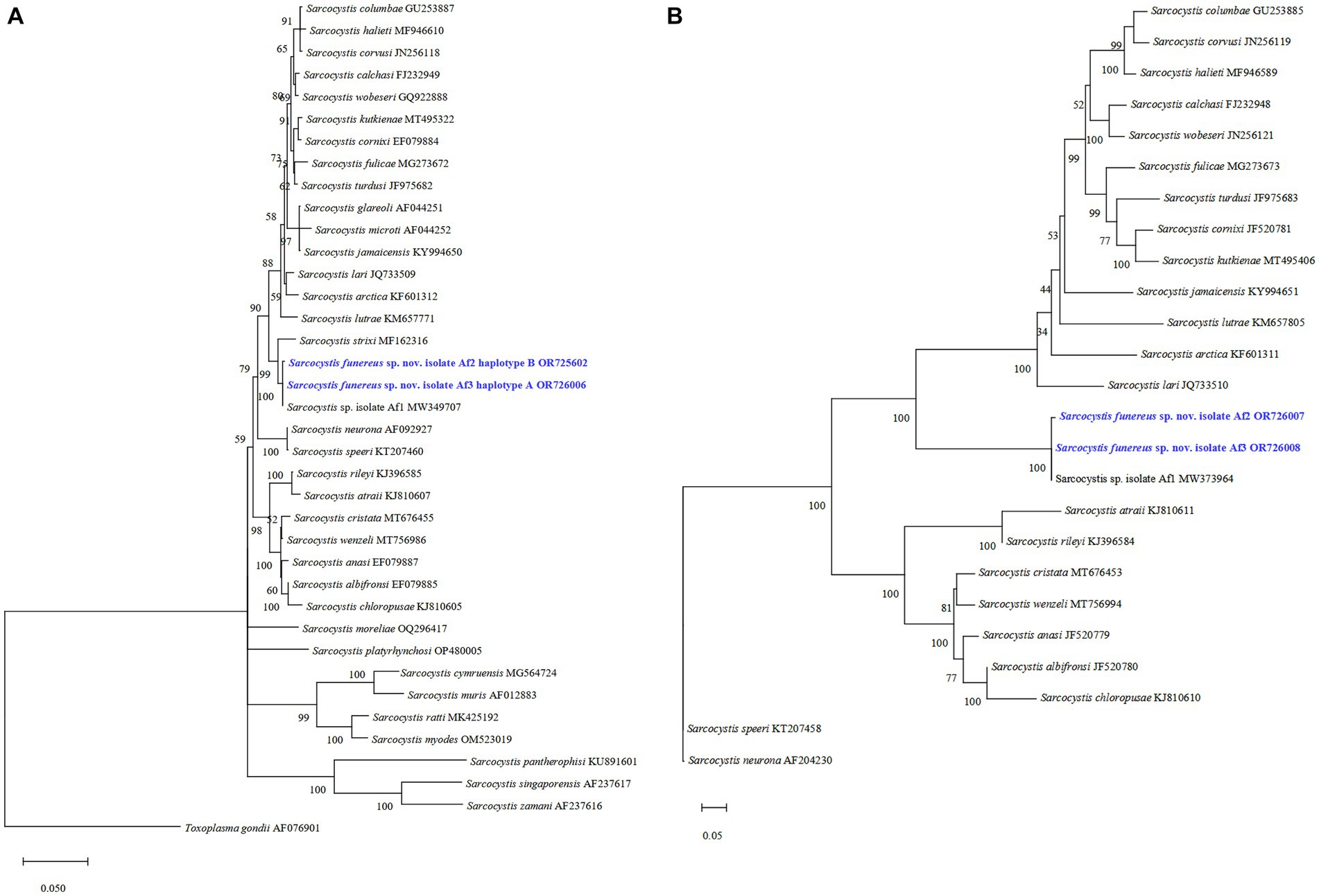
Figure 2. Phylogenetic trees of the related species of Sarcocystis from various hosts based on sequences of the 28S rRNA gene (A) and the ITS1 region (B). The numbers on phylogenetic trees represent bootstrap values based on 1,000 replications. GenBank accession numbers follow Sarcocystis taxa.
Out of the 8 dexamethasone-immunosuppressed BALB/cOlaHsd mice used for the experimental infections, only 2 of the second group, treated with 30 μg/mL dexamethasone and dosed with 10,000 sporocysts were positive to S. funereus sp. nov. after 22- and 24-day post-infection. Both mice showed sarcocysts in the skeletal muscles (diaphragm, leg), but no in the brain nor heart. Infected mice were asymptomatic during the whole infection process, from inoculation to euthanasia.
The 6 sequences (5 from S. araneus, 1 from S. minutus) successfully obtained from the sarcocysts of naturally infected small mammals were molecularly identified by using the ITS1 region marker (unpublished data). Those sequences from S. araneus and S. minutus were 75.23–90.30% (13–26% query cover) and 90.30% (19% query cover), respectively, similar to Sarcocystis sp. isolate Af1 (MW373964) and S. funereus sp. nov. isolate Af3 (OR726008). Negative PCR resulted in the samples of M. rossiaemeridionalis and 2 of S. araneus.
Discussion
While recently reporting the finding of oocysts/sporocysts in the intestine of the Tengmalm’s owl, Máca et al. (15) refrained from specifically naming the Sarcocystis sp. isolate Af1, since its intermediate host and sarcocysts were unknown. This unnamed Sarcocystis was fully characterized by using four loci (18S rRNA, 28S rRNA, ITS1, and CO1), so we decided to only use two of them (28S rRNA and ITS1) because both, but especially the ITS1 region, clearly delimite species using avian hosts than the other two loci (22). Máca et al. (15) mentioned that the ITS1 region is more sensitive to the genetic differences among Sarcocystis species from birds and carnivores as intermediate hosts, while CO1 are considered of limited taxonomic help.
After the experimental infections of mice, the presence of sarcocysts in the skeletal muscles elucidates the determination of such species as new to science and the possible route of the life cycle. The current finding supports the rodent-owl life cycle reported in our previous study (15). The phylogenetic position of S. funereus sp. nov. inferred separately at the 28S rRNA gene and ITS1 region was the same than that obtained by Máca et al. (15); thus, Sarcocystis sp. isolate Af1 should be considered to belong to S. funereus sp. nov.
There is only one previous record of an unnamed Sarcocystis in the Tengmalm’s owl, whose sporocysts and oocysts were not morphologically nor molecularly described [(see 12)]; additionally, this unnamed species was experimentally transmitted to the bank vole, while the new species to immunosuppressed mice. Considering that Sarcocystis are more specific to their rodent intermediate hosts [(see 5)] and that those bank voles examined during this study were free of sarcocysts, they most probably represent two different species. Our opinion supports the finding that S. sebeki of the house mouse was not transmissible to the wood mouse, bank vole or meadow vole (Microtus arvalis) (9).
The Tengmalm’s owl acts as the definitive host for Sarcocystis sp. Wiesner, 1980 [as named by Levine and Ivens (23)] and S. funereus sp. nov., with bank voles and mice, respectively, are its intermediate hosts. However, more information on the former unnamed species is required. The finding of S. halieti and S. lari in the white-tailed sea eagle showed that more than one Sarcocystis species might infect a particular bird of prey species [(see 22)]. As stated by Máca et al. (15), more Tengmalm’s owls and other birds of prey species should be examined to determine the presence of other parasite species or morphospecies of Sarcocystis.
Corticosteroids, such as dexamethasone, have proven to be useful in developing animal models for studying coccidian parasites (24). Dexamethasone induces the depletion of CD4+ T lymphocytes (25) and suppresses T- and natural killer (NK) cell-mediated immunity (26). Similarly, Interferon-gamma (IFN-gamma) gene knockout (KO) mice, often employed to establish Sarcocystis neurona infection in laboratory mice, lack a CD4+ Th1 response. The immunity of gamma-IFN KO mice is either deleted or reduced, allowing the establishment of infection and the development of clinical disease in mice (27). Therefore, we selected dexamethasone immunosuppression to set up Sarcocystis infection. The optimal concentration of dexamethasone in the drinking water was determined to be 30 μg/mL, as a lower concentration likely did not sufficiently suppress the immune response for successful infection establishment.
Máca et al. (15) found S. strixi to cluster with Sarcocystis sp. isolate Af1 (now recognized as S. funereus sp. nov.) and regarded them as distinct species. Interestingly, both have a densely covered wall of the sarcocyst, although S. strixi has knob-like blebs and a thinner wall (< 2 μm). In contrast, S. funereus sp. nov. has longer finger-shaped blebs and a thicker wall (> 2 μm). Additionally, the ends of the sarcocyst (pointed vs. rounded) differ from each other.
Six sequences from wild small mammals were positive for Sarcocystis, but none of them could be considered as S. funereus sp. nov. They probably belong to one or several different species, although more analyses are needed to fully understand whether they represent various species and find their natural definitive hosts. The natural intermediate host of S. funereus sp. nov. is still unknown, but it is very likely that the Eurasian harvest mouse, one of the most common small mammals used as prey by the Tengmalm’s owls (28, 29), plays that important role. Interestingly, after examining several samples of this host (n = 28), no sarcocysts were found. It does not mean that they cannot be parasitize by Sarcocystis, but that the density of intermediate hosts, visiting of sites with contaminated feces, and the climatic seasons are important factors to determine the infection of the definitive hosts, as stated by Hoogenboom and Dijkstra (30). Therefore, further investigations of potential intermediate hosts (wild small mammals) should be done to warrant the identification of the real host. As in other species of Sarcocystis using owls as definitive hosts, mice are essential in the life cycle, as S. espinosai in the northern saw-whet owl and eastern deer mouse [(see 4)].
The present findings show the role of the Tengmalm’s owl in the life cycle of a Sarcocystis species and increase our knowledge of rodent predators as part of the life cycle of parasites. Many more studies are needed to understand the individual relationships between parasites and their intermediate and final hosts. As in other parasite/birds of prey relationships worldwide, we know nothing about the impact of S. funereus sp. nov. on the body condition of fledglings, dispersal, and probability of recruitment to the breeding population and particularly long-term survival of the Tengmalm’s owl. It is important to understand if it is possible to have intestines full of parasites without adversely affect the health of individuals, as in the present study. Thus, more focus and studies are needed to determine how parasites and their hosts interact and influence each other’s lives, either negatively or positively.
Conclusion
This work elucidates the specific identity of the Sarcocystis infecting the Tengmalm’s owl and its experimental intermediate host, although more prey and potential intermediate hosts should be examined to determine their role in the life cycle of the new species. The morphological and molecular characterizations and phylogenetic placement of S. funereus sp. nov. are presented here for the first time and support the erection of the new species.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Ethics statement
The animal study was approved by the Varsinais-Suomen Elinkeino-, Liikenne-ja Ympäristökeskus (permit numbers VARELY/1389/2018 and VARELY/5933/2019), respectively, and welfare laws of the Czech Republic and were approved by the Animal Welfare Committee of the Czech University of Life Sciences Prague (permit number: MSMT-15824/2023-4). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
OM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MK: Conceptualization, Funding acquisition, Investigation, Methodology, Writing – review & editing. IL: Formal analysis, Investigation, Methodology, Supervision, Writing – review & editing. LP: Investigation, Methodology, Writing – review & editing. EK: Investigation, Methodology, Writing – review & editing. DG-S: Conceptualization, Formal analysis, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague provided open access funding. The research project of Tengmalm’s owls in the Kauhava region was financially supported by two grants provided to MK by the Regional Fund of the South Ostrobothnia of the Finnish Cultural Foundation (Business ID 0116947–3 and 10201775).
Acknowledgments
The authors thank all colleagues from the Pathology and Parasitology Department (SVI Prague), especially I. Křičková and K. Sedlák. Thanks are also to T. Bušina, F. Tulis, S. Sovadinová, M. Janouš, and K. Hongisto for their great help during the field-work in the Kauhava study area.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
1. Černá, Z. Multiplication of merozoites of Sarcocystis dispersa Černa, Kolařova et Šulc, 1978 and Sarcocystis cernae Levine, 1977 in the blood stream of the intermediate host. Folia Parasit. (1983) 30:5–8.
2. Černá, Ž, Kolařova, I, and Šulc, P. Contribution to the problem of cyst-producing coccidians. Folia Parasit. (1978) 25:9–16.
3. Munday, BL. A species of Sarcocystis using owls as definitive hosts. J Wildlife Dis. (1977) 13:205–7. doi: 10.7589/0090-3558-13.2.205
4. Espinosa, R, Sterner, M, Blixt, J, and Cawthorn, R. Description of a species of Sarcocystis (Apicomplexa: Sarcocystidae), a parasite of the northern saw-whet owl, Aegolius acadicus, and experimental transmission to deer mice Peromyscus maniculatus. Can J Zool. (1988) 66:2118–21. doi: 10.1139/z88-31
5. Dubey, JP, Calero-Bernal, R, Rosenthal, BM, Speer, CA, and Fayer, R. Sarcocystosis of animals and humans. Boca Raton: CRC Press (2016).
6. Levine, ND, and Tadros, W. Named species and hosts of Sarcocystis (Protozoa: Apicomplexa: Sarcocystidae). Syst Parasit. (1980) 2:41–59. doi: 10.1007/BF00015094
7. Tadros, W, and Laarman, JJ. The tawny owl, Strix aluco as final host of a new species of Sarcocystis with Mus musculus as intermediate host. Trop Geogr Med. (1980) 32:364.
8. Tadros, W, and Laarman, JJ. Sarcocystis and related coccidian parasites: a brief general review, together with a discussion on some biological aspects of their life cycles and a new proposal for their classification. Act Leidens. (1976) 44:1107.
9. Tadros, W, and Laarman, JJ. Current concepts on the biology, evolution and taxonomy of tissue cyst-forming eimeriid coccidia. Adv Parasit. (1982) 20:293–468. doi: 10.1016/S0065-308X(08)60540-0
10. Cawthorn, RJ, Gajadhar, AA, and Brooks, RJ. Description of Sarcocystis rauschorum sp. n. (Protozoa: Sarcocystidae) with experimental cyclic transmission between varying lemmings (Dicrostonyx richardsoni) and snowy owls (Nyctea scandiaca). Can J Zool. (1984) 62:217–25. doi: 10.1139/z84-036
11. Cawthorn, RJ, and Brooks, RJ. Histological observations on precystic merogony and metrocyte formation of Sarcocystis rauschorum (Apicomplexa: Sarcocystidae) in varying lemmings, Dicrostonyx richardsoni. Can J Zool. (1985) 63:2907–12. doi: 10.1139/z85-435
12. Wiesner, J. A new sarcosporidian species of Clethrionomys glareolus inhabiting the owl Aegolius funereus as definitive host. J Protozool. (1980) 27:72A.
13. Kouba, M, Bartoš, L, Bartošová, J, Hongisto, K, and Korpimäki, E. Interactive influences of fluctuations of main food resources and climate change on long-term population decline of Tengmalm’s owls in the boreal forest. Sci Rep. (2020) 10:20429. doi: 10.1038/s41598-020-77531-y
14. Kouba, M, Bartoš, L, Bartošová, J, Hongisto, K, and Korpimäki, E. Long-term trends in the body condition of parents and offspring of Tengmalm’s owls under fluctuating food conditions and climate change. Sci Rep. (2021) 11:18893. doi: 10.1038/s41598-021-98447-1
15. Máca, O, Kouba, M, Korpimäki, E, and González-Solís, D. Molecular identification of Sarcocystis sp. (Apicomplexa, Sarcocystidae) in offspring of Tengmalmʼs owls, Aegolius funereus (Aves, Strigidae). Front Vet Sc. (2021) 8:804096. doi: 10.3389/fvets.2021.804096
16. Kouba, M, Bartoš, L, Tulis, F, Ševčík, M, Sovadinová, S, Bušina, T, et al. Post-fledging survival of Tengmalm’s owl offspring in boreal forests: interactive effects of varying dynamics of main prey and habitat composition. Front Ecol Evol. (2023) 11:1151622. doi: 10.3389/fevo.2023.1151622
17. Kutkienė, L, Prakas, P, Sruoga, A, and Butkauskas, D. The mallard duck (Anas platyrhynchos) as intermediate host for Sarcocystis wobeseri sp. nov. from the barnacle goose (Branta leucopsis). Parasit Res. (2010) 107:879–88. doi: 10.1007/s00436-010-1945-4
18. Katoh, K, Rozewicki, J, and Yamada, KD. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. (2019) 20:1160–6. doi: 10.1093/bib/bbx108
19. Tamura, K, Stecher, G, and Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–7. doi: 10.1093/molbev/msab120
20. Hasegawa, M, Kishino, H, and Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J Mol Evol. (1985) 22:160–74. doi: 10.1007/BF02101694
21. ICZN (International Commission on Zoological Nomenclature). Amendment of articles 8, 9, 10, 21 and 78 of the international code of zoological nomenclature to expand and refine methods of publication. Bull Zool Nom. (2012) 69:1–10. doi: 10.3897/zookeys.219.3994
22. Gjerde, B, Vikøren, T, and Hamnes, IS. Molecular identification of Sarcocystis halieti n. sp., Sarcocystis lari and Sarcocystis truncata in the intestine of a white-tailed sea eagle (Haliaeetus albicilla) in Norway. Int J Parasitol. (2018) 7:1–11. doi: 10.1016/j.ijppaw.2017.12.001
23. Levine, ND, and Ivens, V. The coccidian parasites (Protozoa, Sporozoa) of rodents. Ill Biol Mongr. (1965) 33:1–365. doi: 10.5962/bhl.title.50242
24. Cutler, TJ, MacKay, RJ, Ginn, PE, Gillis, K, Tanhauser, SM, LeRay, EV, et al. Immunoconversion against Sarcocystis neurona in normal and dexamethasone-treated horses challenged with S. Neurona sporocysts. Vet Parasitol. (2001) 95:197–210. doi: 10.1016/s0304-4017(00)00420-9
25. Giles, AJ, Hutchinson, MKND, Sonnemann, HM, Jung, J, Fecci, PE, Ratnam, NM, et al. Dexamethasone-induced immunosuppression: mechanisms and implications for immunotherapy. J Immunother Cancer. (2018) 6:51. doi: 10.1186/s40425-018-0371-5
26. Chen, L, Jondal, M, and Yakimchuk, K. Regulatory effects of dexamethasone on NK and T cell immunity. Inflammopharmacology. (2018) 26:1331–8. doi: 10.1007/s10787-017-0418-0
27. Witonsky, SG, Gogal, RM Jr, Duncan, RB, and Lindsay, DS. Immunopathologic effects associated with Sarcocystis neurona–infected interferon-gamma knockout mice. J Parasitol. (2003) 89:932–40. doi: 10.1645/GE-72R
28. Korpimäki, E. Diet of breeding Tengmalm’s owls Aegolius funereus: long term changes and year to year variation under cyclic food conditions. Ornis Fennica. (1988) 65:21–30.
29. Korpimäki, E, and Hakkarainen, H. The boreal owl: ecology, behaviour and conservation of a forest-dwelling predator. Cambridge: Cambridge University Press (2012).
Keywords: rodents, birds of prey, Europe, sarcocysts, morphology, molecular characterization, oocysts and sporocysts, phylogeny
Citation: Máca O, Kouba M, Langrová I, Panská L, Korpimäki E and González-Solís D (2024) The Tengmalm’s owl Aegolius funereus (Aves, Strigidae) as the definitive host of Sarcocystis funereus sp. nov. (Apicomplexa). Front. Vet. Sci. 11:1356549. doi: 10.3389/fvets.2024.1356549
Edited by:
Alireza Sazmand, Bu-Ali Sina University, IranReviewed by:
Petras Prakas, Nature Research Centre, LithuaniaVikrant Sudan, Guru Angad Dev Veterinary and Animal Sciences University, India
Copyright © 2024 Máca, Kouba, Langrová, Panská, Korpimäki and González-Solís. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David González-Solís, ZGdvbnphbGVAZWNvc3VyLm14
†ORCID: Ondřej Máca https://orcid.org/0000-0002-6311-0851
Marek Kouba https://orcid.org/0000-0003-2262-5733
Iva Langrová https://orcid.org/0000-0001-8731-7470
Lucie Panská https://orcid.org/0000-0002-9746-1633
Erkki Korpimäki https://orcid.org/0000-0001-7596-1955
David González-Solís https://orcid.org/0000-0001-7765-2865