 Getachew Abraham
Getachew Abraham Yisehak Kechero1
Yisehak Kechero1 Dereje Andualem
Dereje Andualem- 1Department of Animal Science, College of Agricultural Sciences, Arba-Minch University, Arba Minch, Ethiopia
- 2Department of Animal Science, College of Agriculture and Natural Resources, Dilla University, Dilla, Ethiopia
Introduction: Developing a technology for fodder trees and shrubs tailored to farmers’ preferences is best done with their input, perceptions, and interests in mind.
Objective: The research aimed to determine farmer preferences for indigenous legumes, fodder trees, and shrubs (ILFTS) and to examine the relationship between feed valuation and scientific parameters.
Methods: A focus group discussion (FGD) was conducted with 10 farmers in each agroecological zone to determine the benchmarks for the preference ratings. The respondent farmers used the preference score sheet to rate all ILFTS on an individual basis. Twenty farmers with extensive experience in ILFTS took part in the preference score rating of each plant species in each agroecosystems. Dry matter (DM), organic matter (OM), ash, crude protein (CP), neutral detergent fiber (NDF), acid detergent fiber (ADF), acid detergent lignin (ADL), metabolizable energy (ME) and condensed tannin (CT) content of the samples were determined. The standard two-stage in vitro Tilley and Terry method was used to measure the in vitro dry matter digestibility (IVDMD) and in vitro organic matter digestibility (IVOMD) of samples. Digestible organic matter in dry matter (DOMD) and ME values were estimated using standard models. Analysis of variance (ANOVA) was used to analyze the variation among the species in agroecosystems. Tukey HSD tests were used for mean separation.
Results and discussions: Farmers evaluated the ILFTS using a variety of parameters, according to the study (feed value, growth rate, biomass output, compatibility, and multifunctionality). The farmers’ ILFTS preference score on the evaluation criteria differed considerably (p<0.05) with species in agroecosystems. The CP, ash, and ME values of ILFTS in the study were moderate to high although exhibited a wide variation among the species in agroecosystems. The CP content was above the minimum requirement (8%) to support the normal function of rumen microorganisms. Moreover, CP content exhibited a positive significant correlation with IVDMD, IVOMD, and DOMD, unlike CT and ADL which exhibited a negative significant correlation. Conversely, the DM, OM, CP, IVDMD, IVOMD, DOMD, and ME were shown a positive significant correlation with farmers’ feed value preference score, unlike the ADL and CT which exhibited a negative significant correlation.
Conclusions: Farmers’ indigenous knowledge of feed value is therefore relevant for judging the nutritive value of the ILFTS and could complement the scientific indicators.
Introduction
Fodder trees, and shrubs have been an essential source of forage for ruminants to complement the critical dry period feed deficit in the tropics (1). In addition to flourishing with a deep root system capable of absorbing water far from the surface, they produce considerable biomass of leaves, twigs, fruits, and pods which can bridge the feed supply gap commonly observed during dry periods (2, 3). Fodder trees and shrubs have high nutrient content and digestibility, although this varies by species and season (4, 5). In particular, the crude protein (CP) content of fodder trees is above the minimum requirement for the normal microbial function of the rumen, so it is usually recommended to supplement poor-quality fiber-based diets (6, 7). Feeding ruminants’ legumes, fodder trees, and shrubs improve the intake and digestibility of low-fiber-based diets by increasing the activity of rumen microorganisms via improving the nitrogen supply which is necessary for their proliferation (8). However, deterring mechanisms related to phenolic compounds, especially high condensed tannin (CT) content, which reduce feed intake, nutrient digestibility, and nitrogen retention, limit their potential as feed resources for herbivores (9). Condensed tannin in low to medium concentrations (below 5 g/kg dry matter) has been found to benefit ruminant production by improving rumen bypass protein and carbohydrates, preventing bloat and helminthiasis, and reducing greenhouse gas emissions (9).
Indigenous fodder trees and shrubs are adaptable to the local environment due to their pest resistance and drought tolerance. Furthermore, they are preferred to exotic browse trees due to their palatability, high nutrient value, biomass yield, readily available planting materials, and local community appreciation (10). Currently, indigenous fodder trees and shrubs have been receiving research attention in Ethiopia and other tropical countries. However, most studies ignore the farmers’ knowledge and rely on on-station agronomic and feeding trials to compare biomass yield and nutritive value of a specific species with various management practices (11, 12). This approach, however, has an impact on the spread of emerging technologies involving trees and shrubs as forage plants. The uptake of technology is determined by the farmers’ knowledge, perceptions, and attitudes, according to Meijer et al. (13). Farmers could have an immense contribution to research since their knowledge and preferences are crucial as potential users of the upcoming technologies (14). Farmers’ perceptions of trees are based on their felt needs, prior experiences, and expectations, which may or may not correspond to scientific reality (13). Boogaard et al. (15) indicated that farmers valued fodder trees based on their knowledge, experience, values, and interests.
Many countries around the world, particularly in tropical and semi-humid regions, have not dealt with studies on farmers’ preferences, nutritional quality, and their correlation with fodder trees and shrubs. However, involving farmers’ knowledge, perceptions, and interests in appraising fodder trees and shrubs and analyzing their correlation with scientifically proven methods could save the time; energy, and cost to produce tailored technologies that effectively address farmers’ problems. Thus, this research was conducted to determine nutritional quality, farmer preferences, and the relationship between local feed valuation and laboratory outcomes of indigenous legume fodder trees and shrubs (ILFTS) in a sub-humid environment.
Materials and methods
Description of the study area
The research was conducted in Gamo zone of southern Ethiopia, which is one of the sub-humid regions of the country. The zone lies between 5′57° – 6′71° North, latitude, and 36°37′–37°98′ East, longitude. The elevation in the Gamo zone ranged between 501 and 4,207 m above sea level, which is the reason for a varied climate and agroecosystems that produced wide biodiversity. Gamo zone is characterized by bimodal rainfall with the mean annual rainfall ranging from 801 to 2,000 mm and the annual mean temperature range from 10.1 to 27.5°C. The terrain has an undulating feature that favors the existence of different agro-climatic zones in close proximity ranging from dry lowlands to wet highlands. Mixed crop-livestock production is a prominent farming system where fruit and vegetable are the dominant crops in the lowland unlike in the highland where Ensete ventricosum predominates (16). Agroforestry is the common practice where trees are an integral component of the farming system that complements the function of land uses and enhances productivity (2) (see Figure 1).
Figure 1. The map of the study districts in Gamo landscape.
Farmers’ preference scoring and sample collection of ILFTS
Following a purposive sampling procedure, 60 experienced and acquainted farmers (20 in each agro-ecological zone) that were participating in the management and utilization of ILFTS were chosen for preference scoring. Focus group discussions (FGD) with 10 farmers were held in each agroecological zone to explore the desired tree characteristics and perceived benefits of ILFTS, which were the foundations to set the benchmarks for preference scoring. As a result, feed value, growth rate, biomass yield, compatibility, and multifunctionality were established as benchmarks for preference scoring of ILFTS. The nutritional preference score was determined based on criteria such as palatability, improvement of body condition, growth, and milk production, improvement of straw diet intake, and improvement in animal health, while the preference score for growth and regrowth potential was determined by criteria such as growth rate after establishment and re-growth potential after frequent cutting or looping. Farmer’s preference for compatibility was primarily based on the absence of crop competition for available soil nutrients and moisture, which improves soil fertility and improves the growth of annual and perennial crops below the canopy. Timber, poles, and other local constructions, as well as fuel wood, fences, medicinal values, shade trees, honey sources, soil stabilization, and farm implements, were incorporated to create multifunctionality indices. The rating of the ILFTS species was done using a preference score sheet. In each agro-ecological zone, preference scoring was carried out for ILFTS species on a point scale ranging from 1 (not preferred) to 4 (highly preferred) (17). Respondent farmers completed the ranking exercise on an individual basis. In each agro-ecological zone, samples of ILFTS, leaves, fruits, and pods were collected and were chemically analyzed. Five to ten individual plants per species were sampled and pooled to obtain a representative sample for each species in each agroecological zone. The samples were air-dried before being taken to the lab for testing.
Chemical analyses of indigenous legume fodder trees and shrubs
The leaf (all ILFTS), pod (Acacia tortilis), and fruits (Acacia albida) samples were collected from the lowland [1,000–1,500 m above sea level (masl)], midland (1,500–2,300 masl) and highland (2,400–3,000 masl) in May and June. The dry matter (DM) content was determined by oven drying the feed samples at 55°C for 72 h for the constant weight (18). Oven-dried feed samples were ground using a Wiley mill to pass through a 1 mm sieve for chemical analyses. Contents of DM, total ash, and crude protein (CP) were analyzed following the standard methods of AOAC (18). The method of Van Soest et al. (19) was used to determine neutral detergent fiber (NDF). Acid detergent fiber (ADF) was determined following the method (20). Accordingly, the NDF and ADF analyses were followed sequentially. The residual ash was included in the NDF and ADF values. By solubilizing cellulose with 72% H2SO4, lignin (ADL) was determined (20). The difference between the percentages of NDF and ADF was used to calculate the hemicellulose (% HC). Total condensed tannins (CT) were determined using a butanol-HCl reagent and 2 percent ferric ammonium sulfate in a 2 N HCl catalyst (21). All chemical analyses were carried out in duplicate.
In vitro dry matter digestibility potential of ILFTS
The two-stage technique was implemented to determine the in vitro dry matter digestibility (IVDMD) of leaf fruit and pod samples (22) as modified by Van Soest and Robertson (20). The rumen liquor was collected from three rumen cannulated steers before the morning feeding that was fed natural pasture hay (5%-6%CP) ad libitum supplemented with about 2 kg of concentrate (69% wheat bran, 30% noug seed cake and 1% salt) mixture per steer/day. The liquor from three steers was mixed on a volume basis and filtered through cheesecloth. The incubation inoculum was prepared by diluting the rumen liquor with a buffer solution (NaHCO3 + Na2HPO4 + KCl + NaCl + MgSO4. 7H2O + CaCl2.2H2O) (1:2 v/v) in a 1:4 (v/v) ratio (22). The mixed inoculum was stirred in a water bath at 39°C with purging CO2 until its use (10–15 min later). About 0.5 g (1 mm ground) of each sample was placed into 50 mL sterile tubes, and 20 mL of the incubation inoculum was added. The tube was stoppered with a Bunsen valve and incubated for 48 h at 39°C. Tubes were gently swirled by hand every 8 h. Each sample was incubated in three replicates. At the end of 48 h of the incubation period, the tube contents were acidified using 6 M HCL to reach a final pH of 1.3–1.5. After the foam subsided, the pepsin powder was added to the final concentration of 0.2% (w/v). Then, the sample was reincubated for 48 h again. The undigested portion of the sample (residue) was transferred into the crucible, and the liquid was filtered out via a sacking machine. The pellets were dried in a forced-air oven at 105°C for 24 h to determine the residual DM weight. Three blanks were included in each run. Then, to determine the ash content, the residues were kept at 550°C for 8 h to estimate ash. The in vitro DM and OM digestibility was determined as the DM and OM which disappeared from the initial weight added into the tube using the following equation.
Digestible organic matter in total dry matter (DOMD) was calculated as 0.95 IVDMD (%) − 2 (23). Metabolizable energy (ME) was estimated from DOMD (24) using the equation indicated below:
where: DOMD = Digestible Organic Matter in the Dry matter.
Statistical analysis
Data on the farmers’ assessment of ILFTS, and feed value preference score in agroecosystems, and the chemical composition including total CT, IVDMD, and ME values among trees shrubs, and fruits/pods in the lowland were subjected to the analysis of variance (ANOVA). The farmers’ preference score between trees and shrubs was analyzed using an independent t-test. Tukey HSD tests were used for mean separation. Mean differences were declared at p < 0.05. The analyses were performed using SPSS statistical software version 21 following the model indicated below:
where,
: Response variable; μ: overall mean effect;: the effect of plant species; dj, the jth effect of agroecology and is the random error.
Pearson correlation coefficient was done to analyze the relationships that exist, if any, between farmers’ feed value preference scores of ILFTS with the relative evaluation derived from laboratory-based indicators of feed quality.
Results
Socioeconomic characteristics’ of farmers
Socioeconomic characteristics’ of the respondents’ exhibited a wide variation among agroecosystems (Table 1). The sex and marital status of the respondents indicated that the majority were male (91.7%) of which 95% were married and the remaining were widowed. There was a variation in age category, with the majority of the respondents aged 41–50 years, followed by 31–40 years, with only 1.67% of the respondents below 30 years of age. The purposive sampling procedure intended to select the most knowledgeable respondents might favor the older and male groups. The educational level of the respondents varied widely, with the majority attending grades 5–8 (28.3%) followed by basic education (25%), where those attending above grade 12 were the least (3.33%). The limited education facilities in the area, which is a common occurrence in most rural parts of the country, might explain the low educational level of the respondents in the study. The land holdings of the majority of the respondents were 0.51–1 ha (36.67%) and 0.26–0.5 ha (30%), respectively. However, 10% of the respondents had less than 0.25 ha.
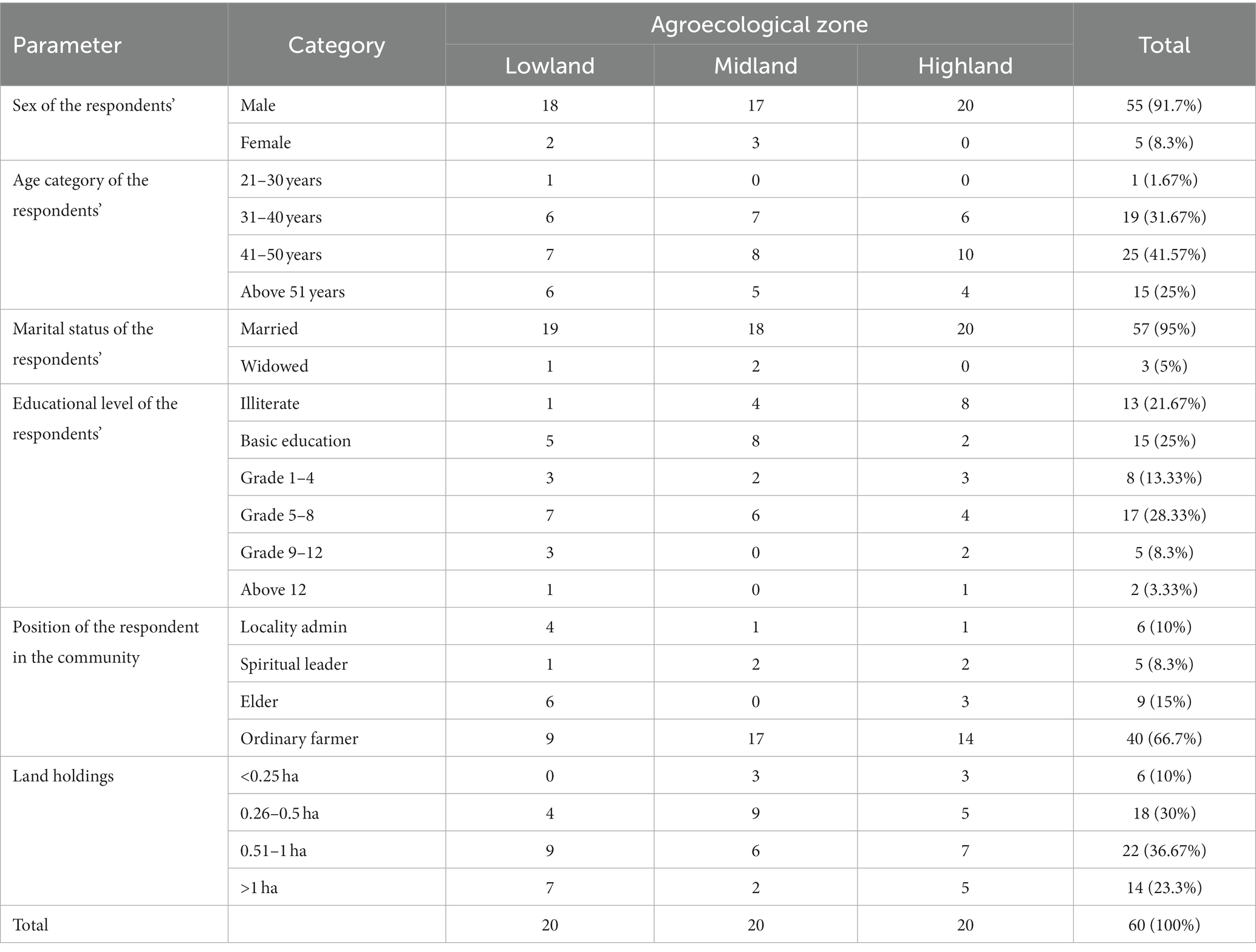
Table 1. The socioeconomic characteristics’ of the respondents’ (N = 60).
Farmers’ preference of ILFTS
The study indicated that farmers used multiple criteria to evaluate the ILFTS and the farmers’ feed value preference score exhibited significant differences between species in all agroecosystems and between shrubs and trees in the lowlands (Tables 2, 3). Among the trees Acacia seyal and Acacia albida (A.albida) excelled in feed value preference score however Acacia nilotica (A.nilotica) scored the least in the lowland. Acacia brevispica (A.brevispica) and Acacia mellifera (A.mellifera) excelled among the shrubs in feed value preference score in the lowland whereas Dichrostachys cinerea scored the least. In both midland and highland agroecosystems, Albizia schimperiana (A.schimperiana) and Erythrina brucei (E.brucei) showed significantly high feed value preference scores.
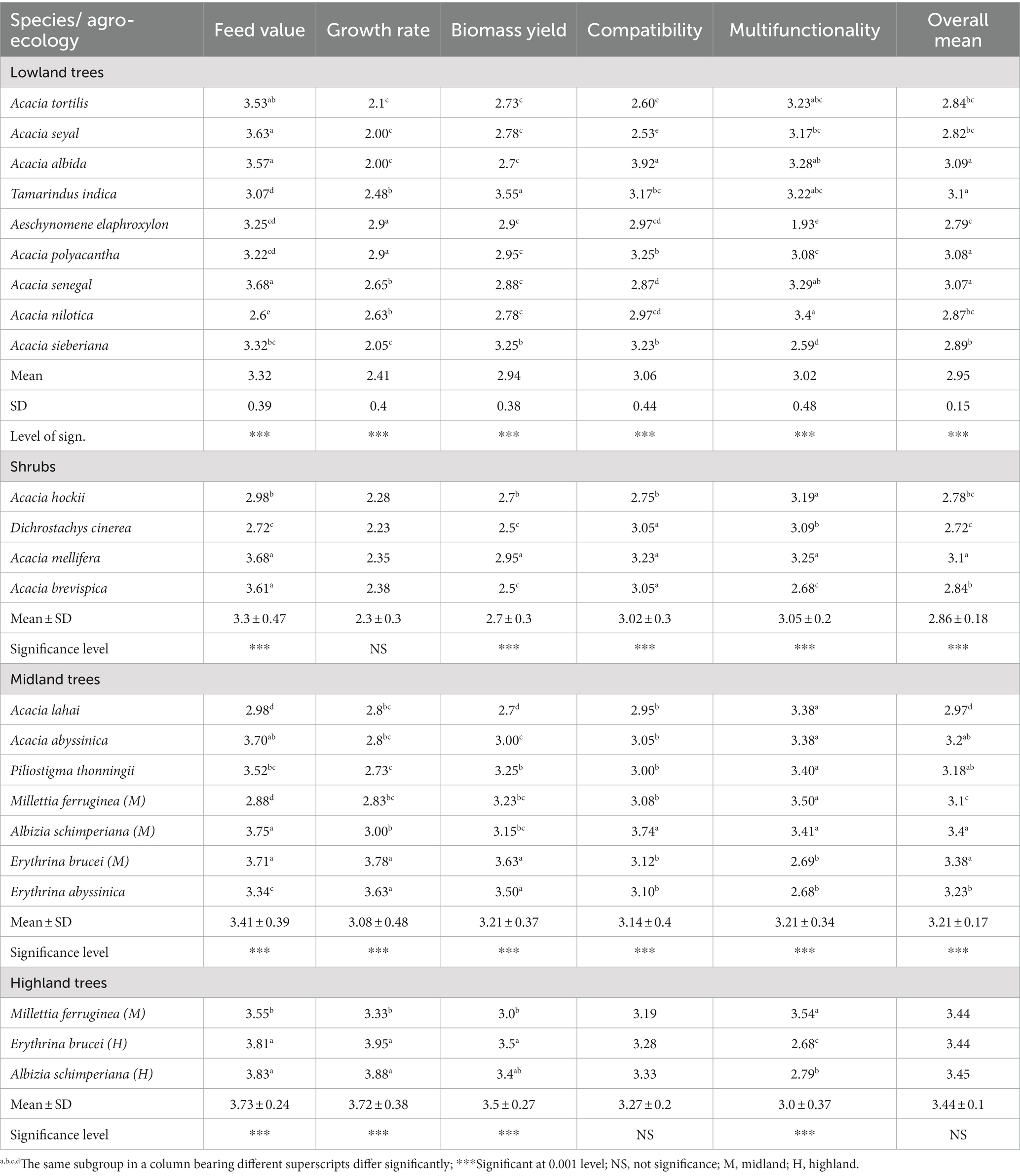
Table 2. Farmer preference score of ILFTS with species and evaluation parameters (N = 60).
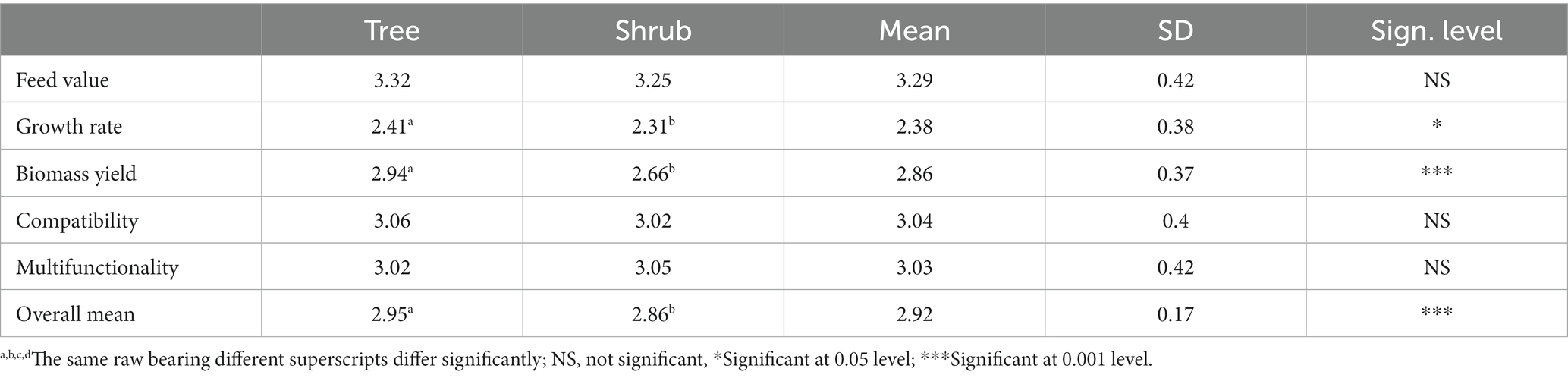
Table 3. Farmers’ preference score between trees and shrubs of ILFTS in the lowland.
Likewise, among the parameters of tree characteristics, the farmers’ preference score for growth rate among the shrubs in the lowland, and compatibility and multifunctionality in the highland exhibited non-significant differences (p < 0.05) with species. The study, however, found a significant difference between shrubs and trees in the lowland (Table 3). Among the trees Aeschynomene elaphroxylon (A.elaphroxylon) and Acacia polyacantha (A.polyacantha) in the lowlands, E.brucei and Erythrina abyssinica (E.abyssinica) in the midland, and E.brucei and A.schimperiana in the highland excelled in growth rate.
Similarly, Tamarindus indica (T.indica) among the trees and A.mellifera among the shrubs in the lowland, E.brucei and E.abyssinica in the midland, and Millettia ferruginea (M.ferruginea) in the highlands revealed the highest significant (p < 0.05) score for biomass yield, whereas A.albida (3.92) among the trees in the lowland and A.schimperiana (3.74) midland scored the highest significant compatibility score although no significant difference was observed among the highland trees and between trees and shrubs in the lowland. According to respondents, A.albida (lowland) and A.schimperiana (midland) improved soil fertility and stability as well as increased crop yield and were therefore preferred in the farmland. Of course, all ILFTS species are likely to enhance crop yield via improved soil fertility and stability due to their ability to fix atmospheric nitrogen (2, 25). Likewise, A.nilotica and A. mellifera among the trees and shrubs, respectively, in the lowland and all midland species except Erythrina species and M. ferruginea (highland) achieved significantly high scores (p < 0.05) for multifunctionality, as the study revealed. However, there is no significant difference between trees and shrubs in multifunctionality (Table 3). ILFTS have been used for multiple functions such as local construction, firewood, charcoal, tool handlers, local furniture, traditional medicine, bee forage, and fencing were among those mentioned by the respondents. Moreover, the overall mean score of the ILFTS revealed significant differences (p < 0.05) with species among trees and shrubs in the lowland and midland. Four ILFTS species, namely A.albida, T.indica, A.polyacantha, and Acacia senegal (A.senegal) among the trees and A.mellifera among the shrubs in the lowland, and two species namely A.schimperiana and E.brucei in the midland exhibited the highest score for overall mean, even though the highland species exhibited no significant difference. Trees excelled shrubs in growth rate, biomass yield, and overall mean in the lowland among farmers’ preference score parameters (Table 3).
Nutritional value parameters
Wide variations in nutritive values were found among the ILFTS species; however, there was no significant variation among trees, shrubs and fruit/pods in the lowlands (Tables 4, 5). As an example, the DM content of the ILFTS species ranged from 838.3 to 948.4 g/kg with A. brevispica leaf exhibiting the highest content followed by E.brucei leaf (M) (midland) and M. ferruginea leaf (H) (highland) respectively; Acacia hockii leaf (A.hockii) showed the lowest. The difference in ash content among the ILFTS was more than fivefold ranging from 25.9 to 134.2 g/kg DM, where P. thonningii leaf and A.sieberiana leaf recorded the highest and lowest among the trees respectively, whereas A. hockii leaf and D. cinerea leaf had the highest and lowest values among the shrubs. A similar tendency was observed in the variation of CP content among the ILFTS which spanned from 81.8 to 314 g/kg DM where E.brucei leaf (M) (midland) exhibited the top, succeeded by the highland M.ferruginea leaf (H) and E.brucei leaf (H) in that order, although A.albida fruit revealed the least. A. mellifera leaf and D. cinerea leaf exhibited the highest and lowest value of CP among the shrubs.
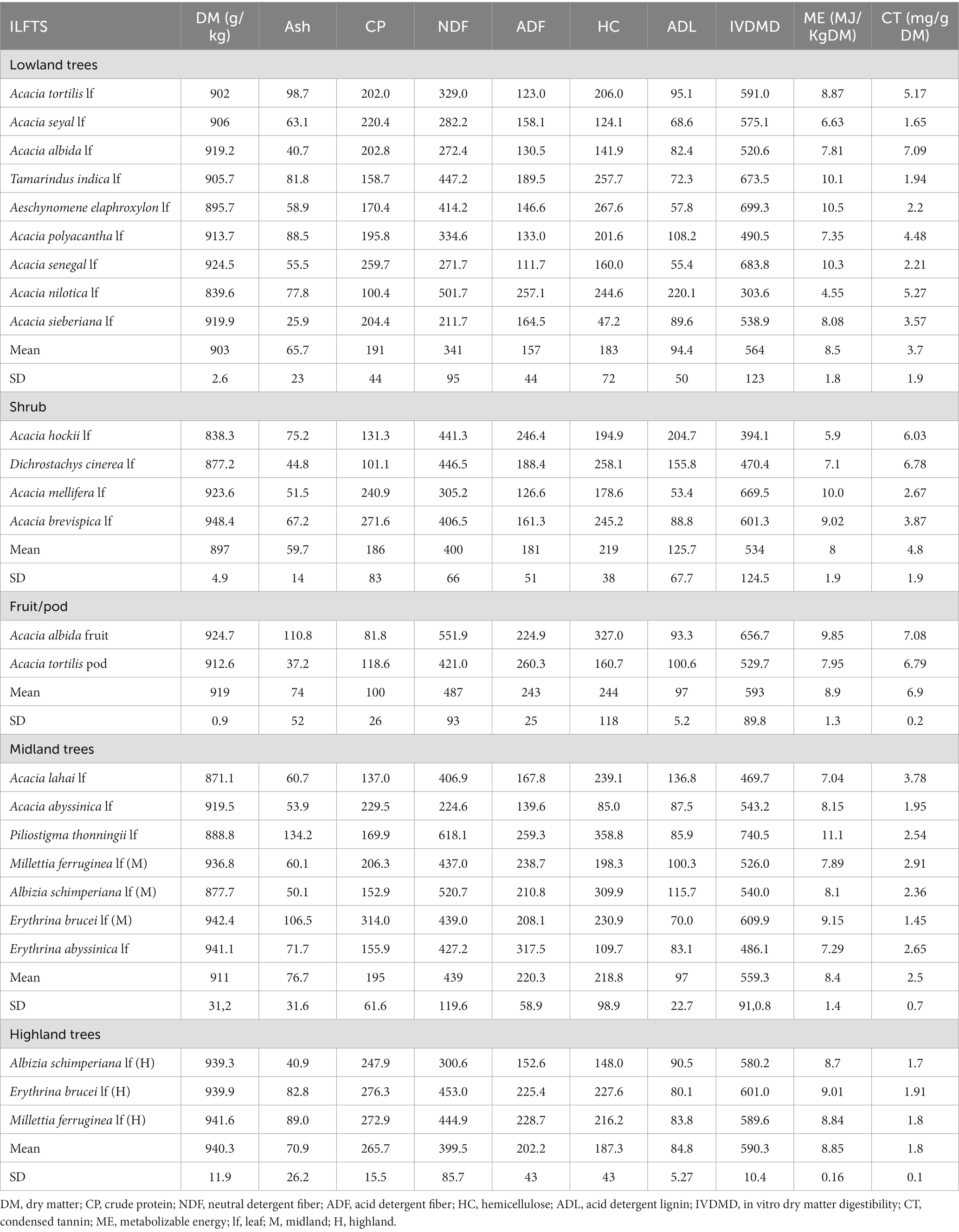
Table 4. Chemical composition and IVDMD of the leaves, fruits, and pods of ILFTS (g/kg DM) by species and agroecosystems.
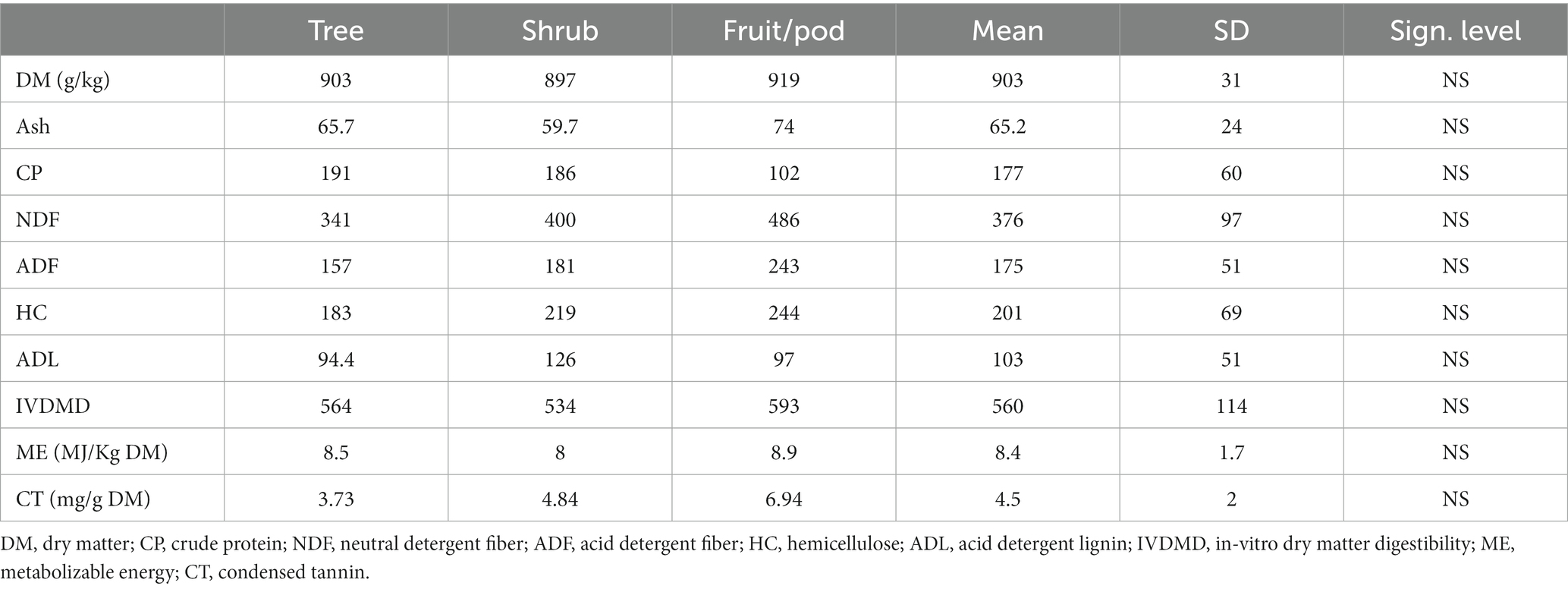
Table 5. The nutritional quality among tree, shrub, and fruit/pod of ILFTS (g/kg DM) in the lowland.
The HC and ADL components had the most variance, fluctuating from 47.2 to 327 and 53.4 to 220.1 g/kg of DM, respectively, tracked by ADF and NDF. The ILFTS NDF value ranged from 300.6 to 618.1 g/kg DM, with P. thonningii leaf revealing the most, accompanied by A. albida fruit and midland A. schimperiana leaf (M), but A. lahai leaf revealing the least. The ILFTS ADF value ranged from 111.7 to 317.5, with E. abyssinica leaf having the greatest value, preceded by A. tortilis pod and Piliostigma thonningii (P. thonningii) leaf, and A. senegal leaf having the lowest. P. thonningii leaf had the highest HC, followed by A. schimperiana leaf (M) (midland) and A. elaphroxylon leaf; however, A. abyssinica leaf (midland) had the least HC. A. nilotica leaf has the highest ADL value, followed by A. hockii leaf and Acacia lahai (A. lahai) leaf, respectively, with A. mellifera leaf having the lowest.
The variation of IVDMD and ME was more than twofold, ranging from 303.6 to 740.5 g/kg DM and 4.55 to 10.5 MJ/kg DM, respectively. In terms of decreasing order of IVDMD, the ranking for the leaves of five ILFTS species was P.thonningii > A.elaphroxylon > A. senegal > T.indica > A.mellifera leaf respectively, however, A.nilotica leaf showed the least. In terms of ME, P.thonningii displayed the highest rate followed by A.elaphroxylon leaf and T.indica leaf, respectively, while A. nilotica leaf displayed the lowest level. Likewise the variation in CT content of ILFTS species forages was more than fourfold which ranged from 1.45 mg/g DM to 7.09 mg/g DM with Acacia albida lf and Acacia albida pod exhibiting the highest content followed by Acacia tortilis pod. However, Erythrina brucei lf (midland) exhibited the least.
The correlation among nutrients and farmers feed value scores
The nutritive parameters of the ILFTS, such as CP (r = 0.768), IVDMD (r = 0.6), IVOMD (r = 0.565), ME (r = 0.6), and DOMD (r = 0.6), had a positive significant correlation with the farmers feed value score, unlike the ADL (r = −0.702) and CT (r = −0.543) which exhibited negative significant correlation (Table 6). Fiber components such as NDF (r = −0.314), ADF (r = −0.332), and HC (r = −0.19) had a negative non-significant connection with the farmers’ feed value score. However, the largest positive and negative Pearson correlation coefficients with farmers’ feed value assessments were found in CP (r = 0.768) and ADL (r = − 0.702) respectively.
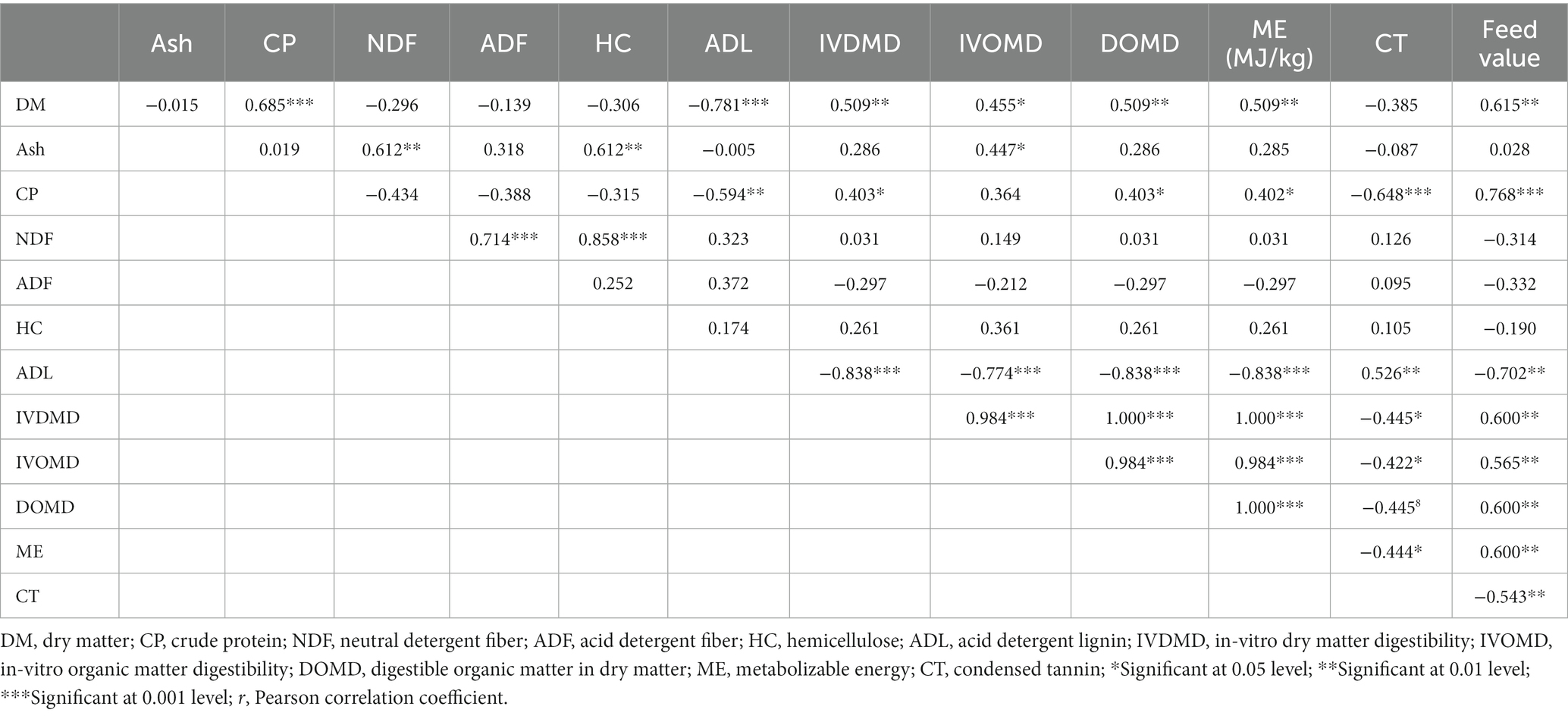
Table 6. Pearson correlation between the nutrients and indigenous knowledge of feed value of the ILFTS in Gamo landscape.
The CP of the ILFTS had a substantial positive connection with DM (r = 0.685), OM (r = 0.507), IVDMD (r = 0.403), DOMD (r = 0.403), and ME (r = 0.402), but a significant negative correlation with NDF (r = −0.434). The IVDMD had a substantial positive connection with DM (r = 0.509) and CP (r = 0.403), whereas the ADL had a significant negative correlation (r = −0.838). The IVOMD showed a positive significant link with DM (r = 0.455) and ash, in contrast to HC (r = 0.361) and ADL (r = −0.838), which showed a negative significant correlation. The ME showed a positive significant association with DM (r = 0.509), IVDMD (r = 1.0), IVOMD (r = 0.984), and DOMD (r = 1.0), but a negative significant link with ADL (r = −0.702).
The ADL had negative significant correlation with DM (r = −0.781), OM (r = −0.589), IVDMD (r = −0.838), IVOMD (r = −0.774), and DOMD (r = −0.984); however, it revealed positive non-significant correlation with the fiber components such as NDF (r = 0.323), ADF (r = 0.372) and HC (0.174).
The CT had a negative non-significant correlation with DM (r = −0.385), Ash (r = −0.087), and OM (r = −0.236) unlike the CP (r = −0.648) which revealed a negative significant correlation. CT exhibited negative significant correlation with IVDMD (r = −0.445), IVOMD (r = −0.422), DOMD (r = −0.445), and ME (r = −0.444) unlike the ADL (r = 0.526) which displayed positive significant correlation, though it showed positive non-significant correlation with NDF (r = 0.126), ADF (r = 0.095) and HC (r = 0.105).
Discussions
Farmers’ preference of ILFTS
The study indicated that farmers used multiple criteria such as feed value, multifunctionality, growth rate, biomass yield, and compatibility to evaluate the ILFTS which marked the farmers’ preference measures for the ILFTS as multifaceted (Table 2). This was due to the multiple functions of ILFTS as substantiated by several studies in the tropics and subtropics (2, 26). In agreement with the current study, various studies in Ethiopia have unveiled multiple criteria employed by farmers to evaluate fodder trees (4, 27); however, some emphasized certain criteria. For instance, availability and feed value were the major criteria to evaluate fodder trees in northwestern Ethiopia (4) and southern Ethiopia (28) due to critical feed shortages during the dry season. Some of the evaluation criteria used for the farmers’ preferential treatment of the ILFTS in the study are similar to those employed in other studies conducted in Ethiopia (4, 12), possibly because of the similarities in sociocultural practices.
The preference score was the farmer’s relative appraisal of one species over another on a given parameter. Distinct species of ILFTS inhabited the lowlands and highlands, even though the midland featured the common species of both agroecological zones, according to the study. The ILFTS species analyzed in the lowland, midland, and highland of the study were 13, 7, and 3, respectively, and hence the higher the species diversity across the agroecological zones, the higher the variation in the perception which in turn affects the preference rating and vice versa. Various research conducted in Ethiopia, in concurrence with the current study, showed that farmers’ choices for fodder trees varied significantly with species and evaluation criteria in agroecosystems (4, 12).
The superior feed value preference score of the respondents for the ILFTS in the lowland (A. tortilis, A.seyal, A.albida, A.senegal, A.mellifera, and A.brevispica), midland and highland (E.brucei and A.schimperiana) were probably due to the high perceived benefit of feeding them to ruminants, which enhances the quantity and quality of the diet as the study unveiled. Fodder trees are rich in CP, energy, and minerals (29, 30), adding to ruminants fed low-quality fiber-based diets enhanced their performance (8, 31). The higher performance of ruminants fed fodder trees and shrubs could potentially be attributed to reduced gastrointestinal distress and effective protein and energy use, because smaller quantities of secondary metabolites, particularly condensed tannins, are susceptible to such activities (reducing methane emission and enhancing bypass protein). The deleterious effect of CT on ruminant nutrition was substantiated by the negative significant correlation between CT and farmers’ feed value scores as revealed in the study. Various studies have discovered that fodder plants and shrubs contain anthelmintic (32, 33) and anti-methane emission constituents (34, 35). A high feed value score was observed for the Acacia species in Lay-Armachuho as well as the Millettia and Acacia species in Sidama (27) which was in contrast with the current study.
A.polyacantha (lowland), E.brucei and E.abyssinica (midland), and E.brucei and A.schimperiana (highland) excelled in growth rate due to their easily establishing nature and high growth potential that allowed reaching harvests (exploitation) early, make them valuable to farmers. Likewise, the ample and sustainable leaf yield of T.indica (lowland), E.brucei and E.abyssinica (midland), and E.brucei and A.schimperiana (highland) might justify their preeminence for biomass yield score. However, in a study conducted in southwestern Ethiopia (12), E. abyssinica had a low growth rate score, which is in contrary to the current study.
The ability of A. albida (lowland), A.schimperiana (midland), and highland species to enrich the soil and boost crop output growing underneath may explain why they are more compatible. Mekoya et al. (27) revealed the supremacy of the Acacia species for compatibility score in northern regions and southern semi-humid lands, which substantiate the current study, even though A. abyssynica in southwestern Ethiopia (12) exhibited significantly lower compatibility scores. In the current study, A. nilotica (lowland) and M. ferruginea (midland and highland) excelled for multifunctionality, likely due to their multiple functions such as poles or posts for construction, firewood, charcoal, farm implements, bee forage, and shade, in contrast to Acacia and Millettia species in northern dry midlands and Sidama (27), which scored slightly lower.
Nutritional value parameters
The study found that there is a wide difference in nutritive value among the browse species, which is most likely due to the species’ inherent nature. Genomic and environmental factors influence the structure and composition of polysaccharides, such as lignin, cellulose, and hemicellulose resulting in differences in cell wall composition among forage species (36–38). The cell wall is comprised of 23%–90% of the plant tissue (37) hence its composition substantially affects the nutritive quality of forage. According to Grant et al. (39) and Ray et al. (40) regional and inter-annual climate variability causes fluctuations in forage nutritive values.
The ash values of most of the ILFTS species reported in the study are within the ranges reported for most native African browse species (10, 30). The ash values of T.indica, A.nilotica, and A. tortilis in the study were greater than the values observed in eastern Ethiopia (41). Likewise, the ash values reported for A.polyacantha in Tanzania are higher than the values found in the current study, yet A.tortilis, A.nilotica, and Dichrostachys species recorded lower amounts (42). The variation of the ash value of the ILFTS in the study compared to other studies is probably due to the soil fertility disparity, the season of harvest, and the harvesting stage of the fodder trees and shrubs (29, 36). Ash value is a potential indicator of the mineral content of forages however some minerals are volatile in nature hence further analysis is necessary to determine their actual value in the forages of ILFTS.
The CP values of most of the ILFTS species reported in the study are within the ranges found for most native African browse tree and shrub species (30, 43). While the CP value of T.indica leaf reported in the present study was similar to that observed in eastern Ethiopia; unlike, A.tortilis leaf and A.nilotica leaf which were higher and lower, respectively, than those in the current study (41). The study conducted in western Ethiopia reported 178 and 240 g/kg DM of CP for A.abyssinica leaf and E.abyssyinica leaf, respectively (12) which is lower and higher than the values found in the current study. The study revealed the minimum CP content of the ILFTS in the study was 81.8 g/kg DM, which was above the CP requirement [70 g/kg DM (44)] for normal rumen microbial function in ruminant livestock. The study substantiates that fodder trees and shrubs have the potential to complement the CP and mineral deficiency commonly observed in poor-quality pastures and crop residues, particularly during dry periods (45, 46).
The IVDMD values of most of the ILFTS reported in the study are within the range reported for native fodder trees and shrub species in Ethiopia and other tropical countries (10, 47). The IVDMD of P.thonningii leaf is the highest in the study, suggesting that it contains the maximum available nutrients among the ILFTS. The fiber of fodder trees and shrubs is more digestible than grass due to its less lignin content (5).
The NDF, ADF, and ADL values of some of the ILFTS species reported in the study are within the range reported for native fodder tree and shrub species in Ethiopia and other African countries (30, 41). For instance, the lignin value reported for T.indica leaf and A.tortilis leaf in the study was less than the value found in eastern Ethiopia (41) however the value for A.nilotica leaf was greater in the current study. The NDF and ADF values reported for A.abyssinica leaf and E.abyssyinica leaf in the study were lower and higher, respectively than the values found in western Ethiopia (12), unlike their ADL values. The variation in the fiber content of fodder trees and shrubs among studies is probably due to the harvesting season and stage of maturity of the browse trees (29). P.thonningii leaf recorded the highest NDF value in the study implying likely to affect feed intake unlike A.nilotica leaf which showed relatively high NDF (520.7 g/kg DM) yet its ME value was the least due to its high ADL value. ADL is most likely to determine the nutritional value of plant fiber by interfering with the digestion of cell-wall polysaccharides by acting as a physical barrier. Li (38), Moore and Jung (48), and Yayneshet et al. (5) stated that lignin is the single most important cell wall constituent that impacts digestibility, which substantiates the present study. NDF, ADF, and ADL are plant cell wall fractions linked and packed together in tight configurations to resist degradation, and hence their nutritional value to animals varies substantially, depending on the composition, structure, and degradability (38).
The correlation between nutrients and farmers’ feed value score
The farmers’ feed value preference score had a positive significant correlation with the nutritive value indicators DM (r = 0.615), OM (r = 0.458), CP (r = 0.768), IVDMD (r = 0.600), IVOMD (r = 0.565), DOMD (r = 0.600) and ME (r = 0.600), implying that farmers’ indigenous knowledge is relevant in judging the protein, energy, and digestibility values of browse species based on the perceived benefits associated with the animals’ performance measures. The greater the impact of feeding a certain browse species on animal performance, the higher the nutritional value, and hence the higher the grade. ADL (r = −0.702) and CT (r = −0.543) had a negative significant correlation with farmers’ feed preference score, indicating their impact on digestibility either through denying access or inhibiting microbial activity against cell wall components. Farmers used several indigenous criteria to judge the nutritional quality of available feed resources, according to Lumu et al. (49), including perceived effects on disease resistance, feed intake, growth/body condition, hair coat appearance, fecal output, and texture, and level of production, among others. Mekoya et al. (27) and Yisehak and Janssens (12) found a significant positive correlation of the farmers’ feed value score with the CP value of fodder trees and shrubs which partly agrees with the current study. The CP and IVDMD were positively correlated with the feed value score of the farmers in the highlands in the study conducted in northwestern Ethiopia (4), which agrees with the present study.
Because of its role in rumen microbial activity, CP showed a positive significant connection with IVDMD, DOMD, and ME. The ILFTS in the study had moderate to high CP values, which improved digestibility by increasing microbial activity. The high CP value of the ILFTS in the study suggests they could be used to supplement the N deficiency observed in ruminants feeding poor quality pastures and crop residues as a basal diet (26, 50).
Unlike ADF, which revealed a negative non-significant association with IVDMD, IVOMD, DOMD, and ME, ADL showed a strong negative significant link with IVDMD, IVOMD, DOMD, and ME. Both the IVDMD and IVOMD were depressed by the high ADL value of some of the indigenous browses in the research. The complex structure of plant cell walls, particularly the physical protection afforded by lignin, covalent connections between lignin and phenolic chemicals, and cell wall polysaccharides, impedes rumen digestion of fibrous plant components (51). Moore and Jung (48) discovered that lignin has a strong inhibitory influence on cell wall digestibility.
CT showed a negative significant connection with IVDMD, IVOMD, and DOMD, showing that it has a digestive inhibitory impact via suppressing microbial activity. Several investigations have revealed that CT is a secondary metabolite that binds the available CP in the rumen (9, 52) and hence lowers rumen microbial activity, affecting DM degradability. The CT and ADL have a positive connection, implying that they have a complimentary biosynthesis where the former is a secondary metabolite and the latter is a structural component.
Conclusion
In the study, the ILFTS had a CP value over 81 g/kg DM confirming that it can be used to supplement ruminant diets that are deficient in N. In spite of this, there is a wide variation in nutritive quality among indigenous browse trees, probably because of the differences between species and agroecological zone which impact farmers’ preferences score. The farmer’s evaluation of ILFTS species was multidimensional, which encompasses the perceived benefits associated with animal performance measures and desired characteristics of a tree as they have been used for multiple purposes. Yet, the preference score for all evaluation parameters varied significantly with species except for growth rate in the lowland and compatibility and overall mean in the highlands. However, significant differences were observed in growth rate, biomass yield, and overall mean between shrubs and trees in the lowlands. The nutrients in ILFTS exhibited various correlations among themselves and with feed value preference scores depending on their distinct nature and biochemical function. Additionally, CP, CT, and ADL values of the ILFTS had significant correlations with IVDMD, thereby affecting the energy and protein supply of the ILFTS forage, as well as its feed value preference score as demonstrated by the animals’ performance measures. Thus, farmers’ indigenous knowledge of feed value may be relevant to for evaluating the nutritional quality of ILFTS forage by envisaging its nutrient content and interaction with other nutrients and may be used to complement scientific indicators.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://doi.org/10.5281/zenodo.6585390.
Author contributions
GA conceptualized the idea of the study, formulated the methodology, curated data, specimens of plant sample collection, performed formal analysis and investigation, wrote the original draft, and administrated the project. YK and DA reviewed and edited the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. FAO. Ethiopia: report on feed inventory and feed balance. Rome, Italy: Food and Agriculture Organization of the United Nations (2018).
2. Abraham, G, Kechero, Y, Andualem, D, and Dingamo, T. Indigenous legume fodder trees and shrubs with emphasis on land use and agroecological zones: identification, diversity, and distribution in semi-humid condition of southern Ethiopia. Vet Med Sci. (2022) 8:2126–37. doi: 10.1002/vms3.858
3. Lelamo, L. A review on the indigenous multipurpose agroforestry tree species in Ethiopia: management, their productive and service roles and constraints. Heliyon. (2021) 7:e07874. doi: 10.1016/j.heliyon.2021.e07874
4. Ayenew, A, Tolera, A, Nurfeta, A, and Assefa, G. Farmers’ preference and knowledge on indigenous multipurpose browse species towards their feed value in north western Ethiopia. Trop Subtrop Agroecosyst. (2021) 24:24. doi: 10.56369/tsaes.3124
5. Yayneshet, T, Eik, LO, and Moe, SR. Seasonal variations in the chemical composition and dry matter degradability of exclosure forages in the semi-arid region of northern Ethiopia. Anim Feed Sci Technol. (2009) 148:12–33. doi: 10.1016/j.anifeedsci.2008.02.003
6. Andualem, D, Gelgele, M, and Bayssa, M. In vitro gas production kinetics of selected multipurpose tree browses in Gelana rangelands. Livest Res Rural Dev. (2021) 33:Article #18 Available at: http://www.lrrd.org/lrrd33/2/a.dere3318.html
7. Brown, D, Ng’ambi, JW, and Norris, D. Effect of tanniniferous Acacia karroo leaf meal inclusion level on feed intake, digestibility and live weight gain of goats fed a Setaria verticillata grass hay-based diet. J Appl Anim Res. (2018) 46:248–53. doi: 10.1080/09712119.2017.1289939
8. Makau, DN, VanLeeuwen, JA, Gitau, GK, McKenna, SL, Walton, C, Muraya, J, et al. Effects of Calliandra and Sesbania on daily Milk production in dairy cows on commercial smallholder farms in Kenya. Vet Med Int. (2020) 2020:1–15. doi: 10.1155/2020/3262370
9. Naumann, HD, Tedeschi, LO, Zeller, WE, and Huntley, NF. The role of condensed tannins in ruminant animal production: advances, limitations and future directions. Rev Bras Zootec. (2017) 46:929–49. doi: 10.1590/S1806-92902017001200009
10. Mekonnen, K, Glatzel, G, and Sieghardt, M. Assessments of fodder values of 3 indigenous and 1 exotic woody plant species in the highlands of Central Ethiopia. Mt Res Dev. (2009) 29:135–42. doi: 10.1659/mrd.1098
11. Dida, MF, Challi, DG, and Gangasahay, KY. Effect of feeding different proportions of pigeon pea (Cajanus cajan) and neem (Azadirachta indica) leaves on feed intake, digestibility, body weight gain and carcass characteristics of goats. Vet Anim Sci. (2019) 8:100079. doi: 10.1016/j.vas.2019.100079
12. Yisehak, K, and Janssens, GPJ. Evaluation of nutritive value of leaves of tropical tanniferous trees and shrubs. Livest Res Rural Dev. (2013) 25:Article #28 Available at: http://www.lrrd.org/lrrd25/2/yise25028.htm
13. Meijer, SS, Catacutan, D, Ajayi, OC, Sileshi, GW, and Nieuwenhuis, M. The role of knowledge, attitudes and perceptions in the uptake of agricultural and agroforestry innovations among smallholder farmers in sub-Saharan Africa. Int J Agric Sustain. (2015) 13:40–54. doi: 10.1080/14735903.2014.912493
14. Haugerud, A, and Collinson, MP. Plants, genes and people: improving the relevance of plant breeding in Africa. Exp Agric. (1990) 26:341–62. doi: 10.1017/S0014479700018500
15. Boogaard, BK, Oosting, SJ, and Bock, BB. Elements of societal perception of farm animal welfare: a quantitative study in the Netherlands. Livest Sci. (2006) 104:13–22. doi: 10.1016/j.livsci.2006.02.010
16. Dires, A, Asefa, M, Ashenafi, A, and Wasihun, W. Discover the land of paradize: Gamo zone travel guide. 2nd ed. Arba-Minch, Ethiopia: Gamo Zone Culture Tourism and Sport Department (2021).
17. Kuntashula, E, and Mafongoya, PL. Farmer participatory evaluation of agroforestry trees in eastern Zambia. Agric Syst. (2005) 84:39–53. doi: 10.1016/j.agsy.2004.06.009
18. AOAC. Official methods of analysis of AOAC International. 18th ed. Rockville, MD: AOAC International (2006).
19. Van Soest, PJ, Robertson, JB, and Lewis, BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds.S0022-0302(91)78551-2
20. Van Soest, PJ, and Robertson, JB. Analysis of forages and fibrous foods: a laboratory manual for animal science. Ithaca, NY (USA): Cornell University (1985).
21. Makkar, H. P. S. (2000). Quantification of tannins in tree foliage. Animal Production and Health Sub-Programme; Joint FAO/IAEA division of nuclear techniques in food and agriculture. Available at: http://www-naweb.iaea.org/nafa/aph/public/pubd31022manual-tannin.pdf
22. Tilley, JMA, and Terry, RA. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci. (1963) 18:104–11. doi: 10.1111/j.1365-2494.1963.tb00335.x
23. AOAC. Official method of analysis of the Association of Official Analytical Chemists. 15th ed. Rockville, MD: Association of Official Analytical Chemists, Inc (1990).
24. McDonald, P, Edwards, RA, Greenhalgh, GFD, Morgan, CA, Sinclair, LA, and Wilkinson, RG. Animal nutrition. 7th ed. London: Pearson Press (2010).
25. Chimphango, SBM, Gallant, LH, Poulsen, ZC, Samuels, MI, Hattas, D, Curtis, OE, et al. Native legume species as potential fodder crops in the mediterranean renosterveld shrubland, South Africa. J Arid Environ. (2020) 173:104015. doi: 10.1016/j.jaridenv.2019.104015
26. Balehegn, M. Silvopasture using indigenous fodder trees and shrubs: the underexploited synergy between climate change adaptation and mitigation in the livestock sector In: W Leal Filho, S Belay, J Kalangu, W Menas, P Munishi, and K Musiyiwa, editors. Climate change adaptation in Africa, climate change management. Cham: Springer (2017). 493–510. doi: 10.1007/978-3-319-49520-0_30
27. Mekoya, A, Oosting, SJ, Fernandez-Rivera, S, and Van der Zijpp, AJ. Multipurpose fodder trees in the Ethiopian highlands: farmers’ preference and relationship of indigenous knowledge of feed value with laboratory indicators. Agric Syst. (2008) 96:184–94. doi: 10.1016/j.agsy.2007.08.001
28. Mitiku, B. Evaluation and demonstration of indigenous fodder trees and shrubs in Alicho-Wriro District, Siltie zone. J Nat Sci Res. (2018) 8:21.
29. Balehegn, M, and Hintsa, K. Effect of maturity on chemical composition of edible parts of Ficus thonningii Blume (Moraceae): an indigenous multipurpose fodder tree in Ethiopia. Livest Res Rural Dev. (2015) 27:Article #233
30. Shenkute, B, Hassen, A, Assafa, T, Amen, N, and Ebro, A. Identification and nutritive value of potential fodder trees and shrubs in the mid rift valley of Ethiopia. J Anim Plant Sci. (2012) 22:1126–32.
31. Franzel, S, Carsan, S, Lukuyu, B, Sinja, J, and Wambugu, C. Fodder trees for improving livestock productivity and smallholder livelihoods in Africa. Curr Opin Environ Sustain. (2014) 6:98–103. doi: 10.1016/j.cosust.2013.11.008
32. Assefa, A, Kechero, Y, Tolemariam, T, Kebede, A, and Shumi, E. Anthelmintic effects of indigenous multipurpose fodder tree extracts against Haemonchus contortus. Trop Anim Health Prod. (2018) 50:727–32. doi: 10.1007/s11250-017-1488-0
33. Birhan, M, Gesses, T, Kenubih, A, Dejene, H, and Yayeh, M. Evaluation of anthelminthic activity of tropical Taniferous plant extracts against Haemonchus contortus. Vet Med Res Rep. (2020) 11:109–17. doi: 10.2147/VMRR.S225717
34. Adejoro, F.A. (2019). The use of condensed tannins and nitrate to reduce enteric methane emission and enhance utilization of high forage diets in sheep (Doctoral Dissertation). University of Pretoria, Pretoria, South Africa.
35. Cardoso-Gutierrez, E, Aranda-Aguirre, E, Robles-Jimenez, LE, Castelán-Ortega, OA, Chay-Canul, AJ, Foggi, G, et al. Effect of tannins from tropical plants on methane production from ruminants: a systematic review. Vet Anim Sci. (2021) 14:100214. doi: 10.1016/j.vas.2021.100214
36. Arigbede, OM, Anele, UY, Südekum, K-H, Hummel, J, Oni, AO, Olanite, JA, et al. Effects of species and season on chemical composition and ruminal crude protein and organic matter degradability of some multi-purpose tree species by West African dwarf rams: chemical composition and degradability of browse trees. J Anim Physiol Anim Nutr. (2012) 96:250–9. doi: 10.1111/j.1439-0396.2011.01146.x
37. Lee, MA. A global comparison of the nutritive values of forage plants grown in contrasting environments. J Plant Res. (2018) 131:641–54. doi: 10.1007/s10265-018-1024-y
38. Li, X. Plant cell wall chemistry: implications for ruminant utilisation. J Appl Anim Nutr. (2021) 9:31–56. doi: 10.3920/JAAN2020.0017
39. Grant, K, Kreyling, J, Dienstbach, LFH, Beierkuhnlein, C, and Jentsch, A. Water stress due to increased intra-annual precipitation variability reduced forage yield but raised forage quality of a temperate grassland. Agric Ecosyst Amp Environ. (2014) 186:11–22. doi: 10.1016/j.agee.2014.01.013
40. Ray, DK, Gerber, JS, MacDonald, GK, and West, PC. Climate variation explains a third of global crop yield variability. Nat Commun. (2015) 6:5989. doi: 10.1038/ncomms6989
41. Derero, A, and Kitaw, G. Nutritive values of seven high priority indigenous fodder tree species in pastoral and agro-pastoral areas in eastern Ethiopia. Agric Food Secur. (2018) 7:68. doi: 10.1186/s40066-018-0216-y
42. Rubanza, CDK, Shem, MN, Otsyina, R, Ichinohe, T, and Fujihara, T. Nutritive Evaluation of Some Browse Tree Legume Foliages Native to Semi-arid Areas in Western Tanzania. Asian-Aus J Anim Sci. (2003) 16:1429–437. doi: 10.5713/ajas.2003.1429
43. Mohameed, A, Kassahun, G, and Ayantu, M. Identification and nutritional evaluation of potential indigenous browse species in Guba Lafto District, North Wollo. Ethiopia J Anim Sci Res. (2020) 4:1–6. doi: 10.16966/2576-6457.144
44. McDonald, P, Edwards, RA, Greenhalgh, JFD, and Morgan, CA. Animal Nutrition. 6th ed. Hoboken, NJ: Prentice Hall (2002).
45. Enri, SR, Probo, M, Renna, M, Caro, E, Lussiana, C, Battaglini, LM, et al. Temporal variations in leaf traits, chemical composition and in vitro true digestibility of four temperate fodder tree species. Anim Prod Sci. (2020) 60:643–58. doi: 10.1071/AN18771
46. Gebremedhin, AT, Gedo, AH, Edo, GY, and Haile, ST. Evaluation of multi-functional fodder tree and shrub species in mid-altitudes of south Omo zone, southern Ethiopia. J Hortic For. (2020) 12:27–34. doi: 10.5897/JHF2019.0617
47. Datt, C, Datta, M, and Singh, NP. Assessment of fodder quality of leaves of multipurpose trees in subtropical humid climate of India. J For Res. (2008) 19:209–14. doi: 10.1007/s11676-008-0035-2
48. Moore, KJ, and Jung, H-JG. Lignin and fiber digestion. J Range Manag. (2001) 54:420–30. doi: 10.2307/4003113
49. Lumu, R, Katongole, CB, Nambi-Kasozi, J, Bareeba, F, Presto, M, Ivarsson, E, et al. Indigenous knowledge on the nutritional quality of urban and peri-urban livestock feed resources in Kampala, Uganda. Trop Anim Health Prod. (2013) 45:1571–8. doi: 10.1007/s11250-013-0401-8
50. Bouazza, L, Bodas, R, Boufennara, S, Bousseboua, H, and López, S. Nutritive evaluation of foliage from fodder trees and shrubs characteristic of Algerian arid and semi-arid areas. J Anim Feed Sci. (2012) 21:521–36. doi: 10.22358/jafs/66126/2012
51. Yu, P, McKinnon, JJ, and Christensen, DA. Hydroxycinnamic acids and ferulic acid esterase in relation to biodegradation of complex plant cell walls. Can J Anim Sci. (2005) 85:255–67. doi: 10.4141/A04-010
Keywords: agroecosystems, farmers’ evaluation, feed value, indigenous knowledge, nutritive value
Citation: Abraham G, Kechero Y and Andualem D (2023) Nutritional quality of indigenous legume browse in southern Ethiopia: farmers’ preference and correlation of local valuation of feed value with scientific indicators. Front. Vet. Sci. 10:1198212. doi: 10.3389/fvets.2023.1198212
Edited by:
Adugna Tolera, Hawassa University, EthiopiaReviewed by:
Nuh Ocak, Ondokuz Mayıs University, TürkiyeMaghsoud Besharati, University of Tabriz, Iran
Lars Olav Eik, Norwegian University of Life Sciences, Norway
Copyright © 2023 Abraham, Kechero and Andualem. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Getachew Abraham, Z2VjaG9hYnJhaGFtQGdtYWlsLmNvbQ==