
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Vet. Sci. , 15 September 2023
Sec. Parasitology
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1188659
This article is part of the Research Topic Parasitism: The Good, The Bad and The Ugly View all 14 articles
 Wendy Wee1
Wendy Wee1 Guillermo Téllez-Isaías2
Guillermo Téllez-Isaías2 Zulhisyam Abdul Kari3,4*
Zulhisyam Abdul Kari3,4* Romalee Cheadoloh5Muhammad Anamul Kabir6Khairiyah Mat3,4
Romalee Cheadoloh5Muhammad Anamul Kabir6Khairiyah Mat3,4 Suniza Anis Mohamad Sukri3,4
Suniza Anis Mohamad Sukri3,4 Mohammad Mijanur Rahman3,4
Mohammad Mijanur Rahman3,4 Nor Dini Rusli3,4
Nor Dini Rusli3,4 Lee Seong Wei3,4*
Lee Seong Wei3,4*Soybean lecithin is extensively used as the dietary supplementation of phospholipids in animal production. Soybean lecithin plays significant roles in aquafeed as growth promoter, feed enhancer, immunity modulator and antioxidant activity stimulator for aquaculture species. Besides, soybean lecithin is also reported to help aquaculture species being resilient to physical and chemical stressors. In this review, common sources, chemical structure and mode of action of lecithin, with highlight on soybean lecithin application in aquaculture over four-decadal studies published between 1983 and 2023, were evaluated and summarized. By far, soybean lecithin is best-known for its beneficial effects, availability yet cost-effective for aquafeed formulation. Findings from this review also demonstrate that although nutritional profile of long-chain polyunsaturated fatty acids and phosphatidylcholine from egg yolk and marine sources are superior to those from plant sources such as soybean, it is rather costly for sustainable application in aquafeed formulation. Moreover, commercially available products that incorporate soybean lecithin with other feed additives are promising to boost aquaculture production. Overall, effects of soybean lecithin supplementation are well-recognized on larval and juvenile of aquaculture species which having limited ability to biosynthesis phospholipids de novo, and correspondingly attribute to phospholipid, a primary component of soybean lecithin, that is essential for rapid growth during early stages development. In addition, soybean lecithin supplementation plays a distinguish role in stimulating maturation of gonadal development in the adults, especially for crustaceans.
Aquafeed is the main production expense of an aquaculture operation, accounting for about 50 to 70% of the total aquaculture operation cost. According to a recent study, aquafeed costs about 65% of the total aquaculture production cost (1). The feed cost is expected to further increase due to excessive reliance on conventional raw materials, especially fish oil and fish meal in aquafeed formulation (2). Sustainability of the aquaculture industry can be greatly impacted by the future shortages of fish oil and fish meal (3). Hence, it is vital to have some animal origin materials gradually replaced by substances derived from plant origin such as soybean lecithin, soybean oil, and soybean meal in the aquafeed formulation.
As a byproduct of the oilseed industry, lecithin is widely introduced into food, cosmetic, pharmaceutical and other non-food industries as the emulsifier and liposomes producer, along with their great nutritional value (4). In the aquaculture industry, lecithin becomes one of the important raw materials to meet essential fatty acid requirements of the targeted species (5). Lecithin production became well-established in the United States in 1940 and expanded rapidly upon the commercial introduction of genetically modified (GMO) soybeans in 1996. On the other hand, lecithin from non–GMO sources like sunflower, rapeseed, and rice bran are favored by the European market (6, 7). Soybean lecithin is currently the primary source available in the worldwide market and offers a comparatively affordable price compared to lecithin from other sources, especially the marine lecithin (8). Commercial soybean lecithin claimed to consist high concentration of phospholipid, 65–75% (9) while some other studies showed that concentration of phospholipids in soybean lecithin ranged between 28–44% (10), 55–57% (11), 47% (12), and 38–45% (13). These differences were probably due to the discrepancies in the purity of soybean lecithin and approaches used to determine the concentration of phospholipid in soybean lecithin.
Aquafeed formulation that offer phospholipid composition resembles to the fish egg is deemed to be an ideal diet for fish larvae to ensure maintenance and functionality of cellular membrane structure, and as a source of energy to the fish (14). Therefore, phospholipid is supplemented in the larval diet of the aquatic animals owing to their limited biosynthetic capacity in the initial stages (15–17). For most of the aquaculture species, a supplementation of 8 to 12% dietary phospholipid is regarded to be optimal to promote growth and survival (18). Nonetheless, requirements for dietary phospholipids vary among species, life stage of the species, and the source and purity of the phospholipids. Phospholipid from different sources of lecithin such as soybean, milk, egg and krill, differs from one source to another in terms of phospholipid classes and fatty acids nature (19). Remarkably, soybean lecithin predominated by phosphatidylcholine, followed by phosphatidylethanolamine, and phosphatidylinositol provides an excellent source of phospholipids and fatty acids which are essential to aquatic animals during their early life stages (20). In this review, common sources, chemical structure and mode of action of lecithin are presented with the examples on how soybean lecithin promotes aquaculture production by improving feed utilization, growth performance, intestinal health, antioxidant capacity and resilience against stressors.
Literally means egg yolk in Greek word lekithos, lecithin was first extracted from the egg yolk by a French chemist, Theodore Nicolas Gobley in 1850. Subsequently, soybean lecithin came into worldwide commercial availability in 1921, almost a decade after the introduction of soybean from China into Europe and North America (21, 22). Global lecithin market size has exceeded USD 2 billion in 2021 and is forecasted to surpass USD 3.4 billion by 2030 (23). Lecithin is a complex mixture of phosphatide fraction available in both plants and animals, especially in the soybean and egg yolk (19). As a result of the esterification of choline, ethanolamine, serine, and inositol to a phosphatidic acid backbone, phospholipids are formed and classified into respective phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylserine (PS) and phosphatidylinositol (PI) with other constituents of fatty acids, triglycerides, and carbohydrates (4). Despite that the term for lecithin specifically refers to phosphatidylcholines in some earlier literatures, it is often used interchangeably with phospholipids since the commercial lecithin is largely made up of phospholipids and vegetable oils (24, 25).
Lecithin can be isolated and characterized from various sources, both from the terrestrial and aquatic (Table 1). These including soybean (48), sunflower (29), rapeseed (26), corn (34), camelina seed (32), canola (31), rice bran (35), egg yolk (37), dairy products (38, 39), and marine products (41, 42) that displaying assorted profile of phospholipids due to variations of two ester-bonded fatty acids at the sn-1 and sn-2 positions of the glycerol backbone. The sn-1 position chiefly carries a saturated fatty acid such as stearic acid or palmitic acid, whereas the sn-2 position carries an unsaturated fatty acid such as oleic acid, linoleic acid, α-linolenic acid, arachidonic acid, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) (49). Lecithin from different sources differs substantially on structural and functional roles depending on its origin, either plant or animal origin, and the extraction process (27, 50). An example of chemical structure and model of soybean lecithin is presented in Figure 1.
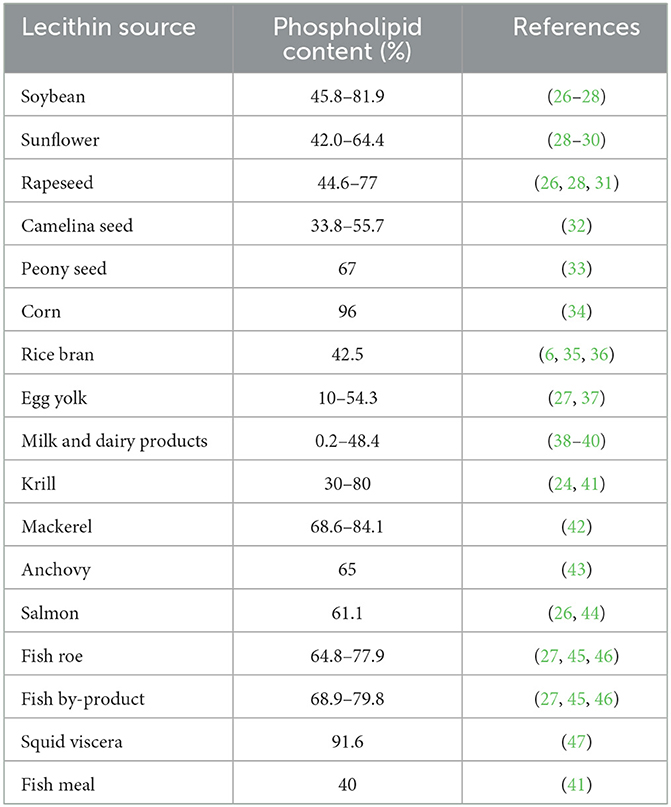
Table 1. Phospholipid content of various lecithin sources.
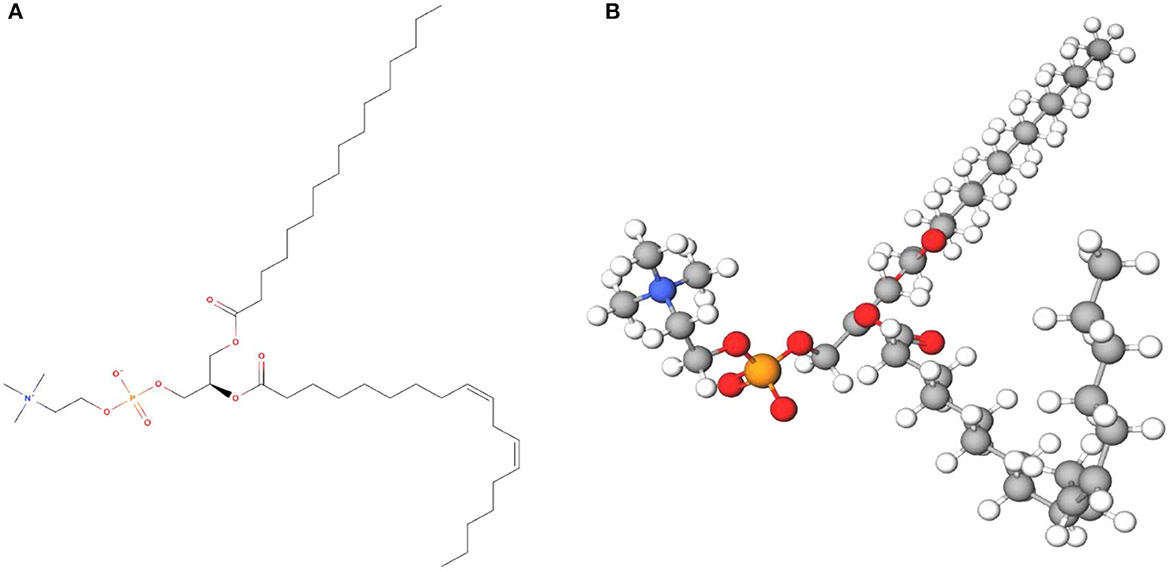
Figure 1. Soybean lecithin (C42H80NO8P) in (A) 2D chemical structure and (B) 3D model (MolView).
Lecithin is commonly extracted by degumming crude vegetable oils and drying the hydrated gums. Soybean oil, for example, contains approximately 60% acetone-insoluble which corresponds to 2–3% of the commercial lecithin and high amounts of phosphorus (12). Compared to sunflower and rapeseed oils, crude soybean oil has the greatest phospholipid concentration that is still present in vegetable oils after extraction (51). Therefore, soybean oil is the main source for the production of commercial lecithin. A typical crude soybean lecithin is reported to contain 18% PC, 14% PE, 9% PI, 5% phosphatidic acid, 2% minor phospholipids, 11% glycolipids, 5% complex sugars and 37% neutral oil (20). However, these properties are subjected to alteration by deoiling the crude lecithin into high-purity refined lecithin products, or by chemical and enzymatic modification of the phospholipids (52). For examples, the refined soybean lecithin has a greater purity of phospholipid (97.6%) than those from the crude soybean lecithin (60%) (53). Similarly, corn lecithin deoiled by supercritical carbon dioxide is claimed to contain 96% of phospholipids (34).
Animal lecithin derived from eggs, milk and bovine brain contains sphingomyelin on top of the major phospholipid classes such as PC, PE, PS and PI. Egg yolk lecithin is distinguished from soybean lecithin by having relatively higher proportion of saturated fatty acids, n-6 and n-3 polyunsaturated fatty acids (PUFAs) such as arachidonic acid and DHA. It is even more oxidatively stable than soybean lecithin (54). However, the application of egg yolk lecithin in aquafeed is not a viable option due its cost and commercial availability (37). On the other hand, marine lecithin derived from cold-water fishes is predominated by PC and PE and characterized by the presence of n-3 PUFAs such as EPA [20:5(n-3)] and DHA [22:6(n-3)] (55). For instance, marine lecithin extracted from salmon head was found to be rich in phospholipids, especially the PC (44). Nonetheless, supplies of the marine lecithin are not sustainable and therefore much more costly than other sources of lecithin (41, 56). Unlike the animal lecithin, plant lecithin derived from oilseeds such as soybean, rapeseed and sunflower contain mainly of PC, PE and PI and characterized by the availability of mono- and poly- unsaturated fatty acids, namely the oleic acid (18:1), linoleic acid [18:2 (n-6)], and α-linolenic acid [18:3(n-3)] (26). In particular, soybean lecithin has been the most widely used phospholipid source in aquaculture feed formulation due to its market availability and the beneficial effects of promoting growth performance and enhancing survival on fish (57).
Soybean lecithin is an essential component of aquafeed that exhibits multi-faceted mode of action and involves diverse mechanisms. Primarily used in aquafeed as an emulsifying agent, phospholipid components of soybean lecithin form a protective layer surrounding the lipid droplets in the feed, thus improves the stability of aquafeed and reduces the leaching rate of water-soluble vitamins and minerals (58, 59). By increasing the surface area of the lipid droplets in the aquafeed, soybean lecithin serves to increase the digestibility of aquafeed by making it more easily accessed by digestive enzymes to break down the lipids into smaller molecules. This is particularly important in carnivorous fish species that require a high level of dietary lipids for optimal growth (60). Phospholipids are important for the maintenance of bio-membrane structure, formation of cell organelles and superior to neutral lipids as the main energy source during early life history stages of some fish species (8, 18, 61). Phospholipid deficiency in fish larval diets has been reported to develop aberrant lipid deposition in intestinal enterocytes due to insufficient chylomicron synthesis and reduced specific activity of lipase (62, 63). Besides, phospholipids are reported to suppress cholesterol absorption in the animal digestive system (64) while facilitate the transport of lipids released from the hepatopancreas into hemolymph and enhance the availability of dietary cholesterol in the prawn Penaeus japonicus (65).
To ensure that a nutritionally complete and balanced diet is delivered, the mechanism of metabolism for soybean lecithin in aquafeed primarily involves the breakdown of its phospholipid components by digestive enzymes, followed by the absorption of fatty acids and nutrients, and the conversion of choline to betaine. Based on the chemical structure of soybean lecithin, free fatty acids are readily to be absorbed by the fish gut and transported to other tissues for storage, energy production, or used as precursors for various metabolic pathways upon the hydrolysis of ester bonds that link the fatty acid chains to the glycerol backbone of phospholipid (8, 66). Unsaturated fatty acids of soybean lecithin, particularly the omega-3 and omega-6 fatty acids, are crucial for fish growth and health (55). However, choline which typically forms the polar head group of soybean lecithin, is metabolized differently depending on fish species and its nutritional requirements. The conversion of choline to betaine is one of the common pathways that facilitate osmoregulation in fish and profoundly affects a series of metabolic processes including DNA methylation, protein synthesis, lipid metabolism, and energy production (67, 68).
Betaine, the oxidized form of choline, usually serves as a methyl group donor that transfers methyl groups (CH3) to metabolic compounds such as DNA, RNA, protein and lipid (69). With the addition of methyl group from betaine and alteration to the structure of DNA molecule, betaine regulates gene expression for growth development and immune responses (70). Furthermore, methylation process is essential for the synthesis of structural proteins and enzymes in aquaculture species. Lacking methyl groups in an organism likely to trigger a condition of under–methylation of DNA and concomitant activation of oncogenes (58, 71). Yet, these methyl groups cannot be synthesized by animals and can only be derived from diet (72). By regulating the activity of enzymes involved in the synthesis and breakdown of fatty acids, betaine impacts on fish lipid metabolism, which modifies the composition of body fat and cell membrane consequently (73). On top of that, betaine is also known to serve as an osmoprotectant in the energy production in fish by lowering the energy demands to regulate ions and cell volume, and supporting the conversion of homocysteine into methionine, an essential amino acid required for protein synthesis (70). Hence, the availability of methyl groups in the diet and functionality of soybean lecithin imparts a number of advantages to aquaculture species.
In addition to its role as the source of fatty acids and choline, soybean lecithin also delivers other important nutrients, such as phosphorus and vitamin E. Phosphorus is an essential mineral that is required for bone formation and other metabolic processes (74), while vitamin E is a powerful antioxidant that can protect cell membranes from oxidative damage (75). By providing a balanced and nutritionally complete diet, soybean lecithin is well-documented to enhance growth performance, survival, proper bone skeletal formation and stress mitigation (76, 77).
Overall, mode of action of soybean lecithin in aquafeed highly relies on its emulsifying properties, as well as its abilities to enhance digestibility and nutritional quality of the aquafeed. Depending on the composition of phospholipid which includes the fatty acids chains, glycerol backbone, phosphate group, and the polar head group, soybean lecithin contributes crucial roles to the growth performance, lipid and carbohydrate metabolisms, nutrient utilization, antioxidant activities and stress resistance in farmed species (78).
Extensive studies have been conducted to evaluate the effect of dietary phospholipid in different aquaculture species at different stages by using egg yolk, soybean and marine lecithin in the aquafeed formulation. Of major interest in aquafeed supplementation are the PC, PE and PI (79, 80). Soybean lecithin has been receiving considerable attention in aquafeed formulation due to their implication in numerous metabolic pathways and regulation processes that improve the feed utilization and growth performance of larval and juvenile fish species such as red sea bream, Pagrus major (81), knifejaw, Oplegnathus fasciatus (81), ayu, Plecoglossus altivelis (82), rainbow trout, Oncorhynchus mykiss (83), Atlantic salmon, Salmo salar (84–86), red drum, Sciaenops ocellatus (87), goldfish, Carassius auratus (88), common carp, Cyprinus carpio (89), seabass, Dicentrarchus labrax (74), Japanese flounder, Paralichthys olivaceus (90), cobia, Rachycentron canadum (91), amberjack Seriola dumerili(92), rohu, Labeo rohita (68), gilthead seabream, Sparus aurata (93, 94), silvery-black porgy, Sparidentex hasta (95), large yellow croaker, Lamichthys crocea (57), Nile tilapia, Oreochromis niloticus (96), hybrid grouper, Epinephelus fuscoguttatus x E. lancolatus (60), and giant grouper, E. lanceolatus (97) (Supplementary Table 1). There are numerous explanations on how the inclusion of soybean lecithin possibly promotes feed utilization and growth performance of the larval aquaculture species. Dietary soybean lecithin, notably phospholipids, has been showing stimulating effects on larval growth and survival when de novo synthesis of phospholipids in larval fish was insufficient to meet the requirements of the fish during their early development (98, 99). High phospholipids content in the dietary soybean lecithin not only serves as a superior energy source to the larvae (8), but also promotes the conversion of phospholipids to other lipids, including diacylglycerol, diphosphatidylglycerol, PUFAs, and cholesterol (100). Comparatively, larval stages are more vulnerable to dietary phospholipid deficiency and require more dietary phospholipids than the juveniles. Phospholipids requirements for larval fish varied from 2 to 12% with higher requirements among the marine larval fish (8, 100). As larval stages are highly sensitive to phospholipid deficiency, inclusion of soybean lecithin in microparticulate diet serves an essential role to satisfy phospholipid requirement for ontogenetic development at the expense of fish oil, fish meal and live food. For instance, linoleic acid, a polyunsaturated fatty acid that fish require but cannot be produced on their own, is provided by soybean lecithin (101). Soybean lecithin is known to be easily digested by fish into lysophosphatidylcholine form and absorbed directly by fish (102). The distribution and digestion of ingested free oleic acid (18:1n-9) in fish larvae were significantly influenced by dietary phosphatidylcholine (103, 104). Furthermore, supplemental emulsifier such as soybean lecithin can stimulate the production of lipoprotein in fish digestive system to enhance feed digestibility (105). High PC composition in soybean lecithin functions as an age-dependent feed attractant in enhancing feeding activity to fish larvae and therefore displaying a notable impact on fish development (106, 107).
Besides finfish species, soybean lecithin also demonstrated significant role in supporting good growth performance in a number of larval and juvenile penaeid species, including Penaeus japonicus (65, 108), Penaeus merguiensis (109), Penaeus monodon (66, 110), Litopenaeus vannamei (53, 111–113), and Macrobrachium rosenbergii (114, 115). The dietary phospholipid is essential for promoting greater rates of cholesterol turnover from the gut to the circulatory system in crustaceans (116). In recent studies, larval mud crab, Scylla serrata (117), juvenile swimming crab, Portunus trituberculatus (118, 119) and Chinese mitten crab, Eriocheir sinensis (120, 121) also benefited from the inclusion of soybean lecithin that enhances feed utilization, improves survival rate, promotes growth performance and molting frequency. Besides growth performance, positive effect of dietary soybean lecithin is gaining attention on gonadal development of the brood stock such as Chinese mitten crab, Eriocheir sinensis (122), swimming crab, Portunus trituberculatus (123, 124), red claw crayfish, Cherax quadricarinatus (125), and adult sea urchin, Strongylocentrotus intermedius (126). Nevertheless, juvenile sea urchin (127) is observed to have reduced weight gain when increased dietary phospholipid levels are provided and these excessive phospholipids eventually converted to neutral lipid in the gut and gonad (Supplementary Table 1).
In feed formulations for juvenile black seabream, Acanthopagrus schlegeli, soybean oil may be used up to 60 to 80% in place of fish oil. However, fully replacement of fish oil with soybean oil in the feed formulation is not recommended as it can reduce growth performance in juvenile black seabream (128). This was supported by the studies of Seiliez et al. (129) that growth performance of larval gilthead seabream, Sparus aurata was affected when soybean lecithin was utilized as a total replacement of live feed whereas overdose of soybean lecithin in the feed formulation (> 35.6 g/ kg diet) lead to decline in growth performance of early juvenile milkfish, Chanos chanos (130). On the other hand, some aquaculture species were found to perform better when marine lecithin was administered instead of soybean lecithin (93, 131). Similar finding was also observed in the study of Salini et al. (132) where juvenile barramundi, Lates calcarifer received marine lecithin from krill showed better growth performance than those received soybean lecithin. This may be attributed to the presence of high concentration of PUFAs mainly the EPA and DHA in marine lecithin which are important in promoting growth performance of aquatic animals in the early stage of life (27). As demonstrated by Liu et al. (133), EPA is needed to improve larval growth and survival when DHA level is high but arachidonic acid (ARA) level is low. Although marine lecithin tends to outperform as growth promoter for aquaculture species, soybean lecithin is favored as an alternative growth promoter because it is more viable and economically wise compared to marine lecithin from krill and fish meal. Moreover, study by Jaxion-Harm (56) established that phospholipids derived from soybean lecithin are not significantly different from marine sources such as krill and fish meal in promoting growth performance of Atlantic salmon, Salmo salar fry (Supplementary Table 1).
Global warming and climate change are issues being highlighted in recent years. These issues have an impact on world aquaculture production in which increased water temperature is one of the abiotic stressors to aquatic animals (134). Sensitivity of fish to xenobiotics is also indirectly influenced by the fluctuations of temperature. Presence of organochlorine pesticides such as endosulfan in the water was found to aggravate the situation by causing a 2.6 to 6°C reduction in thermal tolerance of freshwater fishes (135). Pesticides can seep into the aquatic environment as a result of extensive usage in agricultural activities and it is highly toxic to aquatic animals (136, 137). Thus, the nutritional approach is employed as one of the strategies to enhance the immune system and allow aquatic animals to be resistant to different stressors, especially to thermal stress.
It has been observed that inclusion of dietary soybean lecithin has a beneficial effect on thermal tolerance in aquaculture species which allows the aquaculture species to be more resilient to the fluctuation of water temperature. For instance, Kumar et al. (58) postulated that 1.5 to 2% of soybean lecithin helps milkfish, Chanos chanos coping stress from high temperature up to 46.4°C via protection of antioxidative status and neurotransmitter enzymes. Besides, soybean lecithin was observed to allow fish being resistant to stress caused by low temperature (138). Thermal tolerance of aquatic animals is influenced by many factors such as the presence of toxic in the water (139), species of aquatic animals (140), size of aquatic animals (141) and acclimation temperature (142). In this case, antioxidant defense system of fish may be fortified by soybean lecithin and thereby less susceptible to fluctuations in water temperature. Not only serves as the raw material in the repair of cell damage caused by thermal stress, soybean lecithin also promotes the expression of heat shock protein, which offers protection to the cells against the accumulation of altered proteins caused by high or low temperature stress (143–145).
Other than thermal stress, soybean lecithin is found to be effective in helping aquaculture species to cope with stress caused by hypoxic condition (78, 100), changes in water salinity (57), and the presence of pesticide in the water (68). Therefore, supplementation of lecithin from soybean in the feed formulation for aquaculture species is noteworthy to enhance stress tolerance of aquaculture species toward abiotic stressors (Supplementary Table 1).
Despite those phospholipid and fatty acid components of soybean lecithin are playing important roles in the health maintenance of various aquaculture species, documentation on the effects of soybean lecithin on intestinal health, whole body total lipid content, antioxidant capacity and immunity of aquaculture species are relatively limited in the past. Supplementation of soybean lecithin in the micro diet has been observed to prevent intestinal steatosis in the larval common carp, Cyprinus carpio (146), and promotes intestinal health of juvenile red drum, Sciaenops ocellatus (87), larval largemouth bass, Micropterus salmoides (147), yellow drum, Nibea albiflora (148), and adult Nile tilapia, Oreochromis niloticus (105). In addition, phospholipid in the soybean lecithin helps to regulate lipid metabolism and increases body lipid content of juvenile amberjack, Seriola dumerili (92), large yellow croaker, Larmichthys crocea (63), silvery-black porgy, Sparidentex hasta (95), hybrid grouper, Epinephelus fuscoguttatus × E. lancolatus (149). Higher body lipid content is obtained when fatty acids delivery and uptake in the fish are improved (Supplementary Table 1).
In many organisms, antioxidant system functions to mitigate the effects of reactive oxygen species (ROS) by protecting and repairing cells from oxidative damage. Dietary supplementation of soybean lecithin is capable to trigger antioxidant responses in aquaculture species when dealing with oxidative stress, or the elevation of ROS at intracellular level (150, 151). Activities of radical scavenging enzymes such as the superoxide dismutases (SOD), catalase (CAT), glutathione peroxidase (GPX), glutathione-S-transferase (GST) and glutathione reductase (GR) have been used as effective biomarkers to examine the effects of dietary phospholipid on enzymatic function and biochemical pathways in several studies involving larval Dojo loach, Misgurnus anguillicaudatus (152), common carp, Cyprinus carpio (153), golden mahseer, Tor putitora (138), stellate sturgeon, Acipenser stellatus (75), gilthead seabream, Sparus aurata (94), and hybrid snakehead, Channa argus x C. maculata (154). As a consequence of antioxidant capacity-promoting effect of dietary phospholipid, lipid peroxidation is reduced and survival rate is increased in the fish (63).
Dietary supplementation of soybean lecithin is also associated with the enhancement of fish systemic immunity against bacterial infection. According to Adel et al. (153), better immunostimulatory effect was reflected by common carp, Cyprinus carpio that received 3% soybean lecithin-enriched diet with an increase of mucosal immune parameters including alkaline phosphatase, lysozyme, protease, and esterase activity. These enzymes are involved in the regulatory secretion of antimicrobial peptides to suppress bacterial pathogens such as Aeromonas hydrophila, Streptococcus iniae, Yersinia ruckeri, and Lactococcus garviea. The enhancement on immunity and phagocytic activity is further supported by Jafari et al. (155) with an inclusion of 3.3% soybean lecithin for juvenile stellate sturgeon, Acipenser stellatus, 6–9% of soybean lecithin for pre-spawning Caspian brown trout, Salmo trutta caspius (156), and 0.3 g of soybean lecithin-containing bioemulsifier Lysomax® per kg of diet for the adult Nile tilapia, Oreochromis niloticus (105). In contrast, there is almost no observable impact on immune genes expression in golden mahseer fry when diet is supplemented with soybean lecithin (138). Similarly, diet supplemented with 4% soybean lecithin showed limited effects on the innate immune system and whole-body composition of juvenile channel catfish, Ictalurus punctatus, despite an improvement on feed conversion (157). Further studies need to be carried on different aquaculture species in order to be conclusive on the effect of dietary soybean lecithin on immunity enhancement.
Lecithin is obtainable from a wide variety of sources, including terrestrial and aquatic animals and plants. Different sources of lecithin have different compositions of phospholipids and fatty acids which determine the structural and functional roles of lecithin. Despite the fact that marine lecithin contains a high concentration of PUFAs, especially EPA and DHA, soybean lecithin is leading the role as growth promoter in the production of major aquaculture species because it is more practical and cost-effective for aquafeed formulation. Many studies have been included in this review to evaluate the potential of soybean lecithin supplementation and substitution for marine lecithin and live feed in the micro diet of aquaculture species in their early life stages. As concluded from the literature, proper supplementation of phospholipids from soybean lecithin in the larval and juvenile diet have significantly improves growth, survival, feed utilization, lipid metabolism, skeletal formation, stress tolerance, antioxidant capacity, and immune response in a number of finfish and shellfish species from larval to juvenile stage. However, research on the effect of dietary soybean lecithin on pre-reproductive phase and brood stock of crustacean species are relatively lacking. In this regard, future work that address the application of soybean lecithin in the diet formulation for pre-reproductive and brood stock of crustacean species is strongly recommended, with emphasis on enhancing lipid utilization, gonadal development and seed quality.
Writing—original draft preparation: WW, ZA, and LW. Writing—review and editing: RC, MK, KM, NR, SM, and MR. Supervision: GT-I and LW. All authors contributed to the article and approved the submitted version.
This research was funded by the International Grant from Yala Rajabhat University (grant no.: R/YALA/A0700/00387A/008/2023/01159) and supported in part by funds provided by the USDA-NIFA Sustainable Agriculture Systems, grant no. 2019-69012-29905.
This review article is a collaborative work between Universiti Malaysia Kelantan, Sylhet Agricultural University, Universiti Malaysia Terengganu, and University of Arkansas. The collaboration is part of the planning by Advanced Livestock and Aquaculture Research Group – ALAReG under Faculty of Agro-Based Industry, Universiti Malaysia Kelantan, Jeli Campus.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer RC declared a shared affiliation with the author GT-I to the handling editor at the time of review.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2023.1188659/full#supplementary-material
1. Ekmekci H, Gül M. Economic structure and problems of trout enterprises: a case of Fethiye. Turkish J Agric Food Sci Technol. (2017) 5:33–42. doi: 10.24925/turjaf.v5i1.33-42.806
2. Aponte FR, Tveterås S. On the drivers of cost changes in the Norwegian salmon aquaculture sector: a decomposition of a flexible cost function from 2001 to 2014. Aquac Econ Manag. (2019) 23:276–91. doi: 10.1080/13657305.2018.1551438
3. Turchini GM, Trushenski JT, Glencross BD. Thoughts for the future of aquaculture nutrition: realigning perspectives to reflect contemporary issues related to judicious use of marine resources in aquafeeds. N Am J Aquac. (2019) 81:13–39. doi: 10.1002/naaq.10067
4. Johnson W, Bergfeld WF, Belsito DV, Hill RA, Klaassen CD, Liebler DC, et al. Safety assessment of lecithin and other phosphoglycerides as used in cosmetics. Int J Toxicol. (2020) 39:5S–25S. doi: 10.1177/1091581820953123
5. Hardy RW, Brezas A. “Diet formulation and manufacture,” In: Hardy RW, Kaushik SJ, editors. Fish Nutrition. Amsterdam: Elsevier (2022). p. 643–708.
6. Jala RCR, Prasad RBN. “Rice bran lecithin: compositional, nutritional, and functional characteristics,” In: Moghis UA, Xu X, editors. Polar Lipids: Biology, Chemistry, and Technology. Amsterdam: Elsevier (2015). p. 35–55.
7. List GR. “Soybean lecithin: food, industrial uses, and other applications,” In: Moghis UA, Xu X, editors. Polar Lipids: Biology, Chemistry, and Technology. Amsterdam: Elsevier (2015). p. 1–33.
8. Tocher DR, Bendiksen EÅ, Campbell PJ, Bell JG. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture. (2008) 280:21–34. doi: 10.1016/j.aquaculture.2008.04.034
9. Scholfield CR. Composition of soybean lecithin. J Am Oil Chem Soc. (1981) 58:889–92. doi: 10.1007/BF02659652
10. Hurst WJ, Martin RA. The analysis of phospholipids in soy lecithin by HPLC. J Am Oil Chem Soc. (1984) 61:1462–3. doi: 10.1007/BF02636365
11. Helmerich G, Koehler P. Comparison of methods for the quantitative determination of phospholipids in lecithins and flour improvers. J Agric Food Chem. (2003) 51:6645–51. doi: 10.1021/jf0345088
12. van Nieuwenhuyzen W, Tomás MC. Update on vegetable lecithin and phospholipid technologies. Eur J Lipid Sci. (2008) 110:472–86. doi: 10.1002/ejlt.200800041
13. Pires LN, Brandão GC, Teixeira LSG. Determination of phospholipids in soybean lecithin samples via the phosphorus monoxide molecule by high-resolution continuum source graphite furnace molecular absorption spectrometry. Food Chem. (2017) 225:162–6. doi: 10.1016/j.foodchem.2017.01.019
14. Sargent J, McEvoy L, Estevez A, Bell G, Bell M, Henderson J, et al. Lipid nutrition of marine fish during early development: current status and future directions. Aquaculture. (1999) 179:217–29. doi: 10.1016/S0044-8486(99)00191-X
15. D'Abramo LR, Bordner CE, Conklin DE, Baum NA. Essentiality of dietary phosphatidylcholine for the survival of juvenile lobsters. J Nutr. (1981) 111:425–31. doi: 10.1093/jn/111.3.425
16. Akiyama DM, Dominy WG, Lawrence AL. “Penaeid shrimp nutrition,” In: Fast AW, James Lester L, editors. Marine Shrimp Culture: Principles and Practices. Amsterdam, The Netherlands: Elsevier Science (1992). p. 535–568 doi: 10.1016/B978-0-444-88606-4.50031-X
17. Carmona-Antoñanzas G, Taylor JF, Martinez-Rubio L, Tocher DR. Molecular mechanism of dietary phospholipid requirement of Atlantic salmon, Salmo salar, fry. BBA Mol Cell Biol Lipids. (2015) 1851:1428–41. doi: 10.1016/j.bbalip.2015.08.006
18. Cahu CL, Gisbert E, Villeneuve LAN, Morais S, Hamza N, Wold PA, et al. Influence of dietary phospholipids on early ontogenesis of fish. Aquac Res. (2009) 40:989–99. doi: 10.1111/j.1365-2109.2009.02190.x
19. Rossi M. “Use of lecithin and lecithin fractions,” In: Huopalahti R, López-Fandiño R, Anton M, Schade R, editors. Bioactive Egg Compounds. Berlin Heidelberg: Springer-Verlag (2007). p. 229–239.
20. Wu Y, Wang T. Soybean lecithin fractionation and functionality. J Am Oil Chem Soc. (2003) 80:319–26. doi: 10.1007/s11746-003-0697-x
21. Hertrampf JW, Piedad-Pascual F. “Soya lecithin,” In: Handbook on Ingredients for Aquaculture Fe, eds. Dordrecht: Springer (2000). p. 383–395.
22. Shurtleff W, Aoyagi A. History of Lecithin and Phospholipids (1850-2016). Lafayette, CA: Soyinfo Center (2016), 954.
23. Ahuja K, Malkani T. Lecithin Market Share, Size and Statistics Report 2030. (2022). Available online at: https://www.gminsights.com/industry-analysis/lecithin-market (accessed December 12, 2022).
24. Yu HY, Park SE, Chun HS, Rho JR, Ahn S. Phospholipid composition analysis of krill oil through HPLC with ELSD: development, validation, and comparison with 31P NMR spectroscopy. J Food Compost Anal. (2022) 107:104408. doi: 10.1016/j.jfca.2022.104408
25. Fernandes GD, Alberici RM, Pereira GG, Cabral EC, Eberlin MN, Barrera-Arellano D. Direct characterization of commercial lecithins by easy ambient sonic-spray ionization mass spectrometry. Food Chem. (2012) 135:1855–60. doi: 10.1016/j.foodchem.2012.06.072
26. Arab Tehrany E, Kahn CJF, Baravian C, Maherani B, Belhaj N, Wang X, et al. Elaboration and characterization of nanoliposome made of soya; rapeseed and salmon lecithins: application to cell culture. Colloids Surf B Biointerfaces. (2012) 95:75–81. doi: 10.1016/j.colsurfb.2012.02.024
27. Topuz OK, Aygün T, Alp AC, Yatmaz HA, Torun M, Yerlikaya P. Characterization and emulsifying properties of aquatic lecithins isolated from processing discard of rainbow trout fish and its eggs. Food Chem. (2021) 339:128103. doi: 10.1016/j.foodchem.2020.128103
28. Lončarević I, Pajin B, Petrović J, Zarić D, Sakač M, Torbica A, et al. The impact of sunflower and rapeseed lecithin on the rheological properties of spreadable cocoa cream. J Food Eng. (2016) 171:67–77. doi: 10.1016/j.jfoodeng.2015.10.001
29. Cabezas DM, Madoery R, Diehl BWK, Tomás MC. Emulsifying properties of different modified sunflower lecithins. J Am Oil Chem Soc. (2012) 89:355–61. doi: 10.1007/s11746-011-1915-8
30. Bot F, Cossuta D, O'Mahony JA. Inter-relationships between composition, physicochemical properties and functionality of lecithin ingredients. Trends Food Sci Technol. (2021) 111:261–70. doi: 10.1016/j.tifs.2021.02.028
31. Xie M, Dunford NT. Fractionating of canola lecithin from acid degumming and its effect. Food Chem. (2019) 300:125217. doi: 10.1016/j.foodchem.2019.125217
32. Belayneh HD, Wehling RL, Cahoon E, Ciftci ON. Lipid composition and emulsifying properties of Camelina sativa seed lecithin. Food Chem. (2018) 242:139–46. doi: 10.1016/j.foodchem.2017.08.082
33. Xia ZW, Zhang JG, Ni ZJ, Zhang F, Thakur K, Hu F, Wei ZJ. Functional and emulsification characteristics of phospholipids and derived o/w emulsions from peony seed meal. Food Chem. (2022) 389:133112. doi: 10.1016/j.foodchem.2022.133112
34. Liu H, Liu T, Fan H, Gou M, Li G, Ren H, et al. Corn lecithin for injection from deoiled corn germ: extraction, composition, and emulsifying properties. Eur J Lipid Sci. (2018) 120:1700288. doi: 10.1002/ejlt.201700288
35. Sun X, Zhang L, Tian S, Yang K, Xie J. Phospholipid composition and emulsifying properties of rice bran lecithin from enzymatic degumming. LWT. (2020) 117:108588. doi: 10.1016/j.lwt.2019.108588
36. Pragasam A, Indira TN, Gopala Krishna AG. Preparation and physico chemical characteristics evaluation of rice bran lecithin in relation to soya lecithin. Beverage Food World. (2002) 12:19–22.
37. Palacios LE, Wang T. Egg-yolk lipid fractionation and lecithin characterization. J Am Oil Chem Soc. (2005) 82:571–8. doi: 10.1007/s11746-005-1111-4
38. Rombaut R, Dewettinck K. Properties, analysis and purification of milk polar lipids. Int Dairy J. (2006) 16:1362–73. doi: 10.1016/j.idairyj.2006.06.011
39. Price N, Fei T, Clark S, Wang T. Extraction of phospholipids from a dairy by-product (whey protein phospholipid concentrate) using ethanol. J Dairy Sci. (2018) 101:8778–87. doi: 10.3168/jds.2018-14950
40. Burling H, Graverholt G. Milk - a new source for bioactive phospholipids for use in food formulations. Lipid Technol. (2008) 20:229–31. doi: 10.1002/lite.200800058
41. Schneider M. “Marine phospholipids and their applications: next-generation omega-3 lipids,” In: De Meester F, Watson R, Zibadi S, editors. Omega-6/3 Fatty Acids: Functions, Sustainability Strategies and Perspectives. New Jersey, NJ: Humana Press Inc. (2013), p. 297–308.
42. Asaduzzaman AKM, Chun BS. Quality characteristics of lecithin isolated from deoiled mackerel (Scomber japonicus) muscle using different methods. J Ind Eng Chem. (2015) 21:620–6. doi: 10.1016/j.jiec.2014.03.029
43. Lee S-M, Asaduzzaman AKM, Chun B-S. Characterization of lecithin isolated from anchovy (Engraulis japonica) residues deoiled by supercritical carbon dioxide and organic solvent extraction. J Food Sci. (2012) 77:C773–8. doi: 10.1111/j.1750-3841.2012.02764.x
44. Belhaj N, Arab-Tehrany E, Linder M. Oxidative kinetics of salmon oil in bulk and in nanoemulsion stabilized by marine lecithin. Process Biochemistry. (2010) 45:187–95. doi: 10.1016/j.procbio.2009.09.005
45. Ahmmed MK, Carne A, Ahmmed F, Stewart I, (Sabrina) Tian H, Bekhit AEDA. Positional distribution of fatty acids and phospholipid composition in King salmon (Oncorhynchus tshawytscha) head, roe and skin using nuclear magnetic resonance spectroscopy. Food Chem. (2021) 363:130302. doi: 10.1016/j.foodchem.2021.130302
46. Ahmmed MK, Ahmmed F, Stewart I, Carne A, Tian H (Sabrina), Bekhit AEDA. Omega-3 phospholipids in Pacific blue mackerel (Scomber australasicus) processing by-products. Food Chem. (2021) 353:451. doi: 10.1016/j.foodchem.2021.129451
47. Uddin MS, Kishimura H, Chun BS. Isolation and characterization of lecithin from squid (Todarodes pacificus) viscera deoiled by supercritical carbon dioxide extraction. J Food Sci. (2011) 76:C350–4. doi: 10.1111/j.1750-3841.2010.02039.x
48. Yang B, Zhou R, Yang JG, Wang YH, Wang WF. Insight into the enzymatic degumming process of soybean oil. J Am Oil Chem Soc. (2008) 85:421–5. doi: 10.1007/s11746-008-1225-y
49. Lordan R, Tsoupras A, Zabetakis I. Phospholipids of animal and marine origin: structure, function, and anti-inflammatory properties. Molecules. (2017) 22:1964. doi: 10.3390/molecules22111964
50. Szuhaj BF. “Phospholipids: properties and occurrence,” In: Caballero B, Finglas PM, Toldra F, editors. Encyclopedia of Food and Health. Kidlington, United Kingdom: Academic Press (2016), p. 360–364.
51. Nguyen MT, Van De Walle D, Petit C, Beheydt B, Depypere F, Dewettinck K. Mapping the chemical variability of vegetable lecithins. J Am Oil Chem Soc. (2014) 91:1093–101. doi: 10.1007/s11746-014-2455-9
52. Pokorný J. “Production, separation and modification of phospholipids for use in food.” In: Gunstone FD, editor. Modifying Lipids for Use in Food. Cambridge: Woodhead Publishing (2006), p. 369–390.
53. Gong H, Lawrence AL, Jiang DH, Castille FL, Gatlin DM. Lipid nutrition of juvenile Litopenaeus vannamei: I. Dietary cholesterol and de-oiled soy lecithin requirements and their interaction. Aquaculture. (2000) 190:305–24. doi: 10.1016/S0044-8486(00)00414-2
54. Sun N, Chen J, Wang D, Lin S. Advance in food-derived phospholipids: Sources, molecular species and structure as well as their biological activities. Trends Food Sci Technol. (2018) 80:199–211. doi: 10.1016/j.tifs.2018.08.010
55. Turchini GM, Francis DS, Du Z-Y, Olsen RE, Ringø E, Tocher DR. “The lipids,” In: Hardy RW, Kaushik SJ, editors. Fish Nutrition. Amsterdam: Elsevier (2022). p. 303–467.
56. Jaxion-Harm J. Effects of dietary phospholipids on early stage Atlantic Salmon (Salmo salar) performance: a comparison among phospholipid sources. Aquaculture. (2021) 544:737055. doi: 10.1016/j.aquaculture.2021.737055
57. Zhao J, Ai Q, Mai K, Zuo R, Luo Y. Effects of dietary phospholipids on survival, growth, digestive enzymes and stress resistance of large yellow croaker, Larmichthys crocea larvae. Aquaculture. (2013) 411:122–8. doi: 10.1016/j.aquaculture.2013.05.018
58. Kumar N, Minhas PS, Ambasankar K, Krishnani KK, Rana RS. Dietary lecithin potentiates thermal tolerance and cellular stress protection of milk fish (Chanos chanos) reared under low dose endosulfan-induced stress. J Therm Biol. (2014) 46:40–6. doi: 10.1016/j.jtherbio.2014.10.004
59. Li Y, Gao J, Huang S. Effects of different dietary phospholipid levels on growth performance, fatty acid composition, PPAR gene expressions and antioxidant responses of blunt snout bream Megalobrama amblycephala fingerlings. Fish Physiol Biochem. (2015) 41:423–36. doi: 10.1007/s10695-014-9994-8
60. Huang Y, Xu J, Sheng Z, Chen N, Li S. Integrated response of growth performance, fatty acid composition, antioxidant responses and lipid metabolism to dietary phospholipids in hybrid grouper (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂) larvae. Aquaculture. (2021) 541:736728. doi: 10.1016/j.aquaculture.2021.736728
61. Gisbert E, Villeneuve L, Zambonino-Infante JL, Quazuguel P, Cahu CL. Dietary phospholipids are more efficient than neutral lipids for long-chain polyunsaturated fatty acid supply in European sea bass Dicentrarchus labrax larval development. Lipids. (2005) 40:609–18. doi: 10.1007/s11745-005-1422-0
62. Daprà F, Geurden I, Corraze G, Bazin D, Zambonino-Infante JL, Fontagné-Dicharry S. Physiological and molecular responses to dietary phospholipids vary between fry and early juvenile stages of rainbow trout (Oncorhynchus mykiss). Aquaculture. (2011) 319:377–84. doi: 10.1016/j.aquaculture.2011.07.016
63. Cai Z, Feng S, Xiang X, Mai K, Ai Q. Effects of dietary phospholipid on lipase activity, antioxidant capacity and lipid metabolism-related gene expression in large yellow croaker larvae (Larimichthys crocea). Comp Biochem Physiol B Biochem Mol Biol. (2016) 201:46–52. doi: 10.1016/j.cbpb.2016.06.007
64. Rampone AJ, Machida CM. Mode of action of lecithin in suppressing cholesterol absorption. J Lipid Res. (1981) 22:744–52. doi: 10.1016/S0022-2275(20)37345-4
65. Teshima S, Kanazawa A, Kakuta Y. Growth, survival, and body lipid composition of the prawn larvae receiving several dietary phospholipids. Memoir Faculty Fisheries Kagoshima Univ. (1986) 35:17–27.
66. Paibulkichakul C, Piyatiratitivorakul S, Kittakoop P, Viyakarn V, Fast AW, Menasveta P. Optimal dietary levels of lecithin and cholesterol for black tiger prawn Penaeus monodon larvae and postlarvae. Aquaculture. (1998) 167:273–81. doi: 10.1016/S0044-8486(98)00327-5
67. Ghosh TK, Chauhan YH, Mandal RN. Growth performance of Labeo bata (Hamilton, 1822) in freshwater and its acclimatization in brackish water with betaine as feed additive. Aquaculture. (2019) 501:128–34. doi: 10.1016/j.aquaculture.2018.11.020
68. Kumar N, Jadhao SB, Chandan NK, Kumar K, Jha AK, Bhushan S, et al. Dietary choline, betaine and lecithin mitigates endosulfan-induced stress in Labeo rohita fingerlings. Fish Physiol Biochem. (2012) 38:989–1000. doi: 10.1007/s10695-011-9584-y
69. Frontiera MS, Stabler SP, Kolhouse JF, Allen RH. Regulation of methionine metabolism: Effects of nitrous oxide and excess dietary methionine. J Nutr Biochem. (1994) 5:28–38. doi: 10.1016/0955-2863(94)90006-X
70. Geng H, Yang P, Chen Y, Qin Y, Li X, He C, et al. Dietary choline can partially spare methionine to improve the feeds utilization and immune response in juvenile largemouth bass (Micropterus salmoides): Based on phenotypic response to gene expression. Aquac Rep. (2023) 30:101546. doi: 10.1016/j.aqrep.2023.101546
71. Newberne PM. The methyl-deficiency model: History characteristics and research directions. J Nutr Biochem. (1993) 4:618–24. doi: 10.1016/0955-2863(93)90032-R
72. Kidd MT, Ferket PR, Garlich JD. Nutritional and osmoregulatory functions of betaine. Worlds Poult Sci J. (1997) 53:125–39. doi: 10.1079/WPS19970013
73. Hung SSO, Berge GM, Storebakken T. Growth and digestibility effects of soya lecithin and choline chloride on juvenile Atlantic salmon. Aquac Nutr. (1997) 3:141–4. doi: 10.1046/j.1365-2095.1997.00080.x
74. Cahu CL, Infante JLZ, Barbosa V. Effect of dietary phospholipid level and phospholipid: neutral lipid value on the development of sea bass (Dicentrarchus labrax) larvae fed a compound diet. Br J Nutr. (2003) 90:21–8. doi: 10.1079/BJN2003880
75. Jafari F, Noori F, Agh N, Estevez A, Ghasemi A, Alcaraz C, et al. Phospholipids improve the performance, physiological, antioxidative responses and, lpl and igf1 gene expressions in juvenile stellate sturgeon (Acipenser stellatus). Aquaculture. (2021) 541:736809. doi: 10.1016/j.aquaculture.2021.736809
76. Torfi Mozanzadeh M, Marammazi JG, Yaghoubi M, Agh N, Pagheh E, Gisbert E. Macronutrient requirements of Silvery-Black porgy (Sparidentex hasta): a comparison with other farmed Sparid species. Fishes. (2017) 2:5. doi: 10.3390/fishes2020005
77. Sivaramakrishnan T, Ambasankar K, Kumaraguru Vasagam KP, Syama Dayal J, Sandeep KP, Bera A, et al. Effect of dietary soy lecithin inclusion levels on growth, feed utilization, fatty acid profile, deformity and survival of milkfish (Chanos chanos) larvae. Aquac Res. (2021) 52:5366–74. doi: 10.1111/are.15406
78. Maleki Moghaddam MR, Agh N, Sarvi Moghanlou K, Noori F, Taghizadeh A, Gisbert E. Dietary conjugated linoleic acid (CLA) and lecithin affects levels of serum cholesterol, triglyceride, lipoprotein and hypoxic stress resistance in rainbow trout (Oncorhynchus mykiss). Int J Aquat Biol. (2021) 9:124–33. doi: 10.22034/ijab.v9i2.544
79. Azarm HM, Kenari AA, Hedayati M. Effect of dietary phospholipid sources and levels on growth performance, enzymes activity, cholecystokinin and lipoprotein fractions of rainbow trout (Oncorhynchus mykiss) fry. Aquac Res. (2013) 44:634–44. doi: 10.1111/j.1365-2109.2011.03068.x
80. Taylor JF, Martinez-Rubio L, Del Pozo J, Walton JM, Tinch AE, Migaud H, et al. Influence of dietary phospholipid on early development and performance of Atlantic salmon (Salmo salar). Aquaculture. (2015) 448:262–72. doi: 10.1016/j.aquaculture.2015.06.012
81. Kanazawa A, Teshima S, Inamori S, Matsubara H. Effects of dietary phospholipids on growth of the larval red sea bream and knife jaw. Memoir Faculty Fish Kagoshima Univ. (1983) 32:109–114.
82. Kanazawa A, Teshima S, Kobayashi T, Takae M, Iwashita T, Uehara R. Necessity of dietary phospholipids for growth of the larval ayu. Memoir Faculty Fish Kagoshima Univ. (1983) 32:115–120.
83. Poston HA. Performance of rainbow trout fry fed supplemental soy lecithin and choline. The Progr Fish-Culturist. (1990) 52:218–25.
84. Poston HA. Effect of body size on growth, survival, and chemical composition of Atlantic salmon fed soy lecithin and choline. The Prog Fish-Culturist. (1990) 52:226–30.
85. Poston HA. Response of Atlantic salmon fry to feed-grade lecithin and choline. The Prog Fish-Culturist. (1991) 53:224–8. doi: 10.1577/1548-8640(1991)053<0224:ROASFT>2.3.CO;2
86. De Santis C, Taylor JF, Martinez-Rubio L, Boltana S, Tocher DR. Influence of development and dietary phospholipid content and composition on intestinal transcriptome of Atlantic salmon (Salmo salar). PLoS ONE. (2015) 10:e0140964. doi: 10.1371/journal.pone.0140964
87. Craig SR, Gatlin DM. Growth and body composition of juvenile red drum (Sciaenops ocellatus) fed diets containing lecithin and supplemental choline. Aquaculture. (1997) 151:259–67. doi: 10.1016/S0044-8486(96)01479-2
88. Lochmann R, Brown R. Soybean-lecithin supplementation of practical diets for juvenile goldfish (Carassius auratus). J Am Oil Chem Soc. (1997) 74:149–52. doi: 10.1007/s11746-997-0160-7
89. Geurden I, Charlon N, Marion D, Bergot P. Influence of purified soybean phospholipids on early development of carp. Aquac Int. (1997) 5:137–149.
90. Uyan O, Koshio S, Ishikawa M, Uyan S, Ren T, Yokoyama S, et al. Effects of dietary phosphorus and phospholipid level on growth, and phosphorus deficiency signs in juvenile Japanese flounder, Paralichthys olivaceus. Aquaculture. (2007) 267:44–54. doi: 10.1016/j.aquaculture.2007.01.020
91. Niu J, Liu YJ, Tian LX, Mai KS, Yang HJ, Ye CX, et al. Effects of dietary phospholipid level in cobia (Rachycentron canadum) larvae: growth, survival, plasma lipids and enzymes of lipid metabolism. Fish Physiol Biochem. (2008) 34:9–17. doi: 10.1007/s10695-007-9140-y
92. Uyan O, Koshio S, Ishikawa M, Yokoyama S, Uyan S, Ren T, et al. The influence of dietary phospholipid level on the performances of juvenile amberjack, Seriola dumerili, fed non-fishmeal diets. Aquac Nutr. (2009) 15:550–7. doi: 10.1111/j.1365-2095.2008.00621.x
93. Saleh R, Betancor MB, Roo J, Benítez-Dorta V, Zamorano MJ, Bell JG, et al. Effect of krill phospholipids versus soybean lecithin in microdiets for gilthead seabream (Sparus aurata) larvae on molecular markers of antioxidative metabolism and bone development. Aquac Nutr. (2015) 21:474–88. doi: 10.1111/anu.12177
94. Saleh NE, Wassef EA, Kamel MA, El-Haroun ER, El-Tahan RA. Beneficial effects of soybean lecithin and vitamin C combination in fingerlings gilthead seabream (Sparus aurata) diets on; fish performance, oxidation status and genes expression responses. Aquaculture. (2022) 546:737345. doi: 10.1016/j.aquaculture.2021.737345
95. Pagheh E, Agh N, Marammazi JG, Nouri F, Sepahdari A, Gisbert E, et al. Dietary soybean lecithin affects growth performance, fillet biochemical composition and digestive enzyme activity in Sparidentex hasta juvenile. J Appl Anim Res. (2019) 47:24–33. doi: 10.1080/09712119.2018.1557663
96. El-Naggar K, Mohamed R. El-katcha MI, Abdo SE, Soltan MA. Plant ingredient diet supplemented with lecithin as fish meal and fish oil alternative affects growth performance, serum biochemical, lipid metabolism and growth-related gene expression in Nile tilapia. Aquac Res. (2021) 52:6308–21. doi: 10.1111/are.15494
97. Lin Y-H, Chen K-H, Wu P-C. Effects of the diet based on soybean meal supplemented with soy lecithin on growth, biochemical parameters and digestibility of nutrients in grouper, Epinephelus lanceolatus. Aquac Res. (2022) 53:700–6. doi: 10.1111/are.15593
98. Coutteau P, Geurden I, Camara MR, Bergot P, Sorgeloos P. Review on the dietary effects of phospholipids in fish and crustacean larviculture. Aquaculture. (1997) 155:149–64. doi: 10.1016/S0044-8486(97)00125-7
99. Sargent JR, Tocher DR, Bell JG. “The lipids,” In: Halver JE, Hardy RW, editors. Fish Nutrition. San Diego, CA: Elsevier (Academic Press) (2011). p. 181–257.
100. Tan P, Zhang P, Zhang L, Zhu W, Wang L, Chen R, et al. Effects of soybean lecithin on growth performance, intestine morphology, and liver tissue metabolism in rock bream (Oplegnathus fasciatus) larvae. Front Mar Sci. (2022) 9:942259. doi: 10.3389/fmars.2022.942259
101. Hamza N, Mhetli M, Khemis I. Ben, Cahu C, Kestemont P. Effect of dietary phospholipid levels on performance, enzyme activities and fatty acid composition of pikeperch (Sander lucioperca) larvae. Aquaculture. (2008) 275:274–82. doi: 10.1016/j.aquaculture.2008.01.014
102. Llorente I, Fernández-Polanco J, Baraibar-Diez E, Odriozola MD, Bjørndal T, Asche F, et al. Assessment of the economic performance of the seabream and seabass aquaculture industry in the European Union. Mar Policy. (2020) 117:103876. doi: 10.1016/j.marpol.2020.103876
103. Hadas E, Koven W, Sklan D, Tandler A. The effect of dietary phosphatidylcholine on the assimilation and distribution of ingested free oleic acid (18:1n–9) in gilthead seabream (Sparus aurata) larvae. Aquaculture. (2003) 217:577–88. doi: 10.1016/S0044-8486(02)00431-3
104. Olsen RE, Myklebust R, Kaino T. Ringø E. Lipid digestibility and ultrastructural changes in the enterocytes of Arctic char (Salvelinus alpinus L) fed linseed oil and soybean lecithin. Fish Physiol Biochem. (1999) 21:35–44. doi: 10.1023/A:1007726615889
105. El-Sayed AFM, Tammam MS, Makled SO. Lecithin-containing bioemulsifier boosts growth performance, feed digestion and absorption and immune response of adult Nile tilapia (Oreochromis niloticus). Aquac Nutr. (2021) 27:757–70. doi: 10.1111/anu.13221
106. Koven WM, Parra G, Kolkovski S, Tandler A. The effect of dietary phosphatidylcholine and its constituent fatty acids on microdiet ingestion and fatty acid absorption rate in gilthead sea bream, Sparus auratus, larvae. Aquac Nutr. (1998) 4:39–45. doi: 10.1046/j.1365-2095.1998.00101.x
107. Koven WM, Kolkovski S, Tandler A, Kissil GW, Sklan D. The effect of dietary lecithin and lipase, as a function of age, on n-9 fatty acid incorporation in the tissue lipids of Sparus aurata larvae. Fish Physiol Biochem. (1993) 10:357–64. doi: 10.1007/BF00004502
108. Kanazawa A, Teshima SI, Sakamoto M. Effects of dietary lipids, fatty acids, and phospholipids on growth and survival of prawn (Penaeus japonicus) larvae. Aquaculture. (1985) 50:39–49. doi: 10.1016/0044-8486(85)90151-6
109. Thongrod S, Boonyaratpalin M. Cholesterol and lecithin requirement of juvenile banana shrimp, Penaeus merguiensis. Aquaculture. (1998) 161:315–21. doi: 10.1016/S0044-8486(97)00278-0
110. Kumaraguru Vasagam KP, Ramesh S, Balasubramanian T. Dietary value of different vegetable oil in black tiger shrimp Penaeus monodon in the presence and absence of soy lecithin supplementation: effect on growth, nutrient digestibility and body composition. Aquaculture. (2005) 250:317–27. doi: 10.1016/j.aquaculture.2005.02.035
111. Coutteau P, Camara MR, Sorgeloos P. The effect of different levels and sources of dietary phosphatidylcholine on the growth, survival, stress resistance, and fatty acid composition of postlarval Penaeus vannamei. Aquaculture. (1996) 147:261–73. doi: 10.1016/S0044-8486(96)01387-7
112. Hu Y, Tan B, Mai K, Ai Q, Zhang L, Zheng S. Effects of dietary menhaden oil, soybean oil and soybean lecithin oil at different ratios on growth, body composition and blood chemistry of juvenile Litopenaeus vannamei. Aquac Int. (2011) 19:459–73. doi: 10.1007/s10499-010-9361-4
113. Liou C-H, To V-A, Zhang Z-F, Lin Y-H. The effect of dietary lecithin and lipid levels on the growth performance, body composition, hemolymph parameters, immune responses, body texture, and gene expression of juvenile white shrimp (Litopenaeus vannamei). Aquaculture. (2023) 567:739260. doi: 10.1016/j.aquaculture.2023.739260
114. Briggs MRP, Jauncey K, Brown JH. The cholesterol and lecithin requirements of juvenile prawn (Macrobrachium rosenbergii) fed semi-purified diets. Aquaculture. (1988) 70:121–9. doi: 10.1016/0044-8486(88)90011-7
115. Hien TTT, Hai TN, Phuong NT, Ogata HY, Wilder MN. The effects of dietary lipid sources and lecithin on the production of giant freshwater prawn Macrobrachium rosenbergii larvae in the Mekong Delta region of Vietnam. Fisheries Sci. (2005) 71:279–86. doi: 10.1111/j.1444-2906.2005.00961.x
116. Teshima S, Kanazawa A, Kakuta Y. Effects of dietary phospholipids on growth and body composition of the juvenile prawn. Bullet Soc Sci Fish. (1986) 52:155–8. doi: 10.2331/suisan.52.155
117. Holme MH, Southgate PC, Zeng C. Assessment of dietary lecithin and cholesterol requirements of mud crab, Scylla serrata, megalopa using semi-purified microbound diets. Aquac Nutr. (2007) 13:413–23. doi: 10.1111/j.1365-2095.2007.00492.x
118. Li X, Wang J, Han T, Hu S, Jiang Y, Wang C. Effect of dietary phospholipids levels and sources on growth performance, fatty acid composition of the juvenile swimming crab, Portunus trituberculatus. Aquaculture. (2014) 430:166–72. doi: 10.1016/j.aquaculture.2014.03.037
119. Wang JT, Han T, Li XY, Hu SX, Jiang YD, Wang CL. Effects of dietary phosphatidylcholine (PC) levels on the growth, molt performance and fatty acid composition of juvenile swimming crab, Portunus trituberculatus. Anim Feed Sci Technol. (2016) 216:225–33. doi: 10.1016/j.anifeedsci.2016.03.023
120. Lin Z, Han F, Lu J, Guo J, Qi C, Wang C, et al. Influence of dietary phospholipid on growth performance, body composition, antioxidant capacity and lipid metabolism of Chinese mitten crab, Eriocheir sinensis. Aquaculture. (2020) 516:734653. doi: 10.1016/j.aquaculture.2019.734653
121. Lin Z, Bu X, Wang N, Lei Y, Liu S, Wang X, et al. Dietary phospholipid alleviates the adverse effects of high-lipid diet in Chinese mitten crab (Eriocheir sinensis). Aquaculture. (2021) 531:735899. doi: 10.1016/j.aquaculture.2020.735899
122. Sui LY, Wu XG, Wille M, Cheng YX, Sorgeloos P. Effect of dietary soybean lecithin on reproductive performance of Chinese mitten crab Eriocheir sinensis (H. Milne-Edwards) broodstock. Aquac Int. (2009) 17:45–56. doi: 10.1007/s10499-008-9178-6
123. Ding LY, Jin M, Sun P, Lu Y, Ma HN, Yuan Y, et al. Cloning, tissue expression of the fatty acid-binding protein (Pt-FABP1) gene, and effects of dietary phospholipid levels on fabp and vitellogenin gene expression in the female swimming crab Portunus trituberculatus. Aquaculture. (2017) 474:57–65. doi: 10.1016/j.aquaculture.2017.03.029
124. Song D, Shi B, Ding L, Jin M, Sun P, Jiao L, et al. Regulation of dietary phospholipids on growth performance, antioxidant activities, phospholipid metabolism and vitellogenesis in pre reproductive phase of female swimming crabs, Portunus trituberculatus. Aquaculture. (2019) 511:734230. doi: 10.1016/j.aquaculture.2019.734230
125. Wang L, Zuo D, Lv W, Li J, Wang Q, Zhao Y. Effects of dietary soybean lecithin on gonadal development and vitellogenin mRNA expression in the female redclaw crayfish Cherax quadricarinatus (von Martens) at first maturation. Aquac Res. (2013) 44:1167–76. doi: 10.1111/j.1365-2109.2012.03128.x
126. Zhang F, Ning Y, Yuan R, Ding J, Chang Y, Zuo R. Effects of soya lecithin addition on the growth, antioxidant capacity, gonad development and nutritional quality of adult sea urchin (Strongylocentrotus intermedius). Aquac Rep. (2022) 22:100990. doi: 10.1016/j.aqrep.2021.100990
127. Gibbs VK, Watts SA, Lawrence AL, Lawrence JM. Dietary phospholipids affect growth and production of juvenile sea urchin Lytechinus variegatus. Aquaculture. (2009) 292:95–103. doi: 10.1016/j.aquaculture.2009.03.046
128. Peng S, Chen L, Qin JG, Hou J, Yu N, Long Z, et al. Effects of replacement of dietary fish oil by soybean oil on growth performance and liver biochemical composition in juvenile black seabream, Acanthopagrus schlegeli. Aquaculture. (2008) 276:154–61. doi: 10.1016/j.aquaculture.2008.01.035
129. Seiliez I, Bruant JS, Zambonino Infante JL, Kaushik S, Bergot P. Effect of dietary phospholipid level on the development of gilthead sea bream (Sparus aurata) larvae fed a compound diet. Aquac Nutr. (2006) 12:372–8. doi: 10.1111/j.1365-2095.2006.00436.x
130. Balito-Liboon JS, Ferdinand R, Traifalgar M, Pagapulan MJBB, Mameloco EJG, Temario EE. Dietary soybean lecithin enhances growth performance, feed utilization efficiency and body composition of early juvenile milkfish, Chanos chanos. Israeli J Aquac Bamidgeh. (2018) 70:20297. doi: 10.46989/001c.20927
131. Alves Martins D, Estévez A, Stickland NC, Simbi BH, Yúfera M. Dietary lecithin source affects growth potential and gene expression in Sparus aurata larvae. Lipids. (2010) 45:1011–23. doi: 10.1007/s11745-010-3471-7
132. Salini MJ, Wade N, Bourne N, Turchini GM, Glencross BD. The effect of marine and non-marine phospholipid rich oils when fed to juvenile barramundi (Lates calcarifer). Aquaculture. (2016) 455:125–35. doi: 10.1016/j.aquaculture.2016.01.013
133. Liu J, Caballero MJ, Izquierdo M, El-Sayed Ali T, Hernández-Cruz CM, Valencia A, et al. Necessity of dietary lecithin and eicosapentaenoic acid for growth, survival, stress resistance and lipoprotein formation in gilthead sea bream Sparus aurata. Fisheries Sci. (2002) 68:1165–72. doi: 10.1046/j.1444-2906.2002.00551.x
134. Joshi KD, Das SCS, Pathak RK, Khan A, Sarkar UK, Roy K. Pattern of reproductive biology of the endangered golden mahseer Tor putitora (Hamilton 1822) with special reference to regional climate change implications on breeding phenology from lesser Himalayan region, India. J Appl Anim Res. (2018) 46:1289–95. doi: 10.1080/09712119.2018.1497493
135. Patra RW, Chapman JC, Lim RP, Gehrke PC. The effects of three organic chemicals on the upper thermal tolerances of four freshwater fishes. Environ Toxicol Chem. (2007) 26:1454–9. doi: 10.1897/06-156R1.1
136. Cengiz EI, Unlu E. Histopathological changes in the gills of mosquitofish, Gambusia affinis exposed to endosulfan. Bull Environ Contam Toxicol. (2002) 68:290–6. doi: 10.1007/s001280251
137. Pawar AP, Sanaye SV, Shyama S, Sreepada RA, Dake AS. Effects of salinity and temperature on the acute toxicity of the pesticides, dimethoate and chlorpyrifos in post-larvae and juveniles of the whiteleg shrimp. Aquac Rep. (2020) 16:100240. doi: 10.1016/j.aqrep.2019.100240
138. Ciji A, Akhtar MS, Tripathi PH, Pandey A, Rajesh M, Kamalam BS. Dietary soy lecithin augments antioxidative defense and thermal tolerance but fails to modulate non-specific immune genes in endangered golden mahseer (Tor putitora) fry. Fish Shellfish Immunol. (2021) 109:34–40. doi: 10.1016/j.fsi.2020.11.031
139. Beitinger TL, Bennett WA, McCauley RW. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature. Environ Biol Fishes. (2000) 58:237–75. doi: 10.1023/A:1007676325825
140. Das T, Pal AK, Chakraborty SK, Manush SM, Chatterjee N, Mukherjee SC. Thermal tolerance and oxygen consumption of Indian Major Carps acclimated to four temperatures. J Therm Biol. (2004) 29:157–63. doi: 10.1016/j.jtherbio.2004.02.001
141. Díaz F, Re AD, González RA, Sánchez LN, Leyva G, Valenzuela F. Temperature preference and oxygen consumption of the largemouth bass Micropterus salmoides (Lacépède) acclimated to different temperatures. Aquac Res. (2007) 38:1387–94. doi: 10.1111/j.1365-2109.2007.01817.x
142. Sarma K, Pal AK, Ayyappan S, Das T, Manush SM, Debnath D, et al. Acclimation of Anabas testudineus (Bloch) to three test temperatures influences thermal tolerance and oxygen consumption. Fish Physiol Biochem. (2010) 36:85–90. doi: 10.1007/s10695-008-9293-3
143. Nakano K, Iwama GK. The 70-kDa heat shock protein response in two intertidal sculpins, Oligocottus maculosus and O. snyderi: relationship of hsp70 and thermal tolerance. Comp Biochem Physiol Mol Integr Physiol. (2002) 133:79–94. doi: 10.1016/S1095-6433(02)00115-0
144. Werner I, Smith TB, Feliciano J, Johnson ML. Heat shock proteins in juvenile steelhead reflect thermal conditions in the Navarro River watershed, California. Trans Am Fish Soc. (2005) 134:399–410. doi: 10.1577/T03-181.1
145. Das T, Pal AK, Chakraborty SK, Manush SM, Chatterjee N, Apte SK. Metabolic elasticity and induction of heat shock protein 70 in Labeo rohita acclimated to three temperatures. Asian-Australas J Anim Sci. (2006) 19:1033–9. doi: 10.5713/ajas.2006.1033
146. Fontagné S, Geurden I, Escaffre AM, Bergot P. Histological changes induced by dietary phospholipids in intestine and liver of common carp (Cyprinus carpio L). larvae. Aquaculture. (1998) 161:213–23. doi: 10.1016/S0044-8486(97)00271-8
147. Wang S, Han Z, Turchini GM, Wang X, Fang Z, Chen N, et al. Effects of dietary phospholipids on growth performance, digestive enzymes activity and intestinal health of largemouth bass (Micropterus salmoides) larvae. Front Immunol. (2021) 12:827946. doi: 10.3389/fimmu.2021.827946
148. Tan P, Zhu W, Zhang P, Wang L, Chen R, Xu D. Dietary soybean lecithin inclusion promotes growth, development, and intestinal morphology of yellow drum (Nibea albiflora) larvae. Aquaculture. (2022) 559:738446. doi: 10.1016/j.aquaculture.2022.738446
149. Huang Y, Xu J, Sheng Z, Xie R, Zhang H, Chen N, Li S. Effects of dietary phospholipids on growth performance, fatty acid composition, and expression of lipid metabolism related genes of juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × E. lancolatus ♂). Aquac Rep. (2022) 22:100993. doi: 10.1016/j.aqrep.2021.100993
150. Zhang H, Mu Z, Xu L, Xu G, Liu M, Shan A. Dietary lipid level induced antioxidant response in Manchurian trout, Brachymystax lenok (Pallas) larvae. Lipids. (2009) 44:643–54. doi: 10.1007/s11745-009-3313-7
151. Mourente G, Diaz-Salvago E, Bell JG, Tocher DR. Increased activities of hepatic antioxidant defence enzymes in juvenile gilthead sea bream (Sparus aurata L) fed dietary oxidised oil: attenuation by dietary vitamin E. Aquaculture. (2002) 214:343–61. doi: 10.1016/S0044-8486(02)00064-9
152. Gao J, Koshio S, Wang W, Li Y, Huang S, Cao X. Effects of dietary phospholipid levels on growth performance, fatty acid composition and antioxidant responses of Dojo loach Misgurnus anguillicaudatus larvae. Aquaculture. (2014) 427:304–9. doi: 10.1016/j.aquaculture.2014.02.022
153. Adel M, Gholaghaie M, Khanjany P, Citarasu T. Effect of dietary soybean lecithin on growth parameters, digestive enzyme activity, antioxidative status and mucosal immune responses of common carp (Cyprinus carpio). Aquac Nutr. (2017) 23:1145–52. doi: 10.1111/anu.12483
154. Lin SM Li FJ, Yuangsoi B, Doolgindachbaporn S. Effect of dietary phospholipid levels on growth, lipid metabolism, and antioxidative status of juvenile hybrid snakehead (Channa argus × Channa maculata). Fish Physiol Biochem. (2018) 44:401–10. doi: 10.1007/s10695-017-0443-3
155. Jafari F, Agh N, Noori F, Tokmachi A, Gisbert E. Effects of dietary soybean lecithin on growth performance, blood chemistry and immunity in juvenile stellate sturgeon (Acipenser stellatus). Fish Shellfish Immunol. (2018) 80:487–96. doi: 10.1016/j.fsi.2018.06.023
156. Jenabi Haghparast R, Sarvi Moghanlou K, Mohseni M, Imani A. Effect of dietary soybean lecithin on fish performance, hemato-immunological parameters, lipid biochemistry, antioxidant status, digestive enzymes activity and intestinal histomorphometry of pre-spawning Caspian brown trout (Salmo trutta caspius). Fish Shellfish Immunol. (2019) 91:50–7. doi: 10.1016/j.fsi.2019.05.022
Keywords: lecithin, growth performance, feed utilization, immunity, antioxidant, stressor, sustainable aquaculture
Citation: Wee W, Téllez-Isaías G, Abdul Kari Z, Cheadoloh R, Kabir MA, Mat K, Mohamad Sukri SA, Rahman MM, Rusli ND and Wei LS (2023) The roles of soybean lecithin in aquafeed: a crucial need and update. Front. Vet. Sci. 10:1188659. doi: 10.3389/fvets.2023.1188659
Received: 17 March 2023; Accepted: 28 August 2023;
Published: 15 September 2023.
Edited by:
Rantao Zuo, Dalian Ocean University, ChinaReviewed by:
Roberto Senas Cuesta, University of Arkansas, United StatesCopyright © 2023 Wee, Téllez-Isaías, Abdul Kari, Cheadoloh, Kabir, Mat, Mohamad Sukri, Rahman, Rusli and Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zulhisyam Abdul Kari, enVsaGlzeWFtLmFAdW1rLmVkdS5teQ==; Lee Seong Wei, bGVlc2VvbmdAdW1rLmVkdS5teQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.