 Flor Y. Ramírez-Castillo
Flor Y. Ramírez-Castillo Alma L. Guerrero-Barrera
Alma L. Guerrero-Barrera Francisco J. Avelar-González
Francisco J. Avelar-González
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci., 15 June 2023
Sec. Veterinary Epidemiology and Economics
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1158588
This article is part of the Research TopicLivestock and its role in the emergence, spread, and evolution of antimicrobial resistance: Animal-to-human or animal-to-environment transmissionView all 10 articles
Carbapenem resistance (CR) is a major global health concern. CR is a growing challenge in clinical settings due to its rapid dissemination and low treatment options. The characterization of its molecular mechanisms and epidemiology are highly studied. Nevertheless, little is known about the spread of CR in food-producing animals, seafood, aquaculture, wildlife, their environment, or the health risks associated with CR in humans. In this review, we discuss the detection of carbapenem-resistant organisms and their mechanisms of action in pigs, cattle, poultry, seafood products, companion animals, and wildlife. We also pointed out the One Health approach as a strategy to attempt the emergency and dispersion of carbapenem-resistance in this sector and to determine the role of carbapenem-producing bacteria in animals among human public health risk. A higher occurrence of carbapenem enzymes in poultry and swine has been previously reported. Studies related to poultry have highlighted P. mirabilis, E. coli, and K. pneumoniae as NDM-5- and NDM-1-producing bacteria, which lead to carbapenem resistance. OXA-181, IMP-27, and VIM-1 have also been detected in pigs. Carbapenem resistance is rare in cattle. However, OXA- and NDM-producing bacteria, mainly E. coli and A. baumannii, are cattle's leading causes of carbapenem resistance. A high prevalence of carbapenem enzymes has been reported in wildlife and companion animals, suggesting their role in the cross-species transmission of carbapenem-resistant genes. Antibiotic-resistant organisms in aquatic environments should be considered because they may act as reservoirs for carbapenem-resistant genes. It is urgent to implement the One Health approach worldwide to make an effort to contain the dissemination of carbapenem resistance.
Carbapenems are broad-spectrum beta (β)-lactam antimicrobials primarily used to treat severe human infections. These antibiotics are considered one of the most reliable drugs and the last line of therapy for infections caused by multidrug-resistant Gram-negative and Gram-positive bacteria. Carbapenems possess a broad-spectrum antibacterial activity and have a structure defined by a carbapenem coupled with a β-lactam ring. In addition, these antibiotics contain a carbon instead of a sulfone in the fourth position of the thiazolidine moiety β-lactam ring, which confers protection against most β-lactamases (1).
The widespread use of these antibiotics has increased to a worldwide emergence of carbapenem-resistant organisms (CROs), which constitute a critical growing public health threat, mainly in hospital settings, as their prescription has escalated in recent years and used for treating life-threatening infections. Carbapenem-resistant Enterobacteriaceae (CRE) [i.e., Carbapenem-resistant Klebsiella pneumoniae (CR-Kp), Carbapenem-resistant Escherichia coli (CREc), Enterobacter spp., Serratia spp., and Proteus spp.] are some of the most critical CROs because they are associated with infections that lead to high mortality and have the potential to spread carbapenem resistance via mobile genetic elements (2). In addition, non-fermenting bacteria such as carbapenem-resistant Acinetobacter baumannii (CRAB) and carbapenem-resistant Pseudomonas aeruginosa (CRPA) have also emerged as critical CROs (3–5).
The four significant carbapenem mechanisms of resistance include the presence of β-lactamase enzymes called carbapenemases, which hydrolyze carbapenem antibiotics encoded on chromosomal or plasmid genes, the synergistic effect of other β-lactamases with bacterial cell membrane permeability due to alterations or mutations in porins, the low affinity of penicillin-binding proteins (PBPs) in different species, and the increased efflux pumps (6, 7).
Among carbapenem-producing (CP) microorganisms, different classes of carbapenemases are found under the Ambler classification (Table 1). Class A or serine carbapenemases can hydrolyze all β-lactams, carbapenems, cephalosporins, penicillins, and aztreonam but are inhibited by clavulanate and tazobactam. Additionally, a combination of the newly cephalosporin antibiotic, ceftaroline, and avibactam (ceftaroline/avibactam) has been shown to produce activity against Enterobacteriaceae KPC producers (19). Klebsiella pneumoniae carbapenemase (KPC), not metalloenzyme carbapenemase (NMC-A), imipenem-hydrolyzing beta-lactamase (IMI), and Serratia marcescens enzyme (SME) are representative of this class. KPC enzymes confer resistance to all β-lactamases and other types of antibiotics, such as quinolones and aminoglycosides. They are only partially inhibited by β-lactamase inhibitors such as clavulanic acid, tazobactam, and boronic acid (1–7).
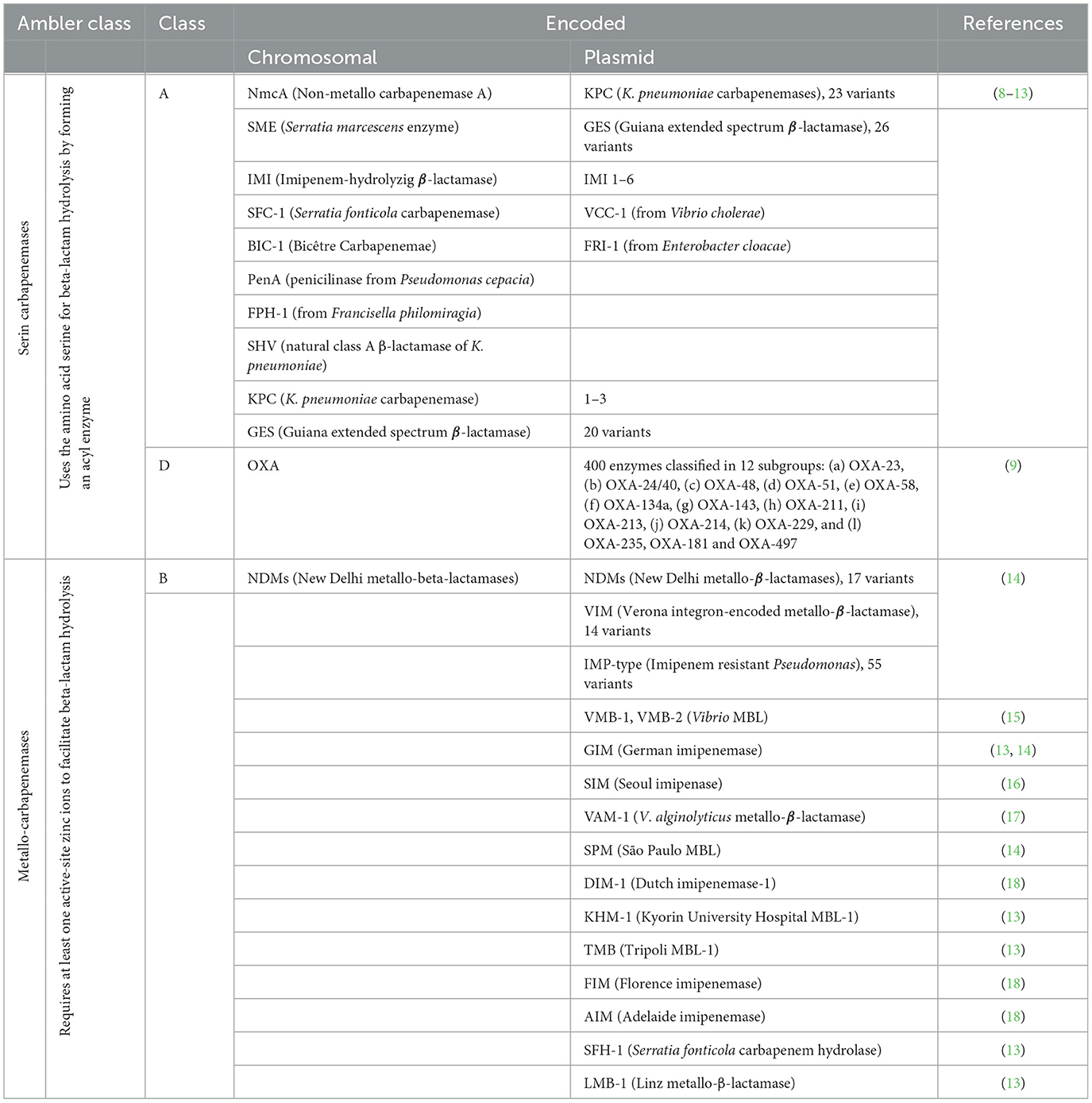
Table 1. Carbapenem-enzymes among carbapenem-resistance organisms on Ambler classification.
Class B or metallo-β-lactamases (MBLs) use a zinc ion (Zn2+) to hydrolyze the β-lactam ring. They confer resistance to all β-lactam antibiotics but are susceptible to aztreonam and β-lactam inhibitors such as ethylenediaminetetraacetic acid (EDTA). Most clinically important MBLs belong to the six different families [imipenem (IMP), Verona integron-encoded metallo-β-lactamase (VIM), New Delhi metallo-β-lactamase (NDM), São Paulo metallo-β-lactamases (SPM), German imipenemase (GIM), and Seoul imipenemase (SIM)]. This type of enzyme has been identified in clinically relevant species such as Enterobacterales, Acinetobacter, and P. aeruginosa. It is commonly expressed from mobile elements such as integrons, plasmids, and transposons (1–7).
Class D serine oxacillinases (OXAs) have been commonly detected worldwide. They have hydrolytic activity against β-lactams, high activity against penicillin, and weak activity against extended-spectrum cephalosporins and carbapenems. OXA-23 and OXA-48 are variants widely dispersed globally. OXA-23 is almost strictly restricted to Acinetobacter spp., wily OXA-48 was found among K. pneumoniae and Enterobacterales (20).
CROs are usually only susceptible to polymyxins (e.g., colistin), fosfomycin, and tigecycline, while colistin resistance in carbapenem-resistance K. pneumoniae (CR-Kp) isolates has been recently reported (21–24). Thus, colistin combination therapy is more frequently used to treat drug-resistant bacteria with significantly lower treatment failure rates (24).
Carbapenems such as imipenem, meropenem, ertapenem, doripenem, biapenem, faropenem, and panipenem have been approved for use in human clinical settings (6). However, carbapenems are not licensed in livestock or veterinary fields; therefore, carbapenem resistance is not commonly tested in animals. However, extended-spectrum beta-lactamases (ESBL), including ceftiofur, cefquinome, cefpodoxime, cefoperazone, and cefovecin, are commonly found in this sector. Additionally, ceftiofur, a third-generation cephalosporin, is the main cephalosporin used in veterinary fields and has been approved for treating bacterial infections in food-producing animals (i.e., pneumonia, arthritis, septicemia, meningitis, metritis, and polyserositis) (25). For this reason, these antibiotics could provide selection pressure that favors the expression of carbapenem-resistant (CR) strains.
Indeed, the author's statement in the Scientific Opinion on carbapenem resistance in food-animal ecosystems that “diagnostic isolates of veterinary origin classified as microbiologically resistant to third- and fourth-generation cephalosporins based on epidemiological cutoff values, should be subjected to phenotypic testing for carbapenem resistance and carbapenemase production and subsequent molecular identification and characterization of carbapenemase production genes because they favor the emergence of carbapenem-resistant isolates” (26).
Many studies have reported CROs in livestock, seafood, companion animals, wildlife, and their environments (7, 27–30). As animals have been identified as a relevant source of multidrug-resistant (MDR) bacteria, they can serve as reservoirs for carbapenem-resistant bacteria, and foodborne routes and emission in the environment through the excreta and the subsequent exposure of humans via the environment serve as transmission pathways for carbapenem-resistance genes from animals to humans and vice versa. Furthermore, their incidence may be underestimated because there is usually no surveillance, which dismisses the potential risks to human health. This review aimed to summarize the occurrence and molecular mechanisms of resistance in carbapenem-resistant organisms in food-producing animals, seafood, companion animals, and wildlife to address the importance of antimicrobial resistance surveillance in these sectors, resistance dissemination to the environment and humans, and potential public health risks.
Carbapenem-resistance genes have been identified in isolates from aquatic environments. Generally, resistance to carbapenems in bacteria from aquatic systems such as Vibrio and Shewanella spp., as well as Enterobacterales, is mainly mediated by the production of carbapenemases encoded by chromosomal genes or by plasmids. However, carbapenemases are variable, with Class B enzymes and the enzymes described only on aquatic species such as VMB-1 from Vibrio alginolyticus (17, 31) and VMB-2 (32) from Vibrio diabolicus on shrimps; as well as the Class A, VCC-1 from non-toxigenic Vibrio cholerae on shrimp (8).
In 2017, in Canada, Brouwer et al. isolated the E. cloacae complex from shrimp (Litopenaeus vannamei) originating in India. The isolated had a ST previously described in companion animals in Japan, ST813, and was positive for the blaIMI − 2 gene, which is located in a plasmid p3443-IMI2, which is closely related to IncFII plasmids and pIMI-6, which was described in an E. cloacae complex clinical isolate from Canada and carries the carbapenemase blaIMI − 6 (33). The same strain displayed the plasmid p3442-FLC-1 that carries the gene encoding a novel class A carbapenemase FLC-1 with close sequence similarity to blaFRI − 1, previously described in imipenem-resistant E. cloacae recovered from a clinical patient in France (33). In 2013, OXA-23-producing A. baumannii, on fish Pagellus acarne harvested in the Mediterranean Sea in Algeria, was reported. The isolate belonged to the widespread sequence type 2 (ST2)/international clone II and harbored aminoglycoside-modifying enzymes [aac(6′)-Ib and aac(3′)-I genes] as well as the naturally occurring blaOXA − 51 − like gene. However, the isolates differed from human clinical strains previously isolated from France and Algeria (34). In 2010, in Brazil, a high percentage of resistance to imipenem (71.43%) in E. coli isolated from aquaculture was detected, including isolates from pond water, shrimp tissues, and pond sediment (35). In 2014 and 2015, the occurrence of VIM-2-producing Pseudomonas fluorescens isolated from squid in Canada (imported from South Korea) and OXA-48-producing bacteria in seafood from China and Korea were described on the bacterial species Stenotrophomonas maltophilia, Myroides odoratimimus, Stenotrophomonas spp., and Pseudomonas putida (36, 37). In 2015, carbapenem-resistant Enterobacter spp., derived from imported retail seafood in Canada were detected, including two Enterobacter cloacae isolated from shrimp imported from Vietnam harboring blaIMI − 1; one Enterobacter aerogenes harboring blaIMI − 2 isolated from shrimp imported from Bangladesh, three E. cloacae harboring blaIMI − 1 isolated from clam imported from Vietnam, and two E. cloacae harboring blaNDM − 1, blaTEM, and blaOXA − 1 from clam samples from Vietnam (38). blaIMI − 2 gene was plasmid-mediated; the plasmid contained the IncFII (Yp) replicon, while blaNDM − 1 plasmid contained IncHI2, IncFIb, and IncFII replicons. Six different sequence types of E. cloacae were assigned (ST479, ST373, ST477, ST478, ST411, and ST412). The authors showed that the human-source E. cloacae ST373 isolate harboring blaIMI − 1 shared >75% similarity with the E. cloacae IMI-1 positive isolated from clam. In 2016, VCC-1-producing Vibrio cholerae isolated from retail shrimp imported from Canada was also identified (8). In 2016, in Italy, one VIM-1 carbapenemase-producing E. coli (ST10) was isolated from a Venus's clam (Ruditapes philippinarum) harvested in the Mediterranean Sea with blaVIM − 1 as part of the variable region of a class I integron embedded in a Tn3-like transposon that also contained the fluoroquinolone resistance gene qnrS1. Interestingly, E. coli ST10 is widespread among clinical and animal samples (39). In 2018, six blaNDM-harboring Enterobacteriaceae (four K. pneumoniae strains and two E. coli strains) from the retail fish market were detected in India, including the variants blaNDM − 5, blaNDM − 2, and blaNDM − 1. The blaNDM-positive E. coli isolates belonged to the multidrug-resistant widespread ST131 clone, representing extra-intestinal pathogenic E. coli. ST131 clone is widely distributed among human clinical isolates from urinary tract infections (UTIs). Moreover, they found that all the isolates were resistant to all β-lactam antibiotics, quinolones, trimethoprim-sulfamethoxazole, chloramphenicol, and tetracycline (40). In China, several isolates of Vibrio spp. were found to be resistant to imipenem and meropenem. Isolates from shrimps of seafood carried the genes blaNDM − 1, blaVIM − 1 (31), and blaVMB − 2 identified on a plasmid-borne composite transposon ISShfr9-ISCR1-blaVMB − 2-blaCARB − 12-aadA1-ISShfr9, where ISShfr9 was found to be disseminated in multidrug-resistant (MDR) pathogens (32); as well as producing the enzymes VMB-1 encoded by a gene blaVMB − 1 located in an integron bearing, highly transmissible IncC type plasmid, namely pVB1796 (15), and VAM-1 located in a conjugative plasmid, namely, pC1579 (17). Other studies in Korea have reported carbapenem-resistant Vibrio spp., isolates from shrimp (41), cockles (42), or hard-shell mussels harboring blaOXA genes (43). In Europe, in 2018 and 2017, Italy reported V. cholerae resistant to meropenem (44) and V. vulnificus isolated from shellfish resistant to imipenem and meropenem (45). KPC-3-producing E. coli in mussels (Mytilus galloprovincialis) and OXA-23-producing A. baumannii ST2 isolates from mussels and oysters (Crassostrea gigas) were also reported in Tunisia in 2016 (46, 47). The blaKPC − 3 gene was identified on an ~180 kb IncFII plasmid carrying Tn4401d transposon and belonged to the ST167 phylogroup A of the ST10 complex. Interestingly, the authors mention that the predominance of blaKPC − 3 in Portugal was also associated with the spread of an IncF plasmid carrying Tn4401d. The ST10 complex was reported previously in a hospital from the US to spread blaKPC genes. In 2018, France reported the isolation of NDM-1-producing V. parahaemolyticus ST864 from a shelled shrimp tail imported from Vietnam, which harbored the epidemic plasmid IncA/C (48). In South America and Ecuador in 2015, Vibrio spp., resistant to imipenem, was isolated from shrimp from seawater (49). In 2020, E. coli, Enterobacter cloacae complex, and K. pneumoniae were found in tilapia fish from Egyptian fish farms carrying blaKPC, blaOXA − 48, and blaNDM (50). In 2018, in Taiwan, carbapenem-resistant Shewanella algae were isolated from small abalone (Haliotis diversicolor) harboring genes encoding OXA-55 and multiple antibiotic-resistance genes including dfrA3 (trimethoprim resistance), tet (35) (tetracycline resistance), and qnrA3 (quinolone resistance); and the pmrCAB operon, which has been shown to mediate resistance to colistin (51). In Taiwan in 2019, carbapenem-resistant S. algae carrying blaOXA − 55 with multiple genes encoding efflux pumps was detected in Asian hard clam (Meretrix lusoria) (52). In 2020 in Italy, OXA-55-like producing S. algae was found (53).
All the previous information demonstrated the presence of CROs in seafood and aquaculture. Even when bacteria in aquatic environments are mainly non-pathogenic, their occurrence highlights the relevance of the food production chain in the global spread of antibiotic-resistance genes. Moreover, it is crucial to consider the seafood market, where countries can consume a specific product imported from a separate region by another country, which could have different regulations for antimicrobial resistance surveillance in food. Canada is one example of this since shrimp consumed by Canadians are imported from Asian countries. However, carbapenemase genes have been detected in these products. Examples include the isolation of Enterobacter cloacae or Enterobacter aerogenes harboring blaIMI − 1, blaIMI − 2, or blaNDM − 1 genes in retail seafood (39, 54).
In addition, it is remarkably the high amount of antibiotics used and their multiple classes in aquaculture for prophylactic purposes or metaphylactic treatment. Quinolones, tetracyclines, amphenicols, and sulfonamides are the most commonly used classes (55); however, they also include aminoglycosides, antimycobacterial (rifampin), beta-lactams (aminopenicillins and cephalosporins), and polymyxins. This factor could promote selective pressure for the emergence of antimicrobial resistance and the selection of multidrug-resistant organisms among seafood animals. Indeed, global antimicrobial consumption is estimated to reach 13,600 tons by 2030 (55). Additionally, antibiotic residues are difficult to eliminate by water treatment plants and can be discharged into water flows, thus being a source of antimicrobial resistance genes.
Moreover, fish do not effectively metabolize antibiotics; thus, the active substance passes into the environment in the feces. Indeed, it has been suggested that ~70–80% of the antibiotics applied in aquaculture are dispersed into water systems (37), which might provide a selection and enrichment mechanism for resistant bacteria (22). Studies reporting the occurrence of carbapenem-resistant genes worldwide in seafood and aquaculture are summarized in Table 2.
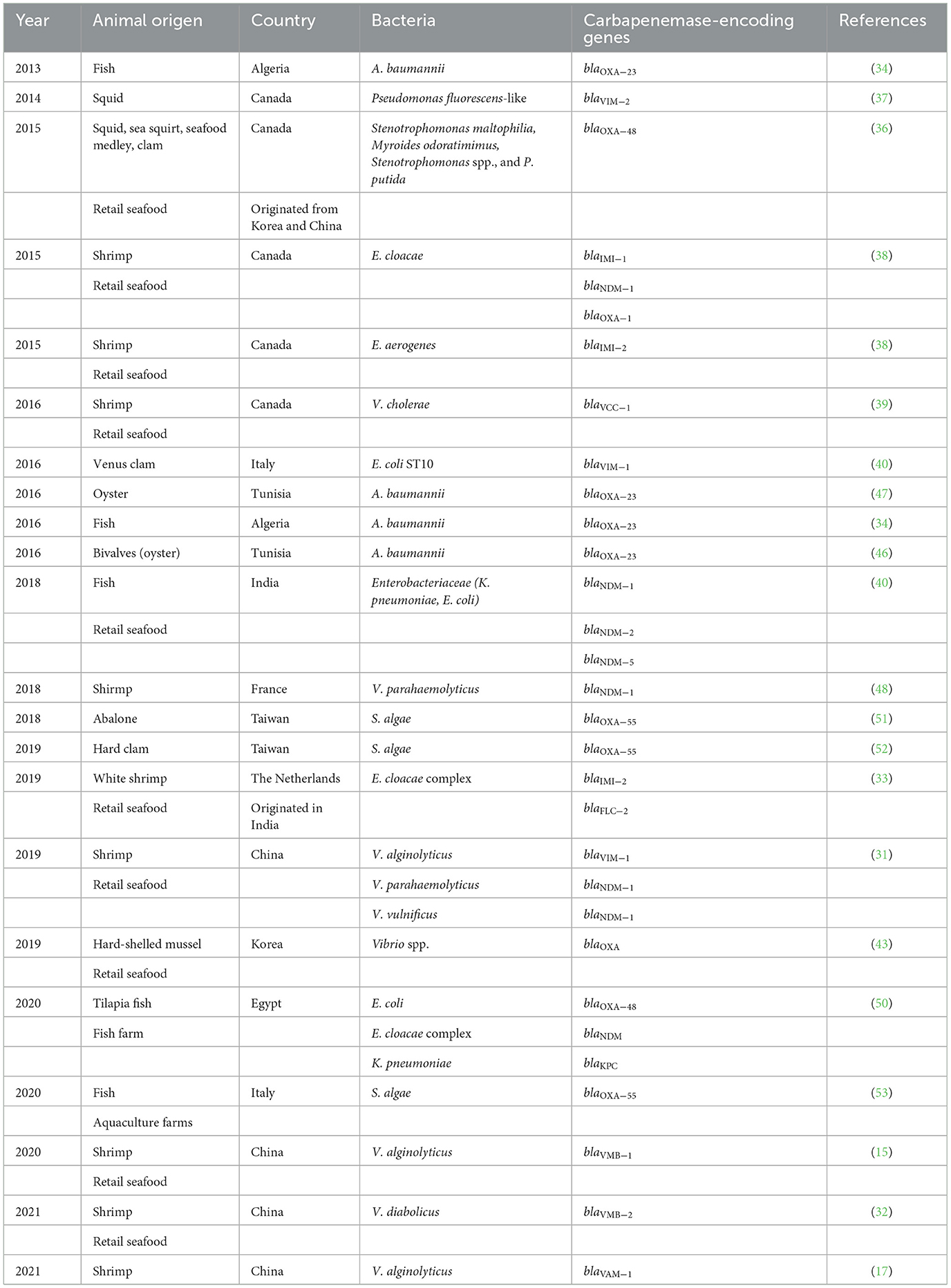
Table 2. Occurrence of carbapenemase-encoding genes in seafood and aquaculture.
Intensive farming has frequently been associated with the excessive use of antimicrobials and drug-resistant microorganisms isolated from food-producing animals that can be transmitted to humans via direct contact with animals or ingestion of derived food products (9). By 2030, global antimicrobial use from human, terrestrial, and aquatic food-producing animal sectors will reach 236,757 tons annually, with an estimated proportion of terrestrial food-producing animal use of 174,549 tons, representing 73.7% of the global consumption of antimicrobials (55). This intensive use of antibiotics creates selective pressure for the emergence of antimicrobial resistance among farmers and the environment. In addition, antibiotics continue to be used in livestock production as prophylaxis and animal growth support (9), which may lead to the emergence of antimicrobial resistance.
The transmission of antimicrobial resistance between food-producing animals and humans can occur via the food chain, by consuming food products contaminated with antimicrobial resistance genes or antimicrobial-resistant bacteria, through direct contact between humans and animals, or through shared environmental sources such as contaminated water (56, 57). In addition, resistance can be transmitted to livestock from environmental sources (i.e., hospital sewage, wastewater treatment plants contaminating water and soil, surface water flow, and wildlife) and biological vectors, such as flies and wild birds (21, 58). Moreover, livestock growth promotion antibiotics may interact with the animal gut microbiota and introduce increased variation in antimicrobial resistance genes (ARGs) in the gut (59), thus increasing their dispersion. Studies reporting the occurrence of carbapenem-resistant genes worldwide in terrestrial food-producing animals are summarized in Table 3.
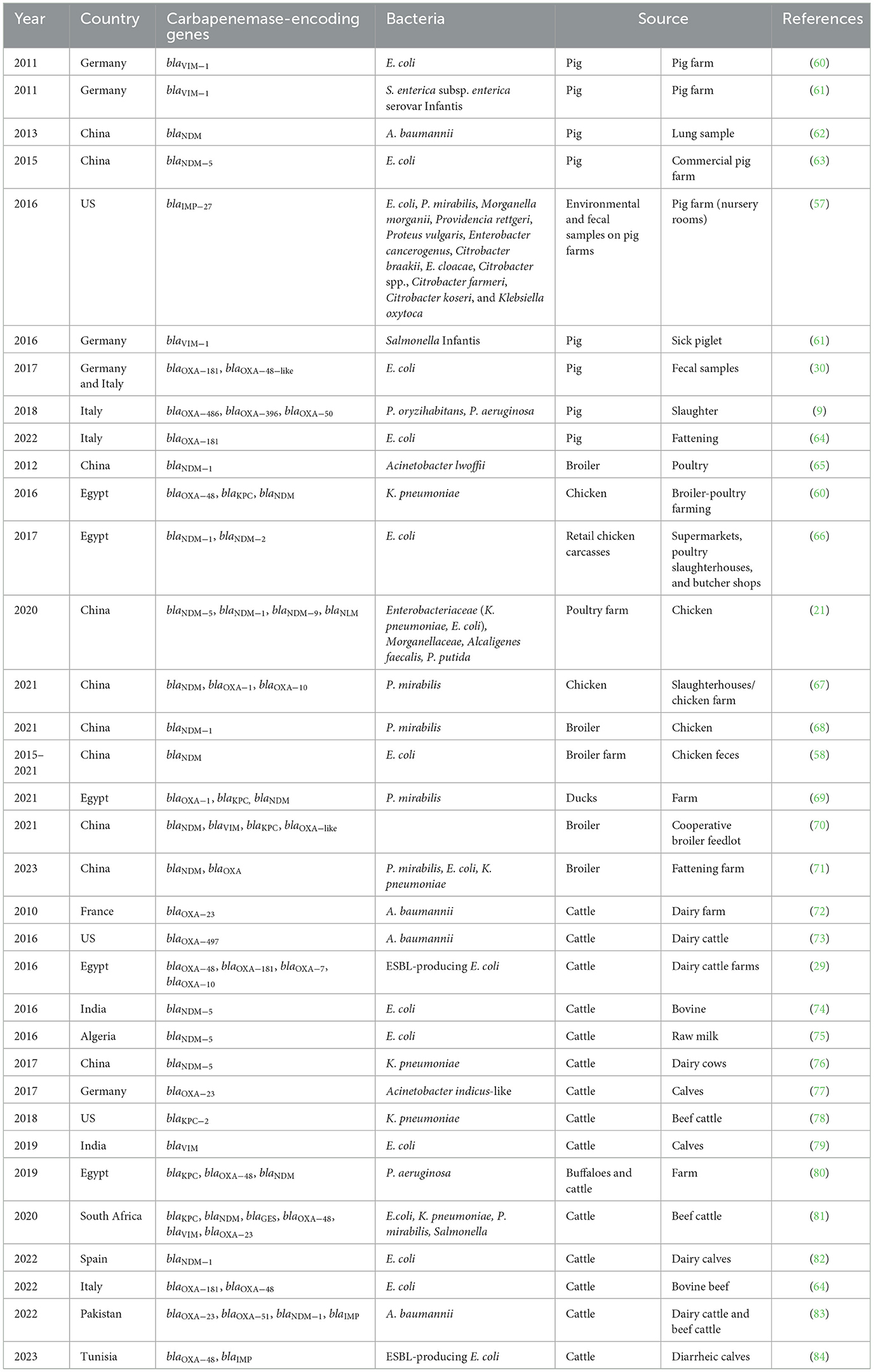
Table 3. Occurrence of carbapenemase-encoding genes in terrestrial food-producing animals.
In pigs, carbapenem resistance has been observed in microorganisms from different bacterial species, including E. coli, Salmonella, P. aeruginosa, and A. baumannii. In 2011, VIM-1-producing E. coli ST88 and Salmonella Infantis harboring blaVIM − 1IncHI2 plasmids were reported in Germany (60, 61). ST88 was also previously identified among chickens, cattle, and humans in Germany. Moreover, class 1 integron harbored by an IncH12 plasmid was found in human strains (60). In addition, during the sampling period in the same year (2011), 35 isolates were positive for blaVIM − 1, indicating that carbapenemase-producing bacteria may persist in livestock farms. Another study reported VIM-1-producing Salmonella Infantis in Germany in 2017 (59). The encoded genes blaVIM − 1 and blaNDM − 5 have reported resistance to third-generation cephalosporins used in animal husbandry. Moreover, Salmonella Infantis is one of the leading causes of human salmonellosis in Europe and a zoonotic pathogen commonly transferred via contaminated food products (59). In 2017, Pulss et al. (30) reported a porcine E. coli isolated carrying OXA-181 carbapenemase and the coexistence of mcr-1 (mobilized colistin resistance gene) and acquired carbapenemase gene blaOXA − 48 − like on isolates originated from Italy farms. blaOXA − 181 gene was located on al IncX3 plasmid (pEcIHIT31346-OXA-181), which presented high nucleotide similarity >99% to previously published plasmids from human sources (plasmid pOXA181_14828 of an E. coli isolated from a human patient in China) and also carried qnrS1 (plasmid-media quinolone resistance gene), thus providing evidence of the possible link between human- and animal-derived carbapenem resistance. In 2015, blaIMP − 27 was detected in Proteus mirabilis, Morganella morganii, Providencia rettgeri, Proteus vulgaris, Enterobacter cancerogenus, Citrobacter braakii, Enterobacter cloacae, Citrobacter spp., Citrobacter farmeri, Citrobacter koseri, and Klebsiella oxytoca in the United States, within an IncQ1 plasmid recovered from the nursery and farrowing barns of a swine production system (57). In 2013 in China, A. baumannii harboring blaNDM − 1 genes isolated from lung samples of pigs with pneumonia and sepsis were identified (62). Meropenem-resistant P. aeruginosa strains carrying blaOXA − 486, blaOXA − 396, blaOXA − 50, and blaPAO were also found in Italy in 2018, as were meropenem-resistant isolates of Pseudomonas oryzihabitans and P. aeruginosa (9). Three other isolates of P. aeruginosa carrying blaPAO, blaOXA − 50, blaOXA − 486, and blaOXA − 488 were detected in animals reared on different farms (85). Interestingly, two isolates of P. aeruginosa ST938 carrying blaPAO and blaOXA − 396 and the resistance genes to aminoglycosides [aph(3′)-IIb], fosfomycin (fosA4), and chloramphenicol (catB7) were detected, one in a pig and another one in 83-year-old patients. However, no epidemiological links were demonstrable between the animal and the patient. Other sequence types found were ST274, ST782, and ST885. The presence of blaOXA − 50 is concerning because this variant confers a decreased susceptibility to ampicillin, ticarcillin, and meropenem. In addition, the OXA-50 family also comprises blaOXA − 396, blaOXA − 486, and blaOXA − 488 genes (9). blaOXA48-like contained no plasmid, and blaOXA − 181-carrying IncX plasmid has also been reported in E. coli isolated from Italian fattening pigs (64). In the study, the authors recovered samples from fattening pigs, cattle, and workers from slaughterhouses. Twenty-four isolates were positive for blaOXA − 181 and one for blaOXA − 48. The isolates presented high ST diversity within ST5229 with higher prevalence. Different plasmid replicons were present in the isolates, with IncX1 and IncX3, and IncF types being the most represented. OXA-48-producing isolates did not contain any plasmid replicon. Furthermore, the authors detected an OXA-181-producing E. coli belonging to ST410 isolated in two fecal samples from fattening pigs, described as a high-risk clone associated with blaOXA − 181 in human patients. Moreover, in China in 2017, E. coli harboring a carbapenem-resistance gene blaNDM − 5 and mcr-1 were detected on IncX3 plasmid with a high degree of diversity of ST. ST156 was also previously reported in a Chinese hospital (63).
Among broiler farms, an increased prevalence of CRE has been shown, mainly in E. coli, K. pneumoniae, and P. mirabilis, harboring blaNDM genes, with blaNDM − 1 and blaNDM − 5 being the most predominant in chicken farm environments. Specifically, farming presented a higher prevalence of CRO among the studies, mainly attributed to the heavy use of antimicrobials on farms, transportation activities, and inadequate farm disinfection and management. Similarly, blaNDM genes are usually carried by the IncX3 plasmid, which is clinically significant because it contributes to disseminating various blaNDM variant genes (86).
In 2010, carbapenem-resistant isolates were detected in eight chicken farms, six duck farms, and one pig slaughterhouse in China. One of these isolates, Acinetobacter lwoffii, was identified as positive for blaNDM − 1, particularly on a 270 kb plasmid (65). In 2021, NDM-producing P. mirabilis was reported in broiler chickens (68). The isolate harbors a plasmid named pSNYG35, a pPrY2001-like plasmid that shares high nucleotide identity with pHFK418-NDM and an NDM-1-encoding plasmid from clinical P. mirabilis. Recently, Su et al. (87) reported isolation rates of 3.57% for carbapenem-resistant E. coli, 10% for carbapenem-resistant P. mirabilis, and 3.03% for carbapenem-resistant K. pneumoniae in six broiler fattening farms in China. Among carbapenem-resistant isolates, six E. coli carried class I integron, one carried class II integron, four P. mirabilis carried class I or II integrons, and one K. pneumoniae carried class 1 integron. All of these isolates harbor blaNDM and blaOXA genes. In 2016 in Egypt, carbapenem-producing K. pneumoniae (CR-Kp) in broiler poultry farming was reported. The authors found that 42% of the isolates from poultry samples carried blaNDM (11 isolates carried blaKPC, blaOXA − 48, and blaNDM; four isolates carried blaKPC, blaNDM or blaOXA − 48 and one isolate carried blaNDM alone) (70). Interestingly, the authors collected 49 fecal samples from workers and veterinarians working in the poultry farm; 56% of the samples were CR-Kp-positive, with all strains carrying the three carbapenemase genes blaKPC, blaOXA − 48, and blaNDM, and 5% of them displayed all the carbapenemase-encoding genes at the same time. Furthermore, the prevalence was higher in farm workers (67%) compared to veterinarians (33%), indicating that transmission could be facilitated by close contact between broilers and humans since the workers are in continuous contact with the animals and lived on the farm during the fattening program. However, the study did not compare clones or plasmids; non-genetic relationships between humans and chickens were found (70). Lately, in China in 2023, He et al. (58) observed the transmission of blaNDM-bearing plasmids of E. coli isolated from chickens between different farms and detected carbapenem-resistant isolates in farmlands, vegetable fields, and the environment of chicken farms. The authors performed a longitudinal study from 2015 to 2021 that demonstrated that the prevalence of blaNDM-positive clones and plasmids varied in different years, which suggested that new strains and plasmids are constantly being introduced into the farms. In 2020 in China, Zhai et al. (21) reported 279 NDM-producing bacteria, including Enterobacteriaceae (K. pneumoniae, E. coli), Morganellaceae, Alcaligenes faecalis, and Pseudomonas putida, with the variants NDM-5, NDM-1, and NDM-9 as well as a novel NDM-like-metallo-β-lactamase (NLM) within IncX3, IncA/C2, and IncFII as major blaNDM-carrying plasmid types among isolates. Moreover, they found the coexistence of mcr-1 or mcr-8 on K. pneumoniae positive for blaNDM − 1. The authors identified 14 sequence types among the E. coli isolates, with ST6751 being the most prevalent. ST6716, ST156, ST69, ST48, and ST10 were also found. STs 6751, 10, 125, and 746 were recovered from chicken and environmental samples (sewage trenches, corridor floors, drooping boards, nipple drinkers, and air). Most of the K. pneumoniae isolates were ST37, followed by ST3410 and ST726. Additionally, in China, Shi et al. (71) reported the presence of the resistance genes blaNDM, blaVIM, blaKPC, and blaOXA − like on broiler, layer, and pig farms with a significative higher relative abundance on blaOXA − like genes from 2016 to 2019. The authors detected a prevalence of 20–30% for blaKPC and blaVIM genes, respectively, and a prevalence of 75% for blaNDM, reflecting the great incidence of carbapenemase-producing genes in farming. Moreover, the study also found the coexistence of colistin resistance gene mcr-1 and blaNDM with pig and chicken farms displaying high prevalence. In 2020 in Egypt, 155 meropenem-resistant isolates were obtained from retail chicken meat, indicating that carbapenem-producing bacteria may enter the food chain. The study reported a single K. pneumoniae ST147 and a single E. coli ST648 producing NDM-1 and NDM-5. This last isolated carried also blaOXA − 1, blaTEM − 1, blaCTX − M−3, and aac(6′)-Ib-cr, while the K. pneumoniae harbored the blaSHV − 1, blaCTX − M−15, and aac(6′)-Ib-cr genes (71). NDM-producing ST648 E. coli has been reported in clinical isolates in India, the United Kingdom, and Australia. NDM-1-producing ST147 K. pneumoniae clone has been reported previously in Iraq, Oman, Tunisia, and Egypt from hospitalized patients (66). A study conducted in China in 2019 reported NDM-1-producing P. mirabilis recovered from commercial broilers in slaughterhouses (67). In 2021 in Egypt, P. mirabilis harbored blaNDM − 1, blaOXA − 1, and blaKPC was isolated from ducks on a duck farm (69).
Carbapenem-resistant bacteria are rare in cattle. However, since 2012, more studies have reported CROs in cattle with OXA- and NDM-producing bacteria leading to carbapenem resistance. In 2010, nine OXA-23-producing Acinetobacter genomospecies 15TU were reported in France, with a Tn2008 as a vehicle for the spread (72). In 2016 in the United States, a novel blaOXA − 497 gene was detected in A. baumannii, which is part of the OXA-51-like enzyme group and displays resistance to ertapenem; however, these enzymes are naturally occurring in A. baumannii (73, 88). In 2017, blaOXA − 23 harboring Acinetobacter indicus-like strains that displayed imipenem, meropenem, and doripenem resistance were isolated from nasal swabs of two calves in Germany. blaOXA − 23 was localized on the chromosome and surrounded by interrupted Tn2008 transposon structures. In addition, genetic relatedness between bovine isolates and Acinetobacter indicus type strains A648T and human clinical A. indicus isolates were found (77). In 2022, 27.7% of CRAB bacteria in Pakistan were reported to harbor blaOXA − 23 and blaOXA − 51 within 17 isolates carrying blaIMP and one isolate carrying blaNDM − 1. The typical sequence types found were ST642 and the international clone ST2 (83). In Egypt in 2019, carbapenem-resistant P. aeruginosa (CRPA) was reported in buffaloes and cattle with a prevalence of 60 and 59% (50 total samples) within isolates harboring blaKPC, blaOXA − 48, and blaNDM. The authors also found carbapenem-resistance genes from drinking water within 67% of prevalence and from stool human samples within 80% of prevalence. Additionally, phylogenetic analysis showed that cattle and water sequences were in one cluster and more related to each other than to human isolates (80). Similarly, in Egypt in 2014, five E. coli carrying blaOXA − 48, and one E. coli carrying blaOXA − 181 were reported in dairy cattle (29). In South Africa, 28–42% of carbapenem resistance was found in isolates such as E. coli, K. pneumoniae, P. mirabilis, and Salmonella spp., carrying blaKPC, blaNDM, blaOXA − 23, blaVIM, blaOXA − 48 and blaGES with different prevalence (81). In Italy, in 2021, the EU harmonized antimicrobial resistance (AMR) monitoring program reported that units of fattening pigs (21/301) and bovines (4/310) were positive for OXA-48-like E. coli (n = 24 OXA-181, n = OXA-48) (64). Most recently, in Tunisia, one isolate of ESBL-producing E. coli from calves with diarrhea carrying blaOXA − 48 and blaIMP were reported (84).
Antimicrobial resistance mediated through NDM enzymes is present in cattle. In 2013 in India, E. coli harboring the blaNDM − 5 gene was detected in milk samples of dairy cattle suffering mastitis (74). NDM-5-producing E. coli isolates from raw milk collected in a dairy farm in Algeria and India in 2016 were again found (75, 89). blaNDM − 1 gene in E. coli isolated from cattle, carried in an IncC plasmid, was reported in 2022 in Spain. The IncC plasmid also carried genes for aminoglycoside, sulphonamide, and trimethoprim resistance (82). K. pneumoniae carrying blaNDM − 5 located on IncX3 plasmid was isolated from dairy cows in China in 2017 (76). The presence of IncX3 plasmid is highly relevant since it mediates the spread of genes encoding resistance to clinically relevant antibiotics. It has been reported to encode qnrB7, qnrS, blaCTX − M−3, blaSHV − 12, blaKPC − 2, blaKPC − 3, blaNDM − 1, blaNDM − 4, blaNDM − 5, blaNDM − 7, blaNDM − 13, blaNDM − 17, and blaOXA − 181 (86). The authors found that the K.pneumoniaeblaNDM − 5 positive belonged to five STs, within ST1661 and ST2108, which were the most prevalent. The blaNDM − 5 gene was located on the ~46 kb IncX3 plasmid. The plasmid shared a similar genetic context and was nearly identical to the human K. pneumoniae plasmid (pNDM-MGR194) previously reported in India. Among beef cattle in the United States in 2018, isolates of K. pneumoniae carrying blaKPC − 2 from feces were detected in 72 samples (78). blaVIM gene located on an Incl1 plasmid of a novel sequence type (ST 297) from E. coli was well-detected among calves from India in 2019 (79).
Antimicrobial resistance genes can colonize wild animals following contact with sewage, human waste, or animal manure (90). Human feces and manure runoff are the primary sources of AMR in wild animals, as intake water polluted with feces could directly or indirectly contaminate other animals and the surrounding environment. Moreover, AMR genes, such as carbapenemase genes, could originate from environmental bacteria, such as the OXA-48 family of enzymes, which occurs naturally in Shewanella spp., a genus that inhabits lake sediments (91, 92), and OXA-23 enzymes, which are almost entirely restricted to A. baumannii and originate from the environmental species Acinetobacter radioresistens (93). Chickens have been proposed as a source of carbapenemase-producing Salmonella enterica in livestock (85).
Several carbapenemase-producing bacteria have been reported worldwide in wildlife including the NDM, IMP, VIM, and OXA enzymes. In Germany, Salmonella corvallis carried blaNDM − 1 belonging to ST1541 isolated from black kites (Milvus migrans) were detected in 2013 (94). blaNDM − 1 gene was located in the IncA/C conjugative plasmid pRH-1738 and contained a fosfomycin-resistance gene (fosA3 gen) (95). In 2016 in Australia, a high prevalence of Salmonella and IMP-4-producing Enterobacteriaceae was reported in silver gulls (96). The authors detected 120 carbapenem-resistant Enterobacteriaceae strains of 10 species, mainly E. coli carrying the blaIMP − 4, blaIMP − 38, and blaIMP − 26 genes, with a prevalence of 40% in the gulls. blaIMP gene was carried by conjugative plasmids of variable sizes and diverse replicons, including HI2-N, HI2, A/C, A/C-Y, L/M, I1, and non-typeable plasmids. The authors showed that isolates from gulls have significant similarities with clinical isolates from Australia, suggesting the human origin of the isolates. In 2017, France reported 22 carbapenem-resistant VIM-1-producing E. coli in yellow-legged gull (Larus michahellis) isolated in 2012 (97). Interestingly, gulls live in close contact with humans; thus, wildlife may be an important transmission route of AMR. In 2018, carbapenem-producing Enterobacteriaceae isolates (two E. coli ST635 and one K. pneumoniae ST13) were reported in fecal samples from wild boars in Algeria, Africa. OXA-48-producing isolates were also resistant to amoxicillin, amoxicillin-clavulanate, tobramycin, ertapenem, and meropenem (98). In 2019, China reported a high frequency of carbapenemase producer isolates (350 isolates) in migratory birds (Anser indicus, Phalacrocorax, and Larus ichthyaetus), while 233 Klebsiella spp. and 2 E. coli isolates were NDM-5-carriers (99). In 2019 in Korea, zoonotic Aeromonas spp., resistant to imipenem and meropenem, were isolated from the nutria (Myocastor coypus). These isolates also carried the cphA gene (Aeromonas hydrophila gene) coding for a carbapenem-hydrolyzing metallo-β-lactamase (100). In the same year, in Algeria, carbapenemase-producing K. pneumoniae was reported in bat guano with OXA-48 and KPC-3 enzymes present in the isolates, as well as the resistance genes blaTEM − 1 (ampicillin resistance) and aac(6′)-lb (aminoglycoside resistance) (101). In 2019, 13 carbapenem-resistant K. pneumoniae were isolated from Barbary deer (Cervus elaphus barbarus) in Akfadou Forest in Algeria. The resistome of these isolates revealed the presence of blaNDM − 1, blaCTX − M−15, blaSHV − 182, blaDHA − 1, blaOXA − 1, aac(3)-IIa, aac(3)-IId (aminoglycoside resistance), aac(6′)-Ib-cr (aminoglycoside-fluoroquinolone resistance), rmtC (rRNA methyltransferase with high-level resistance to aminoglycosides), sul1 (sulfonamides resistance), qnrB9 (plasmid-mediated quinolone resistance), fosA (fosfomycin resistance), tetA (tetracycline resistance), dfrA14 (trimethoprim resistance), catA2, catB3 (chloramphenicol resistance), and mphA (macrolide-resistant phosphotransferase) genes. Five different plasmids, IncA/C2, IncFIA/(HI1), IncFIB(K), IncFII(K), and ColRNAI, were also found (102). Similarly, in 2020, carbapenem-resistant K. pneumoniae carried blaOXA − 48 on an incompatible group L/M plasmid found in seals (Phoca vitulina) (103), reflecting anthropogenic pollution as a source of AMR genes. In 2021, a genomic comparison was performed between E. coli carrying KPC-2 and K. pneumoniae containing KPC-3 isolated from gulls and humans in Alaska. The authors found varying levels of genetic similarity at discrete genetic loci with no evidence of direct transmission of blaKPC between people and gulls; however, the conserved genetic elements surrounding blaKPC suggest a possible exchange between species (104). In 2021 in India, five carbapenem-resistant E. coli were reported isolated from rescued sloth bear (Melursus ursinus). The isolates were positive for blaNDM (60%, 3/5) carbapenemase gene and efflux pump-mediated carbapenem resistance (40%, 2/5), and co-harbor AMR genes blaTEM − 1, blaAmpC, qnrA, qnrB, qnrS, tetA, tetB, and sulI (105). In 2022, carbapenem-resistant P. aeruginosa strains were recovered from the feces of a red deer (Cervus elaphus) from Portugal, which resulted in a high-risk clone belonging to ST274 and co-harboring the genes blaPAO, blaPDC − 24, blaOXA − 486, aph(3′)-lb (aminoglycoside resistance), fosA (fosfomycin resistance), and catB7 (chloramphenicol resistance), which are phenotypically resistant to imipenem and intermediate resistance to meropenem and doripenem (106). In 2022, a high diversity of carbapenem-resistance genes was found in wild birds sampled from Alaska, Chile, Spain, Ukraine, Turkey, and Pakistan. The authors found carbapenemase genes in diverse isolates, including K. pneumoniae carrying KPC, NDM, OXA, and VIM, as well as in hypervirulent CR-Kp isolates from gulls in Spain and Ukraine. Some isolates harbored antimicrobial resistance to up to 10 antibiotic classes, including colistin. OXA-48-producing E. coli in gulls in Alaska and Turkey and CRE from Chile and Spain also harbored colistin-resistance genes. Similarly, the authors found evidence of global temporal and spatial dissemination (107). In 2022 in Brazil, NDM-1-producing E. coli ST162 infecting a pygmy sperm whale (Kogia breviceps) was reported (108). Moreover, the resistome of the isolate carried genes conferring resistance to β-lactams (blaNDM − 1, blaTEM − 1, and blaOXA − 1), aminoglycosides [aph(3′)-lb, aph(3′)-VI], macrolide (ermB, mdfA, and mphA), rifamycin (arr-3), [aac(6′)-Ib-cr, and qnrB6], phenicols (catB3 and floR), sulfonamide (sul1 and sul2), and tetracycline (tetA), and plasmid replicons IncFIB and IncA/C2 were also detected. All the previous studies shown here demonstrate that wild animals are reservoirs of carbapenem-resistant bacteria. They provide a biological mechanism for spreading antibiotic-resistance genes and can facilitate their transmission to humans and livestock.
On the other hand, rivers and water flow are also an environment from the emergence of CROs. For example, Pseudomonas fluorescens was recovered from the Seine River (Paris, France) in 2010, which expressed PF-1, a novel Ambler class A carbapenemase (109). In 2022 in Poland, 301 carbapenem-resistant Acinetobacter strains were isolated from municipal wastewater and river water (110). In 2005, carbapenem-resistant bacteria were reported on water bodies in the United States (111, 112). In 2019, carbapenem-resistant bacteria on water bodies were isolated, including Enterobacter asburiae, Aeromonas veronii, Cupriavidus gilardii, Pseudomonas, and Stenotrophomonas spp. This study found that most strains were carbapenemase producers, and all the isolates of Enterobacter asburiae carried the blaIMP − 2 gene (111). Other studies have also reported the presence of carbapenem-resistant strains in seawater, stormwater, and surface runoff water at Costa locations in Sydney, Australia, in 2020 (113). Therefore, water environments are an important reservoir of bacteria resistant to carbapenems and other antibiotics, including bacteria carrying intrinsic and acquired carbapenemase genes.
Carpabenemase-producing Enterobacterales (CPE) and non-fermenting bacteria have also been reported in companion animals. As we pointed out before, carbapenems are not approved for veterinary use. The prescription is restricted to treating urinary tract infections and respiratory tract infections in dogs and cats originating from multidrug-resistant (MDR) E. coli, K. pneumoniae, and P. aeruginosa bacteria. Additionally, the treatment must be supported by a veterinarian specializing in infectious disease, and by a pharmacologist (114), even though the continuous evidence of carbapenem-producing bacteria in companion animals has been increasing.
Companion animals can acquire carbapenemase-producing bacteria through direct contact with colonized hosts and the through contaminated environments such as veterinary hospitals (115, 116). In this regard, the human-pet bond has favored the silent transmission of carbapenem-producing bacteria to companion animals by a reverse zoonotic route called zooanthroponosis (115–117). Indeed, in Finland in 2015, identically isolates from dogs (with a long history of recurrent otitis externa without carbapenem prescription) and human family members with NDM-5-producing multidrug-resistant ST167 E. coli were reported. In addition, the same family carried an identical extended-spectrum beta-lactamase (ESBL) CTX-M-group 9 E. coli ST69, indicating interspecies transmission (118). In 2018 in Brazil, six VIM-2 carbapenemase-producing P. aeruginosa ST233 isolates were recovered from an infected dog, its owner (with a history of hospitalization), and its domestic environment (sofa, balcony, and water cooler) (116). ST233 has been reported as an international high-risk clone associated with carbapenemase production with resistance to all antimicrobial drugs. It has generally been restricted to human hospital settings (119–121), suggesting a zooanthroponotic transmission of this clone after the patient's hospital discharge. More recently, in 2022 in Guangzhou, China, a large-scale investigation on the prevalence of blaNDM-positive E. coli isolates from companion animals and their healthcare providers in clinical veterinary settings revealed the clonal spread blaNDM-positive ST453 E. coli isolates between both species (122). In France in 2022, OXA-48-producing K. pneumoniae were isolated from companion animals (dogs, cats, horses, cattle, and birds) with 56.2% (59/105 isolates) of the isolates belonging to the human-associated MDR ST11, ST15, and ST307 lineages, suggesting that numerous human-associated clones could infect the animal host (123).
Among carbapenemases on companion animals, NDM-5 and OXA-48-like carbapenemases are the most frequently described enzymes, with E. coli and K. pneumoniae being the main carbapenem-producing Enterobacterales, along with the non-fermenting bacteria A. baumannii (124). OXA-48 has been identified in Enterobacteriaceae from dogs and cats in different countries, such as Germany (2013) and the United States (2009–2013) (27, 125), as well as in an ST38 E. coli isolated from fowl (Gallus domesticus) in 2015 in Lebanon (28). In 2012, in Belgium, two OXA-23-producing Acinetobacter spp. were detected in fecal samples from 20 hospitalized horses, both resistant to imipenem and presented resistance to tetracyclines, sulfonamides, trimethoprim, and gentamicin but were still susceptible to colistin (126).
On the other hand, the KPC enzyme has also been reported. In 2018 in Brazil, in K. pneumoniae and E. coli from dogs, the blaKPC − 2 gene was found in Tn4401 transposons contained in IncN plasmids, which also carried blaCTX − M−15, and other clinically significant resistance determinants conferring resistance to aminoglycosides (aadA5), quinolines (qnrS1), macrolides [mph(A) and erm(B)], sulfonamides (sul1), tetracycline [tet(B)], and trimethoprim (dfrA17), and point of mutation conferring quinolone resistance (127). In Brazil 2021, the KPC-2-producing K. pneumoniae belonging to the high-risk international clone ST11/CG258 in a dog with urinary tract infection carrying the IncN plasmid assigned to ST15 was reported (128). The blaKPC − 4 gene was detected in 2016 in Ohio, US, in an IncHI2 plasmid in the context of the Tn4401b transposon in Enterobacter xiangfangensis isolated from a clinical dog sample with ST171, which has been responsible for major clusters of human CRE infections in the northeastern and upper-midwestern of the United States (129, 130). IMP-4 has been reported in Salmonella enterica serovar Typhimurium isolated from cats in Australia in 2016 (131). NDM-1 was isolated in 2013 in the United States from dogs and cats from E. coli that also carried blaCTX − M−15 and belonged to ST167 (132), as well as in China, with Acinetobacter species carrying blaNDM − 1 and blaOXA − 23 (133), and in Italy from A. radioresistens (134). Recently, in 2022 in China, five blaNDM − 5 harboring E. coli were reported in dogs and cats, all of them multidrogo resistant. The blaNDM − 5 gene was located on 46 kb IncX3 plasmids in the five strains. Additionally, one strais coharbored blaNDM − 5-encoding-IncX3 plasmid along with an mcr-1-IncX4 hybrid plasmid (135). OXA-48 has mainly been described in dogs, cats, and horses and mostly from infections such as urinary tract infections (UTIs) isolates from E. coli, K. pneumoniae, E. cloacae, and K. oxytoca (136, 137). VIM-1 and VIM-2 were also reported in dogs infected with K. pneumoniae and P. aeruginosa in Spain and Korea in 2016 and 2018, respectively (138, 139), as well as OXA-181-producing extra-intestinal pathogenic E. coli ST410 from a dog in Portugal in 2020 (140), OXA-23-mediated carbapenem-resistance A. baumannii ST2 from a cat (141), and OXA-66-producing A. baumannii isolated from cats (124, 142).
Among transmission between animals and humans, few studies have investigated the evidence for established links between human- and animal-derived carbapenem resistance. In 2022, Shen et al. (143) reported 29,799 E. coli isolates recovered from patients at 30 hospitals in China, as well as 61 pig farms and 45 chicken farms in 2017. From human clinical isolates, 631 were defined as carbapenem-resistant E. coli (CREc, 2.1%) with 195 NDM-positive. For livestock production, blaNDM was detected in 73.8% (n = 45) and 62.2% (n = 28) of pig and chicken farms, respectively. Furthermore, they found that human NDM-positive E. coli isolates shared 15 (n = 111), 11 (n = 90), and 10 (n = 96) STs with those from chickens, pigs, and flies, respectively. NDM-positive isolates belonging to ST167, ST206, ST10, and ST48 were recovered from all four origins. Furthermore, the authors found that large proportions of blaNDM genes (>70%) were associated with IncX3 plasmid in both animals (pig, chicken, and fly isolates) and humans. The authors also predicted the origins of 463 NDM-positive isolates. They found that 19% (n = 24), 8.1% (n = 10), and 1.6% (n = 2) of chicken NDM-positive E. coli isolates (n = 123) were predicted to originate from humans, pigs, and flies, respectively. In contrast, 27.3% (27/99) of pig NDM-positive E. coli isolates were predicted to originate from humans. Similarly, 53.8% (n = 105) and 14.9% (n = 29) of human isolates were predicted to have originated from chickens and pigs. All fly-derived isolates (n = 46) were predicted to have originated from humans (n = 5, 10.9%), chickens (n = 22, 47.8%), and pigs (n = 19, 41.3%). These results indicated positive associations and transmission of CREc between animals and humans. Indeed, the authors hypothesize that “CREc first arose in clinical settings and was then introduced into livestock animals, which are favorable hosts for the persistence of CREc. This led to the circulation of CREc between humans and animals, either via the food chain or through environmental vectors”.
In 2019, Li et al. (144) sampled 12 villages in China used as pig production farms [using the household as a single surveillance unit (resident and their backyard animals, including farm and companion animals)] and two commercial pig farms near the villages. The authors collected flies, fecal samples from humans, pigs, chickens, cattle, goats, ducks, one donkey, dogs, and cats across the villages, and additional fecal samples from pigs and farm workers at the two commercial farms. They obtained 88 CREC isolates that contained the blaNDM carbapenemase gene, 17 from humans, 44 from pigs, 12 from chickens, 12 from flies, two from dogs, and one from cattle. No CREC isolates were recovered from workers of pigs at the two nearest commercial pig farms. The authors detected blaNDM − 5, blaNDM − 1, and blaNDM − 9, with most of these blaNDM-genes likely located on IncX3-type plasmids. Indeed, the blaNDM-carrying regions/plasmid (IncX3) in CRE isolates from humans exhibited >99% nucleotide sequence identity to those in isolates from backyard animals and flies. MLST showed that six human CRE-NDM-positive isolates displayed ST48, ST10, ST1114, or ST6910 shared by animal isolates. ST48 was the most prevalent and was associated with isolates from pigs, humans, chickens, and flies. Furthermore, they found that two human isolates displayed only three single-nucleotide polymorphisms (SNPs) with two pig isolates from the same village. They also reported that CREC isolates from flies have human and dog origins, while chicken isolates had a predominant origin from pigs and dogs. In addition, the single cattle-derived isolate was clustered with the chicken isolates. Therefore, many CRE isolates from humans, backyard animals, and flies originated from hosts other than those included in the study.
In 2017, Wang et al. (145) recovered 245 CRE from poultry (chicken farms, slaughterhouses, and supermarkets), dogs, sewage, wild birds, flies, and farmers. The authors identified blaNDM in 21.8% (n = 161) of the E. coli isolates, 7.4% on K. pneumoniae, and 3.9% in E. cloacae, with blaNDM − 5, blaNDM − 9, blaNDM − 1, and blaNDM − 7 variants. Importantly, 23% of CREC isolates were also positive for mcr-1. High rates of CREC were found in dogs' feces (82.4%), flies (25.8%), wild birds' nests (40%), and anal swabs of farmers (50%). The most prevalent STs among blaNDM-positive isolates were ST101, ST156, and ST746. Moreover, MLST analysis showed commonality between strains from chicken farms, slaughterhouses, supermarkets, and humans, typified by genotypes ST10 and ST156. The authors confirm the commonality of ST156 isolates among disparate samples by core-genome single-nucleotide polymorphism (SNP)-based phylogenetic analysis. Additionally, blaNDM-carrying contigs gave three main genomic backbone profiles. The type II backbone included the contigs from 84 isolates derived from chicken cloacae (n = 37), flies (n = 21), dog feces (n = 12), chicken meat from supermarkets (n = 5), sewage from the farm (n = 1), chicken caeca from a slaughterhouse (n = 1), feces from farmers (n = 3), swallows (n = 3), and sewage from a slaughterhouse (n = 1). Type II genomic backbone was found in 26 E. coli isolates and shares >99.9% nucleotide sequence identity with the corresponding region of a 46,253bp IncX3 plasmid pJEG027 from K. pneumoniae isolated from an Australian traveler who was repatriated to Sydney from Myanmar.
The studies above showed a positive association between livestock production and human CREC infections since they identified a close relationship between the genomic profile of carbapenem-resistant isolates from humans and animals. High similarities between isolates from different sources were found. However, the studies only focused on NDM-positive isolates and not on other carbapenemases-encoding genes. Further studies are needed to elucidate the link between humans and animals.
Different strategies to combat antimicrobial resistance have been developed, including the One Health approach, the EU Harmonized AMR Monitoring Program conducted in Italy in 2021, the National Action Plan for Combating Antibiotic-Resistant Bacteria (CARB) by the US (2020–2025), and the implementation of antimicrobial risk assessment.
One Health approach is a term recognized in the EU in 2016 by the United Nations Political Declaration on Antimicrobial Resistance (AMR), which states that human health, animal health, and the environment are interconnected and that disease is transmitted from humans to animals and vice versa. Furthermore, the environment could be a potential source of new resistant microorganisms; therefore, AMR should be addressed in all scenarios (25, 146, 147).
Strictly, One Health is defined as “a collaborative, multisectoral, and trans-disciplinary approach—working at local, regional, national, and global levels—to achieve optimal health (and wellbeing) outcomes recognizing the interconnections between people, animals, plants and their shared environment” (146). Therefore, a multidisciplinary approach is required to prevent the spread and emergence of antimicrobial resistance. Antimicrobial resistance (AMR) surveillance using the One Health approach has been implemented in Europe, the UK, and the US to mitigate the crisis. However, the lack of implementation in most developing countries resulted in the underestimation of the burden of AMR on terrestrial and aquatic animals and the environment (147).
Improper management of antimicrobials, such as inadequate control of infection, use of antimicrobials as growth promoters (long-term, low-dose mass medication), prophylaxis in livestock, farmed fish in aquaculture systems, agricultural debris, environmental pollutants from sewage, pharmaceutical industry waste, manure runoff from farms, use of heavy metals, use of disinfectants, and migration of people and animals infected with resistant bacteria, facilitate the spread of resistance between humans and animals (25, 148). Consequently, the One Health approach is fundamental since it is a multidisciplinary approach that tries to prevent, predict, detect, and respond to AMR (25, 148).
Critical strategies for addressing AMR from the One Health perspective includes: (1) conduct a global campaign to raise awareness of antimicrobial resistance and the damage caused by the overuse and misuse of antibiotics, (2) improve hygiene measures and prevent the spread of infections (i.e., decrease missing of animals from different sources, stress of transport, unsanitary or crowded conditions), (3) reduce the use of antimicrobials in agriculture and their dissemination to the environment (including third-generation cephalosporins, fluoroquinolones, colistin, tetracyclines, and macrolides; i.e., growth promoters such as colistin has been banned in Europe, Canada, Denmark, United States, and other countries) (24), (4) improve global surveillance of drug resistance in order to understand and clarify the new mechanisms of resistance acquisition and predict future threats, (5) promote new and rapid clinical diagnoses, (6) promote the development and use of vaccines and alternatives to antibiotics (i.e., phage therapy, probiotics, antibodies, lysins, among others), (7) improve the number of studies in the field, (8) generated a global innovation fund for early-stage research on new treatments, (9) promote investment in new drugs and in the improvement of existing drugs, and finally, (10) build a global coalition for real action against AMR (25, 148, 149).
All previous studies have highlighted the urgent need to establish a One Health AMR surveillance system to understand the magnitude of the AMR problem, specifically the carbapenem-resistance problem, identify trends, and determine how all scenarios are linked and establish settings to content the widespread carbapenem-resistant organisms and genes. This approach requires the integration of human healthcare, livestock, aquaculture, and the environment, as well as other variety of disciplines and fields (149). Furthermore, the role of infections caused by antimicrobial-resistant organisms in wildlife may also have to be addressed, along with resistant organisms from aquatic environments, as they could possess intrinsic resistance and the possibility of being transmitted horizontally. In addition, carbapenem-resistant in companion animals has to take seriously since the human-pet bond might favor the silent transmission of clinically significant multidrug-resistant bacteria through zooanthroponosis (148).
Moreover, antimicrobial residues in fish products can persist in aquatic environments through excreta. For example, testing foodstuffs for carbapenem-resistant bacteria is not a legal requirement in any country; however, even a low prevalence of carbapenem-resistant genes has been detected in imported shrimp and salmon. In addition, studies have shown that aquaculture and terrestrial farms exhibit significant differences in drug consumption, with the aquaculture sector exhibiting the lowest. However, commensal bacterial flora can act as reservoirs of AMR genes, which may be transferred to microorganisms capable of causing human and animal diseases. Furthermore, it has been documented that animals excrete a significant percentage (75–90%) of antimicrobials without being metabolized and dispersed into the environment (145), which could be taken up by wild animals and function as a reservoir for antimicrobial resistance genes (149).
The reports on carbapenem-resistant organisms published from seafood and aquaculture are still low. Most of these reports must include information on potential sources or transmission between humans, animals, and their environment. Similarly, some of the microorganisms found are of clinical importance. Some examples are carbapenem-producing Vibrio alginolyticus, which causes vibriosis, wound infection, and ear infection; Vibrio cholerae, which causes cholera; Vibrio parahaemolyticus, that cause acute, self-limiting gastroenteritis (150); Shewanella algae, a potential foodborne zoonotic agent in humans that causes necrotizing fasciitis, discitis, meningitis, biliary infection, pneumonia, and endocarditis (151); and Enterobacter cloacae complex that is common in nosocomial settings and capable of producing several infections such as pneumonia, urinary tract infections, and septicemia (152).
On livestock, carbapenem resistance has been observed in microorganisms from different bacterial species, critical in human settings, and associated with significant public health concerns worldwide, including E. coli, Salmonella, P. mirabilis, P. aeruginosa, and A. baumannii. In swine and poultry settings, VIM-1-producing Salmonella Infantis was found. This bacterium is a zoonotic pathogen commonly transferred via contaminated food products that have been implicated in human salmonellosis and foodborne outbreaks associated with egg and chicken meat (153). For instance, A. baumannii is an important opportunistic pathogen for hospital-acquired infections commonly associated with multi-drug resistance. The mortality rate of A. baumannii infection has been estimated to be over 50% (154). In livestock such as cattle and pigs, A. baumannii causes mastitis, pneumonia, and sepsis. In companion, animals cause urinary tract infections (155–157). Whether the presence of OXA-23-producing A. baumannii poses a substantial public health threat is unclear, but the presence of NDM-1 producers in A. lwoffii and A. baumannii isolated from poultry, swine, and cattle, which are clinically relevant to humans, is worrying. Similarly, NDM-producing E. coli and K. pneumoniae, isolated from poultry and cattle, are worrisome since they have been reported in clinical isolates worldwide.
The carbapenem-resistance determinants in wild animals need to be better understood. Wild animals may act as potential environmental reservoirs for bacterial resistance. VIM, NDM, OXA-48, and KPC-producing K. pneumoniae are the more frequent carbapenemases reported, followed by IMP, and NDM-producing E. coli. Contaminated food and water are the main routes of transmission of carbapenem-resistance bacteria to wild animals (158). However, anthropogenic pressure plays an essential role in the emergence of resistance, particularly in this setting. Interestingly, migrating birds (i.e., gulls) have been proposed to serve as a vehicle for disseminating carbapenem-resistance genes (159).
Among food-producing animals, the link between farming practices, animal health, carbapenem-resistant organisms' development and spread to farmers, and the presence of carbapenem-resistant organisms in foodstuffs requires much more investigation. Three studies positively associated livestock production with human CREC infections (143–145). These associations are mainly based on the observation that blaNDM-carrying IncX3 plasmid isolated from humans exhibited between 75 and 99% nucleotide sequence identity to those in isolates from other sources, including chicken, pigs, and fly isolates. Moreover, one article identified a close relationship between the core-genome sequences of NDM-positive E. coli from humans and animals. The source-tracing analysis revealed indistinct boundaries between human- and animal-derived NDM-positive E. coli (143). Several studies have found genetic similarities between carbapenem-producing bacteria from animal and human sources, including the detection of a porcine E. coli isolated carrying OXA-181 carbapenemase located on al IncX3 plasmid with high nucleotide similarity (99%) to previously published plasmid from human sources (30); E. cloacae IMI-1 positive isolated from clam and human-source E. cloacae ST373 isolate harboring blaIMI − 1 sharing >75% similarity (8), and the detection of varying levels of genetic similarity at discrete genetic loci between E. coli carrying KPC-2 and K. pneumoniae containing KPC-3 isolated from gulls and humans in Alaska (104).
In companion animals, carbapenem resistance has been reported. The enzymes NDM-5, VIM, KPC, OXA-48-like, and OXA-23 were detected in E. coli, K. pneumoniae, P. aeruginosa, and A. baumannii (124–142). In this setting, the evidence suggests that zooanthroponosis is the main route of transmission of carbapenem-producing bacteria from humans to companion animals, indicating a cross-species transmission (115–123). Remarkably, in carbapenem-resistant organisms isolated from all the sources presented here (food-producing animals, seafood, aquaculture, wildlife, and companion animals), NDM-1 enzymes are occurring. These enzymes can hydrolyze all β-lactam antibiotics and have a high potential for rapid dissemination, thus, may constitute a public health risk (159).
These examples demonstrated that direct anthropozoonotic or zooanthroponotic transmission might be possible for CRE. However, to estimate the public health relevance of this transmission, more studies are needed to elucidate the problem. The addition of high-throughput technology, such as whole-genome sequencing and next-generation sequencing (NGS), has been permitted to determine the genetic relationship among CRO from different species at gene, plasmid, and strain levels. Similarly, introducing discriminant analysis of principal components (DAPCs) is helpful for tracing carbapenem-producing strains' potential origins.
The data presented in this review confirm the widespread of carbapenemase-producing bacteria and encoding genes in food-producing animals, seafood, aquaculture, companion animals, and wildlife as a cause of representing a severe problem for human and animal health.
Several studies have shown genetic similarities between human and animal carbapenem-resistance isolates, thus, demonstrating the possible cross-species transmission. Nonetheless, epidemiologic and genotypic analysis studies are needed to understand better the dynamics of antimicrobial drug resistance transmission between humans, animals, and the environment. In addition, the presence of CROs in the food chain compromises food safety and security and increases the chance of cross-border transmission of these bacteria.
One Health approach can help to implement global monitoring programs and establish antimicrobial risk assessments for the zoonotic and environmental sectors to address AMR emergencies. It is essential to identify and share best practices and policies globally. Collaboration between governments is needed to address cross-border health threats of AMR.
AG-B conceived and designed the idea for the manuscript. FR-C and AG-B wrote the manuscript. AG-B, FR-C, and FA-G reviewed the manuscript. All authors read and approved the final manuscript.
FR-C received a research grant from CONACYT (grant number: 373176). This study was supported by the internal research project PIB19-3 of the Autonomous University of Aguascalientes.
The authors are grateful to the Universidad Autonoma de Aguascalientes for supporting this study and the Master in Veterinary Science María Magdalena Soto Perezchica for their advice and point of view about the use of antibiotics in livestock. The authors are grateful to the financial support of the National Council of Science and Technology (CONACyT) for the scholarship awarded in the Call for Postdoctoral Stays for Mexico.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Nordmann P, Dortet L, Poirel L. Carbapenem resistance in Enterobacteriaceae: here is the storm! Trends Mol Med (2012) 18:263–72. doi: 10.1016/j.molmed.2012.03.003
2. Gupta N, Limbago BM, Patel JB, Kallen AJ. Carbapenem-resistant Enterobacteriaceae: epidemiology and prevention. Clin Infect Dis. (2011) 53:60–7. doi: 10.1093/cid/cir202
3. Aurilio C, Sansone P, Barbarisi M, Pota V, Giaccari LG, Coppolino F, et al. Mechanisms of action of carbapenem resistance. Antibiotics. (2022) 11:421. doi: 10.3390/antibiotics11030421
4. Codjoe FS, Donkor ES. Carbapenem resistance: a review. Med Sci. (2017) 6. doi: 10.3390/medsci6010001
5. Meletis G, Protonotariou E, Papadopoulou D, Skoura L. Comment on: the carbapenemase menace: do dual mechanisms code for more resistance? Infect Control Hosp Epidemiol. (2016) 37:1392–4. doi: 10.1017/ice.2016.197
6. G.Taggar, Attiq Rheman M, Boerlin P, Diarra MS. Molecular epidemiology of carbapenemases in Enterobacteriales from humans, animals, food and the environment. Antibiotics. (2020) 9:693. doi: 10.3390/antibiotics9100693
7. Anderson REV, Boerlin P. Carbapenemase-producing Enterobacteriaceae in animals and methodologies for their detection. Can J Vet Res. (2020) 84:3–17.
8. Mangat CS, Boyd D, Janecko N, Martz SL, Desruisseau A, Carpenter M, et al. Characterization of VCC-1, a novel Ambler Class A carbapenemase from Vibrio cholerae isolated from imported retail shrimp sold in Canada. Antimicrob Agents Chemother. (2016) 60:1819–25. doi: 10.1128/AAC.02812-15
9. Bonardi S, Cabassi CS, Manfreda G, Parisi A, Fiaccadori E, Sabatino A, et al. Survey on carbapenem-resistant bacteria in pigs at slaughter and comparison with human clinical isolates in Italy. Antibiotics. (2022) 11:777. doi: 10.3390/antibiotics11060777
10. Walther-Rasmussen J, Høiby N, Class A. carbapenemases. J Antimicrob Chemother. (2007) 60:470–82. doi: 10.1093/jac/dkm226
11. Queenan AM, Bush K. Carbapenemases: the versatile beta-lactamases. Clin Microbiol Rev. (2007) 20:440–58. doi: 10.1128/CMR.00001-07
12. Hammoudi Halat D, Ayoub Moubareck C. The current burden of carbapenemases: review of significant properties and dissemination among gram-negative bacteria. Antibiotics. (2020) 9:186. doi: 10.3390/antibiotics9040186
13. Bonnin RA, Jousset AB, Emeraud C, Oueslati S, Dortet L, Naas T. Genetic diversity, biochemical properties, and detection methods of minor carbapenemases in Enterobacterales. Front Med. (2021) 7:616490. doi: 10.3389/fmed.2020.616490
14. Castanheira M, Toleman MA, Jones RN, Schmidt FJ, Walsh TR. Molecular characterization of a beta-lactamase gene, blaGIM-1, encoding a new subclass of metallo-beta-lactamase. Antimicrob Agents Chemother. (2004) 48:4654–61. doi: 10.1128/AAC.48.12.4654-4661.2004
15. Zheng Z, Cheng Q, Chan EW, Chen S. Genetic and biochemical characterization of VMB-1, a novel metallo-β-Lactamase encoded by a conjugative, broad-host range IncC plasmid from Vibrio spp. Adv Biosyst. (2020) 4:e1900221. doi: 10.1002/adbi.201900221
16. Lee K, Yum JH, Yong D, Lee HM, Kim HD, Docquier JD, et al. Novel acquired metallo-beta-lactamase gene, blaSIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother. (2005) 49:4485–91. doi: 10.1128/AAC.49.11.4485-4491.2005
17. Cheng Q, Zheng Z, Ye L, Chen S. Identification of a novel metallo-β-lactamase, VAM-1, in a foodborne Vibrio alginolyticus isolate from China. Antimicrob Agents Chemother. (2021) 65:e0112921. doi: 10.1128/AAC.01129-21
18. Zhao WH, Hu Z-Q. Acquired metallo-β-lactamses and their genetic association with class 1 integrons and ISCR elements in Gram-negative bacteria. Fut Microbiol. (2015) 10. doi: 10.2217/fmb.15.18
19. Castanheira M, Sader HS, Farrell DJ, Mendes RE, Jones RN. Activity of ceftaroline-avibactam tested against Gram-negative organism populations, including strains expressing one or more β-lactamases and methicillin-resistant Staphylococcus aureus carrying various staphylococcal cassette chromosome mec types. Antimicrob Agents Chemother. (2012) 9:4779–85. doi: 10.1128/AAC.00817-12
20. Hansen GT. Continuous evolution: perspective on the epidemiology of carbapenemase resistance among Enterobacterales and other Gram-negative bacteria. Infect Dis Ther. (2021) 10:75–92. doi: 10.1007/s40121-020-00395-2
21. Zhai R, Fu B, Shi X, Sun C, Liu Z, Wang S, et al. Contaminated in-house environment contributes to the persistence and transmission of NDM-producing bacteria in a Chinese poultry farm. Environ Int. (2020) 139:105715. doi: 10.1016/j.envint.2020.105715
22. Nordmann P, Cuzon G, Naas T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect Dis. (2009) 9:228–36. doi: 10.1016/S1473-3099(09)70054-4
23. van Duin D, Doi Y. Outbreak of colistin-resistant, carbapenemase-producing Klebsiella pneumoniae: are we at the end of the road? J Clin Microbiol. (2015) 53:3116–7. doi: 10.1128/JCM.01399-15
24. Shein AMS, Wannigama DL, Higgins PG, Hurst C, Abe S, Hongsing P, et al. High prevalence of mgrB-mediated colistin resistance among carbapenem-resistant Klebsiella pneumoniae is associated with biofilm formation, and can be overcome by colistin-EDTA combination therapy. Sci Rep. (2022) 12:12939. doi: 10.1038/s41598-022-17083-5
25. McEwen SA, Collignon PJ. Antimicrobial resistance: a one health perspective. Microbiol Spectr. (2018) 6. doi: 10.1128/microbiolspec.ARBA-0009-2017
26. EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on Carbapenem resistance in food animal ecosystems. EFSA J. (2013) 11:3501. doi: 10.2903/j.efsa.2013.3501
27. Liu X, Thungrat K, Boothe DM. Occurrence of OXA-48 carbapenemase and other β-lactamase genes in ESBL-producing multidrug resistant Escherichia coli from dogs and cats in the United States, 2009-2013. Front Microbiol. (2016) 7:1057. doi: 10.3389/fmicb.2016.01057
28. Al Bayssari C, Olaitan AO, Dabboussi F, Hamze M, Rolain JM. Emergence of OXA-48-producing Escherichia coli clone ST38 in fowl. Antimicrob Agents Chemother. (2015) 59:745–6. doi: 10.1128/AAC.03552-14
29. Braun SD, Ahmed MF, El-Adawy H, Hotzel H, Engelmann I, Weiss D, et al. Surveillance of extended-spectrum beta-lactamase-producing Escherichia coli in dairy cattle farms in the Nile Delta, Egypt. Front Microbiol. (2016) 7:1020. doi: 10.3389/fmicb.2016.01020
30. Pulss S, Semmler T, Prenger-Berninghoff E, Bauerfeind R, Ewers C. First report of an Escherichia coli strain from swine carrying an OXA-181 carbapenemase and the colistin resistance determinant MCR-1. Int J Antimicrob Agents. (2017) 50:232–6. doi: 10.1016/j.ijantimicag.2017.03.014
31. Zheng Z, Ye L, Chan EW, Chen S. Identification and characterization of a conjugative blaVIM-1-bearing plasmid in Vibrio alginolyticus of food origin. J Antimicrob Chemother. (2019) 74:1842–7. doi: 10.1093/jac/dkz140
32. Liu M, Zhang W, Peng K, Wang Z, Li R. Identification of a novel plasmid-mediated carbapenemase-encoding gene, blaVMB-2, in Vibrio diabolicus. Antimicrob Agents Chemother. (2021) 65:e0020621. doi: 10.1128/AAC.00206-21
33. Brouwer MSM, Tehrani HME, Rapallini M, Geurts Y, Kant A, Harders F, et al. Novel carbapenemases FLC-1 and IMI-2 encoded by an Enterobacter cloacae complex esolated from food products. Antimicrob Agents Chemother. (2019) 63:e02338–18. doi: 10.1128/AAC.02338-18
34. Brahmi S, Touati A, Cadiere A, Djahmi N, Pantel A, Sotto A, et al. First description of two sequence Type 2 Acinetobacter baumannii isolates carrying OXA-23 Carbapenemase in Pagellus acarne fished from the Mediterranean Sea near Bejaia, Algeria. Antimicrob Agents Chemother. (2016) 60:2513–5. doi: 10.1128/AAC.02384-15
35. Vieira RH, Carvalho EM, Carvalho FC, Silva CM, Sousa OV, Rodrigues DP. Antimicrobial susceptibility of Escherichia coli isolated from shrimp (Litopenaeus vannamei) and pond environment in northeastern Brazil. J Environ Sci Health B. (2010) 45:198–203. doi: 10.1080/03601231003613526
36. Morrison BJ, Rubin JE. Carbapenemase producing bacteria in the food supply escaping detection. PLoS ONE. (2015) 10:e0126717. doi: 10.1371/journal.pone.0126717
37. Rubin JE, Ekanayake S, Fernando C. Carbapenemase-producing organism in food, 2014. Emerg Infect Dis. (2014) 20:1264–5. doi: 10.3201/eid2007.140534
38. Janecko N, Martz SL, Avery BP, Daignault D, Desruisseau A, Boyd D, et al. Carbapenem-resistant Enterobacter spp. in retail seafood imported from Southeast Asia to Canada. Emerg Infect Dis. (2016) 22:1675–7. doi: 10.3201/eid2209.160305
39. Roschanski N, Guenther S, Vu TTT, Fischer J, Semmler T, Huehn S, et al. VIM-1 carbapenemase-producing Escherichia coli isolated from retail seafood, Germany 2016. Euro Surveill. (2017) 22:17-00032. doi: 10.2807/1560-7917.ES.2017.22.43.17-00032
40. Das UN, Singh AS, Lekshmi M, Nayak BB, Kumar S. Characterization of bla(NDM)-harboring, multidrug-resistant Enterobacteriaceae isolated from seafood. Environ Sci Pollut Res Int. (2019) 26:2455–63. doi: 10.1007/s11356-018-3759-3
41. Kim DH, Rajapaksha LG, Gunasekara CW, Wimalasena SH, Pathirana HN, Kim SR, et al. Phylogenetic relationships and antibiotic resistance of Vibrio parahaemolyticus isolates related to acute hepatopancreatic necrosis disease in Korea. Aquaculture. (2021) 545:737253. doi: 10.1016/j.aquaculture.2021.737253
42. Dahanayake PS, Hossain S, Wickramanayake M, Heo GJ. Prevalence of virulence and extended-spectrum beta-lactamase (ESBL) genes harbouring Vibrio spp. isolated from cockles (Tegillarca granosa) marketed in Korea. Lett Appl Microbiol. (2020) 71:61–9. doi: 10.1111/lam.13232
43. Hossain S, Wickramanayake M, Dahanayake PS, Heo GJ. Occurrence of virulence and extended-spectrum beta-lactamase leterminants in Vibrio spp. Isolated from marketed hard-shelled mussel (Mytilus coruscus). Microb Drug Resist. (2020) 26:391–401. doi: 10.1089/mdr.2019.0131
44. Ottaviani D, Medici L, Talevi G, Napoleoni M, Serratore P, Zavatta E, et al. Molecular characterization and drug susceptibility of non-O1/O139 V. cholerae strains of seafood, environmental and clinical origin, Italy. Food Microbiol. (2018) 72:82–8. doi: 10.1016/j.fm.2017.11.011
45. Serratore P, Zavatta E, Fiocchi E, Serafini E, Serraino A, Giacometti F, et al. Preliminary study on the antimicrobial susceptibility pattern related to the genotype of Vibrio vulnificus strains isolated in the north-western Adriatic Sea coastal area. Ital J Food Saf. (2017) 6:6843. doi: 10.4081/ijfs.2017.6843
46. Mani Y, Mansour W, Mammeri H, Denamur E, Saras E, Boujâafar N, Bouallègue O, J-Madec Y, Haenni M KPC-3-producing ST167 Escherichia coli from mussels bought at a retail market in Tunisia. J Antimicrob Chemotherapy. (2017) 72:2403–4. doi: 10.1093/jac/dkx124
47. Mani Y, Mansour W, Lupo A, Saras E, Bouallègue O, Madec JY, et al. Spread of blaCTX-M-15-producing Enterobacteriaceae and OXA-23-producing Acinetobacter baumannii sequence type 2 in Tunisian seafood. Antimicrob Agents Chemother. (2018) 62:e00727–18. doi: 10.1128/AAC.00727-18
48. Briet A, Helsens N, Delannoy S, Debuiche S, Brisabois A, Midelet G, et al. NDM-1-producing Vibrio parahaemolyticus isolated from imported seafood. J Antimicrob Chemother. (2018) 73:2578–9. doi: 10.1093/jac/dky200
49. Sperling L, Alter T, Huehn S. Prevalence and antimicrobial resistance of Vibrio spp. in retail and farm shrimps in Ecuador. J Food Prot. (2015) 78:2089–92. doi: 10.4315/0362-028X.JFP-15-160
50. Hamza D, Dorgham S, Ismael E, Siel-Moez A, Elhariri M, Elhelw R, et al. Emergence of β-lactamase- and carbapenemase-producing Enterobacteriaceae at integrated fish farms. Antimicrob Resist Infect Control. (2020) 9. doi: 10.1186/s13756-020-00736-3
51. Huang YT, Cheng JF, Chen SY, Hong YK, Wu ZY, Liu PY. Draft genome sequence of carbapenem-resistant Shewanella algae strain AC isolated from small abalone (Haliotis diversicolor). J Glob Antimicrob Resist. (2018) 14:65–7. doi: 10.1016/j.jgar.2018.06.005
52. Lee YH, Tung KC, Cheng JF, Wu ZY, Chen SY, Hong YK. Genomic characterization of carbapenem-resistant Shewanella algae isolated from Asian hard clam (Meretrix lusoria). Aquaculture. (2019) 500:300–4. doi: 10.1016/j.aquaculture.2018.10.028
53. Zago V, Veschetti L, Patuzzo C, Malerba G, MLleo M. Shewanella algae and Vibrio spp. strains isolated in Italian aquaculture farms are reservoirs of antibiotic resistant genes that might constitute a risk for human health. Mar Pollut Bull. (2020) 154:111057. doi: 10.1016/j.marpolbul.2020.111057
54. Loest D, Uhland FC, Young KM, Li XZ, Mulvey MR, Reid-Smith R, et al. Carbapenem-resistant Escherichia coli from shrimp and salmon available for purchase by consumers in Canada: a risk profile using the Codex framework. Epidemiol Infect. (2022) 150:e148. doi: 10.1017/S0950268822001030
55. Schar D, Klein EY, Laxminarayan R, Gilbert M, Van Boeckel TP. Global trends in antimicrobial use in aquaculture. Sci Rep. (2020) 10:21878. doi: 10.1038/s41598-020-78849-3
56. Mollenkopf DF, Kleinhenz KE, Funk JA, Gebreyes WA, Wittum TE. Salmonella enterica and Escherichia coli harboring blaCMY in retail beef and pork products. Foodborne Pathog Dis. (2011) 8:333–6. doi: 10.1089/fpd.2010.0701
57. Mollenkopf DF, Stull JW, Mathys DA, Bowman AS, Feicht SM, Grooters SV, et al. Carbapenemase-producing Enterobacteriaceae recovered from the environment of a swine farrow-to-finish operation in the United States. Antimicrob Agents Chemother. (2017) 61:e01298-16. doi: 10.1128/AAC.01298-16
58. He W, Gao M, Lv L, Wang J, Cai Z, Bai Y, et al. Persistence and molecular epidemiology of bla(NDM)-positive Gram-negative bacteria in three broiler farms: A longitudinal study (2015-2021). J Hazard Mater. (2023) 446:130725. doi: 10.1016/j.jhazmat.2023.130725
59. Borowiak M, Szabo I, Baumann B, Junker E, Hammerl JA, Kaesbohrer A, et al. VIM-1-producing Salmonella Infantis isolated from swine and minced pork meat in Germany. J Antimicrob Chemotherapy. (2017) 72:2131–3. doi: 10.1093/jac/dkx101
60. Fischer J, San José M, Roschanski N, Schmoger S, Baumann B, Irrgang A, et al. Spread and persistence of VIM-1 Carbapenemase-producing Enterobacteriaceae in three German swine farms in 2011 and 2012. Vet Microbiol. (2017) 200:118–23. doi: 10.1016/j.vetmic.2016.04.026
61. Fischer J, Rodríguez I, Schmoger S, Friese A, Roesler U, Helmuth R, et al. Salmonella enterica subsp. enterica producing VIM-1 carbapenemase isolated from livestock farms. J Antimicrob Chemother. (2013) 68:478–80. doi: 10.1093/jac/dks393
62. Zhang WJ, Lu Z, Schwarz S, Zhang RM, Wang XM, Si W, et al. Complete sequence of the blaNDM-1-carrying plasmid pNDM-AB from Acinetobacter baumannii of food animal origin. J Antimicrob Chemother. (2013) 68:1681–2. doi: 10.1093/jac/dkt066
63. Kong LH, Lei CW, Ma SZ, Jiang W, Liu BH, Wang YX, et al. Various sequence types of Escherichia coli isolates coharboring blaNDM-5 and mcr-1 genes from a commercial swine farm in China. Antimicrob Agents Chemother. (2017) 61:e02167–16. doi: 10.1128/AAC.02167-16
64. Carfora V, Diaconu EL, Ianzano A, Di Matteo P, Amoruso R, Dell'Aira E, et al. The hazard of carbapenemase OXA-181-producing Escherichia coli spreading in pig and veal calf holdings in Italy in the genomics era: Risk of spill over and spill back between humans and animals. Front Microbiol. (2022) 13:1016895. doi: 10.3389/fmicb.2022.1016895
65. Wang Y, Wu C, Zhang Q, Qi J, Liu H, Wang Y, et al. Identification of New Delhi metallo-β-lactamase 1 in Acinetobacter lwoffii of food animal origin. PLoS ONE. (2012) 7:e37152. doi: 10.1371/journal.pone.0037152
66. Sadek M, Poirel L, Nordmann P, Nariya H, Shimamoto T, Shimamoto T. Genetic characterisation of NDM-1 and NDM-5-producing Enterobacterales from retail chicken meat in Egypt. J Glob Antimicrob Resist. (2020) 23:70–1. doi: 10.1016/j.jgar.2020.07.031
67. Zhu X, Zhang Y, Shen Z, Xia L, Wang J, Zhao L, et al. Characterization of NDM-1-Producing Carbapenemase in Proteus mirabilis among Broilers in China. Microorganisms. (2021) 9:2443. doi: 10.3390/microorganisms9122443
68. Xie X, Zhang J, Wang HN, Lei CW. Whole genome sequence of a New Delhi metallo-β-lactamase 1-producing Proteus mirabilis isolate SNYG35 from broiler chicken in China. J Glob Antimicrob Resist. (2021) 24:266–9. doi: 10.1016/j.jgar.2020.12.023
69. Algammal AM, Hashem HR, Alfifi KJ, Hetta HF, Sheraba NS, Ramadan H, et al. atpD gene sequencing, multidrug resistance traits, virulence-determinants, and antimicrobial resistance genes of emerging XDR and MDR-Proteus mirabilis. Sci Rep. (2021) 11:9476. doi: 10.1038/s41598-021-88861-w
70. Hamza E, Dorgham SM, Hamza DA. Carbapenemase-producing Klebsiella pneumoniae in broiler poultry farming in Egypt. J Glob Antimicrob Resist. (2016) 7:8–10. doi: 10.1016/j.jgar.2016.06.004
71. Shi X, Li Y, Yang Y, Shen Z, Cai C, Wang Y, et al. High prevalence and persistence of carbapenem and colistin resistance in livestock farm environments in China. J Hazard Mater. (2021) 406:124298. doi: 10.1016/j.jhazmat.2021.125951
72. Poirel L, Berçot B, Millemann Y, Bonnin RA, Pannaux G, Nordmann P. Carbapenemase-producing Acinetobacter spp. in Cattle, France. Emerg Infect Dis. (2012) 18:523–5. doi: 10.3201/eid1803.111330
73. Webb HE, Bugarel M, den Bakker HC, Nightingale KK, Granier SA, Scott HM, et al. Carbapenem-resistant bacteria recovered from faeces of dairy cattle in the High Plains Region of the USA. PLoS ONE. (2016) 11:e0147363. doi: 10.1371/journal.pone.0147363
74. Ghatak S, Singha A, Sen A, Guha C, Ahuja A, Bhattacharjee U, et al. Detection of New Delhi metallo-beta-lactamse and extended-spectrum beta-lactamse genes in Escherichia coli isolated from mastitic milk samples. Transbound Emerg Dis. (2013) 60:385–9. doi: 10.1111/tbed.12119
75. Yaici L, Haenni M, Saras E, Boudehouche W, Touati A, Madec JY. blaNDM-5-carrying IncX3 plasmid in Escherichia coli ST1284 isolated from raw milk collected in a dairy farm in Algeria. J Antimicrob Chemother. (2016) 71:2671–2. doi: 10.1093/jac/dkw160
76. He T, Wang Y, Sun L, Pang M, Zhang L, Wang R. Occurrence and characterization of blaNDM-5-positive Klebsiella pneumoniae isolates from dairy cows in Jiangsu, China. J Antimicrob Chemother. (2017) 72:90–4. doi: 10.1093/jac/dkw357
77. Klotz P, Göttig S, Leidner U, Semmler T, Scheufen S, Ewers C. Carbapenem-resistance and pathogenicity of bovine Acinetobacter indicus-like isolates. PLoS ONE. (2017) 12:e0171986. doi: 10.1371/journal.pone.0171986
78. Vikram A, Schmidt JW. Functional bla(KPC-2) Sequences are present in US beef cattle feces regardless of antibiotic use. Foodborne Pathog Dis. (2018) 15:444–8. doi: 10.1089/fpd.2017.2406
79. Murugan MS, Sinha DK, Vinodh Kumar OR, Yadav AK, Pruthvishree BS, Vadhana P, et al. Epidemiology of carbapenem-resistant Escherichia coli and first report of blaVIM carbapenemases gene in calves from India. Epidemiol Infect. (2019) 147:e159. doi: 10.1017/S0950268819000463
80. Elshafiee EA, Nader SM, Dorgham SM, Hamza DA. Carbapenem-resistant Pseudomonas aeruginosa originating from farm animals and people in Egypt. J Vet Res. (2019) 63:333–7. doi: 10.2478/jvetres-2019-0049
81. Tshitshi L, Manganyi MC, Montso PK, Mbewe M, Ateba CN. Extended spectrum beta-Lactamase-resistant determinants among carbapenem-resistant Enterobacteriaceae from beef cattle in the North West Province, South Africa: a critical assessment of their possible public health implications. Antibiotics. (2020) 9:820. doi: 10.3390/antibiotics9110820
82. Tello M, Oporto B, Lavín JL, Ocejo M, Hurtado A. Characterization of a carbapenem-resistant Escherichia coli from dairy cattle harbouring blaNDM-1 in an IncC plasmid. J Antimicrob Chemother. (2022) 77:843–5. doi: 10.1093/jac/dkab455
83. Alam M, Rasool MH, Khan I, Khurshid M, Aslam B. Multilocus sequence typing of carbapenem-resistant Acinetobacter baumannii isolates harboring blaOXA-23 and blaIMP in cattle from Punjab, Pakistan. Microb Drug Resist. (2022) 28:997–1002. doi: 10.1089/mdr.2022.0083
84. Ben Haj Yahia A, Tayh G, Landolsi S, Maamar E, Galai N, Landoulsi Z„ et al. First report of OXA-48 and IMP genes among extended-spectrum beta-lactamase-producing Escherichia coli isolates from diarrheic calves in Tunisia. Microb Drug Resist. (2023) 29:150–62. doi: 10.1089/mdr.2022.0129
85. Girlich D, Naas T, Nordmann P. Biochemical characterization of the naturally occurring oxacillinase OXA-50 of Pseudomonas aeruginosa. Antimicrob Agents Chemother. (2004) 48:2043–8. doi: 10.1128/AAC.48.6.2043-2048.2004
86. Liakopoulos A, van der Goot J, Bossers A, Betts J, Brouwer MSM, Kant A, et al. Genomic and functional characterisation of IncX3 plasmids encoding blaSHV-12 in Escherichia coli from human and animal origin. Sci Rep. (2018) 8:7674. doi: 10.1038/s41598-018-26073-5
87. Su Y, Xin L, Zhang F, Peng C, Li Z, Liu C, et al. Drug resistance analysis of three types of avian-origin carbapenem-resistant Enterobacteriaceae in Shandong Province, China. Poult Sci. (2023) 102:102483. doi: 10.1016/j.psj.2023.102483
88. Evans BA, Amyes SG. OXA β-lactamases. Clin Microbiol Rev. (2014) 27:241–63. doi: 10.1128/CMR.00117-13
89. Purkait D, Ahuja A, Bhattacharjee U, Singha A, Rhetso K, Dey TK, et al. Molecular characterization and computational modelling of New Delhi metallo-β-lactamase-5 from an Escherichia coli Isolate (KOEC3) of bovine origin. Indian J Microbiol. (2016) 56:182–9. doi: 10.1007/s12088-016-0569-5
90. Pesapane R, Ponder M, Alexander KA. Tracking pathogen transmission at the human-wildlife interface: banded mongoose and Escherichia coli. Ecohealth. (2013) 10:115–28. doi: 10.1007/s10393-013-0838-2
91. Poirel L, Héritier C, Nordmann P. Chromosome-encoded ambler class D beta-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob Agents Chemother. (2004) 48:348–51. doi: 10.1128/AAC.48.1.348-351.2004
92. Potron A, Poirel L, Nordmann P. Origin of OXA-181, an emerging carbapenem-hydrolyzing oxacillinase, as a chromosomal gene in Shewanella xiamenensis. Antimicrob Agents Chemother. (2011) 55:4405–7. doi: 10.1128/AAC.00681-11
93. Poirel L, Figueiredo S, Cattoir V, Carattoli A, Nordmann P. Acinetobacter radioresistens as a silent source of carbapenem resistance for Acinetobacter spp. Antimicrob Agents Chemother. (2008) 52:1252–6. doi: 10.1128/AAC.01304-07
94. Fischer J, Schmoger S, Jahn S, Helmuth R, Guerra B. NDM-1 carbapenemase-producing Salmonella enterica subsp. enterica serovar Corvallis isolated from a wild bird in Germany. J Antimicrob Chemother. (2013) 68:2954–6. doi: 10.1093/jac/dkt260
95. Qin S, Fu Y, Zhang Q, Qi H, Wen JG, Xu H, et al. High incidence and endemic spread of NDM-1-positive Enterobacteriaceae in Henan Province, China. Antimicrob Agents Chemother. (2014) 58:4275–82. doi: 10.1128/AAC.02813-13
96. Dolejska M, Masarikova M, Dobiasova H, Jamborova I, Karpiskova R, Havlicek M, et al. High prevalence of Salmonella and IMP-4-producing Enterobacteriaceae in the silver gull on Five Islands, Australia. J Antimicrob Chemother. (2016) 71:63–70. doi: 10.1093/jac/dkv306
97. Vittecoq M, Laurens C, Brazier L, Durand P, Elguero E, Arnal A, et al. VIM-1 carbapenemase-producing Escherichia coli in gulls from southern France. Ecol Evol. (2017) 7:1224–32. doi: 10.1002/ece3.2707
98. Bachiri T, Bakour S, Lalaoui R, Belkebla N, Allouache M, Rolain JM, et al. Occurrence of carbapenemase-producing Enterobacteriaceae isolates in the wildlife: first report of OXA-48 in wild boars in Algeria. Microb Drug Resist. (2018) 24:337–45. doi: 10.1089/mdr.2016.0323
99. Liao X, Yang RS, Xia J, Chen L, Zhang R, Fang LX, et al. High colonization rate of a novel carbapenem-resistant Klebsiella lineage among migratory birds at Qinghai Lake, China. J Antimicrob Chemotherapy. (2019) 74:2895–903. doi: 10.1093/jac/dkz268
100. Lim SR, Lee D-H, Park SY, Lee S, Kim HY, Lee M-S, et al. Wild Nutria (Myocastor coypus) is a potential reservoir of carbapenem-resistant and zoonotic Aeromonas spp. in Korea. Microorganisms. (2019) 7:224. doi: 10.3390/microorganisms7080224
101. Gharout-Sait A, Touati A, Ahmim M, Brasme L, Guillard T, Agsous A, de Champs C. Occurrence of carbapenemase-producing Klebsiella pneumoniae in bat guano. Microbial Drug Resist. (2019) 25:1057–62. doi: 10.1089/mdr.2018.0471
102. Mairi A, Barraud O, Muggeo A, de Champs C, Touati A. Genomic analysis of a multidrug-resistant Klebsiella pneumoniae ST11 strain recovered from barbary deer (Cervus elaphus barbarus) in Akfadou forest, Algeria. J Global Antimicrob Resist. (2019) 22:300–4. doi: 10.1016/j.jgar.2020.04.027
103. Duff JP, AbuOun M, Bexton S, Rogers J, Turton J, Woodford N, et al. Resistance to carbapenems and other antibiotics in Klebsiella pneumoniae found in seals indicates anthropogenic pollution. Vet Rec. (2020) 187:154. doi: 10.1136/vr.105440
104. Ahlstrom CA, Frick A, Pongratz C, Spink K, Xavier C, Bonnedahl J, et al. Genomic comparison of carbapenem-resistant Enterobacteriaceae from humans and gulls in Alaska. J Glob Antimicrob Resist. (2021) 25:23–5. doi: 10.1016/j.jgar.2021.02.028
105. VinodhKumar OR, Karikalan M, Ilayaraja S, Sha AA, Singh BR, Sinha DL, et al. Multi-drug resistant (MDR), extended spectrum beta-lactamase (ESBL) producing and carbapenem resistant Escherichia coli in rescued Sloth bears (Melursus ursinus), India. Vet Res Commun. (2021) 45:163–70. doi: 10.1007/s11259-021-09794-3
106. Torres RT, Cunha MV, Ferreira H, Fonseca C, Palmeira JD, A. high-risk carbapenem-resistant Pseudomonas aeruginosa clone detected in red deer (Cervus elaphus) from Portugal. Sci Total Environ. (2022) 829:154699. doi: 10.1016/j.scitotenv.2022.154699
107. Ahlstrom CA, Woksepp H, Sandegren L, Mohsin M, Hasan B, Muzyka D, et al. Genomically diverse carbapenem resistant Enterobacteriaceae from wild birds provide insight into global patterns of spatiotemporal dissemination. Sci Total Environ. (2022) 824:153632. doi: 10.1016/j.scitotenv.2022.153632
108. Sellera FP, Cardoso B, Fuentes-Castillo D, Esposito F, Sano E, Fontana H, et al. Genomic analysis of a highly virulent NDM-1-producing Escherichia coli ST162 infecting a pygmy sperm whale (Kogia breviceps) in South America. Front Microbiol. (2022) 13:915375. doi: 10.3389/fmicb.2022.915375
109. Girlich D, Poirel L, Nordmann P. Novel ambler class A carbapenem-hydrolyzing beta-lactamase from a Pseudomonas fluorescens isolate from the Seine River, Paris, France. Antimicrob Agents Chemother. (2010) 54:328–32. doi: 10.1128/AAC.00961-09
110. Hubeny J, Korzeniewska E, Buta-Hubeny M, Zieliński W, Rolbiecki D, Harnisz M. Characterization of carbapenem resistance in environmental samples and Acinetobacter spp. isolates from wastewater and river water in Poland. Sci Total Environ. (2022) 822:153437. doi: 10.1016/j.scitotenv.2022.153437
111. Harmon DE, Miranda OA, McCarley A, Eshaghian M, Carlson N, Ruiz C. Prevalence and characterization of carbapenem-resistant bacteria in water bodies in the Los Angeles-Southern California area. Microbiol Open. (2019) 8:e00692. doi: 10.1002/mbo3.692
112. Aubron C, Poirel L, Ash RJ, Nordmann P. Carbapenemase-producing Enterobacteriaceae, US rivers. Emerg Infect Dis. (2005) 11:260–4. doi: 10.3201/eid1102.030684
113. Dewi D, Götz B, Thomas T. Diversity and genetic basis for carbapenem resistance in a coastal marine environment. Appl Environ Microbiol. (2020) 86. doi: 10.1128/AEM.02939-19
114. Sellera FP, Da Silva LCBA, Lincopan N. Rapid spread of critical priority carbapenemase-producing pathogens in companion animals: a One Health challenge for a post-pandemic world. J Antimicrob Chemotherapy. (2021) 76:2225–9. doi: 10.1093/jac/dkab169
115. Sellera FP, Lincopan N. Zooanthroponotic transmission of high-risk multi-drug-resistant pathogens: a neglected public health issue. J Infect Public Health. (2019) 12:294–5. doi: 10.1016/j.jiph.2018.12.013
116. Fernandes MR, Sellera FP, Moura Q, Carvalho MP, Rosato PN, Cerdeira L, et al. Zooanthroponotic transmission of drug-resistant Pseudomonas aeruginosa, Brazil. Emerg Infect Dis. (2018) 24:1160–2. doi: 10.3201/eid2406.180335
117. Smith A, Wayne AS, Fellman CL, Rosenbaum MH. Usage patterns of carbapenem antimicrobials in dogs and cats at a veterinary tertiary care hospital. J Vet Intern Med. (2019) 33:1677–85. doi: 10.1111/jvim.15522
118. Grönthal T, Österblad M, Eklund M, Jalava J, Nykäsenoja S, Pekkanen K, et al. Sharing more than friendship – transmission of NDM-5 ST167 and CTX-M-9 ST69 Escherichia coli between dogs and humans in a family, Finland 2015. Euro Surveill. (2018) 23:1700497. doi: 10.2807/1560-7917.ES.2018.23.27.1700497
119. Zafer MM, Al-Agamy MH, El-Mahallawy HA, Amin MA, Ashour EDS. Dissemination of VIM-2 producing Pseudomonas aeruginosa ST233 at tertiary care hospitals in Egypt. BMC Infect Dis. (2015) 15:122. doi: 10.1186/s12879-015-0861-8
120. Wright LL, Turton JF, Livermore DM, Hopkins KL, Woodford N. Dominance of international ‘high-risk clones' among metallo-β-lactamase producing Pseudomonas aeruginosa in the UK. J Antimicrob Chemother. (2015) 70:103–10. doi: 10.1093/jac/dku339
121. Sader HS, Reis AO, Silbert S, Gales AC. IMPs, VIMs and SPMs: the diversity of metallo-β-lactamases produced by carbapenem-resistant Pseudomonas aeruginosa in a Brazilian hospital. Clin Microbiol Infect. (2005) 11:73–6. doi: 10.1111/j.1469-0691.2004.01031.x
122. Wang MG, Fang C, Liu KD, Wang LL, Sun RY, Zhang RM, et al. Transmission and molecular characteristics of blaNDM-producing Escherichia coli between companion animals and their healthcare providers in Guangzhou, China. J Antimicrob Chemother. (2022) 77:351–5. doi: 10.1093/jac/dkab382
123. Garcia-Fierro R, Drapeau A, Dazas M, Saras E, Rodrigues C, Brisse S, et al. Comparative phylogenomics of ESBL-, AmpC- and carbapenemase-producing Klebsiella pneumoniae originating from companion animals and humans. J Antimicrob Chemotherapy. (2022) 77:1263–71. doi: 10.1093/jac/dkac041
124. Silva JMD, Menezes J, Marques C, Pomba CF. Companion animals-an overlooked and misdiagnosed reservoir of carbapenem resistance. Antibiotics. (2022) 11:533. doi: 10.3390/antibiotics11040533
125. Stolle I, Prenger-Berninghoff E, Stamm I, Scheufen S, Hassdenteufel E, Guenther S, et al. Emergence of OXA-48 carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in dogs. J Antimicrob Chemother. (2013) 68:2802–8. doi: 10.1093/jac/dkt259
126. Smet A, Boyen F, Pasmans F, Butaye P, Martens A, Nemec A, et al. OXA-23-producing Acinetobacter species from horses: a public health hazard? J Antimicrob Chemother. (2012) 67:3009–10. doi: 10.1093/jac/dks311
127. Sellera FP, Fernandes MR, Ruiz R, Falleiros ACM, Rodrigues FP, Cerdeira L, et al. Identification of KPC-2-producing Escherichia coli in a companion animal: a new challenge for veterinary clinicians. J Antimicrob Chemother. (2018) 73:2259–61. doi: 10.1093/jac/dky173
128. Sellera FP, Fuga B, Fontana H, Esposito F, Cardoso B, Konno S, et al. Detection of IncN-pST15 one-health plasmid harbouring blaKPC-2 in a hypermucoviscous Klebsiella pneumoniae CG258 isolated from an infected dog, Brazil. Transbound Emerg Dis. (2021) 68:3083–8. doi: 10.1111/tbed.14006
129. Gomez-Simmonds A, Hu Y, Sullivan SB, Wang Z, Whittier S, Uhlemann AC. Evidence from a New York City hospital of rising incidence of genetically diverse carbapenem-resistant Enterobacter cloacae and dominance of ST171, 2007-14. J Antimicrob Chemother. (2016) 71:2351–3. doi: 10.1093/jac/dkw132
130. Daniels JB, Chen L, Grooters SV, Mollenkopf DF, Mathys DA, Pancholi P, et al. Enterobacter cloacae complex Sequence Type 171 Isolates Expressing KPC-4 Carbapenemase Recovered from Canine Patients in Ohio. Antimicrob Agents Chemother. (2018) 62:e01161-18. doi: 10.1128/AAC.01161-18
131. Abraham S, O'Dea M, Trott DJ, Abraham RJ, Hughes D, Pang S, et al. Isolation and plasmid characterization of carbapenemase (IMP-4) producing Salmonella enterica Typhimurium from cats. Sci Rep. (2016) 6:35527. doi: 10.1038/srep35527
132. Shaheen BW, Nayak R, Boothe DM. Emergence of a New Delhi metallo-β-lactamase (NDM-1)-encoding gene in clinical Escherichia coli isolates recovered from companion animals in the United States. Antimicrob Agents Chemother. (2013) 57:2902–3. doi: 10.1128/AAC.02028-12
133. Cui L, Lei L, Lv Y, Zhang R, Liu X, Li M, et al. bla(NDM-1)-producing multidrug-resistant Escherichia coli isolated from a companion dog in China. J Glob Antimicrob Resist. (2018) 13:24–7. doi: 10.1016/j.jgar.2017.10.021
134. Gentilini F, Turba ME, Pasquali F, Mion D, Romagnoli N, Zambon E, et al. Hospitalized pets as a source of carbapenem-resistance. Front Microbiol. (2018) 9:2872. doi: 10.3389/fmicb.2018.02872
135. Kuang X, Yang R, Ye X, Sun J, Liao X, Liu Y, et al. NDM-5-Producing Escherichia coli co-harboring mcr-1 gene in companion animals in China. Animals. (2022) 12:1310. doi: 10.3390/ani12101310
136. Pulss S, Stolle I, Stamm I, Leidner U, Heydel C, Semmler T, et al. Multispecies and clonal dissemination of OXA-48 carbapenemase in Enterobacteriaceae from companion animals in Germany, 2009-2016. Front Microbiol. (2018) 9:1265. doi: 10.3389/fmicb.2018.01265
137. Yousfi M, Touati A, Muggeo A, Mira B, Asma B, Brasme L, et al. Clonal dissemination of OXA-48-producing Enterobacter cloacae isolates from companion animals in Algeria. J Glob Antimicrob Resist. (2018) 12:187–91. doi: 10.1016/j.jgar.2017.10.007
138. González-Torralba A, Oteo J, Asenjo A, Bautista V, Fuentes E, Alós JI. Survey of carbapenemase-producing Enterobacteriaceae in companion dogs in Madrid, Spain. Antimicrob Agents Chemother. (2016) 60:2499–501. doi: 10.1128/AAC.02383-15
139. Hyun JE, Chung TH, Hwang CY. Identification of VIM-2 metallo-β-lactamase-producing Pseudomonas aeruginosa isolated from dogs with pyoderma and otitis in Korea. Vet Dermatol. (2018) 29:186–e68. doi: 10.1111/vde.12534
140. Brilhante M, Menezes J, Belas A, Feudi C, Schwarz S, Pomba C, et al. OXA-181-Producing extraintestinal pathogenic Escherichia coli Sequence Type 410 isolated from a dog in Portugal. Antimicrob Agents Chemother. (2020) 64:e02298-19. doi: 10.1128/AAC.02298-19
141. Pomba C, Endimiani A, Rossano A, Saial D, Couto N, Perreten V. First report of OXA-23-mediated carbapenem resistance in sequence type 2 multidrug-resistant Acinetobacter baumannii associated with urinary tract infection in a cat. Antimicrob Agents Chemother. (2014) 58:1267–8. doi: 10.1128/AAC.02527-13
142. Ewers C, Klotz P, Leidner U, Stamm I, Prenger-Berninghoff E, Göttig S, et al. OXA-23 and ISAba1-OXA-66 class D β-lactamases in Acinetobacter baumannii isolates from companion animals. Int J Antimicrob Agents. (2017) 49:37–44. doi: 10.1016/j.ijantimicag.2016.09.033
143. Shen Y, Hu F, Wang Y, Yin D, Yang L, Chen Y, et al. Transmission of carbapenem resistance between human and animal NDM-positive Escherichia coli strains. Engineering. (2022) 15:24–33. doi: 10.1016/j.eng.2021.07.030
144. Li J, Bi Z, Ma S, Chen B, Cai C, He J, et al. Inter-host transmission of carbapenemase-producing Escherichia coli among humans and backyard animals. Environ Health Perspect. (2019) 127:107009. doi: 10.1289/EHP5251
145. Wang Y, Zhang R, Li J, Wu Z, Yin W, Schwarz S, et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat Microbiol. (2017) 6:16260. doi: 10.1038/nmicrobiol.2016.260
146. One Health High-Level Expert Panel (OHHLEP) Adisasmito WB Almuhairi S Behravesh CB Bilivogui P Bukachi SA . (One Health: A new definition for a sustainable and healthy future. PLoS Pathog. (2022) 18:e1010537. doi: 10.1371/journal.ppat.1010537
147. Das S. The crisis of carbapenemase-mediated carbapenem resistance across the human-animal-environmental interface in India. Infect Dis Now. (2023) 53:104628. doi: 10.1016/j.idnow.2022.09.023
148. Velazquez-Meza ME, Galarde-López M, Carrillo-Quiróz B, Alpuche-Aranda CM. Antimicrobial resistance: One Health approach. Vet World. (2022) 15:743–9. doi: 10.14202/vetworld.2022.743-749
149. Ferri M, Ranucci E, Romagnoli P, Giaccone V. Antimicrobial resistance: a global emerging threat to public health systems. Crit Rev Food Sci Nutr. (2017) 57:2857–76. doi: 10.1080/10408398.2015.1077192
150. Ottaviani D, Leoni F, Serram Serracca L, Decastelli L, Rocchegiani E, Masini L, et al. Nontoxigenic Vibrio parahemolyticus strains causing acute gastroenteritis. J Clin Microbiol. (2012) 50:4141–3. doi: 10.1128/JCM.01993-12
151. Bauer MJ, Stone-Garza KK, Croom D, Andreoli C, Woodson P, Graf PC-F, et al. Shewanella algae infections in United States naval special warfare trainees. Open Forum Infect Dis. (2019) 6:ofz442 doi: 10.1093/ofid/ofz442
152. Annavajhala MK, Gomez-Simmonds A, Uhlemann A-C. (2019). Multidrug-resistant enterobacter cloacae complex emerging as a global, diversifying threat. Front. Microbiol. 10:44. doi: 10.3389/fmicb.2019.00044
153. Pillai SD. Source tracing in Thailand following contamination of dog chew toys with Salmonella. In:Hoorfar J, , editor. Woodhead Publishing Series in Food Science, Technology and Nutrition, Case Studies in Food Safety and Authenticity. Woodhead Publishing. p. 123–9. doi: 10.1533/9780857096937.2.123
154. Bulens SN Yi SH, Walters MS, Jacob JT, Bower C, Reno J, et al. Carbapenem-nonsusceptible Acinetobacter baumannii, 8 US metropolitan areas, 2012–2015. Emerg Infect Dis. (2018) 24:727–34. doi: 10.3201/eid2404.171461
155. Müller S, Janssen T, Wieler LH. Multidrug resistant Acinetobacter baumannii in veterinary medicine - emergence of an underestimated pathogen? BMTW. (2014) 127:435–46.
156. van der Kolk JH, Endimiani A, Graubner C, Gerber V, Perreten V. Acinetobacter in Veterinary Medicine with emphasis on A. baumannii. J Glob Antimicrob Resist. (2018) 16:59–71. doi: 10.1016/j.jgar.2018.08.011
157. Wareth G, Neubauer H, Sprague LD. Acinetobacter baumannii – a neglected pathogen in veterinary and environmental health in Germany. Vet Res Commun. (2019) 43:1–6. doi: 10.1007/s11259-018-9742-0
158. Cole D, Drum DJ, Stalknecht DE, White DG, Lee MD, Ayers S, et al. Free-living Canada geese and antimicrobial resistance. Emerg Infect Dis. (2005) 11:935–8. doi: 10.3201/eid1106.040717
Keywords: carbapenem resistance, One Health approach, food-producing animals, carbapenemase producers, transmission
Citation: Ramírez-Castillo FY, Guerrero-Barrera AL and Avelar-González FJ (2023) An overview of carbapenem-resistant organisms from food-producing animals, seafood, aquaculture, companion animals, and wildlife. Front. Vet. Sci. 10:1158588. doi: 10.3389/fvets.2023.1158588
Received: 04 February 2023; Accepted: 23 May 2023;
Published: 15 June 2023.
Edited by:
João Pedro Rueda Furlan, University of São Paulo, BrazilReviewed by:
Fábio Sellera, University of São Paulo, BrazilCopyright © 2023 Ramírez-Castillo, Guerrero-Barrera and Avelar-González. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alma L. Guerrero-Barrera, bGlsaWFuLmd1ZXJyZXJvQGVkdS51YWEubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.