
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci. , 24 March 2023
Sec. Animal Reproduction - Theriogenology
Volume 10 - 2023 | https://doi.org/10.3389/fvets.2023.1152103
 Samiullah Khan1,2†
Samiullah Khan1,2† Muhammad Ameen Jamal3†
Muhammad Ameen Jamal3† Ibrar Muhammad Khan1*
Ibrar Muhammad Khan1* Irfan Ullah4Abdul Jabbar5Nazir Muhammad Khan6
Irfan Ullah4Abdul Jabbar5Nazir Muhammad Khan6 Yong Liu1*
Yong Liu1*Goats are generally called a “poor man's cow” because they not only provide meat and milk but also other assistance to their owners, including skins for leather production and their waste, which can be used as compost for fertilizer. Multiple ovulation and embryo transfer (MOET) is an important process in embryo biotechnology, as it increases the contribution of superior female goats to breeding operations. The field of assisted reproductive biotechnologies has seen notable progress. However, unlike in cattle, the standard use of superovulation and other reproductive biotechnologies has not been widely implemented for goats. Multiple intrinsic and extrinsic factors can alter the superovulatory response, significantly restricting the practicability of MOET technology. The use of techniques to induce superovulation is a crucial step in embryo transfer (ET), as it accelerates the propagation of animals with superior genetics for desirable traits. Furthermore, the conventional superovulation techniques based on numerous injections are not appropriate for animals and are labor-intensive as well as expensive. Different approaches and alternatives have been applied to obtain the maximum ovarian response, including immunization against inhibin and the day-0 protocol for the synchronization of the first follicular wave. While there are several studies available in the literature on superovulation in cattle, research on simplified superovulation in goats is limited; only a few studies have been conducted on this topic. This review describes the various treatments with gonadotropin that are used for inducing superovulation in various dairy goat breeds worldwide. The outcomes of these treatments, in terms of ovulation rate and recovery of transferrable embryos, are also discussed. Furthermore, this review also covers the recovery of oocytes through repeated superovulation from the same female goat that is used for somatic cell nuclear transfer (SCNT).
Goat production plays an important role in the livelihoods of farmers, especially in developing countries. In the last 20 years, the world's population of goats has increased by 49%, whereas sheep and cattle have shown a slow increase of 15 and 14%, respectively (1). Goats provide various animal by-products such as meat, milk, and hides to poor families; they are also a viable option for commercial farming, making a valuable contribution to the livestock industry (2–4).
The application of multiple ovulation and embryo transfer (MOET) plays a vital role in the global trade of genetic resources, conserving endangered species, minimizing the risk of exotic diseases and the cost of production, and eliminating transportation-related stress (5, 6). This technology made great progress in the 1970s, largely due to its remarkable practice in cattle (4). The sequence of events leading to the ET usually starts with superovulation. Superovulation is an important phase in the MOET program, as it has the potential to increase the number of genetically superior donors, especially when there is a high demand for such animals (7, 8). Under normal circumstances, a goat typically ovulates 1–3 eggs per estrus cycle. However, by administering approximate doses of FSH hormone through superovulation, approximately 10–20 available oocytes with 13 transferrable embryos (8) can be obtained (4). The fundamental idea behind superovulation involves the artificial administration of exogenous gonadotropins, such as FSH or PMSG, to promote the development of a greater number of follicles, ultimately leading to ovulation (4).
However, the widespread implementation of MOET in goats has been hindered due to the high cost of hormones (9) and the unpredictable response of donor animals to superovulation hormones. Ovarian responses in goats can vary and are affected by a number of factors such as breed, age, nutrition, animal management practices and selection, stress, gonadotrophin supply, and seasonal cyclic activity (8, 10–21). These factors could negatively influence superovulation outcomes by reducing the quality of oocytes, which ultimately decreases the number of transferrable embryos for ET. In addition, there are some other disadvantages associated with the use of multiple injections (6–10 injections over 3–5 days), including the time and labor investments required (22).
A robust superovulation approach that meets the requirements of both researchers and producers in terms of ovarian response predictability and dependability has yet to be created. For instance, a simplified superovulatory protocol that involves fewer FSH injections (i.e., single or only two injections) while maintaining the same level of ovarian response in terms of recovered and transferrable embryos would be ideal (21). Further, these simplified protocols will also facilitate easier implementation for a large herd of animals while requiring less time and resources (18–21).
This review examined recent studies on various hormonal protocols for inducing superovulation in goats, including attempts to simplify these protocols, and discusses the outcomes of the approaches as well as the various factors that can affect superovulation. Additionally, new approaches applied to superovulation in goats are explored.
Superovulation can be performed mostly using exogenous gonadotrophins such as PMSG and FSH. However, other rarely used hormones are horse anterior pituitary (HAP) extract, human chorionic gonadotrophin (hCG), or gonadotrophin-releasing hormone (GnRH) (17). PMSG is a unique member of the gonadotropin family, also known as equine chorionic gonadotropin (eCG), which is a complex glycoprotein containing both FSH and LH-like activities (23). This hormone is fairly valuable for inducing superovulation in goats (24, 25). This hormone has an advantage over FSH because it can easily be applied to an open flock in a single subcutaneous or intramuscular injection given 1 or 2 days prior to the last synchronization treatment at a dose of 750–2,000 IU. Owing to its simplicity, a single injection not only causes less stress by minimizing excessive handling but is also cost-effective in comparison to FSH multiple injection protocols (24, 26, 27). However, PMSG_influenced the steroidal hormone pattern, leading to the premature regression of the corpus luteum (28) and an increase in the number of persistent large follicles that eventually failed to ovulate, resulting in reduced ovarian response (27, 29). It also causes follicular steroid secretion, which can interfere with sperm and gamete transport, oocyte maturation, and early preimplantation embryo development, thereby altering the endogenous endocrine environment (30). Due to its long half-life (72 h), PMSG injection can cause a high incidence of anovulatory follicles and early degeneration of the corpus luteum (CL) in goats' ovaries.
FSH is secreted from an anterior lobe of the pituitary, causing the growth and development of small follicles on the ovaries. This hormone is commonly injected multiple times over a period of 3 to 4 days during the follicular phase and 48 h before the sponges are removed. However, as discussed earlier, multiple injections are time-consuming, laborious, and stressful for animals (31, 32), which ultimately decreases the superovulatory response (33).
Comparative studies have shown that FSH is superior to PMSG and produces more transferable embryos (5, 19, 34). In this study, FSH therapy also produced more embryos than eCG therapy, and embryos were recovered from indigenous dairy goat breeds such as Jamunapari, Angora, Jakhrana, Tellicherry, and crossbred (Boer Katjang) goats (17, 19, 34–38) (Table 1).
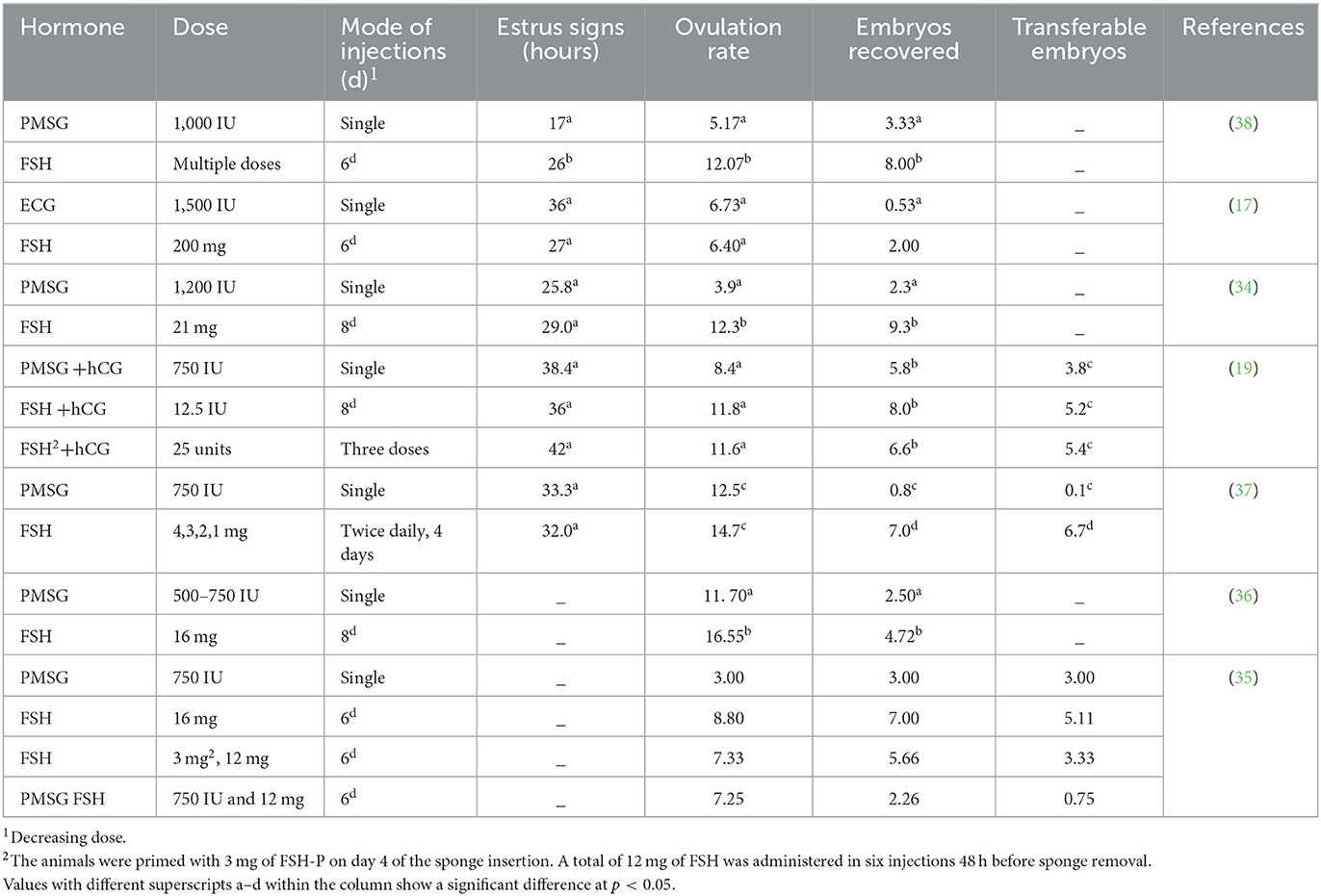
Table 1. Comparison of FSH and PMSG for the induction of the superovulatory response.
Sustained exposure to low levels of FSH hormone in multiple doses is required to achieve superovulation (39). Few successful attempts have been made to simplify these protocols by combining FSH with a low dose of PMSG in a single injection (40) or by using a single injection of porcine FSH (41). In Canindé goats, the mean number of cumulus-oocyte complexes (COCs) recovered did not show any significant difference between the treatments. The number of COCs retrieved was 10.8 oocytes for a single injection of 70 mg Folltropin-V plus 200 IU eCG and 11.7 oocytes for five doses of 120 mg (42).
In contrast, Lehloenya (43) found that using a simplified superovulation treatment with FSH and PMSG was less effective than the traditional protocol based on multiple FSH injections. The lower response to the simplified superovulation treatment could be due to an increase in the number of large follicles during a single injection of FSH and PMSG (44). These contradictory results suggest that the ovarian response may depend on the type of FSH preparation used and the appropriate dosage of FSH and PMSG.
Many studies obtained satisfactory ovarian responses using simplified superovulatory techniques in cattle (45–49). Many research groups also obtained a similar ovarian response in sheep when simplified protocols were compared to multiple injection protocols (32, 39, 41, 50–56). The studies conducted on goats concerning the subsequent stage of embryo yield and quality are limited, suggesting the need for further research, particularly on large-scale embryonic production in the context of the superovulation protocol followed by AI (55, 57). Moreover, the endocrinological bases of simplified protocols are not well understood, leaving considerable room for studying their relationship with the endocrine profile. Such research could potentially reduce the labor and hormone costs associated with large-scale production.
Human chorionic gonadotrophin (hCG) or gonadotrophin-releasing hormone (GnRH) is rarely used in goats, as well as in sheep and cattle. Treating crossbred goats with hCG yielded a higher number of CL and recovered oocytes (10.9 and 3.10, respectively) compared to goats treated with GnRH (1.90 and 0.7, respectively) after supplementing hCG and GnRH with eCG at the rate of 1,500 IU. However, despite the higher number of recovered oocytes, there was no difference in the average number of embryonic recoveries, which made it difficult to enhance embryonic production (58).
Various factors limit the practicability of MOET for goats and other domestic animals. Therefore, it is important to manipulate these factors to improve the ovarian response. Major factors are discussed in this section, including several extrinsic factors (e.g., sources, the purity of gonadotrophins, and their administration) and intrinsic factors (e.g., breed, age, nutrition, and season).
The factors that contribute to the variability in the ovarian response of goats are shown in Figure 1.
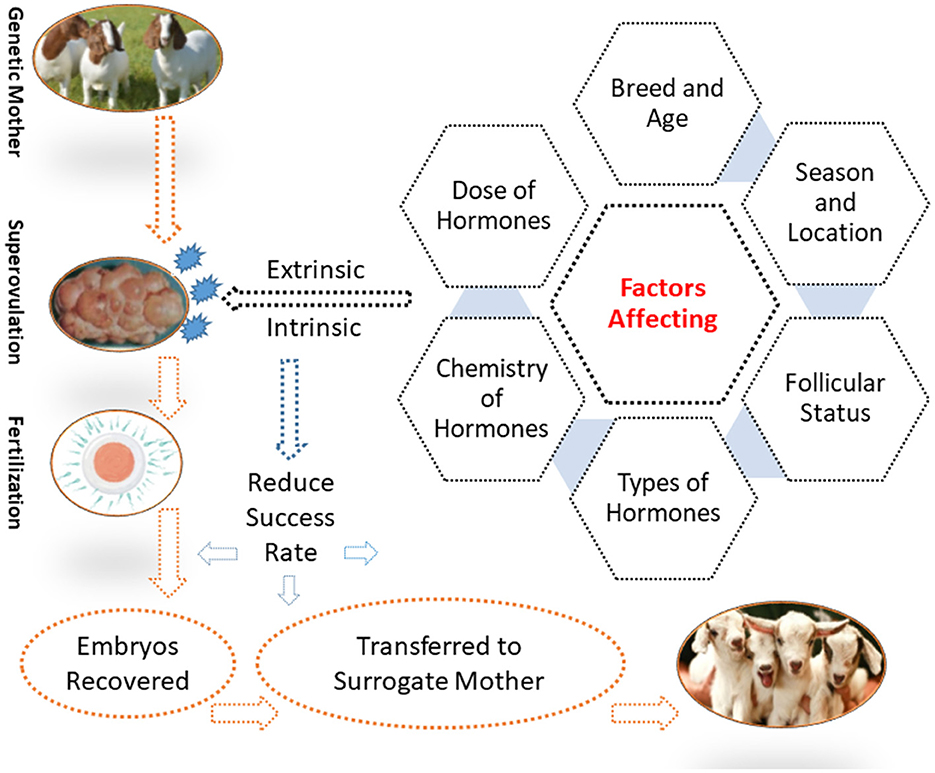
Figure 1. Factors affecting the superovulatory response in goats.
FSH was found to produce higher ovulation rates and transferrable embryos compared to other PMSGs and eCG. The type of hormones and the number of injections may contributed to the superovulatory response. Similar observations for FSH and PMSG were also made in ewes (24, 55, 59, 60). Hormonal protocols and their outcomes are discussed in detail in Table 1.
With varying ovarian responses to hormonal therapies, age may also be regarded as a limiting factor (6, 10). Mahmood et al. (36) reported a higher number of CL and recovered embryos (RE) (19.10 and 5.40) for the 4- to 6-year age group than (13.25 and 4.00) for the 1.5- to 3-year age group. A higher response was also obtained for 3–4-year-old goats than for 1–2-year-old goats (61). Similarly, Chang et al. (10) injected six or seven injections of FSH, given in decreasing doses over a four-day period, to goats of different ages. The results revealed a higher ovarian response, indicated by better ovulation rates, among goats aged 12–23 months compared to those aged 7–12 months. Additionally, a better proportion of transferable embryos was observed in the 1–5-year-old group than in the 0.7–1.0 year age group. In sheep, it was found that older ewes (24–60 months) produced a greater number of recovered and transferrable embryos compared to the younger group (8–12 months) when given a superovulation treatment consisting of eight decreasing doses of 160–200 mg (16).
The higher ovarian response observed in older goats could be due to a fully functional ovary, prior pregnancy experience (1–2 pregnancies), and a large body frame (10, 16). In sheep, a study reported a higher ovulation rate among 1–2-year-old ewes compared to 3–4-year-old ewes. This may be due to young animals experiencing fewer disease incidents and health problems (62).
The effectiveness of the FSH dose was also investigated in relation to an ovarian response (Table 2). Sánchez-Davila et al. (9) reported that administering 145 and 215 mg of FSH in seven doses over 3.5 days produced similar ovarian responses, but significantly higher responses were observed when the dose was reduced to 80 mg using the same protocol. Abdullah et al. (15) found no significant difference in the number of recovered and transferrable embryos while injecting FSH doses of 8.8 mg and 14.08 mg, starting 48 h before CIDR removal. Similarly, Rahman et al. (17) did not find any significant difference in the ovulation rate and transferrable embryos while injecting 5 and 3 mg of FSH/kg body weight, but they did observe a decreased ovarian response while using a higher dose of 8 mg/kg body weight. In sheep, a few studies (55, 60, 63) found no difference with a slight reduction in the FSH dose. These results suggest that a small reduction in dosage may not have a significant impact on ovarian response. However, a large reduction may lead to a decreased ovarian response. Therefore, an optimal reduction in hormonal dose can reduce the cost of hormones without negatively affecting the ovarian response, especially in large-scale in vivo embryonic production.
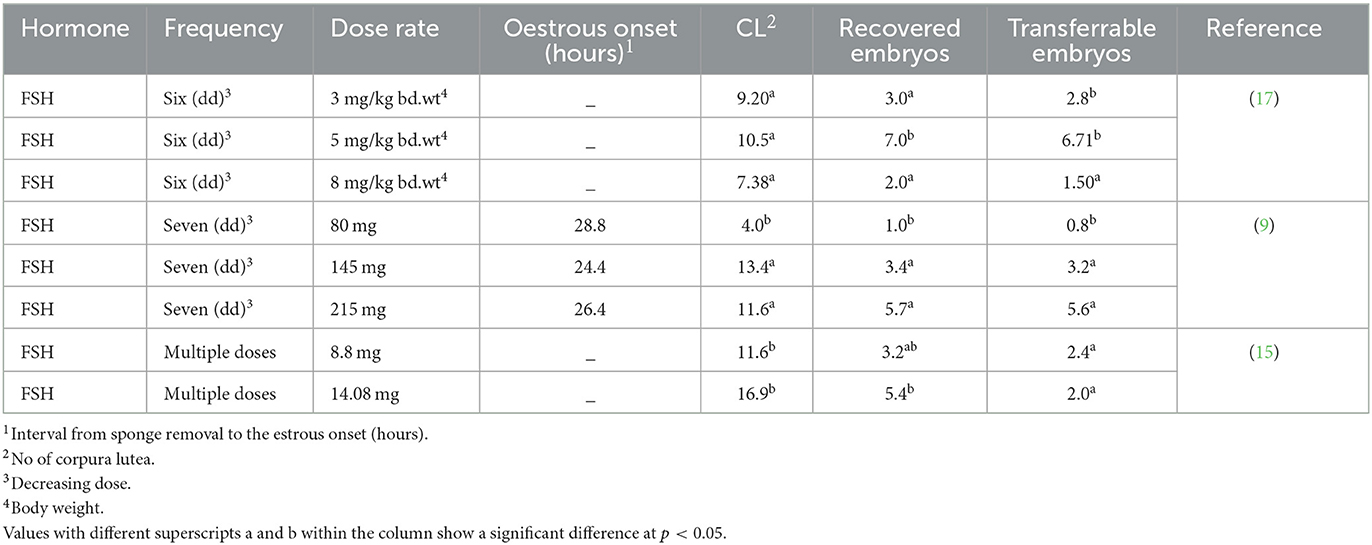
Table 2. Effect of different hormone doses on the superovulatory response.
The ovarian response of the breed to superovulatory hormones was found to be uneven and inconsistent. Nuti et al. (64) found that, when administering a total of 15 mg of FSH through six injections, a higher percentage of ovarian response was observed in Nubian female goats compared to Alpine goats. Conversely, Kiessling et al. (65) found no breed effect on ovarian response in Saanen, Alpine, Nubian, and La Mancha goats when administering superovulation. However, they did not observe limited differences in ovarian response between the Boer and indigenous breeds when using pFSH 200 mg/dose injected in seven decreasing doses. The differences were noted in terms of total CL (12.78 and 14.37), total embryos (8.40 and 8.12), and transferable embryos (6.60 and 6.00) (66).
The administration of different hormones during the superovulation process led to diverse ovarian responses in different breeds of sheep. A significantly higher number of CL and embryos (13.7 and 7.9) were observed for the Rubia del Molar breed compared to the Negra de Colmenar (10 and 4.3) and Manchega ewes (9.8 and 6.7) after administering the same eight decreasing doses of oFSH for superovulation (13). Furthermore, a higher number of CL and transferable embryos were observed in Chios than in Friesian breeds when subjected to multiple doses of pFSH (13, 67). Conversely, there was no difference in ovarian response between Corriedale and Bond donor ewes when injected with a split-single dose of 180 mg of FSH dissolved in 10 mg/ml 750 kDa hyaluronan (56). These results indicated that variation in ovarian response among different breeds subjected to the same hormonal protocol may be due to the varying kinetic behavior of the exogenous gonadotrophin, follicular status, and function, or environmental influences (S.7).
After superovulation protocols, the ovarian response during different seasons was found to be inconsistent (Table 3). Specifically, a significantly higher ovulation rate (28.7) and number of recovered embryos (15.1) were obtained from Nubian goats during the early breeding season compared to the ovulation rate (9.30) and the number of recovered embryos (3.3) during the late breeding season (68). However, Chang et al. (10) obtained a better ovarian response in breeding (17.24) than in non-breeding (10.40) and late breeding (14.32) seasons in Shandong province, China.
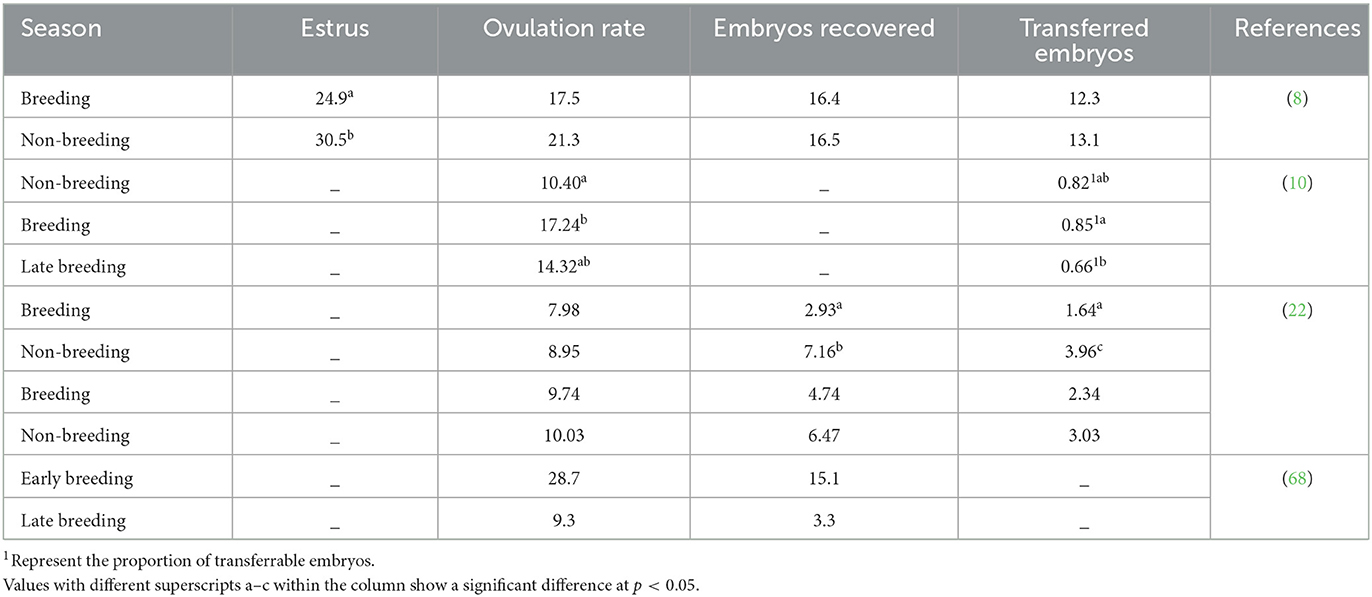
Table 3. Different seasons influence the ovarian response.
Superovulatory treatment and season did not affect the ovulation rate (7.98 vs. 8.95). However, the number of recovered (2.93 vs.7.16) and transferrable (1.64 vs. 3.96 embryos) embryos were higher in the non-breeding season than in the breeding season, respectively (22). There was no significant difference between the ovulation rate (10.2 vs. 9.22) and the total number of embryos (4.10 vs. 5.44) during the breeding season when we administered FSH in seven decreasing doses and three FSH plus one PMSG injection group, respectively. Similarly, the ovulation rate (10.74 and 9.32) and the total number of embryos (5.95 and 7.00) were also similar between these two groups during the non-breeding season when the same superovulatory protocol was used (22). Additionally, there was no significant difference between the breeding and non-breeding seasons when we treated goats using these two protocols. Similarly, similar numbers of recovered embryos (16.4 and 16.5) were obtained when superovulation with multiple dosages of FSH was carried out during the breeding and non-breeding seasons, respectively (8). Baril and Vallet (69) found no difference in ovarian response when inducing superovulation in Alpine goats with porcine FSH during and out of the breeding season. The lack of differences in the number of ovarian structures recovered and transferable embryos between seasons recorded per donor may be due to the similarity of climatic conditions during both seasons (9). It is evident that season has an effect on the time to onset and the duration of the induced estrous period following superovulation in goats, which is important to consider, especially for fixed-time AI (8).
Inhibin, a member of the transforming growth factor beta (TGF-β) superfamily, is a gonadal hormone that has a negative effect on the secretion of FSH from the gonadotropic cells of the pituitary gland (70). The use of immunization against inhibin, along with conventional superovulation, may be considered a viable alternative to achieve maximum output in goats (71). The use of inhibin antiserum in treated goats resulted in a higher ovarian response compared to the control group, with a greater number of follicles (13.5 vs. 5.3) and ovulation rate (4.2 vs. 1.8) (72). There was approximately a 4-fold increase in the ovulation rate (7.6) in goats actively immunized against inhibin compared to the control (1.7) group. The onset of estrus was also shorter (46.8 h) for the inhibin-immunized group than the control group (54.4 h) (73). Holtz et al. (74) reported obtaining 5.5 transferrable embryos using a combination of inhibin immunization and conventional superovulation protocol, which is lower than the 13.1 embryos obtained by Lehloenya et al. (8) using conventional superovulation alone. Studies have shown that the use of immunization against inhibin results in significantly lower ovulation rates and fewer embryos recovered compared to conventional FSH protocols. This finding is consistent with previous studies conducted by Pendleton et al. (37), Lehloenya et al. (8), Palanisamy et al. (38), and a few other studies (75, 76). However, in ewes that were both treated with FSH and actively immunized against porcine inhibin α-subunit, a higher ovulation rate of 12.1 was observed. In contrast, ewes that were treated with the conventional FSH superovulation protocol alone had an ovulation rate of 5.0 (77). In cattle, using inhibin immunization in combination with a superovulation protocol resulted in a higher embryo yield and ovarian response compared to conventional protocols (78, 79). In mice, the use of inhibin antiserum in combination with conventional superovulatory protocols has recently resulted in a one-third increase in the number of oocytes compared to the control group (80). Inhibin immunization, along with conventional or simplified superovulatory methods, has not been broadly studied in goats. Further studies are required to study its effect on the ovulation rate, in vitro fertilization among different breeds, and the further processing of oocytes to observe their quality and quantity.
Recent studies on follicular dynamics have enabled researchers to develop new superovulation protocols for embryonic production, such as the day-0 protocol. The day-0 protocol comprised the synchronization of ovulation and the emergence of the first wave. The day-0 protocol involved inserting CIDR for 5 days, followed by an injection of (PG) F2α to induce luteolysis. To synchronize the ovulation procedure, a dose of approximately 200–300 IU of eCG was injected upon CIDR withdrawal. Additionally, a single dose of GnRH was administered 36 h after CIDR withdrawal to ensure ovulation. Day 0 was estimated as 84 h after CIDR withdrawal (i.e., soon after ovulation). The FSH injections were given two times/day with decreasing doses starting 84 h after CIDR removal, and two half-doses of PGF2a were administered with the 5th and 6th FSH treatments. To synchronize the LH peak and ovulation, the GnRH analog was administered 24 h after the first PGF2a treatment. Timed AI was performed using laparoscopy under moderate sedation with frozen-tawed semen at 16 and 26 h after GnRH administration (14, 81).
In goats, wave-like patterns occur during follicular dynamics (82). The most frequent finding is the occurrence of four follicular waves during an estrous cycle in goats, but high variability among cycles has also been reported. Each follicular wave is preceded by a transient increase in the concentration of FSH. Generally, one to three follicles grow to a diameter of 5 mm after the development of the waves. However, the remaining (medium, 4–5 mm; small, 2–4 mm) enter the atresia. Further aspects of follicular wave patterns in goats have been reviewed by Menchaca et al. (14) and Rubianes and Menchaca (83).
To synchronize ovulation and the emergence of the first follicular wave, Menchaca et al. (81) compared the day-0 protocol with the traditional multiple FSH protocol with six decreasing doses. During the breeding season, the yield of transferable embryos increased from 2.6 in goats treated with the conventional superovulatory protocol to 4.9 in goats treated with the day-0 procedure.
Similarly, during the non-breeding season, the use of the conventional protocol resulted in an ovulation rate of 10.7, embryos recovered at a rate of 7.6, and a yield of transferrable embryos (4.2). In comparison, the day-0 protocol increased the ovulation rate to 14.3, and the number of embryos recovered remained the same at 7.6. However, the yield of transferrable embryos increased to 5.9. The day-0 protocol also yielded a high ovarian response compared to the traditional protocol. However, as there are few studies on day 0 of the superovulatory protocols, further research is warranted to improve embryonic production efficacy.
In goats, the presence of a dominant follicle had a deleterious effect on follicle recruitment and the superovulatory response. However, using the day-0 protocol resulted in a higher percentage of females responding, an increased number of CL, and a higher number of grade 1 and 2 embryos during both breeding and non-breeding seasons in comparison to the traditional protocol. Large follicles at the beginning of the superovulation treatment are associated with the total number of unfertilized ova, while medium follicles (4–5 or 6 mm) at the beginning of the superovulation treatment are directly associated with the number of recovered and viable embryos and transferable embryos (66, 84). The ovarian response in terms of the number of corpora lutea (15.3) was positively correlated to the total number of follicles with a diameter of 2–6 mm at the beginning of the FSH treatment. A high number of larger follicles (≥7 mm) had a negative effect on the ovulation rate (84). The modified (day 0) protocol should be initiated when there is a suitable number of follicles from the second category present on the ovaries (85).
The administration of hormonal treatment at intervals to recover oocytes repeatedly from the same animal is known as “repeated superovulation.” A study conducted by Lehloenya et al. (86) reported a significant reduction in the number of embryos recovered (11.7 vs. 3.8) and the number of transferrable embryos (10.7 vs. 3.8) during the first and repeated superovulation in the natural breeding season; however, the ovulation rate was not affected (14.8 and 16.8). Chang et al. (10) did not observe any reduction in the number of oocytes recovered during the first and second oocyte cycles of superovulation, but significant decreases were observed during the third superovulation cycle. This decrease in ovarian response may be attributed to genital tract adhesion after repeated flushing (10); thus, limited potential for surgically obtaining repeated embryo collections from the same animal is demonstrated (16, 87).
These findings suggested that the decrease in ovarian response during repeated superovulation could be due to the formation of post-operative adhesions in the reproductive tract, which may have an adverse effect on the ovarian function or uterine cells, resulting in a reduced number of embryos recovered during successive treatments (16, 32). It is evident that repeated use of animals for superovulation treatment during the same breeding season may negatively influence the consecutive recovery of oocytes. Proper rest is required for the animal after surgical oocyte removal (16). Another primary problem contributing to a lower number of recovered embryos is the production of high anti-eCG antibodies due to repeated administration of eCG hormones (32). Increasing the interval between repeated recoveries from the same animal and improving embryo recovery methods (16) could improve the potential for surgically recovering more embryos.
Embryos produced through in vivo fertilization generally exhibit greater developmental competence regarding blastocyst formation rate compared to embryos obtained through in vitro fertilization (IVF) and SCNT-derived embryos. This may be because the oocytes used for IVF or SCNT are mostly obtained from an abattoir source, undergo in vitro maturation (IVM), have a relatively hyperoxic environment, and tend to have high oxidative stress levels. Furthermore, the overproduction of ROS induces apoptotic cell death, thereby impairing the quality and developmental potential of oocytes (88, 89). However, these situations do not occur in vivo cultures. Defects in an abnormal epigenetic status have been reported for SCNT-derived embryos due to inadequate remodeling of the donor nucleus.
Enucleated metaphase II oocytes, as recipient cytoplasm (90), are selected from the oocytes. Choosing to collect oocytes from slaughterhouse ovaries is a more convenient and cost-effective option compared to inducing ovulation in donor animals through multiple exogenous gonadotrophin hormone injections (91). Moreover, due to the low efficiency of the SCNT procedure, a relatively large number of oocytes are required to produce live offspring or conduct other meaningful experiments. While selecting oocytes from slaughterhouse ovaries may be easier and cost-effective, there is a lack of proper record-keeping regarding the animals' reproductive performance, age, management practices, and genetic origin.
There was no difference in the number of transferable embryos obtained from the cytoplasts of FSH-stimulated ovaries and embryos from the fusion of cytoplasts from abattoir ovaries (91). In sheep, a higher number of quality oocytes, pregnancy rates, and live kid rates were observed when using reconstructed embryos obtained from enucleated recipient oocytes obtained through FSH treatment compared to using ovaries from slaughterhouses (90). FSH pre-treatment improved oxygen consumption and OCT4 and IFN-τ expression in SCNT embryos, which indicates that FSH has a positive effect on oocyte quality (92). Therefore, it has become important to identify a noninvasive and non-disruptive method for selecting oocytes before culture (93). Oocytes obtained through superovulation may have a key role in obtaining better efficiency for nuclear transfer. In cattle and goats, it has been observed that in vitro-matured oocytes result in high prenatal and postnatal losses, poor embryo developmental competence, and lower pregnancy rates compared to in vitro-matured oocytes (91, 94–97). It was found that the oviductal oocytes (in vivo-matured oocytes) have a greater electrical pulse than the follicular oocytes (in vitro-matured oocytes). One of the 17 recipients delivered a normal live birth, and two pregnancies were achieved by transferring in vivo-matured embryos. On the other hand, no live births were obtained from in vitro-matured oocytes. Similarly, Martins et al. (98) also observed that no SCNT pregnancies reached term with the use of in vitro-matured oocytes, whereas in vivo-matured oocytes resulted in the successful birth of two transgenic cloned kids.
Reggio et al. (91) conducted a study comparing the in vitro developmental potential of nuclear transfer embryos produced by fusing transgenic donor cells with cytoplasts derived from the ovaries of animals that had undergone superovulation induced through multiple injections of FSH and abattoir ovaries. The results showed that the rate of fusion of NT embryos reconstructed from oocytes from either FSH-stimulated or abattoir-derived ovaries was 63 and 57%, respectively, which shows that oocyte source had no effect on embryo development or the overall pregnancy rate. However, the performance of oocytes selected using these vague criteria is often problematic and inaccurate, making it difficult to distinguish oocytes with different levels of developmental competence (93). Another study conducted by Peura et al. (99) reported no significant difference in blastocyst development rates (40.4 and 35.8%) between the high and low diet groups, but there was a significant difference in the established pregnancies (50 and 28.6%), resulting in live births. Multiple studies have shown that, after reprogramming by oocytes, SCNT embryos exhibit distinct gene expression patterns compared to in vivo-derived or in vitro-fertilized (IVF) embryos (100). In sheep, SCNT embryos have been reported to exhibit slightly higher overall methylation levels than IVF embryos. However, the donor cell chromatin showed a conserved distribution when the transferred donor cell nuclei were compared to IVF embryonic nuclei (101).
The superovulation process yields a large number of oocytes. However, their maturation rate can be hampered. Despite this barrier, it was found that the potential of mature oocytes to be fertilized and to develop into blastocysts is not affected by their origin (102). Despite substantial advancements in assisted reproduction technologies in recent years, the pregnancy rate remained low since a large proportion of transferred embryos fail to implant (103). Exogenous gonadotropin treatment leads to greater concentrations of circulating steroids, which may influence either oocyte or embryo quality, as well as the oviductal and/or uterine environment. This can also disrupt the synchronization that typically occurs between the embryo and the endometrium during the implantation process. Hence, a link may exist between the use of gonadotrophins for ovarian stimulation and the observed low implantation rate and gestational problems (104).
Several studies have shown that the yield of embryos produced through the induction of superovulation can be highly variable and negatively influenced by factors such as the effects on the oocyte during follicular growth or directly during embryo development in the oviduct and/or uterus (105, 106). The mechanism by which oocytes and embryos develop under hormonal superstimulation has an effect on abnormal endocrine conditions compared to those developed in unstimulated animals, ultimately leading to low developmental potential (107). Ovarian stimulation, or “superovulation,” induced by exogenous hormones may stimulate follicular development and oocyte maturation, resulting in the production of a greater number of oocytes. However, exogenous hormones affect the natural hormone environment, which is necessary for female reproduction, particularly follicle development and oocyte maturation (108). Superovulation may decrease the number of fertilized oocytes and preimplantation competence in vivo. The lower fertilization rate of oocytes obtained in vivo using superovulation methods may be attributed to sperm or oocyte transportation disturbances in the oviduct (109), alteration in the oviductal function such as the presence of carbohydrate residues on the epithelium of the ampulla (110), and the expression of specific genes (111). The variation in the timing of oocyte maturation between follicles during ovulation could also potentially lead to a reduced fertilization rate (112). The use of superovulation in ewes has been shown to reduce the number of sperm present in the oviduct (109). Previous studies on mice have shown that the COH procedures can cause delayed embryonic development, decreased implantation, and higher post-implantation loss (104). Another study reported that stimulated mouse embryos had delayed blastocyst formation, increased incidence of zonal lysis, and blastocyst collapse compared to naturally cycling controls (113).
Some studies suggest that superovulation can cause alterations in oocyte properties, and the in vitro fertilization rate in cows may be adjusted by altering the timing of FSH administration relative to oocyte harvesting (114). Thus, superovulation is likely to result in the ovulation of abnormal oocytes in at least some instances (109). Studies on mice have demonstrated the detrimental impact of ovarian stimulation on oocyte/embryo developmental competence, where the transfer of embryos from superovulated donors resulted in a considerably lower implantation rate in the control recipients than when embryos from control donors were transferred (104). Implications for the embryo as shown in mice, embryos produced from oocytes retrieved during superovulation may have reduced competence for preimplantation development in vivo (104, 113) in mice and cows (115). The oviduct and its fluid provide favorable conditions for gamete maturation, gamete transit, fertilization, and early embryonic development, which are crucial for mammalian reproduction and are regulated by steroids (115, 116).
These results showed that embryonic development in vivo before the transfer and superovulation was associated with reduced embryo competence for establishing pregnancy in recipients (113), increased fetal loss rates after the establishment of pregnancy (104), placental dysfunction (117), and reductions in fetal weight (117). The effects of superovulation may be influenced by the specific technique used to induce it, as well as other factors such as animal strain. Superovulation has no deleterious influence on embryonic and fetal development in pigs (118). Disruption of the development of embryos from superovulated females could have effects on the oocyte or alterations in the function of the oviduct and endometrium due to high concentrations of ovarian steroids in the blood of superovulated females.
Several studies on mammalian oocytes and embryos have shown that superovulation results in aberrant gene expression, including genes that are believed to be important for oocyte quality, cell cycle regulation, and inhibition (119). The gene expression patterns during blastocytes were found to be different in the embryos derived from the superovulated females. Ovarian stimulation triggers a cascade of hormonal and physiological events that create a different environment for oocyte maturation compared to naturally matured oocytes. This may also result in variations in the timing of ovulation. In addition, endometrial gene expression was altered by superovulation in both cows and humans (104). Embryos that are produced in vivo and those produced in vitro also exhibit differences in their gene expression and patterns of DNA methylation (120, 121). Therefore, the use of exogenous hormone stimulation can result in epigenetic changes in both oocytes and developing embryos. To understand the consequences of these changes during development, it is important to conduct controlled experiments that can dissect the epigenetic alterations that occur.
Successful studies have been conducted on simplified superovulation protocols using a combination of FSH and eCG in cattle and sheep. However, there were insufficient data on the successful simplification of superovulatory protocols with satisfactory ovarian responses in goats. Unlike cattle, further studies on simplification protocols based on the endocrinology profile of goats may not only reduce the cost of hormones but also make this technology more applicable to goats. Studies on immunization against inhibin along conventional superovulation protocols have shown successful results in cattle; however, there are limited data available, indicating a need for further studies to achieve a maximum ovarian response. The synchronization of the first follicular wave using the day-0 protocol showed a satisfactory response in terms of transferable embryos.
However, further studies are required to simplify this protocol by reducing the number of FSH injections from multiple injections to a single injection. A significant number of embryos undergo deterioration, and pregnancy rates remain low because a larger number of transplanted embryos fail to implant. In addition, a validation technique is required to determine whether candidate genes and putative SNP markers may contribute to oocyte quality, cell cycle regulation, inhibition, or a higher concentration of circulating hormones that can compromise the quality of embryos in the oviductal and even uterine environment, resulting in a reduced competency of embryos to establish a pregnancy.
Conceptualization: SK, MJ, and YL. Software: IK. Validation: YL and SK. Resources, supervision, and funding acquisition: YL. Writing—original draft preparation: SK and MJ. Writing—review and editing: IK, AJ, and IU. Visualization: NK. Project administration: YL and IK. All authors have read and agreed to the published version of the manuscript.
Industry-University-Research Project of Fuyang Normal University (HX2021027000 and HX2022048000) funded this study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Cannas A, Tedeschi LO, Atzori AS, Lunesu MF. How can nutrition models increase the production efficiency of sheep and goat operations? Ani Front. (2019) 9:33–44. doi: 10.1093/af/vfz005
2. Iqbal A, Khan B, Tariq M, Mirza M. Goat-A potential dairy animal: present and future prospects. Pak J Agri Sci. (2008) 45:227–30.
3. Babar ME, Hussain T, Nadeem A, Jabeen R, Javed M. Genetic characterization of Azakheli buffalo breed of Pakistan using microsatellite DNA markers. Pak J Zool. (2009) 9:361–6. doi: 10.1007/s11033-011-0808-0
4. Luo J, Wang W, Sun S. Research advances in reproduction for dairy goats. Asian-Australas J Anim Sci. (2019) 32:1284–95. doi: 10.5713/ajas.19.0486
5. Ishwar A, Memon M. Embryo transfer in sheep and goats: a review. Small Ruminant Res. (1996) 19:35–43. doi: 10.1016/0921-4488(95)00735-0
6. González-Bulnes A, Baird DT, Campbell BK, Cocero MJ, García-García RM, Inskeep EK, et al. Multiple factors affecting the efficiency of multiple ovulation and embryo transfer in sheep and goats. Reprod Fert Dev. (2004) 16:421–35. doi: 10.1071/RD04033
7. Khan SU, Jamal MA, Wei HJ, Qing Y, Cheng W. Towards improving the outcomes of multiple ovulation and embryo transfer in sheep, with particular focus on donor superovulation. Vet Sci. (2022) 9:117. doi: 10.3390/vetsci9030117
8. Lehloenya K, Greyling J, Grobler S. Effect of season on the superovulatory response in Boer goat does. Small Rumin Res. (2008) 78:74–9. doi: 10.1016/j.smallrumres.2008.05.003
9. Sánchez-Dávila F, Ledezma-Torres R, Padilla-Rivas G, del Bosque-González A, González Gómez A, Bernal-Barragán H. Effect of three pFSH doses on superovulation and embryo quality in goats during two breeding seasons in North-eastern Mexico. Reprod Domest Anim. (2014) 49:40–3. doi: 10.1111/rda.12350
10. Chang Z, Fan X, Luo M, Wu Z, Tan J. Factors affecting superovulation and embryo transfer in Boer goats. Asian Austral J Anim. (2006) 19:341–6. doi: 10.5713/ajas.2006.341
11. O'callaghan D, Yaakub H, Hyttel P, Spicer L, Boland M. Effect of nutrition and superovulation on oocyte morphology, follicular fluid composition and systemic hormone concentrations in ewes. J Reprod Fertil. (2000) 118:303–14. doi: 10.1530/jrf.0.1180303
12. Terawaki Y, Asada Y. Relationships between distribution of number of transferable embryos and inbreeding coefficient in a MOET dairy cattle population. Asian Austral J Anim. (2002) 15:1686–9. doi: 10.5713/ajas.2002.1686
13. Ammoun I, Encinas T, Veiga-Lopez A, Ros J, Contreras I, Gonzalez-Anover P, et al. Effects of breed on kinetics of ovine FSH and ovarian response in superovulated sheep. Theriogenology. (2006) 66:896–905. doi: 10.1016/j.theriogenology.2006.02.024
14. Menchaca A, Vilariño M, Crispo M, De Castro T, Rubianes E. New approaches to superovulation and embryo transfer in small ruminants. Reprod Fertil Dev. (2009) 22:113–8. doi: 10.1071/RD09222
15. Abdullah R, Shariffah N, Khadijah MR, Wan WE. Effect of oFSH Dosages in Superovulation Protocolon Ovarian Responses in Goat. In SAADC 2011 Strategies and Challenges for Sustainable Animal Agriculture-Crop Systems, Volume III: Full Papers. Proceedings of the 3rd International Conference on Sustainable Animal Agriculture for Developing Countries, Nakhon Ratchasima, Thailand. Princeton, NJ: Citeseer (2011). p. 662–5.
16. Quan F, Zhang Z, An Z, Hua S, Zhao X, Zhang Y. Multiple factors affecting superovulation in Poll Dorset in China. Reprod Domest Anim. (2011) 46:39–44. doi: 10.1111/j.1439-0531.2009.01551.x
17. Rahman M, Rahman M, Khadijah WW, Abdullah R. Comparison of superovulatory effect of equine chorionic gonadotrophin and follicle stimulating hormone on embryo production in crossbred (Boer A-Katjang) goats. Pak J Zool. (2014) 46:819–26. Available online at: http://zsp.com.pk/pdf46/819-826%20_29_%20PJZ-1628-13%2026-5-14%20_Proof%20version_.pdf
18. Greyling J, Van der Nest M, Schwalbach L, Muller T. Superovulation and embryo transfer in South African Boer and indigenous feral goats. Small Ruminant Res. (2002) 43:45–51. doi: 10.1016/S0921-4488(01)00257-7
19. Goel A, Agrawal K. Ovulatory response and embryo yield in Jakhrana goats following treatments with PMSG and FSH. Trop Anim Health Pro. (2005) 37:549–58. doi: 10.1007/s11250-005-4223-1
20. Holtz W. Recent developments in assisted reproduction in goats. Small Ruminant Res. (2005) 60:95–110. doi: 10.1016/j.smallrumres.2005.06.032
21. Rahman A, Abdullah RB, Khadijah WW. A review of reproductive biotechnologies and their application in goat. Biotech. (2008) 7:371–84. doi: 10.3923/biotech.2008.371.384
22. Pintado B, Gutierrez-Adan A, Llano BP. Superovulatory response of Murciana goats to treatments based on PMSG/Anti-PMSG or combined FSH/PMSG administration. Theriogenology. (1998) 50:357–64. doi: 10.1016/S0093-691X(98)00145-9
23. Allen W, Moor R. The origin of the equine endometrial cups. J Reprod. (1972) 29:313–6. doi: 10.1530/jrf.0.0290313
24. Armstrong D, Evans G. Factors influencing success of embryo transfer in sheep and goats. Theriogenology. (1983) 19:31–42. doi: 10.1016/0093-691X(83)90121-8
25. Amoah E, Gelaye S. Superovulation, synchronization and breeding of does. Small Ruminant Res. (1990) 3:63–72. doi: 10.1016/0921-4488(90)90032-2
26. Foote R, Ellington J. Is a superovulated oocyte normal? Theriogenology. (1988) 29:111–23. doi: 10.1016/0093-691X(88)90035-0
27. D'Alessandro A, Martemucci G, Toteda F, Gambacorta M, Manchisi A. Superovulation and embryo production in ewes using a commercial p-FSH. Small Ruminant Res. (1996) 19:255–61. doi: 10.1016/0921-4488(95)00765-2
28. Senthilkumar P, Rajasundaram R, Selvaraju M, Kathiresan D. Effect of inclusion of norgestomet ear implants in the goat superovulation regime. Ind Vet J. (1998) 75:595–7.
29. Armstrong D, Pfitzner A, Warnes G, Ralph M, Seamark R. Endocrine responses of goats after induction of superovulation with PMSG and FSH. Reproduction. (1983) 67:395–401. doi: 10.1530/jrf.0.0670395
30. Gonzalez F, Calero P, Beckers JF. Induction of superovulation in domestic ruminants. In Renaville R, and Burny A, editors. Biotechnology in Animal Husbandry. New York, NY: Kluwer Academic Publishers (2001). p. 209–23.
31. Wang X, El-Gayar M, Knight P, Holtz W. The long-term effect of active immunization against inhibin in goats. Theriogenology. (2009) 71:318–22. doi: 10.1016/j.theriogenology.2008.07.024
32. Forcada F, Amer-Meziane MA, Abecia J, Maurel MC, Cebrián-Pérez J, Muiño-Blanco T, et al. Repeated superovulation using a simplified FSH/eCG treatment for in vivo embryo production in sheep. Theriogenology. (2011) 75:769–76. doi: 10.1016/j.theriogenology.2010.10.019
33. Kimura K. Superovulation with a single administration of FSH in aluminum hydroxide gel: a novel superovulation method for cattle. J Reprod Develop. (2016) 62:423–9. doi: 10.1262/jrd.2016-066
34. Pampukidou A, Alifakiotis T, Avdi M, Ivanova R. Superovulation and embryo transfer in goats by using PMSG or FSH. J Agr Sci Tech. (2011) 3:94–7.
35. Goel A, Agrawal K. Superovulation and embryo collection in Jamunapari goats. Theriogenology. (1990) 33:232. doi: 10.1016/0093-691X(90)90656-E
36. Mahmood S, Koul G, Biswas J. Comparative efficacy of FSH-P and PMSG on superovulation in Pashmina goats. Theriogenology. (1991) 35:1191–6. doi: 10.1016/0093-691X(91)90365-K
37. Pendleton R, Youngs C, Rorie R, Pool S, Memon M, Godke R. Follicle stimulating hormone versus pregnant mare serum gonadotropin for superovulation of dairy goats. Small Rumin Res. (1992) 8:217–24. doi: 10.1016/0921-4488(92)90042-3
38. Palanisamy A, Balasubramanian S, Rangasamy S. Effect of FSH, PMSG on superovulatory response and embryo recovery rate in Tellicherry goats. Ind J Anim Reprod. (2015) 32:17–18.
39. Evans G. Superovulation and embryo recovery in Merino ewes. Theriogenology. (1994) 41:192. doi: 10.1016/S0093-691X(05)80102-5
40. Batt P, Killeen I, Cameron A. Use of single or multiple injections of FSH in embryo collection programmes in goats. Reprod Fertil Dev. (1993) 5:49–56. doi: 10.1071/RD9930049
41. Peebles I, Kidd J. Single treatment superovulation in the cashmere goat using porcine follicle stimulating hormone. Theriogenology. (1994) 41:271. doi: 10.1016/S0093-691X(05)80181-5
42. Avelar SR, Moura RR, Sousa FC, Pereira AF, Almeida KC, Melo CH, et al. Oocyte production and in vitro maturation in Canindé goats following hormonal ovarian stimulation. Anim Reprod Sci. (2018) 9:27–32.
43. Lehloenya KC. Preliminary results evaluating a simplified superovulation protocol in Boer goats. Small Rumin Res. (2013) 113:171–4. doi: 10.1016/j.smallrumres.2013.03.017
44. Gibbons A, Bonnet FP, Cueto MI, Catala M, Salamone DF, Gonzalez-Bulnes A. Procedure for maximising oocytes harvest for invitro embryo production in small ruminants. Reprod Domest Anim. (2007) 42:423–6. doi: 10.1111/j.1439-0531.2006.00802.x
45. Bo G, Hockley D, Nasser L, Mapletoft R. Superovulatory response to a single subcutaneous injection of Folltropin-V in beef cattle. Theriogenology. (1994) 42:963–75. doi: 10.1016/0093-691X(94)90119-4
46. Yamamoto M, Ooe M, Kawaguchi M, Suzuki T. Superovulation in the cow with a single intramuscular injection of FSH dissolved in polyvinylpyrrolidone. Theriogenology. (1994) 41:747–55. doi: 10.1016/0093-691X(94)90184-K
47. Kimura K, Hirako M, Iwata H, Aoki M, Kawaguchi M, Seki M. Successful superovulation of cattle by a single administration of FSH in aluminum hydroxide gel. Theriogenology. (2007) 68:633–9. doi: 10.1016/j.theriogenology.2007.02.016
48. Tríbulo A, Rogan D, Tribulo H, Tribulo R, Alasino RV, Beltramo D, et al. Superstimulation of ovarian follicular development in beef cattle with a single intramuscular injection of Folltropin-V. Anim Reprod Sci. (2011) 129:7–13. doi: 10.1016/j.anireprosci.2011.10.013
49. Chasombat J, Sakhong D, Nagai T, Parnpai R, Vongpralub T. Superstimulation of follicular growth in Thai native heifers by a single administration of follicle stimulating hormone dissolved in polyvinylpyrrolidone. J Reprod Dev. (2012) 59:214–8. doi: 10.1262/jrd.2012-119
50. Maxwell W, Wilson H. Superovulation and embryo recovery in adult and maiden ewes treated with single and multiple injections of pituitary extract. In Proc Aust Soc Reprod Biol. (1990) 22:15.
51. Lopez-Sebastian A, Gomez-Brunet A, Lishman A, Johnson S, Inskeep E. Modification by propylene glycol of ovulation rate in ewes in response to a single injection of FSH. Reproduction. (1993) 99:437–42. doi: 10.1530/jrf.0.0990437
52. Dattena M, Vespignani S, Branca A, Gallus M, Ledda S, Naitana S, et al. Superovulatory response and quality of embryos recovered from anestrus ewes after a single injection of porcine FSH dissolved in polyvinylpyrrolidone. Theriogenology. (1994) 42:235–9. doi: 10.1016/0093-691X(94)90267-4
53. Meinecke-Tillman S, Lewalski H, Meinecke B. Induction of superovulation in Merinolandshaf ewes after single or multiple FSH injectons. Reprod Domest Anim. (1993) 28:433–42. doi: 10.1111/j.1439-0531.1993.tb01020.x
54. Yamada A, Kawana M, Tamura Y, Miyamoto A, Fukui Y. Effect of single or multiple injection of follicle stimulating hormone combined with pregnant mare serum gonadotropin on superovulatory response, and normal and freezable embryos in ewes. J Reprod Develop. (1996) 42:81–7. doi: 10.1262/jrd.42.81
55. Simonetti L, Forcada F, Rivera O, Carou N, Alberio R, Abecia J, et al. Simplified superovulatory treatments in Corriedale ewes. Anim Reprod Sci. (2008) 104:227–37. doi: 10.1016/j.anireprosci.2007.01.020
56. Panyaboriban S, Suwimonteerabutr J, Swangchan-Uthai T, Tharasanit T, Suthikrai W, Suadsong S, et al. A simplified superovulation protocol using split-single administration of Folltropin®-V in hyaluronan: application to purebred sheep. Vet Med (Praha). (2018) 63:321–8. doi: 10.17221/52/2016-VETMED
57. Leoni G, Bogliolo L, Pintus P, Ledda S, Naitana S. Sheep embryos derived from FSH/eCG treatment have a lower in vitro viability after vitrification than those derived from FSH treatment. Reprod Nutr Dev. (2001) 41:239–46. doi: 10.1051/rnd:2001127
58. Rahman M, Rahman M, Khadijah W, Abdullah R. Effect of supplementation of hCG or GnRH on ovulation and subsequent embryo production of eCG superovulated goats. Indian J Anim Res. (2017) 51:438–43. doi: 10.18805/ijar.11166
59. D'Alessandro A, Martemucci G. Efficiency of superovulatory treatment with FSH-p for in vivo embryo production in dairy ewes: multiple versus single dose regimen. J Anim Vet Adv. (2004) 3:388–93.
60. Loiola Filho JB, do Monte APO, dos Santos Souza TT, de Souza Miranda M, Magalhães Barros LC, Souza Costa Barros CH, et al. Effect of pFSH dose reduction on in vivo embryo production in Dorper ewes Semina. Ciências Agrárias. (2015) 36:4215–24. doi: 10.5433/1679-0359.2015v36n6Supl2p4215
61. Lehloenya KC, Greyling JP. The effect of embryo donor age and parity on the superovulatory response in Boer goat does. S Afr J Anim Sci. (2010) 88:38–43. doi: 10.1016/j.smallrumres.2009.11.007
62. Lopes EL, Maia EL, Paula NR, Teixeira DI, Villarroel AB, Rondina D, et al. Effect of age of donor on embryo production in Morada Nova (white variety) ewes participating in a conservation programme in Brazil. Trop Anim Health Pro. (2006) 38:555–61. doi: 10.1007/s11250-006-4344-1
63. Wu W, Yang M, Gong P, Wang F, Tian Y, Xu X, et al. Effect of two follicle stimulating hormone (FSH) preparations and simplified superovulatory treatments on superovulatory response in Xinji fine-wool sheep. Afr J Biotechnol. (2011) 10:15834–7. doi: 10.5897/AJB11.1927
64. Nuti LC, Minhas BS, Baker WC, Capehart JS, Marraek P. Superovulation and recovery of zygotes from Nubian and Alpine dairy goats. TheriogenoIogy. (1987) 28:481–8. doi: 10.1016/0093-691X(87)90252-4
65. Kiessling AA, Hughes WH, Blankevoort MR. Superovulation and embryo transfer in the dairy goat. J Am Vet Med Assoc. (1986) 188:829.
66. Mpebe NA, Gonzalez-Bulnes A, Lehloenya KC. Effect of breed and follicular status on response to superovulation in South African goats. J Appl Anim Res. (2018) 46:141–5. doi: 10.1080/09712119.2016.1277530
67. Boscos C, Vainas E, Kouskoura T, Samartzi F, Vafiadis D, Dellis S. Superovulatory response of Chios and Friesian ewes to two FSH-P dose levels. Reprod Domestic Anim. (1997) 32:195–8. doi: 10.1111/j.1439-0531.1997.tb01281.x
68. Senn BJ, Richardson ME. Seasonal effects on caprine response to synchronization of estrus and superovulatory treatment. Theriogenology. (1991) 37:579–85. doi: 10.1016/0093-691X(92)90138-H
69. Baril G, Vallet JC. Time of ovulation in dairy goats induced to superovulated with porcine follicle stimulating hormone and out ofthe breeding season. Theriogenology. (1990) 34:303–11. doi: 10.1016/0093-691X(90)90523-V
71. Yan L, Li H, Shi Z. Immunization against inhibin improves in vivo and in vitro embryo production. Anim Reprod Sci. (2015) 163:1–9. doi: 10.1016/j.anireprosci.2015.11.001
72. Medan M, Watanabe G, Sasaki K, Nagura Y, Sakaime H, Fujita M, et al. Effects of passive immunization of goats against inhibin on follicular development, hormone profile and ovulation rate. Reproduction. (2003) 125:751–7. doi: 10.1530/rep.0.1250751
73. Medan MS, Watanabe G, Sasaki K, Nagura Y, Sakaime H, Fujita M, et al. Ovarian and hormonal response of female goats to active immunization against inhibin. J Endocrinol. (2003) 177:287–94. doi: 10.1677/joe.0.1770287
74. Holtz W, Wang X, El-Gayar M, Knight PG. The effect of exogenous gonadotropins on ovarian function in goats actively immunized against inhibin. Theriogenology. (2012) 77:253–9. doi: 10.1016/j.theriogenology.2011.07.037
75. Fan BQ, Wang GJ, Yang LG, Ji XP, Wang HH, Cao BK, et al. Superovulation and embryo transf er in Boer goats. Jiangsu J of Agr Sci. (2000) 16:230–2.
76. Yuan Y, An L, Yu B, Yang T, Cheng Y. Effect of follicle stimulating hormone dosage, seasons and treatment frequency on superovulation in goats. Anim Husb Feed Sci. (2011) 3:20–2.
77. D'Alessandro A, Martemucci G, Iaffaldano N. Active immunization with asynthetic fragment of pig inhibin alpha-subunit increases ovulation rate and embryo production in superovulated ewes but season affects its efficiency. Reproduction. (1999) 115:185–91. doi: 10.1530/jrf.0.1150185
78. Li C, Zhu YL, Xue JH, Zhang SL, Ma Z, Shi ZD. Immunization against inhibin enhances both embryo quantity and quality in Holstein heifers after superovulation and insemination with sex-sorted semen. Theriogenology. (2009) 71:1011–7. doi: 10.1016/j.theriogenology.2008.11.003
79. Mei C, Li MY, Zhong SK, Lei Y, Shi ZD. Enhancing embryo yield by immunization against inhibin in superovulated Holstein heifers. Reprod Domest Anim. (2009) 44:735–9. doi: 10.1111/j.1439-0531.2008.01061.x
80. Takeo T, Nakagata N. Superovulation using the combined 576 administration of inhibin antiserum and equine chorionic gonadotropin increases the number of ovulated oocytes in C57BL/6 female mice. PLoS ONE. (2015) 10:e0128330. doi: 10.1371/journal.pone.0128330
81. Menchaca A, Vilariño M, Crispo M, Pinczak A, Rubianes E. Day 0 protocol: Superstimulatory treatment initiated in the absence of a large follicle improves ovarian response and embryo yield in goats. Theriogenology. (2007) 68:1111–7. doi: 10.1016/j.theriogenology.2007.07.020
82. Menchaca A, Miller V, Salveraglio V, Rubianes E. Endocrine, luteal and follicular responses after the use of the shortterm protocol to synchronize ovulation in goats. Anim Reprod Sci. (2007) 102:76–87. doi: 10.1016/j.anireprosci.2006.10.001
83. Rubianes E, Menchaca A. The pattern and manipulation of ovarian follicular growth in goats. Anim Reprod Sci. (2003) 78:271–87. doi: 10.1016/S0378-4320(03)00095-2
84. Gonzalez-Bulnes A, Carrizosa JA, Diaz-Delfa C, Garcia-Garcia RM, Rutia B, Santiago-Moreno J, et al. Effect of ovarian follicular status on superovulatory response of dairy goats to FSH treatment. Small Ruminant Res. (2003) 48:9–14. doi: 10.1016/S0921-4488(02)00294-8
85. Grizelj J, Špoljarić BT, Dobranić M, Lojkić FS, Dávila FS, SamardŽija M, et al. Efficiency analysis of standard and day 0 superovulatory protocols in Boer breed goats. Vet Arh. (2017) 87:473–86. doi: 10.24099/vet.arhiv.160317
86. Lehloenya K, Greyling J, Groble S. Can repeated superovulation and embryo recovery in Boer goats limit donor participation in a MOET programme? S. Afr J Anim Sci. (2009) 39:193–7. doi: 10.4314/sajas.v39i1.61216
87. Ustuner B, Alcay S, Nak Y, Nur Z, Nak D, Tuna B, et al. Repeated superovulation treatments in Kivircik ewes during breeding and nonbreeding seasons. Turk J Vet Anim Sci. (2014) 38:480–4. doi: 10.3906/vet-1401-20
88. Agarwal A, Said TM, Bedaiwy MA, Banerjee J, Alvarez JG. Oxidative stress in an assisted reproductive techniques setting. Fertil Steril. (2006) 86:503–12. doi: 10.1016/j.fertnstert.2006.02.088
89. Combelles CM, Gupta S, Agarwal A. Could oxidative stress influence the in-vitro maturation of oocytes? Reprod Biomed Online. (2009) 18:864–80. doi: 10.1016/S1472-6483(10)60038-7
90. Yuan Y, Liu R, Zhang X, Zhang J, Zheng Z, Huang C, et al. Effects of recipient oocyte source, number of transferred embryos and season on somatic cell nuclear transfer efficiency in sheep. Reprod Domest Anim. (2019) 54:1443–8. doi: 10.1111/rda.13546
91. Reggio BC, James AN, Green HL, Gavin WG, Behboodi E, Echelard Y, et al. Cloned transgenic offspring resulting from somatic cell nuclear transfer in the goat: oocytes derived from both follicle-stimulating hormone-stimulated and nonstimulated abattoir-derived ovaries. Biol Reprod. (2001) 65:1528–33. doi: 10.1095/biolreprod65.5.1528
92. Sugimura S, Kobayashi S, Hashiyada Y, Ohtake M, Kaneda M, Yamanouchi T, et al. Follicular growth-stimulated cows provide favorable oocytes for producing cloned embryos. Cell Reprogram. (2012) 14:29–37. doi: 10.1089/cell.2011.0060
93. Bondioli KR. Cloning of livestock by somatic cell nuclear transfer. Anim Biotechnol. (2018) 2:21–20. doi: 10.1007/978-3-319-92348-2_1
94. Hill JR, Roussel A, Cibelli J, Edwards J, Hooper N, Miller M, et al. Clinical and pathologic features of cloned transgenic calves and fetuses (13 case studies). Theriogenology. (1999) 51:1451–65. doi: 10.1016/S0093-691X(99)00089-8
95. Zakhartchenko V, Durcova-Hills G, Stojkovic M, Schernthaner W, Prelle K, Steinborn R, et al. Effects of serum starvation and re-cloning on the efficiency of nuclear transfer using bovine fetal fibroblasts. J Reprod. (1999) 115:325–31. doi: 10.1530/jrf.0.1150325
96. Hill JR, Winger QA, Long CR, Looney CR, Thompson JA, Westhusin ME. Development rates of male bovine nuclear transfer embryos derived from adult and fetal cells. Biol Reprod. (2000) 62:1135–40. doi: 10.1095/biolreprod62.5.1135
97. Keefer C, Baldassarre H, Keyston R, Wang B, Bhatia B, Bilodeau A, et al. Generation of dwarf goat (Capra hircus) clones following nuclear transfer with transfected and nontransfected fetal fibroblasts and in vitro-matured oocytes. Biol Reprod. (2001) 64:849–56. doi: 10.1095/biolreprod64.3.849
98. Martins LT, Neto SG, Tavares KCS, Calderón CEM, Aguiar LH, Lazzarotto CR, et al. Developmental outcome and related abnormalities in goats: comparison between somatic cell nuclear transfer-and in vivo-derived concepti during pregnancy through term. Cell Reprogram. (2016) 18:264–79. doi: 10.1089/cell.2015.0082
99. Peura TT, Kleemann DO, Rudiger SR, Nattrass GS, McLaughlan CJ, Walker SK. Effect of nutrition of oocyte donor on the outcomes of somatic cell nuclear transfer in the sheep. Biol Reprod. (2003) 68:45–50. doi: 10.1095/biolreprod.102.007039
100. Lee K, Prather RS. Advancements in somatic cell nuclear transfer and future perspectives. Anim Fron. (2013) 3:56–661. doi: 10.2527/af.2013-0034
101. Beaujean N, Taylor J, Gardner J, Wilmut I, Meehan R, Young L. Effect of limited DNA methylation reprogramming in the normal sheep embryo on somatic cell nuclear transfer. Biol Reprod. (2004) 71:185–93. doi: 10.1095/biolreprod.103.026559
102. Lee M, Ahn JI, Lee AR, Ko DW, Yang WS, Lee G, et al. Adverse effect of superovulation treatment on maturation, function and ultrastructural integrity of murine oocytes. Mol Cells. (2017) 40:558. doi: 10.14348/molcells.2017.0058
103. Ramos-Ibeas P, Heras S, Gómez-Redondo I, Planells B, Fernández-González R, Pericuesta E, et al. Embryo responses to stress induced by assisted reproductive technologies. Mol Reprod Dev. (2019) 86:1292–306. doi: 10.1002/mrd.23119
104. Ertzeid G, Storeng R. The impact of ovarian stimulation on implantation and fetal development in mice. Hum Reprod. (2001) 16:221–5. doi: 10.1093/humrep/16.2.221
105. Looney C, Pryor J. Practical applications of new research information in the practice of bovine embryo transfer. Reprod Fert Dev. (2009) 22:145–50. doi: 10.1071/RD09223
106. Sirard M-A, Richard F, Blondin P, Robert C. Contribution of the oocyte to embryo quality. Theriogenology. (2006) 126–36. doi: 10.1016/j.theriogenology.2005.09.020
107. Greve T, Callesen H. Rendez-vous in the oviduct: implications for superovulation and embryo transfer. Reprod Nutr Dev. (2001) 41:451–9. doi: 10.1051/rnd:2001144
108. Xiao P, Nie J, Wang X, Lu K, Lu S, Liang X. Melatonin alleviates the deterioration of oocytes from mice subjected to repeated superovulation. J Cell Physiol. (2019) 234:13413–22. doi: 10.1002/jcp.28018
109. Hansen PJ. Implications of assisted reproductive technologies for pregnancy outcomes in mammals. Ann Rev Anim Biosci. (2020) 8:395–413. doi: 10.1146/annurev-animal-021419-084010
110. Desantis S, Albrizio M, Lacitignola L, Laricchiuta P, Cinone M. Modification of Morphology and Glycan Pattern of the Oviductal Epithelium of Baboon Papio hamadryas during the menstrual cycle. Animals. (2022) 12:2769. doi: 10.3390/ani12202769
111. Małysz-Cymborska I, Andronowska A. Expression of the vascular endothelial growth factor receptor system in porcine oviducts after induction of ovulation and superovulation. Domestic Anim Endocrinol. (2014) 49:86–95. doi: 10.1016/j.domaniend.2014.06.003
112. Taiyeb AM, Muhsen-Alanssari SA, Dees W, Ridha-Albarzanchi MT, Kraemer DC. Improvement in in vitro fertilization outcome following in vivo synchronization of oocyte maturation in mice. Exp Biol Med. (2015) 240:519–26. doi: 10.1177/1535370214549533
113. Van der Auwera I, D'Hooghe T. Superovulation of female mice delays embryonic and fetal development. Hum Reprod. (2001) 16:1237–43. doi: 10.1093/humrep/16.6.1237
114. Dias F, Dadarwal D, Adams G, Mrigank H, Mapletoft R, Singh J. Length of the follicular growing phase and oocyte competence in beef heifers. Theriogenology. (2013) 79:1177–83. e1. doi: 10.1016/j.theriogenology.2013.02.016
115. Gad A, Besenfelder U, Rings F, Ghanem N, Salilew-Wondim D, Hossain M, et al. Effect of reproductive tract environment following controlled ovarian hyperstimulation treatment on embryo development and global transcriptome profile of blastocysts: implications for animal breeding and human assisted reproduction. Hum Reprod. (2011) 26:1693–707. doi: 10.1093/humrep/der110
116. Murray MK, Desouza MM, Messinger SM. Oviduct during early pregnancy: hormonal regulation and interactions with the fertilized ovum. Micro Res Tech. (1995) 31:497–506. doi: 10.1002/jemt.1070310606
117. Mainigi MA, Olalere D, Burd I, Sapienza C, Bartolomei M, Coutifaris C. Peri-implantation hormonal milieu: elucidating mechanisms of abnormal placentation and fetal growth. Biol Reprod. (2014) 90:26; 1–9. doi: 10.1095/biolreprod.113.110411
118. Angel M, Gil M, Cuello C, Sanchez-Osorio J, Gomis J, Parrilla I, et al. The effects of superovulation of donor sows on ovarian response and embryo development after nonsurgical deep-uterine embryo transfer. Theriogenology. (2014) 81:832–9. doi: 10.1016/j.theriogenology.2013.12.017
119. Marshall KL, Rivera RM. The effects of superovulation and reproductive aging on the epigenome of the oocyte and embryo. Mol Reprod Dev. (2018) 85:90–105. doi: 10.1002/mrd.22951
120. Förstl H, Almeida OP, Owen AM, Burns A, Howard R. Psychiatric, neurological and medical aspects of misidentification syndromes: a review of 260 cases. Psychol Med. (1991) 21:905–10. doi: 10.1017/S0033291700029895
Keywords: strategies, factors, superovulation, hormonal protocols, goat
Citation: Khan S, Jamal MA, Khan IM, Ullah I, Jabbar A, Khan NM and Liu Y (2023) Factors affecting superovulation induction in goats (Capra hericus): An analysis of various approaches. Front. Vet. Sci. 10:1152103. doi: 10.3389/fvets.2023.1152103
Received: 27 January 2023; Accepted: 24 February 2023;
Published: 24 March 2023.
Edited by:
Ayman Abdel-Aziz Swelum, Zagazig University, EgyptReviewed by:
Shanshan Yang, Hebei Normal University of Science and Technology, ChinaCopyright © 2023 Khan, Jamal, Khan, Ullah, Jabbar, Khan and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ibrar Muhammad Khan, aWJyYXIucGVzaEBnbWFpbC5jb20=; Yong Liu, bGl1eW9uZ0BmeW51LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.