
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 27 July 2022
Sec. Livestock Genomics
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.951168
This article is part of the Research Topic Genetic Regulation of Meat Quality Traits in Livestock Species View all 10 articles
 Xiaofan Tan1Yu He1Yanchun Qin1Zhiwei Yan1Jing Chen1Ruixue Zhao1Shenglan Zhou1
Xiaofan Tan1Yu He1Yanchun Qin1Zhiwei Yan1Jing Chen1Ruixue Zhao1Shenglan Zhou1 David M. Irwin2
David M. Irwin2 Bojiang Li1*
Bojiang Li1* Shuyi Zhang1*
Shuyi Zhang1*Intramuscular fat (IMF) is an important regulator that determines meat quality, and its content is closely related to flavor, tenderness, and juiciness. Many studies have used quantitative proteomic analysis to identify proteins associated with meat quality traits in livestock, however, the potential candidate proteins that influence IMF in donkey muscle are not fully understood. In this study, we performed quantitative proteomic analysis, with tandem-mass-tagged (TMT) labeling, with samples from the longissimus dorsi (LD) muscle of the donkey. A total of 585,555 spectra were identified from the six muscle samples used in this study. In total, 20,583 peptides were detected, including 15,279 unique peptides, and 2,540 proteins were identified. We analyzed differentially abundant proteins (DAPs) between LD muscles of donkeys with high (H) and low (L) IMF content. We identified 30 DAPs between the H and L IMF content groups, of which 17 were upregulated and 13 downregulated in the H IMF group. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) functional enrichment analysis of these DAPs revealed many GO terms (e.g., bone morphogenetic protein (BMP) receptor binding) and pathways (e.g., Wnt signaling pathway and Hippo signaling pathway) involved in lipid metabolism and adipogenesis. The construction of protein–protein interaction networks identified 16 DAPs involved in these networks. Our data provide a basis for future investigations into candidate proteins involved in IMF deposition and potential new approaches to improve meat quality in the donkey.
Donkeys are domesticated animals that belong to the horse family (1). They are one of the most important livestock animals in many countries in Africa and the Middle East, which are mainly used for farming or other work on large farms (2, 3). With the increase of interest in donkey breeding, it is increasingly being used as a milk and meat-producing animal (4, 5). Donkey milk has been shown to be the best substitute for human milk for children who are allergic to milk proteins (6). Donkey meat is characterized by high-quality protein, vitamins, and minerals, which is the preferred meat for many consumers due to its high protein content (5, 7, 8). According to a previous study, 100 g of donkey meat contains 22.8 g of protein and 2.02 g of fat (2, 9, 10). With improvements in living standards, consumers are becoming more concerned about the quality of livestock meat. In recent years, donkey meat consumption has increased in many countries, including China and Italy, and has undoubtedly become one of the livestock meat choices (6, 11, 12).
Intramuscular fat (IMF), also referred to as marbling, is the amount of fat that accumulates between muscle fibers or inside muscle cells (13). Its main components are phospholipids and triglycerides (14, 15). A previous study reported that IMF content is mainly determined by the number and size of intramuscular adipocytes (16). Furthermore, IMF is a complex quantitative trait, which is influenced by a variety of regulatory factors, such as gene regulation (17), sex (18), age, or body weight (19), as well as environmental conditions, cell signals, and diet (20). A previous study has shown that IMF plays a key role in many meat quality characteristics and quality (21). For example, IMF can improve meat quality by improving flavor, juiciness, and tenderness (22, 23). Therefore, IMF not only plays a very important role in animal husbandry production but also is closely related to a healthy and desirous human food supply. In recent years, the identification of candidate genes for IMF to improve meat quality has become an important research topic in livestock breeding. A large number of studies have investigated candidate genes affecting IMF content in many species, including cattle (24), pigs (25), sheep (26), and goats (27). Genes, such as PHKG1 (28), MYH3 (20), and PLIN1 (29), have been identified as candidates for regulating IMF content in pigs. However, candidate genes and regulatory mechanisms for IMF content in donkeys are not fully resolved.
With recent developments in proteomic technologies, it has become an increasingly important approach to identify candidate proteins related to meat quality in livestock. A previous study identified 127 proteins with differential abundance associated with IMF in the longissimus dorsi (LD) muscle of pigs between days 120 d and 240 of growth (30). Hou et al. (31) identified proteins from the pig related to postmortem meat quality using a TMT (tandem-mass-tagged)-labeled quantitative proteomic. Similarly, proteomics was used to detect proteins associated with meat quality traits in other species including sheep (32), cattle (33), and chickens (34). Current proteomic studies on the donkey have mainly focused on the identification of proteins associated with donkey milk (35, 36). Few studies on proteins related to the regulation of donkey meat quality have been reported.
In this study, expression profiling of proteins was performed using LD muscle samples with divergent IMF content phenotypes (H and L IMF content groups). We used bioinformatic methods to identify the differentially abundant proteins (DAPs) between the H and L IMF groups. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of the DAPs was conducted. The aim of this study was to identify candidate proteins that influence IMF content in donkeys and to provide a basis for improving the quality of donkey meat.
All animal procedures described in this study were conducted according to the animal husbandry guidelines of the Shenyang Agricultural University. The studies on these animals were reviewed and approved by the Ethics Committee and Experimental Animal Committee of Shenyang Agriculture University (No. 202006032).
All animals used in this study were derived from a population of 30 donkeys described in a previous study (37). These animals were raised under the same environmental conditions. At about 15 months of age, all donkeys are slaughtered in the same abattoir and LD muscle tissue samples were collected for IMF determination and protein extraction. Samples for protein extraction were immediately frozen in liquid nitrogen and then stored at−80°C until use. The Soxhlet extraction method (37) was used to determine the IMF content of the samples.
Protein in the samples was isolated after disruption of the tissue in SDT [4% sodium dodecyl sulfate (SDS), 100 mM Tris-HCl,1 mM DTT, pH 7.6] buffer, with the protein concentration quantified with the bicinchoninic acid (BCA) Protein Assay Kit (Bio-Rad, USA). Protein digestion was performed using trypsin according to the filter-aided sample preparation (FASP) method previously described by Matthias Mann (38). Briefly, protein from each sample was incorporated into SDT [4% SDS, 100 mM dithiothreitol (DTT), 150 mM Tris-HCl pH 8.0] buffer. The detergent, DTT, and other low-molecular-weight components were removed using UA buffer (8 M urea, 150 mM Tris-HCl pH 8.0). Iodoacetamide (IAA), 100 μl of 100 mM IAA in UA buffer, was added to block reduced cysteine residues and incubated in the dark for 30 min. The filter was then washed three times with 100 μl UA buffer and then twice with 100 μl 25 mM NH4HCO3 buffer. The eluted protein suspension was then digested overnight at 37°C with 4 μg trypsin (Promega) in 40 μl of 25 mM NH4HCO3 buffer and the resulting peptides were collected as filtrate. Digested peptides for each sample were desalted on C18 cartridges, concentrated by vacuum centrifugation, and reconstituted in 40 μl of 0.1% (v/v) formic acid.
Peptides from the three high (H1, H2, and H3) and three low (L1, L2, and L3) IMF content donkey samples were labeled with 126, 127, 128, 129, 130, and 131 isotope tags, respectively, using TMT reagents according to the manufacturer's instructions (Thermo Scientific, Waltham, USA).
Liquid chromatography–mass spectrometry (LC–MS/MS) analysis was conducted on a Q Exactive mass spectrometer (Thermo Scientific, Waltham, USA). MS data were obtained using a data-dependent top 10 method dynamically choosing the most abundant precursor ions from the survey scan (300–1,800 m/z) for high-energy collisional dissociation (HCD) fragmentation. The automatic gain control (AGC) target was 3 × 106 and the maximum injection time was 10 ms. The dynamic exclusion duration was 40.0 s. Survey scans were obtained at a resolution of 70,000 at m/z 200 and resolution for HCD spectra was set to 17,500 at m/z 200, and isolation width was 2 m/z. The normalized collision energy was 30 eV and the underfill ratio was 0.1%.
The MS raw data for each sample were searched against the donkey UniProt database using the MASCOT engine (Matrix Science, London, UK; version 2.2) embedded into Proteome Discoverer 1.4 software for protein identification and quantitation. Proteins with fold change (FC) >1.2 or <0.833 and p < 0.05 were considered to be significantly DAPs.
The gene ontology (GO) term annotation of selected DAPs was performed using Blast2GO software (39, 40). The DAPs were blasted against the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (http://geneontology.org/) to retrieve their KEGG orthology identifications, which were subsequently mapped to the pathways in KEGG (41). GO and KEGG enrichment was analyzed using the Fisher's exact test. GO and KEGG terms with p < 0.05 were considered significantly enriched.
For protein–protein interactions, we used STRING v11.5 (42) to predict and visualize networks according to default parameters. Protein interactions were illustrated using Cytoscape software (43).
We selected three LD muscle samples with high (H1, H2, and H3) and three with low (L1, L2, and L3) IMF content from a population of 30 individuals. Statistical analysis of the data showed that IMF content was significantly different between H and L samples (3.04 ± 0.12%, 6.39 ± 0.47%; p < 0.01). We further characterized the protein abundance profiles of the six samples using the TMT-labeled proteomic approach. The entire proteomic experimental flow for this study is shown in Figure 1. A total of 585,555 spectra were detected from the six LD muscle samples by LC–MS/MS analysis (Figure 2A). From this, we identified a total of 20,583 peptides, of which 15,279 were unique, corresponding to 2,540 distinct proteins (Figure 2A; Supplementary Table S1). A statistical analysis of the lengths of the identified peptide was conducted, which showed that the peptides were mainly between 5 and 15 amino acids with peptides of 7 and 9 amino acids in length being the highest (Figure 2B). Further analyses of the numbers of peptides identified in the proteins showed that about 50% of the proteins contained 1–3 identified peptides (Figure 2C). The molecular weights of the identified proteins indicated that most of these proteins have molecular weights between 10 and 70 kDa (Figure 2D).
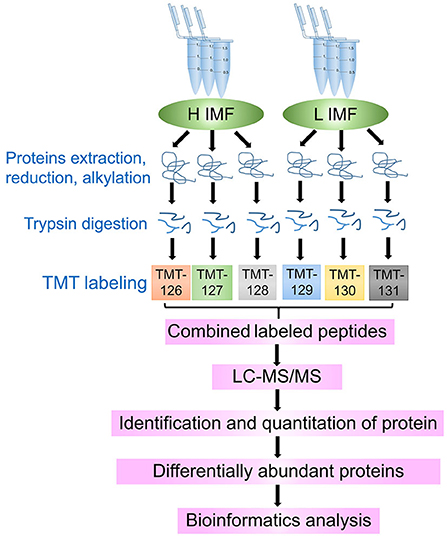
Figure 1. Schematic protocol for the identification of longissimus dorsi muscle proteins from tissue samples with high (H) and low (L) intramuscular fat (IMF) content using tandem-mass-tagged (TMT)-labeled quantitative proteomics. The H and L IMF samples were in triplicate, and each sample used a different TMT label.
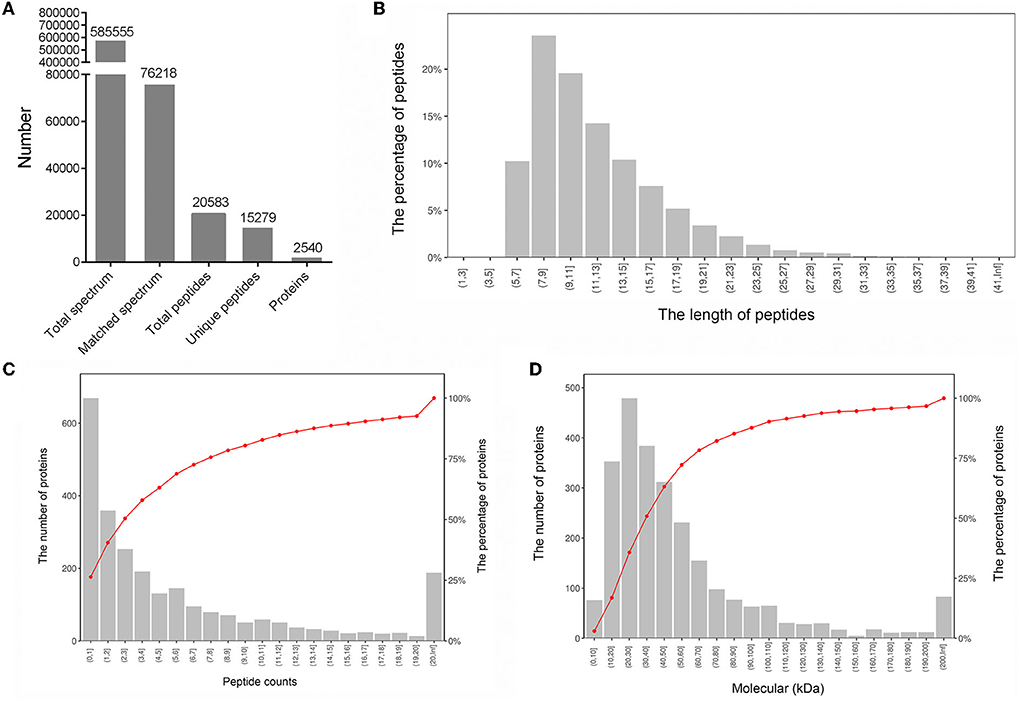
Figure 2. Characterization of proteins from six longissimus dorsi muscle samples. (A) Overview information of identified proteins in this study. (B) Distribution of the lengths of the identified peptides. (C) Distribution of the numbers of identified proteins containing different numbers of peptides. (D) Distribution of the molecular weights of the identified proteins.
To further explore candidate proteins associated with IMF deposition in donkeys, we analyzed the differentially abundant proteins in the high and low IMF content groups. A total of 30 DAPs were identified between the H and L IMF groups, with 13 upregulated and 17 downregulated in the H IMF group (Figure 3A). Summary information on these proteins is listed in Supplementary Table S2. The top upregulated DAP is MAP4K4, while the top downregulated DAP is Rho-associated protein kinase 2 (ROCK2) (Supplementary Table S2). We also examined the abundance patterns of these DAPs in the six samples, which showed clear abundance differences between the high and low IMF group samples (Figure 3B).
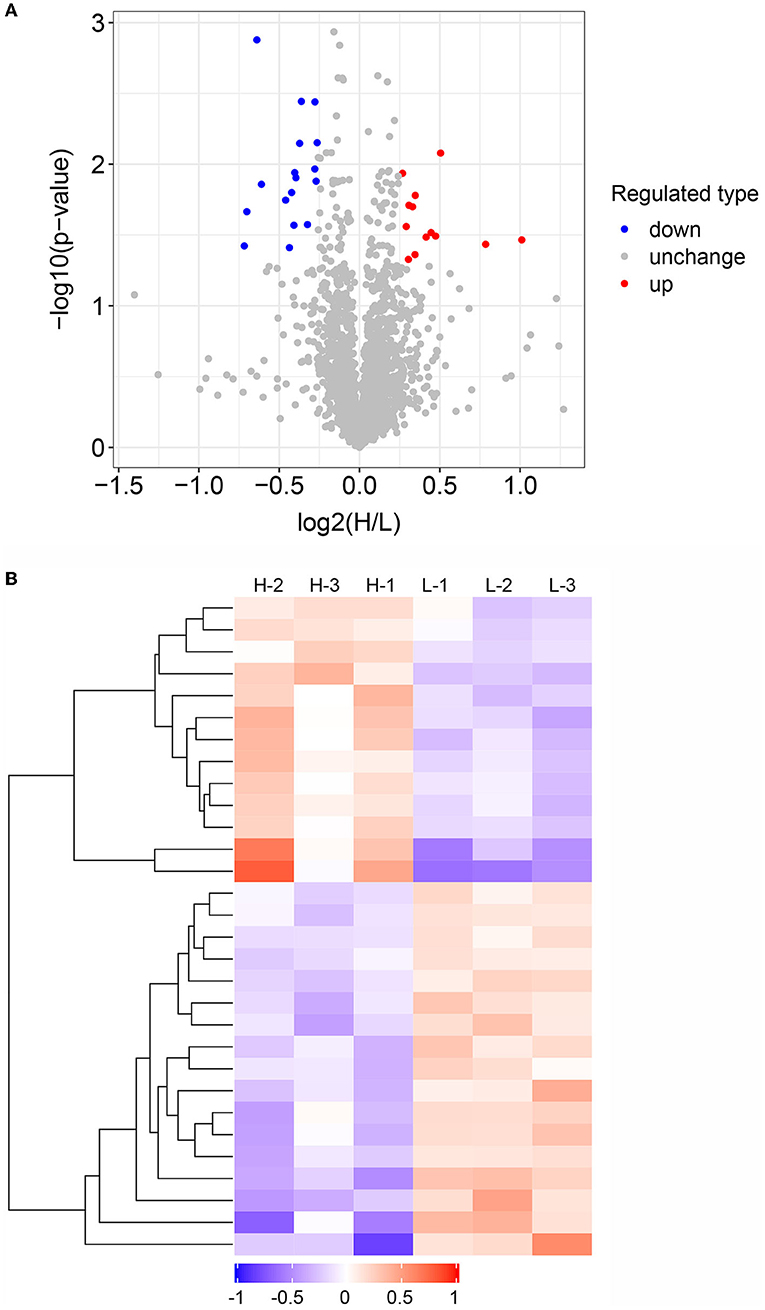
Figure 3. Analysis of differentially abundant proteins (DAPs) between high and low intramuscular fat (IMF) groups. (A) Volcano plot showing DAPs between high and low IMF groups. Red, gray, and blue dots represent upregulated, unchanged, and downregulated proteins, respectively. (B) Heatmap showing the abundance patterns of the DAPs between the three high and three low IMF samples.
To explore the potential biological functions of the DAPs in IMF deposition, we performed GO and KEGG functional enrichment analyses of the DAPs. Here, 227, 27, and 24 GO terms were found to be significantly enriched in biological process, cellular component, and molecular function, respectively (Supplementary Table S3). The top 30 significantly enriched GO terms are shown in Figure 4A. This data suggests that the DAPs are primarily involved in bone morphogenetic protein (BMP) receptor binding, which is associated with lipid metabolism. The KEGG enrichment analysis suggested that the DAPs are significantly enriched in 19 pathways (Figure 4B). Interestingly, some of these KEGG participate in the lipogenic processes, for instance, the Wnt signaling and Hippo signaling pathways. Further, a DAP, ROCK2, is significantly enriched in the Wnt signaling pathway. These results suggest that the enrichment of DAPs in these pathways may be closely related to the lipogenic process.
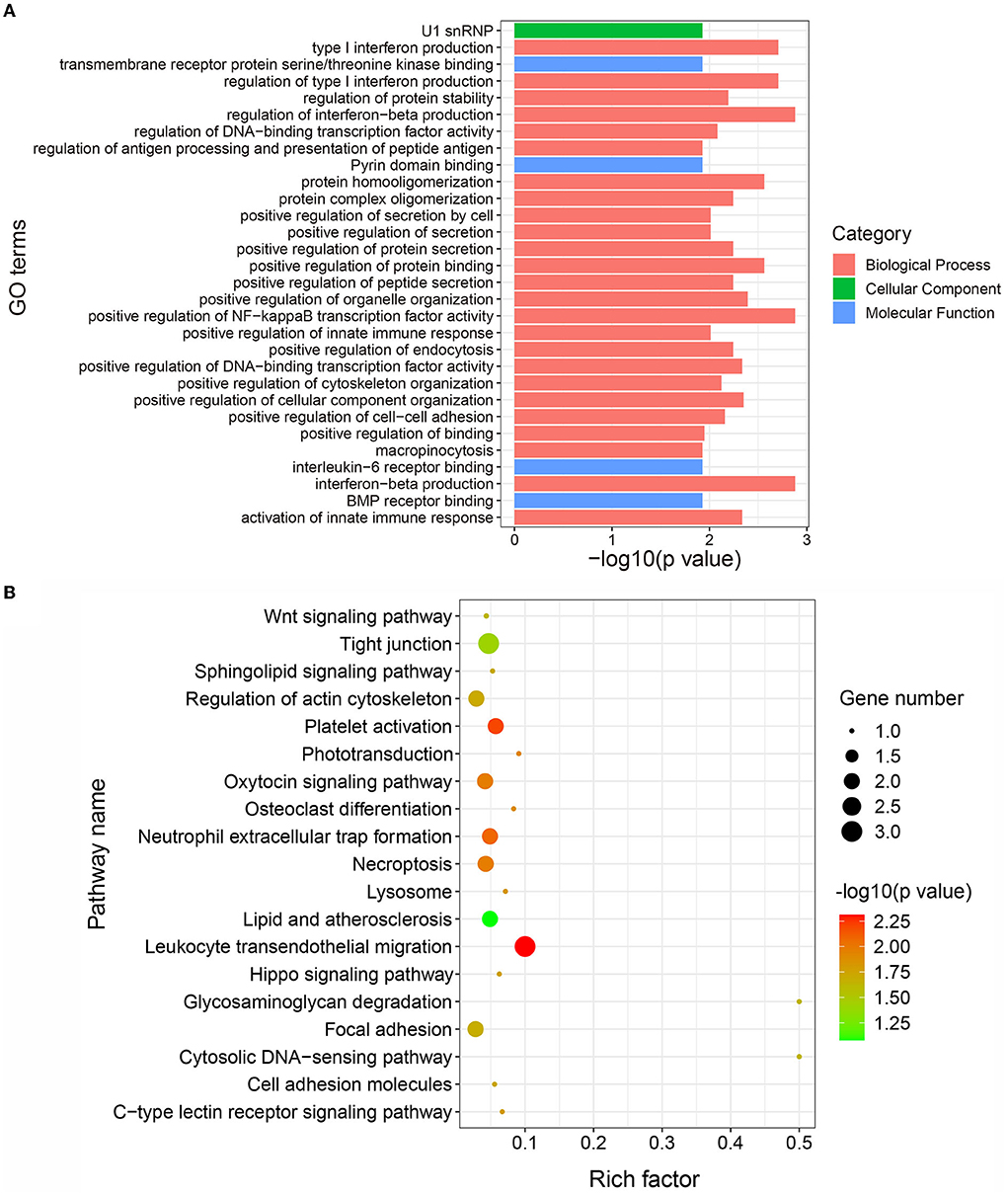
Figure 4. Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) functional enrichment analysis of differentially abundant proteins (DAPs). (A) Top 30 significantly enriched GO terms. The X-axis represents the –log10 (p value) value of the GO terms and the Y-axis represents the GO term name. (B) Significantly enriched pathways of DAPs. The X-axis represents the rich factor and the Y-axis represents the pathway. The size and color of the bubbles represent the number of proteins enriched in the pathway and enrichment significance, respectively.
Next, we examined protein–protein interactions of the 30 DAPs to better understand IMF deposition. A protein–protein interaction analysis of the DAPs was conducted based on the STRING database, which contains details functional relationships between proteins, thus allowing predictions on the functional impact of changing protein abundance (44). The results of this analysis showed that 16 DAPs are involved in a protein–protein interaction network (Figure 5). For example, a DAP, ROCK2, interacts with UBR4, TJP1, MBP, and PTEN. In addition, the ADGRV1 and USH2A proteins interact with each other.
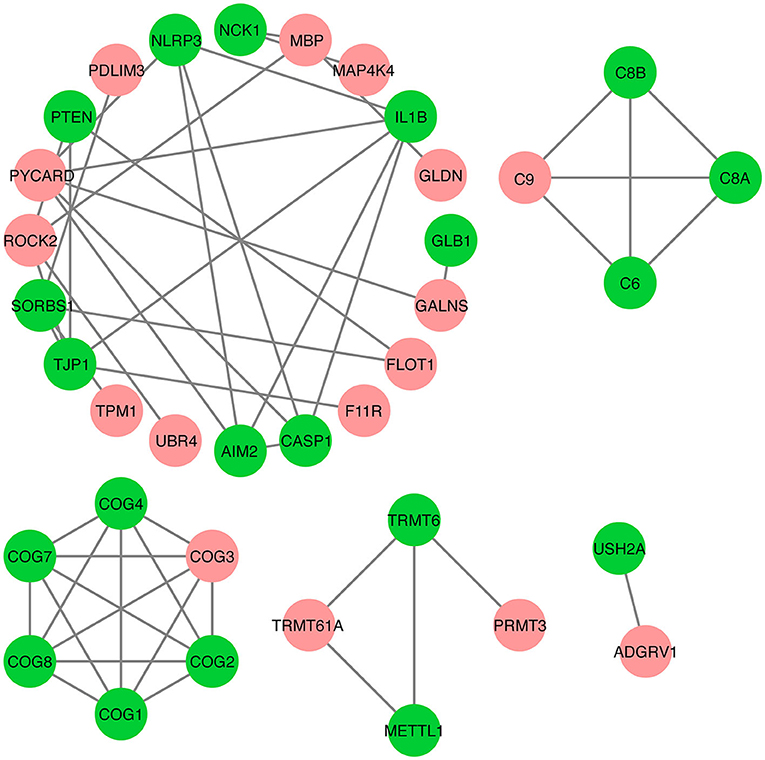
Figure 5. Protein–protein interactions analysis for differentially abundant proteins (DAPs). The red nodes represent DAPs, while the green nodes represent other proteins that interact with DAPs. The edge represents interactions between proteins.
In recent years, consumers have become increasingly interested in donkey meat, with China becoming the world's largest consumer of this meat (45). IMF plays a vital role in the quality of livestock meat, therefore, the identification of candidate proteins affecting IMF is essential for improving meat quality. Previous studies have applied proteomics to identify DAPs of IMF in other species including pigs (30) and goats (46). However, there is a paucity of research on candidate proteins for quality traits in donkey meat. In this study, we identified 30 proteins that were differentially abundant between LD muscle samples with high and low IMF content based on proteomic techniques. Our results suggest that the identified DAPs are candidates for influencing IMF content in donkey meat.
Further, of these DAPs, many are associated with the adipogenic processes. For example, ROCK2, a downregulated protein, inhibits adipogenesis based on knockdown experiments (47). Protein interaction analysis revealed that ROCK2 potentially binds UBR4, TJP1, MBP, and PTEN proteins. These results suggest that the ROCK2 protein might act as a suppressor of IMF deposition in the donkey by binding these proteins. Ahbara et al. (48) revealed the SPAG8 gene is a candidate for growth traits and adipogenesis in sheep. A previous study identified the RPL27A gene as a candidate gene for bovine marbling, with an single nucleotide polymorphism (SNP) in its promoter used as a molecular marker for bovine marbling (49). Tu et al. (50) found that a high-fat diet increases the expression level of TPM1. Another interesting finding is that PRMT3 acts as a co-transcription factor by translocating into the nucleus and binding to LXRα to regulate downstream gene expression, thereby promoting lipogenesis (51). These proteins (i.e., SPAG8, RPL27A, TPM1, MBP, and PRMT3) were identified as DAPs between the H and L IMF groups in this study, suggesting that they are important regulators of IMF. However, the mechanism of action of these DAPs in IMF deposition in donkeys needs to be further demonstrated.
Our study identified a number of GO terms related to the lipogenic process based on enrichment analysis of the DAPs, including tRNA methylation and BMP receptor binding. Zhao et al. (52) showed that RNA methylation has a crucial role and is required for adipose differentiation. In addition, it has been shown that tRNA methylation levels regulate the translation initiation of genes and thus affect protein synthesis (53). These data suggest that some DAPs may regulate IMF deposition by affecting tRNA methylation. BMP signaling is essential for the differentiation of mesenchymal stem cells (MSCs) into the adipocyte lineage (54). Jung et al. (55) found that plasma BMP2 levels were positively correlated with IMF content in steers, suggesting that BMP signaling contributes to meat quality. In addition, a GO term, positive regulation of c-Jun N-terminal kinase (JNK) cascade, was significantly enriched in this study. A previous study demonstrated that JNK signaling inhibits adipose differentiation by suppressing peroxisome proliferator-activated receptor-gamma (PPARγ) and fatty acid binding protein 4 (AP2) expression (56). Taken together, these DAPs may regulate the deposition of IMF through an association with adipogenesis.
In addition, DAPs were enriched in many key signaling pathways (e.g., Wnt and Hippo signaling pathways) involved in the adipogenic process. The Wnt signaling pathway is a highly conserved critical factor in animals that negatively regulates adipose differentiation (57). Mechanistically, Wnt signaling inhibits adipogenesis by suppressing the expression of PPARγ and CCAAT/enhancer-binding protein alpha (C/EBP)α (58). Activation of Wnt signaling, by transgenic overexpression of Wnt10, in mice leads to significantly reduced adipose tissue weight (59). A downregulated protein identified in our study, ROCK2, is significantly enriched in the Wnt signaling pathway, suggesting that it inhibits IMF deposition by mediating the Wnt signaling pathway. The Hippo signaling pathway was also enriched, and it has been shown to be involved in adipocyte proliferation and differentiation in animals (60, 61). In Hippo signaling pathway, serine/threonine kinase 24 (Ste20) family kinases mammalian STE20-like kinase (MST1/2) activate large tumor suppressor (LATS) kinases through phosphorylation, which in turn allows phosphorylation of yes-associated protein (YAP) protein and facilitates the binding of YAP to 14-3-3 proteins (62). Park et al. (63) found that MST1/2 promotes the differentiation of 3T3-L1 cells through the activation of PPARγ. Deng et al. (61) demonstrated that YAP1 inhibits the differentiation of ovine adipocytes by affecting PPARG and RXR alpha levels. These results suggest that the Hippo signaling pathway is implicated in the deposition of IMF in the donkey. However, how DAPs involved in Hippo signaling regulate the deposition of IMF in donkeys still needs to be further explored.
In summary, we identified 30 candidate proteins that might affect IMF content in the LD muscle of donkeys. We provide evidence that some of the DAPs affect IMF deposition as they are involved in adipogenic functions or signaling pathways in other animals. Our data provide evidence for the role of these proteins in IMF content in the donkey and provide new insights into the molecular mechanisms of the regulation of IMF deposition.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author/s.
The animal study was reviewed and approved by the Ethics Committee and Experimental Animal Committee of Shenyang Agriculture University. Written informed consent was obtained from the owners for the participation of their animals in this study.
BL and SZha conceived and designed this study. YH, YQ, and XT performed the experiments. ZY, JC, SZho, and RZ analyzed the data. XT and BL drafted the manuscript. DI and BL revised the manuscript.
This research was supported by grants from the National Natural Science Foundation of China (No. 32002147), the China Postdoctoral Science Foundation (No. 2021MD703855), the Educational Department of Liaoning Province (No. LJKZ0671), the Natural Science Foundation of Liaoning Province (No. 2021-BS-140), the Science and Technology Plan Project of Shenyang City (Nos. 21-116-3-40 and 21-110-3-10), the Organization Department of Liaoning Provincial Committee of China (No. XLYC1907018), and the Shenyang Agricultural University Research Start-up Funding (No. 880418062).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.951168/full#supplementary-material
2. Shi T, Hu W, Hou H, Zhao Z, Shang M, Zhang L. Identification and comparative analysis of long non-coding RNA in the skeletal muscle of two Dezhou Donkey strains. Genes (Basel). (2020) 11:508. doi: 10.3390/genes11050508
3. Polidori P, Vincenzetti S, Cavallucci C, Beghelli D. Quality of donkey meat and carcass characteristics. Meat Sci. (2008) 80:1222–4. doi: 10.1016/j.meatsci.2008.05.027
4. De Palo P, Maggiolino A, Milella P, Centoducati N, Papaleo A, Tateo A. Artificial suckling in Martina Franca donkey foals: effect on in vivo performances and carcass composition. Trop Anim Health Prod. (2016) 48:167–73. doi: 10.1007/s11250-015-0940-2
5. De Palo P, Tateo A, Maggiolino A, Marino R, Ceci E, Nisi A, et al. Martina Franca donkey meat quality: influence of slaughter age and suckling technique. Meat Sci. (2017) 134:128–34. doi: 10.1016/j.meatsci.2017.07.025
6. Polidori P, Pucciarelli S, Ariani A, Polzonetti V, Vincenzetti S A. comparison of the carcass and meat quality of Martina Franca donkey foals aged 8 or 12 months. Meat Sci. (2015) 106:6–10. doi: 10.1016/j.meatsci.2015.03.018
7. Trinchese G, Cavaliere G, De Filippo C, Aceto S, Prisco M, Chun JT, et al. Human milk and donkey milk, compared to cow milk, reduce inflammatory mediators and modulate glucose and lipid metabolism, acting on mitochondrial function and oleylethanolamide levels in rat skeletal muscle. Front Physiol. (2018) 9:32. doi: 10.3389/fphys.2018.00032
8. Zhang X, Li H, Yu J, Zhou X, Ji C, Wu S, et al. Label-free based comparative proteomic analysis of whey proteins between different milk yields of Dezhou donkey. Biochem Biophys Res Commun. (2019) 508:237–42. doi: 10.1016/j.bbrc.2018.11.130
9. Lanza M, Landi C, Scerra M, Galofaro V, Pennisi P. Meat quality and intramuscular fatty acid composition of Sanfratellano and Haflinger foals. Meat Sci. (2009) 81:142–7. doi: 10.1016/j.meatsci.2008.07.008
10. Lorenzo JM, Sarries MV, Tateo A, Polidori P, Franco D, Lanza M. Carcass characteristics, meat quality and nutritional value of horsemeat: a review. Meat Sci. (2014) 96:1478–88. doi: 10.1016/j.meatsci.2013.12.006
11. Seyiti S, Kelimu A. Donkey industry in China: current aspects, suggestions and future challenges. J Equine Vet Sci. (2021) 102:103642. doi: 10.1016/j.jevs.2021.103642
12. Polidori P, Vincenzetti S, Pucciarelli S, Polzonetti V. Comparison of carcass and meat quality obtained from mule and donkey. Animals-Basel. (2020) 10:1620. doi: 10.3390/ani10091620
13. Listrat A, Lebret B, Louveau I, Astruc T, Bonnet M, Lefaucheur L, et al. How muscle structure and composition influence meat and flesh quality. ScientificWorldJournal. (2016) 2016:3182746. doi: 10.1155/2016/3182746
14. da Costa AS, Pires VM, Fontes CM, Mestre Prates JA. Expression of genes controlling fat deposition in two genetically diverse beef cattle breeds fed high or low silage diets. BMC Vet Res. (2013) 9:118. doi: 10.1186/1746-6148-9-118
15. Silva DBD, Fonseca LFS, Pinheiro DG, Muniz MMM, Magalhaes AFB, Baldi F, et al. Prediction of hub genes associated with intramuscular fat content in Nelore cattle. BMC Genom. (2019) 20:1–12. doi: 10.1186/s12864-019-5904-x
16. Li X, Fu X, Yang G, Du M. Review: Enhancing intramuscular fat development via targeting fibro-adipogenic progenitor cells in meat animals. Animal. (2020) 14:312–21. doi: 10.1017/S175173111900209X
17. Plastow GS, Carrion D, Gil M, Garcia-Regueiro JA, Gispert M, Oliver MA, et al. Quality pork genes and meat production. Meat Sci. (2005) 70:409–21. doi: 10.1016/j.meatsci.2004.06.025
18. Skrlep M, Batorek N, Bonneau M, Prevolnik M, Kubale V, Candek-Potokar M. Effect of immunocastration in group-housed commercial fattening pigs on reproductive organs, malodorous compounds, carcass and meat quality. Czech J Anim Sci. (2012) 57:290–9. doi: 10.17221/5964-CJAS
19. Tyra M, Ropka-Molik K, Terman A, Piorkowska K, Oczkowicz M, Bereta A. Association between subcutaneous and intramuscular fat content in porcine ham and loin depending on age, breed and FABP3 and LEPR genes transcript abundance. Mol Biol Rep. (2013) 40:2301–8. doi: 10.1007/s11033-012-2311-7
20. Cho IC, Park HB, Ahn JS, Han SH, Lee JB, Lim HT, et al. A functional regulatory variant of MYH3 influences muscle fiber-type composition and intramuscular fat content in pigs. PLoS Genet. (2019) 15:e1008279. doi: 10.1371/journal.pgen.1008279
21. Gao SZ, Zhao SM. Physiology, affecting factors and strategies for control of pig meat intramuscular fat. Recent Pat Food Nutr Agric. (2009) 1:59–74. doi: 10.2174/2212798410901010059
22. Hausman GJ, Dodson MV, Ajuwon K, Azain M, Barnes KM, Guan LL, et al. Board-invited review: the biology and regulation of preadipocytes and adipocytes in meat animals. J Anim Sci. (2009) 87:1218–46. doi: 10.2527/jas.2008-1427
23. Li QQ, Huang ZY, Zhao WJ, Li MX, Li CC. Transcriptome analysis reveals long intergenic non-coding RNAs contributed to intramuscular fat content differences between Yorkshire and Wei pigs. Int J Mol Sci. (2020) 21:1732. doi: 10.3390/ijms21051732
24. Ochsner KP, MacNeil MD, Lewis RM, Spangler ML. Economic selection index development for Beefmaster cattle I: terminal breeding objective. J Anim Sci. (2017) 95:1063–70. doi: 10.2527/jas.2016.1231
25. Gao Y, Zhang R, Hu X, Li N. Application of genomic technologies to the improvement of meat quality of farm animals. Meat Sci. (2007) 77:36–45. doi: 10.1016/j.meatsci.2007.03.026
26. Mortimer SI, van der Werf JHJ, Jacob RH, Hopkins DL, Pannier L, Pearce KL, et al. Genetic parameters for meat quality traits of Australian lamb meat. Meat Sci. (2014) 96:1016–24. doi: 10.1016/j.meatsci.2013.09.007
27. Pena F, Juarez M, Bonvillani A, Garcia P, Polvillo O, Domenech V. Muscle and genotype effects on fatty acid composition of goat kid intramuscular fat. Ital J Anim Sci. (2011) 10:212–6. doi: 10.4081/ijas.2011.e40
28. Ma JW, Yang J, Zhou LS, Ren J, Liu XX, Zhang H, et al. A splice mutation in the PHKG1 gene causes high glycogen content and low meat quality in pig skeletal muscle. PLoS Genet. (2014) 10:e1004710. doi: 10.1371/journal.pgen.1004710
29. Li BJ, Weng QN, Dong C, Zhang ZK, Li RY, Liu JG, et al. A key gene, PLIN1, can affect porcine intramuscular fat content based on transcriptome analysis. Genes. (2018) 9:194. doi: 10.3390/genes9040194
30. Ma C, Wang WW, Wang YD, Sun Y, Kang L, Zhang Q, et al. TMT-labeled quantitative proteomic analyses on the longissimus dorsi to identify the proteins underlying intramuscular fat content in pigs. J Proteomics. (2020) 213:103630. doi: 10.1016/j.jprot.2019.103630
31. Hou XH, Liu QF, Meng QS, Wang LG, Yan H, Zhang LC, et al. TMT-based quantitative proteomic analysis of porcine muscle associated with postmortem meat quality. Food Chem. (2020) 328:127133. doi: 10.1016/j.foodchem.2020.127133
32. Ma DY, Yu QQ, Hedrick VE, Cooper BR, Sobreira TJP, Oh JH, et al. Proteomic and metabolomic profiling reveals the involvement of apoptosis in meat quality characteristics of ovine M. longissimus from different callipyge genotypes. Meat Sci. (2020) 166:108140. doi: 10.1016/j.meatsci.2020.108140
33. Poleti MD, Regitano LCA, Souza G, Cesar ASM, Simas RC, Silva-Vignato B, et al. Longissimus dorsi muscle label-free quantitative proteomic reveals biological mechanisms associated with intramuscular fat deposition. J Proteomics. (2018) 179:30–41. doi: 10.1016/j.jprot.2018.02.028
34. Zhang J, Cao J, Geng A, Wang H, Chu Q, Yang L, et al. Comprehensive proteomic characterization of the pectoralis major at three chronological ages in Beijing-you chicken. Front Physiol. (2021) 12:658711. doi: 10.3389/fphys.2021.658711
35. Li W, Li M, Cao X, Han H, Kong F, Yue X. Comparative analysis of whey proteins in donkey colostrum and mature milk using quantitative proteomics. Food Res Int. (2020) 127:108741. doi: 10.1016/j.foodres.2019.108741
36. Zhang X, Jiang B, Ji C, Li H, Yang L, Jiang G, et al. Quantitative label-free proteomic analysis of milk fat globule membrane in donkey and human milk. Front Nutr. (2021) 8:670099. doi: 10.3389/fnut.2021.670099
37. Li B, Feng C, Zhu S, Zhang J, Irwin DM, Zhang X, et al. Identification of candidate circular RNAs underlying intramuscular fat content in the donkey. Front Genet. (2020) 11:587559. doi: 10.3389/fgene.2020.587559
38. Wisniewski JR, Zougman A, Nagaraj N, Mann M. Universal sample preparation method for proteome analysis. Nat Methods. (2009) 6:359–62. doi: 10.1038/nmeth.1322
39. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene ontology: tool for the unification of biology. Gene Ontol Consortium Nat Genet. (2000) 25:25–9. doi: 10.1038/75556
40. Gotz S, Garcia-Gomez JM, Terol J, Williams TD, Nagaraj SH, Nueda MJ, et al. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. (2008) 36:3420–35. doi: 10.1093/nar/gkn176
41. Kanehisa M, Goto S, Sato Y, Furumichi M, Tanabe M KEGG. for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. (2012) 40:D109–114. doi: 10.1093/nar/gkr988
42. Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. (2019) 47:D607–13. doi: 10.1093/nar/gky1131
43. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. (2003) 13:2498–504. doi: 10.1101/gr.1239303
44. Liu G, Wong L, Chua HN. Complex discovery from weighted PPI networks. Bioinformatics. (2009) 25:1891–7. doi: 10.1093/bioinformatics/btp311
45. Polidori P, Cammertoni N, Santini G, Klimanova Y, Zhang JJ, Vincenzetti S. Effects of donkeys rearing system on performance indices, carcass, and meat quality. Foods. (2021) 10:3119. doi: 10.3390/foods10123119
46. Du Y, Wang Y, Xu Q, Zhu J, Lin Y. TMT-based quantitative proteomics analysis reveals the key proteins related with the differentiation process of goat intramuscular adipocytes. BMC Genom. (2021) 22:417. doi: 10.1186/s12864-021-07730-y
47. Noguchi M, Hosoda K, Fujikura J, Fujimoto M, Iwakura H, Tomita T, et al. Genetic and pharmacological inhibition of Rho-associated kinase II enhances adipogenesis. J Biol Chem. (2007) 282:29574. doi: 10.1074/jbc.M705972200
48. Ahbara A, Bahbahani H, Almathen F, Al Abri M, Agoub MO, Abeba A, et al. Genome-wide variation, candidate regions and genes associated with fat deposition and tail morphology in Ethiopian indigenous sheep. Front Genet. (2018) 9:699. doi: 10.3389/fgene.2018.00699
49. Yamada T, Sasaki S, Sukegawa S, Miyake T, Fujita T, Kose H, et al. Association of a single nucleotide polymorphism in ribosomal protein L27a gene with marbling in Japanese Black beef cattle. Anim Sci J. (2009) 80:631–5. doi: 10.1111/j.1740-0929.2009.00688.x
50. Tu ZL Yu B, Huang DY, Ojha R, Zhou SK, An HD, et al. Proteomic analysis and comparison of intra and extracranial cerebral atherosclerosis responses to hyperlipidemia in rabbits. Mol Med Rep. (2017) 16:2347–54. doi: 10.3892/mmr.2017.6869
51. Kim DI, Park MJ, Lim SK, Park JI, Yoon KC, Han HJ, et al. PRMT3 regulates hepatic lipogenesis through direct interaction with LXRα. Diabetes. (2015) 64:60. doi: 10.2337/db13-1394
52. Zhao X, Yang Y, Sun BF, Yue S, Xin Y, Xiao W, et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 24:1403–19. doi: 10.1038/cr.2014.151
53. Liu F, Clark W, Luo G, Wang X, He CJC. ALKBH1-mediated tRNA demethylation regulates translation. (2016) 167:233–47. doi: 10.1016/j.cell.2016.11.045
54. Huang H, Song TJ Li X, Hu L, He Q, Liu M, et al. BMP signaling pathway is required for commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc Natl Acad Sci USA. (2009) 106:12670–5. doi: 10.1073/pnas.0906266106
55. Jung DS Baik Baik M Up-regulation of bone morphogenetic protein and its signaling molecules following castration of bulls and their association with intramuscular fat content in Korean cattle. Sci Rep-UK. (2019) 9:1–7. doi: 10.1038/s41598-019-56439-2
56. Sanyal A, Naumann J, Hoffmann LS, Chabowska-Kita A, Ehrlund A, Schlitzer A, et al. Interplay between obesity-induced inflammation and cGMP signaling in white adipose tissue. Cell Rep. (2017) 18:225. doi: 10.1016/j.celrep.2016.12.028
57. Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, et al. Inhibition of adipogenesis by Wnt signaling. Science. (2000) 289:950–3. doi: 10.1126/science.289.5481.950
58. Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol. (2006) 7:885–96. doi: 10.1038/nrm2066
59. Wright WS, Longo KA, Dolinsky VW, Gerin I, Kang S, Bennett CN, et al. Wnt10b inhibits obesity in ob/ob and agouti mice. Diabetes. (2007) 56:295–303. doi: 10.2337/db06-1339
60. Hong JH Hwang ES McManus MT Amsterdam A Tian Y Kalmukova RTAZ a a transcriptional modulator of mesenchymal stem cell differentiation. J Sci. (2005) 309:1074–8. doi: 10.1126/science.1110955
61. Deng K, Ren C, Fan Y, Pang J, Zhang G, Zhang Y, et al. YAP1 regulates PPARG and RXR alpha expression to affect the proliferation and differentiation of ovine preadipocyte. J Cell Biochem. (2019) 120:19578–89. doi: 10.1002/jcb.29265
62. Ibar C, Irvine KD. DC integration of hippo-YAP signaling with metabolism. Dev Cell. (2020) 54:256–67. doi: 10.1016/j.devcel.2020.06.025
Keywords: donkey, IMF, proteomics, differentially abundant proteins, functional analysis
Citation: Tan X, He Y, Qin Y, Yan Z, Chen J, Zhao R, Zhou S, Irwin DM, Li B and Zhang S (2022) Comparative analysis of differentially abundant proteins between high and low intramuscular fat content groups in donkeys. Front. Vet. Sci. 9:951168. doi: 10.3389/fvets.2022.951168
Received: 23 May 2022; Accepted: 05 July 2022;
Published: 27 July 2022.
Edited by:
Anning Li, Northwest A&F University, ChinaReviewed by:
Xing Du, Nanjing Agricultural University, ChinaCopyright © 2022 Tan, He, Qin, Yan, Chen, Zhao, Zhou, Irwin, Li and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bojiang Li, Ym9qaWFuZ2xpQHN5YXUuZWR1LmNu; Shuyi Zhang, c3poYW5nQHN5YXUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.