 Zhigao An1,2†
Zhigao An1,2† Mohamed Abdelrahman1,3†
Mohamed Abdelrahman1,3† Jiayan Zhou1,2†
Jiayan Zhou1,2† Umair Riaz1,2,4Shanshan Gao1,2Shan Gao1,2Gan Luo1
Umair Riaz1,2,4Shanshan Gao1,2Shan Gao1,2Gan Luo1 Liguo Yang1,2,5*
Liguo Yang1,2,5*- 1Key Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Education, College of Animal Science and Technology, Huazhong Agricultural University, Wuhan, China
- 2International Joint Research Centre for Animal Genetics, Breeding and Reproduction (IJRCAGBR), Huazhong Agricultural University, Wuhan, China
- 3Animal Production Department, Faculty of Agriculture, Assuit University, Asyut, Egypt
- 4Faculty of Veterinary and Animal Sciences, The Islamia University of Bahawalpur, Bahawalpur, Pakistan
- 5Hubei Province's Engineering Research Center in Buffalo Breeding and Products, Wuhan, China
The present study aims to evaluate the effects of prepartum maternal supplementation of Capsicum oleoresin (CAP) on colostrum quality and growth performance in newborn buffalo calves. Twelve multiparous buffaloes were randomly assigned to two groups starting from 4 weeks prepartum: the control group with a basal diet (CON) and the treatment group with a basal diet supplemented with 20 mg CAP/kg dry matter (CAP20). After birth, all calves were weighed and received colostrum from their respective dam directly within 2 h. After that, calves received pasteurized milk and starter feed till 56 days of age. The results showed that CAP increased lactose (P < 0.05) in colostrum, and it tended to increase monounsaturated fatty acids; however, it decreased colostrum urea nitrogen (P < 0.10). CAP did not affect colostrum yield and immunoglobulin G and M concentrations. The weekly starter intake was not affected by maternal CAP supplementation during the first 6 weeks of life. There was an increasing tendency in weekly starter intake from weeks 7 and 8 (P < 0.10) in CAP20 compared with CON. At 7 days of age, calves in CAP20 had higher immunoglobulin G (P < 0.05) and a decreased tendency in calves' serum glucose compared with CON. Additionally, maternal CAP supplementation increased calves' serum β-hydroxybutyric acid (P < 0.05) and tended to increase total protein (P < 0.10), while decreased non-esterified fatty acids (P < 0.05) at 56 days of age. Calves in CAP20 had higher final withers height, final heart girth, average withers height, and average heart girth than the CON (P < 0.05). These results suggest that maternal CAP supplementation could improve colostrum quality and positively affect the performance of buffalo calves.
Introduction
The buffalo is a major livestock species that significantly contribute to milk and meat production worldwide. Buffaloes show outstanding productive performance under harsh climatic conditions; it also resistant to various diseases and parasites compared to cattle Naveena and Kiran (1).
The maternal environment plays a critical role in the fetus's growth and development during gestation, and manipulation during this stage can significantly affect the calf's performance (2). Prenatal maternal nutrition may directly affect fetal growth (3), metabolism (4), endocrinology (5), immune system (6), and gene expression (7). At the same time, prenatal nutrition affects colostrum composition and immunoglobulin content (8, 9). Therefore, prenatal nutrition strategies can improve colostrum quality and calf performance.
After birth, calves are confronted with various stressors. In particular, the changes in nutrition from the dam's milk to the calves' digestion and nutrient absorption (10). However, because newborn calves have not yet developed immune systems and have immature digestive systems (11), any changes in their diet might significantly impact their growth (12). Therefore, efficient feeding management is essential to improve their growth and future performance since growing calves are profitable for cattle farms.
Colostrum is the first meal that is crucial for calves' health and survival (13). Besides providing nutrients and growth factors, colostrum provides early immune protection for newborn calves by providing maternal antibodies that cannot pass through the placenta. Thus, it is vital to maintain normal physiological function and growth in newborn calves (14).
It was reported that many factors might affect colostrum quality, including maternal diet management (15). It seems feasible to improve colostrum quality through prepartum nutritional strategies (9, 16). Among feed additives, herbal additives could potentially improve the colostrum quality and affect young animals' health and growth. It has been reported that the inclusion of Rosmarinus officinalis L. essential oils in the prenatal maternal diet could improve colostrum non-milk fat solids and tends to increase colostrum production in ewes (17).
Capsicum oleoresin (CAP) is a commercial feed additive extracted as a component of chili peppers belonging to the genus Capsicum. It has been shown that CAP could enhance the energy of corrective milk and affect the immune system in dairy cows (18). Likewise, dietary supplementation with capsaicin can improve lactating cows' nutritional status and milk production (19). Also, adding Capsicum oleoresins to the sows' diet in late gestation can improve total protein levels in colostrum and increase the proportion of piglets born alive per litter (20).
Therefore, we hypothesized that CAP supplementation would improve colostrum quality and thus positively affects calves' feed intake and growth performance. This study aimed to evaluate the effects of prepartum CAP addition on the dams' colostrum quality and buffalo calves' growth performance.
Materials and methods
Maternal treatments
The experiment was conducted on a commercial buffalo farm from August to October 2021 (Jinniu Animal Husbandry Co., LTD., Hubei, China). Four weeks before the expected date of birth, 12 multiparous pregnant buffaloes were randomly assigned to two treatments diet: the control group (n = 6) was given the basal diet (CON), while the treatment group (n = 6) was given the basal diet supplemented with 20 mg CAP/kg dry matter (CAP20). The commercial CAP product (10.0% capsaicin; Tianxu Food Additive Co. LTD, China) was dissolved in water and mixed into the diet before feeding. The buffaloes' diet was composed of corn silage, straw, corn meal, soybean meal, mineral, and vitamin mix. The nutritional composition is given in Table 1, and the nutrient requirements to meet the requirements of pregnant buffaloes are based on a previous report (21). All buffaloes were fed individually and had free access to water and feed before calving. All calves are born normally without any calving complications.
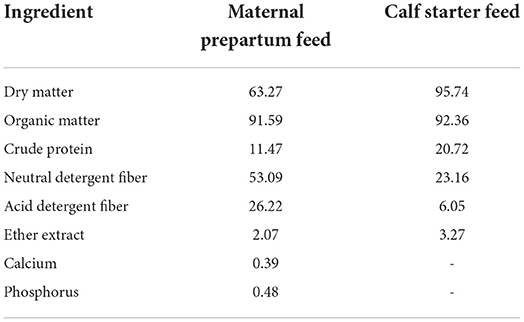
Table 1. Nutrient composition (% dry matter) of maternal prepartum diet and calves starter feed.
Calf management
Buffalo calves were separated immediately after calving from the dams and moved to individual pens (4.5 × 4 m) bedded with straw. Calving day was considered as 0 d of age (DOA). The dams were milked within 2 h of parturition, and the colostrum from respective dams was collected to feed the calves. Starting at two DOA, calves were fed 5 L/day of pasteurized milk twice daily (6 am and 6 pm). All calves were fed individually with starter feed (MG 200, Huludao Meigao Animal Husbandry Co., LTD, Liaoning, China) and freshwater until 56 DOA. The starter feed was composed of corn, soybean meal, bran, vinasse, calcium bicarbonate, rock flour, sodium chloride, and compound premix. The nutritional composition is given in Table 1, and it fulfills buffalo calves' nutrient requirements based on the Nutrient Requirements of Dairy Cattle (22).
Measurements, sample collection, and laboratory analyses
Colostrum collection and analysis
Colostrum samples (100 ml) were collected from each buffalo individually by a milking machine (Asahi Bronte Machinery Co., LTD, Zibo, China) within 2 h after parturition and divided into two tubes of 50 mL. The first colostrum samples from all experimental animals have added potassium dichromate and conserved at 4°C and measured with a milk composition analyzer (CombiFoss FT+, Shanghai Jinmai Instrument Equipment Co., LTD, Shanghai, China) in Dairy Herd Improvement of Hubei province. The second colostrum sample was frozen at −20°C for immunoglobulin G (IgG) and M (IgM) estimations. Then, it was thawed at 4°C and centrifuged at 10,000 × g for 10 min to remove the lipid portion before analysis. The IgG (catalog number RX1600804B) and IgM (catalog number RX1600805B) in colostrum were evaluated using the bovine ELISA kits (Ruixin Biotechnology Co., LTD., Quanzhou, China), according to the method of the previous study (23).
Calf starter intake and feed sample collection
The calf starter intake was calculated by weighing the daily starter offered and refused. Calf starter feed and fresh water were provided ad libitum. For composition analysis, pregnant buffaloes' diet and calf starter feed were collected daily and frozen at −20°C. Then, samples were dried for 48 h at 65°C and crushed through a 1 mm sieve for the analysis of dry matter (24; method 945.15), crude protein (24; method 984.13), ether extract (24; method 920.29), ash (24; method 942.05), phosphorus (24; method 991.25), and calcium (25). According to the previous study (26), neutral detergent fiber (using α-amylase and sodium sulfite) and acid detergent fiber were analyzed. Milk samples from all buffaloes were collected weekly and analyzed by the milk composition analyzer. The average milk composition offered to calves was 6.61% fat, 4.29% protein, 4.84% lactose, and 16.68% total solids.
Calf blood sampling and analyses
Calves' blood samples were collected in vacutainer tubes (EDTAK2) from the jugular vein at 10 am of 7 DOA and 56 DOA. The serum was separated by centrifugation at 3,500 g for 10 min and frozen at −20°C until parameter determination. Serum non-esterified fatty acids (NEFA, catalog number A042-1), glucose (catalog number A154-1), total cholesterol (TC, catalog number A111-1), β-hydroxybutyric acid (BHB, catalog number E030-1), total protein (TP, catalog number A045-4-2), and triglycerides (TG, catalog number A110-1) were measured using commercial assay kits (Nanjing Jian Cheng Bioengineering Institute, Nanjing, China). In addition, serum insulin (RX1600758B) and IgG (RX1600804B) were measured using the ELISA kits (Ruixin Biotechnology Co., LTD., Quanzhou, China).
Growth and skeletal measurements
Bodyweight (BW) was measured at 0 DOA (initial BW) and 56 DOA (final BW) using the electronic scale (ZF8003–T2, Kunshan Jiefeiyuan Measuring Equipment Co., LTD, Jiangsu, China). Average daily gain (ADG) was calculated as the difference between the final and initial BW divided by 56. Feed efficiency was calculated as the ratio of ADG to dry matter intake (DMI, milk dry matter + calf starter dry matter). Body size measurements, including the body length (distance between the points of shoulder and rump), withers height (distance from the base of the front feet to the withers), hip width (distance between the points of hook bones), and heart girth (circumference of the chest) were measured according to a previous study (27).
Statistical analysis
All data were analyzed using the PROC MIXED procedure of SAS 9.4 (SAS Institute Inc.) according to a randomized design unless otherwise stated. Data of 7 DOA blood metabolites, initial BW, and skeletal measurements were analyzed using one-way ANOVA. Weekly starter intake was analyzed using the repeated measures mixed model including the fixed effects of treatment, time, and their interaction, with the calf as a random effect. Degrees of freedom were calculated using the Kenward-Roger approximation option of the MIXED procedure. Significance was determined at P ≤ 0.05, and trends were considered when 0.05 < P ≤ 0.10.
Results
Colostrum quality
The least-square mean of the colostrum yield and composition response to the maternal CAP supplementation are shown in Table 2. No differences in colostrum production, IgG, and IgM were observed between the CON and CAP20 (P > 0.05). In the composition of colostrum, CAP increased lactose (P < 0.05) and tended to increase monounsaturated fatty (MUFA) (P = 0.09), while tending to decrease milk urea nitrogen (MUN) (P = 0.07). There were no differences in colostrum fat, protein, solid not fat (SNF), total solids (TS), polyunsaturated fatty acid (PUFA), saturated fatty acid (SFA), trans fatty acid (TFA), and free fatty acid (FFA) (P > 0.05).
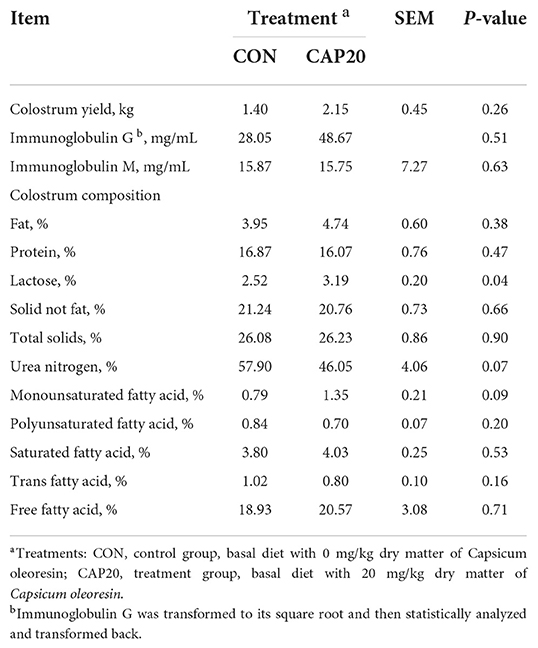
Table 2. Colostrum yield, immunoglobulin G (IgG), Immunoglobulin M (IgM), and colostrum composition data of buffaloes fed either 0 or 20 mg/kg dry matter of Capsicum oleoresin.
BW, calf starter intake, and feed efficiency
Prepartum maternal supplementation of CAP did not affect initial BW, final BW, ADG, total starter intake, and feed efficiency (Table 3). However, as presented in Figure 1, prepartum maternal supplementation of CAP tended to increase calf starter intake in weeks 7 and 8 (P < 0.10). Additionally, starter intake increased with increasing calves' age (time effect: P < 0.001). Also, there was no interaction between maternal CAP supplementation by week on weekly starter intake (P = 0.08).

Table 3. Calf starter intake, body weight, average daily gain, and feed efficiency of calves born from dams supplemented with either 0 or 20 mg/kg dry matter of Capsicum oleoresin.
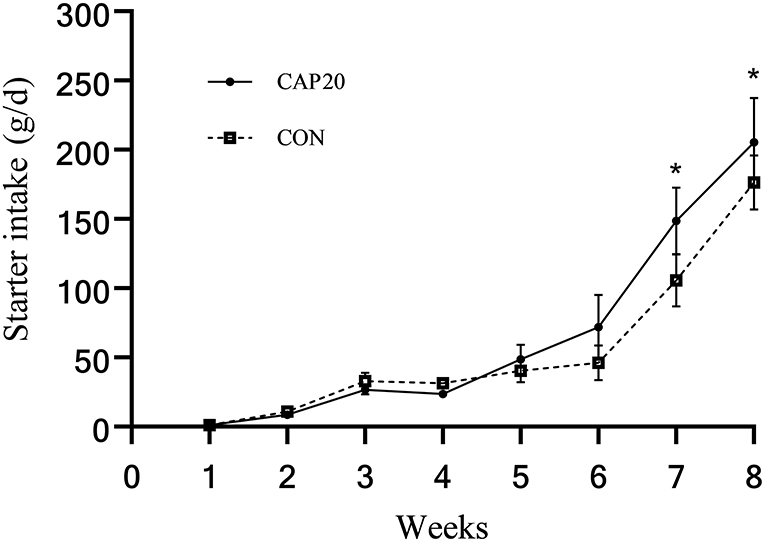
Figure 1. Weekly starter intake of calves born from dams supplemented either 0 or 20mg/kg dry matter of Capsicum oleoresin. Treatment effect, P = 0.36; week effect, P < 0.001; interaction of treatment by week, P = 0.08. *Signifies trends between groups (0.05 < P ≤ 0.10).
Calf blood metabolites
The blood parameters of the calves in both groups are shown in Table 4. At 7 DOA, maternal supplementation of CAP increased IgG (P < 0.05) and tended to decrease the glucose (P = 0.07). No difference in NEFA, BHB, insulin, TC, and TP between the CON and CAP20 (P > 0.10). However, at 56 DOA, maternal supplementation of CAP increased BHB (P < 0.05) and tended to increase TP (P = 0.08), while it resulted in decreased serum concentration of NEFA (P < 0.05), respectively. CAP supplementation of maternal did not affect serum insulin, glucose, TC, and IgG (P > 0.10) at 56 DOA.
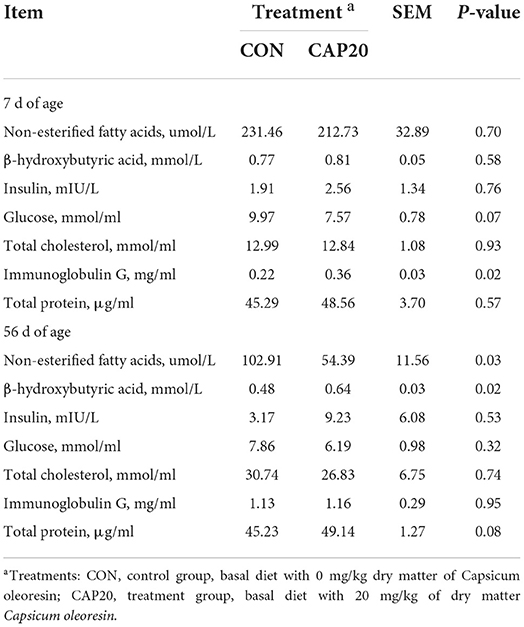
Table 4. Blood parameters of calves born from dams supplemented either 0 or 20 mg/kg dry matter of Capsicum oleoresin.
Calf skeletal growth
The least-square means for calf skeletal growth in both groups are listed in Table 5. There were no differences in initial skeletal measures of body length, wither height, hip width, and heart girth between the CON and CAP20 at 0 DOA (P > 0.05). Calves in CAP20 showed higher final withers height and heart girth than calves in CON (P < 0.05), while no difference was observed differences in final body length and hip width (P > 0.05). In average gain of calves skeletal, the maternal CAP supplementation could increase significantly in withers height and heart girth (P < 0.05), while no differences in body length and hip width.
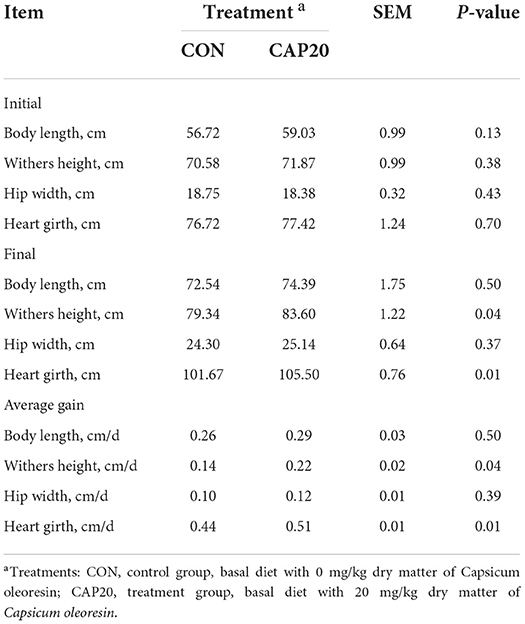
Table 5. Skeletal growth indices of calves born from dams supplemented either 0 or 20 mg/kg dry matter of Capsicum oleoresin.
Discussion
After birth, the calf's first meal of colostrum is critical for transferring IgG and IgM to support calf health and survival. However, the maternal CAP supplementation appears insignificant on colostrum immunoglobulin, which may be due to the low CAP concentration used in this study.
In the present study, there was an increase in colostrum lactose. Lactose is the main carbohydrate and energy source found in colostrum, which could increase glucose levels in newborn calf serum (28). Similarly, rumen-protected Capsicum supplementation could increase milk lactose in the transition period by effectively improving energy status and milk production in Holstein cows (29). Additionally, it was reported that adding essential oils mixture containing capsaicin to the diet before and during lactation could increase lactose in colostrum (30–32). Although there was no difference between SFA and PUFA, MUFA showed an increasing trend in CAP20. It has been reported that capsaicin can alter fat metabolism in prepartum Holstein cows (29), which seems to indicate that CAP was involved in the metabolism of milk fatty acids. In the current study, CAP tended to reduce urea nitrogen in colostrum. Previous reports discussed that capsaicin can cause vasodilation via transient receptor potential cation channel subfamily V member 1 (33, 34) and increase blood flow and permeability to the mammary glands until delivery. This may explain the variation of urea nitrogen in colostrum.
On the other hand, contrary to previous studies that have shown adding capsaicin to the sow's diet could improve piglet weight gain (32), there were no differences detected in total intake, body weight change, or feed efficiency in calves in both groups of the present study. Additionally, weekly starter intake in the CAP20 tended to increase at weeks 7 and 8. The fetus may be directly affected by the partial degradation of capsaicin (35), which caused the same result as buffalo calves. As calves grew, calf starter intake increased drastically, showing the treatment-by-week interaction effect throughout the experiment period. In the early weeks of calves' life, milk mainly covers energy intake and increases starter intake due to growing energy requirements.
In this study, the blood concentration IgG of calves showed an increase in CAP20 at 7 DOA, while there was no difference in IgG concentration in colostrum. However, the colostrum IgG concentration in CAP20 was 48.67 mg/ml vs. 28.05 mg/ml in CON. A significant relationship between IgG concentration and calf health has been reported (36, 37). Additionally, high IgG levels might increase growth rates based on energy directed to growth rather than the immune system (38). Therefore, plasma IgG concentration may correlate with early calves' later growth performance. Due to the difference in weekly starter intake, plasma BHB was higher in CAP20 than CON at 56 DOA (39). BHB is also an indicator of rumen function initiation (40, 41). Therefore, elevated plasma BHB levels may be due to reduced fatty acid oxidation caused by rumen supply of butyrate or body fat mobilization (42). In the present study, the changes in NEFA appear to be contrary to BHB, which suggests a lack of energy due to increased energy requirement intake and body fat mobilization in the CON group (43).
Calves in CAP20 had higher final withers height and heart girth, which was similar to indices of average gain, although it did not affect these measures at birth. Additionally, another reason for this result may be the difference in weekly feed intake. According to a model developed to predict Holstein calf's growth, appetizer intake was one of the factors affecting ADG and BW at weaning (44).
In conclusion, prepartum maternal supplementation of CAP in buffalo improved colostrum quality. It also positively affected the calves' growth performance and increased weekly starter intake.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was reviewed and approved by Huazhong Agricultural University Animal Care and Use Committee.
Author contributions
ZA, MA, and JZ conceived and designed the experiment data curation. ZA and MA wrote the manuscript. ShansG, ShanG, and GL contributed to the animals' arrangement and sample collection. MA, UR, and LY revised the manuscript. All the authors reviewed the manuscript.
Funding
This work was supported by the Modern Agro-industry Technology Research System: CARS-36.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Naveena B, Kiran M. Buffalo meat quality, composition, and processing characteristics: Contribution to the global economy and nutritional security. Animal Front. (2014) 4:18–24. doi: 10.2527/af.2014-0029
2. Reynolds L, Borowicz P, Caton J, Vonnahme K, Luther J, Hammer C, Maddock Carlin K, Grazul-Bilska A, Redmer D. Developmental programming: the concept, large animal models, and the key role of uteroplacental vascular development. J Animal Sci. (2010) 88(Suppl_13):E61–72. doi: 10.2527/jas.2009-2359
3. Funston R, Summers A, Roberts A. Alpharma beef cattle nutrition symposium: implications of nutritional management for beef cow-calf systems. J Anim Sci. (2012) 90:2301–7. doi: 10.2527/jas.2011-4568
4. Niederecker KN, Larson JM, Kallenbach RL, Meyer AM. Effects of feeding stockpiled tall fescue versus summer-baled tall fescue-based hay to late gestation beef cows: I. Cow performance, maternal metabolic status, and fetal growth. J Animal Sci. (2018) 96:4618–32. doi: 10.1093/jas/sky341
5. Alharthi A, Coleman D, Liang Y, Batistel F, Elolimy A, Yambao R, et al. Hepatic 1-carbon metabolism enzyme activity, intermediate metabolites, and growth in neonatal Holstein dairy calves are altered by maternal supply of methionine during late pregnancy. J Dairy Sci. (2019) 102:10291–303. doi: 10.3168/jds.2019-16562
6. Lopes M, Alharthi A, Lopreiato V, Abdel-Hamied E, Liang Y, Coleman D, et al. Maternal supplementation with cobalt sources, folic acid, and rumen-protected methionine and its effects on molecular and functional correlates of the immune system in neonatal Holstein calves. J Dairy Sci. (2021) 104:9340–54. doi: 10.3168/jds.2020-19674
7. Diniz WJ, Bobe G, Klopfenstein JJ, Gultekin Y, Davis TZ, Ward AK, et al. Supranutritional maternal organic selenium supplementation during different trimesters of pregnancy affects the muscle gene transcriptome of newborn beef calves in a time-dependent manner. Genes. (2021) 12:1884. doi: 10.3390/genes12121884
8. Westland A, Martin R, White R, Martin J. Mannan oligosaccharide prepartum supplementation: effects on dairy cow colostrum quality and quantity. Animal. (2017) 11:1779–82. doi: 10.1017/S1751731117000672
9. Morsy A, Eissa M, Anwer M, Ghobashy H, Sallam S, Soltan Y, et al. Colostral immunoglobulin concentration and milk production of ewes fed salt tolerant forages as alternatives to berseem hay. Livest Sci. (2018) 210:125–8. doi: 10.1016/j.livsci.2018.02.012
10. Ballou M, Hanson D, Cobb C, Obeidat B, Sellers M, Pepper-Yowell A, et al. Plane of nutrition influences the performance, innate leukocyte responses, and resistance to an oral Salmonella enterica serotype Typhimurium challenge in Jersey calves. J Dairy Sci. (2015) 98:1972–82. doi: 10.3168/jds.2014-8783
11. Stelwagen K, Carpenter E, Haigh B, Hodgkinson A, Wheeler T. Immune components of bovine colostrum and milk. J Animal Sci. (2009) 87(Suppl_13):3–9. doi: 10.2527/jas.2008-1377
12. Diao Q, Zhang R, Fu T. Review of strategies to promote rumen development in calves. Animals. (2019) 9:490. doi: 10.3390/ani9080490
13. Weaver DM, Tyler JW, VanMetre DC, Hostetler DE, Barrington GM. Passive transfer of colostral immunoglobulins in calves. J Vet Int Med. (2000) 14:569–77. doi: 10.1111/j.1939-1676.2000.tb02278.x
14. Osorio J. Gut health, stress, and immunity in neonatal dairy calves: the host side of host-pathogen interactions. J Anim Sci Biotechnol. (2020) 11:1–15. doi: 10.1186/s40104-020-00509-3
15. Godden S. Colostrum management for dairy calves. Veterinary Clin North Am: Food Animal Pract. (2008) 24:19–39. doi: 10.1016/j.cvfa.2007.10.005
16. Hasan S, Junnikkala S, Peltoniemi O, Paulin L, Lyyski A, Vuorenmaa J, et al. Dietary supplementation with yeast hydrolysate in pregnancy influences colostrum yield and gut microbiota of sows and piglets after birth. PLoS ONE. (2018) 13:e0197586. doi: 10.1371/journal.pone.0197586
17. Smeti S, Joy M, Hajji H, Alabart JL, Muñoz F, Mahouachi M, et al. Effects of R osmarinus officinalis L. essential oils supplementation on digestion, colostrum production of dairy ewes and lamb mortality and growth. J Animal Sci. (2015) 86:679–88. doi: 10.1111/asj.12352
18. Oh J, Giallongo F, Frederick T, Pate J, Walusimbi S, Elias R, et al. Effects of dietary Capsicum oleoresin on productivity and immune responses in lactating dairy cows. J Dairy Sci. (2015) 98:6327–39. doi: 10.3168/jds.2014-9294
19. Rodrigues RO, Cooke RF, Firmino FC, Moura MK, Angeli BF, Ferreira HA, et al. Productive and physiological responses of lactating dairy cows supplemented with phytogenic feed ingredients. Trans Animal Sci. (2019) 3:1133–42. doi: 10.1093/tas/txz108
20. Oguey C, Riu I, Quintilla C, Lopez S. 1017 A standardized blend of capsicum and turmeric oleoresins given during late gestation improves the performance of sows vaccinated against E. coli. J Animal Sci. (2016) 94(Suppl. 5):487. doi: 10.2527/jam2016-1017
21. Bartocci S, Tripaldi C, Terramoccia S. Characteristics of foodstuffs and diets, and the quanti-qualitative milk parameters of Mediterranean buffaloes bred in Italy using the intensive system: an estimate of the nutritional requirements of buffalo herds lactating or dry. Livestock Product Sci. (2002) 77:45–58. doi: 10.1016/S0301-6226(02)00022-2
22. Council NR. Nutrient Requirements of Dairy Cattle: 2001. Washington, DC: National Academies Press (2001).
23. Martin P, Vinet A, Denis C, Grohs C, Chanteloup L, Dozias D, et al. Determination of immunoglobulin concentrations and genetic parameters for colostrum and calf serum in Charolais animals. J Dairy Sci. (2021) 104:3240–9. doi: 10.3168/jds.2020-19423
24. AOAC A. Official Method 990.12 for Aerobic Plate Count in Foods, Official Methods of Analysis. Washington, DC: Association of Official Analytical Chemists (1995).
25. Brooks I, Luster G, Easterly D. A procedure for the rapid determination of the major cations in milk by atomic absorption spectrophotometry. Atom Absorption Newsl. (1970) 9:93–4.
26. Van Soest Pv, Robertson J, Lewis B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds.S0022-0302(91)78551-2
27. Nemati M, Amanlou H, Khorvash M, Mirzaei M, Moshiri B, Ghaffari M. Effect of different alfalfa hay levels on growth performance, rumen fermentation, and structural growth of Holstein dairy calves. J Anim Sci. (2016) 94:1141–8. doi: 10.2527/jas.2015-0111
28. Kekana T, Marume U, Nherera-Chokuda F. Prepartum supplementation of Moringa oleifera leaf meal: Effects on health of the dam, colostrum quality, and acquisition of immunity in the calf. J Dairy Sci. (2022) 105:5813–821. doi: 10.3168/jds.2021-21535
29. Oh J, Harper M, Melgar A, Räisänen S, Chen X, Nedelkov K, etal. Dietary supplementation with rumen-protected capsicum during the transition period improves the metabolic status of dairy cows. J Dairy Sci. (2021) 104:11609–20. doi: 10.3168/jds.2020-19892
30. Matysiak B, Jacyno E, Kawecka M, Kołodziej-Skalska A, Pietruszka A. The effect of plant extracts fed before farrowing and during lactation on sow and piglet performance. S Afr J Anim Sci. (2012) 42:15–21. doi: 10.4314/sajas.v42i1.2
31. Miller H. Effects of sow and piglet dietary supplementation with a plant extract additive on the composition of sow colostrums and milk (day 21) and its effects on piglet development from birth to day 6 postweaning. Final Year Project, Alex Moore. (2003) 27:2003.
32. Medina ER. The effect of including capsaicin and gut microbiota feed additives on growth performance of nursery pigs (MSc Thesis). University of Minnesota, Minneapolis, MN, United States. (2020). Available online at: https://conservancy.umn.edu/bitstream/handle/11299/219291/RosaMedina_umn_0130M_22039.pdf?sequence=1
33. Van der Schueren BJ, Rogiers A, Vanmolkot FH, Van Hecken A, Depre M, Kane SA, et al. Calcitonin gene-related peptide8-37 antagonizes capsaicin-induced vasodilation in the skin: evaluation of a human in vivo pharmacodynamic model. J Pharmacol Experiment Therapeutics. (2008) 325:248–55. doi: 10.1124/jpet.107.133868
34. Munjuluri S, Wilkerson DA, Sooch G, Chen X, White FA, Obukhov AG. Capsaicin and TRPV1 channels in the cardiovascular system: the role of inflammation. Cells. (2021) 11:18. doi: 10.3390/cells11010018
35. Oh J, Bravo D, Wall E, Hristov A. Rumen disappearance of capsaicin and dihydrocapsaicin in lactating dairy cows. J Anim Sci. (2016) 94:801–801. doi: 10.2527/jam2016-1644
36. Conneely M, Berry D, Murphy JP, Lorenz I, Doherty ML, Kennedy E. Effect of feeding colostrum at different volumes and subsequent number of transition milk feeds on the serum immunoglobulin G concentration and health status of dairy calves. J Dairy Sci. (2014) 97:6991–7000. doi: 10.3168/jds.2013-7494
37. Gelsinger S, Heinrichs A. A short review: the immune system of the dairy calf and the importance of colostrum IgG. J Dairy Vet Anim Res. (2017) 5:104–7. doi: 10.15406/jdvar.2017.05.00144
38. Froehlich K, Abdelsalam K, Chase C, Koppien-Fox J, Casper D. Evaluation of essential oils and prebiotics for newborn dairy calves. J Anim Sci. (2017) 95:3772–82. doi: 10.2527/jas.2017.1601
39. Frieten D, Gerbert C, Koch C, Dusel G, Eder K, Kanitz E, et al. Ad libitum milk replacer feeding, but not butyrate supplementation, affects growth performance as well as metabolic and endocrine traits in Holstein calves. J Dairy Sci. (2017) 100:6648–61. doi: 10.3168/jds.2017-12722
40. Naeem A, Drackley J, Stamey J, Loor J. Role of metabolic and cellular proliferation genes in ruminal development in response to enhanced plane of nutrition in neonatal Holstein calves. J Dairy Sci. (2012) 95:1807–20. doi: 10.3168/jds.2011-4709
41. Khan M, Lee H, Lee W, Kim H, Kim S, Ki K, et al. Starch source evaluation in calf starter: I. Feed consumption, body weight gain, structural growth, and blood metabolites in Holstein calves. J Dairy Sci. (2007) 90:5259–68. doi: 10.3168/jds.2007-0338
42. Kesser J, Korst M, Koch C, Romberg F-J, Rehage J, Müller U, et al. Different milk feeding intensities during the first 4 weeks of rearing dairy calves: part 2: effects on the metabolic and endocrine status during calfhood and around the first lactation. J Dairy Sci. (2017) 100:3109–25. doi: 10.3168/jds.2016-11595
43. Schäff CT, Gruse J, Maciej J, Mielenz M, Wirthgen E, Hoeflich A, et al. Effects of feeding milk replacer ad libitum or in restricted amounts for the first five weeks of life on the growth, metabolic adaptation, and immune status of newborn calves. PLoS ONE. (2016) 11:e0168974. doi: 10.1371/journal.pone.0168974
Keywords: Capsicum oleoresin, maternal supplementation, colostrum, buffalo calf, growth performance
Citation: An Z, Abdelrahman M, Zhou J, Riaz U, Gao S, Gao S, Luo G and Yang L (2022) Prepartum maternal supplementation of Capsicum oleoresin improves colostrum quality and buffalo calves' performance. Front. Vet. Sci. 9:935634. doi: 10.3389/fvets.2022.935634
Received: 04 May 2022; Accepted: 09 September 2022;
Published: 04 October 2022.
Edited by:
Yosra Ahmed Soltan, Alexandria University, EgyptReviewed by:
Peter Erickson, University of New Hampshire, United StatesAmr Salah Morsy, Arid Lands Cultivation Research Institute (ALCRI), Egypt
Copyright © 2022 An, Abdelrahman, Zhou, Riaz, Gao, Gao, Luo and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liguo Yang, eWFuZ2xpZ3VvMjAwNkBxcS5jb20=
†These authors have contributed equally to this work