 Norma Hernández-Jardón1
Norma Hernández-Jardón1 Julio César Rojas-Castañeda2*
Julio César Rojas-Castañeda2* Daniel Landero-Huerta2
Daniel Landero-Huerta2 Estefanía Reyes-Cruz1
Estefanía Reyes-Cruz1 Rafael Reynoso-Robles3
Rafael Reynoso-Robles3 María del Lourdes Juárez-Mosqueda4
María del Lourdes Juárez-Mosqueda4 Alfredo Medrano5Fausto Reyes-Delgado6
Alfredo Medrano5Fausto Reyes-Delgado6 Rosa María Vigueras-Villaseñor2*
Rosa María Vigueras-Villaseñor2*- 1Programa Doctorado en Ciencias de la Producción y de la Salud Animal, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 2Laboratorio de Biología de la Reproducción, Instituto Nacional de Pediatría, SS, Mexico City, Mexico
- 3Laboratorio de Morfología Celular y Tisular, Instituto Nacional de Pediatría, SS, Mexico City, Mexico
- 4Departamento de Morfología, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 5Laboratorio de Reproducción Animal, Facultad de Estudios Superiores Cuautitlán, Universidad Nacional Autónoma de México, Cuautitlán Izcalli, Mexico
- 6Banfield Pet Hospital-Universidad Nacional Autónoma de México, Mexico City, Mexico
Cryptorchidism (CO) or undescended testicle is an abnormality of male gonadal development that can generate long-term repercussions in men, such as infertility and germ cell neoplasia in situ (GCNIS). The origin of these alterations in humans is not completely clear, due to the absence of an animal model with similar testicular development as in humans with CO. This work intends to describe the testicular histological development of dogs with congenital CO, and determine whether the species could adequately serve as a study model for this pathology in humans. The study was carried out with 36 dogs, equally distributed in two groups: healthy control (CTRL) and CO groups. The contralateral testis to the undescended one in CO group of the animals was considered and analyzed. Each group was subdivided in three stages of development: (1) peripubertal stage (6–8 months), (2) young adult (9–48 months) and (3) senile (49–130 months). Histological development, the presence of cells with gonocyte morphology, cell proliferation, testicular lipoperoxidation and hormonal concentrations of testosterone, estradiol, FSH and LH were evaluated and described. In the cryptorchid testes, the first histological alterations appeared from the first stage of development and were maintained until the senile stage. A pronounced testicular lipoperoxidation occurred only in the second stage of development. The histological alterations due to CO were markedly evident in the young adult stage. Testosterone concentrations witnessed a decrease starting from in the second stage and kept on until the last stage. The contralateral testes of the CO animals showed alterations that positioned them between the control and CO testes. Testicular development of dogs with CO is similar to that of humans. The results of the study suggest that this species could serve as a suitable model for the study of CO in humans.
Introduction
Cryptorchidism (CO) or undescended testis is the most common abnormality of male reproductive development (1). In premature children, the incidence is 1.1–45.3% (2) while in term newborn and 1-year aged males, it is 3–5 and 1.0–1.5% respectively (3–5). The etiology is associated with environmental, anatomical and genetic factors (6). Its importance resides on the long-term effects, since it predisposes patients to the development of infertility and germ cell neoplasia in situ (GCNIS) (7, 8). GCNIS is the histological lesion that precedes the development of testicular germ cell tumor (TGCT) (9).
Reported fertility in men who underwent CO and orchidopexy is 89.7% for unilateral CO and 65.3% for bilateral CO (2). Moreover, oligospermia is documented in 43 and 73% of patients with unilateral and bilateral CO respectively (10). It is mentioned that the oxidative stress generated by the high temperature in the suprascrotal area can damage the Sertoli cells function (11). This condition deprives the germ cells of the factors necessary for their differentiation, or their binding proteins (12). Although, it is possible that free radicals directly affect germ cells by inhibiting spermatogenesis (13).
The prevalence of GCNIS is estimated to be 2.9% in men with a history of CO (14). Undifferentiated gonocytes with pluripotent capacity are suggested to be the target of malignancy, and thus the origin of this neoplasm (9, 15). A gonocyte is a cell of embryonic origin that differentiates into a spermatogonia stem cell in the last third of gestation period and continues through the first 10 months of postnatal period to give rise to the germ line that will initiate spermatogenesis in the future (16–19).
Knowledge about the etiopathogenesis of GCNIS and infertility in patients with CO is still limited (20). This may be attributed to the current lack of an animal model that show similar testicular development as in human. Inevitably, this situation may have affected the efforts to find alternative ways to reduce the risk of developing testicular cancer and fertility problems in these patients.
The most used animal models for the study of CO in man have been the rat, mouse and rabbit. In these species, CO is induced by the administration of drugs or surgical manipulation, which can generate stress, surgical iatrogenesis and tissue adhesions, thus compromising the results (21). The rat and the mouse have the disadvantages that the period of gonocyte differentiation to spermatogonia (4–10 days of postnatal life) and the so-called “minipuberty” (the first postnatal hours) are short and without quiescence period (22). Therefore, its experimental management implies challenges.
The domestic dog (Canis familiaris) presents the advantage of spontaneous CO development, with a frequency of 6.8% (23). In dogs, the CO presentation can be unilateral or bilateral, the retained testis positioned in inguinal or abdominal the right unilateral inguinal region being the most frequent as in humans (23, 24). In addition, CO in the dog increases the risk of presenting testicular neoplasia from 3.6 to 13.6 times (25). Coupled with this, is the likelihood of developing tumors of the germ cells; such as Sertoli-Leydig cells tumor (26), and classic (26) and spermatocytic seminomas (27). Also, the differentiation period of the gonocytes in dogs, as a target of malignancy, extends up to the first 4 months of age (28).
Based on the above exposed points, the objective of this study was to describe the histological development of the canine testis with spontaneous CO, at different stages of development, to determine if this species can be best suited as a study model for this pathology in humans.
Materials and methods
Animals
Thirty-six small-breed dogs (≤ 10 kg) were included in the study. The animals were from several private veterinary clinics, UNAM-Banfield Veterinary Hospital and the Office of the Mobile Sterilization Unit for Dogs and Cats in Alvaro Obregon borough, Mexico City, Mexico. Animals that presented infections, chronic degenerative diseases, malnutrition, testicular tumors and problems in the reproductive system unrelated to CO were excluded from the study.
For the purposes of the study, the dogs were divided into two equal groups of 18 animals each, consisting of healthy control (CTRL) and unilateral inguinal CO animals, clinically diagnosed by palpation and ultrasound imaging study. Each group was subdivided with an equal number of animals into three stages of development as follows: (1) peripuberal (6–8 months), representing the age of the spermatogenic onset and also the limit for testicular descent into the scrotum (28). The average age of the CTRL group was 6.42 ± 0.48 months and that of the CO group was 5.6 ± 0.41 months. (2) The age range of young adult animals was (9–48 months), which is the age of sexual maturity (26). The average age of the CTRL group was 41.14 ± 5.14 months and that of the CO group was 29.85 ± 5.35 months; and (3) while the age of senile animals ranged 49–130 months (26, 28), which encompasses the age of possible development of neoplasia (29, 30). The average age of the CTRL group was 102.85 ± 6.85 months and that of the CO group was 122.57 ± 11.04 months. In addition, in the CO group, the contralateral testicle (CONTRA) to the cryptorchid one, which was always in scrotal position, was analyzed. The testicular samples were obtained at orquiectomy; from each gonad, three samples of testicular tissue, ~0.25 cm3 each, from the mid-portion, opposite to the testicular mediastinum were obtained for morphological analysis, two samples of ~0.75 cm3 for immunohistochemistry and two of ~0.75 cm3 for the determination of lipoperoxidation. In addition, peripheral blood samples were obtained and used to hormonal determination.
Morphological analysis
The histological alterations induced by canine spontaneous CO condition were used to compare for resemblance with the histological pattern of human CO development.
The evaluation was performed by fixing testicular tissue samples in Karnovsky solution for 24 h, postfixed in 1% OsO4, dehydrated in gradual ethanol solutions and embedded in EPON 812 (Ted Pella, INC. CA, USA). Semi-thin sections of 1 μm thickness were cut using an Ultracut UCT ultramicrotome (Leica, Vienna, Austria) and stained with 0.5% toluidine blue. Histological analysis of the seminiferous tubules was performed using a light microscope (BX 51, Olympus, Tokyo, Japan). From each tissue sample, a slide with 5 tissue sections was obtained. Therefore, 5 cross-sectioned tubules were evaluated, resulting in a total of 15 tubules per animal. The area of the seminiferous epithelium was determined by subtracting the internal area from the external area. The cellular area of at least 15 Leydig cells per animal was also determined using an image analysis system (Image-Pro Plus 6.2. Media Cybernetics, INC. MD, USA). Leydig cells were evaluated using the 100x objective lens and identified by their central location in the interstitial space, its polygonal shape of round nucleus, with an evident nucleolus. The maturation index (MI) or Johnsen index (31) and the histopathological index (HI) were determined (32).
To confirm the presence of gonocytes and cellular alterations, the testicular tissue was sectioned at 60–70 nm thickness. Sections were counterstained with uranyl acetate and lead citrate and analyzed with an electron microscope (JEM-1011, JEOL, Osaka, Japan). The presence of gonocytes was determined through their ultramorphological characteristics, among which are large round cells with the presence of cytoplasmic extensions in some cases, prominent nucleus and nucleolus, and mitochondria peripheral to the nucleus.
Determination of cell proliferation and the functional status of Sertoli cells
Immunoreactivity to proliferating cell nuclear antigen (PCNA) and to vimentin antigen were evaluated using immunohistochemical techniques to determine if the spontaneous CO in the dogs presents cell hypoplasia due to low cell proliferation and alterations in the functional status of Sertoli cells.
The determination was carried out by fixing the two testicular samples per gonad in 4% paraformaldehyde for 2 h. Later, these were dehydrated, clarified and embedded in paraffin. Sections of 4 μm thickness were cut and mounted on slides with poly-L- lysine (Sigma-Aldrich, St. Louis, MO, USA). Tissue sections were deparaffinized with xylene and hydrated with graded ethanol solutions. Subsequently, tissue sections were exposed to sodium citrate (pH 7.6) for 5 min in a microwave oven at 800 W. Afterwards, sections were outlined with PAP pen for immunostaining (Dako, Carpinteria, CA, USA). The tissue sections were then treated with 3% hydrogen peroxide for 10 min. Subsequently, they were placed in 0.1% Tween 20 (Sigma-Aldrich) in phosphate buffered saline (PBS, pH 7.4) for 10 min and blocked with 1% bovine serum albumin in PBS for 2 h. Thereafter, they were incubated overnight at room temperature with mouse monoclonal primary antibody against PCNA (sc-56 B1717, Santa Cruz, Biotechnology, CA, USA) and the primary monoclonal antibody against vimentin (sc-6260, Santa Cruz Biotechnology) both at a dilution of 1:500. This was followed by another incubation with biotinylated anti-mouse IgG (Invitrogen, Carlsbad, CA,USA) at a dilution of 1:100 for 1 h. Finally, the samples were exposed to the streptavidin-horseradish peroxidase conjugate for 1 h (Vector Laboratories, Peterborough, UK). To visualize the immunoreaction, tissue sections were incubated in a peroxidase substrate solution containing 1.6 ml distilled H2O, 20 μl 10x substrate buffer and 40 μl 50x diaminobenzidine (chromogen) (Santa Cruz Biotechnology) and 1% H2O2 (Merck, Darmstadt, Germany) in methanol for 30 min. Subsequently, sections were counterstained with hematoxylin, dehydrated through a graded ethanol series and washed with xylene. All dilutions and washes between steps were done with PBS. The number of immunoreactive cells per transversely cut seminiferous tube was determined. 5 seminiferous tubes per section were evaluated, making a total of 10 tubes evaluated per animal. The distribution pattern of immunoreactivity to vimentin in the cytoplasm of Sertoli cells was also determined. In addition, the Masson trichrome technique was performed on one slide of each animal to show the collagen fibers.
Determination of lipoperoxidation
Malondialdehyde, the final product of testicular lipoperoxidation, was determined using TBARS technique with the objective to know if the spontaneous CO in the dogs presents an increase in lipoperoxidation as an indirect way of oxidative damage.
For this purpose, 1 ml aliquot containing homogenized testicular tissue was added to 2 ml of thiobarbituric acid reagent (TBA; MP, Illkirch, France) (0.375 g of TBA + 15 g of trichloroacetic acid + 2.5 ml of HCl diluted in 100 ml distilled water). The resulting solution (3 ml total volume) was heated in a boiling water bath for 30 min. Samples were cooled on ice and centrifuged at 3,000 g for 15 min. The absorbance was measured in the supernatants by spectrophotometry at 532 nm. The concentrations of substances reactive with thiobarbituric acid were calculated by interpolation on a standard curve of periodic oxidation of malondialdehyde. The final result was expressed as nmol of substances reactive to thiobarbituric acid per mg of protein. Protein content in testicular tissue samples was measured with Folin's phenol reagent (33). Lipid peroxidation results were corrected for protein content in each sample and expressed as pg MDA/mg protein.
Hormonal determination
Peripheral blood (5 ml) was performed at 9:00 AM to avoid circadian fluctuations and collected in sterile BD vacutainer tubes for serum extraction and centrifuged at 3500 rpm for 10 min. The serum obtained was stored at −70°C until analysis. The determination of testosterone, estradiol, LH, and FSH was performed for each individual without replicates and by automated chemiluminescent microparticle immunoassay, using the Architect R 2nd generation (Abbott Diagnostics, Abbott Park, IL, USA), with strict adherence to the manufacturers' instructions. The detection limits and the intra- and inter-assay coefficients of variation for each hormone, respectively, were as follows: testosterone 0.01 ng/ml, 3.10 and 3.60%; estradiol 10.00 pg/ml, 2.98 and 3.60%; LH 0.50 mIU/ml, 2.07 and 3.74%; FSH 0.05 mIU/mL, 2.99 and 3.80%.
Statistical analysis
The data were expressed as the mean ± SEM. A parametric ANOVA test, followed by a multiple comparison using the Tukey test (seminiferous epithelial area, MI, HI, proliferation index, and hormone concentrations) and non-parametric Kruskal–Wallis (lipoperoxidation) test were used for statistical analysis with P values < 0.05 considered statistically significant.
Results
Stage 1: Peripuberal
The CTRL group animals presented different degrees of development of spermatogenesis. The 16.6% of the animals (1/6) presented few gonocytes (verified with electron microscopy), spermatogonia and spermatocytes. In other specimens, it was possible to observe round and elongated spermatids. Slight histological alterations such as scaling were observed (Figure 1A). The ultrastructure showed completely healthy Sertoli and germ cells (Figure 2A).
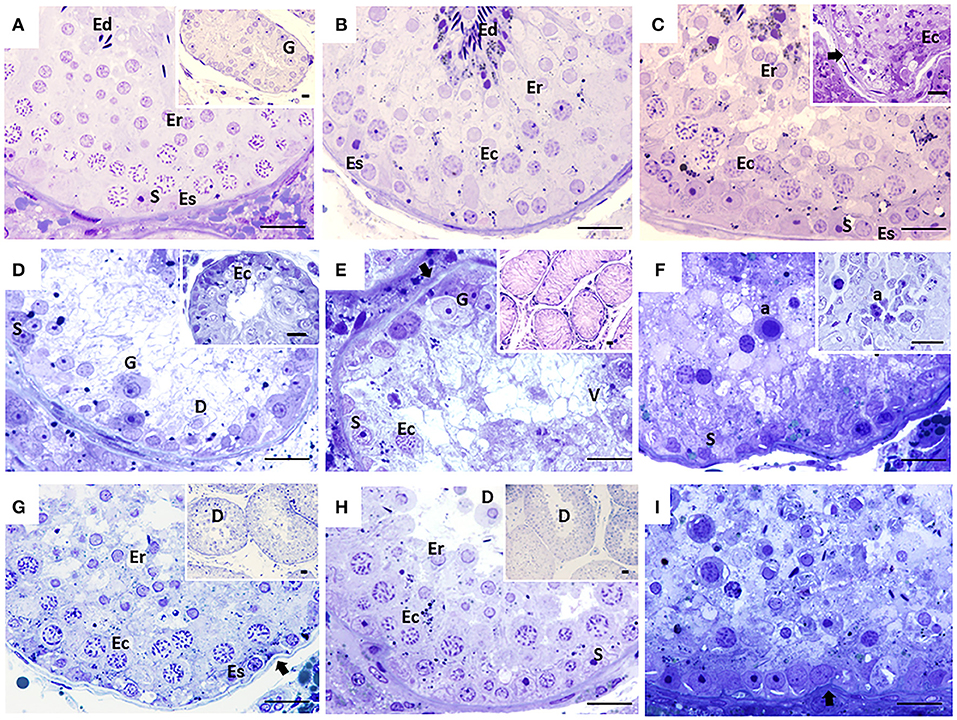
Figure 1. Seminiferous tubules of dogs from the control (CTRL) (A–C), cryptorchid (CO) (D–F) and contralateral (CONTRA) (G–I) groups at peripubertal (first column); young adult (second column) and senile (third column) stages. In CTRL dogs, complete spermatogenesis is observed, although gonocytes were still observed in the peripubertal stage (A). In the senile stage (C), some specimens showed histological alterations such as thickening and folding in the basement membrane (arrow) and even complete disorganization of the seminiferous epithelium (C insert). In animals with CO in the peripubertal stage, different degrees of maturation of the epithelium are observed, from the presence of gonocytes to spermatocytes (D insert). Histological alterations become evident from the young adult stage (E), which continued until senile (F) with vacuolization (V), desquamation (D), folding of the basement membrane (arrow), presence of atypical cells (F), cellular degeneration and arrest of spermatogenesis, as well as Sertoli-cell-only syndrome (E insert). In the contralateral testes, the degree of histological damage was not as severe as in the cryptorchid ones, presenting alterations such as hypoplasia and cellular degeneration, thickening and folding of the basement membrane (arrow) with cellular desquamation (D). The CONTRA group images inserts show a panoramic view of the tubes. Gonocyte (G), Spermatogonia (Es) spermatocyte (Ec), Round spermatid (Er), elongated spermatid (Ed), atypical cells (a), Sertoli cell nuclei (S). Toluidine Blue. Bar scale: 20μm.
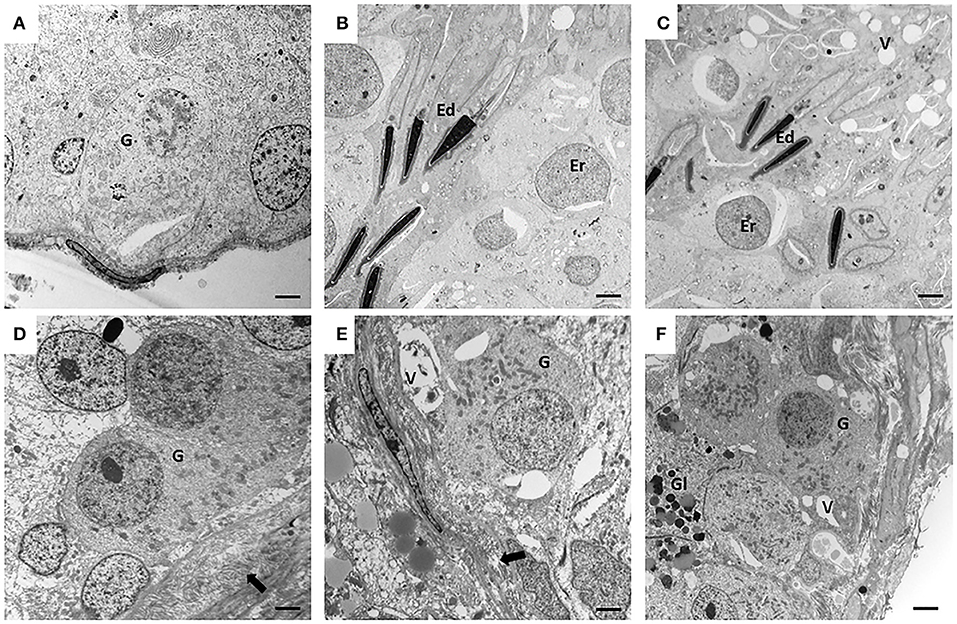
Figure 2. Electron microscopy (EM) of canine testes from the control groups (CTRL) (A–C) and from cryptorchidism (CO) (D–F). It is observed in the peripubertal stage, the presence of gonocytes in migration phase (A) and with small pseudopodia touching the basement membrane (D). In the young adult stage of the second row, a clear spermatogenesis is observed in the CTRL testis (B), unlike the CO testis, which shows the presence of a gonocyte-like cell with thickening of the basement membrane (E). In the senile stage, the CTRL testes still preserve spermatogenesis (C) with disorganization of the epithelium, and cells morphologically similar to gonocytes (F) were observed in the CO. Gonocyte (G), round spermatid (Er), elongated spermatid (Ed), basement membrane (arrow), vacuoles (V), glycogen (Gl). Electron microscopy. Bar scale: 2μm.
Fifty percent (3/6) of the animals of CO group, presented cells with gonocyte morphology (Figure 1D). Spermatogenesis did not develop beyond spermatocytes (Figure 1D) and slight histological alterations, such as tubular hypoplasia and cellular degeneration, mainly in spermatocytes, were observed in their seminiferous tubules. The animals of the CO group presented a significant reduction in the maturation index (MI) of 2.57 times as well as in cell proliferation index 4.1 times when compared with the CTRL group (Table 1; Figures 1D, 3A,D). However, histopathological index (HI) and seminiferous epithelial area did not show differences between both groups (p > 0.05). The result of TBARS technique did not show significant difference (p > 0.05) between both groups, although there was a tendency for greater lipoperoxidation in the CO group (2.07 times more. The analysis of the hormones show that T concentrations experienced a downward trend in the CO group when compared with the CTRL group, nevertheless, this was not significant (Table 2). The same trend was observed with the rest of the hormones (Table 2).
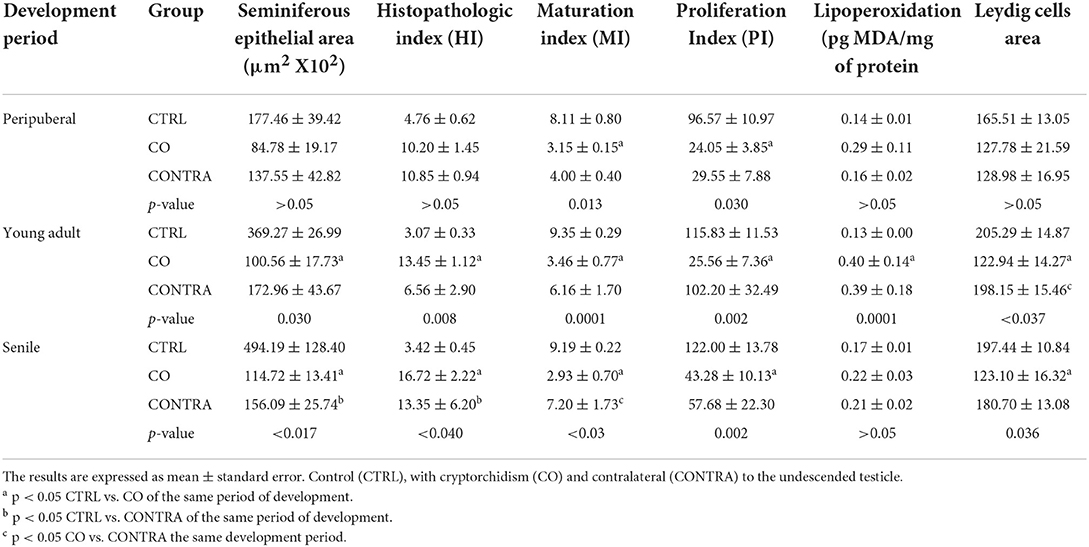
Table 1. Parameters evaluated in the studied groups at different stages of development of the dogs.
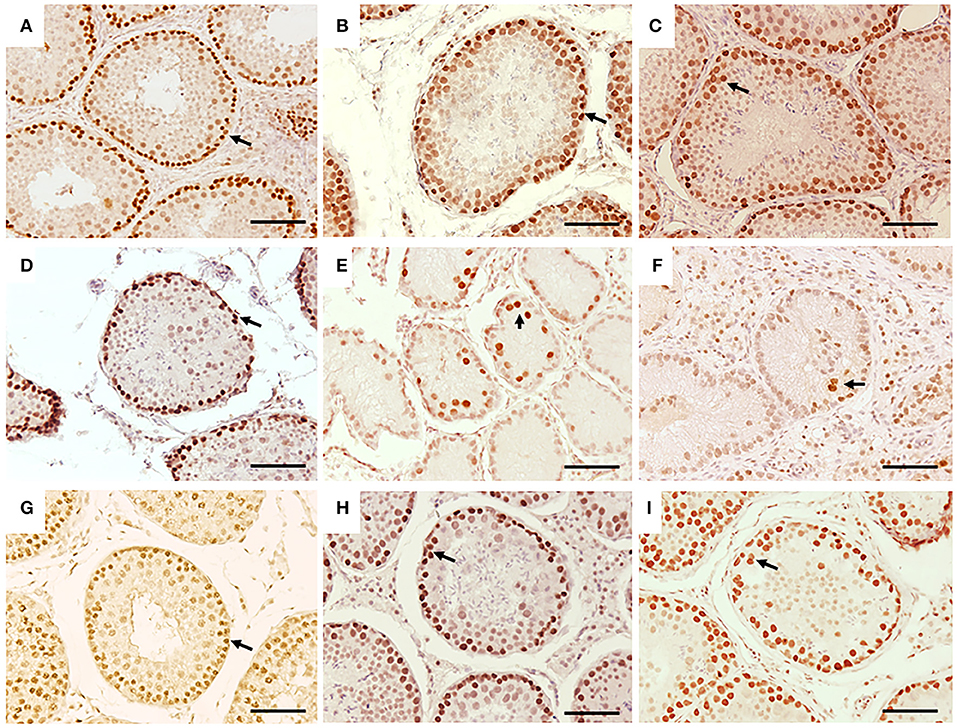
Figure 3. Immunohistochemistry for PCNA in seminiferous tubes of dogs from the control group (CTRL) (A–C); from cryptorchidism (CO) (D–F) and contralateral (CONTRA) (G–I) in the peripubertal stages (first column); young adult (second column) and senile (third column). A marked immunoreactivity (arrow) is observed in the testes of the CTRL animals unlike the CO, mainly in the young adult stages. The testes of the CONTRA group show an immunoreactivity that is neither as marked as in the CTRL nor as weak as in the CO. Bar scale: 50 μm.
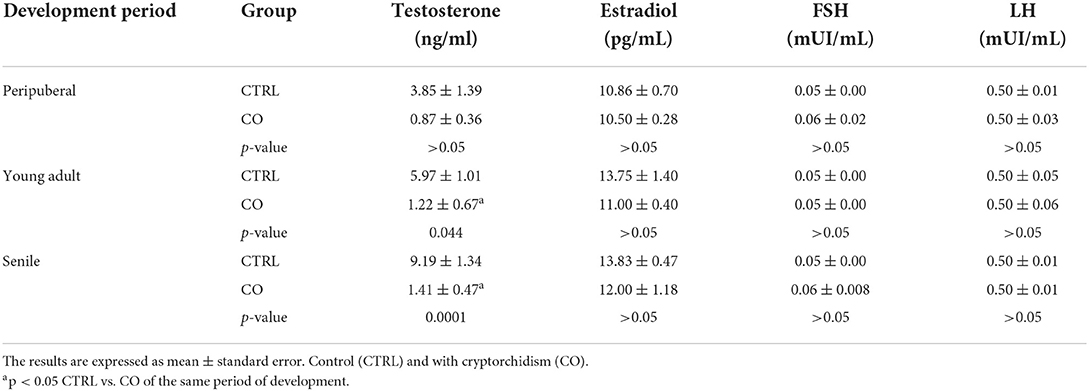
Table 2. Hormone concentrations in the control and cryptorchid groups at different developmental stages in the dog.
In the CO group, the contralateral testes showed seminiferous tubes with different stages of development, which go from gonocytes to round spermatids (Figure 1G). There were no differences in the variables studied when compared to the CTRL and CO groups (p > 0.05) (Table 1; Figure 3G).
In the CTRL, CO groups and the contralateral testes, Leydig cells were observed occupying the central spaces of the tubular interstitium. They were characterized by their polygonal shape of central nuclei that may present lipid droplets (Figures 4A,D,G).
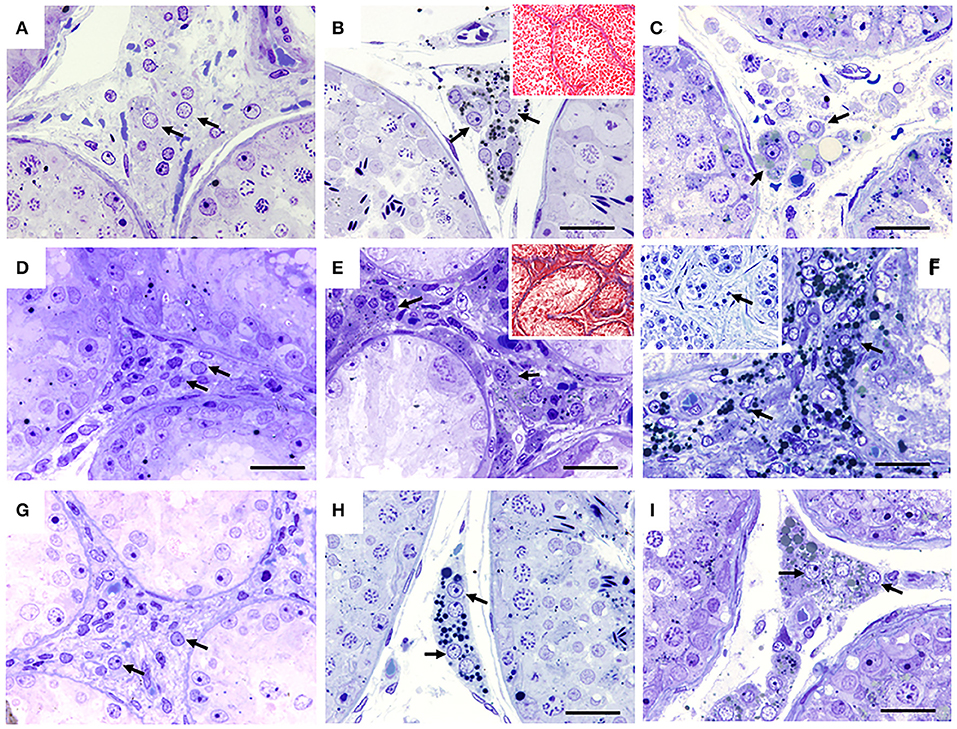
Figure 4. Seminiferous tubules of dogs from the control (CTRL) (A–C), cryptorchid (CO) (D–F) and contralateral (CONTRA) (G–I) groups at peripubertal (first column); young adult (second column) and senile (third column) stages. The interstitial spaces with their characteristic Leydig cells (arrow) are observed. The young adult and senile stage CO group showed small Leydig cell hyperplasia, when compared with the other groups. In some cases in the CO group in the senile stage, completely degenerated seminiferous tubes can be seen with some degenerating or absent Leydig cells (F insert). Toluidine Blue, except (B,C) Masson trichrome technique. Bar scale: 50 μm.
Stage 2: Young adult
The CTRL group complete spermatogenesis with slight histological alterations, such as vacuolization and desquamation, in the seminiferous epithelium were observed. The seminiferous tubules showed marked proliferation (Figure 3B), however, there was absence of gonocytes in all of them (Figures 1B, 2B).
In the CO group, a marked hypoplasia of germ cells was observed in the seminiferous epithelium, vacuolization, desquamation, folding and thickening of the basement membrane with cell degeneration and cytoplasmic extensions of Sertoli cells directed toward the center of the tubules (Figure 1E). Spermatogenesis did not continue beyond the spermatocytes, and in some specimens, no germ cells were observed. Sertoli-cell-only syndrome was identified in 50% of the animals (3/6) (Figure 1E insert). In addition, they depicted the presence of cells with gonocyte morphology in 16.6% of the specimens (1/6) (Figures 1E, 2E). The animals of the CO group presented a significant reduction of 3.67 times in the area of the seminiferous epithelium, de 2.70 times in MI and de 4.53 times in PI (Figure 3E) with a significant increase in HI of 4.38 times when compared with the CTRL group (Table 1).
At this age, there was a reduction of 4.89 times in testosterone concentrations in the CO group, without significant changes in estradiol, LH and FSH concentrations (p > 0.05, Table 2).
The TBARS technique showed a significant increase (p < 0.05) of 3.07 times in lipid peroxidation in CO testes when compared with the CTRL group (Table 1).
The contralateral testes did not show a significant difference in any of the variables studied, when compared to the CTRL and CO groups (Figures 1H, 3H).
In the CTRL groups and contralateral testes to CO, Leydig cells located in the central spaces of the tubular interstices were observed. In the CO group, a clear hyperplasia of this cell type was observed, interspersed between collagen fibers (Figures 4B,E). Leydig cells area were smaller compared to the other groups (Figures 4B,E insert, H).
Stage 3: Senile
In senile CTRL animals the seminiferous tubules presented complete spermatogenesis, slight thickening of the basement membrane, and evident cell proliferation (Figure 3C). However, 33.3% of the specimens (2/6) showed damaged histological features characterized by arrest of spermatogenesis, degenerating cells, thickening and folding of the basement membrane and abundant cell desquamation (Figures 1C insert, 2C) without the presence of gonocytes.
In the senile CO group, the alterations were similar to those observed in young adult CO animals, with arrest of spermatogenesis in spermatocyte stage and the presence of atypical cells (Figure 1F). In 16.6% of the animals (1/6), electron microscopy confirmed the presence of cells with persistent gonocyte morphology (Figure 2F) and in 16.6% (1/6), Sertoli-cell-only syndrome was appreciated. When compared with the CTRL group, a significant reduction in the seminiferous epithelium area of 4.30 times, MI of 3.13 times and proliferation of 2.81 times (Figure 3F) was determined. Contrarily, the HI registered a significant increase of 4.88 times (Table 1) while lipid peroxidation did not show significant difference (p > 0.05) (Table 1). Testosterone concentrations showed a significant reduction of 6.51 times with respect to the CTRL group. The estradiol, LH, and FSH did not present a significant difference (p > 0.05) (Table 2).
In terms of the contralateral testes to the retained one in the CO group (Figure 1I), there was a significant reduction in the area of the seminiferous epithelium of 3.16 times with respect to the CTRL group. On the other hand, the HI increased by 3.90 times with respect to the CTRL group. Histological changes consisted of germ cell hypoplasia, folding and thickening of the basement membrane, some tubes with arrested spermatogenesis in spermatocytes, and other sections of tubules with complete spermatogenesis. Moreover, a significant increase in MI of 2.45 times was appreciated when compared to the CO testes. The rest of the variables were found in a position between the CTRL and CO groups with no significant difference (p > 0.05, Figure 3I).
In all groups, the Leydig cells remained in the same location (Figure 4). However, in the CO group, the cells were observed with hyperplasia and smaller. In the cases where the seminiferous tubules showed clear atrophy, their Leydig cells were observed to be degenerated or absent (Figures 4C,F insert,I), interspersed between a large number of collagen fibers (Figures 4B,E insert).
In all stages of development, vimentin immunoreactivity was present in the cytoplasm of Sertoli cells, mainly perinuclear and in the basal compartment, forming linear columns toward the tubular lumen in the control group. In the CO group, the vimentin immunoreactivity was more evident, due to the absence of germ cells, showing the cytoplasmic extensions of the free Sertoli cells all along the tubular lumen. In the senile stage, organization of Sertoli cells was compromised. In the contralateral testicles, its distribution was similar to the control group (Figure 5).
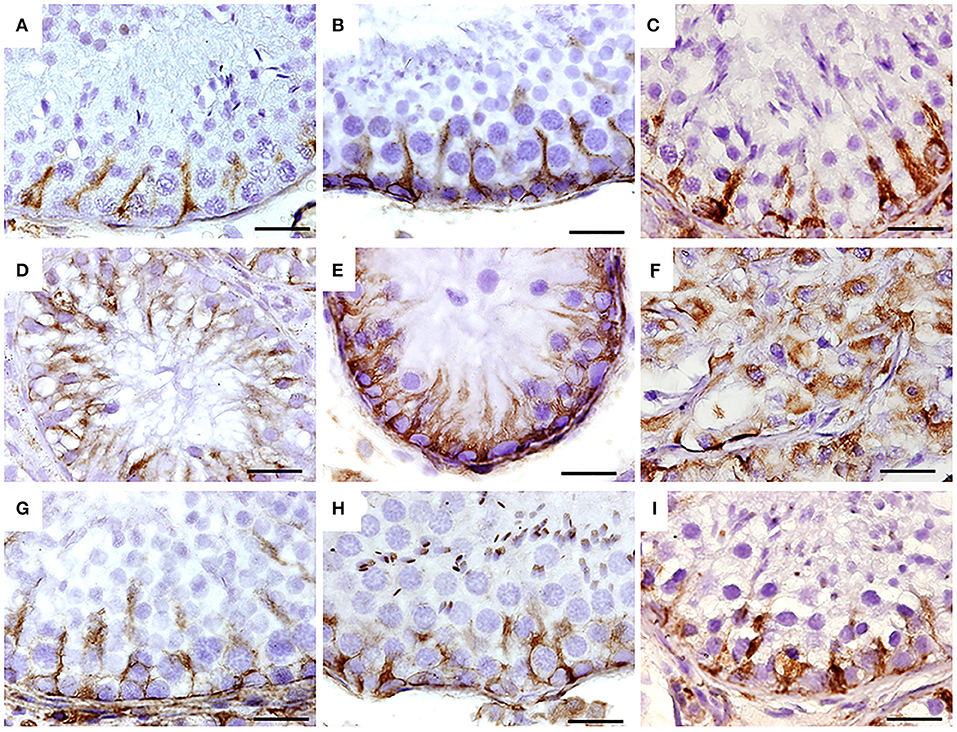
Figure 5. Immunohistochemistry for vimentin in seminiferous tubes of dogs from the control group (CTRL) (A–C); from cryptorchidism (CO) (D–F) and contralateral (CONTRA) (G–I) in the peripubertal stages (first column); young adult (second column) and senile (third column). A marked immunoreactivity is observed in all stages of development with variation in its distribution, mainly in animals with CO. Bar scale: 50 μm.
Discussion
Cryptorchidism occurs in dogs with high frequency (6.8%) (23) and shows anatomical and physiological similarities, as well as risk of infertility and development of neoplasia as in humans (26, 34). Therefore, this species can be considered as a suitable study model of CO.
In this study, the first seminiferous epithelium alterations in the dogs with CO appeared in the peripubertal stage. These alterations were decreased MI and delay in gonocyte differentiation to spermatogonia, evidenced by the presence of gonocytes in 50% of the specimens (3/6). These alterations generated a decrease in the number of germ cells. It has been reported in children with CO that the first histological alterations of the seminiferous epithelium begin, in subtle form, at 18 months of age with cell desquamation, folding of the basement membrane, vacuolization, lack of gonocyte differentiation to spermatogonia, with germ cell hypoplasia from the third year (35–37). Thus, at peripubertal stage, these alterations become evident. However, in this work, we did not include animals younger than 6 months of age. Hence, we were unable to determine the precise moment in which the first histological lesions appear in the pre-peripubertal stage.
In the young adult stage, alterations to the seminiferous epithelium generated by CO were evident, which was in accordance with what was reported for dogs (38, 39). These findings also coincide with what have been reported for seminiferous epithelium in humans where the thickening of the basement membrane was found (40, 41). In addition, various alterations such as germinal hypoplasia, vacuolization, desquamation, cell degeneration, presence of atypical cells, arrest in germ cell maturation (40, 42), reduction in cell proliferation (43), Sertoli-cell-only syndrome (40, 41) leading to the cessation of spermatogenesis with reduction in the MI and epithelial area have been found in humans with CO.
In our study, the presence of CO testes with Sertoli-cell-only syndrome in 22.2% of the dogs (4/18) support that this syndrome is particularly characteristic for men with a history of cryptorchidism (44). The presence of Sertoli-cell-only syndrome has also been reported in 7/10 dogs with CO in young adult animals (39), which is consistent with our results with 3/6 animals with CO in young adult status with this syndrome.
The arrest of spermatogenesis with a reduction in the area of the seminiferous epithelium, as well as in MI and cell proliferation with marked histological and ultrastructural alterations can be related to heat stress, which is known to affect the functioning of Sertoli cells in different species. The heat stress leads to downregulation in the expression of proteins of tight junctions (11, 45); thus, causing failures in spermatogenesis and cell desquamation (11, 45) as was observed in our study. Apoptotic pathways in germ cells can also be activated through redistribution of Bax protein from the cytoplasm to perinuclear of germ cells (46) and expression of the p53 mRNA (47). In this work, we observed in the CO group that spermatogenesis is arrested in spermatocytes. This is consistent with what was reported by Hirano et al. (48) who showed that pachytene spermatocytes are the most sensitive cells to temperature increases, generating DNA double-strand breaks and asynapsis, which leads to discontinuation of meiosis. Consequently, the cells are eliminated by apoptosis.
The effect of heat stress is likely due to oxidative stress generated by the production of superoxide anion, hydroxyl radical, nitric oxide and hydrogen peroxide (49–52). This phenomenon can be determined by increased malondialdehyde, the final product of lipid peroxidation (53) and which leads to the degeneration of germ cells (13). The increase in this compound observed in our animals with CO has also been seen in children with CO (54, 55). Oxidative stress in the CO patient has been shown to participate in germ cell damage (54, 55). Moreover, oxidative stress can be generated by the inactivation of the antioxidant enzymes Cu/Zn superoxide dismutase (SOD) and catalase, demonstrated in animal models (56, 57). Low SOD enzyme activity has also been found in dogs with CO (58).
These alterations are possibly owing to the fact that spermatocytes and spermatids are the germ cells most vulnerable to damage caused by reactive oxygen species because, even though they are capable of converting super oxide anion to hydrogen peroxide, they have difficulty metabolizing peroxide, which saturates its protective system against peroxide that becomes a highly toxic hydroxyl radical (59) when compared to spermatogonia or Sertoli cells. This coincides with what was observed in this work.
From young adult to senile development, CO determined histological alterations remained constant, well above the CTRL group, despite the fact that 33.3% of the CTRL animals (2/6) in this study also were experiencing alterations. None of the control animals showed testicular tumor development. There is controversy about the development of testicular tumor in senile dogs. Some authors mentioned a correlation with tumor development (60) while others assumed a contrary position (61). Seminiferous tubes without germ cells are reported in humans with unilateral and bilateral undescended testicle until old age (62) who, in some cases, did not develop testicular tumor (62).
The presence of cells with morphological and ultrastructural characteristics of gonocytes in 16.6% of the animals (2/12) in the CO dogs in the young and senile stages is uncommon. Gonocytes are not expected to be present in these stages of development; although the presence of gonocytes has been reported in 2/10 young dogs with CO (39). Hence, this event may suggest that gonocytes predisposes the development of testicular tumor (9, 15). It has been reported that in dogs with CO, the main types of testicular tumors that can develop are seminoma and mixed germ cell-sex cord stromal cell tumor, both with germ cell components, and to a lesser extent Sertoli cell tumor and interstitial cell tumor (26).
This opinion is concurrent with the proposition that the failure in the differentiation of the gonocyte to spermatogonia may be the cause of the precursor lesion of GCNIS (9, 15, 19, 63) reported that 5.4% of patients human with CO have germ cells with immunohistochemical characteristics of gonocytes.
Regarding the endocrine behavior of the patient with CO, some authors mentioned a decrease in testosterone concentrations (2, 64), while others found it unchanged (65, 66).
This reduction in testosterone concentrations in boys with cryptorchidism suggests a malfunction in the Leydig cells or the hypothalamic-pituitary-gonadal axis (67–69). Such event may alter the period of “minipuberty,” in which some authors indicate that gonocyte differentiation takes place (19, 70). However, the role of gonadotropins and testicular hormones in this process remains to be clarified (70). In CO dogs, low concentrations of testosterone have been reported from prepubertal to senile stages (71–73), as it was observed in this work, which leads to altered spermatogenesis. Leydig cell hyperplasia in the animals of CO group has been previously described in dogs with CO (30) and coincides with what has been reported for humans with this condition (74). Controverselly, in the current study we observed that the Leydig cells presented a smaller size and signs of degeneration despite being present in large numbers. A reduction in Leydig cell function has been shown in CO testes in rats (75) and humans (76), probably due to depletion of the smooth endoplasmic reticulum as it was described in humans (74).
In humans, 6.33% of patients with unilateral CO present contralateral damage, which was attributed to the impact of this pathology on both gonads (38, 77). In the contralateral testes, the reduction in the area of the seminiferous epithelium when compared with CTRL group and the increase in MI when compared to the CO group suggest that the contralateral position conditions testicular damage. The level of damage places the contralateral testes between the CTRL and CO groups as reported by Kawakami et al. (78) in dogs. However, some authors mentioned that this may be caused by deficient cooling system in the venous and arterial circulation between both gonads, thus, constant heat stress is maintained (75). In any case, in this study, no difference in lipid peroxidation was observed when compared to the CTRL and CO groups. Hence, we suggest that the mechanism of damage may be through another pathway such as Sertoli cell dysfunction determined by low concentrations of transferrin, which has been associated with low concentrations of testosterone, and which has been reported in the contralateral testis of dogs with surgically induced CO (78).
Greater intensity and disorganization of vimentin immunoreactivity have been reported in rats and humans with CO (79–81) and in animals exposed to oxidative stress (81, 82). Although Pecile et al. (83) do not report changes in vimentin immunoreactivity in the testes of dogs with CO, their images show a loss in the linear organization of immunoreactivity to this intermediate filament (83). Damage and restoration of spermatogenesis are related to the disintegration or recovery of these filaments (84).
The results of this work demonstrate that the testicular development of the dog with CO is similar to that of the human with this pathology, provoking alterations of the seminiferous epithelium that lead to problems in fertility and possible risk of developing neoplasia. Therefore, we suggest that this species could be used as a suitable model for the study of CO. This may allow research for compounds that would reduce fertility problems and the risk of developing testicular tumor, as well as provide early risk markers for the development of GCNIS in patients with CO.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by INP: 54/2020. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
Experiments were conceived and designed and the paper was written by RMV-V, JCR-C, and NH-J. Experiments were performed by NH-J, ER-C, and FR-D. Data were analyzed by RMV-V, NH-J, JCR-C, DL-H, MJ-M, RR-R, and AM. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Instituto Nacional de Pediatria (Grant Number INP: 54/2020).
Acknowledgments
We express our gratitude to the Mobile Dog and Cat Sterilization Unit of Alvaro Obregon Borough Office, and to UNAM-Banfield Veterinary Hospital, CDMX. In addition, our thanks go to Pedro Medina Granados and Edna García Cruz for their technical support in processing the samples. In the same, our gratitude to CONACyT for granting the doctoral scholarship to NH-J (Student: 297349) to carry out the research and publication of this article, which is required for her doctorate degree.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Radcliffe J. Hospital Cryptorchidism Study Group. Cryptorchidism: a prospective study of 7500 consecutive male births, 1984-8. John Radcliffe Hospital Cryptorchidism Study Group. Arch Dis Child. (1992) 67:892–9. doi: 10.1136/adc.67.7.892
2. Lee PA, Coughlin MT. Fertility after bilateral cryptorchidism. evaluation by paternity, hormone, and semen data. Horm Res. (2001) 55:28–32. doi: 10.1159/000049960
3. Ghirri P, Ciulli C, Vuerich M, Cuttano A, Faraoni M, Guerrini L, et al. Incidence at birth and natural history of cryptorchidism: a study of 10,730 consecutive male infants. J Endocrinol Invest. (2002) 25:709–15. doi: 10.1007/BF03345105
4. Sijstermans K, Hack WW, Meijer RW, van der Voort-Doedens LM. The frequency of undescended testis from birth to adulthood: a review. Int J Androl. (2008) 31:1–11. doi: 10.1111/j.1365-2605.2007.00770.x
5. Virtanen HE, Toppari J. Epidemiology and pathogenesis of cryptorchidism. Hum Reprod Update. (2008) 14:49–58. doi: 10.1093/humupd/dmm027
6. Mitsui T. Effects of the prenatal environment on cryptorchidism: a narrative review. Int J Urol. (2021) 28:882–9. doi: 10.1111/iju.14600
7. Giwercman A, Grindsted J, Hansen B, Jensen OM, Skakkebaek NE. Testicular cancer risk in boys with maldescended testis: a cohort study. J Urol. (1987) 138:1214–6. doi: 10.1016/s0022-5347[17]43553-1
8. Piltoft JS, Larsen SB, Dalton SO, Johansen C, Baker JL, Cederkvist L, et al. Early life risk factors for testicular cancer: a case-cohort study based on the Copenhagen School Health Records Register. Acta Oncol. (2017) 56:220–4. doi: 10.1080/0284186X.2016.1266085
9. Skakkebaek NE, Berthelsen JG, Giwercman A, Müller J. Carcinoma-in-situ of the testis: possible origin from gonocytes and precursor of all types of germ cell tumours except spermatocytoma. Int J Androl. (1987) 10:19–28. doi: 10.1111/j.1365-2605.1987.tb00161.x
10. Hadziselimovic F, Hoecht B. Testicular histology related to fertility outcome and postpubertal hormone status in cryptorchidism. Klin Padiatr. (2008) 220:302–7. doi: 10.1055/s-2007-993194
11. Yang WR, Li BB, Hu Y, Zhang L, Wang XZ. Oxidative stress mediates heat-induced changes of tight junction proteins in porcine sertoli cells via inhibiting CaMKKβ-AMPK pathway. Theriogenology. (2020) 142:104. doi: 10.1016/j.theriogenology.2019.09.031
12. He C, Sun J, Yang D, He W, Wang J, Qin D, et al. Nrf2 activation mediates the protection of mouse Sertoli Cells damage under acute heat stress conditions. Theriogenology. (2022) 177:183–94. doi: 10.1016/j.theriogenology.2021.10.009
13. Vigueras-Villaseñor RM Ojeda I Gutierrez-Pérez O Chavez-Saldaña M Cuevas O Maria DS . Protective effect of α-tocopherol on damage to rat testes by experimental cryptorchidism. Int J Exp Pathol. (2011) 92:131–9. doi: 10.1111/j.1365-2613.2010.00757.x
14. Rørth M, Rajpert-De Meyts E, Andersson L, Dieckmann KP, Fosså SD, Grigor KM, et al. Carcinoma in situ in the testis. Scand J Urol Nephrol Suppl. (2000) 205:166–86. doi: 10.1080/00365590050509896
15. Hutson JM Li R, Southwell BR, Petersen BL, Thorup J, Cortes D. Germ cell development in the postnatal testis: the key to prevent malignancy in cryptorchidism? Front Endocrinol (Lausanne). (2013) 3:176. doi: 10.3389/fendo.2012.00176
16. Hamilton-Dutoit SJ, Lou H, Pallesen G. The expression of placental alkaline phosphatase (PLAP) and PLAP-like enzymes in normal and neoplastic human tissues. an immunohistological survey using monoclonal antibodies. APMIS. (1990) 98:797–811. doi: 10.1111/j.1699-0463.1990.tb05000.x
17. Huff DS, Hadziselimovic F, Snyder HM, Blythe B, Ducket JW. Histologic maldevelopment of unilaterally cryptorchid testes and their descended partners. Eur J Pediatr. (1993) 152:S11-4.
18. Culty M. Gonocytes, the forgotten cells of the germ cell lineage. Birth Defects Res C Embryo Today. (2009) 87:1–26. doi: 10.1002/bdrc.20142
19. Manku G, Culty M. Mammalian gonocyte and spermatogonia differentiation: recent advances and remaining challenges. Reproduction. (2015) 149:R139–57. doi: 10.1530/REP-14-0431
20. Batool A, Karimi N, Wu XN, Chen SR, Liu YX. Testicular germ cell tumor: a comprehensive review. Cell Mol Life Sci. (2019) 76:1713–27. doi: 10.1007/s00018-019-03022-7
21. Zhang RD, Wen XH, Kong LS, Deng XZ, Peng B, Huang AP, et al. A quantitative (stereological) study of the effects of experimental unilateral cryptorchidism and subsequent orchiopexy on spermatogenesis in adult rabbit testis. Reproduction. (2002) 124:95–105. doi: 10.1530/rep.0.1240095
22. Picut CA, Ziejewski MK, Stanislaus D. Comparative aspects of pre- and postnatal development of the male reproductive system. Birth Defects Res. (2018) 110:190–227. doi: 10.1002/bdr2.1133
23. Yates D, Hayes G, Heffernan M, Beynon R. Incidence of cryptorchidism in dogs and cats. Vet Rec. (2003) 152:502–4. doi: 10.1136/vr.152.16.502
24. Tannouz VGS, Mamprim MJ, Lopes MD, Santos-Sousa CA, Souza Junior P, Babinski MA, et al. Is the right testis more affected by cryptorchidism than the left testis? an ultrasonographic approach in dogs of different sizes and breeds. Folia Morphol (Warsz). (2019) 78:847–52. doi: 10.5603/FM.a2019.0022
25. Hayes HM Jr, Pendergrass TW. Canine testicular tumors: epidemiologic features of 410 dogs. Int J Cancer. (1976) 18:482–7. doi: 10.1002/ijc.2910180413
26. Liao AT, Chu PY, Yeh LS, Lin CT, Liu CH. A 12-year retrospective study of canine testicular tumors. J Vet Med Sci. (2009) 71:919–23. doi: 10.1292/jvms.71.919
27. Grieco V, Riccardi E, Greppi GF, Teruzzi F, Iermanò V, Finazzi M. Canine testicular tumours: a study on 232 dogs. J Comp Pathol. (2008) 138:86–9. doi: 10.1016/j.jcpa.2007.11.002
28. Kawakami E, Tsutsui T, Ogasa A. Histological observations of the reproductive organs of the male dog from birth to sexual maturity. J Vet Med Sci. (1991) 53:241–8. doi: 10.1292/jvms.53.241
29. Taha MB, Noakes DE. The effect of age and season of the year on testicular function in the dog, as determined by histological examination of the seminiferous tubules and the estimation of peripheral plasma testosterone concentrations. J Small Anim. Pract. (1982) 23:351–7. doi: 10.1111/j.1748-5827.1982.tb01677.x
30. Merz SE, Klopfleisch R, Breithaupt A, Gruber AD. Aging and senescence in canine testes. Vet Pathol. (2019) 56:715–24. doi: 10.1177/0300985819843683
31. Johnsen SG. Testicular biopsy score count-a method for registration of spermatogenesis in human testes: normal values and results in 335 hypogonadal males. Hormones. (1970) 1:2–25. doi: 10.1159/000178170
32. Vigueras-Villaseñor RM Molina-Ortiz D Reyes-Torres G del Angel DS Moreno-Mendoza NA Cruz ME . Effect of allopurinol on damage caused by free radicals to cryptorchid testes. Acta Histochem. (2009) 111:127–37. doi: 10.1016/j.acthis.2008.05.004
33. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. (1951) 93:265–75.
34. Ortega-Pacheco A, Rodríguez-Buenfil JC, Segura-Correa JC, Bolio-Gonzalez ME, Jiménez-Coello M, Linde Forsberg C. Pathological conditions of the reproductive organs of male stray dogs in the tropics: prevalence, risk factors, morphological findings and testosterone concentrations. Reprod Domest Anim. (2006) 41:429–37. doi: 10.1111/j.1439-0531.2006.00688.x
35. Mengel W, Wronecki K, Schroeder J, Zimmermann FA. Histopathology of the cryptorchid testis. Urol Clin North Am. (1982) 9:331-8.
36. Huff DS, Hadziselimovic F, Snyder HM 3rd, Duckett JW, Keating MA. Postnatal testicular maldevelopment in unilateral cryptorchidism. J Urol. (1989) 142:546–8. doi: 10.1016/s0022-5347(17)38811-0
37. Verkauskas G, Malcius D, Dasevicius D, Hadziselimovic F. Histopathology of Unilateral Cryptorchidism. Pediatr Dev Pathol. (2019) 22:53–8. doi: 10.1177/1093526618789300
38. Mattos FMR, Simoes-Mattos L, Domingues SSF. Cryptorchidism in dog. Ciencia Animal. (2000) 10:61–70.
39. Veronesi MC, Riccardi E, Rota A, Grieco V. Characteristics of cryptic/ectopic and contralateral scrotal testes in dogs between 1 and 2 years of age. Theriogenology. (2009) 72:969–77. doi: 10.1016/j.theriogenology.2009.06.016
40. Amat P, Paniagua R, Montero J. Seminiferous tubule degeneration in human cryptorchid testes. J Androl. (1985) 6:1–9. doi: 10.1002/j.1939-4640.1985.tb00810.x
41. Govender D, Sing Y, Chetty R. Sertoli cell nodules in the undescended testis: a histochemical, immunohistochemical, and ultrastructural study of hyaline deposits. J Clin Pathol. (2004) 57:802–6. doi: 10.1136/jcp.2004.015982
42. Müller J, Skakkebaek NE, Nielsen OH, Graem N. Cryptorchidism and testis cancer. Atypical infantile germ cells followed by carcinoma in situ and invasive carcinoma in adulthood. Cancer. (1984) 54:629–34. doi: 10.1002/1097-0142(1984)54:4 <629::aid-cncr2820540407>3.0.co;2-e
43. Ofordeme KG, Aslan AR, Nazir TM, Hayner-Buchan A, Kogan BA. Apoptosis and proliferation in human undescended testes. BJU Int. (2005) 96:634–8. doi: 10.1111/j.1464-410X.2005.05698.x
44. Fedder J. History of cryptorchidism and ejaculate volume as simple predictors for the presence of testicular sperm. Syst Biol Reprod Med. (2011) 57:154–61. doi: 10.3109/19396368.2010.550796
45. Cai H, Ren Y, Li XX, Yang JL, Zhang CP, Chen M, et al. Scrotal heat stress causes a transient alteration in tight junctions and induction of TGF-β expression. Int J Androl. (2011) 34:352–62. doi: 10.1111/j.1365-2605.2010.01089.x
46. Yamamoto CM, Sinha Hikim AP, Huynh PN, Shapiro B, Lue Y, Salameh WA, et al. Redistribution of Bax is an early step in an apoptotic pathway leading to germ cell death in rats, triggered by mild testicular hyperthermia. Biol Reprod. (2000) 63:1683–90. doi: 10.1095/biolreprod63.6.1683
47. Absalan F, Movahedin M, Mowla SJ. Germ cell apoptosis induced by experimental cryptorchidism is mediated by molecular pathways in mouse testis. Andrologia. (2010) 42:5–12. doi: 10.1111/j.1439-0272.2009.00947.x
48. Hirano K, Nonami Y, Nakamura Y, Sato T, Sato T, Ishiguro KI, et al. Temperature sensitivity of DNA double-strand break repair underpins heat-induced meiotic failure in mouse spermatogenesis. Commun Biol. (2022) 5:504. doi: 10.1038/s42003-022-03449-y
49. Zini A, Schlegel PN. Cu/Zn superoxide dismutase, catalase and glutathione peroxidase mRNA expression in the rat testis after surgical cryptorchidism and efferent duct ligation. J Urol. (1997) 158:659–63. doi: 10.1016/S0022-5347(01)64578-6
50. Ikeda M, Kodama H, Fukuda J, Shimizu Y, Murata M, Kumagai J, et al. Role of radical oxygen species in rat testicular germ cell apoptosis induced by heat stress. Biol Reprod. (1999) 61:393–9. doi: 10.1095/biolreprod61.2.393
51. Kumagai A, Kodama H, Kumagai J, Fukuda J, Kawamura K, Tanikawa H, et al. Xanthine oxidase inhibitors suppress testicular germ cell apoptosis induced by experimental cryptorchidism. Mol Hum Reprod. (2002) 8:118–23. doi: 10.1093/molehr/8.2.118
52. Ishii T, Matsuki S, Iuchi Y, Okada F, Toyosaki S, Tomita Y, et al. Accelerated impairment of spermatogenic cells in SOD1-knockout mice under heat stress. Free Radic Res. (2005) 39:697–705. doi: 10.1080/10715760500130517
53. Gasparovic AC, Jaganjac M, Mihaljevic B, Sunjic SB, Zarkovic N. Assays for the measurement of lipid peroxidation. Methods Mol Biol. (2013) 965:283–96. doi: 10.1007/978-1-62703-239-1_19
54. Durairajanayagam D, Agarwal A, Ong C. Causes, effects and molecular mechanisms of testicular heat stress. Reprod Biomed Online. (2015) 30:14–27. doi: 10.1016/j.rbmo.2014.09.018
55. Imamoglu M, Bülbül SS, Kaklikkaya N, Sarihan H. Oxidative, inflammatory and immunologic status in children with undescended testes. Pediatr Int. (2012) 54:816–9. doi: 10.1111/j.1442-200X.2012.03695.x
56. Ahotupa M, Huhtaniemi I. Impaired detoxification of reactive oxygen and consequent oxidative stress in experimentally cryptorchid rat testis. Biol Reprod. (1992) 46:1114–8. doi: 10.1095/biolreprod46.6.1114
57. Peltola V, Huhtaniemi I, Ahotupa M. Abdominal position of the rat testis is associated with high level of lipid peroxidation. Biol Reprod. (1995) 53:1146–50. doi: 10.1095/biolreprod53.5.1146
58. Kawakami E, Hirano T, Hori T, Tsutsui T. Testicular superoxide dismutase activity, heat shock protein 70 concentration and blood plasma inhibin-alpha concentration of dogs with a Sertoli cell tumor in a unilateral cryptorchid testis. J Vet Med Sci. (2007) 69:1259–62. doi: 10.1292/jvms.69.1259
59. Bauché F, Fouchard MH, Jégou B. Antioxidant system in rat testicular cells. FEBS Lett. (1994) 349:392–6. doi: 10.1016/0014-5793(94)00709-8
60. Mosier JE. Effect of aging on body systems of the dog. Vet Clin North Am Small Anim Pract. (1989) 19:1–12. doi: 10.1016/s0195-5616(89)50001-9
61. Peters MA, de Rooij DG, Teerds KJ, van Der Gaag I, van Sluijs FJ. Spermatogenesis and testicular tumours in ageing dogs. J Reprod Fertil. (2000) 120:443–52.
62. Kassir R, Dubois J, Berremila SA, Baccot S, Boueil-Bourlier A, Tiffet O, et al. rare variant of inguinal hernia: Cryptorchid testis at the age of 50 years. etiopathogenicity, prognosis and management. Int J Surg Case Rep. (2014) 5:416–8. doi: 10.1016/j.ijscr.2014.03.015
63. Vigueras-Villaseñor RM Cortés-Trujillo L Chávez-Saldaña M Vázquez FG Carrasco-Daza D Cuevas-Alpuche O . Analysis of POU5F1, c-Kit, PLAP, AP2γ and SALL4 in gonocytes of patients with cryptorchidism. Acta Histochem. (2015) 117:752–61. doi: 10.1016/j.acthis.2015.08.004
64. Baker BA, Morley R, Lucas A. Plasma testosterone in preterm infants with cryptorchidism. Arch Dis Child. (1988) 63:1198–200. doi: 10.1136/adc.63.10.1198
65. Barthold JS, Manson J, Regan V, Si X, Hassink SG, Coughlin MT, et al. Reproductive hormone levels in infants with cryptorchidism during postnatal activation of the pituitary-testicular axis. J Urol. (2004) 172:1736–41. doi: 10.1097/01.ju.0000138523.24337.be
66. Iwatsuki S, Kojima Y, Mizuno K, Kamisawa H, Umemoto Y, Sasaki S, et al. Endocrine assessment of prepubertal boys with a history of cryptorchidism and/or hypospadias: a pilot study. J Urol. (2011) 185:2444–50. doi: 10.1016/j.juro.2011.01.018
67. Suomi AM, Main KM, Kaleva M, Schmidt IM, Chellakooty M, Virtanen HE, et al. Hormonal changes in 3-month-old cryptorchid boys. J Clin Endocrinol Metab. (2006) 91:953–8. doi: 10.1210/jc.2004-2318
68. Bay K, Virtanen HE, Hartung S, Ivell R, Main KM, Skakkebaek NE, et al. Nordic Cryptorchidism Study Group, Toppari J. Insulin-like factor 3 levels in cord blood and serum from children: effects of age, postnatal hypothalamic-pituitary-gonadal axis activation, and cryptorchidism. J Clin Endocrinol Metab. (2007) 92:4020–7. doi: 10.1210/jc.2007-0974
69. Hamdi SM, Almont T, Galinier P, Mieusset R, Thonneau P. Altered secretion of Sertoli cells hormones in 2-year-old prepubertal cryptorchid boys: a cross-sectional study. Andrology. (2017) 5:783–9. doi: 10.1111/andr.12373
70. Becker M, Hesse V. Minipuberty: why does it happen? Horm Res Paediatr. (2020) 93:76–84. doi: 10.1159/000508329
71. Pathirana IN, Yamasaki H, Kawate N, Tsuji M, Büllesbach EE, Takahashi M, et al. Plasma insulin-like peptide 3 and testosterone concentrations in male dogs: changes with age and effects of cryptorchidism. Theriogenology. (2012) 77:550–7. doi: 10.1016/j.theriogenology.2011.08.030
72. Hornakova L, Vrbovska T. Pavl'ak M, Valencakova-Agyagosova A, Halo M, Hajurka J. The evaluation of blood concentrations of testosterone, 17β-oestradiol and anti-Mu¨llerian hormone in dogs with cryptorchidism and testicular tumours. Pol J Vet Sci. (2017) 20:677–85. doi: 10.1515/pjvs-2017-0085
73. Kawakami E, Tsutsui T, Saito S, Kakimoto T, Ogasa A. Changes in peripheral plasma luteinizing hormone and testosterone concentrations and semen quality in normal and cryptorchid dogs during sexual maturation. Lab Anim Sci. (1995) 45:258–63.
74. Gotoh M, Miyake K, Mitsuya H. A study of cryptorchidism. I. Light and electron microscopic study of Leydig's cells in the testes of cryptorchid patients. Hinyokika Kiyo. (1984)3:327–47.
75. Aldahhan RA, Stanton PG. Heat stress response of somatic cells in the testis. Mol Cell Endocrinol. (2021) 527:111216. doi: 10.1016/j.mce.2021.111216
76. Regadera J, Martínez-García F, González-Peramato P, Serrano A, Nistal M, Suárez-Quian C. Androgen receptor expression in sertoli cells as a function of seminiferous tubule maturation in the human cryptorchid testis. J Clin Endocrinol Metab. (2001) 86:41321. doi: 10.1210/jcem.86.1.7109
77. Akre O, Pettersson A, Richiardi L. Risk of contralateral testicular cancer among men with unilaterally undescended testis: a mta analysis. Int J Cancer. (2009) 124:687–9. doi: 10.1002/ijc.23936
78. Kawakami E, Hori T, Tsutsui T. Function of contralateral testis after artificial unilateral cryptorchidism in dogs. J Vet Med Sci. (1999) 61:1107–11. doi: 10.1292/jvms.61.1107
79. Abd-El-Hafez MA, El-Shafee MD, Omar SH, Aburahma AA, Kamar SS. The ameliorative effect of curcumin on cryptorchid and non-cryptorchid testes in induced unilateral cryptorchidism in albino rat: histological evaluation. Folia Morphol (Warsz). (2021) 80:596–604. doi: 10.5603/FM.a2020.0084
80. Rogatsch H, Jezek D, Hittmair A, Mikuz G, Feichtinger H. Expression of vimentin, cytokeratin, and desmin in Sertoli cells of human fetal, cryptorchid, and tumour-adjacent testicular tissue. Virchows Arch. (1996) 427:497–502. doi: 10.1007/BF00199510
81. Mohammed M, Elfadeel KA, Abdel-Aziz Fawzy A, Samy W. The effect of chronic stress on the testis of adult albino Rats and the possible protective effect of astaxanthin supplementation (Histological, immunohistochemical and biochemical studies). J Med Histol. (2019) 2:115–30. doi: 10.21608/jmh.2019.7416.1048
82. Liu F, Li XL, Lin T, He DW, Wei GH, Liu JH, et al. The cyclophosphamide metabolite, acrolein, induces cytoskeletal changes and oxidative stress in Sertoli cells. Mol Biol Reprod. (2012) 39:493–500. doi: 10.1007/s11033-011-0763-9
83. Pecile A, Groppetti D, Pizzi G, Banco B, Bronzo V, Giudice C, et al. Immunohistochemical insights into a hidden pathology: canine cryptorchidism. Theriogenology. (2021) 176:43–53. doi: 10.1016/j.theriogenology.2021.09.011
Keywords: cryptorchidism, testis, dog, human, seminiferous epithelium, Leydig cell, vimentin
Citation: Hernández-Jardón N, Rojas-Castañeda JC, Landero-Huerta D, Reyes-Cruz E, Reynoso-Robles R, Juárez-Mosqueda MdL, Medrano A, Reyes-Delgado F and Vigueras-Villaseñor RM (2022) Cryptorchidism: The dog as a study model. Front. Vet. Sci. 9:935307. doi: 10.3389/fvets.2022.935307
Received: 03 May 2022; Accepted: 22 August 2022;
Published: 13 September 2022.
Edited by:
Peter DOVC, University of Ljubljana, SloveniaReviewed by:
Rita Payan Carreira, University of Evora, PortugalAlessandra Pelagalli, University of Naples Federico II, Italy
Caterina Squillacioti, University of Naples Federico II, Italy
Copyright © 2022 Hernández-Jardón, Rojas-Castañeda, Landero-Huerta, Reyes-Cruz, Reynoso-Robles, Juárez-Mosqueda, Medrano, Reyes-Delgado and Vigueras-Villaseñor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosa María Vigueras-Villaseñor, cm12aWd1ZXJhc0B5YWhvby5jb20ubXg=; Julio César Rojas-Castañeda, cm9jYWpjMUB5YWhvby5jb20ubXg=