
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci., 05 December 2022
Sec. Veterinary Infectious Diseases
Volume 9 - 2022 | https://doi.org/10.3389/fvets.2022.1071097
This article is part of the Research TopicGenomics And Proteomics Approaches for The Improved Biologics Against Infectious AgentsView all 6 articles
 Siddiq Ur Rahman1*
Siddiq Ur Rahman1* Hassan Ur Rehman1Inayat Ur Rahman2
Hassan Ur Rehman1Inayat Ur Rahman2 Abdur Rauf3
Abdur Rauf3 Abdulrahman Alshammari4
Abdulrahman Alshammari4 Metab Alharbi4Noor ul Haq1
Metab Alharbi4Noor ul Haq1 Hafiz Ansar Rasul Suleria5
Hafiz Ansar Rasul Suleria5 Sayed Haidar Abbas Raza6,7
Sayed Haidar Abbas Raza6,7Lumpy skin disease virus (LSDV) causes lumpy skin disease (LSD) in livestock, which is a double-stranded DNA virus that belongs to the genus Capripoxvirus of the family Poxviridae. LSDV is an important poxvirus that has spread out far and wide to become distributed worldwide. It poses serious health risks to the host and causes considerable negative socioeconomic impact on farmers financially and on cattle by causing ruminant-related diseases. Previous studies explained the population structure of the LSDV within the evolutionary time scale and adaptive evolution. However, it is still unknown and remains enigmatic as to how synonymous codons are used by the LSDV. Here, we used 53 LSDV strains and applied the codon usage bias (CUB) analysis to them. Both the base content and the relative synonymous codon usage (RSCU) analysis revealed that the AT-ended codons were more frequently used in the genome of LSDV. Further low codon usage bias was calculated from the effective number of codons (ENC) value. The neutrality plot analysis suggested that the dominant factor of natural selection played a role in the structuring of CUB in LSDV. Additionally, the results from a comparative analysis suggested that the LSDV has adapted host-specific codon usage patterns to sustain successful replication and transmission chains within hosts (Bos taurus and Homo sapiens). Both natural selection and mutational pressure have an impact on the codon usage patterns of the protein-coding genes in LSDV. This study is important because it has characterized the codon usage pattern in the LSDV genomes and has provided the necessary data for a basic evolutionary study on them.
Lumpy skin disease virus (LSDV) causes lumpy skin disease (LSD), which is a viral disease affecting the ruminants, while the virus itself belongs to the genus Capripoxvirus of the family Poxviridae (1). This disease assumes economic significance due to the associated financial destruction to farmers it brings along and has a genome that consists of the double-stranded DNA (2). It is genetically associated with the sheeppox and goatpox virus family Poxviridae. The lumpy skin disease causing LSDV is transmitted to animals through blood-sucking arthropods while infecting a human being through direct transmission without the need for a vector (3, 4).
Lumpy skin disease is a viral disease predominantly attacking the bovine population, which presents a plethora of specific clinical signs ranging from subclinical infection to death (4). The main clinical signs and symptoms predictive of this disease are fever, mouth lesions, pharynx, nodules' appearance in the skin, skin edema, and enlargement of the superficial lymph nodes (3–5). The affected livestock instantly start losing weight and produce less milk. However, in the severest of severe cases, the livestock even die from an LSDV infection (1). LSDV gets distributed so widely that it has pervaded almost the entire globe and consequently thereof has set off a wave of serious economic problems for the affected countries worldwide. LSDV was first identified and reported in cattle in an outbreak in Zambia in 1929 (5), and the genome of the LSDV appeared to remain stable there for many years. Only minor genetic changes were noticed between the LSDV field isolates that were recovered over many years in Africa. However, the first outbreaks of this deadly disease surfaced in Egypt in 1988, which further spread to the Middle East in 2012 and later to Europe in 2015. The incidence of the disease has been further identified in a country as far as Russia, prevailing there from 2017 to 2019 (6). Recently, the disease has started wrecking havoc in Asia too, especially in Pakistan, where the prevalence of LSD has been steadily increasing; from 2020 until now, five million dairy farmers have suffered huge losses in the aftermath of LSD's attack on crops (7). Previous studies suggested that LSDV transmission spreads across the countries when the infected cattle are moved from one place to another or by vectors in animal trucks (6). Therefore, it is essential to use genomic analysis based on the knowledge of the distributing LSDV strains while considering the increasing diversity of LSDV in recent years.
Codon usage bias (CUB) refers to a phenomenon in which synonymous codons are not used with an equal frequency during gene translation. CUB is a common phenomenon that is observed in numerous species, including prokaryotes, eukaryotes, and viruses (8, 9). We noticed that a variety of factors affect how codons are used by different organisms. The primary explanations put forth for this variance in codon usage among the genes in these species are attributed to be due to weak natural selection and mutational pressure (10). Extensive research into codon usage patterns across the entire genome is imperative for understanding the fundamental characteristics of a genome's molecular organization. Furthermore, CUB analysis took into account numerous other crucially applied aspects, including heterologous gene expression (11), identifying species origins (12), predicting gene expression levels (13, 14), and predicting gene functions (15). However, the majority of the numerous reports on CUB concentrated on numerous microorganisms and viruses, including Betacoronavirus (16) and Henipavirus (17). For instance, we need to take note of the fact that the most preferred codons in Porcine astrovirus end in A or U (18). On the other hand, genome-wide studies on LSDV are limited.
The pattern of usage of synonymous codons by LSDV is fraught with uncertainty. In this study, we used a multivariate statistical analysis to examine the codon usage patterns of LSDV using the complete coding data. The analysis of the codon usage patterns of LSDV makes it possible for us to understand the underlying mechanism behind the biased usage of synonymous codons and to select suitable host expression systems for an optimal expression of the target genes.
A total of 53 complete coding sequences (CDS) of the lumpy skin disease virus (LSDV) were retrieved from the National Center for Biotechnology Information (NCBI) GenBank database (https://ncbi.nlm.nih.gov/nuccore/?term=Lumpy+Skin+Disease+Virus). The number of nucleotides in the coding sequence was an exact multiple of 3 (19). Complete information about the overall 53 LSDV strains associated with Asian, African, and European countries is listed in Supplementary Table 1.
Here, we employed the CodonW software to determine the total base composition (G, C, A, and T%) and the nucleotide contents at the 3rd codon position (C3, T3, G3, and A3%) for all synonymous codons in LSDV. The GC% contents of all three codon positions (GC1, GC2, and GC3%) were measured. Additionally, the average frequency of G/C at GC12 positions and the overall GC/AT compositions were also determined. Furthermore, only 59 synonymous codons encoding 18 amino acids were considered for the present study, not including the 1st ATG codon, the codon (TGG) encoding tryptophan, and the three end codons (TAG, TAA, and TGA), respectively (12).
The RSCU values indicate the observed codon occurrence to its random occurrence, suggesting that all the identical codons of the LSDV are equal in usage. There are frequently occurring codons that have an RSCU value >1 in the CDS, and less frequently occurring codons that have an RSCU value <1 in the CDS (20). Higher CUB or more frequently used codons were determined through high RSCU. In the coding sequence, the overrepresented codon represents the codon RSCU value >1.6, and the underrepresented codon represents the codon RSCU value <0.6 (21). The RSCU value was determined for each codon by using the following formula (22):
Here, ni is the number of codons for the ith amino acid and Xij denotes the frequency of the jth codon for the ith amino acid.
To determine the proper measurement of a codon bias, ENC (effective number of codons) value, which measures the total usage of the codon in a certain gene, was calculated (30, 31). It clarifies the ratio of codon variation in a gene from the total even usage of codons, which are synonymous. The ENC value varies from 20 (where one amino acid encodes one codon only) to 61 (where each amino acid is used randomly for all codons). The ENC value of <35 implies significant CUB (23, 25). To determine the impact of GC3s composition on the codon usage, a plot is drawn out between ENC and GC3s (23). For each GC3, the expected ENC values were computed by using the following formula:
where n is the total number of observations of the codons for that amino acid and ni is the total number of events of the ith codon for that amino acid.
wherever “s” is the GC3s content of each codon.
The obtained GC3 and GC12 values were plotted to determine and compare the extent of the factors that influence the preference of codon usage in the graph, each point signifying a discrete gene. The line of regression slope between the GC3 and the GC12 indicates that the mutational pressure is the major factor affecting CUB, i.e., for values coming close to 1, although if the value comes close to 0, it indicates that the selection pressure has been the main factor in defining CUB (26, 27).
The codon adaptation index (CAI) is applied to calculate the gene expression level depending on its codon-based sequence. The value of the CAI varies from 0 to 1; a value near 1 indicates higher levels of codon usage bias (CUB) (28). The CAI was determined through an online tool used for the CAI calculation (http://genomes.urv.es/CAIcal) (29), where the Bos taurus and Homo sapiens genomes were used as reference sources. Furthermore, the e-CAI (expected CAI) was analyzed using the online tool “http://genomes.urv.es/CAIcal/.” The RSCU values for the Bos taurus and Homo sapiens genomes were retrieved from the codon usage database.
The correspondence analysis (COA) is a multidimensional critical method that is used to resolve important developments in the codon usage patterns of CDS through codon RSCU values (12, 30, 31). To create the COA plot, the RSCU values of 59 codons were considered. To study the tendencies in the deviation of the use of codon, relative inertia was used to hold a specific position in the graph.
The phylogenetic tree was constructed using the maximum-likelihood (ML) method in Clustal × 2 (http://www.clustal.org/clustal2/). The phylogenetic tree was designed using the online tool, the Interactive Tree of Life (iTOL) version 3 (http://itol.embl.de/). A total of 53 strains were used in this study.
To illustrate the relationships between the nucleotide content and codon usage patterns, an LSDV correlation analysis was performed. These analyses were conducted by using Spearman's rank correlation method (32). All processes were executed using the R corrplot package. For codon usage index analysis, CodonW (1.4.4) software was applied (33, 34) to simplify the Multivariate analysis (correspondence analysis) of codon and amino acid usage.
The similarity index (SiD), which was used to measure how the overall codon usage pattern of the host affects the overall codon usage of the virus, was determined as follows:
where the R(A, B) denotes the degree of similarity between the overall codon usage patterns of the host and LSDV, which is defined as the cosine value of the angle between A and B. Among the 59 synonymous codons in the LSDV, ai is defined as the RSCU value for a particular codon. The RSCU value for the host's identical codon is known as bi. The value of D(A,B) ranges from 0 to 1.0 and shows the potential impact of the host's total codon usage on that of LSDV (35).
Codon usage bias (CUB) can be considerably predisposed by the general base composition of genomes. The nucleotide contents of 53 LSDV strains were studied and are presented in Table 1. Here, our results showed that the mean A (41.0%) and T (32.37%) were maximum, tailed by G (14.8%) and C (11.73%), across all genomes (Figure 1A). The mean A3 (39.83%) and T3 (40.2%) occurred at a maximum level higher than the G3 (10.1%) and C3 (9.81%) (Figure 1B). The total AT and GC compositions were found to be 73.42 and 26.57%, respectively, suggesting that LSDV strains have strong AT (Figure 1C). This finding is similar to the previous research on Porcine astrovirus (18), Hantaan virus (36), and Bluetongue virus (12) which were enriched with A and T. However, the biological significance of this condition still needs to be clarified, and therefore, it is essential to explore the causes for increased AT contents and decreased GC contents in the virus genomes (37).
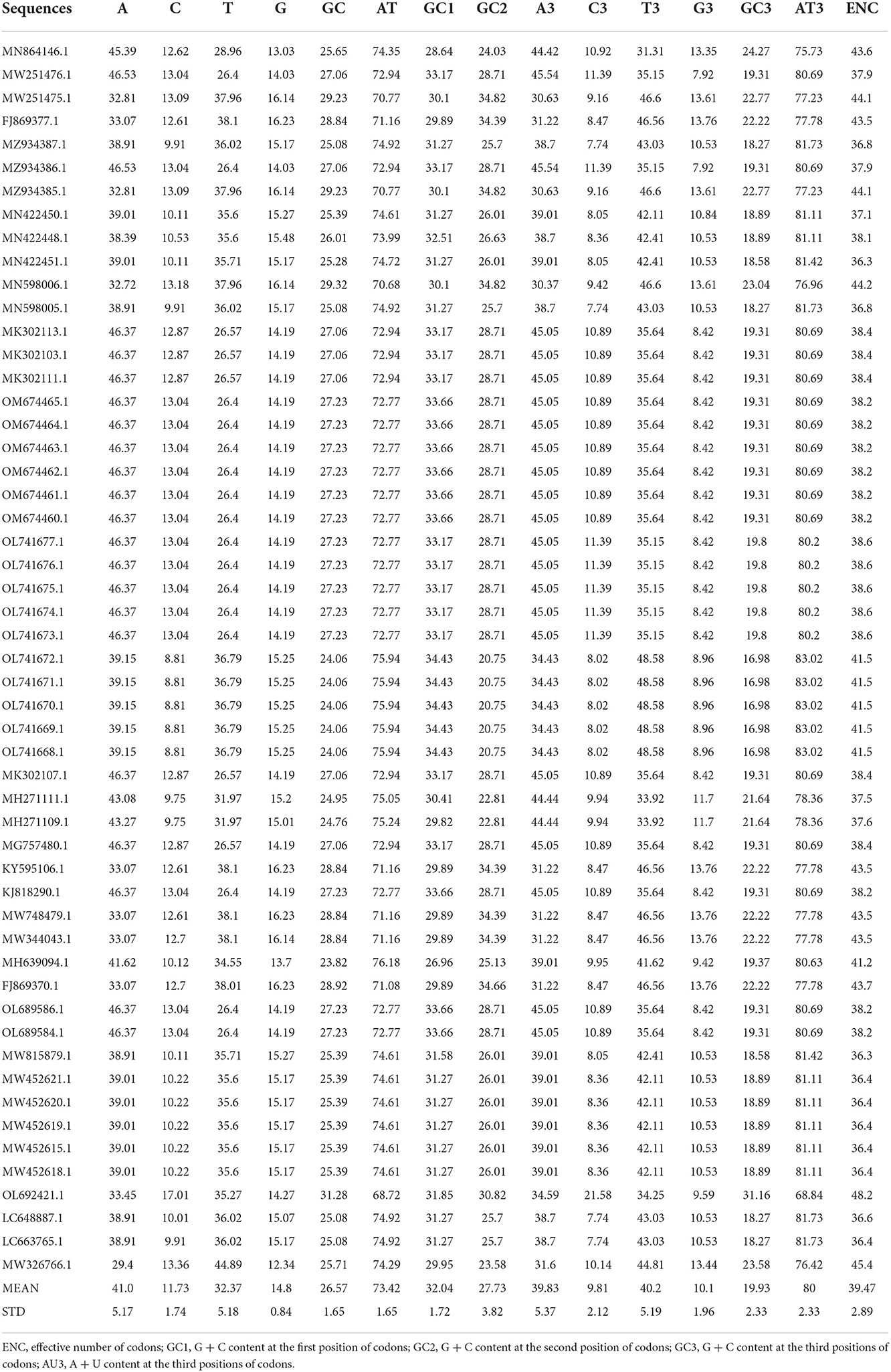
Table 1. Nucleotide compositional analysis of lumpy skin disease virus (LSDV) coding sequences (%).

Figure 1. Nucleotide composition analysis: (A) The average values of the A, T, G, and C nucleotide composition of the entire viral genome. (B) The average values of the nucleotide composition at the 3rd codon position, indicating A/T richness followed G/C richness. (C) The mean frequency of GC and AT composition. (D) The mean frequency of GC and AT composition at the 3rd codon position, indicating that AT3 is more common than GC3.
Nucleotide content analysis at the 1st, 2nd, and 3rd synonymous codon positions disclosed that the values of GC1 ranged from 28.29 to 41.64% (mean: 32.04%; SD: 1.72), while the values of GC2 ranged from 23.38 to 46.77% (mean: 27.73%; SD: 3.82). Whereas, the values of GC3 ranged from 16.28 to 51.64% (mean: 19.93%; SD: 2.33), which are similar to previous studies on the Crimean–Congo hemorrhagic fever (CCHF) virus (38). In contrast, the values of AU3 ranged from 48.36 to 83.72% (mean: 78.11%; SD: 6.36) (Figure 1D). These data further supported the notion that an extensive area of LSDV is self-possessed of A/T contents (Table 1). This study supports the previous studies on the Alongshan virus and Zika virus (24, 39), while it seems to be inconsistent with a recent study on the hepatitis E virus (HEV) (40).
To describe and define the codon usage bias of LSDV, an RSCU analysis was carried out to determine why A/U nucleotides were favored at the third codon position. For 53 LSDV strains, the RSCU values of all synonymous codons were determined and compared to those of their hosts. The result showed that all the 18 most abundantly used codons in LSDV [TTT (Phe), TTA (Leu), ATT (Ile), GTT (Val), AGT (Ser), CCT (Pro), ACT (Thr), GCT (Ala), CAT (His), TAT (Tyr), CAA (Gln), AAT (Asn), AAA (Lys), GAT (Asp), TGT (Cys), CGT (Arg), and GGT (Gly)] ended with T or A (T: 14; A: 3). Interestingly, none of the preferred codons was G/C-ended. Thus, the A or T-end codon bases are shared more in the genome of LSDV, which is similar to an earlier research that T/A-ended codons have a high amount in the virus genome, such as chikungunya virus and Crimean–Congo hemorrhagic fever virus (34, 38). Furthermore, from the RSCU analysis, we found that the overrepresented (>1.6) codons are rarely seen in the genome of LSDV. Nearly all the ideal and non-ideal codons are located in the range of 0.6–1.6. We observed that most codons ending in A/T were overrepresented (>1.6), while codons ending in G/C were under-represented (<0.6) (Table 2), revealing that mutational pressure was the primary factor influencing codon usage patterns in LSDV, which was consistent with those given in previous studies (41, 42). From both the nucleotide content and RSCU analysis, we assumed that the selection of the preferred codons is generally inclined by compositional restraints, which determine the existence of mutational pressure. We are unsure of the fact that the compositional pressure could not be the single aspect related to LSDV patterns of codon usage, as although the total values of RSCU could disclose the pattern of codon usage for the genomes, it may conceal the codon usage variation among distinct genes in a genome (43).
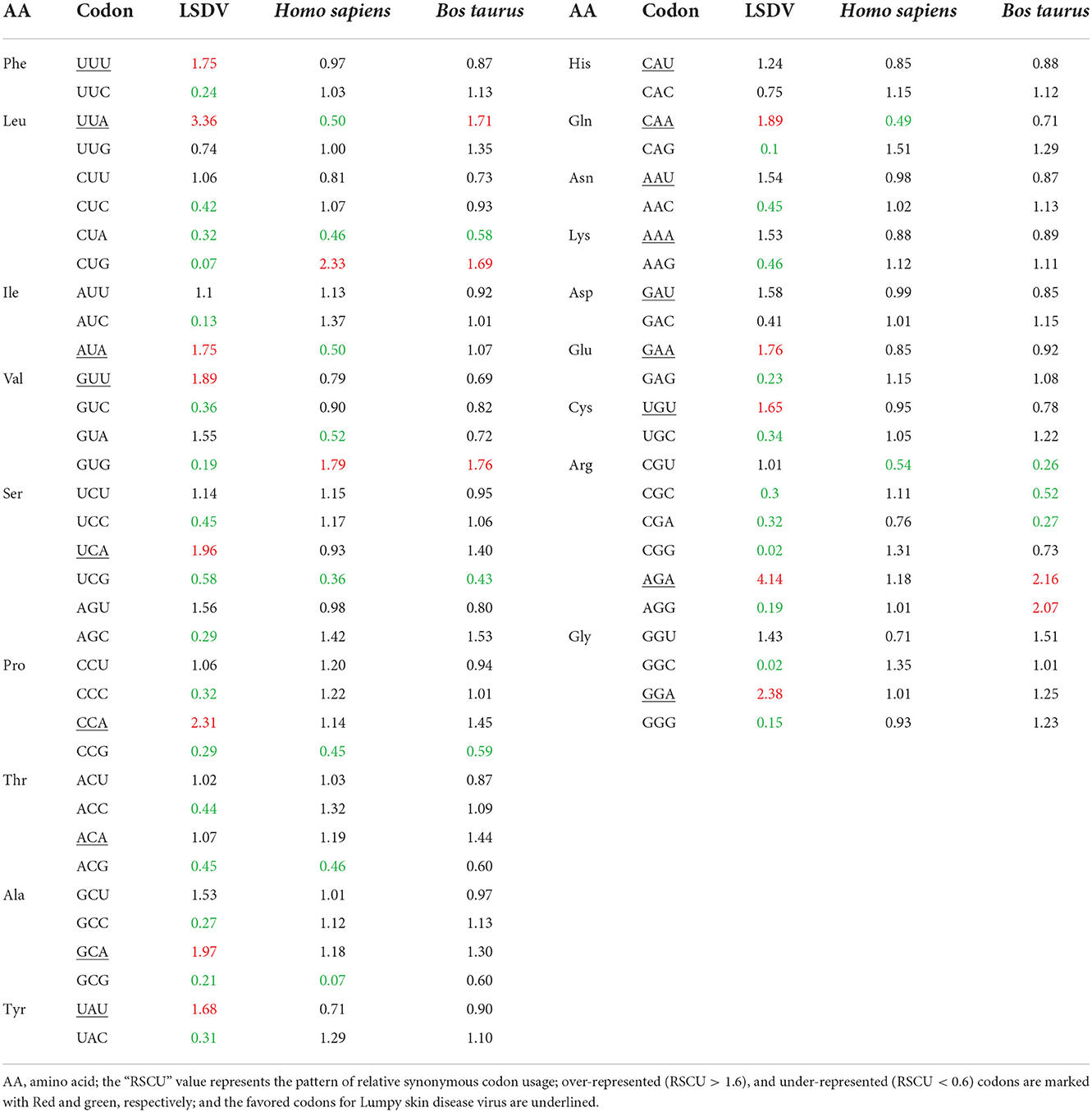
Table 2. The relative synonymous codon usage frequency of Lumpy skin disease virus (LSDV) and its natural hosts (Homo sapiens and Bos taurus).
Additionally, to determine whether the codon usage bias of LSDV can be constrained by its hosts (Bos taurus and Homo sapiens), all their codon RSCU values were also calculated (Table 2). This study indicated that 12 of 59 synonymous codons of LSDV are equivalent to those of Homo sapiens, individually, and that 09 of 59 synonymous codons are equivalent to those of Bos taurus (Figure 2, Table 2). Here, the role of selection from the Bos taurus in shaping codon usage patterns of LSDV is different from that of the host Homo sapiens. It was suggested that the similarity of codon usage patterns between LSDV and Bos taurus/Homo sapiens enhances the efficiency of translation in the virus genome (44).
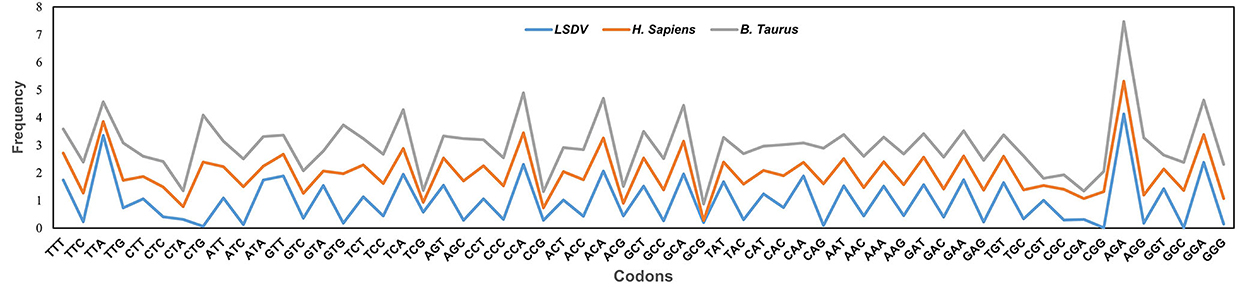
Figure 2. Comparative analysis of relative synonymous codon usage (RSCU) patterns between lumpy skin disease virus (LSDV), and its hosts Bos taurus and Homo sapiens. X-axis represents the Codons, while the Y-axis represents the Frequency.
To determined the magnitude of CUB within LSDV coding sequences, the gene's ENC value was assessed and plotted next to the GC content at the 3rd codon position (GC3) (Table 1). Here, the values of ENC were shown to vary from 36.28 to 48.18, indicating a high level of genetic difference in the codon's usage. Nevertheless, the average value of ENC was 39.47 > 20, implying that the whole CUB was low (Table 1), which was also observed in Human cytomegalovirus (45), Ebola virus (46), and Porcine astrovirus (18). The ENC analysis revealed that a low codon bias was seen along with the position of natural selection on the genes (32, 47). Therefore, it seems that the evolution of a low codon bias within LSDV coding sequences has enabled LSDV survival within the host, even though the host possesses codon usage preferences distinct from those of LSDV.
Next, to determine the codon usage of the genes, a distribution plot that deviated from a similar usage of indistinguishable codons was employed. Here, ENC values were used against the GC3s. If the GC subject of the gene exhibits mutational pressure, all the points in this plot lie below or close to the expected curve, indicating a random codon usage. However, if there is selection pressure on the gene, all the points lie on or below the expected curve. Here, we plotted the ENC values of each gene against the GC3 content (Figure 3). The results reveal that mutational pressure and natural selection both influence the codon usage pattern of an LSDV genome, as the majority of the points fall below the expected curve and just a few points beyond it (48, 49).
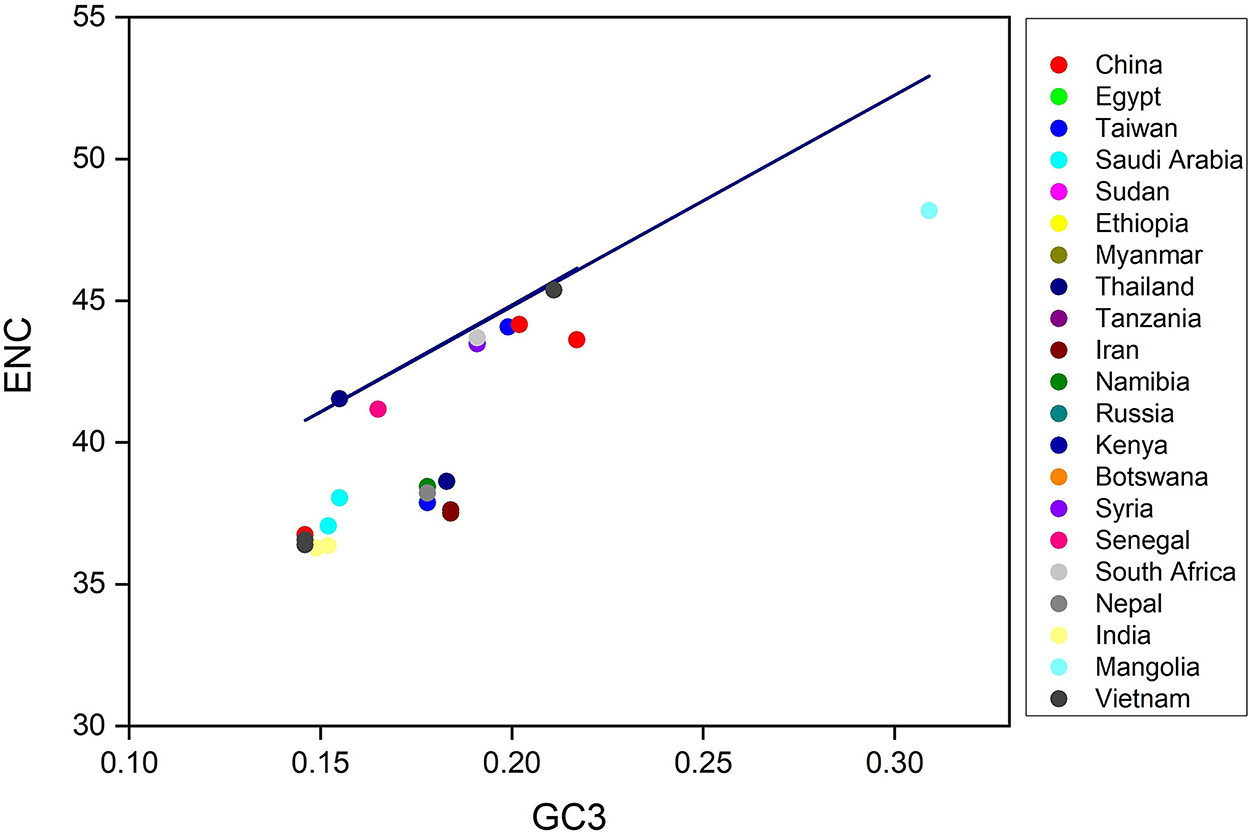
Figure 3. ENC-GC3 plots of 53 lumpy skin disease virus (LSDV) strains: The effective number of codons (ENC-values, Y-axis) is plotted against the GC content at the third synonymous codon positions (GC3-values, X-axis).
A neutrality plot analysis was performed, which implied the bond between GC1/2 and GC3 composition, to determine the position of mutation and selection pressure that has an impact on the form of the codon usage bias (CUB). To observe the association, we programmed a paradigm on the plot of neutrality between GC3 and GC1/2 for the LSDV genome. Here, the said plot shows that no significant relation was found between GC3 and GC1/2 contents because the regression value and link are P > 0.05 and r = 0.003 (Figure 4). Finally, we suggested that both natural selection and mutational pressure have an impact on the codon usage shaping of LSDV. This phenomenon is similar to those given in the previous studies (41, 50, 51).
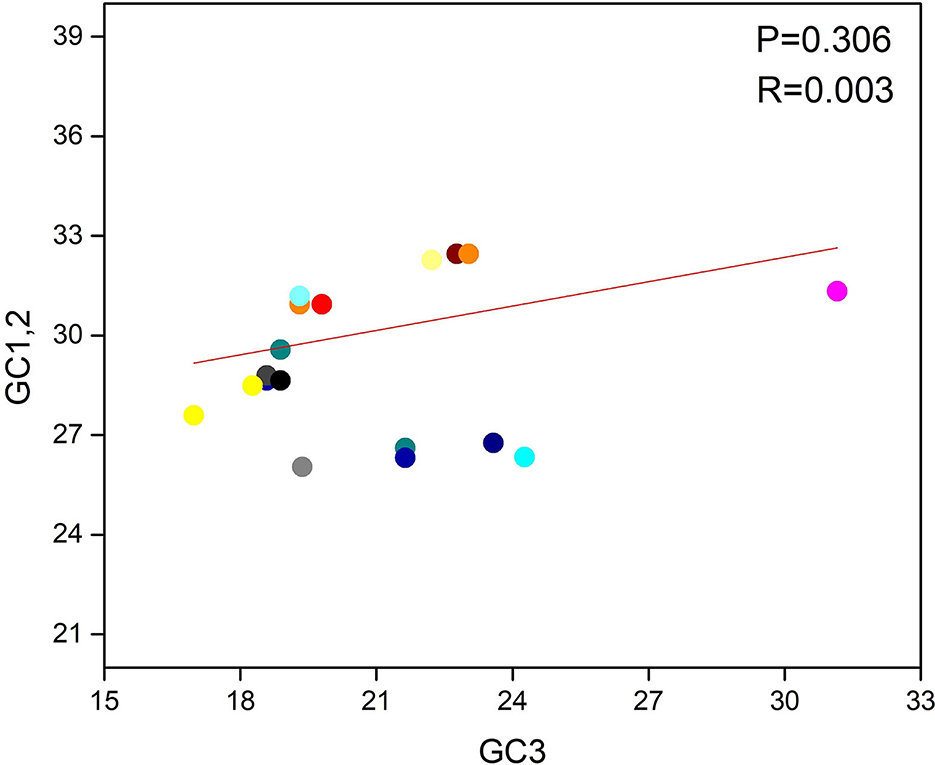
Figure 4. Neutrality plot between (GC3 vs. GC1, 2) for the entire coding sequence of lumpy skin disease virus (LSDV). GC1, 2 represent GC at 1st and 2nd codon positions. While GC3 represents GC at 3rd codon position. And the orange solid line represents the regression analysis of GC1, 2 against GC3.
The codon adaptation index (CAI) analysis was performed to regulate the optimization of codon usage and LSDV adaptation to its hosts (52). The values of CAI range from 0 to 1; a value near 1 indicates higher levels of codon usage bias (13). For all codons, the CAI values were measured through the reference of Bos taurus and Homo sapiens codon usage. We determined that, as regards Bos taurus and Homo sapiens, the mean CAI values of LSDV coding regions were 0.59 and 0.68 (>0.5), which revealed that LSDV has a good adaptation to its hosts and a minimal translation pressure (Supplementary Figure 1) (24, 53). The tendency of Homo sapiens to move toward a high CAI value recommends that the selection pressure from Homo sapiens should impact the LSDV codon usage, and that the codon usage evolution in LSDV should permit it to use the translation machinery of Homo sapiens more capably. Our result was consistent with the published work (54).
To check if the observed statistically significant differences arose in the values of CAI (29, 38), the values of expected CAI (e-CAI) were considered for LSDV CDS with Bos taurus and Homo sapiens codon usage sets. The results of the e-CAI values were 0.70 and 0.79 (P < 0.05) in relation to Bos taurus and Homo sapiens, revealing that the generated sequences keep to a normal distribution. The outcomes of this study about the preferences of codon usage are comparable with those of previous research (12, 55).
The Correspondence (CO) analysis describes the discrepancy in the usage of codons. The changes that occur in the patterns of codon usage are revealed through RSCU values. In the plot of CO analysis, axes 1 and 2 are the two main factors of general discrepancy (30, 34, 56). We used the values of these two axes to draw COA plots, where each strain is represented by a point, and the distance between strains gives a degree of similarity or dissimilarity in the codon usage patterns. The principal axes 1 and 2 accounted for the total variation: 65.66 and 34.34% (Figure 5). These results propose the fact that axis 1 signifies the LSDV strains and axis 2 signifies the countries where the LSDV arises. Scattered data on the main axis represent various geographical ancestries and their relationships. All the LSDV strains were found to be placed in groups using COA (Figure 5). The entire Iran LSDV strains were assembled into one clade, while strains from Taiwan, Nepal, Namibia, and Thailand were present in an alternative clade. Moreover, China, Saudi Arabia, and Sudan strains were divided into separate groups (Figure 5). These studies reveal that the topographical sites play a major role in the evolution of LSDV and in a synonymous codon usage pattern, where in the future such investigations may assist in discovering the essence of rising LSDV strains. Furthermore, present outcomes also show that more than one widespread genetic lineage was found in every infected country.
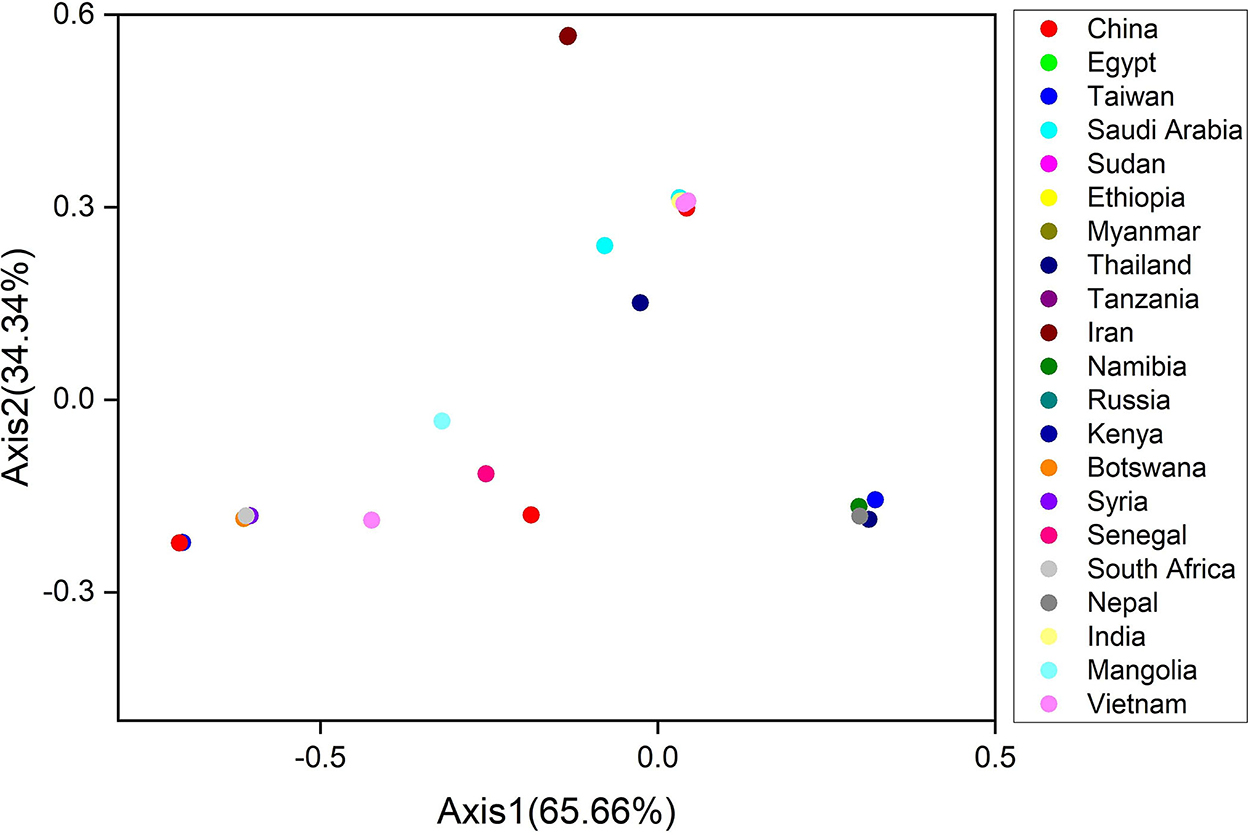
Figure 5. The correspondence analysis (COA) of the genes in lumpy skin disease virus (LSDV) genomes. Each point represents a gene corresponding to the coordinates of the first and second axes of variation generated from the correspondence analysis.
To assess the consequence of evolutionary procedures in the LSDV codon usage pattern, a phylogenetic analysis was carried out using the maximum-likelihood (ML) method. The entire LSDV that separates is dispersed throughout the world, as evidenced by the phylogenetic tree, which shows that no strains form a cluster among different individual countries (Figure 6). The study suggested that this virus might have become altered due to some specific geographical effects such as climatic changes and environmental changes, which support the main outcome of evolutionary processes and topographical dispersal on codon usage patterns. The current study further exposed the signs of recombination and genome reassortment during single host co-infection, signifying the potential for the upcoming arrival of the novel alternates (57, 58).
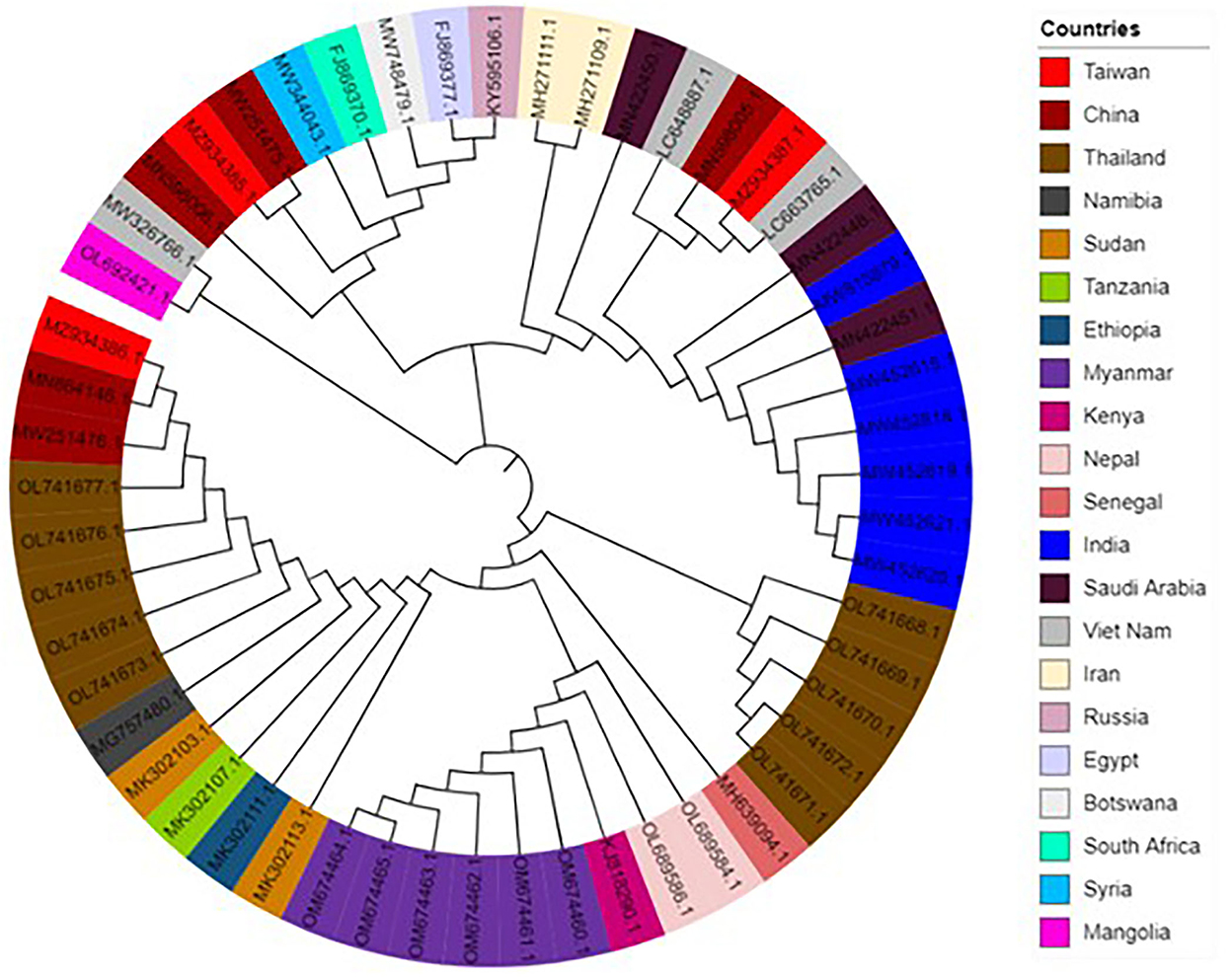
Figure 6. Phylogenetic tree based on the polyprotein-coding regions of 53 lumpy skin disease virus (LSDV) strains. The tree was generated by the maximum-likelihood (ML) method using Clustal X2. The tree was designed by using the online tool “iTOL”.
Here, we consider two factors: natural selection and mutational pressure, to determine the codon usage bias (CUB) in LSDV. Accordingly, we performed a correlation analysis between total nucleotide contents (A, G, C, T), nucleotide contents at 3rd codon position, GC contents (1st, 2nd, 3rd), GC12, and ENC. The ENC values of the LSDV sequences seemed to show a positive relationship with T, C, G, T3, C3, G3, GC, GC2, GC3, and GC12, except A, A3, AT, GC1, and AT3, which have a negative relationship that probably affects the LSDV codon usage pattern (Figure 7). Previous studies have suggested that when we have the base compositions at the 3rd position of codon, mutational bias is mostly explained, while with base compositions at the 1st and 2nd positions, selective pressure is mostly validated (45, 59).
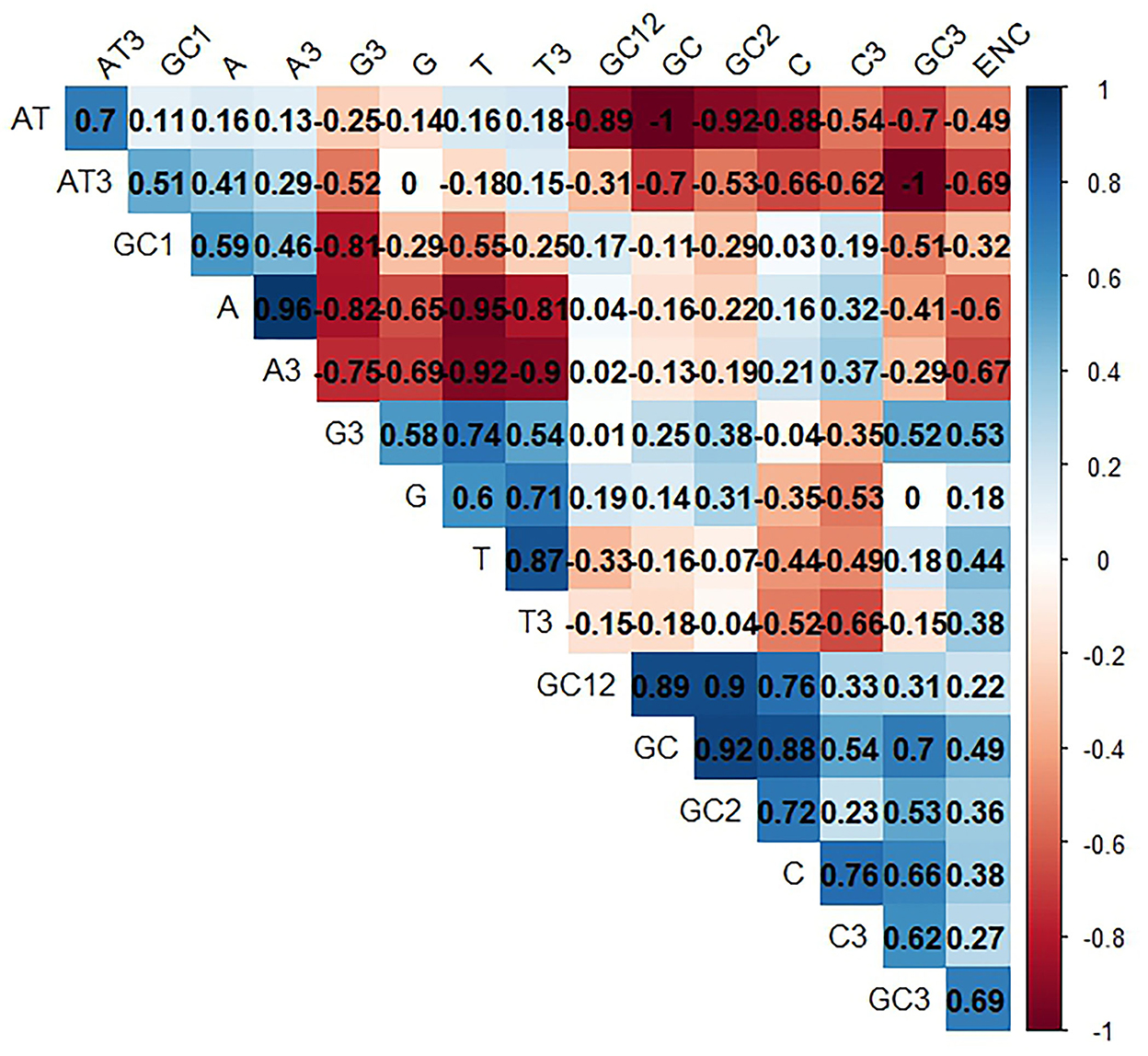
Figure 7. Correlation analysis among different nucleotide contents of lumpy skin disease virus (LSDV). The dark blue means positive correlation, and the dark red means negative correlation; the larger the value means more significant correlation.
Such an impact was also observed among A, G, T, C, GC, AT, GC3, AT3, A3, C3, G3, and T3 with GC12. The A, G, C, G3, GC, GC3, A3, and C3 have a positive correlation with ENC, while the T, AT, AT3, and T3 have a negative correlation with ENC. This result implies the significance of mutational and selection pressure in getting the LSDV codon usage pattern (Figure 7). Additionally, it also suggests that the contents of a nucleotide have an impact on the codon usage pattern of LSDV (60).
The similarity index (SiD) analysis was carried out to assess the potential impact of Bos taurus and Homo sapiens codon usage patterns on the evolution of the codon usage patterns of the LSDV. The SiD was found to be nearly similar in both Bos taurus (0.63) and Homo sapiens (0.68), indicating that both of them have a dominant influence on the formation of LSDV codon usage patterns (Figure 8). Previous studies showed that Bos taurus is thought to be the principal reservoir and a host of LSDV. It is likely that the virus has stabilized its genetic traits to better adapt to the environment of its primary host (35, 61).
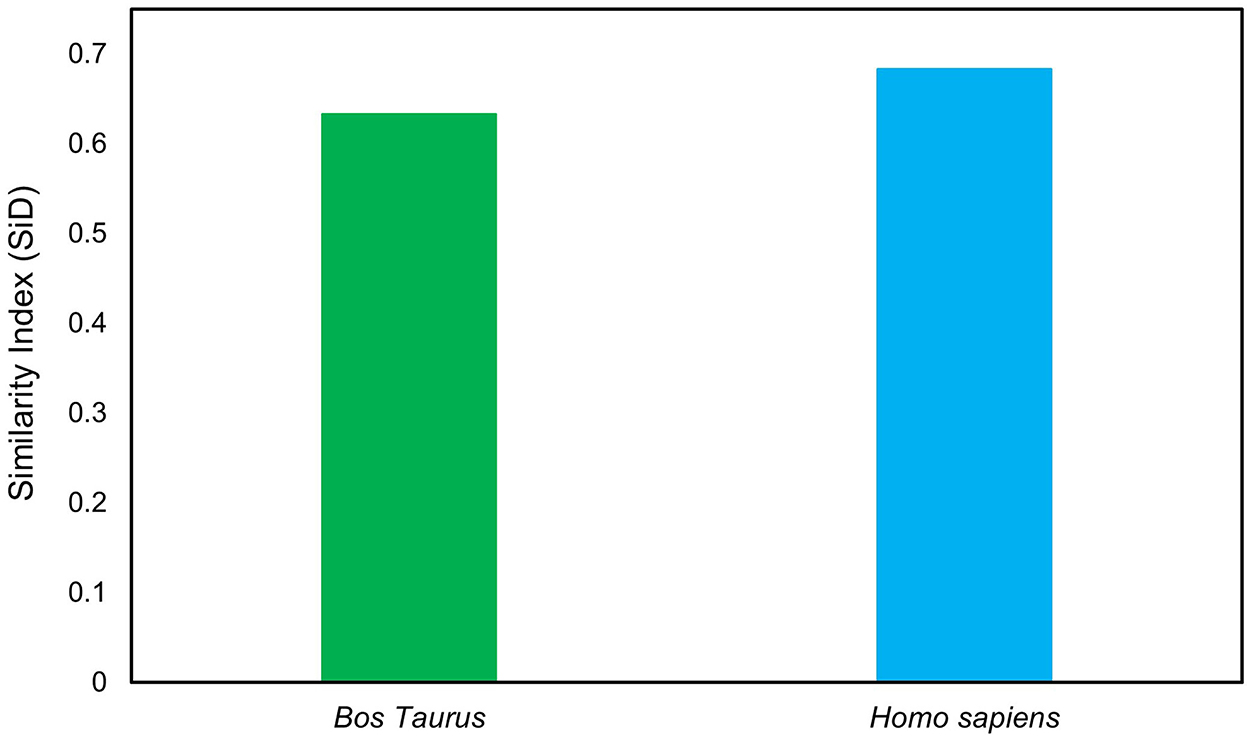
Figure 8. Similarity index (SiD) analysis of the codon usage between lumpy skin disease virus (LSDV) and its hosts.
Publicly available datasets were analyzed in this study. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
SRah and IR: conceptualization, methodology, software, data curation, and writing—original draft preparation. AR, SRaz, and NH: helped in write-up and editing and validation. HR: methodology, visualization, and validation. AA, MA, and HS: reviewing and editing and validation. All authors contributed to the article and approved the submitted version.
Authors are thankful to the Researchers Supporting Project number (RSP2022R491), King Saud University, Riyadh, Saudi Arabia.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2022.1071097/full#supplementary-material
Supplementary Figure 1. Codon adaptation index (CAI) of lumpy skin disease virus (LSDV) to its hosts (Bos taurus and Homo sapiens).
Supplementary Table 1. Detailed information of lumpy skin disease virus studied in this study.
1. Badhy SC, Chowdhury MGA, Settypalli TBK, Cattoli G, Lamien CE, Fakir MAU, et al. Molecular characterization of lumpy skin disease virus (LSDV) emerged in Bangladesh reveals unique genetic features compared to contemporary field strains. BMC Vet Res. (2021) 17:7122–30. doi: 10.1186/s12917-021-02751-x
2. Tulman ER, Afonso CL, Lu Z, Zsak L, Kutish GF, Rock DL. Genome of lumpy skin disease virus. J Virol. (2001) 75:7122–30. doi: 10.1128/JVI.75.15.7122-7130.2001
3. Ahmed Kamal S. Comparative studies on lumpy skin disease virus in human. Med Clin Arch. (2019) 3:1–8. doi: 10.15761/MCA.1000161
4. Sprygin A, Pestova Y, Wallace DB, Tuppurainen E, Kononov AV. Transmission of lumpy skin disease virus: a short review. Virus Res. (2019) 269:197637. doi: 10.1016/j.virusres.2019.05.015
5. Tuppurainen ESM, Stoltsz WH, Troskie M, Wallace DB, Oura CAL, Mellor PS, et al. A potential role for ixodid (hard) tick vectors in the transmission of lumpy skin disease virus in cattle. Transbound Emerg Dis. (2011) 58:93–104. doi: 10.1111/j.1865-1682.2010.01184.x
6. Namazi F, Khodakaram Tafti A. Lumpy skin disease, an emerging transboundary viral disease: a review. Vet Med Sci. (2021) 7:888–96. doi: 10.1002/vms3.434
7. Azeem S, Sharma B, Shabir S, Akbar H, Venter E. Lumpy skin disease is expanding its geographic range: a challenge for Asian livestock management and food security. Vet J. (2022) 279:105785. doi: 10.1016/j.tvjl.2021.105785
8. Rahman SU, Nawaz S, Ullah S, Rahman IU, Haq MIU, Khan MA, et al. Study of codon usage patterns and influencing factors in rice yellow mottle virus based on coding sequence data. Agronomy. (2022) 12:1990. doi: 10.3390/agronomy12091990
9. Nguyen TH, Wang D, Rahman SU, Bai H, Yao X, Chen D, et al. Analysis of codon usage patterns and influencing factors in rice tungro bacilliform virus. Infect Genet Evol. (2021) 90:104750. doi: 10.1016/j.meegid.2021.104750
10. Hershberg R, Petrov DA. Selection on codon bias. Annu Rev Genet. (2008) 42:287–99. doi: 10.1146/annurev.genet.42.110807.091442
11. Kane JF. Effects of rare codon clusters on high-level expression of heterologous proteins in Escherichia coli. Curr Opin Biotechnol. (1995) 6:494–500. doi: 10.1016/0958-1669(95)80082-4
12. Yao X, Fan Q, Yao B, Lu P, Rahman SU, Chen D, et al. Codon usage bias analysis of bluetongue virus causing livestock infection. Front Microbiol. (2020) 11:655. doi: 10.3389/fmicb.2020.00655
13. Gupta SK, Bhattacharyya TK, Ghosh TC. Synonymous codon usage in Lactococcus lactis: mutational bias versus translational selection. J Biomol Struct Dyn. (2004) 21:527–36. doi: 10.1080/07391102.2004.10506946
14. Naya H, Romero H, Carels N, Zavala A, Musto H. Translational selection shapes codon usage in the GC-rich genome of Chlamydomonas reinhardtii. FEBS Lett. (2001) 501:127–30. doi: 10.1016/S0014-5793(01)02644-8
15. Lin K, Kuang Y, Joseph JS, Kolatkar PR. Conserved codon composition of ribosomal protein coding genes in Escherichia coli, Mycobacterium tuberculosis and Saccharomyces cerevisiae: lessons from supervised machine learning in functional genomics. Nucleic Acids Res. (2002) 30:2599–607. doi: 10.1093/nar/30.11.2599
16. Castells M, Victoria M, Colina R, Musto H, Cristina J. Genome-wide analysis of codon usage bias in bovine coronavirus. Virol J. (2017) 14:1–7. doi: 10.1186/s12985-017-0780-y
17. Khandia R, Singhal S, Kumar U, Ansari A, Tiwari R, Dhama K, et al. Analysis of nipah virus codon usage and adaptation to hosts. Front Microbiol. (2019) 10:886. doi: 10.3389/fmicb.2019.00886
18. Wu H, Bao Z, Mou C, Chen Z, Zhao J. Comprehensive analysis of codon usage on porcine astrovirus. Viruses. (2020) 12:991. doi: 10.3390/v12090991
19. Karumathil S, Raveendran NT, Ganesh D, Kumar Ns S, Nair RR, Dirisala VR. Evolution of synonymous codon usage bias in west African and central African strains of monkeypox virus. Evol Bioinform. (2018) 14:1176934318761368. doi: 10.1177/1176934318761368
20. Sharp PM, Li WH. Codon usage in regulatory genes in Escherichia coli does not reflect selection for “rare” codons. Nucleic Acids Res. (1986) 14:7737–7749. doi: 10.1093/nar/14.19.7737
21. Wong EHM, Smith DK, Rabadan R, Peiris M, Poon LLM. Codon usage bias and the evolution of influenza A viruses. Codon usage biases of influenza virus. BMC Evol Biol. (2010) 10:253. doi: 10.1186/1471-2148-10-253
22. Liu J, Zhu D, Ma G, Liu M, Wang M, Jia R, et al. Genome-Wide analysis of the synonymous codon usage patterns in Riemerella anatipestifer. Int J Mol Sci. (2016) 17:1304. doi: 10.3390/ijms17081304
23. Wright F. The “effective number of codons” used in a gene. Gene. (1990) 87:23–9. doi: 10.1016/0378-1119(90)90491-9
24. Rahman SU, Abdullah M, Khan AW, Haq MIU, Haq N ul, Aziz A, et al. A detailed comparative analysis of codon usage bias in alongshan virus. Virus Res. (2022) 308:198646. doi: 10.1016/j.virusres.2021.198646
25. Comeron JM, Aguade M. An evaluation of measures of synonymous codon usage bias. J Mol Evol. (1998) 47:268–274. doi: 10.1007/PL00006384
26. Guan DL, Ma L Bin, Khan MS, Zhang XX, Xu SQ, Xie JY. Analysis of codon usage patterns in Hirudinaria manillensis reveals a preference for GC-ending codons caused by dominant selection constraints. BMC Genomics. (2018) 19:542. doi: 10.1186/s12864-018-4937-x
27. Sueoka N. Directional mutation pressure and neutral molecular evolution. Proc Natl Acad Sci USA. (1988) 85:2653–7. doi: 10.1073/pnas.85.8.2653
28. Chakraborty A, Uechi T, Higa S, Torihara H, Kenmochi N. Loss of ribosomal protein L11 affects zebrafish embryonic development through a p53-dependent apoptotic response. PLoS ONE. (2009) 4:e4152. doi: 10.1371/journal.pone.0004152
29. Puigbò P, Bravo IG, Garcia-Vallve S. CAIcal: a combined set of tools to assess codon usage adaptation. Biol Direct. (2008) 3:38. doi: 10.1186/1745-6150-3-38
31. Cristina J, Fajardo A, Sonora M, Moratorio G, Musto H. A detailed comparative analysis of codon usage bias in Zika virus. Virus Res. (2016) 223:147–52. doi: 10.1016/j.virusres.2016.06.022
32. Wu Y, Zhao D, Tao J. Analysis of codon usage patterns in herbaceous peony (Paeonia lactiflora Pall.) based on transcriptome data. Genes. (2015) 6:1125–39. doi: 10.3390/genes6041125
33. Singh NK, Tyagi A. A detailed analysis of codon usage patterns and influencing factors in Zika virus. Arch Virol. (2017) 162:1963–73. doi: 10.1007/s00705-017-3324-2
34. Butt AM, Nasrullah I, Tong Y. Genome-wide analysis of codon usage and influencing factors in chikungunya viruses. PLoS ONE. (2014) 9:e90905. doi: 10.1371/journal.pone.0090905
35. Butt AM, Nasrullah I, Qamar R, Tong Y. Evolution of codon usage in Zika virus genomes is host and vector speci fi c. Nat Publ Gr. (2016) 5:e107. doi: 10.1038/emi.2016.106
36. Ata G, Wang H, Bai H, Yao X, Tao S. Edging on mutational bias, induced natural selection from host and natural reservoirs predominates codon usage evolution in hantaan virus. Front Microbiol. (2021) 12:699788. doi: 10.3389/fmicb.2021.699788
37. van Hemert F, Berkhout B. Nucleotide composition of the Zika virus RNA genome and its codon usage. Virol J. (2016) 13:95. doi: 10.1186/s12985-016-0551-1
38. Rahman SU, Yao X, Li X, Chen D, Tao S. Analysis of codon usage bias of crimean-congo hemorrhagic fever virus and its adaptation to hosts. Infect Genet Evol. (2018) 58:1–16. doi: 10.1016/j.meegid.2017.11.027
39. Rahman SU, Mao Y, Tao S. Codon usage bias and evolutionary analyses of Zika virus genomes. Genes Genomics. (2017) 39:855–66. doi: 10.1007/s13258-017-0549-0
40. Li B, Wu H, Miao Z, Lu Y. Using codon usage analysis to speculate potential animal hosts of hepatitis E virus: an exploratory study. Infect Genet Evol. (2022) 101:105284. doi: 10.1016/j.meegid.2022.105284
41. Yang X, Ma X, Luo X, Ling H, Zhang X, Cai X. Codon usage bias and determining forces in Taenia solium genome. Korean J Parasitol. (2015) 53:689–97. doi: 10.3347/kjp.2015.53.6.689
42. Le TH, McManus DP, Blair D. Codon usage and bias in mitochondrial genomes of parasitic platyhelminthes. Korean J Parasitol. (2004) 42:159–67. doi: 10.3347/kjp.2004.42.4.159
43. Hassan S, Mahalingam V, Kumar V. Synonymous codon usage analysis of thirty two mycobacteriophage genomes. Adv Bioinformatics. (2009) 2009:316936. doi: 10.1155/2009/316936
44. Ma X-X, Feng Y-P, Bai J-L, Zhang D-R, Lin X-S, Ma Z-R. Nucleotide composition bias and codon usage trends of gene populations in Mycoplasma capricolum subsp. capricolum and M. Agalactiae. J Genet. (2015) 94:251–60. doi: 10.1007/s12041-015-0512-2
45. Hu C, Chen J, Ye L, Chen R, Zhang L, Xue X. Codon usage bias in human cytomegalovirus and its biological implication. Gene. (2014) 545:5–14. doi: 10.1016/j.gene.2014.05.018
46. Cristina J, Moreno P, Moratorio G, Musto H. Genome-wide analysis of codon usage bias in ebolavirus. Virus Res. (2015) 196:87–93. doi: 10.1016/j.virusres.2014.11.005
47. Ma YP, Liu ZX, Hao L, Ma JY, Liang ZL, Li YG, et al. Analysing codon usage bias of cyprinid herpesvirus 3 and adaptation of this virus to the hosts. J Fish Dis. (2015) 38:665–73. doi: 10.1111/jfd.12316
48. Li X, Wang X, Gong P, Zhang N, Zhang X, Li J. Analysis of codon usage patterns in giardia duodenalis based on transcriptome data from GiardiaDB. Genes. (2021) 12:1169. doi: 10.3390/genes12081169
49. Zhou J-H, Gao Z-L, Zhang J, Ding Y-Z, Stipkovits L, Szathmary S, et al. The analysis of codon bias of foot-and-mouth disease virus and the adaptation of this virus to the hosts. Infect Genet Evol. (2013) 14:105–10. doi: 10.1016/j.meegid.2012.09.020
50. Chen L, Liu T, Yang D, Nong X, Xie Y, Fu Y, et al. Analysis of codon usage patterns in Taenia pisiformis through annotated transcriptome data. Biochem Biophys Res Commun. (2013) 430:1344–8. doi: 10.1016/j.bbrc.2012.12.078
51. Yang X, Luo X, Cai X. Analysis of codon usage pattern in Taenia saginata based on a transcriptome dataset. Parasit Vectors. (2014) 7:527. doi: 10.1186/s13071-014-0527-1
52. Sharp PM, Li WH. An evolutionary perspective on synonymous codon usage in unicellular organisms. J Mol Evol. (1986) 24:28–38. doi: 10.1007/BF02099948
53. Subramanian A, Rup Sarkar R. Data in support of large scale comparative codon usage analysis in Leishmania and Trypanosomatids. Data Br. (2015) 4:269–72. doi: 10.1016/j.dib.2015.06.003
54. Pan X, Welti R, Wang X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography–mass spectrometry. Nat Protoc. (2010) 5:986–92. doi: 10.1038/nprot.2010.37
55. Maldonado LL, Stegmayer G, Milone DH, Oliveira G, Rosenzvit M, Kamenetzky L. Whole genome analysis of codon usage in Echinococcus. Mol Biochem Parasitol. (2018) 225:54–66. doi: 10.1016/j.molbiopara.2018.08.001
56. Kumar N, Bera BC, Greenbaum BD, Bhatia S, Sood R, Selvaraj P, et al. Revelation of influencing factors in overall codon usage bias of equine influenza viruses. PLoS ONE. (2016) 11:e0154376. doi: 10.1371/journal.pone.0154376
57. Chamberlain J, Cook N, Lloyd G, Mioulet V, Tolley H, Hewson R. Co-evolutionary patterns of variation in small and large RNA segments of crimean-congo hemorrhagic fever virus. J Gen Virol. (2005) 86:3337–41. doi: 10.1099/vir.0.81213-0
58. Hewson R, Gmyl A, Gmyl L, Smirnova SE, Karganova G, Jamil B, et al. Evidence of segment reassortment in crimean-congo haemorrhagic fever virus. J Gen Virol. (2004) 85:3059–70. doi: 10.1099/vir.0.80121-0
59. RoyChoudhury S, Mukherjee D. A detailed comparative analysis on the overall codon usage pattern in herpesviruses. Virus Res. (2010) 148:31–43. doi: 10.1016/j.virusres.2009.11.018
60. Chen H, Sun S, Norenburg JL, Sundberg P. Mutation and selection cause codon usage and bias in mitochondrial genomes of ribbon worms (Nemertea). PLoS ONE. (2014) 9:e85631. doi: 10.1371/journal.pone.0085631
Keywords: lumpy skin disease virus, codon usage bias (CUB), effective number of codons (ENC), mutation pressure, natural selection
Citation: Rahman SU, Rehman HU, Rahman IU, Rauf A, Alshammari A, Alharbi M, Haq Nu, Suleria HAR and Raza SHA (2022) Analysis of codon usage bias of lumpy skin disease virus causing livestock infection. Front. Vet. Sci. 9:1071097. doi: 10.3389/fvets.2022.1071097
Received: 15 October 2022; Accepted: 10 November 2022;
Published: 05 December 2022.
Edited by:
Farhan Anwar Khan, University of Agriculture, PakistanReviewed by:
Mueed Rahman, Shanghai Jiao Tong University, ChinaCopyright © 2022 Rahman, Rehman, Rahman, Rauf, Alshammari, Alharbi, Haq, Suleria and Raza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Siddiq Ur Rahman, c2lkZGlxdXIucmFobWFuQGtra3VrLmVkdS5waw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.