 Naomi Ariyama1
Naomi Ariyama1 Belén Agüero1Valentina Valdés1Felipe Berrios1Sergio Bucarey2
Belén Agüero1Valentina Valdés1Felipe Berrios1Sergio Bucarey2 Sunil Mor3
Sunil Mor3 Barbara Brito4
Barbara Brito4 Victor Neira1*
Victor Neira1*- 1Departamento de Medicina Preventiva Animal, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile, Santiago, Chile
- 2Departamento de Ciencias Biológicas, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile, Santiago, Chile
- 3Veterinary Population Medicine Department, College of Veterinary Medicine, University of Minnesota, St. Paul, MN, United States
- 4The iThree Institute, University of Technology Sydney, Sydney, NSW, Australia
Porcine Circovirus 2 (PCV2) can cause multiple clinical conditions known as porcine circovirus-associated diseases (PCVAD). Before the wide availability of PCV2 vaccines, PCVAD resulted in significant losses to the global swine industry. PCV2's rapid evolutionary dynamics are comparable to single-stranded RNA viruses. Thus, shifts in the dominance and distribution of different genotypes may frequently occur, resulting in the emergence and spread of varying PCV2 genotypes and recombinant strains in swine. This study aims at identifying the PCV2 genotypes currently circulating in Chile. Seven hundred thirty-eight samples were obtained from 21 swine farms between 2020 and 2021. The samples were tested using PCR for species detection and genotyping. Sequencing and phylogenetic analyses were conducted in selected samples. PCV2 was detected in 26.9% of the PCR reactions and 67% of the sampled farms. The genotypes were determined in nine farms, PCV2a in one farm, PCV2b in four, and PCV2d in five, with PCV2b and PCV2d co-circulating in one farm. The phylogenetic analysis of twelve ORF2 sequences obtained (PCV2a = 5; PCV2b = 4; PCV2d = 3) showed a PCV2a Chilean strains monophyletic cluster; closely related to Chilean viruses collected in 2012 and 2013. Of the three different PCV2b sequenced viruses, two viruses were close to the root of the PCV2b group, whereas the remaining one grouped with a South Korean virus. PCV2d sequences were closely related to Asian viruses. A previously reported PCV2a/PCV2d recombinant strain was not detected in this study. Our results suggest the emergence and potential shift to PCV2d genotype in Chilean farms.
Introduction
Porcine circoviruses (PCVs) are small (12–23 nm), non-enveloped, icosahedral, circular single-stranded DNA viruses (1). Porcine Circovirus 2 (PCV2) is the most studied PCVs species due to its pathogenic potential, associated with multiple clinical conditions (2). These are known as porcine circovirus-associated diseases (PCVAD); including the PCV2 systemic disease, formerly post-weaning multisystemic wasting syndrome (PMWS), and diverse respiratory, reproductive, intestinal, and neurological syndromes (3–5). Although nowadays, the most common PCV2 manifestation is subclinical (5). Currently, PCV2 is widely distributed in domestic and wild swine, negatively affecting pork production (1, 2). Before the availability of vaccines, PCVAD was a major problem for the global swine industry. When uncontrolled, PCV2 losses were estimated at £88 million/year in England only, with subclinical infections causing higher costs than clinical manifestations (5, 6).
PCV2 exhibits evolutionary dynamics similar to RNA viruses, with a nucleotide substitution rate of 1.2 × 10−3 substitutions/site/year and frequent recombination events (7, 8). According to this and the growing number of viral sequences, PCV2 genotype classification is continually changing, and the latest genotyping method assigned eight genotypes: PCV2a to PCV2h (9). Periodic waves of PCV2 genotypes or “genotype shifts” have affected swine; this also occurs in some PCV2 recombinant viruses (8, 10). Initially, the most frequent genotype in affected pigs was PCV2a until the 2000 s, then outclassed by PCV2b and afterward by PCV2d (1). Additionally, the local and global movement of sub-clinical animals allowed the rapid PCV2 spread (7, 8). The remaining genotypes have limited virulence or distribution (8, 9).
The first official detection of PCV2 in Chile occurred in 2006 (11). Between 2005 and 2013, Neira et al. reported the high occurrence of PCV2b (52%) and a PCV2a/PCV2d recombinant lineage (34%) in Chilean swine intensive farms (10). Monitoring the circulating genotypes is critical to understand changes that could lead to lower vaccine protection or higher occurrence of clinical or subclinical disease. Due to the rapid emergence and spread of PCV2 genotypes, this study aims to review the current distribution of PCV2 genetic types in Chile.
Materials and Methods
Samples
The samples were obtained from submissions for PCV2 diagnostic at Universidad de Chile Animal Virology Laboratory between March 2020 to July 2021. The samples were originated for the diagnosis of animals with suspected clinical PCVAD (69) or active molecular surveillance of PCV2 (669). A total of 640 sera, 92 oral fluids, and 6 lung samples were included in this study. Samples were obtained from 21 commercial swine farms (A-U), which represent >90% of the commercial swine companies in the country. Swine farms are located in Valparaiso, Metropolitan, O'Higgins, Maule, and Ñuble regions in Central Chile (Table 1, Supplementary Figure 1). All farms had an active vaccination program against PCV2 in piglets, and in general, did not evidence clinical PCVAD during the sampling. Sera were collected from at least 16 pigs aged 10–20 weeks old in each farm. The oral fluids were collected at a similar age. Samples were stored at 4°C until processing at Animal Virology Laboratory, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile. Sample collection is summarized in Table 1.
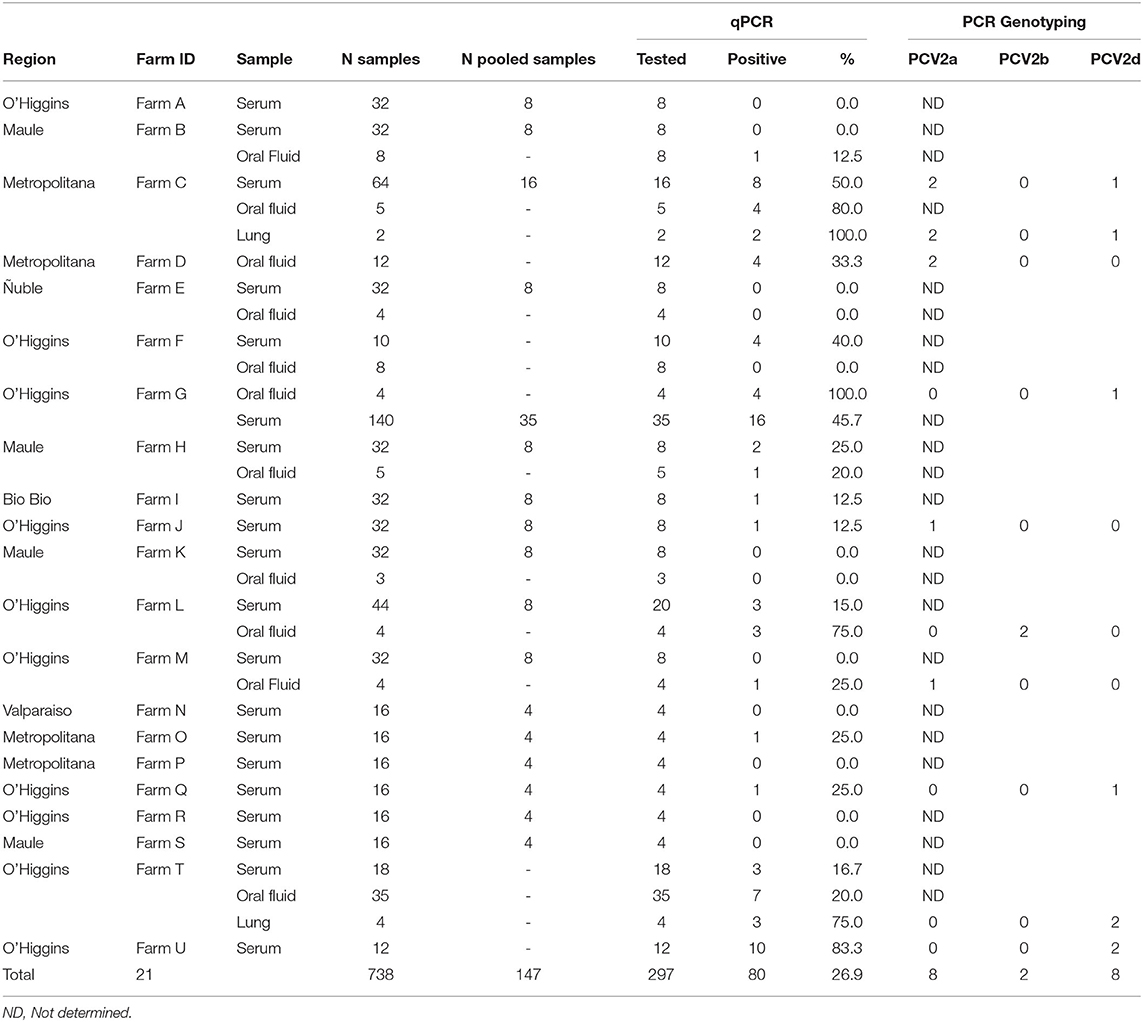
Table 1. Summary of sample collection and PCR results.
Testing and Sequencing
The samples were tested by real-time PCR, genotyped, and sequenced. Briefly, samples were centrifugated at 2,500 g for 10 min. The DNA was extracted individually in lungs and oral fluids samples, whereas sera were pooled (4 samples per pool) before DNA extraction. The extraction was performed using PureLink Genomic DNA Mini Kit (Invitrogen, K182001, CA, USA) following the manufacturer's instructions. All samples were tested by real-time PCR and Ct <40 was considered positive (12). A genotype-specific PCR previously described was used to identify PCV2a, PCV2b, and PCV2d in samples where real-time PCR was Ct < 35 (13). Additionally, in 15 samples selected based on Ct value per case (Ct range: 11.8 to 39.4) ORF2 sequencing was attempted at Veterinary Diagnostic Laboratory, University of Minnesota (12).
Phylogenetic Analysis
Sequences were aligned with publicly available reference sequences from all PCV2 genotypes (a-h) using MUSCLE (14). Two phylogenetic analyses were performed: one including ORF2 partial and complete sequences from Chile (excluding recombinants) and another with only complete ORF2 sequences. The Maximum likelihood (ML) tree was estimated using RAxML (15) with 1,000 bootstrap replications, in Geneious Prime 2021.2 (16) and then visualized in Figtree (http://tree.bio.ed.ac.uk/software/figtree/).
Results and Discussion
Overall results are summarized in Table 1. PCV2 was detected in 14 out of 21 farms, distributed in all the regions in the study, except for Ñuble and Valparaiso. This geographic distribution is similar to previous results (10). Eighty out of 297 PCR reactions (26.9%) were positive for PCV2, corresponding to 25/92 oral fluids (27.2%), 50/199 pooled serum samples (25.1%), and 5/6 lung samples (83.3%). During the study, PCV2 was predominantly detected in oral fluids from 100 to 120 days-old pigs, with and without apparent clinical signs. These results agree with a previous study, where PCV2 detection was higher in oral fluids compared to serum samples from 14 to 16 weeks-old pigs (17). In 19 samples with Ct < 35, belonging to 13 positive farms, PCR genotyping was attempted. Two viruses were identified as PCV2a, eight as PCV2b, eight as PCV2d, and nine samples did not amplify, probably due to the low quantity/quality of PCV2 genetic material. Thus, in nine out of 13 farms at least one of the circulating genotypes was identified. In one farm, two PCV2 genotypes (PCV2b and PCV2d) were identified.
Sequencing was successful in 12 samples from 11 different farms, sequences were deposited in GenBank (Accession numbers OK322701-OK322712). The ORF2 sequencing was successful in all samples with Ct < 30. The phylogenetic tree grouped five isolates into the PCV2a cluster, four in PCV2d, and three in PCV2b clusters (Figure 1, Supplementary Figure 2). Sequences belonging to PCV2a and PCV2d were collected from 4 farms each, and PCV2b sequences from two farms. The PCV2a isolates are grouped in a monophyletic cluster with PCV2a isolates sequenced in Chile in 2012–2013 and are 2.00–6.85% different to reference PCV2a sequences. Interestingly, the current observed PCV2a Chilean isolates were collected from farms related geographically and by pig flow. Two of the three PCV2b sequences were close to the root of the PCV2b genotype cluster, and the remaining one grouped with a sequence from South Korea (EU450638). On the other hand, PCV2d sequences were highly similar (99.43% identity) and grouped with Asian viruses. In contrast with PCV2a, PCV2d viruses were collected from three unrelated farms. Interestingly, we did not detect the PCV2a/PCV2d recombinant strain detected in Chile in a previous study. However, considering the low number of samples sequenced, the current circulation of this recombinant strain cannot be ruled out.
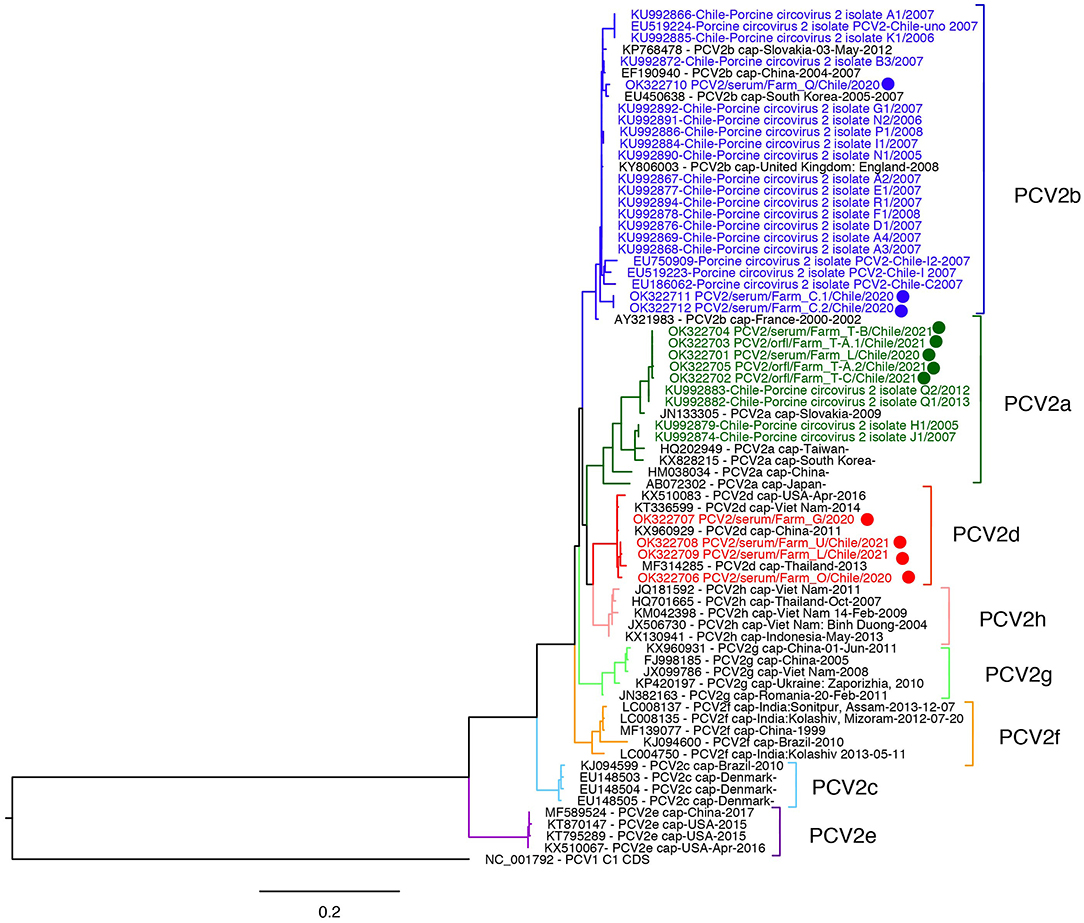
Figure 1. Maximum likelihood phylogenetic tree of Porcine circovirus 2 ORF2 sequences from Chile. The analysis involved 72 sequences, 36 corresponded to PCV2 genotypes (a-h) reference sequences, sequences from Chile from 2005 to 2013 (colored), and sequences from this study (dots). PCV1 (NC001792) was incorporated as an outgroup.
According to a previous report, the PCV2 genotypes occurrence was: 52% of PCV2b from 14 out of 20 intensive farms, 35% of a PCV2a/PCV2d recombinant lineage from seven farms, and 14% of PCV2a from three farms (10). Here, the recombinant strains were not detected, and PCV2b was detected in four farms and successfully sequenced only in two. Also, we observed the emergence of PCV2d and the maintenance of the PCV2a genotype.
In this study, we observed some limitations, essentially the lower detection of PCV2 and the low number of sequences obtained. In this sense, the distribution and occurrence of the detected genotypes may be underestimated in our results. Neira et al. (10), performed the sequencing from PCV2 positive tissues collected from PMWS/PCVAD-affected pigs, but nowadays is difficult to observe the clinical symptoms of PCV2. The lower presence of PMWS/PCVAD-affected pigs is most likely the result of vaccination protocols performed at Chilean farms (11). Vaccination also reduces viremia and shedding during PCV2 infection, limiting viral detection (18).
Overall results of detection and sequencing suggest the potential genotype shift to PCV2d in Chile, although the low number of genotyped samples restricts this conclusion. Similar findings have been reported in recent years in several countries such as the USA, Korea, and Austria (13, 19, 20). This genotype shift may be influenced by natural selection but also by vaccination pressure (21, 22). In this sense, widely administered PCV2 vaccines in Chilean swine since 2008 may have contributed to PCV2 evolution. As previously mentioned, PCV2 genotype shift has been reported in several countries. However, in South America, PCV2d has been reported on three occasions: one during an outbreak of PMWS/PCVAD in vaccinated pigs from Brazil, that suggested a vaccine failure; another from vaccinated and unvaccinated pigs from Uruguay; and the last from unvaccinated pigs with and without clinical signs in Colombia (23–25). Further research on these viruses is necessary to understand the impact of these genotype shifts in the swine industry.
In conclusion, we did not detect a previously reported prevalent PCV2a/PCV2d recombinant strain. Our results suggest the emergence and potential shift to PCV2d genotype in Chilean farms, as in other parts of the world, highlighting the rapid evolution of PCV2 and the relevance of pathogen surveillance and monitoring to the early detection of new genotypes.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval was not required for the animal study because samples were obtained from clinical cases by veterinary practitioners; therefore, ethical approval was not considered. Written informed consent for participation was not obtained from the owners because samples were obtained from clinical cases by veterinary practitioners; therefore, consent forms were not considered.
Author Contributions
NA, FB, BA, and VN: study design and conceptualization. SB and VN: funding and resources. BA, FB, and VV: samples collection and processing. BA, FB, VV, BB, and SM: performed the essays. NA, BB, and VN: data analysis and NA, BA, BB, and VN: wrote the paper. All authors critically evaluated the paper.
Funding
This study was partially funded by the Animal Virology Laboratory, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile; Programa Fondecyt N° 11170877 and N° 1211517 to VN; Programa Fondef N° ID19I10135 to SB and VN.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to the staff of the Animal Virology Laboratory, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile, for all their support in sample processing, especially to Benjamin Bennett and Vanesa Mendieta Reis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.789491/full#supplementary-material
References
1. Segalés J, Allan GM, Domingo M. “Circoviruses,” in Diseases of Swine. Wiley. (2019) 473–87. doi: 10.1002/9781119350927.ch30
2. Opriessnig T, Karuppannan AK, Castro AMMG, Xiao CT. Porcine circoviruses: current status, knowledge gaps and challenges. Virus Res. (2020) 286:198044. doi: 10.1016/j.virusres.2020.198044
3. Palinski R, Piñeyro P, Shang P, Yuan F, Guo R, Fang Y, et al. A novel porcine circovirus distantly related to known circoviruses is associated with porcine dermatitis and nephropathy syndrome and reproductive failure. Virol J. (2017) 91: doi: 10.1128/JVI.01879-16
4. Zhai S-L, Chen S-N, Xu Z-H, Tang M-H, Wang F-G, Li X-J, et al. Porcine circovirus type 2 in China: an update on and insights to its prevalence and control. Virol J. (2014) 11:88. doi: 10.1186/1743-422X-11-88
5. Segalés J. Porcine circovirus type 2 (PCV2) infections: clinical signs, pathology and laboratory diagnosis. Virus Res. (2012) 164:10–9. doi: 10.1016/j.virusres.2011.10.007
6. Alarcon P, Rushton J, Wieland B. Cost of post-weaning multi-systemic wasting syndrome and porcine circovirus type-2 subclinical infection in England–an economic disease model. Prev Vet Med. (2013) 110:88–102. doi: 10.1016/j.prevetmed.2013.02.010
7. Firth C, Charleston MA, Duffy S, Shapiro B, Holmes EC. Insights into the evolutionary history of an emerging livestock pathogen: porcine circovirus 2. J Virol. (2009) 83:12813–21. doi: 10.1128/JVI.01719-09
8. Franzo G, Cortey M, Segalés J, Hughes J, Drigo M. Phylodynamic analysis of porcine circovirus type 2 reveals global waves of emerging genotypes and the circulation of recombinant forms. Mol Phylogenet Evol. (2016) 100:269–80. doi: 10.1016/j.ympev.2016.04.028
9. Franzo G, Segalés J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE. (2018) 13:1–12. doi: 10.1371/journal.pone.0208585
10. Neira V, Ramos N, Tapia R, Arbiza J, Neira-Carrillo A, Quezada M, et al. Genetic analysis of porcine circovirus type 2 from pigs affected with PMWS in Chile reveals intergenotypic recombination. J Virol. (2017) 14. doi: 10.1186/s12985-017-0850-1
11. Estrada A. Síndrome de desmedro multisistémico posdestete (PMWS). Boletín Veterinario Oficial, Servicio Agrícola y Ganadero (SAG). (2009). Available online at: https://www2.sag.gob.cl/Pecuaria/bvo/BVO_9_I_semestre_2009/articulos/PMWS.pdf (Accessed December 17, 2020)
12. Davies B, Wang X, Dvorak CMT, Marthaler D, Murtaugh MP. Diagnostic phylogenetics reveals a new porcine circovirus 2 cluster. Virus Res. (2016) 217:32–7. doi: 10.1016/j.virusres.2016.02.010
13. Kwon T, Lee DU, Yoo SJ, Je SH, Shin JY, Lyoo YS. Genotypic diversity of porcine circovirus type 2 (PCV2) and genotype shift to PCV2d in Korean pig population. Virus Res. (2017) 228:24–9. doi: 10.1016/j.virusres.2016.11.015
14. Edgar RC, MUSCLE. Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. (2004) 32:1792–7. doi: 10.1093/nar/gkh340
15. Stamatakis A. {RAxML} version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. (2014) 30:1312–3. doi: 10.1093/bioinformatics/btu033
16. Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. (2012) 28:1647–9. doi: 10.1093/bioinformatics/bts199
17. Oliver-Ferrando S, Segalés J, López-Soria S, Callén A, Merdy O, Joisel F, Sibila M. Evaluation of natural porcine circovirus type 2 (PCV2) subclinical infection and seroconversion dynamics in piglets vaccinated at different ages. Veterinary Research. (2016) 47:1–11. doi: 10.1186/s13567-016-0405-2
18. Fort M, Sibila M, Pérez-Martín E, Nofrarías M, Mateu E, Segalés J. One dose of a porcine circovirus 2 (PCV2) sub-unit vaccine administered to 3-week-old conventional piglets elicits cell-mediated immunity and significantly reduces PCV2 viremia in an experimental model. Vaccine. (2009) 27:4031–7. doi: 10.1016/j.vaccine.2009.04.028
19. Xiao C-T, Halbur PG, Opriessnig T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. Journal of General Virology. (2015) 96:1830–41. doi: 10.1099/vir.0.000100
20. Weissenbacher-Lang C, Kristen T, Mendel V, Brunthaler R, Schwarz L, Weissenböck H. Porcine circovirus type 2 (PCV2) genotyping in Austrian pigs in the years 2002 to 2017. BMC Veterinary Research. (2020) 16:1–8. doi: 10.1186/s12917-020-02413-4
21. Franzo G, Tucciarone CM, Cecchinato M, Drigo M. Porcine circovirus type 2 (PCV2) evolution before and after the vaccination introduction: A large scale epidemiological study. Sci Rep. (2016) 6:39458. doi: 10.1038/srep39458
22. Reiner G, Hofmeister R, Willems H. Genetic variability of porcine circovirus 2 (PCV2) field isolates from vaccinated and non-vaccinated pig herds in Germany. Vet Microbiol. (2015) 180:41–8. doi: 10.1016/j.vetmic.2015.07.026
23. Salgado RL, Vidigal PMP, de Souza LFL, Onofre TS, Gonzaga NF, Eller MR, et al. Identification of an emergent porcine circovirus-2 in vaccinated pigs from a Brazilian farm during a postweaning multisystemic wasting syndrome outbreak. Genome Announcements. (2014) 2. doi: 10.1128/genomeA.00163-14
24. Almario-Leiva G, Suarez-Mesa R, Uribe-García F, Rondón-Barragán I. Detection and characterization of porcine circovirus type 2 (PCV2) circulating in pigs of the departments of Tolima and Huila, Colombia. Revista de Investigaciones Veterinarias del Peru. (2020) 31:e17553. doi: 10.15381/rivep.v31i1.17553
Keywords: PCV2, genetic diversity, PCV2 genotypes, Chile, genotype shift
Citation: Ariyama N, Agüero B, Valdés V, Berrios F, Bucarey S, Mor S, Brito B and Neira V (2021) Update of Genetic Diversity of Porcine Circovirus Type 2 in Chile Evidences the Emergence of PCV2d Genotype. Front. Vet. Sci. 8:789491. doi: 10.3389/fvets.2021.789491
Received: 05 October 2021; Accepted: 23 November 2021;
Published: 17 December 2021.
Edited by:
Jesus Hernandez, Consejo Nacional de Ciencia y Tecnología (CONACYT), MexicoReviewed by:
Javier Martinez Lobo, Universitat de Lleida, SpainJean-Pierre Frossard, Animal and Plant Health Agency, United Kingdom
Giovanni Franzo, University of Padua, Italy
Copyright © 2021 Ariyama, Agüero, Valdés, Berrios, Bucarey, Mor, Brito and Neira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Victor Neira, dmljdG9ybmVpcmFAdS51Y2hpbGUuY2w=; dm5laXJhcmFtQGdtYWlsLmNvbQ==