 Nara Santos dos Santos1†
Nara Santos dos Santos1† Flaviane Alves de Pinho1,2†Nicole Regina Capacchi Hlavac2
Flaviane Alves de Pinho1,2†Nicole Regina Capacchi Hlavac2 Talyta Lins Nunes2Nádia Rossi Almeida3Manuela Silva Solcà3Bruno Milen Varjão1
Talyta Lins Nunes2Nádia Rossi Almeida3Manuela Silva Solcà3Bruno Milen Varjão1 Ricardo Wagner Portela4
Ricardo Wagner Portela4 Jeronimo Nunes Rugani5
Jeronimo Nunes Rugani5 Felipe Dutra Rêgo5
Felipe Dutra Rêgo5 Stella Maria Barrouin-Melo1,2*‡
Stella Maria Barrouin-Melo1,2*‡ Rodrigo Pedro Soares5*‡
Rodrigo Pedro Soares5*‡- 1Laboratory of Veterinary Infectious Diseases, Teaching Hospital of Veterinary Medicine, Federal University of Bahia (UFBA), Salvador, Brazil
- 2Department of Veterinary Anatomy, Pathology and Clinics of the School of Veterinary Medicine and Zootechny, Federal University of Bahia, Salvador, Brazil
- 3Department of Veterinary Preventive Medicine and Animal Production of the School of Veterinary Medicine and Zootechny, Federal University of Bahia, Salvador, Brazil
- 4Laboratory of Immunology and Molecular Biology, Institute of Health Sciences, Federal University of Bahia, Salvador, BA, Brazil
- 5Instituto Rene Rachou, Fundação Oswaldo Cruz, Belo Horizonte, Brazil
Zoonotic leishmaniasis caused by Leishmania infantum is a disease of One Health concern since human and animal cases and environmental damage are interconnected. L. infantum has a complex epidemiological cycle with multiple hosts, including mammals—humans, domestic, and wild animals—and arthropod vectors. Knowledge on mammal infections in endemic areas is crucial for developing control strategies. This work aimed to detect and characterize L. infantum infection in domestic cats from areas where human and canine leishmaniasis cases occur. No cases of feline leishmaniasis (FeL) had been previously reported in those areas. Five municipalities from Bahia state were chosen, comprising 2,480.8 km2 with 1,103,866 inhabitants. Ninety domiciliated and/or sheltered cats underwent clinical examination and serology by a rapid reference test recommended by the Brazilian government. Cytology, PCR, and parasite DNA sequencing were performed in bone marrow samples. Rapid tests detected antibodies in 5.6% (5/90) of the cats. Leishmania infantum infection was confirmed in 7.8% (7/90) of the cats by PCR, sequencing, and parasite isolation. Three out of the five municipalities (60%) had infected cats, and PCR positivity varied from 6.9 to 29%. One cat was categorized as harboring active L. infantum infection with amastigote forms in bone marrow smears. No clinical signs were detected at the first clinical exam, but 1 month later the cat developed severe FeL. The cat isolate was grown in culture, typed and its DNA sequence was homologous to the L. infantum reference strain (PP75). In conclusion, cats are potential hosts and may acquire L. infantum in endemic areas where canine and human cases occur. For cats, the need for surveillance, differential diagnosis and clinical care is highly recommended since a fast clinical progression of FeL developed in a subclinical animal. An accurate standardized immunodiagnostic assay for FeL is warranted.
Introduction
Leishmaniasis is a vector-borne disease affecting humans and animals caused by more than 20 species of the protozoan Leishmania (1). They are endemic in 92 countries, where more than one billion people are at risk of infection (2). Depending on the region, Leishmania is transmitted by different phlebotomine sand fly vectors (3). There is no human vaccine (4, 5) and the prevalence of leishmaniasis is closely linked to the human development index and environmental degradation (6, 7). This zoonosis remains a serious public health problem and is recognized as one of the 20 major neglected tropical diseases (8). The World Organization for Animal Health also lists leishmaniasis among the diseases that require worldwide notification (9). Visceral leishmaniasis (VL) is the most fatal form of leishmaniasis in humans caused by Leishmania donovani/Leishmania infantum (Old World) and Leishmania chagasi (same as L. infantum) in the New World (10).
In southern Europe, Africa, Asia, and America, L. infantum causes zoonotic leishmaniasis in humans (VL) (11–13) and in domestic dogs (CanL) (14, 15). Leishmania infantum infection has also been detected in wild animals (16, 17) and other domestic species including cats, horses, cattle, sheep, and goats (18–20). Natural infection and disease have been described in domestic cats to a lesser extent than in dogs (21–23). A recent meta-analysis including 36 years of cross-sectional studies from 12,635 cats in Leishmania endemic areas worldwide found an estimated seroprevalence of 11% and an overall PCR positivity of 10%. At least 63% of the infections were caused by L. infantum (24). Other studies detected 3–4% of cats with L. infantum in Italy, Greece, and German by serology (25, 26). Those recent studies have demonstrated that Leishmania is also circulating and cats and this should be taken as an alert in the regions where this parasite occurs.
In South America, the epidemiological cycle of L. infantum is complex and involves wild and domestic vertebrate and invertebrate hosts (27–30) as a result of social/economic inequality and environmental changes (31). CanL is a severe zoonosis because the dog is highly susceptible exhibiting elevated parasite loads and disease. Besides, it is very close to humans acting as a competent domestic reservoir (32, 33). In Brazil, seroprevalence in dogs and cats ranges from 2.4 to 32% (34) and 0.7 to 30.48%, respectively (35–37). However, the role of cats in L. infantum epidemiology is still in its infancy and should be more explored.
Recent investigations have focused on domestic cats' potential in L. infantum transmission (22, 38). Standardized serological tests for cats and a more detailed knowledge on their immune responses against L. infantum infection are needed (39) since coinfections may predispose cats to develop feline leishmaniasis (FeL) and other diseases (18, 40). These relevant aspects would improve the pivotal role of cats as L. infantum hosts in the context of One Health. As part of a wider study on leishmaniasis epidemiology, natural feline L. infantum infection from VL and CanL endemic areas is reported. Also, the clinical presentation and natural evolution of an infected cat is provided in detail.
Materials and Methods
Study Area, Animals, and Ethical Aspects
This study was conducted from July 2018 to December 2019 in five municipalities of Bahia state, Brazil. Those included Camaçari, Feira de Santana, Muritiba, São Félix, and Simões Filho. They are located within a 100 km radius from Salvador, the state's capital (Figure 1), comprising an area of 2,480,800 km2 containing a population of 1,103,866 inhabitants as estimated in 2020 (Table 1) (41). In 2010, their Human Development Index (HDI) ranged from of 0.660 to 0.712 (Table 1) (41) and, from 2009 to 2018, total human VL cases ranged from 3 to 101 (42). Feline and canine populations in the area are estimated at 2,800 to 120,000 and at 1,700 to 72,000, respectively (43).
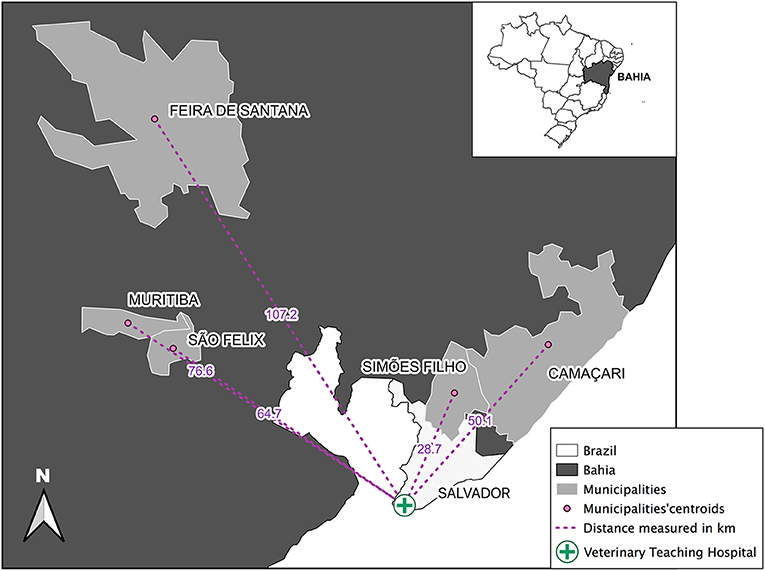
Figure 1. Geographical location of the surveyed municipalities (Feira de Santana, Muritiba, São Felix, Simões Filho, and Camaçari) in the State of Bahia, Brazil. Distance in kilometers (km) from each municipalities' centroids (pink dots) to the Federal University of Bahia Veterinary Teaching Hospital (green cross) were measured (pink dotted lines). Source: IBGE 2010, Datum: SIRGAS, 2000.
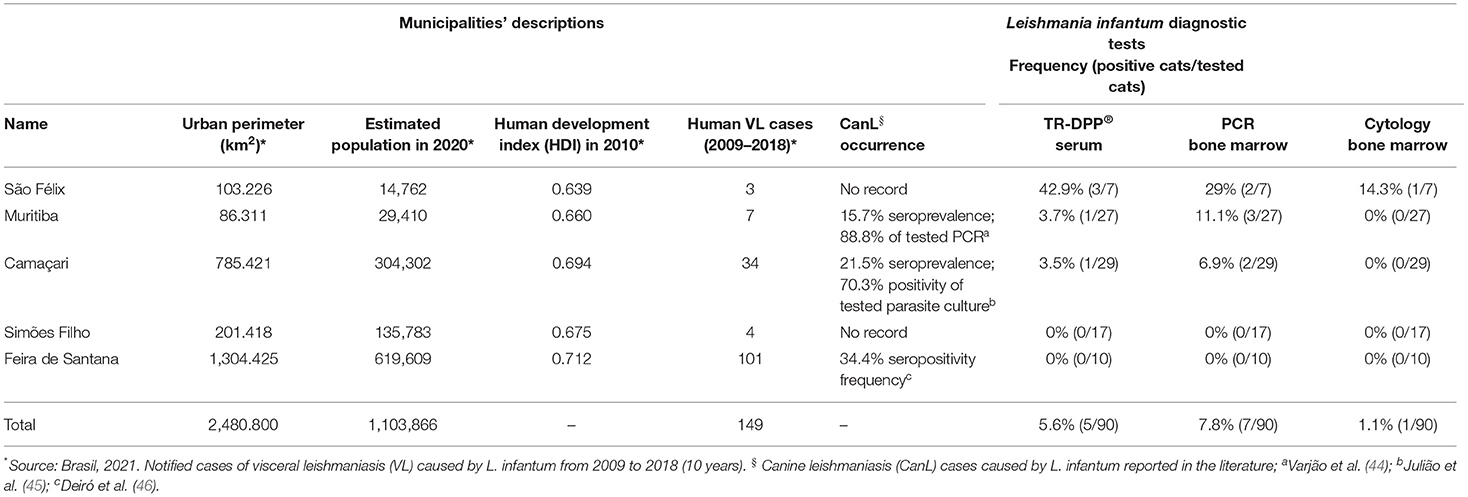
Table 1. Demographic parameters and feline infection (%) by L. infantum by different diagnostic methods performed in 90 cats from leishmaniasis endemic regions in Bahia state, Brazil between July, 2018 and December, 2019.
In these municipalities, VL cases have been officially reported from 2009 to 2018 (42). According to the Health Ministry classification (47), Feira de Santana is the only considered an area of intense VL transmission, whereas the others are sporadic. However, CanL cases amongst the veterinary community of all municipalities is a concern.
A convenience sample of 90 cats including domiciliated (75) and sheltered (14), regardless of breed and sex, was evaluated. An active search was conducted, and 29 households and one animal shelter were included. Inclusion in the study was based on the proximity of a given CanL case either within the household/shelter or in a neighboring house. Eligibility criteria for cats in the study were: at least 6 months of age, and a signed informed consent by the owner. The exclusion criteria were: pregnancy, body weight under 2 kg and/or irascible behavior during handling.
All procedures were approved by the Committee for Ethics on the Use of Animals of the School of Veterinary Medicine and Zootechny from the Federal University of Bahia (Permission number CEUA-13/2017). The study was designed and conducted following the Brazilian Council of Animal Experimentation guidelines, and there was strict adherence to the Brazilian law for “Procedures for the Scientific Use of Animals” (No. 11.794/2008).
Clinical Examination and Sample Collection
The cats underwent clinical evaluation including anamnesis regarding their habits and previous health conditions. Also, physical examination assessing FeL clinical signs as described by the LeishVet group was performed (48). Peripheral blood samples were collected for serology. After sedation with acepromazine (0.05 mg/kg) and meperidine (3 mg/kg), local anesthesia with lidocaine was used for bone marrow aspiration sampling at the iliac crest (49). Part of the bone marrow aspirates was used for cytological preparation; the rest was stored in RNAse and DNAse free microtubes at −20°C prior to DNA extraction. Cats with enlarged superficial lymph nodes and/or skin lesions underwent sampling by fine-needle aspiration.
Cytological Examination
Smears of fine-needle aspiration of bone marrow, lymph nodes, and skin lesions were stained by the Romanowsky method (Panótico Rápido®, Laborclin, São Paulo) for optical microscopic search of Leishmania amastigote forms under 1,000 × magnification (Olympus, CX22LED).
Serology
Because there are no standardized serological tests for cats, the reaction of anti-Leishmania antibodies was assessed using the canine Dual-Path Platform immunochromatographic test (TR-DPP® Biomanguinhos, Fiocruz, BR). It is a reference assay recommended by the Brazilian Ministry of Agriculture for the serodiagnosis of CanL having a sensitivity and specificity of 86 and 94%, respectively (50). It detects the reaction of total IgG against fixed recombinant L. infantum k28 protein as antigen using staphylococcal A protein linked to colloidal gold as conjugate (51). Protein A is also capable of binding to cat IgGs (52). A previous study reported the use of the TR-DPP® test for the serodiagnosis of Leishmania infection in feline sera samples (53). Reactions were conducted according to the manufacturer's instructions. We interpreted TR-DPP® positive results as an exposure to L. infantum but not as markers of active infection or disease.
Molecular Procedures
Total DNA from bone marrow samples and parasite cultures were extracted using Wizard® Genomic DNA Purification Kit (Promega®, USA), according to the manufacturer's recommendations. Bone marrow DNA samples were used to amplify the endogenous control glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (54) and L. infantum kinetoplast DNA with the RV1 and RV2 primers (55). Amplified fragments were observed in a 2% agarose gel stained with ethidium bromide (0.5 μg/mL). RV1-RV2 amplicons were sequenced and the phylogenetic trees were built using MEGA X (56). To identify the isolated strains, HSP70 and ITS1 PCR-RFLP fragments were performed (57, 58). Leishmania reference strains for outgroup comparison included Leishmania braziliensis (MHOM/BR/75/M2903), Leishmania guyanensis (MHOM/BR/75/M4147), Leishmania major (MHOM/IL/81/Friedlin), Leishmania amazonensis (IFLA/BR/67/PH8), and L. infantum (MHOM/BR/74/PP75) (59).
Data Analysis
The data obtained in this study were tabulated in Microsoft Excel 2000 to calculate variable frequencies and perform descriptive analysis. A positive criterion for FeL was the amplification of parasite DNA by PCR in bone marrow samples. Active infection in PCR positive cats was defined by the identification of amastigotes (bone marrow, lymph node or skin) during cytological examination. Maps were constructed using the QGIS v3.8 program to display studied areas where FeL cases occurred.
Feline L. infantum Active Infection
One cat with an active infection detected by cytological examination during the first investigation (T1) was clinically followed. It was evaluated at 1 month (T2) and three months (T3) after the second evaluation totalizing 4 months. At T2 and T3, hemogram, biochemical profile, and urinalysis were performed. Serological, parasitological, and molecular diagnosis of L. infantum infection were performed as described above, with the addition of a conjunctival swab collected with a sterile brush for cytological examination. Parasite load was assessed at T1 using duplex qPCR for simultaneous detection of L. infantum kinetoplast DNA (kDNA) and a conserved region of the housekeeping gene 18S rRNA. The results were expressed as parasites/mL as previously reported (60). Leishmania infantum isolated from bone marrow aspirates was grown in NNN-Schneider and expanded in M199 medium (61, 62) prior to DNA extraction and sequencing. The cat was also tested for feline viral leukemia virus (FeLV) and feline immunodeficiency virus (FIV) by a commercial test (SNAP Combo Test, IDEXX Laboratories) at the three evaluation points.
Results
Cat Population, L. infantum Infection, and Sequencing
All 90 samples from bone marrow were analyzed by PCR and 7.8% (7/90) were positive for L. infantum DNA. Those cats were then diagnosed as infected. The serological test TR-DPP® detected anti-Leishmania antibodies in the sera of 5.6% (5/90) of the cats. However, L. infantum infection was not identified by cytological and/or molecular techniques in four of them. The frequency of PCR positive results within municipalities ranged from zero to 29%. Table 1 shows the results of diagnostic tests with different types of examined biological samples. In the cat population, 77.8% (70/90) were domiciliated, 52% (47/90) males, 86.7% (77/90) adults, 96.7% (87/90) were mixed breed, and 65.6% (59/90) exhibited at least one clinical sign of disease. Table 2 shows the phenotypic characteristics and health status during physical examination of the 90 cats per locality.
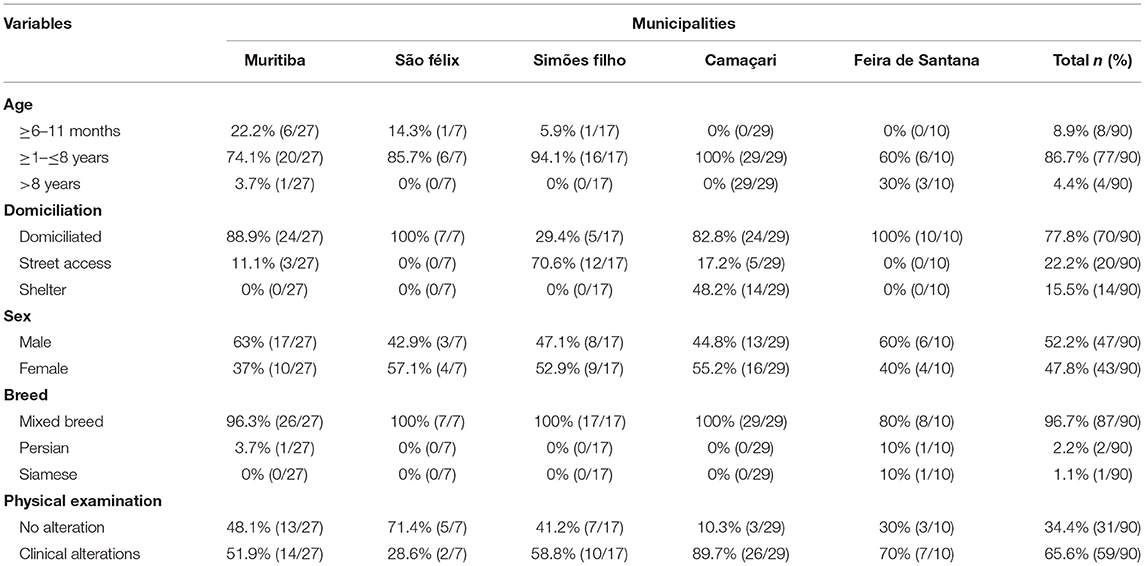
Table 2. Parameters of the feline population in the five surveyed municipalities for L. infantum in the state of Bahia, Brazil (July, 2018 to December, 2019).
One of the PCR and DPP positive cats had Leishmania amastigote forms in its bone marrow aspirate (Figure 3D). This cat was the only categorized as having L. infantum active infection. There were no amastigote forms in the cytological examinations (bone marrow, lymph nodes and skin) of the remaining 89 cats. By using DNA extracted from bone marrow samples, PCR was performed using RV1–RV2 primers (Figure 2A). All positive samples exhibited the expected 145-bp fragment. After sequencing and phylogenetic analysis, those products clustered together with the L. infantum control (strain PP75). To confirm the taxonomic status of the Leishmania species infecting the only cat that we could isolated the parasite (CAT 1), two additional PCR-RFLP were performed (ITS1 and HSP70) (Figures 2B,C). Confirming the previous phylogenetic tree, the isolated strain (lane 6) exhibited the same restriction profile as the L. infantum reference strain PP75 (lane 5).
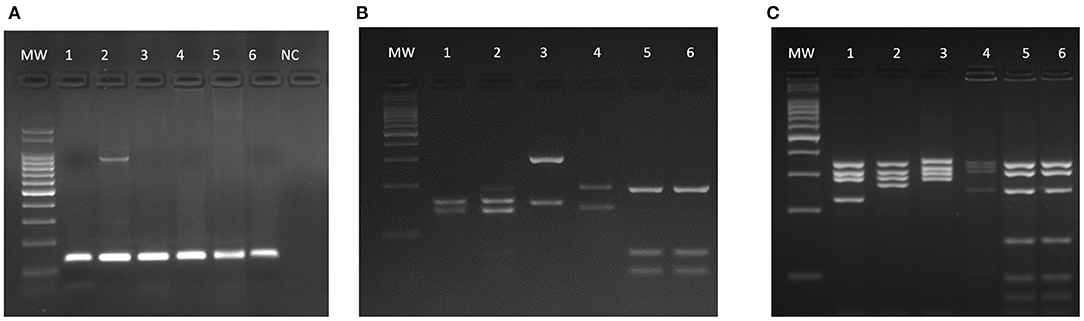
Figure 2. Molecular identification of L. infantum isolated from a cat (CAT1). Agarose gels showing the expected 145-bp fragment for RV1-RV2 PCR (A) and PCR-RFLP restriction profiles of ITS1 (B) and HSP70 (C) fragments. MW −100-bp molecular weight; 1–L. braziliensis (MHOM/BR/75/M2903); 2–L. guyanensis (MHOM/BR/75/M4147); 3–L. major (MHOM/IL/81/Friedlin); 4–L. amazonensis (IFLA/BR/67/PH8); 5–L. infantum (MHOM/BR/74/PP75); 6–CAT1 (isolated from cat); NC–negative control.
Clinical Characterization of the Studied Cats
Three out of the seven cats with confirmed L. infantum infection by PCR had at least one suggestive clinical sign of FeL during physical examination. These alterations included weight loss, dehydration, crusted lesions on the tips of the ears, and/or areas of alopecia (Figure 3A) in the dorsal cervical region, sneezing, enlarged popliteal lymph nodes, gingivitis, and ulcers in the oral cavity suggesting gingivitis-stomatitis complex (Figure 3B).
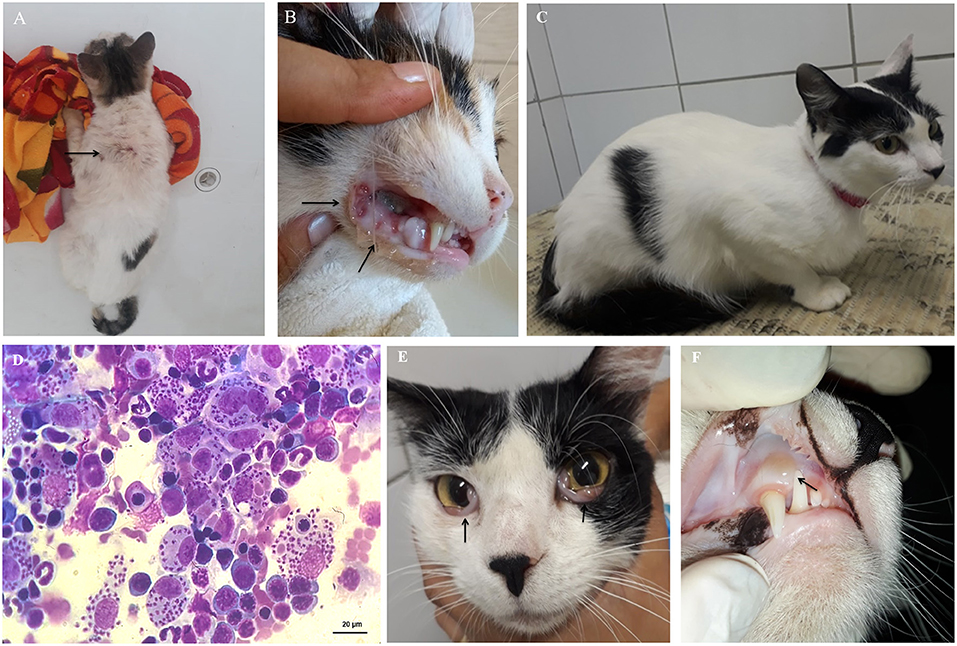
Figure 3. Clinical signs of feline leishmaniasis. (A) Areas of alopecia in the dorsal cervical region (arrow), and (B) ulcers in the oral cavity, suggesting gingivitis-stomatitis complex (arrows). (C) First physical exam of a cat with active infection without clinical signs but presenting Leishmania detection in bone marrow aspirates by cytological examination (D). After 4 months, this cat had (E) bilateral oedema and hyperaemia of the conjunctiva (arrows) and (F) aphthous stomatitis in the upper region of the right canine tooth (arrow).
Clinical assessment (anamnesis and physical examination) of the 83 cats diagnosed as not infected by Leishmania—since they were not positive either in the TR-DPP® or in the PCR test—resulted in 26.5% (22/83) of the cats with no alteration, and 73.5% (61/83) with variable clinical alterations. Signs of disease included cutaneous changes (33/83, 39.8%) characterized by focal alopecia, ulcers, crusts, abrasions, and furfuraceous dermatosis, dehydration (22/83, 26.5%), or weight loss (20/83, 24.0%) among others less frequent.
Clinical and laboratory findings in the female, mixed-breed, neutered 3-year-old pet cat described as having developed an active infection are displayed in the Supplementary Table 1, representing data from evaluations performed at three times, namely T1, T2, and T3. The first evaluation (T1) was carried out in the field. The cat was retrospectively diagnosed as having subclinical infection (PCR positivity but no clinical signs) (Figure 3C). Because there were numerous visible amastigote forms of Leishmania in the cytological examination of the bone marrow sample (Figure 3D), and a high parasitic load was found in the qPCR (7.7 ×108 parasites/mL), the cat was referred to a thorough clinical diagnosis and the owner was instructed to keep a sandfly repellent collar in the cat. One month after the field evaluation, the cat was brought to the clinical examination (T2) exhibiting signs of dehydration, lymph node enlargement and a skin ulcer. Cytological preparations of bone marrow and conjunctival samples were positive for amastigote forms, and serum biochemistry showed increased total serum proteins [7.9 g/dL; reference range (RR) = 5.4 to 7.8 g/dL] and gamma-glutamyl transferase (GGT) (6.3 U/L; RR = 1.5 to 5.3 U/L) (63, 64) at T2. Although the cat was referred for treatment of FeL, the owner did not return until 3 months later (T3). At T3, the cat exhibited bilateral oedema and hyperaemia of the conjunctiva (Figure 3E), presence of aphthous stomatitis (Figure 3F), and enlarged lymph nodes. Hemogram evidenced lymphocytosis and serum biochemistry, hyperproteinaemia associated with hyperglobulinemia and increased serum alkaline phosphatase. At T3, Leishmania amastigotes were found in the cytological examination of multiple samples such as lymph nodes, palpebral conjunctiva, and bone marrow. Serological test TR-DPP® was performed during the three evaluations but resulted positive only for the serum sample collected at T2. The cat was also negative for FIV and FeLV retroviruses as tested by the lateral flow assay (SNAP Combo Test, IDEXX Laboratories) in serum samples collected at T1, T2, and T3.
Discussion
This is the first study to report the infection by L. infantum in cats in the state of Bahia, Brazil, where 7.8% of cats were infected by L. infantum. The finding of L. infantum DNA by PCR in the bone marrow of the tested cats indicating parasite circulation simultaneously with CanL and VL that have been notified in the last 12 years (44, 65, 66). Comparatively, in other endemic Brazilian regions for both VL and CanL with similar socioeconomic and environmental characteristics, the frequency of L. infantum infection in cats ranged from 4.0 to 31.9% suggesting that it my vary depending on the region (21, 37, 67, 68).
Brazilian health authorities declared one of the studied municipalities (Feira de Santana) as an area of intense VL transmission, and an average of 20.2 human cases have been reported from 2014 to 2018 (42). Endemicity of zoonotic leishmaniasis in that municipality had been assessed in a former study reporting 0.6 to 2.1% of “a global prevalence of canine cases” between 1995 and 2000 (69). A recent study on CanL serology reported a seroprevalence of 34.4% in the same area (46). Although this municipality has favourable characteristics for harbouring FeL, a low adherence from the households to participate in the study was noticed. Only one pet-owner agreed to participate, and his cat was negative. This might be a result of the better knowledge of the control measures used for positive dogs. Since this is an intense VL/CanL transmission area, probably the households were aware of Brazilian health authorities' control measures. This would favour underreporting and hinder conclusions in a particular area.
There were positive cats in municipalities considered sporadic VL transmission including São Félix, Muritiba, and Camaçari (Table 1). In Muritiba, a recent study reported on a seroprevalence of 15.7% of dogs with CanL (44) whereas in Camaçari, where 11 VL cases were notified, CanL seroprevalence was 19.8% (70). In São Félix and Simões Filho, where <3 VL cases between 2014 and 2018 were notified, no available data on CanL was found. Despite gaps in notification by the health authorities, there is strong evidence of endemic zoonotic circulation of the parasite in these municipalities where local veterinarians have been increasingly reporting CanL. Consistent with these observations, the highest frequency of L. infantum infection found in the present study was in one of these localities (São Félix). Perhaps the lack of positivity in two of the municipalities investigated in this study was because of the low number of animals and households evaluated.
PCR was chosen as a confirmatory method of infection due to its high sensitivity and ability to confirm parasite species (71, 72). Among the 90 examined cats from a highly endemic area, the frequency found of L. infantum positivity was 7.9%. Moreover, only one out of seven infected cats developed FeL, suggesting that they may not be as susceptible to infection as dogs. In this context, other studies suggested that felines appear to be resistant to infection by L. infantum (73, 74). A lower tissue parasitism in cats compared to dogs might hinder cytological examination less accurate during diagnosis. It has been demonstrated in previous studies on dogs that the spleen is the best choice for a more sensitive parasitological or molecular detection of L. infantum (14, 71, 75). Moreover, studies have showed that a higher parasitic load in the canine host is associated with more intense disease, and that these more susceptible dogs are more infectious to the phlebotomine vectors (33, 76). Factors such as the parasite's tissue tropism and intrinsic pathophysiological mechanisms make organs such as the spleen more intensely parasitised than other ones in infected dogs (77–79). Thus, in dogs, the use of samples from such organs is likely to improve diagnostic sensitivity of methods based on the detection of parasites and/or its DNA (71, 75, 80). All these aspects have crucial importance under the view of One Health and remain to be more investigated in cats in the diverse endemic regions for L. infantum infection not only in Brazil, but also in other countries. Here, we advocate that feline bone marrow could be one of the target organs for FeL since we detected a higher parasite load in infected macrophages (Figure 3C). Bone marrow is commonly a chosen target for sampling tissue for PCR diagnosis because it is known to be consistently parasitised in dogs (71) and humans (81) with Leishmania active infection. Bone marrow has been shown to be a suitable sample for the diagnosis of L. infantum infection in cats by PCR (73, 82). Bone marrow collection under proper sedation also causes a minor impact on the welfare of cats (49). Consistent with those observations, the present study reports the identification of L. infantum in different cats by using several PCR techniques and sequencing.
Five cats tested positive in TR DPP®, but no parasite DNA was detected in the bone marrow in four of them. These data may suggest a natural lesser susceptibility in cats whose residual antibody response might only indicate a transient parasite exposure. Besides, cross-reactions with other pathogens should be considered, as already reported for dogs (83, 84). Susceptible canine hosts develop high antibody levels as the infection progresses, together with the development of clinical alterations and a higher chance to infect vectors (66, 79, 85, 86). Based on our data, this does not seem to happen in cats. Furthermore, the TR-DPP® used in the present study might not have detected feline immunoglobulins against L. infantum which would certainly exist as a consequence of the constant exposure to infected vectors (87). Indeed, among the seven cats that were positive for L. infantum DNA detection in the PCR test, only one was seropositive in the TR DPP®. Moreover, the only cat with active infection described herein had a negative result in the TR DPP® assay at the very moment when it was exhibiting overt signs of severe FeL. We emphasise that we have evaluated cats that were highly exposed to infection because of their close coexistence with infected dogs in areas of high transmission. Comparatively, a study on L. infantum naturally infected cats reported a considerable 75%-seropositivity for anti-Leishmania immunoglobulins as detected by an in-house indirect ELISA test (22). Thus, the lateral flow assay used in this study does not seem to be accurate for detecting feline antibodies against Leishmania antigens. This reinforces the need of more studies on immune responses during FeL and the development of more specific tests for L. infantum detection in cats.
Typically, because of the PCR's high sensitivity to detect L. infantum DNA, a positive result can be obtained with minimum amounts of parasites from a host sample (88). In this sense, a positive PCR result may indicate an initial infection or even an infection under the host immune system's resolution process. To characterize an active infection that may progress to the clinical disease, cytological examinations were performed on different tissue samples from the seven PCR positive cats, and of the remaining four cats that were positive only in the lateral flow assay. The molecular approaches confirmed the isolated strain to be L. infantum. This reinforces that several diagnostic methods should be employed for a correct diagnosis not only in dogs but especially in cats when a specific method warrants further development.
Three L. infantum-infected (PCR-positive) cats showed clinical signs ranging from cutaneous to systemic alterations suggestive of FeL as previously described (18). However, there was no laboratory evidence of active infection as established in our diagnosis criteria. One of these cats exhibited gum lesions indicative of a gingivitis-stomatitis complex. This cat lived in a shelter in close contact with other cats and dogs, some with diverse diseases. Living in an overpopulated environment is a well-described stress factor for cats. This favours immunosuppression and susceptibility to pathogens and other non-infectious morbidities (89). Likewise, clinical signs observed in the other two infected cats might have been associated with other pathogens since they had no Leishmania in their cytological samples and their condition improved upon further examinations and symptomatic treatment.
Among the substantial majority (92.2%) of PCR negative cats for L. infantum infection, a high proportion (73.5%) exhibited clinical signs of disease that could be attributed to FeL in a presumptive approach (18). We emphasise that the region is endemic for many other infectious diseases, and that these cats are subjected to the same conditions as their owners, namely undernourishment, high demographic densities, and precarious sanitation infrastructure. Therefore, the clinical presentation of unhealthy cats living in endemic areas for L. infantum does not necessarily reflect an active infection by this parasite. Accordingly, a careful assessment linking PCR positivity to clinical signs must be pursued during the clinical evaluation of a presumptive feline case. Indeed, in a different context of coinfections, a recent study performed in an endemic region for CanL in Spain, on a population of clinically healthy stray cats, still found a 5.6% positivity in blood samples using qPCR (90).
The clinical course of the cat with an active infection in this study suggests a natural pathophysiological evolution in a susceptible mammal host (91, 92). In the first examination, the cat had no clinical alteration, but its bone marrow was already heavily infected. No proper clinical follow-up and treatment could be performed because of the owner's non-compliance. Therefore, after the cat develop FeL, the owner looked for assistance again. Then, we could describe the natural infection progression. Some studies relate FeL onset to immunosuppressive factors such as retroviral infections, such as FeLV or FIV (18, 40). There was no evidence of a pre-existing infectious immunosuppressive condition in the cat with FeL. In the fourth month after the first PCR diagnosis, clinical signs consistent with FeL were also apparent in different bodily systems, confirming the disease's systemic character. Lesions in tissues such as ocular conjunctiva, oral mucosa, in addition to enlargement of lymph nodes, as observed in the present cat, have also been previously described by other authors (18, 93, 94). Hyperproteinaemia associated with hyperglobulinemia was the most evident clinical pathological finding when the cat exhibited overt clinical FeL. This finding agrees with other authors that considered hyperglobulinemia as the most relevant alteration found in FeL (95). Importantly, the proportions of gamma globulins in cats with FeL have been described as variable and often not detectable in serological tests, probably due to a predominantly cellular immune response (18, 96, 97).
The cat identified with active FeL herein lived in close contact with a dog previously diagnosed with CanL, and this dog could have been a parasite source for vectors. Indeed, a recent study on sheltered dogs and cats showed that dogs could be more infectious to sandflies due to higher parasite loads than cats with the same PCR-positivity rates. Moreover, the study argued that cats were more frequently seropositive than dogs (22). Despite xenodiagnoses techniques already proving the transmission of the parasite in sand fly feeding on infected cats, no consistent data has demonstrated that L. infantum-infected cats can maintain the parasite circulation in the environment (48, 98, 99). Besides, there is no robust data estimating the transmission risk for humans who cohabit with infected felines (38).
Brazilian municipalities with the highest number of VL reports low human development indexes (7, 100). In this context, as expected, cats of low-income individuals do not receive proper health care. They may be more susceptible to developing active infections with high parasite loads and therefore possibly contributing to the parasite transmission. As we mentioned before, the current study has some limitations especially due to underreporting in some areas. The studied feline population sample that could be assessed did not allow a precise prevalence estimation of the parasite infection. In the locality with the highest notification numbers of VL, a low number of cats could be evaluated, and these animals resided in the same household, so there is a possibility that the negative results found might not reflect reality. Yet, we can assume that TR-DPP® was not a reliable method for assessing humoral responses L. infantum-infected cats. Nevertheless, despite all limitations, the present study confirmed that L. infantum is circulating among cats using parasitological and molecular methods.
Since the evolution of the infection in cats is often subclinical and with a low parasitic burden, more detailed studies with representative samples are needed to ascertain L. infantum prevalence in endemic areas (73, 94, 101). Different from dogs, cats exhibit a more nocturnal behaviour and may walk longer distances and environments. This may increase the chances to encounter a sand fly in nature and acquire and/or transmit the parasite. More importantly, co-infections with other feline endemic pathogens including viruses (FIV, FeLV) (102), bacteria (Mycoplasma haemofelis, Ehrlichia spp) (103, 104) other protozoans (Cytauxzoon spp.) (105), and, helminths (enteric or lung worms) (106) may lead to immunosuppression of the cats. They would be more susceptible to L. infantum infection thus favouring higher parasite burdens. Indeed, in a recent study in Italy and Greece, where L. infantum may also occurs in cats and dogs, Aelurostrongylus abstrusus and Troglostrongylus brevior were found in cats and in a fatal case in a dog (107–109). Those worms have been increasingly reported as important in feline medicine and co-infections would impact animal management. Thus, further investigations should attempt to characterize hosts and co-infections in the epidemiological cycle of zoonotic leishmaniasis. This would promote better control and prevention measures to achieve the principles of One Health.
Conclusions
Molecular identification and sequencing of L. infantum from naturally infected cats indicates that the parasite is circulating in areas where CanL and VL are endemic. These data demonstrate the feline's exposure to the parasite and reinforce the need for further epidemiological investigations to understand how cats participate in the cycle of this zoonotic leishmaniasis. Additionally, the clinical evolution of a subclinical infection in a cat toward the development of patent feline leishmaniasis (FeL) illustrates a picture of active infection. A specific immunodiagnostic assay to assess feline humoral response to L. infantum is needed.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The animal study was reviewed and approved by Committee for Ethics on the Use of Animals of the School of Veterinary Medicine and Zootechny of the Federal University of Bahia (Permission number CEUA-13/2017). Written informed consent was obtained from the owners for the participation of their animals in this study.
Author Contributions
SB-M and FP conceived and designed the study. NS, BV, TN, JR, and FR performed the investigation and drafted the manuscript. NH, NA, and RP provided laboratory material resources and validation of experiments. NS, FP, NH, NA, RP, MS, SB-M, and RS analyzed the data. FP and MS conducted statistical analyses, data curation, and visualization. SB-M, FP, and RS provided funding acquisition, project management, experiments supervision, and manuscript editing. All authors contributed to the writing of the manuscript and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Dr. Tiago Peixoto, Dr. Maisa Fonseca, Dr. Rafaela Gonçalves, and Marina Rossi for technical support. This work was supported by Fundação de Amparo à Pesquisa do Estado de Minas Gerais (Award number(s): PPM-00202-18); Fundação de Amparo à Pesquisa do Estado da Bahia (Award number(s): PRONEM 498/2011-PNE 0002/2011, BOL0435/2018); Conselho Nacional de Desenvolvimento Científico e Tecnológico (Award number(s): 302972/2019-6, 307813/2018-5, 313350/2019-1); Ministério da Educação (Award number(s): PaexDoc-UFBA/2019, 11792, 11436).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.734916/full#supplementary-material
References
1. Alvar J, Vélez ID, Bern C, Herrero M, Desjeux P, Cano J, et al. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE. (2012) 7:e35671. doi: 10.1371/journal.pone.0035671
2. World Health Organization. Leishmaniasis. (2021). Avaliable online at: https://www.who.int/health-topics/leishmaniasis#tab=tab_1 (accessed May 4, 2021).
3. Claborn D. The biology and control of leishmaniasis vectors. J Glob Infect Dis. (2010) 2:127–34. doi: 10.4103/0974-777X.62866
4. Lage DP, Ribeiro PAF, Dias DS, Mendonça DVC, Ramos FF, Carvalho LM, et al. A candidate vaccine for human visceral leishmaniasis based on a specific T cell epitope-containing chimeric protein protects mice against Leishmania infantum infection. NPJ Vaccines. (2020) 5:75. doi: 10.1038/s41541-020-00224-0
5. Mohammed ASA, Tian W, Zhang Y, Peng P, Wang F, Li T. Leishmania lipophosphoglycan components: a potent target for synthetic neoglycoproteins as a vaccine candidate for leishmaniasis. Carbohydr Polym. (2020) 237:116120. doi: 10.1016/j.carbpol.2020.116120
6. Nunes BEBR, Leal TC, de Paiva JPS, da Silva LF, Do Carmo RF, Machado MF, et al. Social determinants of mortality due to visceral leishmaniasis in Brazil (2001-2015): an ecological study. Rev Soc Bras Med Trop. (2020) 53:e20190262. doi: 10.1590/0037-8682-0262-2019
7. Lima ID, Lima ALM, Mendes-Aguiar C de O, Coutinho JFV, Wilson ME, Pearson RD, et al. Changing demographics of visceral leishmaniasis in northeast Brazil: lessons for the future. PLoS Negl Trop Dis. (2018) 12:e0006164. doi: 10.1371/journal.pntd.0006164
8. World Health Organization. Neglected Tropical Diseases(2021). Avaliable online at: https://www.who.int/news-room/q-a-detail/neglected-tropical-diseases (accessed May 4, 2021).
9. Word Organization for Animal Health. OIE-Listed Diseases, Infections and Infestations in Force in 2020. (2021). Avaliable online at: https://www.oie.int/en/what-we-do/animal-health-and-welfare/animal-diseases/old-classification-of-diseases-notifiable-to-the-oie-list-b/ (accessed May 4, 2021).
10. Lukeš J, Mauricio IL, Schönian G, Dujardin JC, Soteriadou K, Dedet JP, et al. Evolutionary and geographical history of the leishmania donovani complex with a revision of current taxonomy. Proc Natl Acad Sci USA. (2007) 104:9375–80. doi: 10.1073/pnas.0703678104
11. Quinnell RJ, Courtenay O. Transmission, reservoir hosts and control of zoonotic visceral leishmaniasis. Parasitology. (2009) 136:1915–34. doi: 10.1017/S0031182009991156
12. Ferroglio E, Battisti E, Zanet S, Bolla C, Concialdi E, Trisciuoglio A, et al. Epidemiological evaluation of Leishmania infantum zoonotic transmission risk in the recently established endemic area of Northwestern Italy. Zoonoses Public Health. (2018) 65:675–82. doi: 10.1111/zph.12477
13. Tabbabi A. Review of leishmaniasis in the middle east and North Africa. Afr Health Sci. (2019) 19:1329–37. doi: 10.4314/ahs.v19i1.4
14. Barrouin-Melo SM, Larangeira DF, de Andrade Filho FA, Trigo J, Julião FS, Franke CR, et al. Can spleen aspirations be safely used for the parasitological diagnosis of canine visceral leishmaniosis? A study on assymptomatic and polysymptomatic animals. Vet J. (2006) 171:331–9. doi: 10.1016/j.tvjl.2004.11.010
15. Solano-Gallego L, Miró G, Koutinas A, Cardoso L, Pennisi M, Ferrer L, et al. LeishVet guidelines for the practical management of canine leishmaniosis. Parasite Vect. (2011) 4:1–16. http://parasitesandvectors.biomedcentral.com/articles/10.1186/1756-3305-4-86. doi: 10.1186/1756-3305-4-86
16. Mohebali M, Moradi-Asl E, Rassi Y. Geographic distribution and spatial analysis of leishmania infantum infection in domestic and wild animal reservoir hosts of zoonotic visceral leishmaniasis in Iran: a systematic review. J Vector Borne Dis. (2018) 55:173–83. doi: 10.4103/0972-9062.249125
17. Trüeb I, Portela RD, Franke CR, Carneiro IO, Ribeiro GJ, Soares RP, et al. Trypanosoma cruzi and Leishmania sp. infection in wildlife from urban rainforest fragments in northeast Brazil. J Wildl Dis. (2018) 54:76–84. doi: 10.7589/2017-01-017
18. Pennisi M, Cardoso L, Baneth G, Bourdeau P, Koutinas A, Miró G. LeishVet update and recommendations on feline leishmaniosis. Parasites Vect. (2015) 8:302. doi: 10.1186/s13071-015-0909-z
19. Soares IR, Silva SO, Moreira FM, Prado LG, Fantini P, Maranhão RPA, et al. First evidence of autochthonous cases of Leishmania (Leishmania) infantum in horse (Equus caballus) in the Americas and mixed infection of Leishmania infantum and Leishmania (Viannia) braziliensis. Vet Parasitol. (2013) 197:665–9. doi: 10.1016/j.vetpar.2013.06.014
20. Vioti G, Leonel JAF, Lemes KM, Pereira VF, Ferreira HL, Keid LB, et al. Molecular detection of Leishmania spp. in cattle from Brazil by means of PCR using internal transcribed spacer 1. Rev Bras Parasitol Vet. (2019) 28:303–5. doi: 10.1590/s1984-29612019003
21. Metzdorf IP, Sebastião M, Junior L, Fatima M De, Matos C, Francisco A, et al. Molecular characterization of Leishmania infantum in domestic cats in a region of Brazil endemic for human and canine visceral leishmaniasis. Acta Trop. (2016) 66:121–5. doi: 10.1016/j.actatropica.2016.11.013
22. Baneth G, Nachum-Biala Y, Zuberi A, Zipori-Barki N, Orshan L, Kleinerman G, et al. Leishmania infection in cats and dogs housed together in an animal shelter reveals a higher parasite load in infected dogs despite a greater seroprevalence among cats. Parasites Vect. (2020) 13:4–11. doi: 10.1186/s13071-020-3989-3
23. Fernandez-Gallego A, Feo Bernabe L, Dalmau A, Esteban-Saltiveri D, Font A, Leiva M, et al. Feline leishmaniosis: diagnosis, treatment and outcome in 16 cats. J Feline Med Surg. (2020) 22:993–1007. doi: 10.1177/1098612X20902865
24. Asfaram S, Fakhar M, Teshnizi SH. Is the cat an important reservoir host for visceral leishmaniasis? A systematic review with meta-analysis. J Venom Anim Toxins. (2019) 25:e20190012. doi: 10.1590/1678-9199-jvatitd-2019-0012
25. Morelli S, Colombo M, Dimzas D, Barlaam A, Traversa D, Di Cesare A, et al. Leishmania infantum seroprevalence in cats from touristic areas of Italy and Greece. Front Vet Sci. (2020) 7:616566. doi: 10.3389/fvets.2020.616566
26. Schäfer I, Kohn B, Volkmann M, Müller E. Retrospective evaluation of vector-borne pathogens in cats living in Germany (2012–2020). Parasit Vect. (2021) 14:2–9. doi: 10.1186/s13071-021-04628-2
27. Lainson R, Rangel BF. Lutzomyia longipalpis and the eco-epidemiology of American visceral leishmaniasis, with particular reference to Brazil - a review. Mem Inst Oswaldo Cruz. (2005) 100:811–27. doi: 10.1590/S0074-02762005000800001
28. Lara-Silva F de O, Michalsky ÉM, Fortes-Dias CL, Fiuza V de OP, Pessanha JEM, Regina-Silva S, et al. Epidemiological aspects of vector, parasite, and domestic reservoir in areas of recent transmission and no reported human cases of visceral leishmaniasis in Brazil. Acta Trop. (2015) 148:128–36. doi: 10.1016/j.actatropica.2015.04.002
29. Salgado-Almario J, Hernández CA, Ovalle-Bracho C. Geographical distribution of Leishmania species in Colombia, 1985-2017. Biomedica. (2019) 39:278–90. doi: 10.7705/biomedica.v39i3.4312
30. Barrios SPG, Pereira LE, Casaril AE, Infran JDOM, De Oliveira Moura Infran J, Oshiro ET, et al. Phlebotominae (Diptera: Psychodidae) and biomes in the state of Mato Grosso do Sul, Brazil. J Med Entomol. (2020) 57:1882–904. doi: 10.1093/jme/tjaa127
31. Lima ÁLM, de Lima ID, Coutinho JFV, de Sousa ÚPST, Rodrigues MAG, Wilson ME, et al. Changing epidemiology of visceral leishmaniasis in northeastern Brazil: a 25-year follow-up of an urban outbreak. Trans R Soc Trop Med Hyg. (2017) 111:440–7. doi: 10.1093/trstmh/trx080
32. Courtenay O, Carson C, Calvo-bado L, Garcez LM, Quinnell RJ. Heterogeneities in Leishmania infantum infection : using skin parasite burdens to identify highly infectious dogs. PLoS Negl Trop Dis. (2014) 8:e2583. doi: 10.1371/journal.pntd.0002583
33. Magalhães-Junior JT, Mota TF, Porfirio-Passos G, Larangeira DF, Franke CR, Barrouin-Melo SM. Xenodiagnosis on dogs with visceral leishmaniasis: canine and sand fly aspects related to the parasite transmission. Vet Parasitol. (2016) 223:120–6. doi: 10.1016/j.vetpar.2016.04.031
34. Lins TNB, Souza IB, Barros GMM do R, Santos CVB, Silva RP, Alves LC, et al. Seroprevalence and spatial distribution of canine leishmaniasis in an endemic region in Brazil: how has the situation changed after 10 years? Rev Soc Bras Med Trop. (2018) 51:680–2. doi: 10.1590/0037-8682-0087-2018
35. Bresciani KDS, Serrano ACM, Matos LVS de, Savani ESMM, D'Auria SRN, Perri SH V, et al. Ocorrência de Leishmania spp. em felinos do município de Araçatuba, SP. Rev Bras Parasitol Veterinária. (2010) 19:127–9. doi: 10.1590/S1984-29612010000200012
36. Cardia DFF, Camossi LG, Neto LS, Langoni H, Bresciani KDS. Prevalence of Toxoplasma gondii and Leishmania spp. infection in cats from Brazil. Vet Parasitol. (2013) 197:634–7. doi: 10.1016/j.vetpar.2013.07.017
37. Rocha AVOV, Moreno BFS, Cabral AD, Louzeiro NM, Miranda LM, Santos VMB, et al. Diagnosis and epidemiology of Leishmania infantum in domestic cats in an endemic area of the amazon region, Brazil. Vet Parasitol. (2019) 273:80–5. doi: 10.1016/j.vetpar.2019.08.007
38. Dalvi APR, Carvalho TDG, Werneck GL. Is there an association between exposure to cats and occurrence of visceral leishmaniasis in humans and dogs? Vector Borne Zoonotic Dis. (2018) 18:335–42. doi: 10.1089/vbz.2017.2162
39. Iatta R, Furlanello T, Colella V, Tarallo VD, Latrofa S, Brianti E, et al. A nationwide survey of Leishmania infantum infection in cats and associated risk factors in Italy. PLoS Negl Trop Dis. (2019) 13:e0007594. doi: 10.1371/journal.pntd.0007594
40. Sobrinho LSV, Rossi CN, Vides JP, Braga ET, Gomes AAD, de Lima VMF, et al. Coinfection of Leishmania chagasi with Toxoplasma gondii, feline immunodeficiency virus (FIV) and feline leukemia virus (FeLV) in cats from an endemic area of zoonotic visceral leishmaniasis. Vet Parasitol. (2012) 187:302–6. doi: 10.1016/j.vetpar.2012.01.010
41. Instituição Brasileiro de Geografia e Estatística. Cidades e Estados. (2021). Avaliable online at: https://www.ibge.gov.br/cidades-e-estados.html?view=municipio (accessed May 4, 2021).
42. Brasil. Secretária de Saúde da Bahia. 2009 a 2018 – Incidência e Casos Absolutos de Leishmaniose Tegumentar e Visceral Por NRS. Bahia. (2021). Avaliable online at: http://www.saude.ba.gov.br/suvisa/vigilancia-epidemiologica/doencas-de-transmissao-vetorial/leishmaniose/ (accessed May 20, 2021).
43. Junqueira ANN, Galera PD. Characteristics of the population of dogs and cats in Brazil. Acta Vet Bras. (2019) 13:77–86. doi: 10.21708/avb.2019.13.2.8028
44. Varjão BM, Pinho FA, Solcà MS, Silvestre R, Fujimori M, Goto H, et al. Spatial distribution of canine leishmaniasis by Leishmania infantum in a municipality with endemic human leishmaniasis in eastern Bahia, Brazil. Rev Bras Parasitol Vet. (2021) 30:e022620. doi: 10.1590/s1984-29612021034
45. Julião FS, Souza BMPS, Freitas DS, Oliveira LS, Laranjeira DF, et al. Investigation of risk areas as complemental methodology for the control of canine visceral leishmaniasis. Pesqui Vet Bras. (2007) 27:319–324. doi: 10.1590/S0100-736X2007000800001
46. Deiró AGJ, Montargil SMA, Carvalho FS, Munhoz AD, Albuquerque GR. Antibody occurrence of Anti- Toxoplasma gondii, Leishmania sp. and Ehrlichia canis in dogs in Bahia State Ocorrência de anticorpos Anti- Toxoplasma gondii, Leishmania spp. e Ehrlichia canis em cães do Estado da Bahia. Univer Estadual Londrina. (2018) 39:199–210. doi: 10.5433/1679-0359.2018v39n1p199
47. Brasil Ministério da Saúde Secretaria de Vigilância em Saúde Departamento de Vigilância Epidemiológica. Manual de Vigilância e Controle da Leishmaniose Visceral, Ministério da Saúde, Secretaria de Vigilância em Saúde, Departamento de Vigilância Epidemiológica. (2014). Avaliable online at: http://www.saude.ba.gov.br/wp-content/uploads/2019/05/2014-Manual-de-vigil%C3%A2ncia-e-controle-da-leishmaniose-visceral.pdf (accessed May 4, 2021).
48. Pennisi MG, Hartmann K, Lloret A, Addie D, Belák S, Boucraut-baralon C, et al. Leishmaniasis in cats: ABCD guidelines on prevention and management. J Feline Med Surg. (2013) 15:638–42. doi: 10.1177/1098612X13489229
49. Woods GA, Simpson M, Boag A, Paris J, Piccinelli C, Breheny C. Complications associated with bone marrow sampling in dogs and cats. J Small Anim Pract. (2021) 62:209–15. doi: 10.1111/jsap.13274
50. Fraga DB, Pacheco LV, Borja LS, Tuy PG, Bastos LA, Solcà Mda S, et al. The rapid test based on Leishmania infantum chimeric rK28 protein improves the diagnosis of canine visceral leishmaniasis by reducing the detection of false-positive dogs. PLoS Negl Trop Dis. (2016) 5:e0004333. doi: 10.1371/journal.pntd.0004333
51. Pattabhi S, Whittle J, Mohamath R, El-Safi S, Moulton GG, Guderian JA, et al. Design, development and evaluation of rK28-based point-of-care tests for improving rapid diagnosis of visceral leishmaniasis. PLoS Negl Trop Dis. (2010) 4:e822. doi: 10.1371/journal.pntd.0000822
52. Lindmark R, Thorén-Tolling K, Sjöquist J. Binding of immunoglobulins to protein A and immunoglobulin levels in mammalian sera. J Immunol Methods. (1983) 62:1–13. doi: 10.1016/0022-1759(83)90104-7
53. Silva RBS, Leite DPSBM, Filho EM, Silva LFFE, De Almeida Braz BM, De Melo MA.(2018). Anticorpos anti-Leishmania sp. em felis catus domesticus atendidos no hospital veterinário da UFCG, patos-PB. In: Proceedings of the 54° Congresso da Sociedade Brasileira de Medicina Tropical; Sept 2-5. Pernambuco. Avaliable online at: http://www.adaltech.com.br/anais/medtrop2018/arquivos/todos%20os%20trabalhos.pdf.4 (acessed May 4, 2021).
54. Birkenheuer AJ, Levy MG, Breitschwerdt EB. Development and evaluation of a seminested pcr for detection and differentiation of Babesia gibsoni (asian genotype) and B. canis DNA in canine. Blood Sampl. (2003) 41:4172–77. doi: 10.1128/JCM.41.9.4172-4177.2003
55. Lachaud L, Marchergui-hammami S, Chabbert E, Dedet JP, Bastien P. Comparison of six pcr methods using peripheral blood for detection of canine visceral leishmaniasis. J Clin Microbiol. (2002) 40:210–5. doi: 10.1128/JCM.40.1.210-215.2002
56. Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. (2018) 35:1547–9. doi: 10.1093/molbev/msy096
57. Schönian G, Nasereddin A, Dinse N, Schweynoch C, Schallig HDFH, Presber W, et al. PCR diagnosis and characterization of Leishmania in local and imported clinical samples. Diagn Microbiol Infect Dis. (2003) 47:349–58. doi: 10.1016/S0732-8893(03)00093-2
58. Garcia L, Kindt A, Bermudez H, Llanos-Cuentas A, De Doncker S, Arevalo J, et al. Culture-independent species typing of neotropical Leishmania for clinical validation of a pcr-based assay targeting heat shock protein 70 genes. J Clin Microbiol. (2004) 42:2294–7. doi: 10.1128/JCM.42.5.2294-2297.2004
59. Rocha RF, Menezes E V, Xavier AREO, Royo VA, Oliveira DA, Júnior AFM, et al. Standardization of a molecular method for epidemiologic identification of Leishmania strains. Genet Mol Res. (2016) 4:1–12. doi: 10.4238/gmr.15048854
60. Rampazzo R de CP, Solcà M da S, Santos LCS, Pereira L de N, Guedes JCO, Veras PST, et al. A ready-to-use duplex qPCR to detect Leishmania infantum DNA in naturally infected dogs. Vet Parasitol. (2017) 246:100–7. doi: 10.1016/j.vetpar.2017.09.009
61. Soares RPP, Macedo ME, Ropert C, Gontijo NF, Almeida IC, Gazzinelli RT, et al. Leishmania chagasi: lipophosphoglycan characterization and binding to the midgut of the sand fly vector Lutzomyia longipalpis. Mol Biochem Parasitol. (2002) 121:213–24. doi: 10.1016/S0166-6851(02)00033-6
62. Santos RCD, Pinho FAD, Passos GP, Larangeira DF, Barrouin-Melo SM. Isolation of naturally infecting Leishmania infantum from canine samples in Novy-MacNeal-Nicolle medium prepared with defibrinated blood from different animal species. Vet Parasitol. (2018) 15:10–4. doi: 10.1016/j.vetpar.2018.05.010
63. Jerry Kaneko J, Harvey JJ, Bruss ML. Clinical Biochemistry of Domestic Animals. Burlington, MA: Elsevier; Academic Press (2008). 936p.
65. Leça Júnior NF, Guedes PEB, Santana LN, Almeida V dos A, Carvalho FS, Albuquerque GR, et al. Epidemiology of canine leishmaniasis in southern Bahia, Brazil. Acta Trop. (2015) 148:115–9. doi: 10.1016/j.actatropica.2015.04.008
66. Solcà M da S, Arruda MR, Leite BMM, Mota TF, Rebouças MF, de Jesus MS, et al. Immune response dynamics and lutzomyia longipalpis exposure characterize a biosignature of visceral leishmaniasis susceptibility in a canine cohort. PLoS Negl Trop Dis. (2021) 15:e0009137. doi: 10.1371/journal.pntd.0009137
67. Mendonça IL, Batista JF, Ribeiro IMM, Rocha FSB, Silva SO, Melo MN. Leishmania infantum in domestic cats from the municipality of Teresina, state of Piauí, Brazil. (2017) 3: 1–8. doi: 10.1017/pao.2017.1
68. Leonel JAF, Vioti G, Alves ML, Benassi JC, Silva DT, Spada JCP, et al. Leishmaniasis in cat shelters: A serological, molecular and entomological study. Cambridge Core. (2020) 67:2013–9. doi: 10.1111/tbed.13544
69. de Oliveira SS, de Araújo TM. Evaluation of control measures for visceral leishmaniasis (kala azar) in an endemic area in Bahia, Brazil (1995-2000). Cad saúde pública. (2003) 19:1681–90. doi: 10.1590/S0102-311X2003000600012
70. Gonçalves MB. Prevalência, Distribuição e Identificação de Prováveis Fatores de Risco Para Leishmaniose Visceral Canina em Camaçari-BA. (Dissertation), Fundação Oswaldo Cruz, Centro de Pesquisas Gonçalo Moniz (2014). Available online at: https://www.arca.fiocruz.br/bitstream/icict/9476/2/Marcelo%20Bordoni%20Gon%C3%A7alves.%20Prevalencia.2014.pdf (acessed May 4, 2021).
71. Solcà MDS, Bastos LA, Guedes CES, Bordoni M, Borja LS, Larangeira DF, et al. Evaluating the accuracy of molecular diagnostic testing for canine visceral leishmaniasis using latent class analysis. PLoS ONE. (2014) 9:e0103635. doi: 10.1371/journal.pone.0103635
72. Francino O, Altet L, Sánchez-Robert E, Rodriguez A, Solano-Gallego L, Alberola J, et al. Advantages of real-time PCR assay for diagnosis and monitoring of canine leishmaniosis. Vet Parasitol. (2006) 137:214–21. doi: 10.1016/j.vetpar.2006.01.011
73. Chatzis MK, Andreadou M, Leontides L, Kasabalis D, Mylonakis M, Koutinas AF, et al. Cytological and molecular detection of Leishmania infantum in different tissues of clinically normal and sick cats. Vet Parasitol. (2014) 202:217–25. doi: 10.1016/j.vetpar.2014.02.044
74. Soares CSA, Duarte SC, Sousa SR. What do we know about feline leishmaniosis? J Feline Med Surg. (2015) 18:435–42. doi: 10.1177/1098612X15589358
75. Barrouin-Melo SM, Larangeira DF, Trigo J, Aguiar PHP, Dos-Santos WLC, Pontes-De-Carvalho L. Comparison between splenic and lymph node aspirations as sampling methods for the parasitological detection of Leishmania chagasi infection in dogs. Mem Inst Oswaldo Cruz. (2004) 99:195–7. doi: 10.1590/S0074-02762004000200014
76. Travi BL, Cordeiro-da-Silva A, Dantas-Torres F, Miró G. Canine visceral leishmaniasis: Diagnosis and management of the reservoir living among us. PLoS Neglected Tropical Diseases. (2018) 12:e0006082. doi: 10.1371/journal.pntd.0006082
77. Giunchetti RC, Silveira P, Resende LA, Leite JC, Melo-Júnior OA de O, Rodrigues-Alves ML, et al. Canine visceral leishmaniasis biomarkers and their employment in vaccines. Vet Parasitol. (2019) 271:87–97. doi: 10.1016/j.vetpar.2019.05.006
78. Reis AB, Martins-Filho OA, Teixeira-Carvalho A, Carvalho MG, Mayrink W, França-Silva JC, et al. Parasite density and impaired biochemical/hematological status are associated with severe clinical aspects of canine visceral leishmaniasis. Res Vet Sci. (2006) 81:68–75. doi: 10.1016/j.rvsc.2005.09.011
79. Gonçalves RDS, de Pinho FA, Dinis-Oliveira RJ, Azevedo R, Gaifem J, Larangeira DF, et al. Mathematical modelling using predictive biomarkers for the outcome of canine leishmaniasis upon chemotherapy. Microorganisms. (2020) 8:745. doi: 10.3390/microorganisms8050745
80. Martínez V, Quilez J, Sanchez A, Roura X, Francino O, Altet L. Canine leishmaniasis: the key points for qPCR result interpretation. Parasites Vect. (2011) 4:1–5. doi: 10.1186/1756-3305-4-57
81. Noronha TR De, Fock RA. Hematology, transfusion and cell therapy images in clinical hematology visceral leishmaniasis: amastigotes in the bone marrow. Hematol Transfus Cell Ther. (2019) 41:99–100. doi: 10.1016/j.htct.2018.06.003
82. Marcondes M, Hirata KY, Vides JP, Sobrinho LS V, Azevedo JS, Vieira TSWJ, et al. Infection by Mycoplasma spp., feline immunodeficiency virus and feline leukemia virus in cats from an area endemic for visceral leishmaniasis. Parasites Vect. (2018) 11:131. doi: 10.1186/s13071-018-2716-9
83. Luciano RM, Zampoli M. Avaliação da reatividade cruzada entre antígenos de Leishmania spp e Trypanosoma cruzi na resposta sorológica de cães pela técnica de imunofluorescência indireta (RIFI). Braz J Vet Res An Sci. (2009) 181–7. doi: 10.11606/issn.1678-4456.bjvras.2009.26765
84. Alves AS, Mouta-Confort E, Figueiredo FB, Oliveira RVC, Schubach AO, Madeira MF. Evaluation of serological cross-reactivity between canine visceral leishmaniasis and natural infection by Trypanosoma caninum. Res Vet Sci. (2012) 93:1329–33. doi: 10.1016/j.rvsc.2012.07.006
85. Reis AB, Teixeira-Carvalho A, Vale AM, Marques MJ, Giunchetti RC, Mayrink W, et al. Isotype patterns of immunoglobulins: Hallmarks for clinical status and tissue parasite density in brazilian dogs naturally infected by Leishmania (Leishmania) chagasi. Vet Immunol Immunopathol. (2006) 112:102–16. doi: 10.1016/j.vetimm.2006.02.001
86. Aslan H, Oliveira F, Meneses C, Castrovinci P, Gomes R, Teixeira C, et al. New insights into the transmissibility of leishmania infantum from dogs to sand flies: experimental vector-transmission reveals persistent parasite depots at bite sites. J Infect Dis. (2016) 213:1752–61. doi: 10.1093/infdis/jiw022
87. de Santana Martins Rodgers M, Bavia ME, Fonseca EOL, Cova BO, Silva MMN, Carneiro DDMT, et al. Ecological niche models for sand fly species and predicted distribution of Lutzomyia longipalpis (Diptera: Psychodidae) and visceral leishmaniasis in Bahia state, Brazil. Environ Monit Assess. (2019) 191:331. doi: 10.1007/s10661-019-7431-2
88. Sundar S, Singh OP. Molecular diagnosis of visceral leishmaniasis. Mol Diagnosis Ther. (2018) 22:443–57. doi: 10.1007/s40291-018-0343-y
89. Pesavento PA, Murphy BG. Common and emerging infectious diseases in the animal shelter. Vet Pathol. (2014) 51:478–91. doi: 10.1177/0300985813511129
90. Alcover MM, Basurco A, Fernandez A et al. A cross-sectional study of Leishmania infantum infection in stray cats in the city of Zaragoza (Spain) using serology and PCR. Parasites Vect. (2021) 14:178. doi: 10.1186/s13071-021-04682-w
91. Menna-Barreto RFS. Cell death pathways in pathogenic trypanosomatids: lessons of (over)kill. Cell Death Disease. (2019) 10:93. doi: 10.1038/s41419-019-1370-2
92. Brianti E, Celi N, Napoli E, Abbate JM, Arfuso F, Gaglio G, et al. Treatment and long-term follow-up of a cat with leishmaniosis. Parasites Vect. (2019) 12:121. doi: 10.1186/s13071-019-3388-9
93. Gramiccia M. Recent advances in leishmaniosis in pet animals : epidemiology, diagnostics and anti-vectorial prophylaxis. Vet Parasitol. (2011) 181:23–30. doi: 10.1016/j.vetpar.2011.04.019
94. Basso MA, Marques C, Santos M, Duarte A, Pissarra H, Carreira LM, et al. Successful treatment of feline leishmaniosis using a combination of allopurinol and N-methyl-glucamine antimoniate. J Feline Med Surg Open Rep. (2016) 1:1–7. doi: 10.1177/2055116916630002
95. Chatzis MK, Xenoulis PG, Leontides L, Kasabalis D, Mylonakis ME, Andreadou M, et al. Evaluation of clinicopathological abnormalities in sick cats naturally infected by Leishmania infantum. Heliyon. (2020) 6:e05177. doi: 10.1016/j.heliyon.2020.e05177
96. Persichetti MF, Solano-gallego L, Vullo A, Masucci M, Marty P, Delaunay P, et al. Diagnostic performance of ELISA, IFAT and western blot for the detection of anti- Leishmania infantum antibodies in cats using a Bayesian analysis without a gold standard. Parasites Vect. (2017) 10:119. doi: 10.1186/s13071-017-2046-3
97. Urbani L, Tirolo A, Salvatore D, Tumbarello M, Segatore S, Battilani M, et al. Serological, molecular and clinicopathological findings associated with Leishmania infantum infection in cats in Northern Italy. J Feline Med Surg. (2020) 22:935–43. doi: 10.1177/1098612X19895067
98. Maroli M, Grazia M, Di T, Khoury C, Gradoni L, Gramiccia M. Infection of sandflies by a cat naturally infected with Leishmania infantum. Vet Parasitol. (2007) 145:357–60. doi: 10.1016/j.vetpar.2006.11.009
99. Da Silva SM, Rabelo PFB, Gontijo NF, Ribeiro RR, Melo MN, Ribeiro VM, et al. First report of infection of Lutzomyia longipalpis by Leishmania (Leishmania) infantum from a naturally infected cat of Brazil. Vet Parasitol. (2010) 174:150–4. doi: 10.1016/j.vetpar.2010.08.005
100. Cavalcante ÍJM, Vale MR. Aspectos epidemiológicos da leishmaniose visceral (calazar) no Ceará no período de 2007 a 2011. Rev Bras Epidemiol. (2014) 17:911–24. doi: 10.1590/1809-4503201400040010
101. Pennisi MG, Persichetti MF. Feline leishmaniosis : is the cat a small dog? Vet Parasitol. (2018) 251:131–7. doi: 10.1016/j.vetpar.2018.01.012
102. Poffo D, Almeida ABPF, Nakazato L, Dutra V, Correa SHR, Mendonça AJ, et al. Feline immunodeficiency virus (FIV), feline leukaemia virus (FeLV) and Leishmania sp. in domestic cats in the Midwest of Brazil. Pesqui Vet Bras. (2017) 37:491–4. doi: 10.1590/s0100-736x2017000500011
103. Munhoz AD, Simões IGPC, Calazans APF, Macedo LS, Cruz RDS, Lacerda LC, et al. Hemotropic mycoplasmas in naturally infected cats in Northeastern Brazil. Rev Bras Parasitol Vet. (2021) 27:446–54. doi: 10.1590/s1984-296120180074
104. Guimarães A, Raimundo JM, Rodrigues RB, Peixoto MP, Santos HA, André MR, et al. Ehrlichia spp. infection in domestic cats from Rio de Janeiro State, southeast Brazil. Rev Bras Parasitol Vet. (2019) 28:180–5. doi: 10.1590/s1984-296120180088
105. Maia LMP, Cerqueira A de MF, de Barros Macieira D, de Souza AM, Moreira NS, da Silva AV, et al. Cytauxzoon felis and “Candidatus Mycoplasma haemominutum” coinfection in a Brazilian domestic cat (Felis catus). Braz J Vet Parasitol. (2013) 22:289–91. doi: 10.1590/S1984-29612013000200049
106. Ramos N de V, Silva ML e, Barreto MS, Barros LA, Mendes-de-Almeida F. Endoparasites of household and shelter cats in the city of Rio de Janeiro, Brazil. Rev Bras Parasitol Vet. (2020) 29:1–15. doi: 10.1590/s1984-29612019110
107. Di Cesare A, Morelli S, Colombo M, Simonato G, Veronesi F, Marcer F, et al. Is angiostrongylosis a realistic threat for domestic cats? Front Vet Sci. (2020) 7:195. doi: 10.3389/fvets.2020.00195
108. Morelli S, Crisi PE, Di Cesare A, De Santis F, Barlaam A, Santoprete G, et al. Exposure of client-owned cats to zoonotic vector-borne pathogens: clinic-pathological alterations and infection risk analysis. Comp Immunol Microbiol Infect Dis. (2019) 66:101344. doi: 10.1016/j.cimid.2019.101344
Keywords: bone marrow cytology, cats, leishmaniasis, one health, PCR, zoonosis
Citation: Santos NSd, Pinho FAd, Hlavac NRC, Nunes TL, Almeida NR, Solcà MS, Varjão BM, Portela RW, Rugani JN, Rêgo FD, Barrouin-Melo SM and Soares RP (2021) Feline Leishmaniasis Caused by Leishmania infantum: Parasite Sequencing, Seropositivity, and Clinical Characterization in an Endemic Area From Brazil. Front. Vet. Sci. 8:734916. doi: 10.3389/fvets.2021.734916
Received: 02 July 2021; Accepted: 02 August 2021;
Published: 25 August 2021.
Edited by:
Donato Traversa, University of Teramo, ItalyReviewed by:
Barbara Paoletti, University of Teramo, ItalyAmanda Cavalcanti, National Institute of Traumatology and Orthopaedics, Brazil
Copyright © 2021 Santos, Pinho, Hlavac, Nunes, Almeida, Solcà, Varjão, Portela, Rugani, Rêgo, Barrouin-Melo and Soares. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rodrigo Pedro Soares, cm9kcmlnb3NvYXJlczI4QGhvdG1haWwuY29t; Stella Maria Barrouin-Melo, YmFycm91aW5AdWZiYS5icg==
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share last authorship