
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci., 28 January 2021
Sec. Zoological Medicine
Volume 8 - 2021 | https://doi.org/10.3389/fvets.2021.588697
 Netanya Bernitz1
Netanya Bernitz1 Tanya J. Kerr1Wynand J. Goosen1Josephine Chileshe1Roxanne L. Higgitt1
Tanya J. Kerr1Wynand J. Goosen1Josephine Chileshe1Roxanne L. Higgitt1 Eduard O. Roos1†
Eduard O. Roos1† Christina Meiring1Rachiel Gumbo1Candice de Waal1Charlene Clarke1Katrin Smith1Samantha Goldswain1
Christina Meiring1Rachiel Gumbo1Candice de Waal1Charlene Clarke1Katrin Smith1Samantha Goldswain1 Taschnica T. Sylvester1
Taschnica T. Sylvester1 Léanie Kleynhans1
Léanie Kleynhans1 Anzaan Dippenaar1†
Anzaan Dippenaar1† Peter E. Buss2David V. Cooper3
Peter E. Buss2David V. Cooper3 Konstantin P. Lyashchenko4
Konstantin P. Lyashchenko4 Robin M. Warren1
Robin M. Warren1 Paul D. van Helden1
Paul D. van Helden1 Sven D. C. Parsons1
Sven D. C. Parsons1 Michele A. Miller1*
Michele A. Miller1*Wildlife tuberculosis is a major economic and conservation concern globally. Bovine tuberculosis (bTB), caused by Mycobacterium bovis (M. bovis), is the most common form of wildlife tuberculosis. In South Africa, to date, M. bovis infection has been detected in 24 mammalian wildlife species. The identification of M. bovis infection in wildlife species is essential to limit the spread and to control the disease in these populations, sympatric wildlife species and neighboring livestock. The detection of M. bovis-infected individuals is challenging as only severely diseased animals show clinical disease manifestations and diagnostic tools to identify infection are limited. The emergence of novel reagents and technologies to identify M. bovis infection in wildlife species are instrumental in improving the diagnosis and control of bTB. This review provides an update on the diagnostic tools to detect M. bovis infection in South African wildlife but may be a useful guide for other wildlife species.
Mycobacterium bovis is a member of the Mycobacterium tuberculosis complex (MTBC), a group of genetically related mycobacterium species that cause tuberculosis in a range of mammals (1). Of all MTBC members, M. bovis has the widest host range and causes bovine tuberculosis (bTB) in domestic animals, livestock, wildlife, and humans (2). Globally, the eradication of M. bovis is hampered by the existence of wildlife reservoirs that serve as recurrent sources of infection, posing a threat for spillover to livestock at the livestock-wildlife interface, and other sympatric wildlife species (3).
In South Africa, M. bovis is the most common cause of wildlife tuberculosis, with two of the largest wildlife reserves, the Kruger National Park (KNP) and Hluhluwe-iMfolozi Park (HiP), declared endemic for M. bovis (2, 4). The KNP and HiP are adjacent to communal lands where livestock graze freely (5, 6), risking the spillover of bTB from wildlife to domestic livestock. This may have regulatory consequences and subsequent imposed trade restrictions (6, 7). In addition, the detection of bTB in wildlife can lead to quarantine of wildlife premises and threaten conservation and tourism, which can have environmental and socio-economic implications (8).
The development and optimisation of diagnostic tools to identify M. bovis infection is a crucial step to identify affected individuals to manage transmission. However, the validation of diagnostic tests for wildlife bTB is limited by access to large numbers of high-quality samples from confirmed infected and uninfected species (9–11). Due to logistical challenges, it can be difficult to confirm M. bovis infection, especially from suspected cases using antemortem samples.
With the advent of new techniques and tools to detect M. bovis infection in wildlife, our understanding of bTB continues to evolve. To date, M. bovis infection has been detected in 24 mammalian wildlife species in South Africa (Table 1) with most cases in these species identified within the last decade (Figure 1). The development of diagnostic tools to identify infected hosts is essential to limit the spread of bTB and to control the disease. The aim of this review is to provide an update on recent diagnostic developments for wildlife bTB in South Africa.
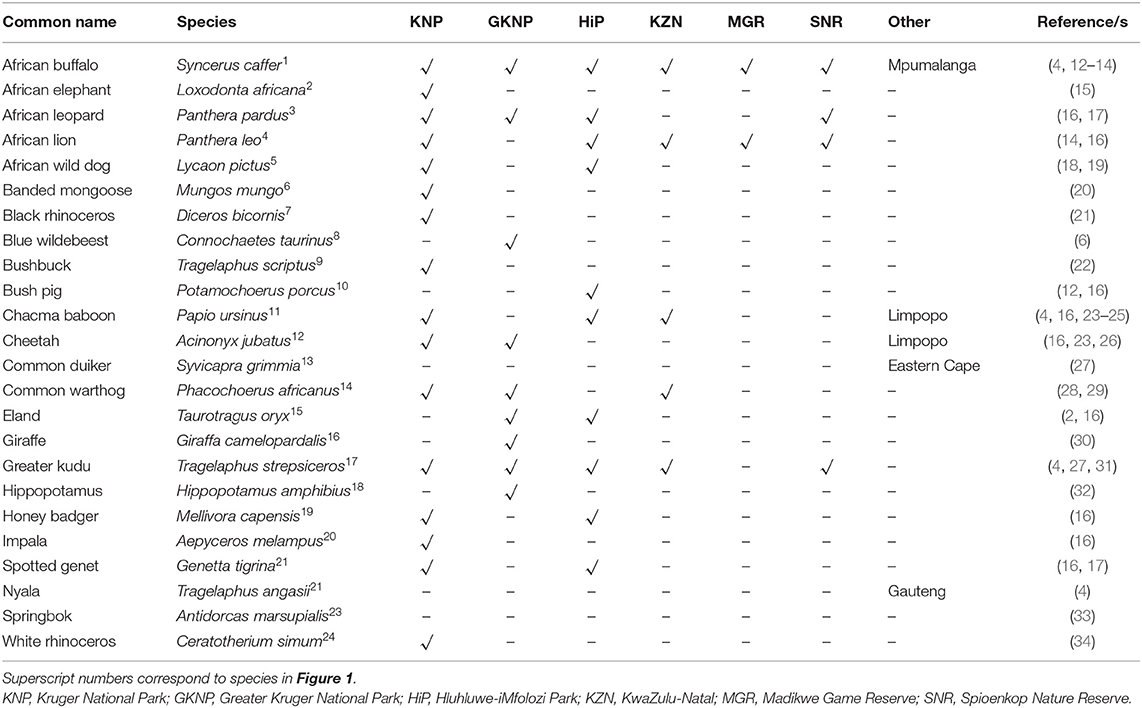
Table 1. Free-ranging wildlife species confirmed to be infected by Mycobacterium bovis in South Africa, based on location, along with references.

Figure 1. Index cases of Mycobacterium bovis infection in free-ranging wildlife species in South Africa over the last century (35). Species correspond to the superscript numbers in Table 1.
Direct staining of sample material with Ziehl-Neelsen stain can provide presumptive diagnosis of mycobacterial infection. Impression smears from lesions in tissues or secretions can provide a rapid screening technique in cases in which bTB is suspect (21). Staining of tissues is also useful for postmortem and antemortem diagnosis in limited cases in which biopsy or other relevant samples such as lymph node may be available (36). Although the presence of acid-fast bacteria provides a presumptive diagnosis, paucibacillary infection may result in a false-negative result using this method (11, 37). In addition, non-tuberculous mycobacteria and non-mycobacterial organisms, such as Nocardia, can stain positive, with a false positive result (38). Therefore, methods that provide differentiation from NTM and speciation within the Mycobacterium tuberculosis complex are important for a definitive diagnosis (39, 40).
Mycobacterial culture is the gold standard of M. bovis infection with speciation confirmed by polymerase chain reaction (PCR). In recent years, there have been advances in techniques to improve direct detection of M. bovis infection in wildlife; however, most procedures still require growing the organism to measurable levels using different mycobacterial culture methods. Due to the inherent slow growth of mycobacteria, the development of improved culture techniques using different media has been investigated.
Mycobacterium bovis has been isolated from both livestock and wildlife samples using solid and liquid culture media. In a study by the National Veterinary Services Laboratory (USA), the liquid BACTEC 12B media supported significantly higher detection of M. bovis compared to the liquid MGIT 960 (93.1 vs. 81.9%, respectively) (both Becton Dickinson, Franklin Lakes, NJ, USA), and both outperformed solid Lowenstein-Jensen and Middlebrook media (41). However, the authors concluded that MGIT 960 media was favored due to increased specific recovery of MTBC and decreased time to positivity.
The BACTEC™ MGIT™ is an automated mycobacterial growth detection system used in routine human tuberculosis diagnostic settings, that has been used to culture MTBC organisms from postmortem tissue samples as well as antemortem bronchoalveolar lavage, elephant trunk wash fluid, and oropharyngeal swab samples from wildlife (42–44). Application of these techniques has permitted antemortem diagnosis of M. bovis infection in African lion (Panthera leo), African wild dog (Lycaon pictus) and white rhinoceros (Ceratotherium simum) (36, 42, 45).
More recently, TiKa-MGIT (TiKa Diagnostics, UK), a novel specialized culture medium (containing a mild decontamination step and growth enhancer steps), with the unique ability to stimulate MTBC growth when used together with the BACTEC™ MGIT™ system, has been used to improve sensitivity of mycobacterial culture, even from samples with low mycobacterial numbers (46). Greater use of this technique may enhance detection of MTBC infection in wildlife in which paucibacillary samples or those heavily contaminated with other environmental bacteria have resulted in false-negative culture results using conventional methods.
Various PCR-based methods have been developed and adapted to identify specific MTBC organisms, based on detecting the presence of mycobacterial DNA, from either cultured isolates or directly from ante or postmortem samples (43, 47). Two emerging PCR-based molecular tools for wildlife samples include VetMAX™ M. tuberculosis complex PCR kit (Thermo Fischer Scientific, Waltham, MA, USA) and cartridge-based GeneXpert® (Cepheid, Sunnyvale, CA, USA) technology.
The VetMAX™ MTBC PCR kit was developed for rapid detection of M. bovis in cattle lymph nodes and other tissues. The real-time PCR targets the IS6110 insertion element found in MTBC, and therefore, should be applicable to samples from any animal species. Use with samples from South African wildlife has been described with successful detection of MTBC DNA in lymph node tissue homogenates and respiratory secretions from buffaloes, white rhinoceros, and African elephants (47). Although useful for diagnosis of MTBC infection in species with limited host-based diagnostic tests, the performance of the assay requires DNA extraction of samples and a facility that can perform PCR, which restricts its use to diagnostic or research laboratories.
The GeneXpert® is a cartridge-based automated screening system endorsed by the World Health Organization (48). Two assays, Xpert® MTB/Rif and Xpert® MTB/Rif Ultra have been developed for the detection of Mycobacterium tuberculosis infection and resistance to rifampicin in humans via PCR (43). This technology allows the detection of MTBC deoxyribonucleic acid (DNA) using a simplified and standardized method and is effective in high throughput diagnostic or research settings. In addition, it is often available in regions where the human tuberculosis burden is high and in regions where laboratories for mycobacterial culture may not exist.
This rapid, simple-to-use, automated platform is currently being adapted and optimized to detect M. bovis from wildlife samples (49) including postmortem tissue homogenates and antemortem bronchoalveolar lavage samples, trunk wash fluids, and oral swabs from elephants, rhinoceroses, and African buffaloes (47). The Xpert® MTB/RIF Ultra assay has also been used to confirm the presence of MTBC DNA directly from tissue homogenates from cheetah (Acinonyx jubatus) (26).
The implementation of GeneXpert® has already played a major role in implementing centralized human tuberculosis diagnosis, increased the number of detected cases, and shortened the diagnostic delay, despite the limitation that it only detects MTBC DNA rather than viable bacilli (48). Therefore, this benchtop system, which can be used with a range of sample types, may provide an alternative or ancillary diagnostic technique for tuberculosis detection in South African wildlife as well as other populations globally, especially when mycobacterial culture is unavailable.
The most common molecular typing tools used to genetically differentiate M. bovis are: (i) region of difference (RD); (ii) spacer oligonucleotide typing (spoligotyping); (iii) variable number of tandem repeats (VNTR) typing of mycobacterial interspersed repetitive units (MIRU); and (iv) next generation sequencing (50–53).
Region of difference analysis is used to speciate MTBC members and allows rapid identification of M. bovis in culture isolates (54). In wildlife, this technique has resulted in differentiation of animal adapted MTBC strains, such as M. mungi in mongooses, M. suricate in meerkats, and M. orygis in a buffalo (55–57). This speciation is important for understanding the epidemiology of infection in different animal hosts.
Spoligotyping is commonly used to detect and identify the genotype of MTBC isolates to determine the phylogenetic relationships between organisms from specific geographical regions and to track sources of infection (Table 2). Previously, data suggested that a single dominant strain of M. bovis circulated within wildlife in specific geographical locations in South Africa. Two geographically distinct spoligotype patterns existed in South African wildlife and were believed to have been caused by different progenitor strains (12, 16). A ‘Kruger’ spoligotype (SB0121) and a ‘KwaZulu Natal’ spoligotype (SB0140) were isolated in wildlife in Greater Kruger National Park (GKNP) and KwaZulu Natal, respectively (12). In a recent study, Sichewo et al. [2020] analyzed samples from cattle and wildlife in KwaZulu Natal and found that although SB0130 was the dominant pattern, these M. bovis isolates were comprised of 29 different strains based on VNTR (58). These results suggest a high level of genetic diversity in this area. These results also support the hypothesis that there are several different M. bovis strains circulating in South Africa. Since spoligotypes SB0120 and SB0121 are the predominant patterns circulating in animals worldwide, as well as finding SB0140 in Asia, Europe and America, it is highly likely that there were multiple introductions with movement of livestock between countries, and inter-species transmission at livestock-wildlife interfaces in South Africa (59). The genetic diversity of M. bovis in South Africa may also be indicative of genetic divergence, ongoing transmission, as well as movement of infected animals (4), but further epidemiological studies are required.
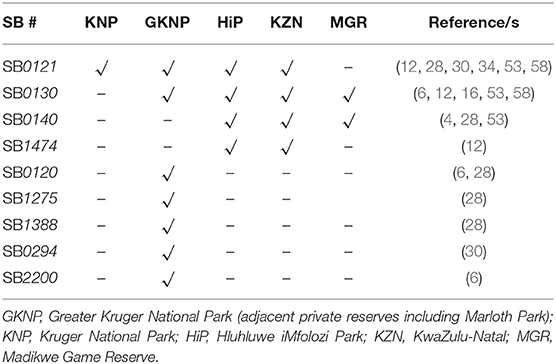
Table 2. Spoligotype (SB) numbers of Mycobacterium bovis isolates identified in free-ranging wildlife in South Africa, based on location of the infected species, with references.
Variable number tandem repeat (VNTR) typing of M. bovis is also used to evaluate transmission by assessing the genetic homology of different isolates. For example, the same VNTR genotype (and spoligotype) of M. bovis was found in cattle and wildlife species in the Greater Kruger National Park Complex, providing evidence that spillover had occurred (7). Studies have shown that VNTR typing is more discriminatory than spoligotyping and often used in combination. These techniques were used to characterize M. bovis isolates from wildlife in Hluhluwe-iMfolozi Park, which is endemic for M. bovis (12). Using VNTR and spoligotyping, the patterns suggested that multiple introductions had occurred and provided a basis for evaluating the epidemiology of bTB in the park. A larger study used both techniques to describe the high genetic diversity of M. bovis strains circulating in livestock and wildlife populations around South Africa and provided data that showed inter-species transmission especially at wildlife/livestock interfaces (6, 58). Molecular characterization of M. bovis isolates is crucial for epidemiological investigation of outbreaks, especially involving multiple species.
Next generation sequencing (NGS) of MTBC has increased resolution and discriminatory power compared to other molecular-based genotyping methods. The generation of whole genome sequences (WGS) allows distinct genetic profiles to be compared at the nucleotide level, and MTBC molecular epidemiology and genetic diversity can be investigated with greater resolution (53). In common warthogs (Phacochoerus africanus), NGS permitted the differentiation of MTBC isolates revealing two distinct clades of M. bovis that were not differentiated using spoligotyping (28); therefore, NGS could improve molecular epidemiological investigations of MTBC infections by resolving transmission events or shedding light on genetic divergence.
A study that analyzed WGS data of 17 M. bovis isolates from different host species in different locations within South Africa revealed distinct regional genomic characteristics (53). Different host species in the KNP clustered together suggesting the introduction of a single progenitor M. bovis strain which led to clonal expansion (53).
High quality WGS is currently dependent on mycobacterial culture due to the requirement for large quantities of DNA for WGS library preparation. Furthermore, a key limitation of WGS is that unless large sample sizes are used, directionality of transmission between species cannot be determined (4, 60–62). The technology for producing diagnostic-quality MTBC WGS directly from specimens is being developed, although still in its infancy. The greater resolution offered by NGS, in combination with a growing availability of M. bovis isolates with WGS, will provide a database for future epidemiological investigations in wildlife, similar to what is being done to trace livestock tuberculosis outbreaks (63).
Despite improvements in directly identifying MTBC organisms, which provides a definitive diagnosis by confirming the presence of the pathogen in a sample or animal, detection of host immune responses remains the principal method used to diagnose M. bovis infection in wildlife. Indirect methods rely on measuring the anamnestic cell-mediated or humoral immune responses to M. bovis antigens in different species (Table 3).
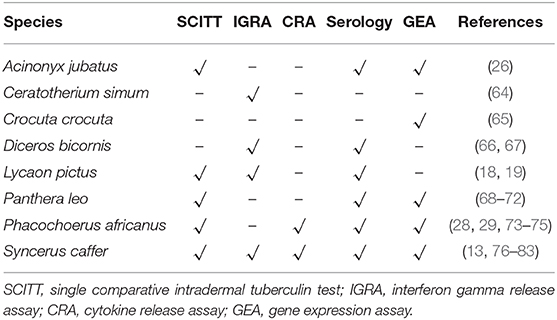
Table 3. Indirect diagnostic tools to detect immune sensitization to Mycobacterium bovis in free-ranging wildlife species in South Africa.
Assays that detect humoral responses use samples (sera) that are typically more readily available than assays based on cell-mediated immunity (CMI), which often require timely processing of fresh whole blood. However, CMI-based assays have shown greater sensitivity in many species (10, 68) and may detect mycobacterial infections earlier than serological assays (84). This may vary with species as more recent studies have shown that humoral responses are better indicators of MTBC infection in some species such as common warthogs and African elephants (29, 85).
The MTBC antigens used to elicit in vitro immune responses in wildlife can either be complex antigens, such as M. bovis purified protein derivative (PPD) or specific mycobacterial peptides (culture filtrate protein 10 and early secretory antigenic target 6 protein) like those in the QuantiFERON® TB-Gold Plus (QFT) system (Qiagen, Venlo, Limburg, Netherlands). The QFT system is a set of tubes, with standardized antigenic peptides and controls, that has been used with whole blood to stimulate MTBC-specific cytokine production in vitro from a number of wildlife species including African buffaloes, common warthogs, African lions, African wild dogs, white rhinoceros and cheetahs (9, 43, 65, 69, 73, 76, 86). These samples can be processed in most veterinary clinics, avoiding the need to get samples to a laboratory within the required 8–10 h for subsequent cytokine release assays. However, species-specific cytokine detection methods, such as enzyme-linked immunosorbent assays (ELISA) or gene expression assays (GEA), still need to be developed and validated for many wildlife species. In addition, cytokine biomarkers for tuberculosis often vary between species; for example, interferon-gamma (IFN-γ) release assays (IGRA) are available for African wild dogs, white rhinoceroses, and African buffaloes, but CXCL9 and CXCL10 gene expression assays are used as diagnostic assays for African lion, cheetah, and common warthog (43, 65, 69, 73).
In vivo CMI tests and the presentation of clinical disease may also be used as indirect methods of detecting bTB. However, clinical assessment is an insensitive tool for diagnosis as clinical disease is typically only observed in advanced stages and may only be practical for use in captive wildlife (68, 87).
The single comparative intradermal tuberculin test (SCITT) relies on stimulating a CMI response in vivo with the intradermal injection of bovine and avian PPDs, after which a memory response is measured by a delayed-type hypersensitivity reaction. The SCITT is applied across multiple wildlife species including African buffalo, African wild dog, African lion, cheetah, leopard, antelope, and common warthog, however, validation has only been performed in a few of these species (70, 74, 77).
The need to calculate a SCITT species-specific cut-off value with appropriate study cohorts was highlighted in a study in which previously published criteria used to interpret SCITT for lions led to false positive results in 54% of animals tested (70, 71). The study by Roos et al. (74) in a bTB endemic cohort of common warthogs demonstrated that M. bovis-infected animals develop a measurable delayed-type hypersensitivity reaction to M. bovis PPD and the sensitivity and specificity of the SCITT, using common warthog-specific cut-off values, were 81 and 100%, respectively (74). Currently, the SCITT is performed in African buffaloes using the interpretation criteria for cattle, leading to suboptimal sensitivity (78). In some species, such as elephants and rhinoceroses, the SCITT is unreliable and not recommended for use, although the response to intradermal injection of specific antigens instead of PPDs should be investigated as a potential option (88). Due to the cost and logistical problems associated with performing the SCITT in wild animals, blood-based assays, namely cytokine release assays or GEA, that require only a single immobilization and blood collection and can deliver a result in <72 h, would be valuable wildlife tools for bTB surveillance and diagnosis.
The cytokine IFN-γ is the most used biomarker to quantify in vitro CMI responses to MTBC in humans, livestock, and wildlife. Commercially available ELISAs have been evaluated and optimized to detect cytokine production in specific wildlife species; the Bovigam® and Bovigam® 2G ELISAs (Prionics AG, Schlieren-Zurich, Switzerland), ruminant cattletype® IFN-gamma ELISA (INDICAL, Inc., San Francisco CA, USA), and Mabtech bovine interferon-gamma (IFN-γ) ELISAPRO kit (Mabtech, Nacka Strand, Sweden) detect African buffalo IFN-γ (76, 77, 79), the Bovigam® detects nyala IFN-γ (89), the Quantikine® canine IFN-γ ELISA (R&D Systems, Inc., Minneapolis, MN, USA) detects wild dog IFN-γ (45), and the equine IFN-γ ELISAPRO (Mabtech) detects white rhinoceros IFN-γ (9). Moreover, alternative biomarkers to IFN-γ have been investigated, with the chemokine IFN-γ-inducible protein-10 (IP-10) shown to be a biomarker of immune activation in African buffaloes and common warthogs (13, 75).
Since no single cytokine release assay has perfect sensitivity and specificity (80), biomarker panels are being investigated to improve diagnostic performance. The parallel measurement of IFN-γ and IP-10 has been shown to maximize the detection of M. bovis-infected African buffaloes while maintaining specificity (13). In some wildlife species, it may be difficult to develop cytokine release assays due to the lack of species-specific reagents; in these cases, GEA may be an alternative method of determining antigen-specific immune responses.
Gene expression analysis quantifies a change in antigen-specific cytokine gene transcription reflecting an immune response. These assays have shown utility in the diagnosis of M. bovis infection in African lions and cheetahs by measuring the expression of CXCL9 mRNA (26, 69), in African buffaloes by measuring the expression of IFN-γ mRNA (81) as well as in warthogs (73) and white rhinoceros (90) by measuring the expression of CXCL10 mRNA. Furthermore, GEA has been used to detect immune sensitization to M. bovis in spotted hyena (Crocuta crocuta), by measuring the expression of CXCL9 and CXCL11 mRNA (65). In species such as African elephants, this approach could be used to circumvent the requirement to produce unique species-specific reagents, such as antibodies to elephant cytokines. Published cytokine gene primer sequences may facilitate use of these assays by additional laboratories without the requirement to share non-commercially available reagents.
Antigen-specific circulating antibodies can be used to differentiate between M. bovis-infected and -uninfected individuals in some species. In common warthogs, an indirect in-house PPD ELISA and the TB ELISA-VK® kit (Vacunek, Bizkaia, Spain) were able to detect M. bovis-specific antibodies with high sensitivity and specificity (29). However, variable test performance was observed when a commercial TB ELISA (IDEXX Laboratories Inc., Westbrook, ME, USA) was used to detect M. bovis-specific antibodies in naturally infected African buffaloes from herds with different bTB prevalence (82). The low sensitivity of serological assays for buffaloes was also observed when serum antibodies were detected to specific mycobacterial peptides using rapid lateral flow assays (91), which suggests that these may not be useful diagnostic tests for this species. In contrast, the Dual Path Platform Vet TB Assay (DPP® VetTB) for Cervids® (Chembio Diagnostic Systems, Medford, NY, USA) has been shown to be a sensitive test to diagnose M. bovis infection in common warthogs (29) and the Elephant TB STAT-PAK® Assay (Chembio Diagnostic Systems) and the DPP® VetTB for Elephants (Chembio Diagnostic Systems) have been used to determine the seroprevalence of MTBC infection in African elephants in KNP (85). The Elephant TB STAT-PAK® and DPP VetTB® assays have also been shown to distinguish between M. bovis-infected and -unexposed African lion populations (68, 69, 72). These species-non-specific rapid tests have also been useful for identifying M. bovis-infected white and black rhinoceros, and a cheetah (26, 34, 66). Sensitivity of serological tests varies between species, although providing a useful tool for retrospective surveys (68, 85).
Diagnostic tests are used by veterinary clinicians, researchers, and animal health regulatory staff to detect infection with M. bovis in individual animals as well as screen herds or perform surveillance in populations. A recent review described current antemortem and postmortem diagnostic tests for wildlife from a global perspective (92). Therefore, the current review focused on the use of diagnostic test specifically for South African wildlife species since application of techniques can vary between hosts as well as geographical location (10, 93). In addition, to accurately interpret results, a diagnostic test should be validated and “fit for purpose,” according to the OIE (11). For example, the OIE has validated the tuberculin skin test in cattle to permit international trade (11) and in South Africa, this has also been approved for certifying African buffaloes for translocation. Although many routine tests for bTB (developed in domestic animals) are used in wildlife, most have not been validated for these species (11).
In general, tests which rely on direct detection of M. bovis are less likely to vary significantly in test performance between species and those validated in a domestic species should be relevant for other related species. However, it is important to be aware of variation in the amount and distribution of the pathogen in different hosts (87). Histological examination may reveal differences in specific pathological changes between species, but the presence of a granuloma, especially with acid-fast positive bacteria present, is a presumptive diagnosis for bTB (10, 11, 94). In addition, collection of respiratory secretions by nasal swabs/lavage, oropharyngeal swabs, and bronchoalveolar lavage for mycobacterial culture or mycobacterial PCR have been applied across a wide range of wildlife species, with positive results confirming infection (36, 42, 43). However, the location of the infection, sporadic shedding, suboptimal sensitivity of mycobacterial culture, and inability of PCR to distinguish between live and dead bacteria are limitations to direct detection techniques for bTB diagnosis, especially antemortem. According to OIE guidelines, “detection of infection in a wildlife population requires bacteriological investigation or the use of a valid testing method for the species involved (the tuberculin test is not effective in all species) together with epidemiological analysis of information” (11).
In contrast to direct detection methods, indirect detection relies on the host's immune responses which can vary significantly between species. This requires knowledge of the immunology of each host species, which is often limited. In addition, understanding of pathogenesis and changes in associated immunological responses are important in appropriate application of indirect tests. One of the major limitations of these assays is that they often require species-specific reagents, which may not be available, and are costly and time-consuming to develop. Acknowledging that validation of tests for use in wildlife may be challenging due to access to adequate numbers of samples from known infected and uninfected individuals, the OIE has provided a validation pathway for provisional recognition of tests that can be used in specific applications (11). Modification of pre-existing tests that have been validated for a domestic species may be provisionally validated in a related wildlife species using a minimum of 10 positive and 10 negative well-characterized reference samples, while for a new test, the numbers increase to 30 each (11). The provisional validation of tests such as IGRAs for white rhinoceros, African buffaloes, and wild dogs demonstrates that this can be accomplished (64, 79, 95). The OIE criteria make it feasible to provide sufficient information for animal health authorities to approve the use of the test in a specified host species for a specific purpose (such as testing animals being moved or surveillance). Therefore, this review of indirect, blood-based assays that can accurately detect M. bovis infection in specific species demonstrates the potential these tests have for being approved tests for wildlife in the future.
Diagnostic tests for wildlife in other countries are like those described for South African wildlife (10, 93, 96, 97). Direct detection techniques for wildlife TB do not vary significantly between countries. There may be some species in which M. bovis is shed in non-respiratory secretions or infection is localized in specific organs, however the methods used to detect bacilli (i.e., microscopy, culture, PCR) are applicable to different sample types (3, 87, 97). In contrast, immunological assays can vary between species, and may be more limited due to lack of species-specific or cross-reactive reagents. The tuberculin skin test is commonly used in a wide range of wildlife species but rarely validated (10, 96, 97). Non-specific or cross-reactions may be more common in some species such as tapirs, camelids, and orangutans, which leads to recommendations against using the test in some groups of animals (97). Interferon gamma release assays have been used in badgers, cervids, bovids, and primates with variable sensitivity and specificity (10, 93, 96, 97). Serological assays, such as STAT-PAK, DPP, and multi-antigen print immunoassay (MAPIA), have also been used in a range of species, however, the humoral response is often only detected later in disease development although elephants, camelids, and suids appear to develop robust early responses (97–100). Therefore, the clinician selecting tests for diagnosing TB in wildlife should base the choice on knowledge of species-specific immune responses and pathogenesis. With the criteria provided by the OIE, more assays will potentially be provisionally approved which will facilitate TB screening of wildlife for movement and surveillance.
During the last decade, an increasing number of wildlife species have been confirmed to be infected with M. bovis in South Africa (Figure 1). The identification of additional host species infected with M. bovis may be due to an increase in transmission or an improvement in detection, the latter being at the forefront of research. Direct detection of M. bovis remains difficult and time-consuming due to the paucity of bacilli in most antemortem samples, together with the slow growth of mycobacteria. However, development of rapid PCR assays may improve direct detection of MTBC in the future, particularly since some of the technology can be performed in a field setting.
Indirect detection of M. bovis through quantification of immune responses elicited by infection, especially in vitro tests, has key advantages in wild animals, and remains the cornerstone of wildlife bTB testing. The development of species-specific reagents and the identification of cross-reactive reagents, as well as standardized kits, will further improve the detection of M. bovis in wildlife species.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This work was partially supported by the South African government through the South African Medical Research Council, National Research Foundation of South Africa (SARChI grant 86949), American Association of Zoo Veterinarians Wild Health Fund, Harry Crossley Foundation, AD and RW acknowledge the Tuberculosis Omics Research (TORCH) Consortium headed by Prof. Annelies van Rie for financial support. AD also acknowledges financial support from the European & Developing Countries Clinical Trial Partnership (TMA2018CDF-2374). Funding to TK was provided through NRF SAMRC CTB Postdoctoral Fellowship. Funding to WG was provided by National Geographic Society Explorer Grant (S006337).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Gagneux S. Ecology and evolution of Mycobacterium tuberculosis. Nat Rev Microbiol. (2018) 16:202–13. doi: 10.1038/nrmicro.2018.8
2. Michel AL, Bengis RG, Keet DF, Hofmeyr M, de Klerk LM, Cross PC, et al. Wildlife tuberculosis in South African conservation areas: Implications and challenges. Vet Microbiol. (2006) 112:91–100. doi: 10.1016/j.vetmic.2005.11.035
3. Fitzgerald SD, Kaneene JB. Wildlife reservoirs of bovine tuberculosis worldwide: hosts, pathology, surveillance, and control. Vet Pathol. (2013) 50:488–99. doi: 10.1177/0300985812467472
4. Hlokwe TM, de Klerk-Lorist L-M, Michel AL. Wildlife on the move: a hidden tuberculosis threat to conservation areas and game farms through introduction of untested animals. J Wildl Dis. (2016) 52:837–43. doi: 10.7589/2015-10-281
5. De Garine-Wichatitsky M, Caron A, Kock R, Tschopp R, Munyeme M, Hofmeyr M, et al. A review of bovine tuberculosis at the wildlife–livestock–human interface in sub-Saharan Africa. Epidemiol Infect. (2013) 141:1342–56. doi: 10.1017/S0950268813000708
6. Hlokwe TM, van Helden PD, Michel AL. Evidence of increasing intra and inter-species transmission of Mycobacterium bovis in South Africa: Are we losing the battle? Prev Vet Med. (2014) 115:10–7. doi: 10.1016/j.prevetmed.2014.03.011
7. Musoke J, Hlokwe T, Marcotty T, du Plessis BJA, Michel AL. Spillover of Mycobacterium bovis from wildlife to livestock, South Africa. Emerg Infect Dis. (2015) 21:448–51. doi: 10.3201/eid2103.131690
8. Meiring C, van Helden PD, Goosen WJ. TB control in humans and animals in South Africa: a perspective on problems and successes. Front Vet Sci. (2018) 5:1–7. doi: 10.3389/fvets.2018.00298
9. Chileshe J, Goosen WJ, Buss PE, van Helden PD, Warren R, Parsons SDC, et al. A commercial ELISA for detection of interferon gamma in white rhinoceros. J Vet Diagnostic Investig. (2019) 31:531–6. doi: 10.1177/1040638719843955
10. Maas M, Michel AL, Rutten VPMG. Facts and dilemmas in diagnosis of tuberculosis in wildlife. Comp Immunol Microbiol Infect Dis. (2013) 36:269–85. doi: 10.1016/j.cimid.2012.10.010
11. Oie.int. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019: OIE – World Organisation For Animal Health. (2019). Available online at: https://www.oie.int/standard-setting/terrestrial-manual/access-online (accessed December 28, 2020).
12. Hlokwe TM, Jenkins AO, Streicher EM, Venter EH, Cooper DV, Godfroid J, et al. Molecular characterisation of Mycobacterium bovis isolated from African buffaloes (Syncerus caffer) in Hluhluwe-iMfolozi Park in KwaZulu-Natal, South Africa. Onderstepoort J Vet Res. (2011) 78:1–6. doi: 10.4102/ojvr.v78i1.232
13. Bernitz N, Kerr TJ, Goosen WJ, Clarke C, Higgitt R, Roos EO, et al. Parallel measurement of IFN-γ and IP-10 in QuantiFERON®-TB Gold (QFT) plasma improves the detection of Mycobacterium bovis infection in African buffaloes (Syncerus caffer). Prev Vet Med. (2019) 169:104700. doi: 10.1016/j.prevetmed.2019.104700
14. Bengis RG, Kriek NP, Keet DF, Raath JP, de Vos V, Huchzermeyer HF. An outbreak of bovine tuberculosis in a free-living African buffalo (Syncerus caffer-Sparrman) population in the Kruger National Park: a preliminary report. Onderstepoort J Vet Res. (1996) 63:15–8.
15. Miller MA, Kerr TJ, de Waal CR, Goosen WJ, Streicher EM, Hausler G, et al. Detection of Mycobacterium bovis infection in free-ranging African elephants (Loxodonta africana) and importance of mycobacterial speciation in understanding epidemiology of TB. Emerg Infect Dis. (2021). in press.
16. Michel AL, Coetzee ML, Keet DF, Maré L, Warren R, Cooper DV, et al. Molecular epidemiology of Mycobacterium bovis isolates from free-ranging wildlife in South African game reserves. Vet Microbiol. (2009) 133:335–43. doi: 10.1016/j.vetmic.2008.07.023
17. Michel AL. Implications of tuberculosis in African wildlife and livestock. Ann N Y Acad Sci. (2002) 969:251–5. doi: 10.1111/j.1749-6632.2002.tb04387.x
18. Higgitt RL. Characterization of Mycobacterium bovis specific immune responses in African wild dogs (Lycaon pictus) (MSc thesis). Stellenbosch University, Stellenbosch, South Africa (2018).
19. Higgitt RL, van Schalkwyk OL, de Klerk-Lorist L-M, Buss PE, Caldwell P, Rossouw L, et al. An interferon gamma release assay for the detection of immune of immune sensitization to Mycobacterium bovis in African wild dogs (Lycaon pictus). J Wildl Dis. (2019) 55:529. doi: 10.7589/2018-03-089
20. Brüns AC, Tanner M, Williams MC, Botha L, O'Brien A, Fosgate GT, et al. Diagnosis and implications of Mycobacterium bovis infection in banded mongooses (Mungos mungo) in the Kruger National Park, South Africa. J Wildlife Dis. (2017) 53:19–29. doi: 10.7589/2015-11-318
21. Miller MA, Buss PE, van Helden PD, Parsons SDC. Mycobacterium bovis in a free-ranging black rhinoceros, Kruger National Park, South Africa, 2016. Emerg Infect Dis. (2017) 23:557–8. doi: 10.3201/eid2303.161622
22. Michel AL, Bengis RG. The African buffalo: a villain for inter-species spread of infectious diseases in southern Africa. Onderstepoort J Vet Res. (2012) 79:453. doi: 10.4102/ojvr.v79i2.453
23. Keet DF, Kriek NP, Penrith ML, Michel A, Huchzermeyer H. Tuberculosis in buffaloes (Syncerus caffer) in the Kruger National Park: spread of the disease to other species. Onderstepoort J Vet Res. (1996) 63:239–44.
24. Keet DF, Kriek NP, Bengis RG, Grobler DG, Michel A. The rise and fall of tuberculosis in a free-ranging chacma baboon troop in the Kruger National Park. Onderstepoort J Vet Res. (2000) 67:115–22.
25. Michel AL, Hlokwe TM, Espie IW, van Zijll Langhout M, Koeppel K, Lane E. Mycobacterium tuberculosis at the human/wildlife interface in a high TB burden country. Transbound Emerg Dis. (2013) 60:46–52. doi: 10.1111/tbed.12099
26. Kerr TJ, Gumbo R, Goosen WJ, Rogers P, Last RD, Miller MA. Novel techniques for detection of Mycobacterium bovis infection in a cheetah. Emerg Infect Dis. (2020) 26:630–1. doi: 10.3201/eid2603.191542
27. Paine R, Martinaglia G. Tuberculosis in wild buck living under natural conditions. J Comp Pathol Ther. (1929) 42:1–8. doi: 10.1016/S0368-1742(29)80001-X
28. Roos EO. Detection and characterization of mycobacterial infections occurring in Phacochoerus africanus (Gmelin, 1788) (Common Warthog) (PhD Thesis). Stellenbosch University, Stellenbosch, South Africa (2018).
29. Roos EO, Buss PE, de Klerk-Lorist L-M, Hewlett J, Hausler GA, Rossouw L, et al. Test performance of three serological assays for the detection of Mycobacterium bovis infection in common warthogs (Phacochoerus africanus). Vet Immunol Immunopathol. (2016) 182:79–84. doi: 10.1016/j.vetimm.2016.10.006
30. Hlokwe TM, Michel AL, Mitchel E, Gcebe N, Reininghaus B. First detection of Mycobacterium bovis infection in giraffe (Giraffa camelopardalis) in the Greater Kruger National Park complex: role and implications. Transbound Emerg Dis. (2019) 66:2264–70. doi: 10.1111/tbed.13275
31. Bengis RG, Keet DF, Michel AL, Kriek NP. Tuberculosis, caused by Mycobacterium bovis, in a kudu (Tragelaphus strepsiceros) from a commercial game farm in the Malelane area of the Mpumalanga Province, South Africa. Onderstepoort J Vet Res. (2001) 68:239–41.
32. Kerr TJ, Goosen WJ, Gumbo R, de Klerk-Lorist L-M, Pretorius O, Buss PE, et al. Diagnosis of Mycobacterium bovis infection in free-ranging common hippopotamus (Hippopotamus amphibius). Transb Emerg Dis. (2021). doi: 10.1111/tbed.13989. [Epub ahead of print].
33. Gouws TA, Williams MC. The pathology of tuberculosis caused by Mycobacterium tuberculosis in a herd of semi-free-ranging springbok (Antidorcas marsupialis). Onderstepoort J Vet Res. (2009) 76:419–41. doi: 10.4102/ojvr.v76i4.26
34. Miller MA, Buss PE, Parsons SDC, Roos EO, Chileshe J, Goosen WJ, et al. Conservation of white rhinoceroses threatened by bovine tuberculosis, South Africa, 2016–2017. Emerg Infect Dis. (2018) 24:2373–5. doi: 10.3201/eid2412.180293
36. Michel AL, Lane EP, de Klerk-Lorist L-M, Hofmeyr M, van der Heijden EMDL, Botha L, et al. Experimental Mycobacterium bovis infection in three white rhinoceroses (Ceratotherium simum): susceptibility, clinical and anatomical pathology. PLoS ONE. (2017) 12:1–16. doi: 10.1371/journal.pone.0179943
37. Miller M, Buss P, de Klerk-Lorist L-M, Hofmeyr J, Hausler G, Lyashchenko K, et al. Application of rapid serological tests for detection of Mycobacterium bovis infection in free-ranging warthogs (Phacochoerus africanus) – implications for ante-mortem disease screening. J Wildl. Dis. (2016) 52:180–2. doi: 10.7589/2015-07-186
38. Malama S, Munyeme M, Mwanza S, Muma JB. Isolation and characterization of non-tuberculous mycobacteria from humans and animals in Namwala District of Zambia. BMC Res Notes. (2014) 7:622. doi: 10.1186/1756-0500-7-622
39. Miller M, Terrell S, Lyashchenko K, Greenwald R, Harris B, Thomsen B, et al. Mycobacterium kansasii infection in a bontebok (Damaliscus pygargus dorcas) herd: Diagnostic challenges in differentiating from the Mycobacterium tuberculosis complex. J Zoo Wildl Med. (2011) 42:468–72.
40. Love DM, Garner MM, Lyashchenko KP, Sikar-Gang A, Bradway DS, Robbe-Austerman S, et al. Tuberculosis caused by Mycobacterium orygis in a greater one-horned rhinoceros (Rhinoceros unicornis): first report in the western hemisphere. J Zoo WildlMed. (2020) 50:1000–4. doi: 10.1638/2018-0084
41. Robbe-Austerman S, Bravo DM, Harris B. Comparison of the MGIT 960, BACTEC 46 TB and solid media for isolation of Mycobacterium bovis in United States veterinary specimens. BMC Vet Res. (2013) 9:74–9. doi: 10.1186/1746-6148-9-74
42. Miller MA, Buss PE, Hofmeyr J, Olea-Popelka F, Parsons SDC, van Helden PD. Antemortem diagnosis of Mycobacterium bovis infection in free-ranging African lions (Panthera leo) and implications for transmission. J Wildl Dis. (2015) 51:493–7. doi: 10.7589/2014-07-170
43. Goosen WJ, Kerr TJ, Kleyhans L, Warren RM, van Helden PD, Persing DH, et al. The Xpert MTB/RIF Ultra assay detects Mycobacterium tuberculosis complex DNA in white rhinoceros (Ceratotherium simum) and African elephants (Loxodonta africana). Sci Rep. (2020) 10:14482. doi: 10.1038/s41598-020-71589-9
44. Meiring C, Higgitt R, Dippenaar A, Roos E, Buss P, Hewlett J, et al. Characterizing epidemiological and genotypic features of Mycobacterium bovis infection in wild dogs (Lycaon pictus). Transb Emerg Dis. (2020). doi: 10.1111/tbed.13947
45. Higgitt RL, van Schalkwyk OL, de Klerk-Lorist L-M, Buss PE, Caldwell P, Rossouw L, et al. Mycobacterium bovis infection in African wild dogs, Kruger National Park, South Africa. Emerg Infect Dis. (2019) 25:1425–7. doi: 10.3201/eid2507.181653
46. Bull TJ, Munshi T, Mikkelsen H, Hartmann SB, Sørensen MR, Garcia JS, et al. Improved culture medium (TiKa) for Mycobacterium avium subspecies paratuberculosis (MAP) matches qPCR sensitivity and reveals significant proportions of non-viable MAP in lymphoid tissue of vaccinated MAP challenged animals. Front Microbiol. (2017) 7:1–8. doi: 10.3389/fmicb.2016.02112
47. Goosen WJ, Kerr TJ, Kleynhans L, Buss PE, Cooper DV, Warren RM, et al. The VetMAXTM M. tuberculosis complex PCR kit detects MTBC DNA in antemortem and postmortem samples from white rhinoceros (Ceratotherium simum), African elephants (Loxodonta africana) and African buffaloes (Syncerus caffer). BMC Vet Res. (2020) 16:220. doi: 10.1186/s12917-020-02438-9
48. World Health Organisation. Global Tuberculosis Report. (2015). Available online at: www.who.int/tb/publications/global_report/gtbr15_main_text.pdf (accessed May 5, 2020).
49. Hlokwe TM, Mogano RM. Utility of xpert® MTB/RIF ultra assay in the rapid diagnosis of bovine tuberculosis in wildlife and livestock animals from South Africa. Prev Vet Med. (2020) 177:104980. doi: 10.1016/j.prevetmed.2020.104980
50. Kamerbeek J, Schouls L, Kolk A, van Agterveld M, van Soolingen D, Kuijper S, et al. Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology. J Clin Microbiol. (1997) 35:907–14.
51. Brudey K, Gutierrez MC, Parsons LM, Salfinger M, Rastogi N, Sola C. Mycobacterium africanum genotyping using novel spacer oligonucleotides in the direct repeat locus. J Clin Microbiol. (2004) 42:5053–7. doi: 10.1128/JCM.42.11.5053
52. Supply P, Allix C, Lesjean S, Cardoso-Oelemann M, Rusch-Gerdes S, Willery E, et al. Proposal for standardization of optimized mycobacterial interspersed repetitive unit-variable-number tandem repeat typing of Mycobacterium tuberculosis. J Clin Microbiol. (2006) 44:4498–510. doi: 10.1128/JCM.01392-06
53. Dippenaar A, Parsons SDC, Miller MA, Hlokwe T, Gey van Pittius NC, Adroub SA, et al. Progenitor strain introduction of Mycobacterium bovis at the wildlife-livestock interface can lead to clonal expansion of the disease in a single ecosystem. Infect Genet Evol. (2017) 51:235–8. doi: 10.1016/j.meegid.2017.04.012
54. Warren RM, Gey van Pittius NC, Barnard M, Hesseling A, Engelke E, de Kock M, et al. Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference. Int J Tuberc Lung Dis. (2006) 10:818–22.
55. Alexander KA, Laver PN, Michel AL, Williams M, van Helden PD, Warren RM, et al. Novel Mycobacterium tuberculosis complex pathogen, M. mungi. Emerg Infect Dis. (2010) 16:1296–9. doi: 10.3201/eid1608.100314
56. Parsons SDC, Drewe JA, Gey van Pittius NC, Warren RM, van Helden LPD. Novel cause of tuberculosis in meerkats, South Africa. Emerg Infect Dis. (2013) 19:2004–7. doi: 10.3201/eid1912.130268
57. Gey van Pittius NC, Perrett KD, Michel AL, Keet DF, Hlokwe T, Streicher EM, et al. Infection of African buffalo (Syncerus caffer) by oryx bacillus, a rare member of the antelope clade of the Mycobacterium tuberculosis complex. J Wildl Dis. (2012) 48:849–57. doi: 10.7589/2010-07-178
58. Sichewo PR, Hlokwe TM, Etter EM, Michel AL. Tracing cross species transmission of Mycobacterium bovis at the wildlife/livestock interface in South Africa. BMC Microbiol. (2020) 20:1–9. doi: 10.1186/s12866-020-01736-4
59. Ghavidel M, Mansury D, Nourian K, Ghazvini K. The most common spoligotype of Mycobacterium bovis isolated in the world and the recommended loci for VNTR typing; A systematic review. Microb Pathog. (2018) 118:310–5. doi: 10.1016/j.micpath.2018.03.036
60. Biek R, O'Hare A, Wright D, Mallon T, McCormick C, Orton RJ, et al. Whole genome sequencing reveals local transmission patterns of Mycobacterium bovis in sympatric cattle and badger populations. PLoS Pathog. (2012) 8:e1003008. doi: 10.1371/journal.ppat.1003008
61. Crispell J, Zadoks RN, Harris SR, Paterson B, Collins DM, De-Lisle GW, et al. Using whole genome sequencing to investigate transmission in a multi-host system: bovine tuberculosis in New Zealand. BMC Genomics. (2017) 18:180. doi: 10.1186/s12864-017-3569-x
62. Rodríguez-Prieto V, Martínez-López B, Barasona J, Acevedo P, Romero B, Rodriguez-Campos S, et al. A Bayesian approach to study the risk variables for tuberculosis occurrence in domestic and wild ungulates in South Central Spain. BMC Vet Res. (2012) 8:148. doi: 10.1186/1746-6148-8-148
63. Orloski K, Robbe-Austerman S, Stuber T, Hench B, Schoenbaum M. Whole genome sequencing of Mycobacterium bovis isolated from livestock in the United States, 1989–2018. Front Vet Sci. (2018) 5:1–10. doi: 10.3389/fvets.2018.00253
64. Chileshe J, Roos EO, Goosen WJ, Buss PE, Hausler G, Rossouw L, et al. An interferon-gamma release assay for the diagnosis of the Mycobacterium bovis infection in white rhinoceros (Ceratotherium simum). Vet Immunol Immunopathol. (2019) 217:109931. doi: 10.1016/j.vetimm.2019.109931
65. Higgitt RL, Buss PE, Van Helden PD, Miller MA, Parsons SDC. Development of gene expression assays measuring immune responses in the spotted hyena (Crocuta crocuta). African Zool. (2017) 52:99–104. doi: 10.1080/15627020.2017.1309300
66. Miller MA, Greenwald R, Lyashchenko KP. Potential for serodiagnosis of tuberculosis in black rhinoceros (Diceros bicornis). J Zoo Wildl Med. (2015) 46:100–4. doi: 10.1638/2014-0172R1.1
67. Morar D, Schreuder J, Mény M, van Kooten PJS, Tijhaar E, Michel AL, et al. Towards establishing a rhinoceros-specific interferon-gamma (IFN-γ) assay for diagnosis of tuberculosis. Transbound Emerg Dis. (2013) 60:60–6. doi: 10.1111/tbed.12132
68. Miller MA, Joubert J, Mathebula N, de Klerk-Lorist L-M, Lyashchenko KP, Bengis RG, et al. Detection of antibodies to tuberculosis antigens in free-ranging lions (Panthera leo) infected with Mycobacterium bovis in Kruger National Park, South Africa. J Zoo Wildl Med. (2012) 43:317–23. doi: 10.1638/2011-0171.1
69. Olivier TT, Viljoen IM, Hofmeyr J, Hausler GA, Goosen WJ, Tordiffe ASW, et al. Development of a gene expression assay for the diagnosis of Mycobacterium bovis infection in African lions (Panthera leo). Transbound Emerg Dis. (2017) 64:774–81. doi: 10.1111/tbed.12436
70. Keet DF, Michel AL, Bengis RG, Becker P, van Dyk DS, van Vuuren M, et al. Intradermal tuberculin testing of wild African lions (Panthera leo) naturally exposed to infection with Mycobacterium bovis. Vet Microbiol. (2010) 144:384–91. doi: 10.1016/j.vetmic.2010.01.028
71. Viljoen IM, Sylvester TT, Parsons SDC, Millar RP, Helden PD van, Miller MA. Performance of the tuberculin skin test in Mycobacterium bovis-exposed and -unexposed African lions (Panthera leo). J Wildl Dis. (2019) 55:537. doi: 10.7589/2018-06-163
72. Miller MA, Buss P, Sylvester TT, Lyashchenko KP, deKlerk-Lorist L-M, Bengis R, et al. Mycobacterium bovis in free-ranging lions (Panthera leo) – Evaluation of serological and tuberculin skin tests for detection of infection and disease. J Zoo Wildl Med. (2019) 50:7–15. doi: 10.1638/2017-0187
73. Roos EO, Scott LA, Ndou S, Olea-Popelka F, Buss PE, de Klerk-Lorist L-M, et al. Cytokine gene expression assay as a diagnostic tool for detection of Mycobacterium bovis infection in warthogs (Phacochoerus africanus). Sci Rep. (2019) 9:16525. doi: 10.1038/s41598-019-53045-0
74. Roos EO, Olea-Popelka F, Buss PE, Hausler GA, Warren R, van Helden PD, et al. Measuring antigen-specific responses in Mycobacterium bovis-infected warthogs (Phacochoerus africanus) using the intradermal tuberculin test. BMC Vet Res. (2018) 14:360. doi: 10.1186/s12917-018-1685-8
75. Roos EO, Olea-Popelka F, Buss PE, de Klerk-Lorist L-M, Cooper DV, Warren RM, et al. IP-10: A potential biomarker for detection of Mycobacterium bovis infection in warthogs (Phacochoerus africanus). Vet Immunol Immunopathol. (2018) 201:43–8. doi: 10.1016/j.vetimm.2018.05.007
76. Bernitz N, Clarke C, Roos EO, Goosen WJ, Cooper DV, van Helden PD, et al. Detection of Mycobacterium bovis infection in African buffaloes (Syncerus caffer) using QuantiFERON® -TB Gold (QFT) tubes and the Qiagen cattletype® IFN-gamma ELISA. Vet Immunol Immunopathol. (2018) 196:48–52. doi: 10.1016/j.vetimm.2017.12.010
77. Bernitz N, Goosen WJ, Clarke C, Kerr TJ, Higgitt R, Roos EO, et al. Parallel testing increases detection of Mycobacterium bovis-infected African buffaloes (Syncerus caffer). Vet Immunol Immunopathol. (2018) 204:40–3. doi: 10.1016/j.vetimm.2018.09.004
78. Smith K, Bernitz N, Cooper D, Kerr TJ, de Waal CR, Clarke C, et al. Optimisation of the tuberculin skin test for detection of Mycobacterium bovis in African buffaloes (Syncerus caffer). Prev Vet Med. (2021). doi: 10.1016/j.prevetmed.2020.105254
79. Smith K, Bernitz N, Goldswain S, Cooper DV, Warren RM, Goosen WJ, et al. Optimized interferon-gamma release says for detection of Mycobacterium bovis infection in African buffaloes (Syncerus caffer). Vet Immunol Immunopath. (2021) 231:110163. doi: 10.1016/j.vetimm.2020.110163
80. Bernitz N, Kerr TJ, de Waal C, Cooper DV, Warren RM, van Helden PD, et al. Test characteristics of assays to detect Mycobacterium bovis infection in high-prevalence African buffalo (Syncerus caffer) herds. J Wildl Dis. (2020) 56:462–5. doi: 10.7589/2019-06-173
81. Parsons SDC, Menezes AM, Cooper DV, Walzl G, Warren RM, van Helden PD. Development of a diagnostic gene expression assay for tuberculosis and its use under field conditions in African buffaloes (Syncerus caffer). Vet Immunol Immunopathol. (2012) 148:337–42. doi: 10.1016/j.vetimm.2012.04.025
82. van der Heijden EM, Cooper DV, Rutten VP, Michel AL. Mycobacterium bovis prevalence affects the performance of a commercial serological assay for bovine tuberculosis in African buffaloes. Compar Immun Microbiol Infect Dis. (2020) 70:101369. doi: 10.1016/j.cimid.2019.101369
83. van der Heijden EMDL, Jenkins AO, Cooper DV, Rutten VPMG, Michel AL. Field application of immunoassays for the detection of Mycobacterium bovis infection in the African buffalo (Syncerus caffer). Vet Immunol Immunopathol. (2016) 169:68–73. doi: 10.1016/j.vetimm.2015.12.003
84. de la Rua-Domenech R, Goodchild AT, Vordermeier HM, Hewinson RG, Christiansen KH, Clifton-Hadley RS. Ante mortem diagnosis of tuberculosis in cattle: A review of the tuberculin tests, γ-interferon assay and other ancillary diagnostic techniques. Res Vet Sci. (2006) 81:190–210. doi: 10.1016/j.rvsc.2005.11.005
85. Kerr TJ, de Waal CR, Buss PE, Hofmeyr J, Lyashchenko KP, Miller MA. Seroprevalence of Mycobacterium tuberculosis complex in free-ranging African elephants (Loxodonta africana) in Kruger National Park, South Africa. J Wildl Dis. (2019) 55:923–7. doi: 10.7589/2018-12-292
86. Parsons SDC, Gous TA, Warren RM, de Villiers C, Seier JV, van Helden PD. Detection of Mycobacterium tuberculosis infection in chacma baboons (Papio ursinus) using the QuantiFERON-TB Gold (In-Tube) assay. J Med Primatol. (2009) 38:411–7. doi: 10.1111/j.1600-0684.2009.00367.x
87. Renwick AR, White PCL, Bengis RG. Bovine tuberculosis in southern African wildlife: a multi-species host–pathogen system. Epidemiol Infect. (2007) 135:529–40. doi: 10.1017/S0950268806007205
88. Flores-Villalva S, Suárez-Güemes F, Espitia C, Whelan AO, Vordermeier M, Gutiérrez-Pabello JA. specificity of the tuberculin skin test is modified by use of a protein cocktail containing ESAT-6 and CFP-10 in cattle naturally infected with Mycobacterium bovis. Clin Vaccine Immunol. (2012) 19:797–803. doi: 10.1128/CVI.05668-11
89. Roux L, McCall AJ, Michel AL. Detection of native interferon-γ in nyala (Tragelaphus angasii): towards diagnosing tuberculosis. Onderstepoort J Vet Res. (2019) 86:a1796. doi: 10.4102/ojvr.v86i1.1796
90. Chileshe J, Kerr TJ, Kinnear C, Buss PE, van Helden PD, Warren RM, et al. Cytokine biomarker discovery in the white rhinoceros (Ceratotherium simum). Vet Immunol Immunopath. (2021) 232:110168. doi: 10.1016/j.vetimm.2020.110168
91. Lyashchenko KP, Sridhara AA, Johnathan-Lee A, Sikar-Gang A, Lambotte P, Esfandiari, et al. Differential antigen recognition by serum antibodies from three bovid hosts of Mycobacterium bovis infection. Compar Immun Microbiol Infect Dis. (2020) 69:101424. doi: 10.1016/j.cimid.2020.101424
92. Lekko YM, Ooi PT, Omar S, Mazlan M, Ramanoon SZ, Jasni S, et al. Mycobacterium tuberculosis complex in wildlife: Review of current applications of antemortem and postmortem diagnosis. Vet World. (2020) 13:1822–36. doi: 10.14202/vetworld.2020.1822-1836
93. Bezos J, Casal C, Romero B, Schroeder B, Hardegger R, Raeber AJ, et al. Current ante-mortem techniques for diagnosis of bovine tuberculosis. Res Vet Sci. (2014) 97:S44–52. doi: 10.1016/j.rvsc.2014.04.002
94. Michel AL, de Klerk-Lorist L- .M, Buss P, Hofmeyr M, Cooper D, Bengis RG. Tuberculosis in South African wildlife: lions, African buffalo and other species. In: Tuberculosis, Leprosy and Mycobacterial Diseases of Man and Animals: The Many Hosts of Mycobacteria. Boston: CABI (2015). p. 365–85.
95. Rodwell TC, Kriek NP, Bengis RG, Whyte IJ, Viljoen PC, de Vos V, et al. Prevalence of bovine tuberculosis in African buffalo at Kruger National Park. J Wildl Dis. (2001) 37:258–64. doi: 10.7589/0090-3558-37.2.258
96. Chambers MA. Review of the diagnosis of tuberculosis in non-bovid wildlife species using immunological methods – an update of published work since 2009. Transb Emerg Dis. (2013) 60:S14–27. doi: 10.1111/tbed.12094
97. Miller M, Lyashchenko K. Mycobacterial infections in other zoo animals. In: Tuberculosis, Leprosy and Mycobacterial Diseases of Man and Animals: The Many Hosts of Mycobacteria. Boston: CABI (2015). p. 277–95.
98. Lyashchenko KP, Gortazar C, Miller MA, Waters WR. Spectrum of antibody profiles in tuberculous elephants, cervids and cattle. Vet Microbiol. (2018) 214:89–92. doi: 10.1016/j.vetmic.2017.12.013
99. Krajewska-Wedzina M, Didkowska A, Sridhara AA, Elahi R, Johnathan-Lee A, Radulski L, et al. Transboundary tuberculosis: importation of alpacas infected with Mycobacterium bovis from the United Kingdom to Poland and potential for serodiagnostic assays in detecting tuberculin skin test false-negative animals. Transb Emerg Dis. (2020) 67:1306–14. doi: 10.1111/tbed.13471
Keywords: bovine tuberculosis, diagnostics, Mycobacterium bovis, South African wildlife, immunological assays, direct detection of mycobacteria, cytokine release assays, gene expression assays
Citation: Bernitz N, Kerr TJ, Goosen WJ, Chileshe J, Higgitt RL, Roos EO, Meiring C, Gumbo R, de Waal C, Clarke C, Smith K, Goldswain S, Sylvester TT, Kleynhans L, Dippenaar A, Buss PE, Cooper DV, Lyashchenko KP, Warren RM, van Helden PD, Parsons SDC and Miller MA (2021) Review of Diagnostic Tests for Detection of Mycobacterium bovis Infection in South African Wildlife. Front. Vet. Sci. 8:588697. doi: 10.3389/fvets.2021.588697
Received: 29 July 2020; Accepted: 06 January 2021;
Published: 28 January 2021.
Edited by:
Arvo Viltrop, Estonian University of Life Sciences, EstoniaReviewed by:
Jane Budd, Breeding Centre for Endangered Arabian Wildlife, United Arab EmiratesCopyright © 2021 Bernitz, Kerr, Goosen, Chileshe, Higgitt, Roos, Meiring, Gumbo, de Waal, Clarke, Smith, Goldswain, Sylvester, Kleynhans, Dippenaar, Buss, Cooper, Lyashchenko, Warren, van Helden, Parsons and Miller. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michele A. Miller, bWlsbGVyQHN1bi5hYy56YQ==
†Present address: Eduard O. Roos, The Pirbright Institute, Pirbright, United Kingdom
Anzaan Dippenaar, Tuberculosis Omics Research Consortium, Department of Epidemiology and Social Medicine, Faculty of Medicine and Health Sciences, Institute of Global Health, University of Antwerp, Antwerp, Belgium
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.