
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci., 22 January 2021
Sec. Animal Behavior and Welfare
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.612504
This article is part of the Research TopicRuminant Grazing Behavior: A Tool to Improve Product Quality and Ecosystem ServicesView all 13 articles
 Madeline Koczura1,2*†
Madeline Koczura1,2*† Bruno Martin1
Bruno Martin1 Marilena Musci3
Marilena Musci3 Martina Di Massimo3
Martina Di Massimo3 Matthieu Bouchon4
Matthieu Bouchon4 Germano Turille5Michael Kreuzer2
Germano Turille5Michael Kreuzer2 Joel Berard6,7
Joel Berard6,7 Mauro Coppa8
Mauro Coppa8In the mountains, autochthonous and robust breeds are often used to valorize biodiverse grasslands. Along with their lower nutrient requirements, compared to specialized dairy breeds, they are expected to be better adapted to complex environments and valorize grasslands into dairy products of high quality. Therefore, the aim of the present study was to investigate the grazing selection of three contrasting dairy breeds on a biodiverse mountain pasture, and its consequences on milk fatty acid (FA) profile and prevalence of individual terpenes. A dual-purpose breed from the Italian Alps, the Valdostana Red Pied (Va), was compared to Montbéliardes (Mo), more specialized in milk production, and the highly specialized Holsteins (Ho). Diet selection was measured by scan-sampling, calculating selectivity indexes, and collecting simulated bites during two consecutive days in June (end of first grazing cycle) and July (second grazing cycle). Milk samples were collected at each milking during these experimental periods. Yield of milk and its fat and protein contents were measured. Milk FA and terpenes were analyzed by gas chromatographic methods. We tested the effects of breed, period and their interaction in a repeated mixed model, and calculated Pearson's correlations between behavioral data and milk FA as well as terpenes. The Va grazed less mature vegetation than Ho, but this difference was not sufficient to lead to a major breed effect on milk FA profile and prevalence of terpenes. However, the proportion of α-linolenic acid (C18:3 n-3) was always higher in the milk fat of Va than Ho (Mo were intermediary), but this without any correlation to grazing selection. This could be a consequence from a different metabolism concerning ruminal biohydrogenation, but must be further investigated. Finally, we confirmed previous studies that highlighted a link between milk quality and cows' grazing behavior, but here without differences among breeds. All cows adapted their behavior to the herbage evolution during the season, leading to higher proportions of unsaturated FA in July than June milks. Our study suggests that under mountain grazing conditions (biodiverse pasture and cows in late lactation), milk quality depends more on herbage composition than on cow breed.
Politics and consumers are increasingly concerned about the impact of livestock systems on environment and animal welfare, which triggers intensive research activities. Livestock systems need to be multi-functional. This means that they have to perform well in ecology, animal welfare and economics while producing healthy and quality food. The valorization of biodiverse pastures by ruminants is one of the key tools toward establishing this kind of livestock systems, characterized by higher resilience and fewer inputs than common systems (1). In mountain grazing dairy systems, autochthonous or robust cow breeds are commonly used to valorize these grasslands (2). Robustness is defined as an ability to adapt and carry on casual activities while facing environmental constraints (3). In the mountains, robust breeds with lower nutrient requirements, compared to specialized breeds, are expected to develop a natural resilience to their local environment and might better adapt to utilize the local grass resource, typically characterized by low nutritional value, and to valorize these resources by generating high-quality dairy products.
The ability to efficiently exploit local natural grasslands depends on grazing behavior, which can in turn influence the quality and specificity of dairy products (4). Plant secondary compounds (PSC) play a specific role in this context, such as phenols and terpenes, which are present in greater quantity in upland biodiverse grasslands (5–7). Besides species diversity, also vegetation stage and environmental conditions are involved in influencing the synthesis of terpenoids by the plants. For instance, Fraisse et al. (8) identified 170 different compounds on the same pasture, of which only 30 were common to all vegetation species and growth stages. These PSC may be transferred to the milk (9). Accordingly, milk from pastures rich in dicotyledons contains a greater quantity and wider diversity of terpenes than milk from pastures consisting mainly of grass species (10–12). Some PSC are also active in inhibiting the final reduction step of the biohydrogenation of fatty acids (FA) in the rumen (13–15). Therefore, when cows are grazing on biodiverse mountain pastures, their milk is richer in monounsaturated FA (MUFA), polyunsaturated FA (PUFA) and, especially, in conjugated linoleic acids (CLA) and n-3 FA (16–18). Terpenes and FA in milk are of interest for the sensory properties of milk and other quality attributes of the processed cheese (19).
Dairy cattle grazing on biodiverse pastures express preference or aversion for some specific plants that can vary according to their nutrient requirements and experience. Therefore, the actual diet ingested by the animals might differ from the average vegetation available on the pasture (20, 21), and this according to individual or breed-specific behavior (22, 23). Consequently, FA and terpene composition of the milk might vary among animals grazing the same biodiverse grassland. We reported a first part of the present study in Koczura et al. (24), showing that specialized dairy breeds with high nutrient requirements like Holstein cows (Ho) selected more grasses than dual-purpose breeds like Valdostana Red Pied cows (Va), an autochthonous Italian alpine breed with low requirements, when grazing on heterogenous and biodiverse pastures. The Va were generally less selective toward forbs and mature vegetation. Hence, the aim of the present study was to (i) deepen the analysis of grazing behavior of the three contrasting breeds by quantifying their diet selection on the biodiverse pasture and (ii) further investigate the consequences of this behavior on FA profile and prevalence of terpenes in the milk. We hypothesized that, as a result of our behavioral observations (24), the ingested diet of autochthonous Va cows would be more diverse than that of highly specialized breeds like Ho. Diet of Montbéliarde cows (Mo), which are less specialized for high milk production than Ho and supposedly better adapted to grazing on upland pastures, would be intermediate. We expected that the milk of the individual cows and breeds selecting less grasses and more forbs would have a higher prevalence of MUFA, PUFA, n-3 FA and terpenes.
The present experiment was carried out in 2017 at Marcenat, INRAE's experimental farm. Other aspects and details of the experiment were described in Koczura et al. (24), in which data obtained in the beginning of June 2017 had also been included. Twelve late-lactating dairy cows (four Ho, four Mo, and four Va) grazing on a biodiverse pasture since the beginning of June were monitored during two consecutive days in the end of June (end of first grazing cycle) and then in July (beginning of the second grazing cycle). Briefly, cows grazed (extensive continuous grazing conditions) a natural and highly biodiverse pasture (65 species; 48% of grasses, 13% of legumes, and 39% of forbs on ground cover), dominated by Festuca gr. rubra (18%), and Agrostis capillaris (15%). Before the start of the experiment, the botanical composition (% of ground cover) was determined using the vertical point-quadrat method (25) (Table 1). The characteristics of the herbage on the experimental plots is described in Table 2. Cows did not receive any supplementation with concentrate, and had free access to water and mineral supplements. The individual cow's potential intake capacity, calculated according to INRA (26), was used as an estimation of herbage intake.

Table 1. Botanical composition of the experimental plot.
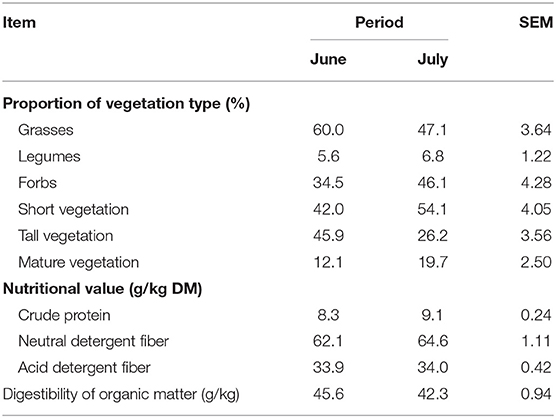
Table 2. Characterization of vegetation offered on the experimental plots.
During the two-day experimental periods, behavioral observations were performed each day by scan-sampling of the cows' bites at 5-min intervals. From these data, Jacob's index of selectivity (IS) was calculated as described in detail by Koczura et al. (24). These indices range from −1 (aversion) to +1 (preference). Additionally, on the same observation days, simulated bites were collected, according to the procedure described by Coppa et al. (27). Individual simulated bites were sampled several times for each cow during the days of observation, and immediately stored at 4°C. The different simulated bite samples were pooled to constitute one sample per period per cow, in order to be representative for the grazed herbage. From this herbage sample, two sub-samples were created: one of them was oven-dried at 60°C for 72 h and analyzed for crude protein (CP) (28), neutral detergent fiber (NDF) and acid detergent fiber (ADF) contents (29), as well as solubility in pepsin and cellulase (30) as an estimate of the organic matter digestibility (OMD). The second sub-sample was sorted into green and dry vegetation. Then, the green herbage was sorted into the three main botanical groups: grasses, legumes and forbs. All fractions were separately oven-dried (in the same conditions as these applied to the first sub-sample) and weighed. The proportion of each fraction to total dry matter (DM) was calculated and was referred to potential intake capacity to estimate the relative intake.
In each experimental period, the individual milk yield of the cows was monitored at each milking during the two consecutive observation days, and 100-mL milk samples were collected. A 30-mL sub-sample was preserved with bronopol-B2, stored at 4°C and analyzed for fat and protein contents by Fourier transformed infrared spectroscopy [MilkoScan 4000, Foss System, Hillerød, Denmark, (31)] following the International Dairy Federation (32). Another sub-sample was stored at −20°C until further analyses.
A 3-mL milk sample was stored at −20°C before lyophilization (Thermovac TM-20, Froilabo, Ozoir-La-Ferrière, France) for FA analysis, performed as described by Ferlay et al. (33). Lipids were directly methylated using 2 mL of 0.5 M Sodium methoxide plus 1 mL of hexane at 50°C for 5 min, followed by cooling with the addition of 75μl of 12M HCl at room temperature for 10 min. The FA methyl esters were recovered in 3 mL hexane and washed with 3 mL water. Samples were injected by auto-sampler into a TraceGC 2000 series gas chromatograph equipped with a flame ionization detector (Thermo Finnigan, Les Ulis, France). Methyl esters from all the samples were separated on a 100 m 30.25 mm i.d. fused-silica capillary column (CP-Sil 88, Chrompack, Middelburg, The Netherlands). The injector temperature was maintained at 250°C and the detector temperature at 255°C. The initial oven temperature was held at 70°C for 1 min, increased by 5°C/min to 100°C (held for 2 min), and then increased by 10°C/min to 175°C (held for 40 min), and 5°C/min to a final temperature of 225°C (held for 15 min). The carrier gas was hydrogen. Identification of trans isomers of 18:1, non-conjugated 18:2, and CLA isomers was as described in Loor et al. (34). A reference standard butter (CRM 164, Commission of the European Communities, Community Bureau of Reference, Brussels, Belgium) was used to estimate correction factors for short-chain FA (C4:0 to C10:0). The de novo synthesized FA were defined as the sum of individual FA synthesized in the mammary gland.
A balanced sample of milk from the four 50-mL samples from morning and evening milking that had been stored in 200-mL glass bottles at −20°C was thawed at ambient temperature for 6 h. The supernatant was then collected and centrifuged at 20,000 rpm and 25°C for 2 h. The anhydrous fat of these samples was collected and stored in 2 mL glass vials until further analysis for terpene composition. Then, headspace-solid phase microextraction (HS-SPME) was carried out using a 50/30 μm divinylbenzene-carboxen-polydimethylsiloxane (DVB/Carboxen/PDMS) fiber (Supelco, Bellefonte, PA, USA). The extraction conditions were the following: equilibration temperature: 60°C for 15 min; extraction temperature: 60°C for 60 min. The fiber was exposed into a gas chromatograph-mass spectrometer (GC-MS) injector for 2 min at 230°C, to desorb the terpenes. Analyses were performed on a Thermo Scientific Trace 1,300 gas chromatograph coupled to a Thermo Scientific ISQ single quadrupole mass spectrometer (both Thermo Fisher, Waltham, Massachusetts), equipped with an electronic impact source. All samples were injected in splitless mode, maintaining the valve closed for 3 min. The carrier gas was helium, with a total flow of 1 mL/min. The separation was performed on a SUPELCOWAX 10 capillary column (Supelco, Bellefonte, PA, USA, 30 m × 0.25 mm × 0.25 μm), using the following temperature program: starting at 35°C for 8 min; increasing by 4°C/min up to 60°C; increasing by 20°C/min up to 200°C; maintaining this final temperature for 10 min. The transfer line temperature was 230°C. The signal acquisition mode was SIM (93, 121, 136 m/z for the first 20 min, from 20 to 20,49 min were 93, 105, 133, 164, 204 m/z; from 20,50 min to the end were 93, 121, 147, 204 m/z). Terpenes identification was performed by comparing their retention times with those of pure standard compounds injected under the same chromatographic conditions, and comparing ratios between selected ions intensity with those of pure compounds.
Statistical analysis was performed with SAS software (version 8.6, SAS Institute Inc., Cary, NC). A repeated mixed model was applied, in which breed (Ho, Mo, Va), period (June, July) and their interaction were included as fixed factors. The repeated factor was the period, with the individual cow as subject. Results are presented as Least Square means and standard errors of the mean (SEM). In addition, Pearson's correlation coefficients between the variables describing grazing behavior and those of milk composition were calculated in order to be able to distinguish among individual cows.
Only a few differences were observed among breeds in terms of grazing behavior (Table 3). Based on the IS, both Ho and Mo avoided mature vegetation whereas Va were indifferent to it. In June, Ho tended to express a stronger aversion to legumes than the other breeds (IS Ho = −0.78, IS Mo = −0.38, and IS Va = −0.25, p = 0.054; data not shown in table). No changes in the botanical and nutritional composition of the simulated bites was observed, except for a tendency of Mo to select herbage with a higher ADF content in June (+23 and +22 compared to that selected by Ho and Va cows, respectively, p = 0.093; data not shown in table). These tendencies observed in June among breeds were not observed in July. The daily potential intake capacity of Va was 4.2 kg of DM lower than those of Ho and Mo, and, as a consequence, their daily estimated grasses intake was lower (3.6 kg of DM). No difference among breeds were observed for legumes and forbs intake. The intake of dry material was also estimated higher for Ho compared to Mo and Va (+2.0 kg of DM). Regardless of breed, grazing behavior of all cows changed between the two experimental periods. In July, the IS for forbs and mature vegetation decreased by 0.31 and 0.62, respectively, whereas it increased by 0.17 and 0.25 for legumes and short vegetative herbage. The proportion of the different vegetation types in the selected bites did not differ between June and July, except for a tendency for increased proportion of dry vegetation (+12.5%, p = 0.059). Overall, the selected bites had a higher crude protein content (+38 g/kg DM) and a lower ADF content (−24 g/kg DM) in July compared to June. The estimated OMD tended to be higher by 2.9 g/kg DM in July than in June (p = 0.086). The proportion of legumes in the selected bites was negatively correlated to ADF and NDF contents (−0.609** and −0.612**, respectively), and positively correlated to the crude protein content (0.673**) and estimated OMD (0.591**). The proportion of forbs in the simulated bites was negatively correlated to the NDF content (−0.462*). The proportion of dry material was negatively correlated to the ADF content (−0.432*).
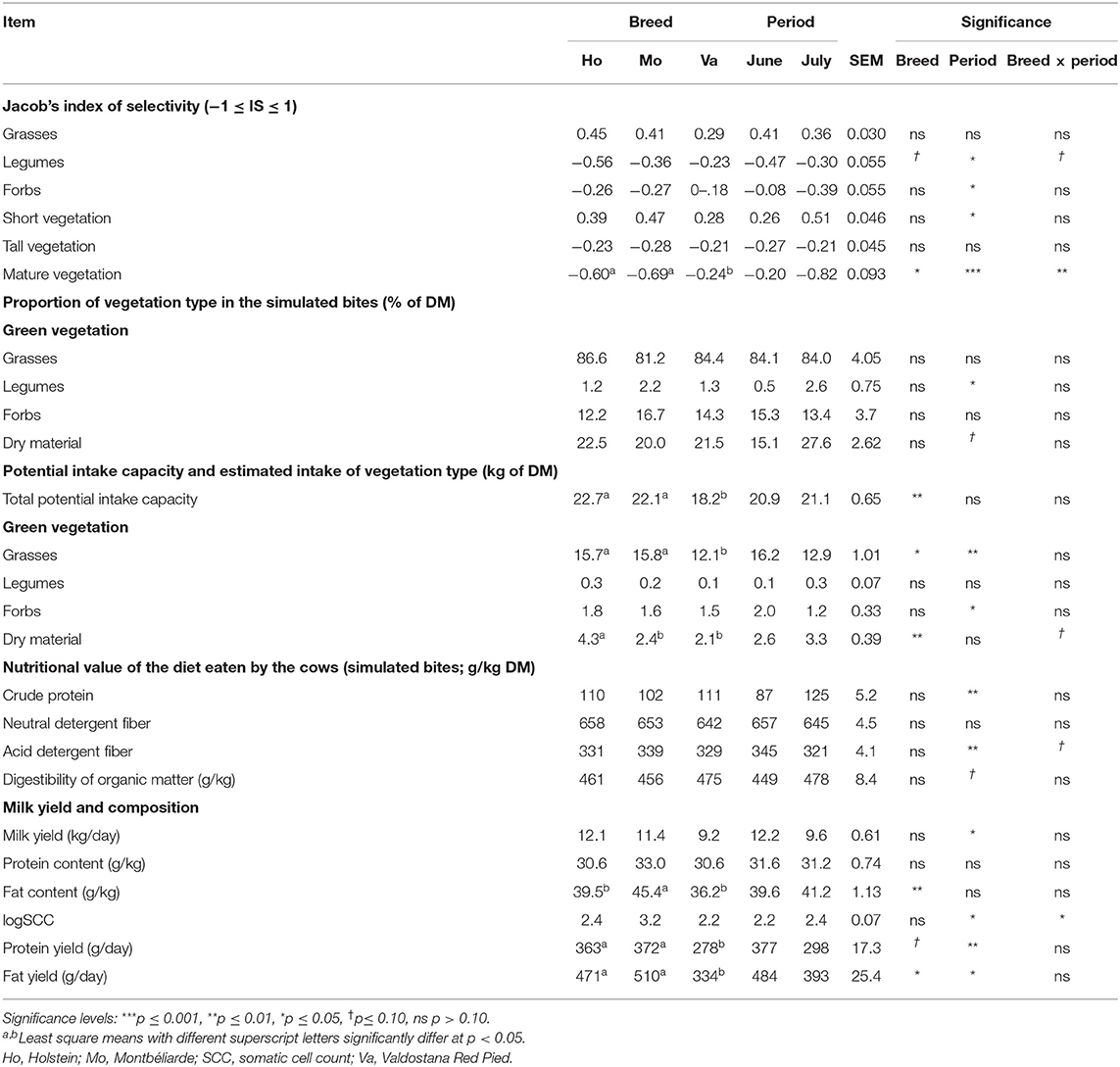
Table 3. Indices of selectivity, characterization of vegetation eaten (simulated bites) by the cows, and performance distinguished by breed and period.
Milk yield and protein content did not significantly differ among breeds. In Mo, the milk fat content was higher than in Ho and Va, by 5.9 and 9.2 g/kg, respectively. The daily milk fat yield was similar between Ho and Mo and lower by 126 g/day in average for Va. There was a similar tendency for the daily protein yield (−89 g/day in average for Va compared to both other breeds, p = 0.077). In June, the SCC of Mo's milk was lower than that of Ho and Va by 0.5 and 0.3 log units, respectively. Regardless of breed, the milk yield decreased by 2.6 kg from June to July. Concomitantly, the daily fat and protein yields decreased by 79 and 91 g/day, respectively. Milk yield and composition were not correlated to the proportions of the different botanical groups in the bites of the cows, but milk yield was positively correlated to the estimated grasses intake (0.594**), as well as milk fat and protein yields (0.620** and 0.575**, respectively). However, both yields were also positively correlated to the total potential intake capacity (0.508* and 0.575**, respectively), as well as milk fat content (0.406*). Concerning herbage composition, fat and protein yield were positively correlated to the ADF content (0.466* and 0.497*, respectively). The SCC (log unit) of the milk was negatively correlated to the ADF and NDF contents of the herbage (−0.564** and −0.457*, respectively).
The milk fat of Va tended to have a higher content of C18:2 n-6 (+0.11 g/100 g of total FA, p = 0.070) and had a higher content of C18:3 n-3 (+0.12 g/100 g of total FA) than the milk fat of both other breeds (Table 4). Milk fat of Mo tended to have the lowest content of total n-6 FA (−0.14 g/100 g of total FA in average, p = 0.097), and the n-6 to n-3 and C18:2n-6 to C18:3n-3 ratios were 10% higher in milk fat of Ho than that of the other breeds. In June, the milk fat of Ho tended to have a higher content of C16:0 (p = 0.071) than that of Mo (+0.30 g/100 g of total FA), and then Va (+1.5 g/100 g of total FA), but this was no longer the case in July (data not shown in table). Most of the changes in milk FA profile occurred between June and July. In June, the milk fat of all cows had a higher content of saturated FA (total SFA +2.9 g/100 g of total FA) than in July. More specifically, the content of C6:0, C8:0, C10:0, C12:0, C14:0, C15:0, and C16:0 was higher. On the contrary, the content of mono-unsaturated FA was higher in milk fat in July (+2.4 g/100 g of total FA), in particular that of C18:1c9 (+1.97 g/100 g of total FA), as well as the sum of all C18:1 cis isomers (+2.1 g/100 g of total FA). The content of poly-unsaturated FA in milk fat was also higher in July (+0.41 g/100 g of total FA), in particular that of n-3 FA (+0.09 g/100 g of total FA). Accordingly, the ratio n-6 to n-3 decreased in July. There was a decrease in the proportion of even-chained FA in July (−2.94 g/kg), whilst that of the odd-chained FA increased (+0.26 g/kg). The C14:1c9 to C14:0 and C18:1c9 to C16:0 ratios increased by 0.02 and 0.17 from June to July, respectively. De novo synthesized FA decreased in July, by 1.79 g/kg. Overall, milk FA proportions had little correlation with the proportions of the botanical groups in the diet of the cows. The proportion of grasses in the simulated bites was correlated only to the proportion of one FA, C18:0 (0.444*). The estimated grasses intake was also negatively correlated to C18:3n-3 (−0.482*) and n-3 FA (−0.412*) concentration in milk. In the same way, legume proportion was only correlated to the proportion of C18:3 n-6 (0.408*). Forb proportion was positively correlated to proportions of C12:0 and C14:0 (0.419* and 0.423*, respectively), and negatively correlated to proportions of C14:1t9 and C18:0 (−0.444* and −0.470*, respectively). The proportion of dry material in the bites was positively correlated to proportions of C18:1c9 (0.487*), MUFA (0.552**), sum of C18:1 cis isomers (0.490*), and to the C14:1c9 to C14:0 and C18:1c9 to C18:0 ratios (0.445* and 0.462*, respectively). It was negatively correlated to the proportions of C5:0 (−0.407*), C6:0 (−0.458*), total SFA (−0.558**) and de novo FA (−0.441*). Two correlations were also found between milk FA profile and the nutritional value of the diet as calculated from the simulated bites: the crude protein content of the herbage was positively correlated to C18:3 n-6 proportion (0.531*) and the NDF content was negatively correlated to C12:1c9 proportion (−0.461*).
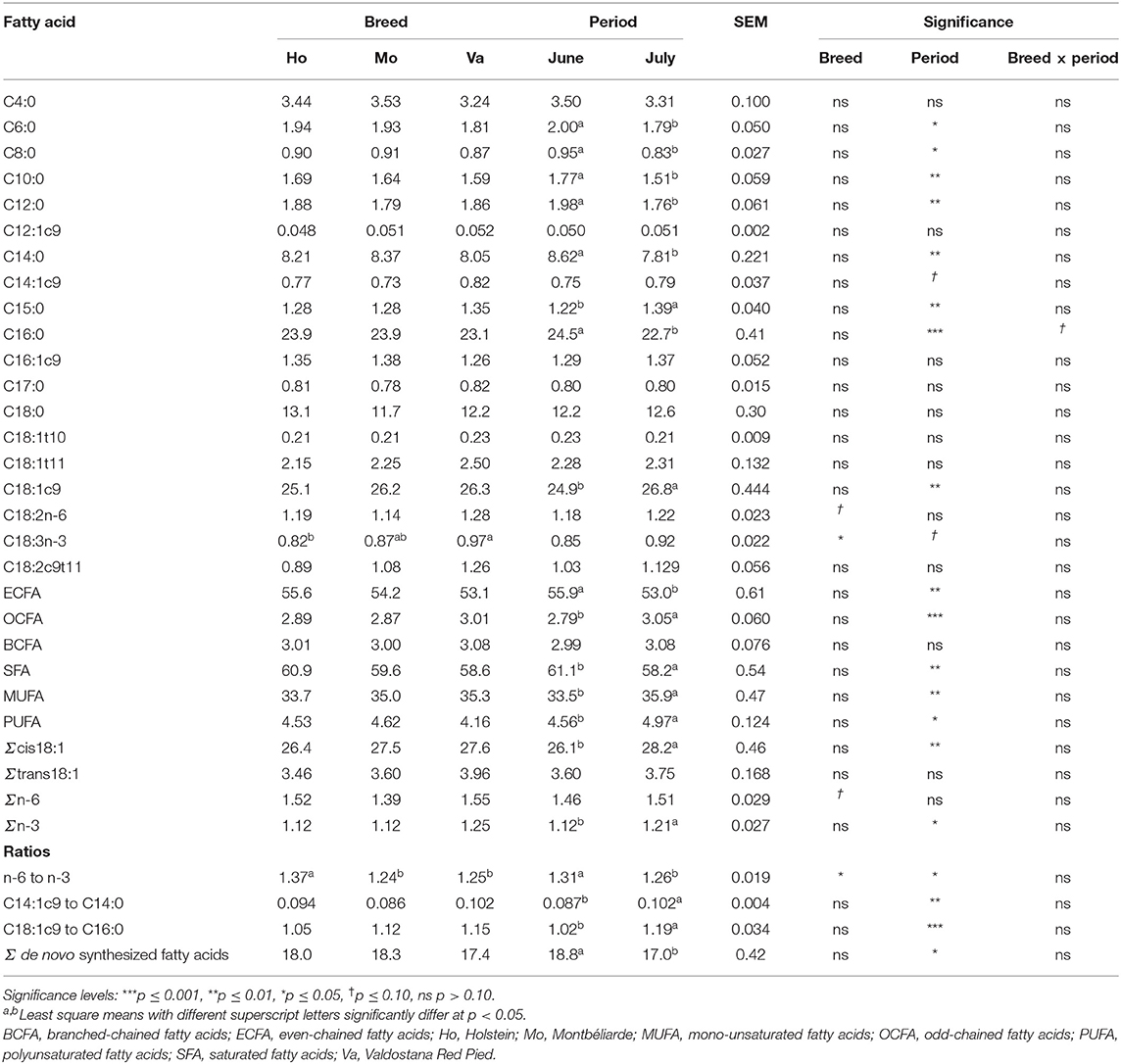
Table 4. Proportions of fatty acids (g/100 g total fatty acids) in milk fat distinguished by breed and period.
A total of 16 different terpenes were identified in the milk of the three breeds, whereof the 11 most frequently found terpenes are presented in Table 5. From these 16 terpenes, α- and β-pinene, camphene, γ-terpinene, α- and β-caryophyllene were found in all 24 samples. Sabinene was detected in 22 samples out of 24, limonene in 21, β-myrcene and terpinolene in 19, and α-phellandrene in 16. The other compounds occasionally detected were α-thujene, δ3-carene, ocimene and linalool. Regardless of breed or period, the three most abundant terpenes found in milk were limonene, β-caryophyllene and γ-terpinene, with in average 24.6, 23.8, and 10.2 %, respectively. Milk terpenes did not vary among breeds, but a few differences occurred between the two experimental periods. This was Specifically an increase in camphene (+2.98 %) and α-caryophyllene (+2.64 %) prevalence in July compared to June. Terpinolene and β-caryophyllene tended to increase in July, too (+1.50 and +9.18%, p = 0.096 and 0.088, respectively), while β-myrcene tended to decrease (−1.53%, p = 0.068). The milk terpenes were not correlated to the proportions of different botanical groups in the bites of the cows, except for α-caryophyllene that was negatively correlated with the estimated grasses intake (−0.475*). It was also negatively correlated to the ADF content of the bites (−0.454*).
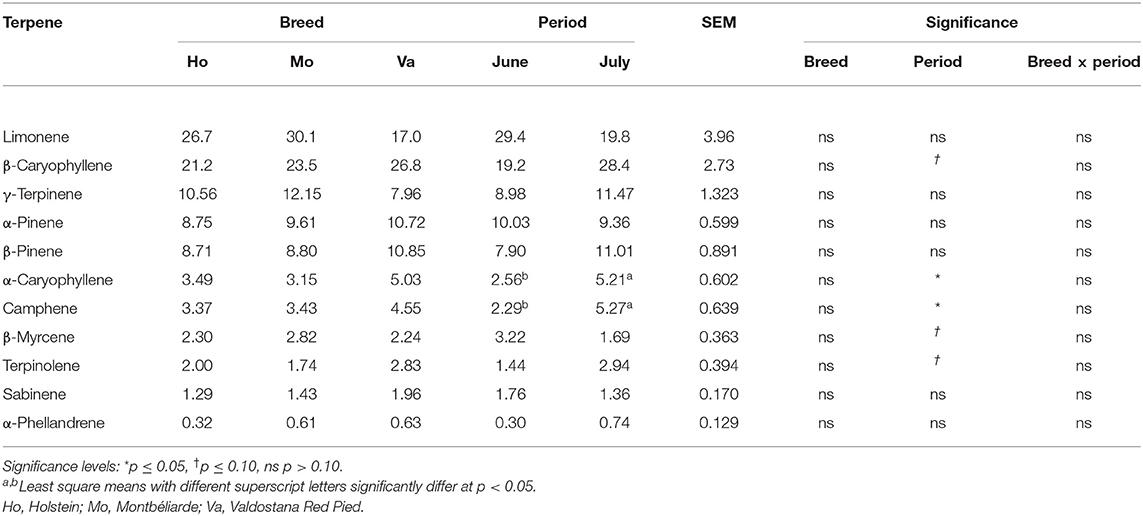
Table 5. Prevalence of the most abundant terpenes in milk (proportion of individual peak area over total peak area, %) distinguished by breed and period.
In continuous grazing conditions on a biodiverse pasture, according to the IS, cows of the least specialized breed (Va) were less selective on pasture than those of the highly specialized breed (Ho). This was particularly the case with mature herbage. Grazing behavior of Mo seems to be more similar to that of Ho, even though they were a little less selective. However, these differences in grazing selection were minimal compared to those expressed within breeds on pastures with different botanical composition or under different grazing management (21, 27), as was also observed in the present study, when forage of different vegetation growth stage was offered in June and July. Still, the breed differences in grazing selection were lower than expected and were in contrast to the results of Farruggia et al. (35), who showed that lactating cattle with high nutrient requirements grazed more selectively than dry cows, and Pauler et al. (22, 23), who found that more productive Angus × Holstein cattle grazed more selectively than Highland cattle. However, Dumont et al. (36) also did not find relevant differences in grazing behavior among traditional and specialized breeds. The few differences among cow breeds highlighted by the IS did not lead to a significant difference in composition and estimated digestibility of the simulated bites in our experiment. This indicates that, even though cows selected or avoided some species while grazing, the corresponding changes in DM quantities found in the simulated bites remained too low to make a clear difference in their proportion of the total daily diet. Concerning season and evolution of the herbage, cows avoided forbs and mature vegetation less in June than in July, probably because they already overgrazed the preferred patches with grasses during the beginning of the grazing cycle (24, 37, 38). In July, they selected the vegetative regrowth and avoided legumes less. This can be explained by the observation that the latter botanical group regrows rapidly (21) and therefore was more accessible at that time than at the end of the first grazing cycle. These changes in grazing selection and the increased small proportion of legumes are coherent with the higher crude protein content in the simulated bites, together with their lower ADF content, in July compared to June. Finally, cows seem to have adapted their grazing behavior to what was offered during the respective season, regardless of breed. Further studies would be required to strengthen our findings over a larger number of animals.
The differences among cow breeds in milk yield and gross composition were previously discussed in Koczura et al. (24). Briefly, the lower milk fat content in Va milk compared to Mo is typical of Va breed (39, 40), as this dual-purpose breed is not specifically selected for a high milk fat content. The lower fat content of Ho compared to Mo is in accordance with previous results comparing Ho and Mo reared in the same conditions (41). The lower fat and protein yields of Ho compared to Mo could be due to the lower adaptability of Ho breed to cover their high requirements without concentrate supplements, resulting in a negative energy balance, and lower fat and protein yield (24). Regarding FA composition of milk fat, several studies already investigated the link between the composition of diverse pastures and milk quality [e.g., (16, 18, 42)], but only a few allowed to link the selective behavior that ruminants exhibit on pasture with their milk composition (27, 38). These authors illustrated how the ability of an animal to feed on forbs instead of grasses has an effect on the ruminal microbial population, with PSC partially inhibiting the biohydrogenation of dietary PUFA, entirely or at certain steps. This results in milk richer in these FA, and particularly in n-3 FA (43). However, this effect is especially visible when comparing selection on highly biodiverse pastures with grasslands with very different proportions of grasses and forbs (18). In the present experiment, all cows grazed on the same biodiverse pasture. The results, therefore, suggest that, even though some preferences were expressed, the differences in the material eaten among breeds remained too low to affect the milk FA profile. This is consistent with the similar FA profile of the milk found for the three breeds, especially in terms of MUFA, PUFA, and CLA proportions of the milk fat. These groups of FA were actually correlated to the herbage evolution over the season and, more specifically, to the proportion of dry material in the simulated bites, regardless of breed. The cause for that is the advanced herbage phenological stage in June compared to July measurement period, which is known to increase MUFA and decrease PUFA and CLA proportions of the milk fat (18, 42). Only one particular FA, the major n-3 FA C18:3 n-3, was found to be always higher in Va milk fat compared to Ho milk fat, without any correlation to diet selection or period. This FA is a substrate for ruminal biohydrogenation, leading to the production of C18:1t11, ultimately being saturated to C18:0 (43). Although not significant, C18:1t11 was also numerically higher in Va than Ho milk fat. This suggests that C18:3 n-3 is less biohydrogenated in the rumen of Va than Ho cows, and this could probably be explained by breed differences in the rumen microbial population and metabolism rather than grazing behavior. This aspect should be further investigated, especially on a larger number of animals.
To our knowledge, this is the first study directly linking diet selection of dairy cows and the occurrence of terpenes in their milk. Terpenes are absorbed directly from the diet (9), and most of them are transferred directly into milk and are subject to no or minor changes (44, 45). Some authors demonstrated that terpenes can also be partially biohydrogenated and isomerized by the rumen microbial population, leading to additional terpenes in milk (46). In the present experiment, the investigated pasture was rich in forbs, such as plants from the Apiaceae family and Thymus serpyllum, both containing pinenes and caryophyllenes (47, 48). The presence of this kind of compounds in the milk of the experimental cows suggests that pinenes and caryophyllenes indeed originated from the diet. Several previous studies already identified β-caryophyllene in the milk of grazing cows (10, 11). However, unlike our hypothesis, it seems difficult to directly relate terpene prevalence in milk and grazing behavior of the cows. Accordingly, in the present study no significant correlations were found between terpene prevalence and the proportion of the different botanical groups in the simulated bites. This could be partially due to the limited number of animals in our experiment. However, if a biological link was underlying between grazing selection and milk composition, significant correlations would have been expected even with few animals, probably with a poor correlation coefficient that would have been improved by increasing the number of animals, but it was not the case. This suggests that the selection of a single species or family could be responsible for the transfer of such molecules (5), rather than the overall proportion of forbs or mature vegetation in the sward. Besides, Lejonklev et al. (49) showed that terpenes from essential oils can be transferred to milk by both ingestion and inhalation. Cows may therefore also have taken in terpenes while breathing, which would explain why the latter are not correlated to the simulated bites. The few differences that occurred in milk terpene prevalence were related to the grazing period: the vegetative stage of the pasture evolved after the first grazing cycle and most probably led to a different terpene composition of the herbage (5, 50). Indeed, a parallel increase was observed by Tornambé et al. (10) between milk terpenes concentration and the variation in the phenological stage of the herbage. Even though concentrations of terpenes found in milk are low, it would be interesting to manufacture cheeses with the milk of the three breeds and investigate the link between terpene profile, microbial development and potential further influences on cheese sensory properties. Such an effect could be expected because some terpenes have been found to have antimicrobial effects. Rivas da Silva et al. (47) for instance demonstrated that positive enantiomers of α- and β-pinene used in synergy (250 μg/mL) can have a bactericidal effect against methicillin-resistant Staphylococcus aureus.
The overall small differences among the autochthonous, more robust Va, the intermediate Mo and the Ho highly specialized for milk production may suggest that in low-input mountain grazing systems, the individual animal's adaptability in the short term could actually be more important than its breed. It has to be mentioned, though, that all cows were in their late lactation and therefore even the Ho were not in a situation of high nutrient requirements. Breed differences could be clearer in other stages of lactation. Breed also seems to have less influence on milk quality than herbage composition. Other criteria than grazing behavior should be investigated in order to assess the role of autochthonous breeds in the multi-performance of future low-input grazing systems. Indeed, individual adaptation in the short term may result in different long-term breed responses, i.e., in reproduction or productive lifespan. Further investigation on a larger number of animals would reinforce our findings.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
No ethical approval was required because no invasive samples have been taken on the animals and all procedures carried out followed the Certificate of Authorization to Experiment on Living Animals No. D 15-114-01 delivered by the French government to Marcenat INRAE experimental facility.
MKo, MC, BM, MKr, and JB contributed to the conception and design of the study. MKo, MB, and GT organized the experiment. MKo supervised the experiment. MKo, MB, and BM participated in behavioral observation and milk sampling. MC performed the fatty acids analysis. MMu and MMa performed the terpene analysis. MKo and MC processed the data and equally contributed to the writing of the manuscript. MC performed the statistical analysis. MKr and JB supervised MKo in the frame of her doctoral thesis. All the authors contributed to manuscript revision, and all read and approved the submitted version.
This research received funding from the French Government IDEX-ISITE initiative 16-IDEX-0001 (CAP 20-25).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank the staff of the INRAE farm at Marcenat (UE Herbipôle) with special thanks to Carole Cirie; Agnès Cornu for advice and help with the extraction of milk terpenes; and Cyril Labonne, Jean-Baptiste Auger, Victoria Verdel, Précilia Bouchy, Laurent Lanore and Isabelle Constant for their participation in the collection of behavioral observations data and treatment of the samples.
1. Leiber F, Jouven M, Martin B, Priolo A, Coppa M, Prache S, et al. Potentials and challenges for future sustainable grassland utilisation in animal production. Options Méditerranéennes. Série A: Séminaires Méditerranéens, A. (2014) 109:33–48. Available online at: http://om.ciheam.org/om/pdf/a109/00007678.pdf
2. Sturaro E, Marchiori E, Cocca G, Penasa M, Ramanzin M, Bittante G. Dairy systems in mountainous areas: farm animal biodiversity, milk production and destination, and land use. Livest Sci. (2013) 158:157–68. doi: 10.1016/j.livsci.2013.09.011
3. Friggens NC, Blanc F, Berry DP, Puillet L. Review: deciphering animal robustness. A synthesis to facilitate its use in livestock breeding and management. Animal. (2017) 11:2237–51. doi: 10.1017/S175173111700088X
4. Coppa M, Chassaing C, Sibra C, Cornu A, Harstad OM, Verbič J, et al. Forage system is the key driver of mountain milk specificity. J Dairy Sci. (2019) 102:10483–99. doi: 10.3168/jds.2019-16726
5. Cornu A, Carnat AP, Martin B, Coulon JB, Lamaison JL, Berdague JL. Solid-phase microextraction of volatile components from natural grassland plants. J Agric Food Chem. (2001) 49:203–9. doi: 10.1021/jf0008341
6. Falchero L, Coppa M, Fossi A, Lombardi G, Ramella D, Tava A. Essential oil composition of lady's mantle (Alchemilla xanthochlora Rothm.) growing wild in alpine pastures. Nat Prod Res. (2009) 23:1367–72. doi: 10.1080/14786410802361438
7. Reynaud A, Fraisse D, Cornu A, Farruggia A, Pujos-Guillot E, Besle JM, et al. Variation in content and composition of phenolic compounds in permanent pastures according to botanical variation. J Agric Food Chem. (2010) 58:5485–94. doi: 10.1021/jf1000293
8. Fraisse D, Carnat A, Viala D, Pradel P, Besle JM, Coulon JB, et al. Polyphenolic composition of a permanent pasture: variations related to the period of harvesting. J Sci Food Agric. (2007) 87:2427–35. doi: 10.1002/jsfa.2918
9. Serrano E, Cornu A, Kondjoyan N, Figueredo G, Agabriel J, Micol D. Terpene accumulation in muscle and fatty tissues of calves supplemented with essential oils. J Anim Feed Sci. (2007) 16:168–79. doi: 10.22358/jafs/66736/2007
10. Tornambé G, Cornu A, Pradel P, Kondjoyan N, Carnat AP, Petit M, et al. Changes in terpene content in milk from pasture-fed cows. J Dairy Sci. (2006) 89:2309–19. doi: 10.3168/jds.S0022-0302(06)72302-5
11. Agabriel C, Cornu A, Journal C, Sibra C, Grolier P, Martin B. Tanker milk variability according to farm feeding practices: vitamins A and E, carotenoids, color, and terpenoids. J Dairy Sci. (2007) 90:4884–96. doi: 10.3168/jds.2007-0171
12. De Noni I, Battelli G. Terpenes and fatty acid profiles of milk fat and Bitto cheese as affected by transhumance of cows on different mountain pastures. Food Chem. (2008) 109:299–309. doi: 10.1016/j.foodchem.2007.12.033
13. Khiaosa-ard R, Soliva CR, Kreuzer M, Leiber F. Influence of alpine forage either employed as donor cow's feed or as incubation substrate on in vitro ruminal fatty acid biohydrogenation. Livest Sci. (2011) 140:80–7. doi: 10.1016/j.livsci.2011.02.012
14. Cabiddu A, Molle G, Decandia M, Spada S, Fiori M, Piredda G, et al. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep: part 2: effects on milk fatty acid profile. Livest Sci. (2009) 123:230–40. doi: 10.1016/j.livsci.2008.11.019
15. Cabiddu A, Salis L, Tweed JS, Molle G, Decandia M, Lee MRF. The influence of plant polyphenols on lipolysis and biohydrogenation in dried forages at different phenological stages: in vitro study. J Sci Food Agric. (2010) 90:829–35. doi: 10.1002/jsfa.3892
16. Leiber F, Kreuzer M, Nigg D, Wettstein HR, Scheeder MRL. A study on the causes for the elevated n-3 fatty acids in cows' milk of alpine origin. Lipids. (2005) 40:191–202. doi: 10.1007/s11745-005-1375-3
17. Wehrmueller K, Schmid A, Walther B. Health benefits of omega-3-fatty acids and the contribution of products from mountain pastures to the supply. Ernährungs-Umschau. (2008) 55:655–61. Available online at: https://www.ernaehrungs-umschau.de/print-artikel/13-11-2008-health-benefits-of-omega-3-fatty-acids-and-the-contribution-of-products-from-mountain-pastures-to-the-supply/
18. Renna M, Ferlay A, Lussiana C, Bany D, Graulet B, Wyss U, et al. Relative hierarchy of farming practices affecting fatty acid composition of permanent grasslands and of the derived bulk milk. Anim Feed Sci Technol. (2020) 267:114561. doi: 10.1016/j.anifeedsci.2020.114561
19. Giaccone D, Revello-Chion A, Galassi L, Bianchi P, Battelli G, Coppa M, et al. Effect of milk thermisation and farming system on cheese sensory profile and fatty acid composition. Int Dairy J. (2016) 59:10–9. doi: 10.1016/j.idairyj,.2016.02.047
20. Prache S, Gordon IJ, Rook AJ. Foraging behaviour and diet selection in domestic herbivores. Annales De Zootechnie. (1998) 47:335–45. doi: 10.1051/animres:19980502
21. Coppa M, Farruggia A, Pradel P, Lombardi G, Martin B. An improved grazed class method to estimate species selection and dry matter intake by cows at pasture. Ital J Anim Sci. (2011) 10:58–65. doi: 10.4081/ijas.2011.e13
22. Pauler CM, Isselstein J, Suter M, Berard J, Braunbeck T, Schneider MK. Choosy grazers: influence of plant traits on forage selection by three cattle breeds. Funct Ecol. (2020) 34:980–92. doi: 10.1111/1365-2435.13542
23. Pauler CM, Isselstein J, Berard J, Braunbeck T, Schneider MK. Grazing allometry: anatomy, movement, and foraging behavior of three cattle breeds of different productivity. Front Vet Sci. (2020) 7:494. doi: 10.3389/fvets.2020.00494
24. Koczura M, Martin B, Bouchon M, Turille G, Berard J, Farruggia A, et al. Grazing behaviour of dairy cows on biodiverse mountain pastures is more influenced by slope than cow breed. Animal. (2019) 13:2594–602. doi: 10.1017/S175173111900079X
25. Daget P, Poissonet J. Une méthode d'analyse phytosociologique des prairies. Ann Agron. (1971) 22:5–41.
27. Coppa M, Farruggia A, Ravaglia P, Pomiès D, Borreani G, Le Morvan A, et al. Frequent moving of grazing dairy cows to new paddocks increases the variability of milk fatty acid composition. Animal. (2015) 9:604–13. doi: 10.1017/S1751731114003000
28. Sweeney RA, Rexroad PR. Comparison of LECO FP-228 “nitrogen determinator” with AOAC copper catalyst Kjeldahl method for crude protein. J Assoc Off Anal Chem. (1987) 70:1028–30. doi: 10.1093/jaoac/70.6.1028
29. Van Soest PJ, Robertson JB, Lewis BA. Methods of dietary fiber, neutral detergent fiber and non-polysaccharides in relation to animal nutrition. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds.S0022-0302(91)78551-2
30. Aufrère J, Michalet-Doreau B. In vivo digestibility and prediction of digestibility of some by-products. In: Boucqué CV, Fimes LO, Cottyn GB, editors. Feeding Value of By-Products and their Use by Beef Cattle—EEC Seminar EUR 8918 EN. Brussels: Commission of the European Union (1983). p. 25–35.
31. AOAC. Official Methods of Analysis, 16th ed. Gaithersburg, MD: Association of Official Analytical Chemists (1997).
32. International Dairy Federation. Whole Milk—Determination of Milkfat, Protein and Lactose Content—Guidance on the Operation of Mid-infrared Instruments. IDF 141C:2000. Brussels, Belgium: International Dairy Federation (2000).
33. Ferlay A, Martin B, Lerch S, Gobert M, Pradel P, Chilliard Y. Effects of supplementation of maize silage diets with extruded linseed, vitamin E and plant extracts rich in polyphenols, and morning v. evening milking on milk fatty acid profiles in Holstein and Montbéliarde cows. Animal. (2010) 4:627–40. doi: 10.1017/S1751731109991224
34. Loor JJ, Ferlay A, Ollier A, Doreau M, Chilliard Y. Relationship among trans conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J Dairy Sci. (2005) 88:726–40. doi: 10.3168/jds.S0022-0302(05)72736-3
35. Farruggia A, Dumont B, D'hour P, Egal D, Petit M. Diet selection of dry and lactating beef cows grazing extensive pastures in late autumn. Grass Forage Sci. (2006) 61:347–53. doi: 10.1111/j.1365-2494.2006.00541.x
36. Dumont B, Rook AJ, Coran Ch, Röver KU. Effects of livestock breed and grazing intensity on biodiversity and production in grazing systems. 2. Diet selection. Grass Forage Sci. (2007) 62:159–71. doi: 10.1111/j.1365-2494.2007.00572.x
37. Adler PB, Raff DA, Lauenroth WK. The effect of grazing on the spatial heterogeneity of vegetation. Oecologia. (2001) 128:465–79. doi: 10.1007/s004420100737
38. Farruggia A, Pomiès D, Coppa M, Ferlay A, Verdier-Metz I, Le Morvan A, et al. Animal performances, pasture biodiversity and dairy product quality: how it works in contrasted mountain grazing systems. Agric Ecosyst Environ. (2014) 185:231–44. doi: 10.1016/j.agee.2014.01.001
39. Coppa M, Gorlier A, Lonati M, Martin B, Russo EM, Lombardi G. The management of the transition from hay- to pasture-based diets affects milk fatty acid kinetics. Dairy Sci. Technol. (2012) 92:279–95. doi: 10.1007/s13594-012-0065-6
40. Manzocchi E, Koczura M, Coppa M, Turille G, Kreuzer M, Berard J. Grazing on upland pastures part-time instead of full-time affects the feeding behavior of dairy cows and has consequences on milk fatty acid profiles. Animals. (2019) 9:908. doi: 10.3390/ani9110908
41. Balandraud N, Mosnier C, Delaby L, Dubief F, Goron JF, Martin B, et al. Holstein ou Montbéliarde: des différences phénotypiques aux conséquences économiques à l'échelle de l'exploitation. INRA Prod Anim. (2018) 31:337–52. doi: 10.20870/productions-animales.2018.31.4.2394
42. Coppa M, Ferlay A, Borreani G, Revello-Chion A, Tabacco E, Tornambé G, et al. Effect of phenological stage and proportion of fresh herbage in cow diets on milk fatty acid composition. Anim Feed Sci Technol. (2015) 208:66–78. doi: 10.1016/j.anifeedsci.2015.07.006
43. Chilliard Y, Glasser F, Ferlay A, Bernard L, Rouel J, Doreau M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur J Lipid Sci Technol. (2007) 109:828–55. doi: 10.1002/ejlt.200700080
44. Viallon C, Martin B, Verdier-Metz I, Pradel P, Garel JP, Coulon JB, et al. Transfer of monoterpenes and sesquiterpenes from forages into milk fat. Lait. (2000) 80:12–6. doi: 10.1051/lait:2000150
45. Bugaud C, Buchin S, Coulon JB, Hauwuy A, Dupont D. Influence of the nature of alpine pasture on plasmin activity, fatty acid and volatile compound composition of milk. Lait. (2001) 81:401–14. doi: 10.1051/lait:2001140
46. Schlichterle-Cerny H, Imhof MI, Fernandez Garcia E, Bosset JO. From pasture to cheese: changes in terpenes composition. In: Cadwallader KR, Drake M, McGorrin R, editors. Flavor of Dairy Products. Washington, DC: American Chemical Society (2007). p. 123–33. doi: 10.1021/bk-2007-0971.ch007
47. Rivas da Silva AC, Monteiro Lopes P, Barros de Azevedo MM, Machado Costa DC, Sales Alviano C, Sales Alviano D. Biological activities of α-pinene and β-pinene enantiomers. Molecules. (2012) 17:6305–16. doi: 10.3390/molecules17066305
48. Cutillas AB, Carrasco A, Martinez-Gutierrez R, Tomas V, Tudela J. Thyme essential oils from Spain: aromatic profile ascertained by GC-MS, and their antioxidant, anti-lipoxygenase and antimicrobial activities. J Food Drug Anal. (2018) 26:529–44. doi: 10.1016/j.jfda.2017.05.004
49. Lejonklev J, Løkke MM, Larsen MK, Mortensen G, Petersen MA, Weisbjerg MR. Transfer of terpenes from essential oils into cow milk. J Dairy Sci. (2013) 96:4235–41. doi: 10.3168/jds.2012-6502
Keywords: grazing behavior, simulated bites, late-lactating, Holstein, Montbéliarde, Valdostana Red Pied, α-linolenic acid (C18:3 n-3)
Citation: Koczura M, Martin B, Musci M, Massimo MD, Bouchon M, Turille G, Kreuzer M, Berard J and Coppa M (2021) Little Difference in Milk Fatty Acid and Terpene Composition Among Three Contrasting Dairy Breeds When Grazing a Biodiverse Mountain Pasture. Front. Vet. Sci. 7:612504. doi: 10.3389/fvets.2020.612504
Received: 30 September 2020; Accepted: 24 December 2020;
Published: 22 January 2021.
Edited by:
Tamara Alejandra Tadich, University of Chile, ChileReviewed by:
Einar Vargas-Bello-Pérez, University of Copenhagen, DenmarkCopyright © 2021 Koczura, Martin, Musci, Massimo, Bouchon, Turille, Kreuzer, Berard and Coppa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madeline Koczura, bWFkZWxpbmUua29jenVyYUBpbnJhZS5mcg==
†Present address: Madeline Koczura, Université Clermont Auvergne, INRAE, VetAgro Sup, UMR0874 Ecosystème Prairial, Clermont-Ferrand, France
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.