 Mercedes Fernández-Escobar1
Mercedes Fernández-Escobar1 Rafael Calero-Bernal1*
Rafael Calero-Bernal1* Javier Regidor-Cerrillo2Raquel Vallejo3
Javier Regidor-Cerrillo2Raquel Vallejo3 Julio Benavides3Esther Collantes-Fernández1
Julio Benavides3Esther Collantes-Fernández1 Luis Miguel Ortega-Mora1*
Luis Miguel Ortega-Mora1*- 1Salud Veterinaria y Zoonosis (SALUVET) Group, Animal Health Department, Faculty of Veterinary Sciences, Complutense University of Madrid, Madrid, Spain
- 2SALUVET-innova S.L., Faculty of Veterinary Sciences, Complutense University of Madrid, Madrid, Spain
- 3Mountain Livestock Institute, Consejo Superior de Investigaciones Científicas-Universidad de León (CSIC-ULE), León, Spain
The present study aimed to isolate and perform molecular and phenotypic characterization of Toxoplasma gondii strains infecting Iberian pigs bred under semi-free conditions and destined for human consumption. Blood and heart tissue samples from 361 fattening pigs from 10 various herds selected in the main areas of Iberian pig production were collected at a slaughterhouse; the sera were tested for anti-T. gondii antibodies using a commercial indirect ELISA kit, and a mouse bioassay was carried out using heart muscle of seropositive individual representatives from each geographical location. Seventy-nine (21.9%) of the 361 animals tested positive for anti-T. gondii antibodies according to the serology test. Fifteen samples of myocardial tissue were subjected to bioassay and 5 isolates (TgPigSp1 to TgPigSp5) were obtained. The isolates were characterized by using 11 PCR-RFLP genetic markers; three isolates had a ToxoDB #3 genotype (3/5) and two isolates had a ToxoDB #2 genotype (2/5). The TgPigSp1 and TgPigSp4 isolates were selected for virulence in mice characterization as instances of each different RFLP-genotype found. The TgPigSp1 isolate (#2 genotype) was virulent in mice with notable cumulative mortality (87.5%) and morbidity rates (100%); the TgPigSp4 (#3) was nonvirulent and triggered mild clinical signs in 42.1% of seropositive mice. Infection dynamics and organ distribution of both isolates were analyzed; the data revealed significant differences, including substantially higher parasite load in the lung during the acute phase of infection, in mice infected with TgPigSp1 than in the case of TgPigSp4 (median parasite load 7.6 vs. 0 zoites/mg, respectively; p < 0.05). Furthermore, degrees of severity of detected histopathological lesions appeared to be related to higher parasite burdens. Taking into account the unexpectedly high mortality rate and parasite load associated with the clonal genotype III, which is traditionally considered nonvirulent in mice, the need for further investigation and characterization of the T. gondii strains circulating in any host in Europe is emphasized.
Introduction
The eukaryotic parasite Toxoplasma gondii (Apicomplexa) can infect virtually all warm-blooded animals and constitutes a specific risk for food safety in the European Union (1–3); additionally, it is considered the second causal agent of foodborne illness in the USA (4). Generally, human infections are mainly acquired after ingestion of raw or undercooked meat containing viable T. gondii tissue cysts (5). Infections are dramatically associated with reproductive failure in pregnant women, neurological signs in immunocompromised patients, and ocular disease in otherwise healthy humans. Thus, control of T. gondii presence in meat destined for consumption is of major interest (2, 6). After chicken, pork meat is the most frequently consumed protein source in western countries, and Spain is the top producer of pork within the EU (7).
Black Iberian pigs (Sus scrofa) constitute a traditional and well-adapted pig breed whose production is linked to highly valuable meat products, especially cured ham and sausages. These animals are usually reared in extensive systems in southwestern areas of the Iberian Peninsula (covering Portugal and Spain), within a favorable ecosystem, called Dehesa, composed mostly of acorn Mediterranean forest with a high natural biodiversity, which is ideal for swine breeding (8) in sympatry with ruminant livestock and a number of other wild animals, such as Cervidae and wild boar.
Natural infections in pigs are usually asymptomatic. In Iberian pigs raised in Spain, anti-T. gondii antibodies have been detected with a frequency from 9.5 to 58.2% (9, 10); extensive management systems and facilities with outdoor access are associated with higher seroprevalence (11, 12); thus, parasite isolation and characterization of the circulating strains are of major interest. Currently, only a few studies reported the genotypes and virulence degrees of T. gondii strains infecting domestic pigs in Europe; thus, the aim of the present study was to isolate and characterize T. gondii strains in domestic Iberian pigs bred in the semifree systems in the traditional raising area of Dehesa located in the southwestern part of the Iberian Peninsula.
Materials and Methods
Ethical Statement
Animal procedures for the T. gondii isolation by bioassay in mice and evaluation of virulence degree (PROEX 274/16) were approved by the Animal Welfare Committee of the Community of Madrid, Spain, following proceedings described in Spanish and EU regulations (Law 32/2007, R.D. 53/2013, and Council Directive 2010/63/EU). All animals used in this study were handled in strict accordance with good clinical practices, and all efforts were made to minimize the suffering. As a humane endpoint, mice with a severe loss of body condition or nervous clinical signs were sacrificed to limit unnecessary suffering.
Mice
Seven-week-old female Swiss/CD1 mice were obtained from a commercial supplier (Janviers Labs, Le Genest-Saint-Isle, France). The animals were free from common viral, parasite, and bacterial pathogens according to the results of the routine screening analyses performed by the manufacturer. Mice were housed with ad libitum access to food and water in a controlled environment with 12-h light and 12-h dark cycles, and the experimental procedures were performed at 8 weeks of age.
Sample Collection and Serological Diagnosis for Tissue Selection
A total of 361 paired blood and myocardial tissue samples were collected from December, 2017 to June, 2018 from fattening Iberian pigs slaughtered for human consumption at an authorized slaughterhouse in Salamanca Province (western Spain) (Table 1). The blood samples were collected using BD PLUS serum tubes (Vacutainer; BD, Franklin Lakes, USA) at the bleeding step after animal stunning (Council Regulation (EC) No 1099/2009), and the apical part of the heart was sampled during the evisceration process; the samples were stored individually at 4°C until analysis. The serum was obtained after blood centrifugation and stored frozen (−20°C) until serological testing.
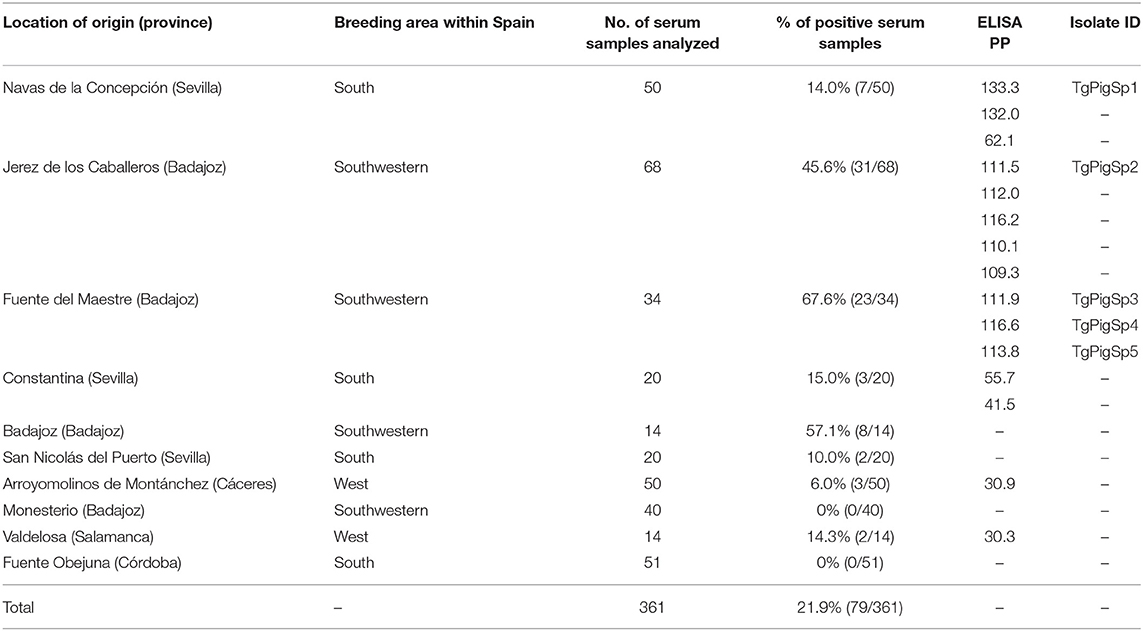
Table 1. Summarized data on geographical area of origin, number of samples collected, ELISA results and isolate obtaining from Iberian fattening pigs.
Toxoplasma gondii-specific IgG antibody levels in swine serum samples were measured using a commercial ELISA kit (PrioCHECK® Toxoplasma Ab SR, Prionics Schlieren-Zurich, Switzerland) (cut-off at 20 for percentage of positivity -ELISA PP-). Only myocardial tissues associated with the highest ELISA PP values and representative of the widespread locations (farms of origin) (Table 1) were selected for isolation by mouse bioassay (n = 15).
Bioassay in Mice and in vitro Cultivation
Portions of the heart muscle (50 g/each) from selected seropositive animals were subjected to acid-pepsin artificial digestion (13) prior to subcutaneous inoculation into 3 female Swiss/CD1 mice (Janvier-Labs, Le Genest-Saint-Isle, France). Mice were observed daily, and clinical signs were scored. Procedures of confirmation of mouse infection and in vitro isolation in cell culture were carried out as described in reference (14).
Genetic Characterization of T. gondii
Genomic DNA was extracted from the cell culture-derived T. gondii tachyzoites of all five isolates obtained using the Maxwell® 16 mouse tail DNA purification kit (Promega, Alcobendas, Spain). Genotyping was performed by the PCR-restriction fragment length polymorphisms (RFLP) method using 11 markers as described by earlier (15, 16). Clonal type reference strains of T. gondii were also included in the genotyping (TgRH, type I; TgMe49, type II; and TgNED type III). RFLP-genotype numbers were assigned according to the ToxoDB database (https://toxodb.org/toxo/).
Assays of Virulence in Mice
Two in vivo experiments were set for the evaluation of cumulative morbidity and mortality rates at 42 dpi (days post-inoculation) (assay A), and the tropism and burden reached by the isolates in mouse tissues during the acute and chronic stages of the infection (assay B).
Parasites and Inocula Preparation
The isolates TgPigSp1 and TgPigSp4 were selected as the instances of each RFLP-genotype detected in the study. Tachyzoites used for in vivo assays were harvested at low passages (p7 and p8, respectively) from cultures of Vero cells, when the majority of the parasites were still intracellular, and purified by filtration through a 3-μm polycarbonate filter (IpPORE®, IT4IP, Louvain-la-Neuve, Belgium) as described previously (17). The quantity and viability of tachyzoites were determined by Trypan blue exclusion followed by direct counting in a Neubauer chamber. Serial dilutions in PBS were performed to obtain the doses from 105 to 1 tachyzoite(s) of each isolate per 200 μl.
Assay A
Each dose was intraperitoneally (IP) inoculated into five 8-week-old female Swiss/CD1 mice. Five control mice were inoculated with 200 μl of PBS. Mice were monitored twice daily for 6 weeks and clinical signs were recorded. Cumulative morbidity rate was evaluated establishing a clinical sign scoring adapted from reference (18). Cumulative mortality rate was calculated based on the ratio of casualties to the total number of infected mice (17). Serum samples from mice, which were humanely euthanized, presented sudden death, or reached the end of the experiment at 6 weeks post-inoculation, were collected and stored at −20°C until anti-T. gondii antibodies detection by IFAT to confirm the infection. Alternative procedures to evidence animal infection were carried out as described in reference (14).
Assay B
An additional group of 10 mice per isolate was IP-inoculated with 103 tachyzoites; five animals were sacrificed at 7 dpi and the remaining five mice were sacrificed at 30 dpi to mimic the acute and chronic phases of the infection, respectively. Selected organs were collected during necropsies for T. gondii DNA detection and quantification. Briefly, mice were bled and the right cerebral hemisphere, the right eye, the right lung, half of the heart, a piece of a liver lobe, and the right kidney from each mouse were transferred immediately following euthanasia to clean 1.5 mL tubes and stored at −80°C until DNA extraction. Samples from the brain, lung, heart, liver, kidney, quadriceps femoris muscle, and tongue were fixed in 10% buffered formalin and processed for conventional histological examination. After staining with haematoxylin/eosin, lesions in the samples were subjectively categorized from 0 (no lesion) to 3 (the most severe grade within observed lesions). Serum samples were also collected and stored at −20°C until analysis.
Indirect Fluorescent Antibody Test (IFAT)
Detection of anti-T. gondii IgG antibodies in the sera was carried out by indirect fluorescent antibody test (IFAT) (19), using an anti-mouse IgG conjugated to FITC (Sigma-Aldrich) diluted 1:64 in Evans blue (Sigma-Aldrich); the cut-off was set at 1:25. Serum samples from previously known positive and negative animals were included as the controls. Tachyzoites of the Me49 strain were used as the coating antigen.
DNA Extraction, Detection, and Quantification
For parasites quantification, genomic DNA was extracted from 50 to 80 mg of selected tissues using a Maxwell® 16 mouse tail DNA purification kit (Promega). DNA samples were adjusted to 20 ng/μl. Screening for the T. gondii DNA presence was carried out by single-tube nested PCR (nPCR) amplification of the specific ITS-1 region as described previously (20). In samples with confirmed T. gondii presence according to nPCR, parasites were quantified by qPCR using primer pairs for the 529-bp repeat element of T. gondii (21). Reactions were performed in a final volume of 25 μl using GoTaq® qPCR master mix (Promega), 10 pmol of each primer and 100 ng of DNA in an Applied Biosystems 7500 FAST real-time PCR system (Applied Biosystems, Foster City, CA, USA). Amplification was performed according to a standard protocol (10 min at 95°C, 40 cycles at 95°C for 15 s, and 60°C for 1 min). The number of T. gondii tachyzoites was calculated by interpolating the average Ct values on a standard curve equivalent to 10−1 to 105 tachyzoites generated by 10-fold serial dilutions of parasite DNA in a solution of mouse genomic DNA at 20 ng/μl. Parasite load was expressed as parasite number/mg of mouse tissue. Standard curves for T. gondii had a consistent average slope close to −3.3 with a R2 > 0.98.
In addition, due to the unexpected mortality rates observed, we conducted nested PCR for CS3 locus amplification (16) over Toxoplasma-positive DNA samples from lung tissues of mice infected by TgPigSp1 (n = 3) and TgPigSp4 (n = 4) isolates (sacrificed at 7 or 30 dpi, respectively) for inocula genotype confirmation. PCR products were subjected to Sanger sequencing and obtained sequences were analyzed as described previously (14). CS3 sequences alignment to previously obtained TgPigSp1 (MW132600), TgPigSp4 (MW132601) and other reference strains (TgRH, MW151245; TgMe49, MW151246; TgNED, MW151247) sequences was carried out using the Clustal Omega software (https://www.ebi.ac.uk/Tools/msa/clustalo/).
Statistical Analyses
Variations in parasite burden in the target tissues between the inoculated mice groups were analyzed by pairwise comparisons using the Mann-Whitney U test. Lethal outcome in mice was analyzed by the Kaplan–Meier survival method, and the Mantel-Cox log-rank test was used to compare the resulting survival curves. The significance for these analyses was established at p < 0.05. GraphPad Prism 6 v.6.01 (San Diego, CA, USA) software was used to perform all statistical analyses and graphical illustrations.
Results
Serological Screening in Iberian Pigs and Parasite Isolation
Toxoplasma gondii-specific IgG antibodies were detected in 21.9% (79/361) of serum samples collected from pigs raised in 10 various locations (Table 1); 15 myocardial tissues of representative animals of various origins with the highest ELISA PP values were subjected to bioassay; subsequently, five isolates (TgPigSp1 to TgPigSp5) were obtained (Table 2). The bioassay success rate was 33.3% (5/15). The isolation rate appeared to increase at higher ELISA PP values; parasites were isolated from 0 of 6 pigs with PP values <110 and from 5 of 9 pigs with PP values > 110.
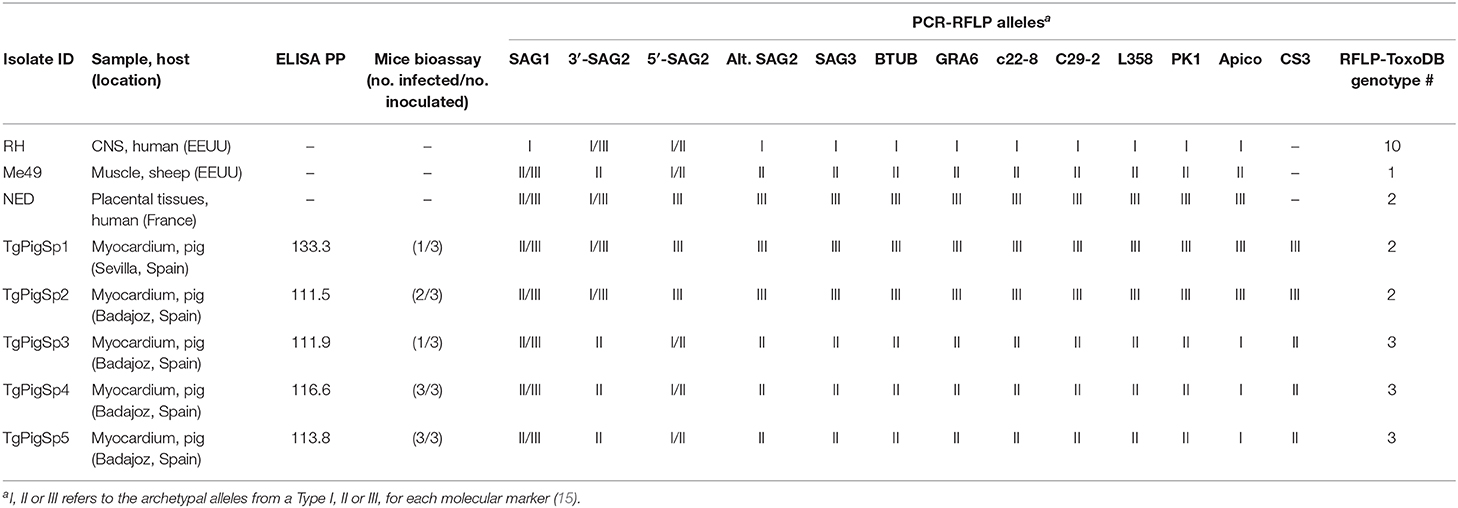
Table 2. PCR-RFLP genotyping of Toxoplasma gondii isolates from Iberian pigs.
Genetic Characterization
Cell culture-derived tachyzoites from all five isolates were successfully characterized as having two different genotypes: ToxoDB #3 (3/5 isolates) and ToxoDB #2 (2/5). The CS3 marker was previously reported to have a high predictive value on virulence in mice (16), and was present in type II alleles in all isolates with the ToxoDB #3 genotype; type III alleles were detected in all isolates with the ToxoDB #2 genotype (Table 2).
Pathogenicity Characterization
In assay A, the cumulative mortality rate was 87.5% for TgPigSp1 and 0% for the TgPigSp4 isolates, as shown in the survival curves (Figure 1A). Clear and statistically significant differences were observed between the survival curves of the two isolates at the doses starting from 102 tachyzoites/mouse (p < 0.01). Regarding morbidity rate, maximum clinical signs score reached by each animal is shown in Figure 1. In detail, the TgPigSp1 isolate caused clinical signs in 100% (16/16) of infected mice at a very acute phase of the infection at the doses starting from 102, rapidly inducing rounded back and noticeable loss of body conditions in the majority of mice (93.8%, 15/16); severe weight loss and development of neurological signs (with consequential humane euthanasia) (12.5%, 2/16) were observed with frequent cases of sudden death (75%, 12/16) (Figure 1A). All sudden deaths or humane euthanasia episodes occurred before 15 dpi, except an animal that died at 26 dpi. On the other hand, the TgPigSp4 isolate triggered clinical signs in 42.1% (8/19) of infected mice, specifically including ruffled coat and ascites in all cases during a very acute phase of the infection; however, the clinical signs receded completely at 13 dpi (Figure 1B).
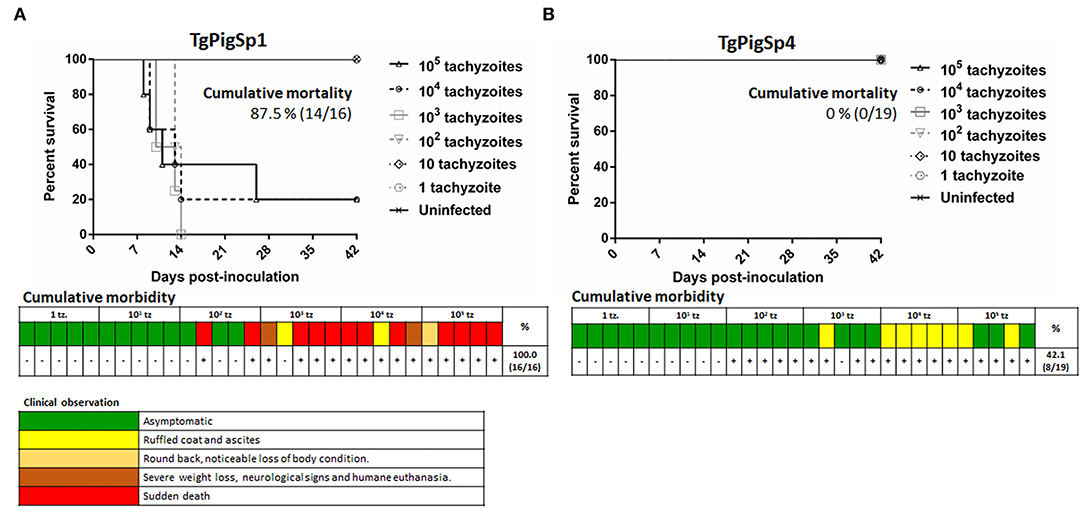
Figure 1. Cumulative mortality and morbidity parameters in the evaluation of virulence degree in two Toxoplasma gondii isolates from Iberian pigs. Survival curves and gradation of clinical signs observed in mice infected/inoculated with TgPigSp1 (A) and TgPigSp4 (B) isolates are shown. Colored boxes represent the most severe clinical sign observed in each individual mouse challenged with each dose according to clinical signs scoring included down below. +: mice with a confirmed Toxoplasma-infection by IFAT or tissue imprints of brain or lung. -: mice without a confirmed Toxoplasma-infection. Cumulative mortality rates are indicated.
Parasite burden reached by both isolates in the different organs studied at 7 and 30 dpi (assay B) is shown in Figure 2. All mice from the negative control group were seronegative and no Toxoplasma-DNA was detected by PCR in their tissue samples. CS3 sequences amplified from Toxoplasma-positive DNA samples from lung tissues of infected mice were aligned to previously obtained sequences from TgPigSp1 (MW132600), TgPigSp4 (MW132601) and other reference strains (TgRH, MW151245; TgMe49, MW151246; TgNED, MW151247). 100% identity was found between CS3 sequences got from TgPigSp1-infected tissues, that from the original isolate and TgNED sequence; likewise, all CS3 sequences from TgPigSp4-infected tissues were identical to TgPigSp4 isolate original one and to TgMe49 sequence, confirming the correct genotype of each inoculum. Only 2 out of 5 mice infected by TgPigSp1 isolate and scheduled for sacrifice at 30 dpi survived until that point; however, they were not seroconverted and did not have parasite DNA in their tissues. This fact revealed a low success rate of infection achieved, probably due to variations inherent to proceedings (e.g., slight differences in inocula or animal individual susceptibility), and supported the high mortality rate exposed previously in the case of TgPigSp1, which made not possible to compare the parasite burdens reached by each isolate at 30 dpi. At 7 dpi, all tested organs had higher average parasite burden in mice infected with TgPigSp1 than that in mice infected with TgPigSp4 isolate (Figure 2H); nevertheless, due to high variance of the data, differences were significant only between the parasite loads in the lung tissues, where TgPigSp1 infection had a median parasite load of 7.6 zoites/mg, whereas no T. gondii DNA was detected in mice infected with TgPigSp4 (p < 0.05) (Figure 2B). In the eye, at the acute stage, T. gondii-DNA was detected only in a mouse infected with TgPigSp1 (0.5 zoites/mg). Usually, T. gondii eye infection is not bilateral; detection by nPCR was performed only in the right eye of each animal thus limiting the frequency of detection. Statistically significant differences were detected in mice infected with TgPigSp4 at 7 vs. 30 dpi in terms of parasite burden in the brain (median 0 vs. 5 zoites/mg; p < 0.01), lung (median 0 vs. 54.7 zoites/mg; p < 0.01) and ocular tissues (median 0 vs. 7.7 zoites/mg; p < 0.05); no significant differences were detected in the case of heart, liver, and kidney tissues. Overall, total parasite burden reached in mice infected with TgPigSp4 was significantly increased from 7 to 30 dpi (median 0.7 vs. 83.3 zoites/mg; p < 0.05).
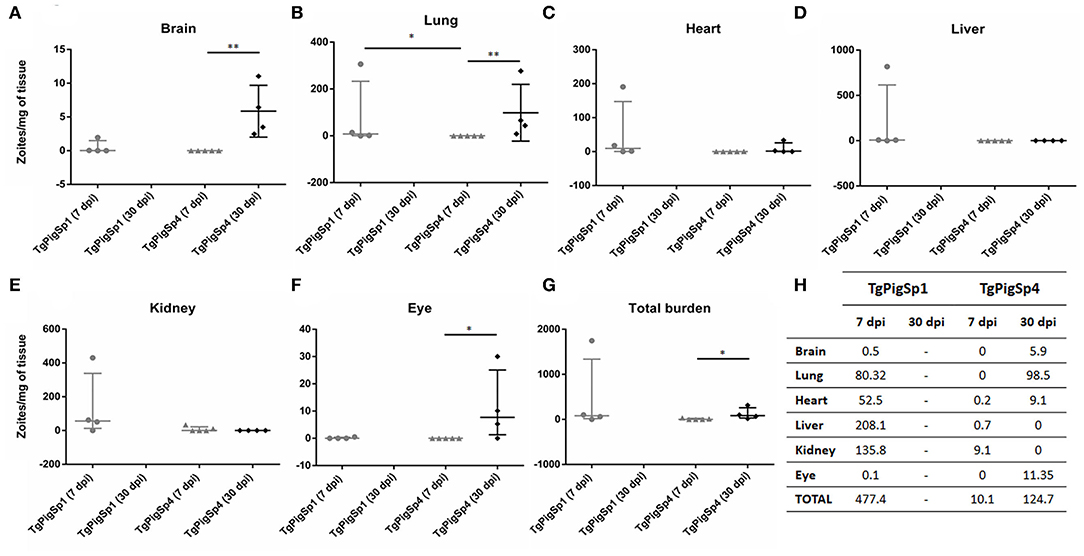
Figure 2. Parasite load (zoites/mg of tissue) found in six different murine organs (A–F) and total parasite burden per mouse (G) at 7- or 30-days post inoculation with 1,000 Toxoplasma gondii tachyzoites of TgPigSp1 and TgPigSp4 isolates. Median and interquartile values are represented. U-Mann-Whitney test; *p < 0.05; **p < 0.01. (H) Parasite burden (zoites/mg of tissue) mean values reached in each organ studied at 7 and 30 dpi by TgPigSp1 and TgPigSp4 isolates were tabulated.
In all histologically evaluated organs, lesions were mainly observed in the brain, liver, and lung, where multifocal aggregates of mononuclear inflammatory cells were detected (Figure 3). No lesions were found in the kidney, quadriceps femoris muscle or tongue tissue. Similar to the parasite load assessment, due to the high mortality rate caused by TgPigSp1, it was not possible to compare the tissue lesion patterns triggered by each isolate at 30 dpi. Organs of three seropositive mice scheduled to be sacrificed at 30 dpi that experienced early sudden dead or were humanely culled, were included for histopathological evaluation to provide further evidence of TgPigSp1 infection course; however, the results were not useful for comparison. In the liver of mice infected with TgPigSp1 lesions were subjectively more severe at an early phase of infection (taking into account the tissue samples from mice sacrificed at 7 dpi and samples from mice who died early at 11–14 dpi (four mice in total) than of those infected with TgPigSp4 (only one animal had grade 3 lesions at 7 dpi). Moreover, the only mouse with heart lesions (i.e., multifocal nonpurulent myocarditis, Figure 3) was infected with TgPigSp1 and died at 14 dpi. On the other hand, the lesions in the brain were distinguished by glial foci and perivascular infiltration of inflammatory cells, and were detected in two mice infected with TgPigSp4; this phenomenon can be explained by development of chronic infection induced by TgPigSp4 isolate unlike acute infection caused by TgPigSp1. Regarding the lung, only 2 mice (one infected by each isolate) had histopathological lesions at 7 dpi; the lesions were more notable in the case of the TgPigSp1 infection. Furthermore, a TgPigSp1-infected animal with evidence of myocarditis (died at 14 dpi) also presented inflammatory lesions in the lung. Finally, 4 out of 5 mice inoculated with TgPigSp4 isolate had multifocal aggregates of mononuclear cells in the lung at 30 dpi.
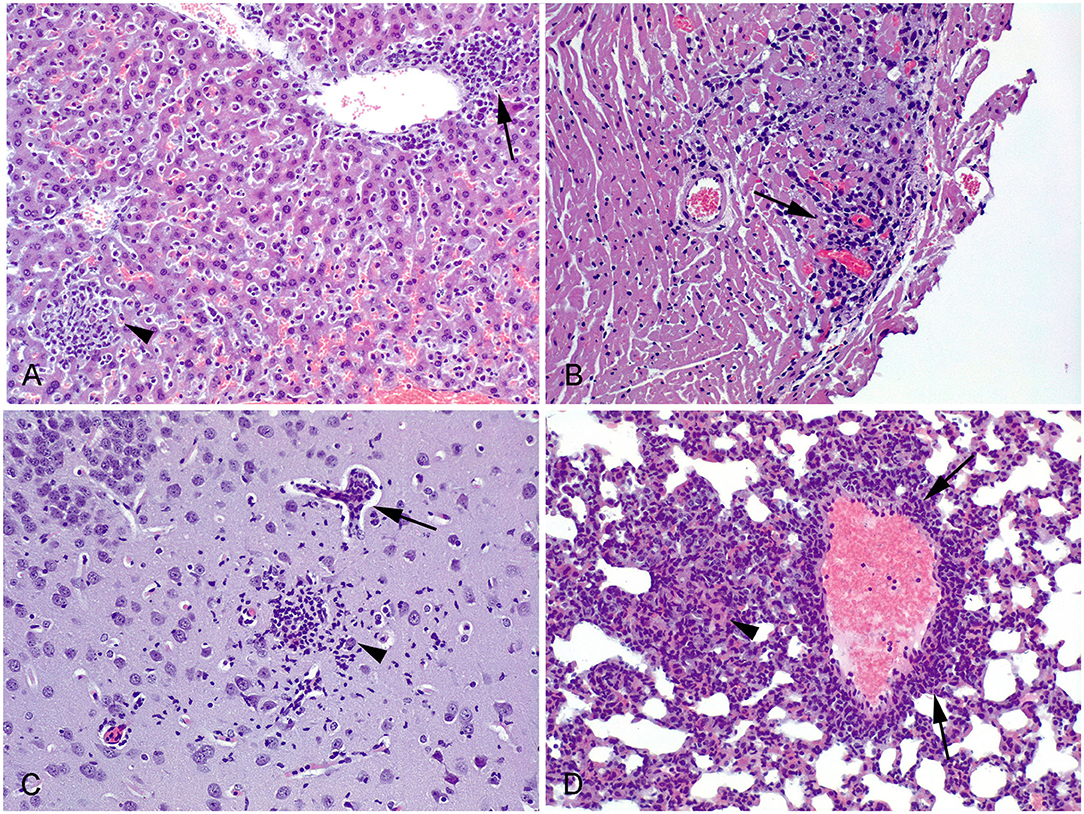
Figure 3. Histological lesions observed in Toxoplasma gondii infected mice. HE. 20 x. (A) Liver. Mouse infected with TgPigSp4, 7 dpi. Aggregation of mononuclear inflammatory cells at the hepatic parenchyma (arrowhead) and in relation with a central vein (arrow). (B) Heart. Mouse infected with TgPigSp1, 14 dpi. Foci of non-purulent myocarditis (arrow). (C) Brain. Mouse infected with TgPigSp4, 30 dpi. There is a glial focus at the neuropile (arrowhead) close to a small vessel showing non-purulent vasculitis (arrow). (D) Lung. Mouse infected with TgPigSp4, 30 dpi. Perivascular infiltration of nonpurulent inflammatory cells (arrows). A focal thickening of the alveolar wall, caused by the infiltration of inflammatory cells, could also be observed (arrowhead).
Discussion
The present study aimed to isolate, genotype, and evaluate the virulence degree in a normalized mouse model of the T. gondii strains infecting Iberian pigs. Seroprevalence values observed in the present study are in agreement with the previous data (9.5–58.2%) reported in the same host and breeding systems in Spain (9, 10, 22). Age and management systems are important factors related to an increase in the seroprevalence levels (11), which makes free-ranging fattening Iberian pigs a perfect model for T. gondii isolation and for evaluation of consumer risk when trends in organic pig production are concerned with natural breeding and welfare. The observed higher success of parasite isolation from the animals with higher ELISA titers should be considered in the future surveys.
Genetic characterization based on PCR-RFLP identified 2 genotypes, ToxoDB #3 (type II PRU-variant) and #2 (clonal type III), predominantly identified in the European domestic livestock (reviewed in (14)), wildlife (23) and humans (24–26). In Europe, the available data on genotyping of T. gondii in infected pigs is scarce; recently, an study (27) summarized the major facts, including the presence of all three clonal lineage alleles with an apparent predominance of type II; however, it should be noted that in most of studies, only one or a few molecular markers was used thus limiting the resolution of the results. A unique study using the same 11 molecular markers reported unexpected combinations of I, II and III alleles in 11 tissue samples from organic pigs in Italy (28). Notable frequency of type III alleles has been also described in Portuguese and Italian studies (29, 30); nonetheless, the studies were based only on SAG2 PCR-RFLP and five microsatellite loci (TUB2, TgM-A, W35, B17, and B18) or on B1-PCR-HRM (high resolution melting) genotyping assays, respectively. Similarly, a recent Serbian study reported that 22.2% (2/9) of detected strains correspond to the genotype III according to PCR-RFLP of GRA6, alt. SAG2, PK-1, BTUB, C22-8, CS3 and Apico markers (31); nevertheless, one of the isolates had a recombinant type II allele in the C22-8 marker. A Polish survey detected high prevalence of type III alleles in T. gondii DNA extracted from retail raw pork meat products (32). Similar results were obtained in the myocardial tissues of wild boars ranging in the southwestern locations in Spain (33) close to the sampling areas covered in the present study. Surprisingly, a high frequency of type I alleles was described in European literature (27, 34, 35) although type I were not detected in the present study except Apico marker identified within the Pruginaud (PRU) variant of the clonal genotype II. Remarkably, ToxoDB #3 genotype (type II PRU-variant) had not been described previously in studies using similar methodologies in infected European pigs (28, 31, 36). Different typing methodologies and sampling efforts used in the available references of the literature complicate the conclusions; however, it appears that overlapping strains from the three clonal genotypes are infecting European pig livestock, and recombination events within feline population are possible (28, 35, 36). This fact may be favored by the production systems involving pigs raised in semi-free ranging conditions that are exposed to potential sources of T. gondii, e.g., oocyst-contaminated environment or animal carcasses (37).
Currently, only two studies (29, 31) reported isolation of T. gondii from domestic swine in Europe; a third study (38) reported the seroconversion and DNA detection in the brain of mice inoculated with the homogenates of meat from 16 seropositive pigs reared in the indoor systems in the Aragon region of Spain. In wild swine, viable T. gondii were isolated from 21 wild boar heart tissue samples in France (23); all samples were identified as genotype II by PCR-RFLP (SAG1, SAG2, and GRA7) and microsatellite loci analysis (TUB2, TgM-A, W35, B17, B18, and M33). No additional data were reported thus increasing the interest in the present investigation along with the fact that no previous assays aimed to determine the virulence of European isolates from pigs.
The CS3 locus has been described previously as a highly predictive marker of mortality in mice inoculated with T. gondii isolates (16). Our CS3 typing results disagree with previous Brazilian and Chinese studies reporting high mortality rates associated with the type I or II alleles of the CS3 gene and low or null rates associated with the type III alleles (16, 39, 40). However, opposite results have been reported in studies from Brazil (41, 42) and Spain (14). Clearly, additional investigations are required to determine the role of this locus and its interaction with related well-known virulence factors, such as ROP5 and ROP18 (43), in Toxoplasma virulence in mice and other hosts.
In this study, an apparent correlation between the genotype and mortality and morbidity rates in mice is described; nonetheless, further studies are needed. Toxoplasma gondii clonal lineages I, II and III have been traditionally classified according to their virulence in outbred laboratory mice into highly virulent (type I), intermediate virulent (type II) and nonvirulent (type III) (44–46). Nevertheless, current population structure of T. gondii is not limited to the three clonal lineages and is considerably more complex with at least 16 haplogroups known worldwide (47, 48); moreover, pre-established virulence classification is apparently also disputed. In the present study, an isolate was classified as clonal type III according to 11 RFLP-markers (TgPigSp1) and therefore was expected to be nonvirulent; however, the isolate had 87.5% cumulative mortality in the standardized mouse model, whereas a type II isolate (genotype #3, TgPigSp4) was completely nonvirulent while it has been described as moderately virulent previously. In the case of the TgPigSp1 isolate, already starting from the dose of 102 tachyzoites/mouse, the mortality rates were 80–100%. High mortality rate detected in the present study is in agreement with a previous report from Japan, where oral doses of 102-cyst of a type III cat isolate were found to cause 100% mortality in mice; nevertheless, no clinical signs of infection were seen when micro minipigs were infected in a similar manner (49). Although a relative correspondence between the virulence of T. gondii strains in mice and their virulence in humans is traditionally assumed (50), there are many studies that point out different behavior of the parasite in different hosts (49, 51, 52). Thus, the mouse virulence model should be interpreted as a relative and not an absolute characterization method; therefore, the mortality values described here will be valid for a mouse model but not for assessing the virulence degree of the TgPigSp1 and TgPigSp4 isolates in a swine model. Increasing evidence of a host-dependent virulence and a broken linkage with genotype, emphasize the need to investigate still unknown T. gondii strains virulence factors.
The parasite load is associated with the histological lesions detected in the tested mouse tissues, indicating a higher degree of virulence of the TgPigSp1 isolate compared to that of the TgPigSp4 isolate thus supporting the results of the cumulative mortality and morbidity rates. A high incidence of the lesions in the tissues of mice infected by TgPigSp4 at 30 dpi, especially in the brain and lung, corresponds to the development of a chronic infection similar to mice that were infected with low virulence type II isolates (53); however, high cumulative mortality of TgPigSp1 does not permit any additional comparison.
In addition to the cumulative morbidity and mortality rates, the virulence degree can be evaluated by nonlethal infection parameters; a pioneering study identified the intra-genotype variations in the weight gain and both anti-T. gondii IgG antibodies and haptoglobin levels in the serum of mice infected with a panel of type II isolates (54). Interestingly, variations in these parameters were observed in strains of the same genotype isolated from different hosts. Recent evaluation of the virulence degree in Caribbean, Brazilian and European T. gondii strains of six different genotypes showed a wide variability in the mortality rate and in parasite burden in the mouse tissues at 8 dpi (55). Additional studies aiming to combine genotypic and phenotypic characterization of the European isolates will be of major interest to determine the possible presence of hidden intragenotype variations within the clonal T. gondii strains, which were formerly classified as moderately virulent (type II) and nonvirulent (type III).
It should be noted that the isolates included in the present experiments on phenotypical characterization have been obtained recently (low number of cell culture passages), fact that enables to conserve their in vivo biological behavior avoiding the adaptation to the cell culture conditions. This feature is important since various factors, such as life stage of the parasite or the number of passages in mice or in cell culture, have been repeatedly demonstrated in the literature to notably influence the evaluation of the virulence parameters (17, 49, 52).
The present study provides new information on the T. gondii strains circulating in swine in Europe and opens interesting avenues toward the epidemiological importance of trending organic farming and semi-free systems with regard to food safety, especially when domestic pigs are raised in sympatry with wildlife; in this sense, studies on the circulation of T. gondii from a One Health approach are of major interest. Considerable sampling and isolation efforts along with genotyping improvements were made to change the paradigm of the genetic structure of T. gondii population (56); similarly, implementation of virulence/phenotypical characterization of a large number of T. gondii strains by accurate models, including mortality and evaluation of nonlethal infection parameters, may let us to solve the raised controversy.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by Animal Welfare Committee of the Community of Madrid (PROEX 274/16).
Author Contributions
MF-E, RC-B, EC-F, and LO-M conceived and designed the laboratory tests. MF-E, RC-B, JR-C, RV, and JB performed experiments. MF-E, RC-B, RV, JR-C, and EC-F analyzed the data. LO-M, EC-F, and JB contributed reagents, materials, analysis tools. MF-E, RC-B, JR-C, JB, EC-F, and LO-M drafted the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the projects funded by the Spanish Ministry of Science and Innovation (AGL2016-75935-C2-R) and the Community of Madrid (PLATESA2-CM-P2018/BAA-4370). MF-E was funded by UCM-Santander/2017 pre-doctoral grants. RC-B, EC-F, and LO-M are part of the TOXOSOURCES consortium supported by the funding from the European Union's Horizon 2020 Research and Innovation Programme under the grant agreement No 773830: One Health European Joint Programme.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Authors thank Dr. Gereon Schares (Friedrich Loeffler Institut, Germany) for providing us with the Toxoplasma gondii reference strains (TgRH, TgMe49, and TgNED).
References
1. De Berardinis A, Paludi D, Pennisi L, Vergara A. Toxoplasma gondii, a foodborne pathogen in the swine production chain from a European perspective. Foodborne Pathog Dis. (2017) 14:637–48. doi: 10.1089/fpd.2017.2305
2. European Food Safety Authority (EFSA). Surveillance and monitoring of Toxoplasma in humans, food and animals. EFSA J. (2007) 583:1–64.
3. European Food Safety Authority (EFSA). Scientific Opinion on the public health hazards to be covered by inspection of meat (swine). EFSA J. (2011) 9:2351. doi: 10.2903/j.efsa.2011.2351
4. Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson MA, Roy SL, et al. Foodborne illness acquired in the United States–major pathogens. Emerg Infect Dis. (2011) 17:7–15. doi: 10.3201/eid1701.P11101
5. Cook AJ, Gilbert RE, Buffolano W, Zufferey J, Petersen E, Jenum PA, et al. Sources of toxoplasma infection in pregnant women: European multicentre case-control study. BMJ. (2000) 321:142–7. doi: 10.1136/bmj.321.7254.142
6. Opsteegh M, Schares G, van der Giessen J. On Behalf of the Consortium, Relationship Between Seroprevalence in the Main Livestock Species and Presence of Toxoplasma gondii in Meat (GP/EFSA/BIOHAZ/2013/01). An Extensive Literature Review. Final report. EFSA Supporting Publication 2016: EN-996 (2016). 294 p. doi: 10.2903/sp.efsa.2016.EN-996
7. Ministerio de Agricultura Pesca y Alimentación (MAPA). Información del Sector Porcino. (2019). Available online at: https://www.mapa.gob.es/es/ganaderia/temas/produccion-y-mercados-ganaderos/sectores-ganaderos/porcino/informacion-del-sector/default.aspx (accessed September 1, 2020).
8. Garrido-Fernández A, León-Camacho M. Assessing the effect of season, montanera length and sampling location on Iberian pig fat by compositional data analysis and standard multivariate statistics. Food Chem. (2019) 295:377–86. doi: 10.1016/j.foodchem.2019.05.123
9. Hernández M, Gómez-Laguna J, Tarradas C, Luque I, García-Valverde R, Reguillo L, et al. A serological survey of Brucella spp., Salmonella spp., Toxoplasma gondii and Trichinella spp. in Iberian fattening pigs reared in free-range systems. Transbound Emerg Dis. (2014) 61:477–81. doi: 10.1111/tbed.12049
10. Castillo-Cuenca JC, Díaz-Cao JM, Martínez-Moreno Á, Cano-Terriza D, Jiménez-Ruiz S, Almería S, et al. Seroepidemiology of Toxoplasma gondii in extensively raised Iberian pigs in Spain. Prev Vet Med. (2019) 175:104854. doi: 10.1016/j.prevetmed.2019.104854
11. Pablos-Tanarro A, Ortega-Mora LM, Palomo A, Casasola F, Ferre I. Seroprevalence of Toxoplasma gondii in Iberian pig sows. Parasitol Res. (2018) 117:1419–24. doi: 10.1007/s00436-018-5837-3
12. García-Bocanegra I, Dubey JP, Simon-Grifé M, Cabezón O, Casal J, Allepuz A. Seroprevalence and risk factors associated with Toxoplasma gondii infection in pig farms from Catalonia, north-eastern Spain. Res Vet Sci. (2010) 89:85–7. doi: 10.1016/j.rvsc.2010.01.017
14. Fernández-Escobar M, Calero-Bernal R, Benavides J, Regidor-Cerrillo J, Guerrero-Molina MC, Gutiérrez-Expósito D. Isolation and genetic characterization of Toxoplasma gondii in Spanish sheep flocks. Parasit Vectors. (2020) 13:396. doi: 10.1186/s13071-020-04275-z
15. Su C, Shwab EK, Zhou P, Zhu XQ, Dubey JP. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology. (2010) 137:1–11. doi: 10.1017/S0031182009991065
16. Pena HF, Gennari SM, Dubey JP, Su C. Population structure and mouse-virulence of Toxoplasma gondii in Brazil. Int J Parasitol. (2008) 38:561–9. doi: 10.1016/j.ijpara.2007.09.004
17. Saraf P, Shwab EK, Dubey JP, Su C. On the determination of Toxoplasma gondii virulence in mice. Exp Parasitol. (2017) 174:25e30. doi: 10.1016/j.exppara.2017.01.009
18. Arranz-Solís D, Benavides J, Regidor-Cerrillo J, Fuertes M, Ferre I, Ferreras MC, et al. Influence of the gestational stage on the clinical course, lesional development and parasite distribution in experimental ovine neosporosis. Vet Res. (2015) 46:19. doi: 10.1186/s13567-014-0139-y
19. Álvarez-García G, Collantes-Fernández E, Costas E, Rebordosa X, Ortega-Mora LM. Influence of age and purpose for testing on the cut-off selection of serological methods in bovine neosporosis. Vet Res. (2003) 34:341–52. doi: 10.1051/vetres:2003009
20. Castaño P, Fuertes M, Ferre I, Fernández M, Ferreras MC, Moreno-Gonzalo J, et al. Placental thrombosis in acute phase abortions during experimental Toxoplasma gondii infection in sheep. Vet Res. (2014) 45:9. doi: 10.1186/1297-9716-45-9
21. Castaño P, Fuertes M, Regidor-Cerrillo J, Ferre I, Fernández M, Ferreras MC. Experimental ovine toxoplasmosis: influence of the gestational stage on the clinical course, lesion development and parasite distribution. Vet Res. (2016) 47:43. doi: 10.1186/s13567-016-0327-z
22. García-Bocanegra I, Simon-Grifé M, Dubey JP, Casal J, Martín GE, Cabezón O, et al. Seroprevalence and risk factors associated with Toxoplasma gondii in domestic pigs from Spain. Parasitol Int. (2010) 59:421–6. doi: 10.1016/j.parint.2010.06.001
23. Richomme C, Aubert D, Gilot-Fromont E, Ajzenberg D, Mercier A, Ducrot C, et al. Genetic characterization of Toxoplasma gondii from wild boar (Sus scrofa) in France. Vet Parasitol. (2009) 164:296–300. doi: 10.1016/j.vetpar.2009.06.014
24. Ajzenberg D, Yera H, Marty P, Paris L, Dalle F, Menotti J, et al. Genotype of 88 Toxoplasma gondii isolates associated with toxoplasmosis in immunocompromised patients and correlation with clinical findings. J Infect Dis. (2009) 199:1155–67. doi: 10.1086/597477
25. Jokelainen P, Murat JB, Nielsen HV. Direct genetic characterization of Toxoplasma gondii from clinical samples from Denmark: not only genotypes II and III. Eur J Clin Microbiol Infect Dis. (2018) 37:579–86. doi: 10.1007/s10096-017-3152-z
26. Herrmann DC, Maksimov P, Hotop A, Groß U, Däubener W, Liesenfeld O, et al. Genotyping of samples from German patients with ocular, cerebral and systemic toxoplasmosis reveals a predominance of Toxoplasma gondii type II. Int J Med Microbiol. (2014) 304:911–16. doi: 10.1016/j.ijmm.2014.06.008
27. Vergara A, Marangi M, Caradonna T, Pennisi L, Paludi D, Papini R, et al. Toxoplasma gondii lineages circulating in slaughtered industrial pigs and potential risk for consumers. J Food Prot. (2018) 81:1373–78. doi: 10.4315/0362-028X.JFP-17-496
28. Bacci C, Vismarra A, Mangia C, Bonardi S, Bruini I, Genchi M, et al. Detection of Toxoplasma gondii in free-range, organic pigs in Italy using serological and molecular methods. Int J Food Microbiol. (2015) 202:54–6. doi: 10.1016/j.ijfoodmicro.2015.03.002
29. de Sousa S, Ajzenberg D, Canada N, Freire L, da Costa JMC, Dardé ML, et al. Biologic and molecular characterization of Toxoplasma gondii isolates from pigs from Portugal. Vet Parasitol. (2006) 135:133–6. doi: 10.1016/j.vetpar.2005.08.012
30. Papini R, di Ciccio P, Marangi M, Ghidini S, Zanardi E, Vergara A, et al. Occurrence of Toxoplasma gondii in carcasses of pigs reared in intensive systems in northern Italy. J Food Prot. (2017) 80:515–22. doi: 10.4315/0362-028X.JFP-16-314
31. Kuruca L, Uzelac A, Klun I, Lalošević V, Djurković-Djaković O. Toxoplasma gondii genotypes circulating in domestic pigs in Serbia. Acta Vet Hung. (2019) 67:204–11. doi: 10.1556/004.2019.022
32. Sroka J, Bilska-Zajac E, Wójcik-Fatla A, Zajac V, Dutkiewicz J, Karamon J, et al. Detection and molecular characteristics of Toxoplasma gondii DNA in retail raw meat products in Poland. Foodborne Pathog Dis. (2019) 16:195–204. doi: 10.1089/fpd.2018.2537
33. Calero-Bernal R, Saugar JM, Frontera E, Pérez-Martín JE, Habela MA, Serrano, et al. Prevalence and genotype identification of Toxoplasma gondii in wild animals from southwestern Spain. J Wildl Dis. (2015) 51:233–8. doi: 10.7589/2013-09-233
34. Turčeková L, Antolová D, Reiterová K, Spišák F. Occurrence and genetic characterization of Toxoplasma gondii in naturally infected pigs. Acta Parasitol. (2013) 58:361–6. doi: 10.2478/s11686-013-0154-6
35. Battisti E, Zaneta S, Trisciuoglio A, Bruno S, Ferroglio E. Circulating genotypes of Toxoplasma gondii in Northwestern Italy. Vet Parasitol. (2018) 253:43–7. doi: 10.1016/j.vetpar.2018.02.023
36. Slany M, Reslova N, Babak V, Lorencova A. Molecular characterization of Toxoplasma gondii in pork meat from different production systems in the Czech Republic. Int J Food Microbiol. (2016) 238:252–5. doi: 10.1016/j.ijfoodmicro.2016.09.020
37. Stelzer S, Basso W, Benavides J, Ortega-Mora LM, Maksimov P, Gethmann J, et al. Toxoplasma gondii infection and toxoplasmosis in farm animals: risk factors and economic impact. Food Waterborne Parasitol. (2019) 15:e00037. doi: 10.1016/j.fawpar.2019.e00037
38. Herrero L, Gracia MJ, Pérez-Arquillué C, Lázaro R, Herrera M, Herrera, et al. Toxoplasma gondii: pig seroprevalence, associated risk factors and viability in fresh pork meat. Vet Parasitol. (2016) 224:52–9. doi: 10.1016/j.vetpar.2016.05.010
39. Wang L, Cheng HW, Huang KQ, Xu YH, Li YN, Du J, et al. Toxoplasma gondii prevalence in food animals and rodents in different regions of China: isolation, genotyping, and mouse pathogenicity. Parasit Vectors. (2013) 6:273. doi: 10.1186/1756-3305-6-273
40. Rocha DS, Nilsson MG, Maciel BM, Pena HFJ, Alves BF, Silva AV, et al. Genetic diversity of Toxoplasma gondii isolates from free-range chickens in Bahia, Brazil. J Parasitol. (2018) 104:377–82. doi: 10.1645/18-9
41. Langoni H, Matteucci G, Medici B, Camossi LG, Richini-Pereira VB, Silva RC. Detection and molecular analysis of Toxoplasma gondii and Neospora caninum from dogs with neurological disorders. Rev Soc Bras Med Trop. (2012) 45:365–8. doi: 10.1590/S0037-86822012000300016
42. Rêgo WMF, Costa JGL, Baraviera RCA, Pinto LV, Bessa GL, Lopes REN, et al. Association of ROP18 and ROP5 was efficient as a marker of virulence in atypical isolates of Toxoplasma gondii obtained from pigs and goats in Piauí, Brazil. Vet Parasitol. (2017) 247:19–25. doi: 10.1016/j.vetpar.2017.09.015
43. Behnke MS, Khan A, Lauron EJ, Jimah JR, Wang Q, Tolia NH, et al. Rhoptry proteins ROP5 and ROP18 are major murine virulence factors in genetically divergent south American strains of Toxoplasma gondii. PLoS Genet. (2015) 11:e1005434. doi: 10.1371/journal.pgen.1005434
44. Sibley LD, Boothroyd JC. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature. (1992) 359:82–5. doi: 10.1038/359082a0
45. Howe DK, Sibley LD. Toxoplasma gondii comprises three clonal lineages: correlation of parasite genotype with human disease. J Infect Dis. (1995) 172:1561–6. doi: 10.1093/infdis/172.6.1561
46. Khan A, Taylor S, Ajioka JW, Rosenthal BM, Sibley LD. Selection at a single locus leads to widespread expansion of Toxoplasma gondii lineages that are virulent in mice. PLoS Genet. (2009) 5:e1000404. doi: 10.1371/journal.pgen.1000404
47. Su C, Khan A, Zhou P, Majumdar D, Ajzenberg D, Dardé ML, et al. Globally diverse Toxoplasma gondii isolates comprise six major clades originating from a small number of distinct ancestral lineages. Proc Natl Acad Sci USA. (2012) 109:5844–9. doi: 10.1073/pnas.1203190109
48. Lorenzi H, Khan A, Behnke MS, Namasivayam S, Swapna LS, Hadjithomas M, et al. Local admixture of amplified and diversified secreted pathogenesis determinants shapes mosaic Toxoplasma gondii genomes. Nat Commun. (2016) 7:10147. doi: 10.1038/ncomms10147
49. Taniguchi Y, Appiah-Kwarteng C, Murakami M, Fukumoto J, Nagamune K, Matsuo T, et al. Atypical virulence in a type III Toxoplasma gondii strain isolated in Japan. Parasitol Int. (2018) 67:587–92. doi: 10.1016/j.parint.2018.05.010
50. Shwab EK, Jiang T, Pena HFJ, Gennari SM, Dubey JP, Su C. The ROP18 and ROP5 gene allele types are highly predictive of virulence in mice across globally distributed strains of Toxoplasma gondii. Int J Parasitol. (2016) 46:141–6. doi: 10.1016/j.ijpara.2015.10.005
51. Niedelman W, Gold DA, Rosowski EE, Sprokholt JK, Lim D, Arenas AF, et al. The rhoptry proteins ROP18 and ROP5 mediate Toxoplasma gondii evasion of the murine, but not the human, interferon-gamma response. PLoS Pathog. (2012) 8:e1002784. doi: 10.1371/journal.ppat.1002784
52. Sánchez-Sánchez R, Ferre I, Regidor-Cerrillo J, Gutiérrez-Expósito D, Ferrer LM, Arteche-Villasol N, et al. Virulence in mice of a Toxoplasma gondii type II isolate does not correlate with the outcome of experimental infection in pregnant sheep. Front Cell Infect Microbiol. (2019) 8:436. doi: 10.3389/fcimb.2018.00436
53. Ingram WM, Goodrich LM, Robey EA, Eisen MB. Mice infected with low-virulence strains of Toxoplasma gondii lose their innate aversion to cat urine, even after extensive parasite clearance. PLoS ONE. (2013) 8:e75246. doi: 10.1371/journal.pone.0075246
54. Jungersen G, Jensen L, Rask MR, Lind P. Non-lethal infection parameters in mice separate sheep Type II Toxoplasma gondii isolates by virulence. Comp Immunol Microbiol Infect Dis. (2002) 25:187–95. doi: 10.1016/S0147-9571(01)00039-X
55. Hamilton CM, Black L, Oliveira S, Burrells A, Bartley PM, Melo RPB, et al. Comparative virulence of Caribbean, Brazilian and European isolates of Toxoplasma gondii. Parasit Vectors. (2019) 12:104. doi: 10.1186/s13071-019-3372-4
Keywords: Toxoplasma gondii, Iberian pigs, isolates, genotypes, virulence
Citation: Fernández-Escobar M, Calero-Bernal R, Regidor-Cerrillo J, Vallejo R, Benavides J, Collantes-Fernández E and Ortega-Mora LM (2020) Isolation, Genotyping, and Mouse Virulence Characterization of Toxoplasma gondii From Free Ranging Iberian Pigs. Front. Vet. Sci. 7:604782. doi: 10.3389/fvets.2020.604782
Received: 10 September 2020; Accepted: 26 October 2020;
Published: 20 November 2020.
Edited by:
Damer Blake, Royal Veterinary College (RVC), United KingdomReviewed by:
Chunlei Su, The University of Tennessee, Knoxville, United StatesGeoff Hide, University of Salford, United Kingdom
Copyright © 2020 Fernández-Escobar, Calero-Bernal, Regidor-Cerrillo, Vallejo, Benavides, Collantes-Fernández and Ortega-Mora. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rafael Calero-Bernal, ci5jYWxlcm9AdWNtLmVz; Luis Miguel Ortega-Mora, bHVpcy5vcnRlZ2FAdWNtLmVz