Corrigendum: Extracorporeal Shock Wave Therapy Enhances the In Vitro Metabolic Activity and Differentiation of Equine Umbilical Cord Blood Mesenchymal Stromal Cells
 Ramés Salcedo-Jiménez1
Ramés Salcedo-Jiménez1 Judith B. Koenig1*
Judith B. Koenig1* Olivia J. Lee2
Olivia J. Lee2 Thomas W. G. Gibson1
Thomas W. G. Gibson1 Pavneesh Madan2
Pavneesh Madan2 Thomas G. Koch2
Thomas G. Koch2- 1Department of Clinical Studies, University of Guelph, Guelph, ON, Canada
- 2Department of Biomedical Sciences, University of Guelph, Guelph, ON, Canada
Extracorporeal shock wave therapy (ESWT) has been shown to induce different biological effects on a variety of cells, including regulation and stimulation of their function and metabolism. ESWT can promote different biological responses such as proliferation, migration, and regenerations of cells. Recent studies have shown that mesenchymal stromal cells (MSCs) secrete factors that enhance the regeneration of tissues, stimulate proliferation and differentiation of cells, and decrease inflammatory and immune reactions. Clinically, the combination of these two therapies has been used as a treatment for tendon and ligament lesions in horses; however, there is no scientific evidence supporting this combination of therapies in vivo. Therefore, the objectives of the study were to evaluate the effects of ESWT on equine umbilical cord blood mesenchymal stromal cells (CB-MSCs) proliferative, metabolic, migrative, differentiation, and immunomodulatory properties in vitro. Three equine CB-MSC cultures from independent donors were treated using an electrohydraulic shock wave generator attached to a water bath. All experiments were performed as triplicates. Proliferation, viability, migration and immunomodulatory properties of the cells were evaluated. Equine CB-MSCs were induced to evaluate their trilineage differentiation potential. ESWT treated cells had increased metabolic activity, showed positive adipogenic, osteogenic, and chondrogenic differentiation, and showed higher potential for differentiation toward the adipogenic and osteogenic cell fates. ESWT treated cells showed similar immunomodulatory properties to none-ESWT treated cells. Equine CB-MSCs are responsive to ESWT treatment and showed increased metabolic, adipogenic and osteogenic activity, but unaltered immunosuppressive properties. In vivo studies are warranted to determine if synergistic effects occur in the treatment of musculoskeletal injuries if ESWT and equine CB-MSC therapies are combined.
Introduction
Extracorporeal shock wave therapy (ESWT) is one of the leading treatments of certain orthopedic diseases in humans such as plantar fasciitis and lateral epicondylitis; more recently, it has been used to treat Achilles and patellar tendinopathies (1, 2). ESWT is also a common treatment for tendon and ligament injuries in the horse particularly suspensory ligament desmitis (3). At present, it is also used as adjunct therapy for the management of osteoarthritis in horses (4). Medically used shock waves are generated in a fluid medium, by one of three different generators: electrohydraulic, piezoelectric, or electromagnetic. These acoustic waves are generated outside the body and can be focused to a specific point within the body. The shape of the generated acoustic wave is characterized by an initial positive rapid phase, of high amplitude, followed by a sudden phase of mild negative pressure, and then returns to the ambient values. Their peak pressure is high—up to 100 mpa (500 bar) with a rapid rise (<10 ns) in pressure, of short duration (<10 ms) and a broad range of frequency (5, 6).
A variety of cell types, including mesenchymal stromal cells (MSCs), were shown to be responsive to ESWT. After focused ESWT, human bone marrow stromal cells (hBMSCs) had increased proliferation and migration, the rate of apoptosis activation was reduced, and all treated cells maintained their differentiation potentials (7). Rat adipose-derived stromal cells (ASCs) responded to ESWT with an elevation of mesenchymal markers and a higher capacity to differentiate toward adipogenic and osteogenic lineages with minimal changes in proliferation (8). When equine adipose tissue-derived mesenchymal stromal cells were treated with ESWT, they showed increased proliferation; however, ESWT did not change their differentiation potential (9). The reported increased proliferation after shockwave treatment may be mediated through purinergic receptors via downstream ErK1/2 signals following activation by extracellular ATP (10).
MSCs are multipotent cells capable of differentiating into osteogenic, adipogenic and chondrogenic lineages (11). Bone marrow aspirates and adipose tissues are the most studied and used sources of MSCs for clinical cases in the horse (12). It has been suggested that MSCs from umbilical cord blood (CB-MSCs) might have superior immune tolerance, proliferative potential, and differentiation potency (13). MSCs have both progenitor and non-progenitor functions. Their progenitor function is characterized by their ability to differentiate into different cell types, and their non-progenitor function allows them to have immunosuppressive effects (14). Recent studies have shown that MSCs secrete factors that enhance regeneration of injured tissue, stimulate proliferation and differentiation of endogenous cells, and decrease inflammatory and immune reactions (14–16).
Clinically, the combination of ESWT and MSC therapies has been used as a treatment for tendon and ligament lesions in horses without scientific evidence supporting this approach. We hypothesize that ESWT will affect the progenitor functions and non-progenitor functions of equine CB-MSCs. The aim of this study was to evaluate if ESWT affects equine CB-MSCs' proliferative, metabolic, migrative, differentiation, and immunomodulatory properties in vitro.
Materials and Methods
Culture of Equine CB–MSCs and in vitro Extracorporeal Shock Wave Treatment
The equine CB-MSC were established as previously described by Koch et al. (17). Briefly, the mononuclear cell fraction (MNCF) was cultured and non-adherent cells were removed through successive complete medium changes. When cell numbers allowed, the cells were passaged and expanded until they were cryopreserved for later use.
Three cryovials of equine CB-MSC isolated from different donors were used in these studies. The cryovials had been stored in liquid nitrogen from 1 to 9 months prior to the study and all contained cells at passage 3. The cells were thawed, seeded at a cell density of 5,000 cells/cm2 culture surface area and expanded until ~80% confluence to obtain sufficient cell numbers for all the experiments and assays of these studies.
The in vitro shock wave treatment was performed as described by Holfeld et al. (18), using an electrohydraulic shock wave generator (VersaTron, Pulse Veterinary Technologies, Alpharetta GA, USA). Equine CB-MSCs at passage 5 were treated at a confluence of 90% in T-25 cell culture flasks. The flasks were placed in front of the applicator at a distance of 5 cm from the probe to the cell layer inside the water container in direct contact with pre-warmed (37°C) water. The focused shock waves were applied with the following parameters: an energy flux density of 0.1 mJ/mm2, frequency of 3 Hz and 300 impulses (Figure 1).
FIGURE 1

Figure 1. Water bath for ESWT application. The bottom of the culture flask containing the adherent cells are facing the ESWT probe.
The control group was maintained in the same culture conditions, consisting of equilibrated expansion media or differentiation media without shock wave exposure. All cell culture experiments were performed as triplicates from 3 different donors. The process is shown in flow the chart (Figure 2).
FIGURE 2
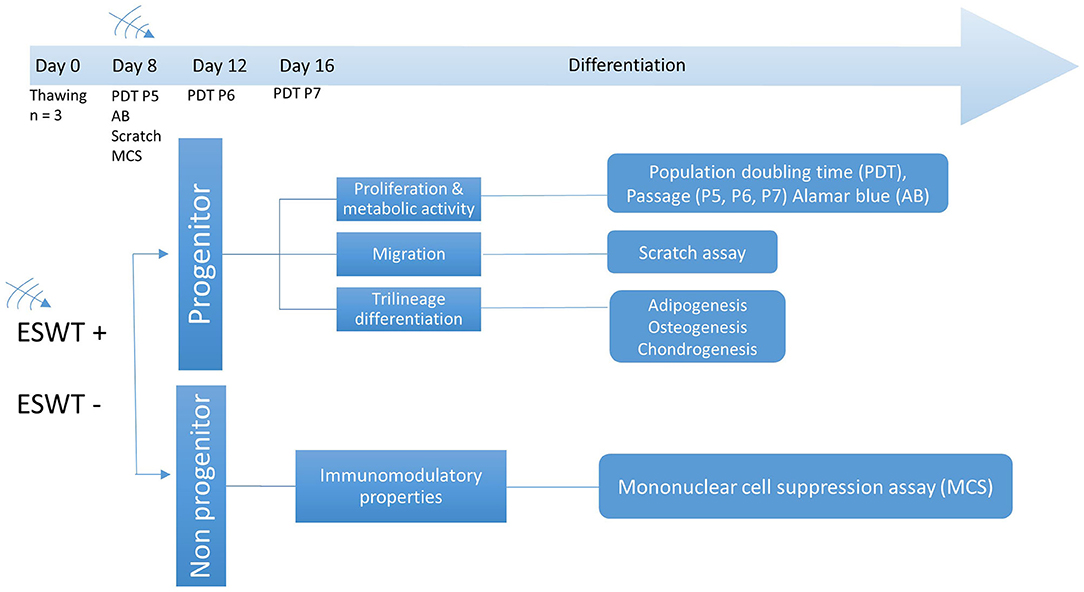
Figure 2. Experimental design. Time points for ESWT, respective passage numbers (P5, P6, P7) and population doubling time (PDT), Alamar blue assay (AB), scratch (migration) assay, and mononuclear cell suppression assay (MCS). Differentiation was initiated after treatment on day 8.
Proliferation
Cells were detached using trypsin 0.025% (Lonza, Walkersville, MD, USA) and re-seeded in 6 well culture flasks. To document cell morphology, digital images were obtained prior to detachment at passage 5, 6, and 7 using phase-contrast microscopy and Q-Capture software (Q-Imaging, Surrey, BC, Canada).
Doubling times for passage 5, 6 and 7 were calculated according to Tessier et al. (19):
1. Cell–doubling number (CD) = In(Nh/Ns)/In 2
In = Natural Logarithm
Nh = harvest cell number
Ns = seed cell number
2. Cell doubling time (DT) = CT/CD
CT = cell culture time (19).
Metabolic Activity
Equine CB-MSC were seeded in 96-well plates at 5,000 cells/cm2 in 300 μl of culture media. The following day the culture media was changed. On day 4, the culture media was replaced with 300 μl of 10% resazurin (Sigma Aldrich, Oakville, ON, Canada) in phosphate-buffered saline (PBS), incubated at 37°C for 4 h protected from light and read by a plate reader (Spectramax i3, Molecular Devices, Sunnyvale, CA, USA) at 585 nm using an excitation wavelength of 555 nm.
Migration (Scratch Assay)
For the scratch migration assay, equine CB-MSC were seeded at 4 x 104 cells in 70 μl into each dish (Culture-Insert 2 well in μ-Dish 35 mm, high Ibidi cat# 80206) and incubated, cells were grown to confluence. The Culture-Insert 2 well was removed, and the cell layer was washed with PBS to remove debris and 2 ml of culture medium were added. Images were taken at 0, 12, 24, 36, and 48-h time points or until complete closure of the scratch wound was observed using a motorized inverted microscope (IX81, Olympus America Inc., Melville, NY, USA). The dishes were incubated between each time point. After all images were acquired, measurements of the surface area of each scratch at each time points were performed with Photoshop (Adobe Systems Incorporated, NY, USA).
Trilineage Differentiation Studies
For adipogenesis, equine CB-MSC were seeded at 2.1 x 104 cells/ml in 6 well plates, media was changed every 2–3 days. After the cells reached 100% confluence the cells were exposed to adipogenic induction media (BulletKit, Adipogenesis, Lonza). The induction medium consisted of 1 mM dexamethasone, 0.5 mM 3-isobutyl-1- methyl-xanthine (IBMX), 10 mg/ml recombinant human (rh) insulin, 0.2 mM indomethacin and 10% fetal calf serum (FCS) in DMEM. Media was changed every 2–3 days as previously described, Koch et al. (17). The adipogenic potential was assessed after 14 days of exposure to induction media by Oil Red O staining to observe lipid droplets and AdipoRed™ assay (Lonza).
For Oil Red O staining, all the media was aspirated off the wells, wash once with 2 ml of PBS, then replaced with 2 ml of 10% formalin (Fischer Scientific, Whitby, ON, Canada) for 30 mins at room temperature to fixate the cells, the formalin was replaced with 2 ml of sterile water (Milli-Q water, Millipore, Mississauga, ON, Canada) for 2 mins and then replaced with 60% isopropanol (Fischer Scientific) for 5 mins, the isopropanol was replaced with 2 ml of Oil Red O (Sigma-Aldrich,) working solution [3 parts (300 mg Oil Red O in 100 ml of 99%o isopropanol) mixed with 2 parts sterile water], after 5 mins the Oil red O solution was washed of indirectly with tap water. Two ml of Harris hematoxylin (Sigma Aldrich) were added to the wells for 1 min before being aspirated and the wells washed with warm tap water.
The AdipoRed Assay (Lonza) was performed according to the manufacturer's protocol for 96 well plates. Briefly, equine CB-MSC were induced for 14 days. The supernatant was removed and rinsed with 200 μl PBS. Then each well was filled with 200 μl of PBS, 5 ul of reagent were injected followed by shaking for 1 s. The fluorescence was measured with excitation at 485 nm and emission at 520 nm as previously described by Koch et al. (17).
Chondrogenesis differentiation was performed in pellet culture as previously described by Co et al. (20). ~2.5 x 105 CB-MSCs were suspended in 200 μl of the chondrogenic medium, containing High Glucose Dulbecco's modified Eagle medium (HG-DMEM) (Sigma Aldrich; Oakville, ON), 200 mM Glutamax (Invitrogen; Burlington, ON, Canada), 100 mM sodium pyruvate (Invitrogen), 1 ABAM (Invitrogen), 0.1 mM dexamethasone (Sigma Aldrich), 100 mg/ml ascorbic acid-2 phosphate (Sigma Aldrich), 40 mg/ml proline (Sigma Aldrich), 1 ITS (BD Biosciences; Mississauga, ON, Canada), and 10 ng/ml TGF-β3 (R&D Systems; Minneapolis, MN, USA), the cells were plated in V-bottom polypropylene 96 well plates (Phenix #MPG-651201) and centrifuged at 200x g for 10 mins and incubated for 3 weeks. The media was changed every 2–3 days. Chondrogenic potential was assessed by Toluidine Blue to visualize glycosaminoglycan (GAG) and biochemistry.
For Toluidine blue stain, the pellets were fixed in 10% formalin, dehydrated in isopropanol and embedded in paraffin and sectioned (5 μM). Slides were deparaffinized and rehydrated prior to staining. The stock solution of Toluidine blue contained 1 g powder (Sigma Aldrich) in 100 ml 70% EtOH diluted to 10 % with 1% NaCl. Sections were stained in Toluidine blue working solution for 2 mins and washed in three changes of distilled water.
For biochemistry evaluation, the pellets were digested in 40 mg/ml papain (Invitrogen; Burlington, ON, Canada), 20 mM ammonium acetate, 1 mM EDTA, and 1 mM dithiothreitol (DTT) (Sigma Aldrich) for 48 h at 65°C vortexing at 24 h. Stored at −40°C until further analysis. The proteoglycan content of the digest was estimated by quantifying the amount of sulfated glycosaminoglycans (GAG), using the dimethylmethylene blue dye and spectrophotometry with a wavelength of 525 nm. The standard curve for the proteoglycan content assay was generated using chondroitin sulfate (Sigma Aldrich). DNA content was assessed using a commercial kit: DNA quantitation kit e Fluorescence assay (Sigma Aldrich) in a 96-well plate. The assay was carried out according to the manufacturer's instructions.
For osteogenesis, equine CB-MSC were seeded at a density of 3,000 cells/cm2 in 6 well plates, media was changed every 2–3 days. After the cells reached 100% confluency the cells were exposed to osteogenic induction media (BulletKit, osteogenic, Lonza). The induction medium consisted of DMEM-LG, 10% FBS, 1% L-glutamine, 1% antibiotic-antimycotic (ABAM), 100 nM dexamethasone, 10 mM β-glycerophosphate, and 0.05 mM ascorbic acid-2-phosphate. The media was changed every 3–4 days. The osteogenic potential was assessed by Alizarin Red S staining for calcium deposition and semi-quantitative alkaline phosphatase enzyme activity assay.
For Alizarin Red S staining, the pH of the Alizarin solution was adjusted to 4.1–4.3 with ammonium hydroxide. The media was aspirated from each well and the cells were fixed in ice-cold 70% ethanol for 5 mins at room temperature. Alcohol was aspirated and the wells were rinsed twice (5 mins) with water, the water was aspirated, and 1 ml of 2% Alizarin Red S solution was added, it was incubated for 3 mins at room temperature and removed, wells were washed 5 times with 2 ml of water.
For Alkaline phosphatase enzyme activity assay, the culture wells were rinsed twice with PBS, 0.5 ml of PBS was added to the well and the cells were scraped off and transferred to a 1.5 ml Eppendorf tube. The wells were rinsed with 0.5 ml PBS, which was transferred to the Eppendorf tube as well. The sample was centrifuged at 5000 Å~ g for 8 s. All the PBS was aspirated, and the cell pellet was resuspended in 0.1 ml lysis buffer by pipetting the cells up and down several times. The lysis buffer consisted of (500 ml lysis buffer: 250 mg sodium deoxycholate, 5 mg phenylmethyl sulfonyl fluoride (PMSF), 5 mg aprotinin, 500 ml nonidet P-40, 500 ml 10% (wt/vol) SDS). The Eppendorf tubes were left on ice for 5 mins and vortexed for 30 s, then they were microcentrifuged at maximum speed for 10 mins at 6°C. Fifty microliters of the supernatant were added in triplicate from each osteo-induced and control sample to a well in a 96 well plate, then the kit, p-Nitrophenyl Phosphate Liquid Substrate System (Sigma-Aldrich), was used and 50 μl of p-nitrophenyl phosphate (pNPP) were added to each well. The absorbance was read at 405 nm on a microplate reader as soon as possible after adding the pNPP (time 0) and subsequent every 5 mins for 20 mins. The absorbance was read against 100 μl of undiluted pNPP. Covering it with aluminum foil screened the microplate from light between readings.
Mononuclear Cell Suppression Assay (MSA)
A pooled peripheral blood mononuclear cell (MNC) from five donors MSA was used to evaluate equine CB-MSCs for their ability to suppress MNC proliferation in vitro. Briefly, MNCs from five unrelated donors were mixed in equal ratios and stimulated with 5 ug/mL mitogen concanavalin A (Sigma Aldrich). Negative control pooled MNCs were not stimulated with mitogen. ESWT treated and non-treated CB-MSC cultures were irradiated (20 Gy) and 50,000 MSCs were seeded in each well of a 48 well plates. Stimulated, pooled MNCs were added to MSCs at a ratio of 1:10 (CB-MSC:MNC) in MNC media (RPMI 1640 with 10% horse serum, 1% L-glutamine and 1% pen/strep) and subsequently incubated at 38°C in 5% CO2. After three days of co-culturing, cells were stained with Bromodeoxyuridine (BrdU) and assessed with a BrdU ELISA kit on the fourth day (Roche, Mississauga, ON, Canada) following the manufacturer's protocol. The amount of MNC proliferation in each treatment condition was quantified by the amount of absorbance at 370 nm.
*Average Absorbance (370 nm) of each individual MNC donor = the average absorbance of the five MNC donors that were mixed in equal ratios cultured individually in separate wells.
Statistical Analysis
For statistical analysis, the effect of ESWT delivered on equine CB-MSCs was evaluated. A statistical program (SAS 9.2, Proc MIXED, SAS Institute Inc., Cary, North Carolina, USA) was used to fit a general linear mixed model. For proliferation, the design was a two-factor factorial in a randomized complete block design with subsampling. Fixed effect factors were treatment and passage. Interactions were also included in the model. Random effects were cell culture, treatment by cell culture by passage and then rep nested within treatment, cell culture and passage. For metabolic activity, the data was divided by a million to facilitate statistical analysis. The design was also a randomized complete block design with subsampling, were the fixed effect is treatment and the random effect is cell culture. The experimental unit is cell culture by treatment. For migration, the design was a two-factor factorial in a randomized complete block design with fixed effect factors treatment and time. To accommodate time being a repeated measure, the following correlation structures (offered by SAS) were attempted: ar(1), arh(1), toep, toep(2)-toep(5), toeph toeph(2)-toeph(5) un un(2)-un(5), and one was chosen based on Akaike Information Criterion (AIC). For adipogenesis, the design was a two-factor factorial in a randomized complete block design by subsampling with fixed effect factors cell culture and treatment. And for osteogenesis to accommodate time being a repeated measure AIC was chosen. Repeated measures were nested in cell culture by treatment. Again, these error structures were attempted: ar(1) arh(1) toep toep(2)-toep(4) toeph toeph(2)-toeph(4) un un(2)-un(4). For the mononuclear cell suppression assay, a one-way ANOVA was performed with Tukey's multiple comparisons test. Differences were considered significant at p ≤ 0.05.
To assess the ANOVA assumptions, comprehensive residual analyses were performed. The assumption of normality was formally tested by use of Shapiro-Wilk, Kolmogorov-Smirnov, Cramér-von Mises, and Anderson-Darling tests. Residuals were plotted against the predicted values and explanatory variables that were used in the models to look for outliers, bimodal distributions, the need for data transformations, or other issues that should be addressed.
Results
Proliferation
Treated and untreated equine CB-MSC showed an elongated, spindle-shaped fibroblast-like morphology. There were no obvious morphological differences between the treated and the untreated equine CB-MSCs (Figure 3).
FIGURE 3
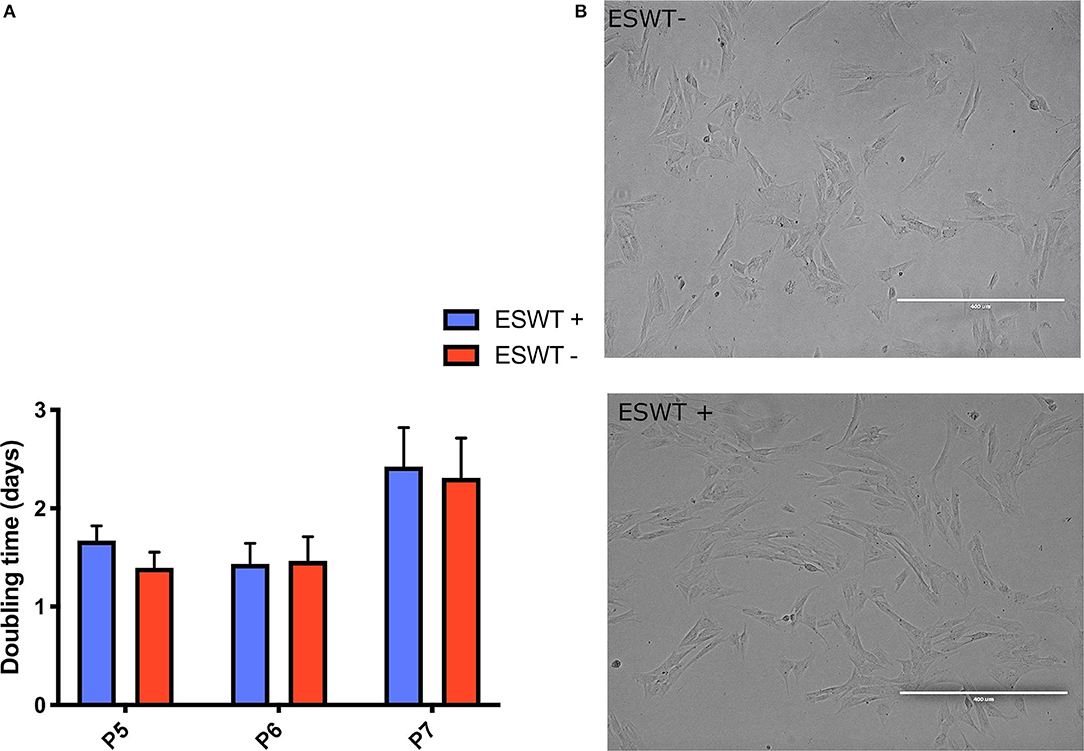
Figure 3. Proliferation. (A) Population doubling times (days) for treated (ESWT+) and untreated (ESWT–) equine CB-MSCs for passage 5 (P5), 6 (P6), and 7 (P7). Results are shown as mean and CI 95% (p > 0.05). No differences were observed. (B) There were no obvious morphological differences between the treated (ESWT+) and untreated (ESWT–) equine CB-MSCs.
Population-doubling time was measured from passage 5 to 7. Mean (95% confidence interval (CI) doubling time in days was as follows: passage 5, 1.67 days (0.83–3.14 days) for treated equine CB-MSC and 1.39 days (0.80–3.03 days) for untreated CB-MSC, for passage 6, 1.43 days (0.68–2.41 days) for treated equine CB-MSC and 1.46 days (0.67–2.39 days) for untreated equine CB-MSC and for passage 7, 2.42 days (1.11–4.30 days) for treated equine CB-MSC and 2.39 days (1.11–4.35 days) for untreated equine CB-MSC. There were no differences (p > 0.05) between treated and untreated equine CB-MSC (Figure 3).
Metabolic Activity
Treated equine CB-MSC had higher metabolic activity than untreated equine CB-MSC as evaluated by the Alamar blue assay (p = 0.0002). Mean relative fluorescent units (RFU) was 102.29 (84.95–119.62 RFU) for treated equine CB-MSC and 65.10 RFU (47.76–82.43 RFU) for untreated equine CB-MSC (Figure 4).
FIGURE 4
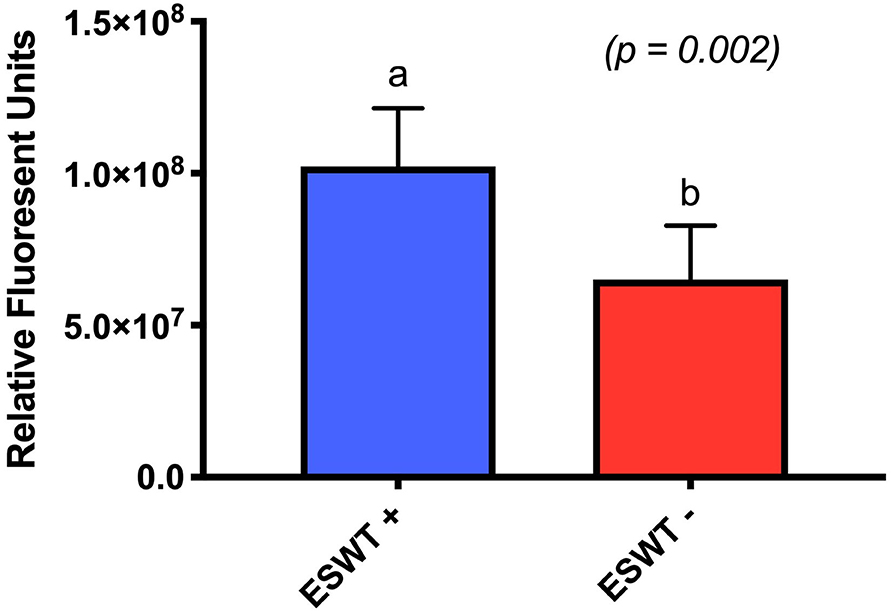
Figure 4. Metabolic activity. Relative fluorescent units for treated (ESWT+) and untreated (ESWT–) equine CB-MSCs. Results are shown as mean and CI 95% (p = 0.0002). Values with different letters are statistically different.
Migration (Scratch Assay)
Data was mildly non-normal, unequal variances in time were accommodated, and the assumptions were not well met. For the scratch assay, the results obtained at the 60-h time point were removed from the analysis as most of the scratches were already closed. For all the experiments, the time to closure among groups was similar. There was no difference for each area measured between the treated and the untreated equine CB-MSC (Figure 5).
FIGURE 5
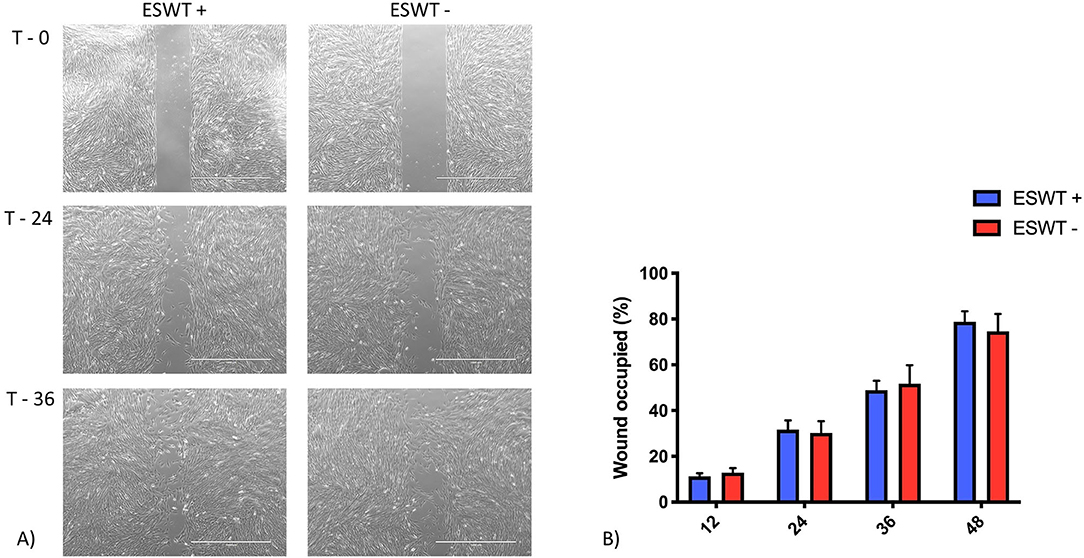
Figure 5. Migration. (A) Representative scratch assay for treated (ESWT+) and untreated (ESWT–) equine CB-MSC. Images were obtained using a motorized inverted microscope until closure of the scratch in the cell layer. (B) Percentage of wound occupied, calculated by dividing the none recovered area at 12, 24, 36, and 48 h, respectively by the initial wound area at 0 h and subtracting this value as a percentage from 100%. There were no differences (p > 0.05) between the treated (ESWT+) and untreated (ESWT–) equine CB-MSC at each time point.
Trilineage Differentiation
Positive trilineage potency was observed for the treated and the untreated equine CB-MSC, no subjective differences were observed for the differentiation capacities when evaluated by Oil Red O, Alizarin Red S, and Toluidine blue staining, respectively. Increased adipogenic differentiation occurred in the treated equine CB-MSC compared to the untreated cells based on a higher intracellular lipid formation (p = 0.0002). Higher osteogenic differentiation occurred in the treated equine CB-MSC compared to the untreated cells as demonstrated by a higher alkaline phosphatase activity in the initial minutes (p < 0.0001) (Figure 6). There was no difference for chondrogenesis as evaluated on biochemistry and wet mass [mean 1.2 mg (0.83–1.57 mg) for treated equine MSC and 1.0 mg (0.82–1.37 mg) for untreated equine MSC].
FIGURE 6
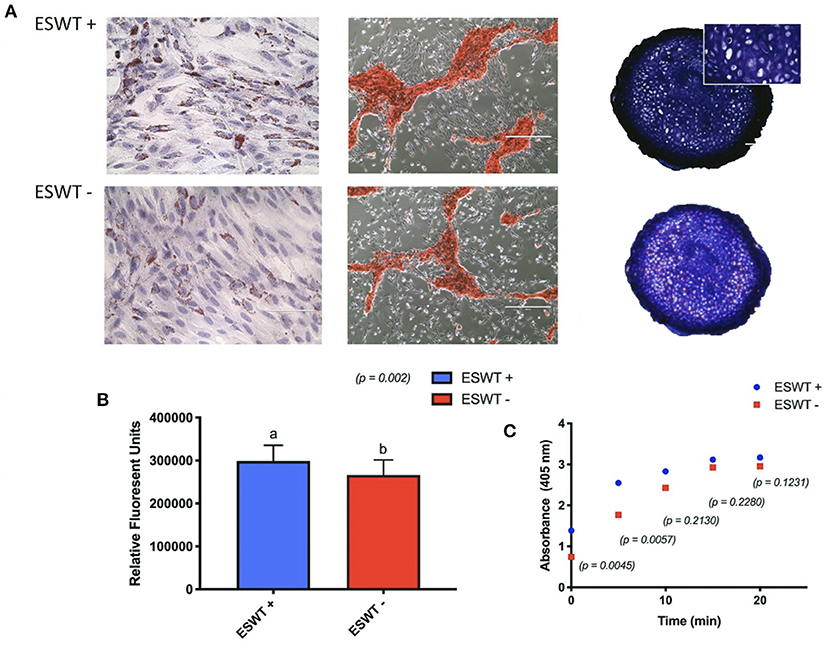
Figure 6. Trilineage differentiation. (A) Positive trilineage potency for adipogenesis (Oil Red O), osteogenesis (Alizarin Red S), and chondrogenesis (Toluidine blue) for treated (ESWT+) and untreated (ESWT–) equine CB-MSC. (B) Quantification of intracellular lipid droplets by relative fluorescent units for treated (ESWT+) and untreated (ESWT–) equine CB-MSC. Results are shown as mean and CI 95% (p = 0.0002). Values with different letters are statistically different (p = 0.002). (C) Alkaline phosphatase activity measured in relative fluorescent units in treated (ESWT+) and untreated (ESWT–) equine CB-MSC. Differences were found at time 0 (p = 0.0045) and 5 mins (p = 0.0057).
Mononuclear Cell Suppression Assay
To investigate the immunomodulatory potency of equine CB-MSC treated with ESWT, we used a pooled mononuclear cell suppression assay (pMSA) to compare MNC suppression of equine CB-MSC with and without ESWT. Treated and non-treated equine CB-MSC were co-culture with pooled MNCs for 96 h and MNC proliferation was assessed with BrdU ELISA. Both treated and non-treated equine CB-MSC were able to suppress MNC relative to the positive control. The ESWT treated and non-treated MSCs suppressed MNCs similarly (Figure 7).
FIGURE 7
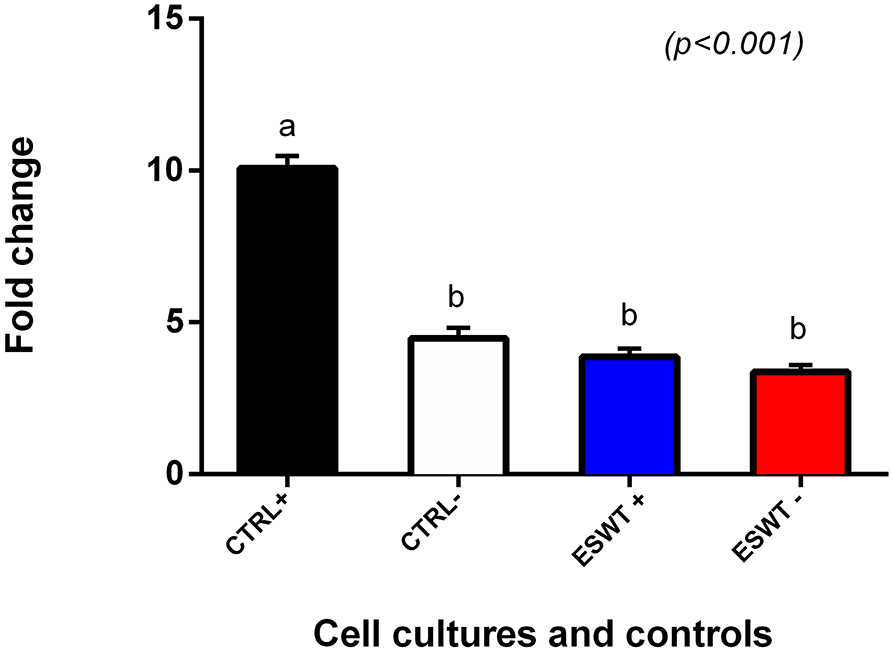
Figure 7. Mononuclear cell suppression assay. MNC suppression assay to determine MNC proliferation suppression capacity of equine CB-MSC. Both treated (ESWT+) and untreated (ESWT–) equine CB-MSC were able to suppress the proliferation of stimulated lymphocytes to the level of unstimulated lymphocytes (CTRL–). Values with different letters are statistically different.
Discussion
In the present study, we demonstrated that equine CB-MSCs respond to ESWT stimulation. The proliferation of these cells was not adversely influenced by shockwave treatment, but the metabolic activity of the treated cells was increased. This increased metabolic activity observed in equine CB-MSCs after ESWT is in agreement with other studies that reported increased proliferation, migration, and metabolic activity of human and rat ASCs (8, 10). Moreover, we have shown that equine CB-MSCs maintain their multilineage differentiation potential after treatment with ESWT. Treated cells do not only maintain but also increase their potency toward adipogenic and osteogenic lineages. There is growing evidence suggesting that ESWT has an effect on MSCs differentiation (7, 9, 10).
Until now, only one experiment reported the combination of equine MSCs and ESWT in vitro; the authors reported that shockwaves stimulate adipose derived stem cells to a higher potential for differentiation toward adipogenic, osteogenic and chondrogenic lineages (9). In contrast, Suhr et al. (7) found that applying shockwaves to human bone marrows MSCs reduced the osteogenic and chondrogenic differentiation, and the adipogenic differentiation was unaffected. Similar to our results, Leone et al. (21) found an improved differentiation toward adipogenic and osteogenic lineages in human tendon-derived stem/progenitor cells. As cell signaling studies have shown, same types of cells respond differently to the same stimulus (22), we believe this variation in response to treatment to be the reason why MSCs from different sources, have a different biological behavior after ESWT.
In the search of MSCs with an increased rate of survival after transplantation, Heneidi et al. (23) isolated and characterized a new population of ASCs named multilineage differentiating stress enduring cells. The cells were isolated using severe stress conditions in an attempt to improve the low rates of post-transplant cell survival. In our study after exposing the equine CB-MSCs to ESWT, an increase in adipogenic potency was observed in addition to an increase in metabolic activity. It is possible that ESWT can precondition the cells, allowing them a superior performance when transplanted. Furthermore, the combination of ESWT with ASCs has been shown beneficial, as observed in experimental studies of brain damage in rats where this combination of ESWT and MSCs treatment reduces the effect of local and systemic inflammatory immune reactions, oxidative stress and apoptosis and even reduced damage to the mitochondria in the brain (24).
The contribution of the immunomodulatory ability of MSCs to tissue regeneration and healing has long been discussed. Using a MSA based on pooled MNCs, we observed that ESWT did not affect the MSCs' ability to suppress MNC proliferation in vitro. Our study is the first to examine the effect that equine CB-MSCs treated with ESWT has on MNC suppression. A previous study compared the surface markers and cytokine secretions of macrophages and demonstrated that the expression of classical pro-inflammatory M1 markers, CD80 and COX2, decreased after ESWT, shifting the monocytes into the anti-inflammatory M2 phenotype. ESWT had also reduced the secretion of pro-inflammatory cytokines by M1 macrophages (25). ESWT may influence MNCs subpopulations when applied directly, but whether ESWT can affect MNCs indirectly through modifying MSCs still requires more investigation. It is possible that ESWT can influence MSCs' surface marker expression and cytokine levels, which would subsequently affect MNCs. Varying the energy level of the ESWT could change the outcome and promote the acquisition of an anti-inflammatory profile as previously described by Sukubo et al. (25).
In the current study, shockwaves were applied via a water bath. The water bath was specifically designed to allow propagation of the shock waves after passing through the cell culture, as it has been suggested that waves that are not propagated will get reflected at the culture medium and will disrupt the upcoming waves as well as the cell monolayer (18). Conflicting information about the in vitro application of shock waves is published. Some authors recommend the application of the shock waves to flasks with an MSC cell monolayer in a water bath as we reported here; while others suggest applying shockwaves directly to tubes or flasks containing an MSC cell suspension through an interface (water-cushion, pigskin) (7, 26). Using the water bath in this study allowed us to standardize conditions, like temperature and distance, as well as its being reported as a model that simulates the in vivo conditions present during treatment with ESWT (18). One of the disadvantages of the method used in our study, is that some detachment of the cells in the monolayer can occur, and the cells located at the periphery might receive lower doses of ESWT than in the center (27).
The effects on differentiation and proliferation of equine CB-MSCs after ESWT observed in our study can be explained by mechanotransduction; which is how cells respond to mechanical stimulation and convert these stimulations into biochemical signals that can alter growth factor expression and cellular adaptation (28). The complete mechanism of action still remains unknown but there is evidence that ESWT can stimulate the mitogen–activated protein kinase (MAPK) cascade (2, 29). As reported by Weihs et al. (10), shock wave treatment initiates the release of ATP activating purinergic receptors that enhance proliferation by increasing ERK ½ signaling.
Our study was performed in an in vitro model and we have a relative low number of equine cultured cell lines investigated. The exact energy of ESWT that needs to be applied to equine CB-MSCs is unknown. Despite the fact that the study showed a significant increase in metabolic activity and increased adipogenic and osteogenic potential, the results are limited to the specific experimental settings selected here and should not be applied to clinical cases. Prospective clinical trials are needed to evaluate the effects of ESWT on different sources of MSCs in vitro and in vivo.
In conclusion, our findings indicate that ESWT can enhance the metabolic activity without adversely affecting the proliferation and immunomodulatory properties of equine CB-MSCs. For trilineage differentiation, we showed that ESWT allowed equine CB-MSCs to maintain their differentiation capacity; furthermore, treated cells showed an increased potency toward adipogenic and osteogenic lineages. The combined effect of these therapies remains to be seen, but the anecdotal clinical impression of improved healing after the combination of these two therapies can be partially explained through the increased metabolic activity and no effect on their immunomodulatory properties observed in this study. Further in vitro and in vivo studies will be required to fully investigate the effects of ESWT on CB-MSCs and the mechanisms by which the combination of these treatments appear to improve healing of soft tissue injuries.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The study was reviewed and approved by the University of Guelph Animal Care Committee AUP 3717.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
Pulse Vet provided the shockwave equipment for this study. The study was funded by Equine Guelph. RS's doctorate tuition was funded by a grant from CONACYT Mexico.
Conflict of Interest
TK acts in a volunteer capacity as non-executive Director, Scientific Affairs (ex officio) of eQcell therapies Inc., Aurora, ON, Canada, a company for which TK's research laboratory provides equine stem cell isolation and storage services. TK holds a minor non-controlling share in eQcell therapies Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank William Sears for his assistance with the statistical analysis. An abstract from this research was presented at the Annual Meeting of the American College of Veterinary Surgeons in Phoenix, Arizona, October 25, 2018 and presented at the North American Veterinary Regenerative Society Conference in Niagara Falls, Ontario, September 10, 2019. This manuscript has been released as a pre-print at bioRxiv (30).
References
1. Ioppolo F, Rompe JD, Furia JP, Cacchio A. Clinical application of shock wave therapy (SWT) in musculoskeletal disorders. Eur. J. Phys. Rehabil. Med. (2014) 50:217–30.
2. Chamberlain GA, Colborne GR. A review of the cellular and molecular effects of extracorporeal shockwave therapy. Vet. Comp. Orthop. Traumatol. (2020) 29:99–107. doi: 10.3415/VCOT-15-04-0057
3. Bosch G, de Mos M, van Binsbergen RV, van Schie HTM, van de Lest CHA, van Weeren PR. The effect of focused extracorporeal shock wave therapy on collagen matrix and gene expression in normal tendons and ligaments. Equine Vet. J. (2009) 41:335–41. doi: 10.2746/042516409X370766
4. Revenaugh MS. Extracorporeal shock wave therapy for treatment of osteoarthritis in the horse: clinical applications. Vet. Clin. North Am. (2005) 21:609–25. doi: 10.1016/j.cveq.2005.09.001
5. Ogden JA, Tóth-Kischkat A, Schultheiss R. Principles of shock wave therapy. Clin. Orthop. Relat. Res. (2001) 387:8–17. doi: 10.1097/00003086-200106000-00003
6. McClure S, Weinberger T. Extracorporeal shock wave therapy: clinical applications and regulation. Clin. Tech. Equine Pract. (2003) 2:358–67. doi: 10.1053/j.ctep.2004.04.007
7. Suhr F, Delhasse Y, Bungartz G, Schmidt A, Pfannkuche K, Bloch W. Cell biological effects of mechanical stimulations generated by focused extracorporeal shock wave applications on cultured human bone marrow stromal cells. Stem Cell Res. (2013) 11:951–64. doi: 10.1016/j.scr.2013.05.010
8. Schuh CMAP, Heher P, Weihs AM, Banerjee A, Fuchs C, Gabriel C, et al. In vitro extracorporeal shock wave treatment enhances stemness and preserves multipotency of rat and human adipose-derived stem cells. Cytotherapy. (2014) 16:1666–78. doi: 10.1016/j.jcyt.2014.07.005
9. Raabe O, Shell K, Goessl A, Crispens C, Delhasse Y, Eva A, et al. Effect of extracorporeal shock wave on proliferation and differentiation of equine adipose tissue-derived mesenchymal stem cells in vitro. Am. J. Stem Cells. (2013) 2:62–73.
10. Weihs AM, Fuchs C, Teuschl AH, Hartinger J, Slezak P, Mittermayr R, et al. Shock wave treatment enhances cell proliferation and improves wound healing by ATP release-coupled extracellular signal-regulated kinase (ERK) activation. J. Biol. Chem. (2014) 289:27090–104. doi: 10.1074/jbc.M114.580936
11. Caplan AI, Bruder SP. Mesenchymal stem cells: building blocks for molecular medicine in the 21st century. Trends Mol. Medi. (2001) 7:259–64. doi: 10.1016/S1471-4914(01)02016-0
12. Gutierrez-Nibeyro SD. Commercial cell-based therapies for musculoskeletal injuries in horses. Vet. Clin. North Am. (2011) 27:363–71. doi: 10.1016/j.cveq.2011.04.001
13. Li T, Xia M, Gao Y, Chen Y, Xu Y. Human umbilical cord mesenchymal stem cells: an overview of their potential in cell-based therapy. Exp. Opin. Biol. Ther. (2015) 15:1293–306. doi: 10.1517/14712598.2015.1051528
14. Mattar P, Bieback K. Comparing the immunomodulatory properties of bone marrow, adipose tissue, and birth-associated tissue mesenchymal stromal cells. Front. Immunol. (2015) 6:560. doi: 10.3389/fimmu.2015.00560
15. Chang Lo C, Chen HH, Chen KH, Chiang JY, Li YC, Lin HS, et al. Adipose-derived mesenchymal stem cell-derived exosomes markedly protected the brain against sepsis syndrome induced injury in rat. Am. J. Transl. Res. (2019) 11:3955–71.
16. Johnson V, Webb T, Norman A, Coy J, Kurihara J, Regan D, et al. Activated mesenchymal stem cells interact with antibiotics and host innate immune responses to control chronic bacterial infections. Sci. Rep. (2017) 7:9575. doi: 10.1038/s41598-017-08311-4
17. Koch TG, Heerkens T, Thomsen PD, Betts DH. Isolation of mesenchymal stem cells from equine umbilical cord blood. BMC Biotechnol. (2007) 7:26. doi: 10.1186/1472-6750-7-26
18. Holfeld J, Tepeköylü C, Kozaryn R, Mathes W, Grimm M, Paulus P. Shock wave application to cell cultures. J. Vis. Exp. (2014) 86:e51076. doi: 10.3791/51076
19. Tessier L, Bienzle D, Williams LB, Koch TG. Phenotypic and immunomodulatory properties of equine cord blood-derived mesenchymal stromal cells. PLoS ONE. (2015) 10:e0122954. doi: 10.1371/journal.pone.0122954
20. Co C, Vickaryous MK, Koch TG. Membrane culture and reduced oxygen tension enhances cartilage matrix formation from equine cord blood mesenchymal stromal cells in vitro. Osteoarthr. Cartil. (2014) 22:472–80. doi: 10.1016/j.joca.2013.12.021
21. Leone L, Raffa S, Vetrano M, Ranieri D, Malisan F, Scrofani C, et al. Extracorporeal shock wave treatment (ESWT) enhances the in vitro-induced differentiation of human tendon-derived stem/progenitor cells (hTSPCs). Oncotarget. (2016) 7:6410–23. doi: 10.18632/oncotarget.7064
22. Tay S, Hughey JJ, Lee TK, Lipniacki T, Quake SR, Covert MW. Single-cell NF-B dynamics reveal digital activation and analogue information processing. Nature. (2010) 466:267–71. doi: 10.1038/nature09145
23. Heneidi S, Simerman AA, Keller E, Singh P, Li X, Dumesic DA, et al. Awakened by cellular stress: isolation and characterization of a novel population of pluripotent stem cells derived from human adipose tissue. PLoS ONE. (2013) 8:e64752. doi: 10.1371/journal.pone.0064752
24. Chen KH, Hsiao HY, Wallace CG, Lin KC, Li YC, Huang TH, et al. Combined adipose-derived mesenchymal stem cells and low-energy extracorporeal shock wave therapy protect the brain from brain death-induced injury in rat. J. Neuropathol. Exp. Neurol. (2019) 78:65–77. doi: 10.1093/jnen/nly108
25. Sukubo NG, Tibalt E, Respizzi S, Locati M, D'Agostino MC. Effect of shock waves on macrophages: a possible role in tissue regeneration and remodeling. Int. J. Surg. (2015) 24(Pt B):124–30. doi: 10.1016/j.ijsu.2015.07.719
26. Zhao Y, Wang J, Wang M, Sun P, Chen J, Jin X, et al. Activation of bone marrow-derived mesenchymal stromal cells-a new mechanism of defocused low-energy shock wave in regenerative medicine. Cytotherapy. (2013) 15:1449–57. doi: 10.1016/j.jcyt.2013.08.012
27. Rinella L, Marano F, Berta L, Bosco O, Fraccalvieri M, Fortunati N, et al. Extracorporeal shock waves modulate myofibroblast differentiation of adipose-derived stem cells. Wound Repair Regen. (2016) 24:275–86. doi: 10.1111/wrr.12410
28. d'Agostino MC, Craig K, Tibalt E, Respizzi S. Shock wave as biological therapeutic tool: from mechanical stimulation to recovery and healing, through mechanotransduction. Int. J. Surg. (2015) 24:147–53. doi: 10.1016/j.ijsu.2015.11.030
29. Chen YJ, Kuo YR, Yang KD, Wang CJ, Chen SMS, Huang HC, et al. Activation of extracellular signal-regulated kinase (ERK) and p38 kinase in shock wave-promoted bone formation of segmental defect in rats. Bone. (2004) 34:466–77. doi: 10.1016/j.bone.2003.11.013
Keywords: shock wave, mesenchymal stromal cells, equine stem cells, umbilical cord, immunomodulatory
Citation: Salcedo-Jiménez R, Koenig JB, Lee OJ, Gibson TWG, Madan P and Koch TG (2020) Extracorporeal Shock Wave Therapy Enhances the In Vitro Metabolic Activity and Differentiation of Equine Umbilical Cord Blood Mesenchymal Stromal Cells. Front. Vet. Sci. 7:554306. doi: 10.3389/fvets.2020.554306
Received: 21 April 2020; Accepted: 11 November 2020;
Published: 04 December 2020.
Edited by:
Martin Andreas Vidal, Cave Creek Equine Surgical and Diagnostic Imaging Center, United StatesReviewed by:
Takashi Taguchi, Western University of Health Sciences, United StatesCristina Esteves, University of Edinburgh, United Kingdom
Copyright © 2020 Salcedo-Jiménez, Koenig, Lee, Gibson, Madan and Koch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Judith B. Koenig, amtvZW5pZ0B1b2d1ZWxwaC5jYQ==