
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 09 September 2020
Sec. Veterinary Infectious Diseases
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.00628
 Alicia Romanò1,2†
Alicia Romanò1,2† Alessandra Gazzola3*†
Alessandra Gazzola3*† Valentina Bianchini1
Valentina Bianchini1 Claudia Cortimiglia1Antonio M. Maisano1
Claudia Cortimiglia1Antonio M. Maisano1 Paola Cremonesi4Hans U. Graber2Fausto Vezzoli1
Paola Cremonesi4Hans U. Graber2Fausto Vezzoli1 Mario Luini1,4
Mario Luini1,4Staphylococcus aureus is one of the major pathogens responsible for intramammary infections in small ruminants, causing severe economic losses in dairy farms. In addition, S. aureus can contaminate milk and dairy products and produce staphylococcal enterotoxins, being responsible for staphylococcal food poisoning. Currently, data on the population structure and the virulence gene patterns of S. aureus strains isolated from goat milk is limited. Therefore, this study aimed at defining Ribosomal Spacer PCR (RS-PCR) genotypes, clonal complexes (CC), spa types, and virulence gene profiles of S. aureus isolated from goat milk samples from Lombardy region of Italy. A total of 295 S. aureus isolates from 65 goat bulk tank milk samples were genotyped by RS-PCR. spa typing and virulence gene patterns of a subgroup of 88 isolates were determined, and MLST was performed on a further subgroup of 39 isolates, representing all the spa types identified during the analysis. This study revealed 7 major genotypic clusters (CLR, CLAA, CLZ, CLAW, CLBW, CLS, and CLI), of which S. aureus CLR (19.8%) was the most common. A total of 26 different spa types were detected, the most prevalent types were t1773 (24%), t5428 (22.7%), and t2678 (12.5%). Overall, 44.3% of all isolates harbored at least one enterotoxin gene. The most prevalent was the combination of sec-sel genes (35.2%). Based on their MLST, isolates were assigned to 14 different CC, with majority grouped as CC133 (24%), CC130 (19.6%), and CC522 (19.6%). The caprine S. aureus population was depicted with a minimum spanning tree and an evolutionary analysis based on spa typing and MLST, respectively. Then, the variability of such strains was compared to that of bovine strains isolated in the same space-time span. Our results confirmed that S. aureus isolates from goats have wide genetic variability and differ from the bovine strains, supporting the idea that S. aureus from small ruminants may constitute a distinct population.
Staphylococcus aureus is one of the major pathogens responsible for clinical and subclinical mastitis in goats, causing significant economic losses due to the decrease of quality and quantity of milk and to the treatment losses for staphylococcal mastitis. Furthermore, S. aureus can contaminate both raw milk and dairy products, being a source of food poisoning (1, 2).
The prevalence of S. aureus in dairy goat herds, estimated by analyzing bulk tank milk, varies between 16.7 and 96.2% in different countries (3–5). In Italy, the prevalence of S. aureus in dairy goat herds was reported to be 43.1% in the Lombardy Region (6) and 76.9% in Sardinia (7).
Ribosomal Spacer PCR (RS-PCR) is an accurate, rapid, inexpensive genotyping method with high discriminatory power (8). It has been used to classify S. aureus isolates into genotypes, that are inferred from the electrophoresis profile using the Mahal software, according to Fournier et al. (9) and Graber (10). In particular, an electrophoretic profile differing in more than one band from all identified genotypes is considered as a new genotype. Genotypes are named and extended leading to the genotypes GTA to GTZ, followed by the genotypes GTAA to GTAZ, GTBA to GTBZ, and so on. Genotypic variants, which differ in only one band from the genotype after the electrophoretic analysis, were indicated with roman superscripts (e.g., GTRI) (10). RS-PCR genotypes, in turn, can be grouped into clusters (CL), each of which consists of a genotype and all its variants (e.g., GTR and GTRI, GTRII, GTRIII, etc.).
Several studies performed RS-PCR to genotype S. aureus strains isolated from bovine milk, and showed a great genotypic variety, differing in their contagiousness (8, 9, 11, 12). S. aureus CLB was found to be highly contagious and has been associated with high within-herd prevalence of intramammary infections (IMI), together with CLR and CLS (8, 9, 12). In contrast, S. aureus CLC appears to be less problematic because it generally affects individual cows and only one quarter of the mammary gland is affected (11, 13). Moreover, each genotype presented a different pattern of virulence genes (9, 14, 15). In spite of this detailed knowledge on dairy-associated S. aureus genotypes, no data on S. aureus genotypes isolated from goats is currently available.
Previous studies suggest that the lineages CC130, CC133, and CC522 represent the major CC among S. aureus isolates from milk of small ruminants (4, 15–17). S. aureus isolates from caprine and ovine hosts have been spa typed previously (15–18). In goats and sheep, fewer spa types were found compared to cows (15). Merz et al. (17) reported t1773 (corresponding to CC130) as the most prevalent spa type in caprine isolates.
S. aureus produces a wide range of virulence factors, which are essential for a successful infection. At least 25 different toxins (e.g., enterotoxins SEA to SEQ, toxic shock syndrome toxin-1 tst1, exfoliative toxins ETA and ETB), 15 microbial surface components recognizing adhesive matrix molecules (such as clumping factor A clfA), 20 immune evasion molecules (such as protein A, coagulase, hemolysins, and leukocidins), among others are known S. aureus virulence factors (19). The most prevalent enterotoxin genes detected in S. aureus isolated from small ruminants were sec and sel, whereas the sea gene was found exclusively in caprine isolates (15, 17). Merz et al. (17) reported that all the isolates harboring the tst1 gene also harbored the combination of genes sec-sel and were assigned exclusively to CC130 and CC133. Significantly higher prevalence rates of sec, sel, and tst1 genes were observed in S. aureus from small ruminants than among bovine isolates (15, 17).
While the population structure and the genomic characteristics of S. aureus isolated from bovine milk are very well-described, similar data on caprine S. aureus is very limited. Therefore, this study aimed at defining RS-PCR genotypes, clonal complexes (CC), spa types, and virulence gene profiles of S. aureus isolated from goat milk in the Lombardy region in northern Italy.
S. aureus isolated from goat bulk tank milk (BTM) collected by Cortimiglia et al. (6) was used for this study, thus the sampling criteria and bacteriological analysis have already been described. Briefly, BTM samples were collected from 197 different dairy goat farms located in Lombardy (northern Italy) between July and October 2012. After collection, milk samples were transported at +4°C to the laboratory and stored at −20°C till analysis, which were conducted within 3 months. Bacteriological analysis was performed with standard techniques for S. aureus isolation, as previously described by Cortimiglia et al. (6). In particular, milk samples were spread on blood agar supplemented with 5% defibrinated sheep blood and on Baird Parker agar supplemented with rabbit plasma fibrinogen (BP-RPF; Oxoid Ltd., Basingstoke, UK), plates were incubated at 37°C and analyzed after 24 and 48 h. Colonies developing a typical coagulase halo on BP-RPF agar were considered suspected S. aureus, as were hemolytic colonies on BA testing positive in a tube coagulase test (Coagulase plasma-EDTA, Biolife srl, Milan, Italy).
DNA was extracted using the DNA isolation system kit (Clonit, Medical System, Genova, Italy) according to the manufacturer's guidelines. DNA quality and quantity were measured using a NanoDrop ND-1000 spectrophotometer (Nano-Drop Technologies, Wilmington, DE), and stored at −20°C till further use.
If present, five S. aureus isolates from each BTM positive sample were genotyped by RS-PCR, according to Fournier et al. (9) and Graber (10). When different cultural morphologies were detected (pigmentation and hemolysis on blood agar or coagulase and lecitinase halos on Baird Parker agar), four colonies per morphology were selected in each sample.
The PCR products were analyzed using the miniaturized electrophoresis system DNA 7500 LabChip [Agilent Technologies, Santa Clara, CA; (9, 10)]. Genotypes were inferred from the electrophoresis profile using the Mahal software, which is freely available online (10)1.
Typing of spa was performed on a subset of strains, representing all the S. aureus RS-PCR genotypes found in each milk sample (i.e., herd). In detail, one or more isolates were selected from each BTM sample, depending on the number of different genotypes identified with the RS-PCR. Typing was performed according to Shopsin et al. (20), and spa types were assigned using the code system described on Ridom SpaServer2.
To depict the frequency and genetic relatedness among the isolates, the spa typing results were analyzed, and a minimum spanning tree (MST) was constructed using the BioNumerics 7.6 software (Applied Maths, Sint-Martens-Latem, Belgium).
PCR amplifications were performed on the same strains selected for spa typing to investigate the presence of 22 virulence factors contributing to S. aureus pathogenicity: enterotoxins (from sea to see, and from seg to sel) (21, 22), toxic shock syndrome (tst) and exfoliative (eta, etb) toxins (21), PVL (lukS-lukF) (23), leucocidin M (lukM), and leukotoxin ED (lukE-lukD) (24), cell-wall associated protein clfA (clumping factor A) (21), cna (collagen-binding protein) (25), and fmtB (cell-wall protein), scn, chp, and sak belonging to the immune evasion cluster (IEC) (26). The primers and protocols are listed in the Supplementary Table.
Multi Locus Sequence Typing (MLST) was performed as previously described (27) on one representative strain for each combination of spa typing and RS-PCR genotyping result. Allele numbers and sequence type (ST) were assigned as per the MLST database3 Clonal analysis of the STs was performed using the clustering algorithm e-BURST4
An evolutionary analysis was carried out using goeBURST5 (Algorithm 3.0), a java implementation of the eBURST algorithm rules proposed by Feil et al. (28), using a graphic matroid approach that ensures an optimal solution for the placement of links between STs.
The degree of similarity between the distributions of RS-PCR genotypes among caprine strains and bovine S. aureus strains in the same geographical area and during the same time period (12) was estimated using the Czekanowski index or Proportional Similarity Index (PSI). It is calculated by
in which pi and qi represent the proportion of genotypes in the two distinct populations (caprine and bovine S. aureus strains), while i represents the total numerosity of the two populations. The values for PSI range from 1 for identical frequency distributions of the variable of interest to 0 for no similarities between the data sets (29).
A total of 295 S. aureus isolates from 65 out of 197 examined goat BTM samples were genotyped by RS-PCR. The resulting genotypes with their variants were grouped into clusters, obtaining 29 genotypes grouped into 18 clusters (Table 1). A single genotype was detected in 43 samples (66.2%), whereas 2 and 3 different genotypes were identified in 18 (27.7%) and 4 (6.2%) samples, respectively. The analyzed isolates were grouped into 7 major CL, in the following order of prevalence: CLR (18/65; 27.7%), CLAA (16/65; 24.6%), CLZ (13/65; 20%); CLAW (11/65; 16.9%); CLBW (9/65; 13.8%), CLS (7/65; 10.8%), and CLI (2/65; 3.1%). Each one of the remaining 11 minor CL was isolated in only one sample.
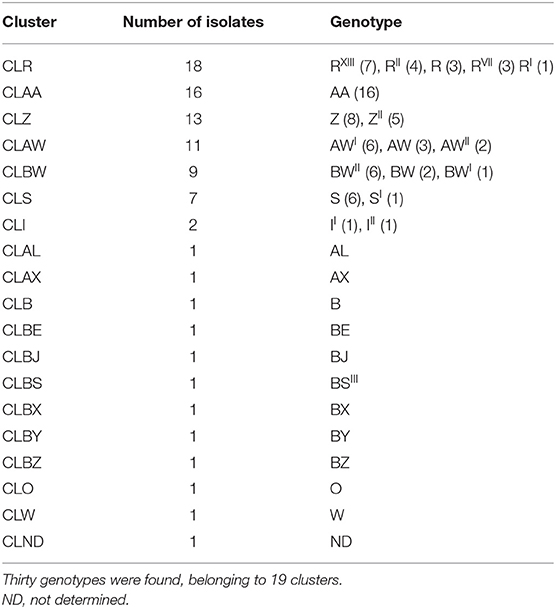
Table 1. RS-PCR genotypes and genotypic clusters of Staphylococcus aureus isolated from goat bulk tank milk samples, sorted by frequency.
Typing of spa was performed on a subset of 88 strains. A total of 26 different spa types were detected and the most prevalent types were t1773 (n = 21; 23.9%), t5428 (n = 20; 22.7%), and t2678 (n = 11; 12.5%). Other spa types, such as t7630, t899 (n = 4; 4.5%), t1180, t988 (n = 3; 3.4%), and t524, t13986 (n = 2; 2.3%), were less frequent, and each of the remaining 17 spa types was detected only once (Table 2).
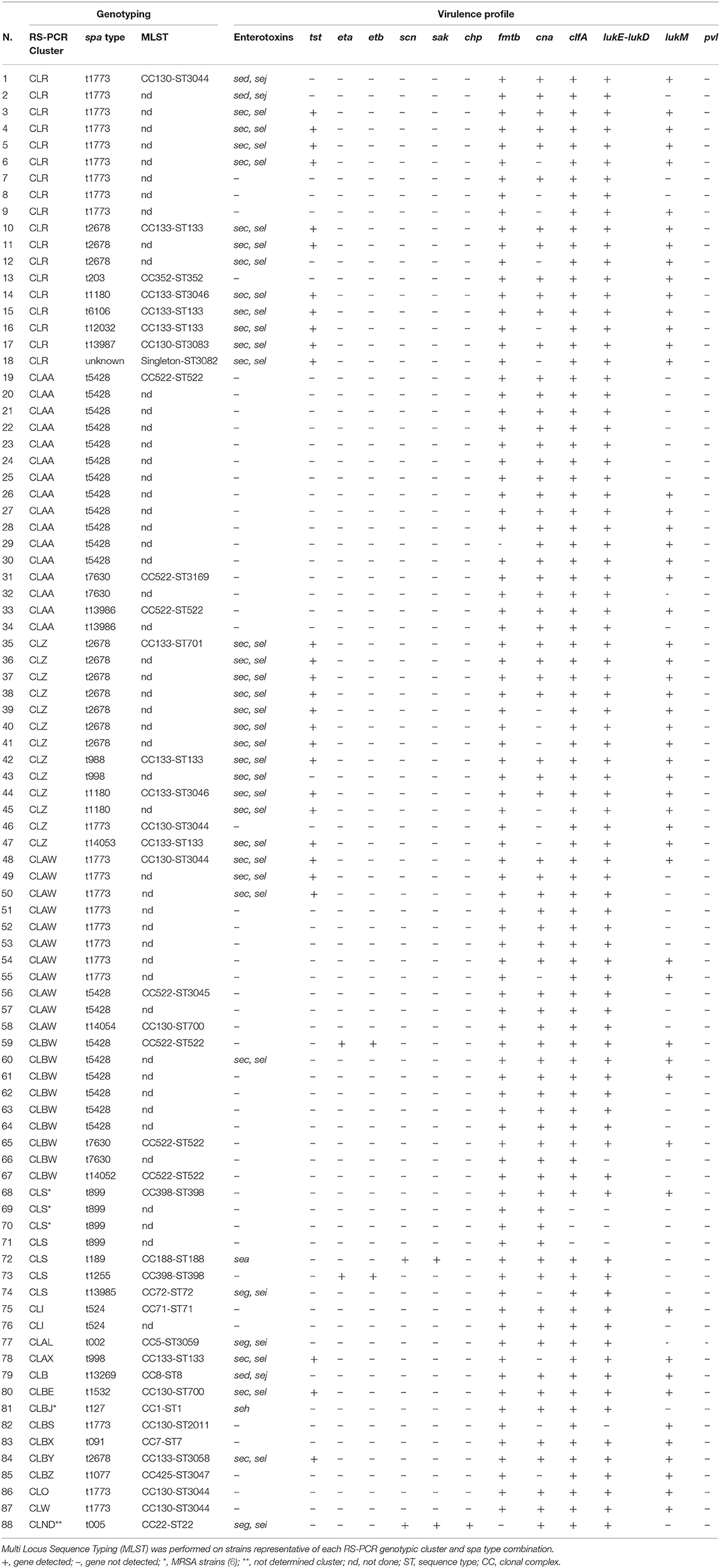
Table 2. Molecular characteristics of the 88 Staphylococcus aureus strains isolated from goat bulk tank milk.
Using the observed spa types, a MST was constructed. MST revealed 3 major spa types (t1773, t2678, and t5428) that were linked to a few secondary and tertiary spa types. In between there was a set of rarely observed spa types, some of them were involved in connecting the 3 main types (Figure 1).
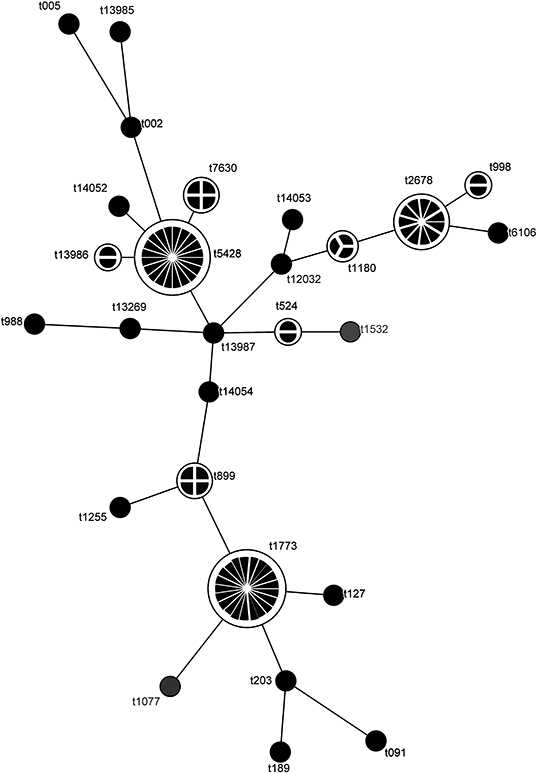
Figure 1. Minimum spanning tree for Staphylococcus aureus strains isolated from goat milk. The analysis was based on spa typing results and elaborated with BioNumerics 7.6 (Applied Maths, Sint-Martens-Latem, Belgium). The size of the circles and the number of sections reflect the numerosity of each spa type.
An overview of the prevalence of the most important virulence genes carried by the 88 considered strains is provided in Table 2. Overall, 44.3% (n = 39) of all isolates harbored at least one enterotoxin gene. The most frequent was the combination sec-sel, which was present in 35.2% (n = 31) of the isolates, followed by sed-sej (n = 3; 3.4%), seg-sei (n = 3; 3.4%), sea (n = 1; 1.1%), and seh (n = 1; 1.1%). The molecular analysis showed that tst, eta, and etb genes were detected in 31.8, 2.3, and 2.3% of the isolates, respectively. Most of the isolates was positive for genes related to host adhesion and invasion, such as fmtB (n = 86; 98%), clfA (n = 85; 96.6%), and cna (n = 71; 80.6%), whereas few isolates carried genes of the immune evasion cluster (scn, n = 2, 2.3%; sak, n = 2, 2.3%; chp, n =1, 1.1%). Genes encoding lukE-lukD and lukM were observed in 94.3% (n = 83) and 60.2% (n = 53) of isolates, respectively. None harbored the gene encoding Panton-Valentine leukocidin.
Thirty-nine isolates were selected to perform a MLST analysis. The isolates were assigned to 23 different ST, which belonged to 14 different CC. One ST was a singleton (Table 2). The most frequent CC were CC133 (n = 10; 25.6%), CC130 (n = 9; 23.1%), and CC522 (n = 7; 17.9%). CC398 was detected in 5.1% of isolates, whereas each of the remaining CC was assigned to only one isolate.
The evolutionary relationships between the different goat S. aureus strains are displayed in Figure 2. Three central ST were observed (ST133, ST522, and ST700), corresponding to CC133, CC522, and CC130, respectively. These clones are the founders of additional single locus variants (Figure 2), reflecting a further evolution away from the central clones. All the other ST/clones remain unrelated.
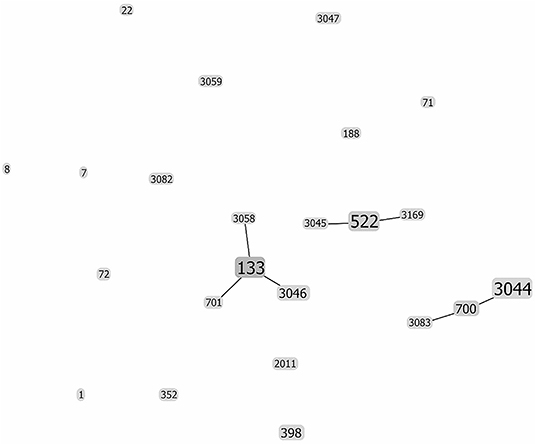
Figure 2. Evolutionary relationship between the different ST of S. aureus strains isolated from goat milk. The analysis was performed with goeBURST (Algorithm 3.0), which predicts the founding (ancestral) genotype of each ST. The size of each square reflects the number of strains. The ST located in the periphery of the founder CC are the corresponding single locus variants. The remaining STs are unrelated.
PSI was estimated by comparing the RS-PCR genotypes of the strains (18 different S. aureus CL), isolated from the 65 caprine BTM samples in our study, with the 471 strains (44 different CL) isolated from 398 bovine BTM samples in a recent study (12). The index was 0.311, indicating a very low correlation between the distribution of caprine and bovine S. aureus genotypes.
Several studies used RS-PCR to genotype S. aureus strains isolated from bovine milk, yet no information on S. aureus from goat milk is currently available. For the first time, we genotyped via RS-PCR 295 S. aureus isolates, which grouped in 18 genotypic clusters. The major S. aureus CL have also been detected in bovine milk, except for CLAW. Other minor CL were here identified and reported for the first time, such as CLW, CLAL, CLBS, and CLBX. Previous studies demonstrated that S. aureus CLB, which is the most prevalent among bovine isolates, is highly contagious and associated with high within-herd prevalence of IMI (8, 11, 12). On the contrary, we isolated S. aureus CLB in a single BTM from a dairy goat farm located in the province of Sondrio. Furthermore, S. aureus CLR and CLS have also been reported to be associated with high within-herd prevalence of IMI (8, 12). S. aureus CLR (27.7%) was the most frequently isolated among the analyzed herds. Albeit the lack of details about the sampled farms, we cannot exclude the possibility that some of these farms reared goats and cattle in close proximity, facilitating the spread between the two animal populations.
Most of the isolates were assigned to CC133, CC130, and CC522, so that these lineages may be the major CC among caprine S. aureus isolates in Italy as well. Our results are in agreement with previous studies reporting the predominance of these CC in S. aureus isolated from milk of small ruminants (4, 15–17).
Typing of spa showed a remarkable heterogeneity among S. aureus isolates. The most prevalent spa types were t1773, t5428, and t2678, corresponding to CC130, CC522, and CC133, respectively, as already reported (16, 17, 30). Comparing RS-PCR genotypes to spa types, frequently Staph. aureus belonging to CLAW (73%) or CLR (50%), especially to its variants RIII and RVII, corresponded to t1773. Similarly, S. aureus CLAA (75%) or CLBW (67%) corresponded to t5428.
Several studies consider S. aureus from goats and sheep to be part of the same population based on CC, since they show pronounced adaptation to small ruminants, but not especially to sheep or goats (17, 30). Nevertheless, different S. aureus spa types are reported to be associated with the two species. For example, among CC133 S. aureus, spa types t544, t3583, and t7304 were predominantly associated with caprine mastitis, whereas t2678 and t9088 with ovine mastitis (16, 18, 30). However, these spa types are not exclusively associated with one host species (30). Actually, CC133/t2678 was one of the most prevalent spa types among our strains, whereas we did not find any CC133/t544 or CC133/t3585. Previous studies have described CC133/t544 isolates in sheep as well (16, 18). These contrasting results support the idea that S. aureus lineages found among sheep and goats are highly similar (17, 31) and this might be due to the fact that in many countries sheep and goats are farmed together, as previously suggested by (31).
Bar-Gal et al. (15) demonstrated that S. aureus isolates from small ruminants differ from the bovine strains in their virulence gene patterns. In fact, cow isolates showed a higher rate of the tested virulence genes than goat and sheep isolates. In our study, 44.3% of all isolates carried at least one enterotoxin gene. All isolates harboring tst also carried the genes sec and sel, and were assigned exclusively to CC130 and CC133, in agreement with previous findings (17, 32). These genes are located on the S. aureus pathogenicity island SaPIov1 (33). Genes coding for toxins usually associated with human diseases, such as eta and etb (responsible for the staphylococcal scalded-skin syndrome), or pvl (leukocidin produced by S. aureus that causes leukocyte destruction and tissue necrosis) were rarely or never detected. Moreover, we found high prevalence of leukocidins or genes related to host adhesion and invasion, suggesting their important role as virulence factors for S. aureus strains in small ruminants.
The comparison between S. aureus strains isolated from goat and bovine milk showed that S. aureus from goat milk have wide genetic variability and differ from the bovine strains.
Caprine S. aureus is characterized by 3 major spa types and a set of rarely observed types, representing a limited genetic variation among caprine strains. Except for t002 and t005, all the observed spa are observed frequently, yet are rarely present in the spa type database [Ridom SpaServer (see text footnote 2)] implying some uniqueness of the strains analyzed in the present study. Indeed, except for t002 and t005, all remaining spa types do not match with those typically observed for human and bovine strains (34, 35). We suspect that t002 and t005 are likely a result of human milk contamination that can occasionally occur during milking and/or equipment cleaning operations. The most prevalent human spa types include t032, t003, t002, t008, t011, and t127 (see text footnote 2), whereas those most often observed in bovine milk include t529, t2953, t1403, t524, t024, and t164 (35).
Our study supports the idea that S. aureus from small ruminants constitutes a distinct population in northern Italy, in agreement with previous findings reported worldwide. It also provides for the first time insights to the S. aureus RS-PCR genotypes circulating in goat herds in this area, allowing a comparison to bovine strains and adding useful data for epidemiological studies. Further information about S. aureus strains isolated from small ruminants is required, as they are a major problem for animal health and may also represent a serious concern for human health, as many dairy products are made from raw milk.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Staphylococcus aureus is the major pathogen responsible for intramammary infections in goats, causing severe economic losses in dairy farms. Furthermore, it can contaminate milk and dairy products, being a source of food poisoning. Since data on the genetic traits and population structure of S. aureus from goats are still very limited, our work aimed at determining genetic lineages and virulence traits of S. aureus isolated from goat milk in northern Italy. Previous studies used the rapid screening method RS-PCR to genotype S. aureus strains from bovine milk, showing a great variety of genotypes differing in their contagiousness. For the first time, we performed RS-PCR on S. aureus isolated from goat milk, in addition to more-recognized genotyping methods, such as spa typing, MLST, and we investigated their virulence profiles. S. aureus isolates showed a wide genetic variability and they differed from bovine strains. Our results support the idea that S. aureus from goats may constitute a distinct population, in agreement with previous findings reported worldwide. Our study provides for the first time an overview of the S. aureus RS-PCR genotypes circulating in goat herds in northern Italy, adding useful data for further epidemiological studies.
ML conceived and planned the experiments. AR, VB, CC, PC, and AG carried out the experiments. AM and FV contributed to the experiments and the interpretation of the results. HG and AR performed the statistical and population study analyses. AG and AR wrote the manuscript in consultation with ML and PC. All authors discussed the results, critically revised, and approved the final manuscript.
This work was funded by Italian Ministry of Health, Ricerca Corrente IZSLER 2010/03.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank Lorenza Sala and Francesca Pozzi (Istituto Zooprofilattico della Lombardia e Dell'Emilia Romagna, Lodi, Italy) for the valuable technical assistance in bacteriological and PCR analysis, Valentina Gualdi (Genomic Platform of Parco Tecnologico Padano, Lodi, Italy) for sequencing, and the Lombardy Regional Breeder Association (ARAL) for providing the BTM samples and the data on the farms.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2020.00628/full#supplementary-material
1. Akineden O, Hassan AA, Schneider E, Usleber E. Enterotoxigenic properties of Staphylococcus aureus isolated from goats' milk cheese. Int J Food Microbiol. (2008) 124:211–6. doi: 10.1016/j.ijfoodmicro.2008.03.027
2. Johler S, Giannini P, Jermini M, Hummerjohann J, Baumgartner A, Stephan R. Further evidence for staphylococcal food poisoning outbreaks caused by egc-encoded enterotoxins. Toxins. (2015) 7:997–1004. doi: 10.3390/toxins7030997
3. Muehlherr JE, Zweifel C, Corti S, Blanco JE, Stephan R. Microbiological quality of raw goat's and ewe's bulk-tank milk in Switzerland. J Dairy Sci. (2003) 86:3849–56. doi: 10.3168/jds.S0022-0302(03)73992-7
4. Jørgensen HJ, Mørk T, Høgåsen HR, Rørvik LM. Enterotoxigenic Staphylococcus aureus in bulk milk in Norway. J Appl Microbiol. (2005) 99:158–66. doi: 10.1111/j.1365-2672.2005.02569.x
5. Chu C, Yu C, Lee Y, Su Y. Genetically divergent methicillin-resistant Staphylococcus aureus and sec-dependent mastitis of dairy goats in Taiwan. BMC Vet Res. (2012) 8:39. doi: 10.1186/1746-6148-8-39
6. Cortimiglia C, Bianchini V, Franco A, Caprioli A, Battisti A, Colombo L, et al. Short communication: Prevalence of Staphylococcus aureus and methicillin-resistant S. aureus in bulk tank milk from dairy goat farms in Northern Italy. J Dairy Sci. (2015) 98:2307–11. doi: 10.3168/jds.2014-8923
7. Spanu V, Scarano C, Virdis S, Melito S, Spanu C, De Santis E. Population structure of Staphylococcus aureus isolated from bulk tank goat's milk. Foodborne Pathog Dis. (2013) 10:310–5. doi: 10.1089/fpd.2012.1356
8. Cremonesi P, Pozzi F, Raschetti M, Bignoli G, Capra E, Graber HU, et al. Genomic characteristics of Staphylococcus aureus strains associated with high within-herd prevalence of intramammary infections in dairy cows. J Dairy Sci. (2015) 98:6828–38. doi: 10.3168/jds.2014-9074
9. Fournier C, Kuhnert P, Frey J, Miserez R, Kirchofer M, Kaufmann T, et al. Bovine Staphylococcus aureus: association of virulence genes, genotypes and clinical outcome. Res Vet Sci. (2008) 85:439–88. doi: 10.1016/j.rvsc.2008.01.010
10. Graber HU. Genotyping of Staphylococcus Aureus by ribosomal spacer PCR (RS-PCR). J Vis Exp. (2016) 117:54623. doi: 10.3791/54623
11. Cosandey A, Boss R, Luini M, Artursson K, Bardiau M, Breitenwieser F, et al. Staphylococcus aureus genotype B and other genotypes isolated from cow milk in European countries. J Dairy Sci. (2016) 99:529–40. doi: 10.3168/jds.2015-9587
12. Gazzola A, Maisano AM, Bianchini V, Vezzoli F, Romanò A, Graber HU, et al. Short communication: characterization of Staphylococcus aureus from bulk tank milk of dairy cattle in Lombardy (northern Italy). J Dairy Sci. (2020) 103:2685–92. doi: 10.3168/jds.2019-17181
13. Graber HU, Bodmer M. Staphylococcus aureus and its genotypes as a mastitis pathogen in dairy cattles - a review. Schweiz Arch Tierheilkd. (2019) 161:611–7. doi: 10.17236/sat00223
14. Graber HU, Naskova J, Studer E, Kaufmann T, Kirchofer M, Brechbühl M, et al. Mastitis-related subtypes of bovine Staphylococcus aureus are characterized by different clinical properties. J Dairy Sci. (2009) 92:1442–51. doi: 10.3168/jds.2008-1430
15. Bar-Gal GK, Blum SE, Hadas L, Ehricht R, Monecke S, Leitner G. Host-specificity of Staphylococcus aureus causing intramammary infections in dairy animals assessed by genotyping and virulence genes. Vet Microbiol. (2015) 176:143–54. doi: 10.1016/j.vetmic.2015.01.007
16. Porrero MC, Hasman H, Vela AI, Fernández-Garayzábal JF, Domínguez L, Aarestrup FM. Clonal diversity of Staphylococcus aureus originating from the small ruminant goats and sheep. Vet Microbiol. (2012) 156:157–61. doi: 10.1016/j.vetmic.2011.10.015
17. Merz A, Stephan R, Johler S. Staphylococcus aureus isolates from goat and sheep milk seem to be closely related and differ from isolates detected from bovine milk. Front Microbiol. (2016) 7:319. doi: 10.3389/fmicb.2016.00319
18. Eriksson J, Espinosa-Gongora C, Stamphøj I, Larsen AR, Guardabassi L. Carriage frequency, diversity and methicillin resistance of Staphylococcus aureus in Danish small ruminants. Vet Microbiol. (2013) 163:110–5. doi: 10.1016/j.vetmic.2012.12.006
19. Monistero V, Graber HU, Pollera C, Cremonesi P, Castiglioni B, Bottini E, et al. Staphylococcus aureus isolates from bovine mastitis in eight countries: genotypes, detection of genes encoding different toxins and other virulence genes. Toxins. (2018) 10:247. doi: 10.3390/toxins10060247
20. Shopsin B, Gomez M, Montgomery SO, Smith DH, Waddington M, Dodge DE, et al. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J Clin Microbiol. (1999) 37:3556–63.
21. Akineden O, Annemüller C, Hassan AA, Lämmler C, Wolter W, Zschock M. Toxin genes and other characteristics of Staphylococcus aureus isolates form milk of cows with mastitis. Clin And Diagn Lab Immu. (2001) 2001:959–64. doi: 10.1128/CDLI.8.5.959-964.2001
22. Cremonesi P, Luzzana M, Brasca M, Morandi S, Lodi R, Vimercati C, et al. Development of a multiplex PCR assay for the identification of Staphylococcus aureus enterotoxigenic strains isolated from milk and dairy products. Mol Cell Probes. (2005) 19:299–305. doi: 10.1016/j.mcp.2005.03.002
23. Lina G, Piémont Y, Godail-Gamot F, Bes M, Peter MO, Gauduchon V, et al. Involvement of panton-valentine leukocidin–producing Staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis. (1999) 29:11281132. doi: 10.1086/313461
24. Jarraud S, Mougel C, Thioulouse J, Lina G, Meugnier H, Forey F, et al. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun. (2002) 70:631–41. doi: 10.1128/iai.70.2.631-641.2002
25. Zecconi A, Binda E, Borromeo V, Piccinini R. Relationship between some Staphylococcus aureus pathogenic factors and growth rates and somatic cell counts. J Dairy Res. (2005) 72:203–8. doi: 10.1017/s0022029905000841
26. Sung JM-L, Lloyd DH, Lindsay JA. Staphylococcus aureus host specificity: comparative genomics of human vs. animal isolates by multi-strain microarray. Microbiology. (2008) 154:1949–59. doi: 10.1099/mic.0.2007/015289-0
27. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. (2000) 38:1008–15. doi: 10.1128/JCM.38.3.1008-1015.2000
28. Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol. (2004) 186:1518–30. doi: 10.1128/JB.186.5.1518-1530.2004
29. Feinsinger P, Spears EE, Poole RW. A simple measure of niche breadth. Ecology. (1981) 62:27–32. doi: 10.2307/1936664
30. Hoekstra J, Rutten VPMG, van den Hout M, Spaninks MP, Benedictus L, Koop G. Differences between Staphylococcus aureus lineages isolated from ovine and caprine mastitis but not between isolates from clinical or subclinical mastitis. J Dairy Sci. (2019) 102:5430–7. doi: 10.3168/jds.2018-16196
31. Smith EM, Needs PF, Manley G, Green LE. Global distribution and diversity of ovine-associated Staphylococcus aureus. Infect Genet Evol. (2014) 22:208–15. doi: 10.1016/j.meegid.2013.09.008
32. Ben Said M, Abbassi MS, Bianchini V, Sghaier S, Cremonesi P, Romanò A, et al. Genetic characterization and antimicrobial resistance of Staphylococcus aureus isolated from bovine milk in Tunisia. Lett Appl Microbiol. (2016) 63:473–81. doi: 10.1111/lam.12672
33. Guinane CM, Ben Zakour NL, Tormo-Mas MA, Wienert LA, Lowder BV, Cartwright RA, et al. Evolutionary genomics of Staphylococcus aureus reveals insights into the origin and molecular basis of ruminant host adaptation. Genome Biol Evol. (2010) 2:454–66. doi: 10.1093/gbe/evq031
34. Grundmann H, Aanensen DM, van den Wijngaard CC, Spratt BG, Harmsen D, Friedrich AW, et al. Geographic distribution of Staphylococcus aureus causing invasive infections in Europe: a molecular-epidemiological analysis. PLoS Med. (2010) 7:e1000215. doi: 10.1371/journal.pmed.1000215
Keywords: Staphylococcus aureus, mastitis, goat, small ruminants, genotyping, virulence gene profile, Italy
Citation: Romanò A, Gazzola A, Bianchini V, Cortimiglia C, Maisano AM, Cremonesi P, Graber HU, Vezzoli F and Luini M (2020) Staphylococcus aureus From Goats Are Genetically Heterogeneous and Distinct to Bovine Ones. Front. Vet. Sci. 7:628. doi: 10.3389/fvets.2020.00628
Received: 06 June 2020; Accepted: 03 August 2020;
Published: 09 September 2020.
Edited by:
Jorge Blanco, University of Santiago de Compostela, SpainReviewed by:
Jesús Santos, Universidad de León, SpainCopyright © 2020 Romanò, Gazzola, Bianchini, Cortimiglia, Maisano, Cremonesi, Graber, Vezzoli and Luini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alessandra Gazzola, YWxlZ2F6em9sYUBob3RtYWlsLml0
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.