
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Vet. Sci. , 27 August 2020
Sec. Veterinary Infectious Diseases
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.00567
 Manrico Sebastiano1*
Manrico Sebastiano1* Daniele Canestrelli2
Daniele Canestrelli2 Roberta Bisconti2
Roberta Bisconti2 Anne Lavergne3
Anne Lavergne3 Kévin Pineau4
Kévin Pineau4 Olivier Chastel1
Olivier Chastel1 Vincent Lacoste3,5
Vincent Lacoste3,5 David Costantini6
David Costantini6Since 2005, we have recorded annual episodes of alphaherpesvirus outbreaks in chicks of magnificent frigatebird Fregata magnificens on the Ile du Grand Connétable Nature Reserve in French Guiana. In 2009, we found sooty terns, Onychoprion fuscatus, that live sympatrically with frigatebirds, with visible clinical signs of a potential viral infection. To determine if the symptoms observed in sooty terns could be associated with an alphaherpesvirus previously identified in frigatebirds, we carried out molecular screening of samples collected from seven individuals. We identified and characterized a novel viral sequence from five birds. BLAST searches, pairwise nucleotide, and amino acid sequence comparisons, as well as phylogenetic analyses confirmed that the sequence belonged to the Herpesviridae family, of the Alphaherpesvirinae subfamily. We observed that it clustered with strains isolated from Podargidae (Caprimulgiformes), Columbiformes, and Falconiformes, but was distinct from the frigatebird herpesvirus. We have tentatively named it Onychoprion fuscatus alphaherpesvirus 1, (OfusAHV1). These two sequences, although found syntopic on the Ile du Grand Connétable, belong to two distinct alphaherpesvirus strains. Thus, the clinical symptoms showed by sooty terns do not likely result from a cross-species transmission event. Future work is needed to better characterize the virus and to investigate herpesvirus prevalence in healthy, free-ranging sooty terns, and to assess the impact of the virus on population viability.
Herpesviruses are DNA viruses found in many animal species, from invertebrates to mammals (1). Herpesviruses are thought to have evolved in association with their hosts. However, some studies reported cases of cross-species transmission, indicating that such events could occur more frequently than previously thought (2–4). These “spillover” infections in alternative hosts can result in dramatic outbreaks of disease (5–7). Because of their ability to establish a latent infection, herpesviruses do not generally pose a threat to their host species. However, some viruses can cause severe diseases and induce high mortality rates in their natural hosts (8, 9). This is the case for avian herpesviruses that remain one of the major causes of fatal infectious diseases in many bird species (10–12).
In 2005, we found several chicks of magnificent frigatebird Fregata magnificens on the Ile du Grand Connétable Nature Reserve (4°49' 36” N, 51°56' 38” W), a rocky island located off the coast of French Guiana, that showed clinical cutaneous signs or were found dead (13). In particular, chicks showed nodular proliferative skin lesions in legs and in the neck, and hyperkeratosis (13). A few years later, we characterized a novel alphaherpesvirus sequence from those chicks (13). In the following years, we have started a monitoring program of the population of frigatebirds, and have found that the disease is widespread in chicks, causing a number of physiological alterations associated with a high mortality rate (14–16). Since the first appearance of clinical signs in frigatebirds, we have also started annual monitoring programs for the other species that breed sympatrically in the natural reserve. On the 30th of April 2009, we found several dead or dying adult sooty terns Onychoprion fuscatus showing similar clinical signs of frigatebirds (bone frailty, hyperkeratosis) as described previously (13, 16, 17). Our goal was to determine if the observed symptoms could be due to a cross-species transmission of the alphaherpesvirus that affect magnificent frigatebirds or the results of an infection with an unknown herpesvirus.
To determine if the observed symptoms could be due to a cross-species transmission we collected biological material (i.e., tracheal swabs and blood) from sick birds, while small tissue samples (i.e., trachea, brain, lungs, liver, and heart) from dead birds were additionally collected. Trachea (3 samples), brain (6 samples), lung (1 sample), liver (4 samples) heart (1 sample), and whole blood (4 samples) for a total of 19 samples were collected and placed in 2 mL Eppendorf tubes. Blood was centrifuged in the field and all samples were subsequently frozen in dry ice while in the field and were then kept in a −80°C freezer until laboratory analyses.
We extracted DNA by a classical phenol, phenol-chloroform (1:1 vol/vol), and chloroform technique and precipitated it by isopropanol. Then, we washed the DNA with 70% ethanol and resuspended it in TE buffer containing 10 mM Tris (pH 8.0) and 1 mM EDTA. We carried out molecular screening by semi-nested PCR amplifications with degenerate consensus primers targeting highly conserved amino acid motifs of the herpesvirus DNA polymerase gene. To this end, we used two sets of primers [First set: Freg1F: GTGTTCGATTTTGCCAGCCTGTATCC, Freg1R: ATGTTCCTTCCTATGGTCGTTACC, Freg2R: ACGTGCAGACACGGCAGAAG; Second set: as explained in (18)] targeting the same region of the gene, but with different levels of degeneracy. This was done for each DNA sample in separate reactions for the first-round PCR (Freg1F/Freg1R or DFASA/GDTD1B) and second-round PCR (Freg1F/Freg2R or VYGA/GDTD1B). The initial round of PCR contained 500 ng of genomic DNA, 30 pmoles of degenerate primers, 2 mM MgCl2, 0.2 mM each dNTP, 5 μL of 10 × PCR buffer, and 0.5 μL of AmpliTaq Gold DNA polymerase in a volume of 50 μL. We used 2 μL of this reaction in the semi-nested reaction. The PCR cycling conditions were as follows: after the DNAs were denaturated at 94°C for 10 min, the reaction mixtures were cycled five times at 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s, followed by 30 cycles at 94°C for 30 s, 46°C for 30 s, and 72°C for 30 s. We made an extension of 10 min at 72°C on the last cycle (GeneAmp PCR system 9600 thermal cycler; Perkin-Elmer). Amplification products of the expected size (about 250 and 350 base pair, respectively) were cloned into pCR4-TOPO vectors using a TA cloning kit from Invitrogen and sent them for sequencing to Genewiz (https://www.genewiz.com/). For each PCR product, three clones of the “screening amplicons” were sequenced on both strands.
Raw sequences were analyzed and edited in MEGA 5.05 (19). The nucleotide sequence was 307 bp in size, excluding primers. We then carried out sequence homology analyses using the BLAST program at the National Center of Biotechnology Information (NCBI) (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Then, a multiple sequence alignment was constructed using ClustalW with previously published avian herpesvirus sequences and representative sequences for each genus or subfamily retrieved from GenBank (http://www.ncbi.nlm.nih.gov/nucleotide) (13, 20). The alignment was checked manually.
We analyzed the phylogenetic relationships among herpesviruses using the Bayesian inference (BI) approach implemented in Beast 1.8.4 (21), based on a final alignment including 37 unique sequences of 104 amino acid positions. We assessed the best-fit model of amino acid evolution for the dataset using the smart model selection (SMS) approach (22) as implemented in the PhyML environment (23). We ran BI analyses with LG+G+I model of aminoacidic substitution (24), an uncorrelated relaxed molecular clock model with a log-normal distribution (25), and a Yule tree prior. Then, we analyzed the results from two independent runs of 10 million generations, sampled every 1,000 generations, using Tracer 1.7.1 to check that the effective sample sizes for all parameters that exceeded 200 (26) and to assess the appropriate number of initial trees to discard as burn-in. Then, we combined the two runs using Logcombiner 1.8.4 [BEAST package, (27)]. We computed the Maximum Clade Credibility (MCC) tree summarizing the post-burn-in trees using TreeAnnotator 1.8.4, and we visualized the tree using FigTree 1.4.4 (26).
This study aimed at assessing the presence of a herpesvirus from dead and dying adult sooty terns and to determine its relationship with other members of the Herpesviridae family. Out of the 19 tissue samples, 7 samples (4 blood samples and 3 brain samples, collected from both dead and dying birds) tested positive for herpesvirus. A unique and novel viral sequence was obtained from five out of the seven individuals with PCR positive results. This sequence is tentatively designated as Onychoprion fuscatus alphaherpesvirus 1 (OfusAHV1) in the Alphaherpesvirinae subfamily. From a phylogenetic perspective, OfusAHV1 clustered with strains detected from Podargidae (Caprimulgiformes), Columbiformes, and Falconiformes (posterior probability = 0.86; Figure 1). We also found that this novel alphaherpesvirus sequence of sooty terns was distinct from the Frigatebird herpesvirus (nucleotide p-distance: 0.28), which belonged to a different well-supported monophyletic lineage. These results seem to indicate that no cross-species transmission has occurred between frigatebirds and sooty terns. This is not surprising given that herpesviruses are usually associated with a single host species (28), and co-evolve with their host over long periods of time (28).
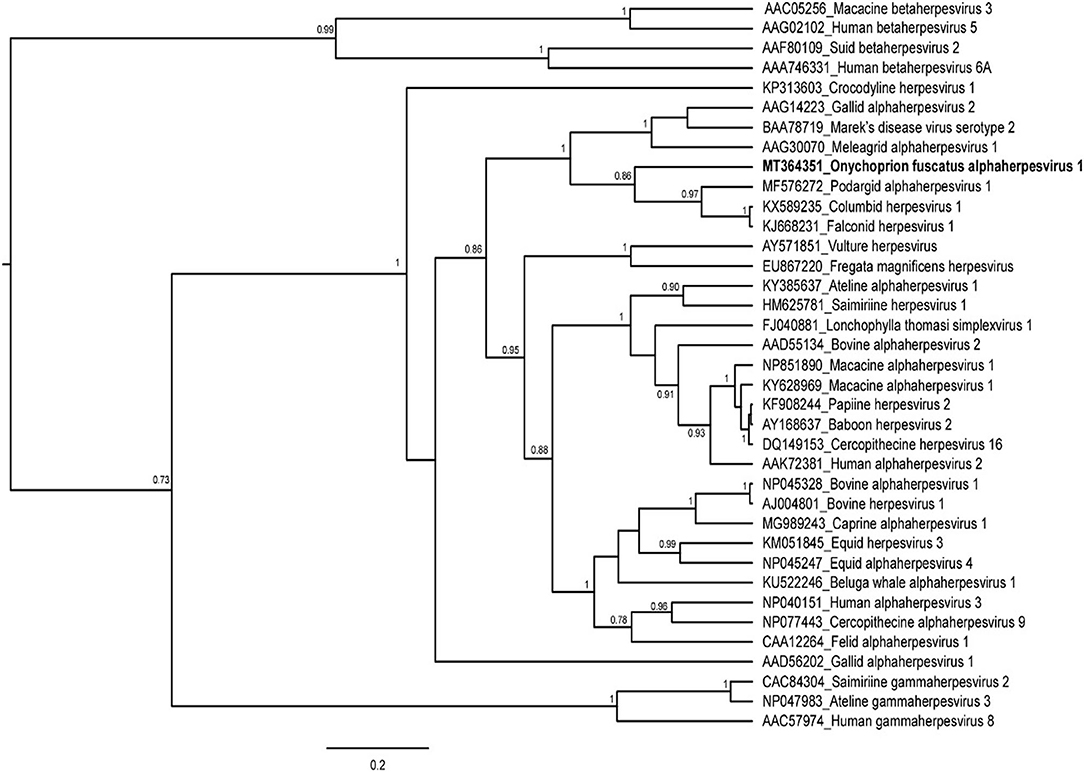
Figure 1. Bayesian inference of the phylogenetic relationships of the herpesvirus sequence identified in the sooty tern Onychoprion fuscatus. Bayesian posterior probabilities are shown at the nodes, when above 0.70. The sequence of Onychoprion fuscatus alphaherpesvirus 1 is in boldface. Alphanumerical codes represent the Genbank accession numbers of each herpesvirus sequence used in this study.
Because herpesviruses establish latent infections and have a high prevalence in natural hosts (11), symptoms of herpesvirus infection and the associated appearance of clinical signs may only occur when birds undergo a stressful situation (29). This raises the question of whether sooty terns were undergoing any form of stress and/or immune suppression. The severe clinical signs found in dying birds with positive PCR results may also suggest that sooty terns had no prior contact with this specific virus. Although very little is known about this population, recent work showed that sympatrically breeding frigatebirds have high blood concentrations of mercury while sooty terns showed very low mercury concentrations (30, 31). Mercury exposure can therefore likely be ruled out as a plausible candidate stressor for this population. However, suppression of the immune system in these birds might also be due to malnutrition, as has previously been suggested and recently corroborated in frigatebirds (13, 17).
This study does not conclusively prove the causal link between this herpesvirus and the occurrence of clinical signs and mortality in this population of sooty terns. The number of tissue samples and birds included in the present study was limited, and additional work would prove beneficial for a more solid interpretation of the results. However, the viral sequence here reported is novel and may be well-specific to this species, further supporting the fact that distinct avian populations are naturally infected with distinct herpesviruses. Seabirds aggregate at high densities during the breeding season, which may favor viral spread among conspecific and may lead to severe outbreaks in wild populations. We do not know the effect that this virus may have on this and other populations of sooty terns in terms of reproductive success and survival. However, given that herpesviruses in wild animals are usually only detected when they cause disease outbreaks, future works are needed to investigate herpesvirus prevalence in healthy, free-ranging seabirds. Seabirds are currently facing a strong decline in food resources (32, 33) and an increasing exposure to environmental contaminants and plastic pollution (34–36) which may increase their susceptibility to viral outbreaks.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/genbank/ (MT364351).
The animal study was reviewed and approved by Prefet de la region Guyane.
MS wrote the article. DCa, RB, and DCo performed phylogenetic analyses. AL and VL performed virus identification analyses. KP and OC contributed to data collection. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the CEBC (project SENTINEL to OC), the GEPOG, and DEAL Guyane, for funding, logistic support, and access to the Grand Connétable Nature Reserve. We are especially grateful to Grand Connétable staff for their great help in the field.
1. Davison AJ. Herpesvirus systematics. Vet Microbiol. (2010) 143:52–69. doi: 10.1016/j.vetmic.2010.02.014
2. Ehlers B, Dural G, Yasmum N, Lembo T, De Thoisy B, Ryser-Degiorgis MP, et al. Novel mammalian herpesviruses and lineages within the Gammaherpesvirinae: cospeciation and interspecies transfer. J Virol. (2008) 82:3509–16. doi: 10.1128/JVI.02646-07
3. Leendertz FH, Deckers M, Schempp W, Lankester F, Boesch C, Mugisha L, et al. Novel cytomegaloviruses in free-ranging and captive great apes: phylogenetic evidence for bidirectional horizontal transmission. J General Virol. (2009) 90:2386–94. doi: 10.1099/vir.0.011866-0
4. Escalera-Zamudio M, Rojas-Anaya E, Kolokotronis S-O, Taboada B, Loza-Rubio E, Méndez-Ojeda ML, et al. Bats, primates, and the evolutionary origins and diversification of mammalian gammaherpesviruses. MBio. (2016) 7:e01425–16. doi: 10.1128/mBio.01425-16
5. Huff J, Barry P. B-Virus (Cercopithecine herpesvirus 1) infection in humans and macaques: potential for zoonotic disease. Emerg Infect Dis. (2003) 9:246–50. doi: 10.3201/eid0902.020272
6. Parrish CR, Holmes EC, Morens DM, Park EC, Burke DS, Calisher CH, et al. Cross-species virus transmission and the emergence of new epidemic diseases. Microbiol Mol Biol Rev. (2008) 72:457–70. doi: 10.1128/MMBR.00004-08
7. Tischer BK, Osterrieder N. Herpesviruses–a zoonotic threat? Vet Microbiol. (2010) 140:266–70. doi: 10.1016/j.vetmic.2009.06.020
8. Longa CS, Bruno SF, Pires AR, Romijn PC, Kimura LS, Costa CHC. Human herpesvirus 1 in wild marmosets, Brazil, 2008. Emerg Infect Dis. (2011) 17:1308–10. doi: 10.3201/eid1707.100333
9. Long SY, Latimer EM, Hayward GS. Review of elephant endotheliotropic herpesviruses and acute hemorrhagic disease. Ilar J. (2016) 56:283–96. doi: 10.1093/ilar/ilv041
10. Tomaszewski E, Wilson VG, Wigle WL, Phalen DN. Detection and heterogeneity of herpesviruses causing Pacheco's disease in parrots. J Clin Microbiol. (2001) 39:533–8. doi: 10.1128/JCM.39.2.533-538.2001
11. Kaleta EF, Docherty DE. Avian Herpesviruses. In: Thomas NJ, Hunter DB, Atkinson CT, editors. Infectious Diseases of Wild Birds. New Jersey, NJ: Wiley (2008). p. 63–86. doi: 10.1002/9780470344668.ch3
12. Dhama K, Kumar N, Saminathan M, Tiwari R, Karthik K, Kumar MA, et al. Duck virus enteritis (duck plague) – a comprehensive update. Veterinary Quart. (2017) 37:57–80. doi: 10.1080/01652176.2017.1298885
13. De Thoisy B, Lavergne A, Semelin J, Pouliquen JF, Blanchard F, Hansen E, et al. Outbreaks of disease possibly due to a natural avian herpesvirus infection in a colony of young Magnificent Frigatebirds (Fregata magnificens) in French Guiana. J Wildl Dis. (2009) 45:802–7. doi: 10.7589/0090-3558-45.3.802
14. Sebastiano M, Eens M, Abd Elgawad H, Thoisy BD, Lacoste V, Pineau K, et al. Oxidative stress biomarkers are associated with visible clinical signs of a disease in frigatebird nestlings. Sci Rep. (2017) 7:1599. doi: 10.1038/s41598-017-01417-9
15. Sebastiano M, Eens M, Angelier F, Pineau K, Chastel O, Costantini D. Corticosterone, inflammation, immune status and telomere length in frigatebird nestlings facing a severe herpesvirus infection. Conserv Physiol. (2017) 5:cow073. doi: 10.1093/conphys/cow073
16. Sebastiano M, Eens M, Messina S, Abdelgawad H, Pineau K, Beemster GTS, et al. Resveratrol supplementation reduces oxidative stress and modulates the immune response in free-living animals during a viral infection. Funct Ecol. (2018) 32:2509–19. doi: 10.1111/1365-2435.13195
17. Sebastiano M, Eens M, Pineau K, Chastel O, Costantini D. Food supplementation protects Magnificent frigatebird chicks against a fatal viral disease. Conserv Lett. (2019) 12:e12630. doi: 10.1111/conl.12630
18. Rose TM, Strand KB, Schultz ER, Schaefer G, Rankin GW Jr, Thouless ME, et al. Identification of two homologs of the Kaposi's sarcoma-associated herpesvirus (human herpesvirus 8) in retroperitoneal fibromatosis of different macaque species. J Virol. (1997) 71:4138–44. doi: 10.1128/JVI.71.5.4138-4144.1997
19. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. (2011) 28:2731–9. doi: 10.1093/molbev/msr121
20. Cardoso M, Hyatt A, Selleck P, Lowther S, Prakash V, Pain D, et al. Phylogenetic analysis of the DNA polymerase gene of a novel alphaherpesvirus isolated from an Indian Gyps vulture. Virus Genes. (2005) 30:371–81. doi: 10.1007/s11262-005-6781-8
21. Drummond AJ, Suchard MA, Xie D, Rambaut A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol Biol Evol. (2012) 29:1969–73. doi: 10.1093/molbev/mss075
22. Lefort V, Longueville JE, Gascuel O. SMS: smart model selection in PhyML. Mol Biol Evol. (2017) 34:2422–4. doi: 10.1093/molbev/msx149
23. Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol. (2010) 59:307–21. doi: 10.1093/sysbio/syq010
24. Le Q, Gascuel O. An improved general amino acid replacement matrix. Mol Biol Evol. (2008) 25:1307–20. doi: 10.1093/molbev/msn067
25. Drummond AJ, Ho SYW, Phillips MJ, Rambaut A. Relaxed phylogenetics and dating with confidence. PLoS Biol. (2006) 4:e88. doi: 10.1371/journal.pbio.0040088
26. Rambaut A, Drummond AJ, Xie D, Baele G, Suchard MA. Posterior summarization in bayesian phylogenetics using tracer 1.7. Syst Biol. (2018) 67:901–4. doi: 10.1093/sysbio/syy032
27. Suchard MA, Lemey P, Baele G, Ayres DL, Drummond AJ, Rambaut A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. (2018) 4:vey016. doi: 10.1093/ve/vey016
28. Davison AJ. Evolution of the herpesviruses. Vet Microbiol. (2002) 86:69–88. doi: 10.1016/S0378-1135(01)00492-8
29. Goldberg DR, Yuill TM, Burgess EC. Mortality from duck plague virus in immunosuppressed adult mallard dUCKS. J Wildl Dis. (1990) 26:299–306. doi: 10.7589/0090-3558-26.3.299
30. Sebastiano M, Bustamante P, Costantini D, Eulaers I, Malarvannan G, Mendez-Fernandez P, et al. High levels of mercury and low levels of persistent organic pollutants in a tropical seabird in French Guiana, the Magnificent frigatebird, Fregata magnificens. Environ Pollut. (2016) 214:384–93. doi: 10.1016/j.envpol.2016.03.070
31. Sebastiano M, Bustamante P, Eulaers I, Malarvannan G, Mendez-Fernandez P, Churlaud C, et al. Trophic ecology drives contaminant concentrations within a tropical seabird community. Environ Pollut. (2017) 227:183–93. doi: 10.1016/j.envpol.2017.04.040
32. Wagner EL, Boersma PD. Effects of fisheries on seabird community ecology. Rev Fisheries Sci. (2011) 19:157–67. doi: 10.1080/10641262.2011.562568
33. Cahill AE, Aiello-Lammens ME, Fisher-Reid MC, Hua X, Karanewsky CJ, Ryu HY, et al. How does climate change cause extinction? Proc R Soc B Biol Sci. (2013) 280:20121890. doi: 10.1098/rspb.2012.1890
34. Vo A-TE, Bank MS, Shine JP, Edwards SV. Temporal increase in organic mercury in an endangered pelagic seabird assessed by century-old museum specimens. Proce Natl Acad Sci USA. (2011) 108:7466–71. doi: 10.1073/pnas.1013865108
35. Bond AL, Hobson KA, Branfireun BA. Rapidly increasing methyl mercury in endangered ivory gull (Pagophila eburnea) feathers over a 130 year record. Proc R Soc B Biol Sci. (2015) 282:20150032. doi: 10.1098/rspb.2015.0032
Keywords: alphaherpesvirus, French Guiana, Onychoprion fuscatus, seabirds, disease
Citation: Sebastiano M, Canestrelli D, Bisconti R, Lavergne A, Pineau K, Chastel O, Lacoste V and Costantini D (2020) Detection and Phylogenetic Characterization of a Novel Herpesvirus in Sooty Terns Onychoprion fuscatus. Front. Vet. Sci. 7:567. doi: 10.3389/fvets.2020.00567
Received: 18 May 2020; Accepted: 16 July 2020;
Published: 27 August 2020.
Edited by:
Armanda Bastos, University of Pretoria, South AfricaReviewed by:
Carlos G. Das Neves, Norwegian Veterinary Institute (NVI), NorwayCopyright © 2020 Sebastiano, Canestrelli, Bisconti, Lavergne, Pineau, Chastel, Lacoste and Costantini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manrico Sebastiano, TWFucmljby5TZWJhc3RpYW5vQHVhbnR3ZXJwZW4uYmU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.