 Ling Li1†
Ling Li1† Ziqiang Cheng
Ziqiang Cheng Guihua Wang
Guihua Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 03 April 2020
Sec. Veterinary Infectious Diseases
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.00152
This article is part of the Research Topic The Epidemiology, Diagnosis and Prevention of Infectious Diseases in Livestock View all 46 articles
Emerging evidence suggests that the tripartite motif containing 62 (TRIM62), a member of the TRIM family, plays an important role in antiviral processes. The objective of the study was to explore the role of TRIM62 in reticuloendotheliosis virus (REV) infection and its potential molecular mechanism. We first demonstrated that the REV infection affected the TRIM62 expression first upregulated and then downregulated in CEF cells. Next, we evaluated the effect of TRIM62 on viral replication. Overexpression of TRIM62 decreased REV replication. On the contrary, silencing of endogenously expressed TRIM62 increased viral replication. Then, to explore the necessity of domains in TRIM62's negative regulation on viral replication, we transfected CEF cells with TRIM62 domain deletion mutants. Deletion domain partially abolished TRIM62's antiviral activity. The effect of SPRY domain deletion was the highest and that of coiled-coil was the lowest. Further, we identified 18 proteins that coimmunoprecipitated and interacted with TRIM62 by immunocoprecipitation and mass spectrometry analysis. Strikingly, among which, both Ras-related protein Rab-5b (RAB5B) and Arp2/3 complex 34-kDa subunit (ARPC2) were involved in actin cytoskeletal pathway. Altogether, these results strongly suggest that chicken TRIM62 provides host defense against viral infection, and all domains are required for its action. RAB5B and ARPC2 may play important roles in its negative regulation processes.
TRIM62 (tripartite motif containing 62) is a member of the TRIM family proteins and is also known as DEAR1 (ductal epithelium-associated RING chromosome) (1). TRIM family proteins, also known as RBCC proteins, contain conserved RING finger, B-box, coiled-coil domains, and a variable C terminus (2). Despite their common domain structure, TRIM proteins play critical roles in distinct cellular processes such as intercellular signaling, innate immunity, transcription, autophagy, and carcinogenesis (3, 4). Members of the TRIM family of E3 ligases exhibit antiviral activities (5, 6). More than 20 TRIM proteins, which affected the entry or release of retrovirus such as human immunodeficiency virus 1 (HIV), murine leukemia virus (MIV), or avian leucosis virus (ALV), were screened (6). Expression of low amounts of TRIM62 enhanced HIV gene expression and release, and the E3 mutant of TRIM62 inhibited HIV release more potently than the wild-type protein (6). However, TRIM62 from orange-spotted grouper (EcTRIM62) negatively regulated the innate antiviral immune response against fish RNA viruses (7).
Reticuloendotheliosis virus (REV) is an avian retrovirus that can induce immunosuppression, runting syndrome, lymphomas, and acute reticulum cell neoplasia (8, 9). The occurrence of reticuloendotheliosis (RE) has an immunosuppressive effect and REV as the contaminant within vaccines against Marek's Disease (MD) (10), fowlpox (11, 12), and Gallid herpesvirus 2 (GaHV-2) (13), which may lead to vaccination failures and co-incidence of RE with other secondary infectious agents. Since breeder and layer flocks are commonly vaccinated against MD, the possible congenital transmission of REV between chickens was also be taking into account. The occurrence of RE has major economic importance. So far, no effective vaccines have been developed against RE; thus, the only protection remains flock renewal with elimination of affected birds or application of experimental antiviral treatment. In a previous study, we have confirmed that TRIM62 possesses restriction of avian leukosis virus subgroup J (ALV-J) replication (14). ALV-J is another avian retrovirus. At present, no data are available regarding the role of TRIM62 from chicken in REV infection.
To explore the role of TRIM62 in REV infection, in the present study, we detected and analyzed the association of TRIM62 expression with viral replication. Then, we evaluated the effects of TRIM62 on viral replication by overexpression, silencing, and domain deletion of TRIM62 in CEF cells with REV infection. Further, with TRIM62 overexpression, we screened key proteins that interacted with TRIM62. Our study provided evidence that chicken TRIM62 negative regulated the REV replication.
The CEF cells' cultural protocol was conducted as described in a previous study (14). CEF cells were incubated with a diluted stock of SNV strain of REV (China strain: JX0927, maintained in our laboratory) at a multiplicity of infection (MOI) of 0.1. The mRNA expression of TRIM62 and REV in CEF cells were detected at different times (12, 24, 48, 72, and 96 h post-infection). The mRNA expression of TRIM62 in the supernatant was also detected in parallel at the same sampling times.
To identify the antiviral function of TRIM62, CEF cells were seeded on six-well-plates for 12 h and transfected with plasmids using a lentiviral vector for TRIM62 overexpression and silence. The transfected plasmids containing chicken TRIM62 (pTRIM62)/domain deletion mutants were for TRIM62 overexpression, and those containing short hairpin RNA targeting TRIM62 (shTRIM62) were for TRIM62 silence. The gene sequence of chicken TRIM62 was obtained from GenBank (XM_015297235.2) (14). These plasmids fused to a Flag tag. The transfected empty vector CEF cells were used as control. The pTRIM62/shTRIM62/pTRIM62-ΔR/B/C/S and empty vector were purchased from Jikai Gene Technology, Inc. (Shanghai, China). After stably expressing TRIM62/shTRIM62 for 12 h/24 h, the transfected CEF cells were incubated with REV. After 72 h, the TRIM62 and viral mRNA/protein levels were detected by qRT-PCR/WB.
Total RNA from CEF cells was isolated using the Tiangen RNeasy mini kit according to the manufacturer's instructions. RNA was reverse transcribed to cDNA using the TaqMan Gold Reverse Transcription kit (Applied Biosystems). qRT-PCR was performed according to a previously described protocol (15, 16). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a control for basal RNA levels. Primer sequences are listed in Table 1.

Table 1. Primers used for quantitative reverse transcription-PCR.
The CEF cells were lysed in RIPA lysis buffer [25 mM Tris (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1% NP-40, 5% glycerol] containing protease and phosphatase inhibitor cocktails (Novasygen, Beijing, China). The lysis was separated by SDS-PAGE and transferred to PVDF membranes (Millipore, Bedford, USA) as reported previously (17, 18). The target proteins were detected with specific primary antibodies against TRIM62 (19) and REV env (primary antibodies were prepared by our laboratory, anti-REV gp90) at a 1:200, 1:500, and 1:3,000 dilution, respectively. The secondary antibodies were horseradish peroxidase (HRP)-conjugated enhanced chemiluminescence (ECL) goat. The blots were visualized by the ECL-enhanced chemiluminescence kit (Roche, Basel, Switzerland).
According to the instructions of Pierce Co-Immunoprecipitation (Co-IP) Kit (Thermo, Thermo Fisher Scientific, Massachusetts, USA), the 12-h pTRIM62 transfected CEF cells were subjected to REV infection for 72 h. The REV-infected CEF cells were then harvested and lysed. The anti-Flag label antibody was incubated with AminoLink coupling resin for 2 h at room temperature for antibody immunobilization. After centrifugation of cell lysates, supernatants were immunoprecipitated with coupling antibody. After another centrifugation of immunoprecipitated supernatants, the protein complex was washed three times with RIPA lysis buffer. Immunoprecipitates were incubated with elution buffer for 5 min at room temperature and centrifuged. The prey complex was collected and stored at −80°C. CEF cells were transfected with empty vector infected with REV as control.
Ten micrograms of the above prey complex was incubated in SDS-PAGE sample buffer (Solarbio, Beijing, China) for 5 min at 95°C. Proteins of cell lysates were separated by SDS-PAGE gels for 10 min and formed into a line. Each of the SDS-PAGE gel lanes was cut and subjected to trypsin digestion. Mass spectrometry (MS) analysis was performed according to a previously reported protocol (20) for detecting polypeptide sequence of proteins. Independent triplicate samples were analyzed. Polypeptide sequence was identified against using the Gallus (chicken) database and viral database (uniport-Gallusgalluschicken−REV−Combined.fasta) using Proteome Discoverer 1.4 (Thermo Fisher Scientific, Massachusetts, USA) software.
Results are presented as the means ± standard deviations (SD) of at least three sample replicates. Statistical analysis was performed using SPSS 19.0 statistical software, and p < 0.05 was considered statistically significant.
To assess association between the expression of TRIM62 and REV, we measured the TRIM62 expression and REV replication in REV-infected CEF cells at the transcriptional and translational level. Time course infection of REV in CEF cells showed that the REV mRNA levels were significantly increased (p < 0.001) from 48 to 96 h post-infection (Figure 1A), and the TRIM62 mRNA levels were also significantly upregulated at 48 h (p < 0.1) and 72 h (p < 0.1) as compared to that in uninfected CEF cells. However, compared with control, there were no changes of TRIM62 mRNA levels at another time point (Figure 1B). The REV mRNA expression was upregulated and reached the plateau at 96 h post-infection in CEF cells (17).
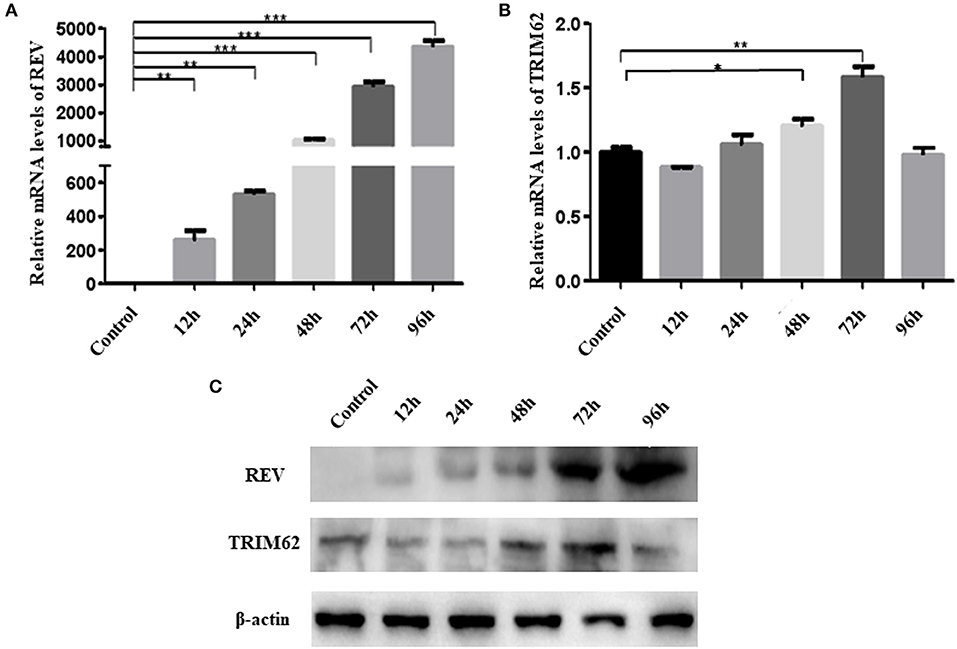
Figure 1. Effect of REV infection on TRIM62 expression. CEF cells were infected with REV. The RNA expression levels of REV (A) and of TRIM62 (B) at 12, 24, 48, 72, and 96 h post-infection in CEF cells were assessed by qRT-PCR. (C) The protein expression levels of REV and TRIM62 were detected by WB. Uninfected CEF cells were used as the control. *p < 0.05, **p < 0.01, and ***p < 0.001.
The protein levels of REV and TRIM62 were detected by WB, which were consistent with the dynamic changes of mRNA levels. Compared with control, the viral protein levels were significantly increased (p < 0.001) from 48 to 96 h post-infection, and TRIM62 protein levels were also significantly upregulated at 48 and 72 h (p < 0.01). However, there were no changes of TRIM62 protein levels at another time point observed (Figure 1C).
We overexpressed or silenced TRIM62 to detect the role of TRIM62 in REV replication in infected CEF cells. Compared with non-transfected cells, despite REV infection, the TRIM62 mRNA level was significantly greater (p < 0.01) in pTRIM62-transfected CEF cells (Figure 2A) and lower (p < 0.01) in shTRIM62-transfected CEF cells (Figure 2C). The TRIM62 overexpression decreased the REV mRNA expression in transfected cells (Figure 2B), whereas silence of TRIM62 increased the mRNA expression of REV (Figure 2D). These results were confirmed in protein levels by Western blotting (Figures 2A–D). Our results strongly suggest the role of TRIM62 in restricting REV infection.
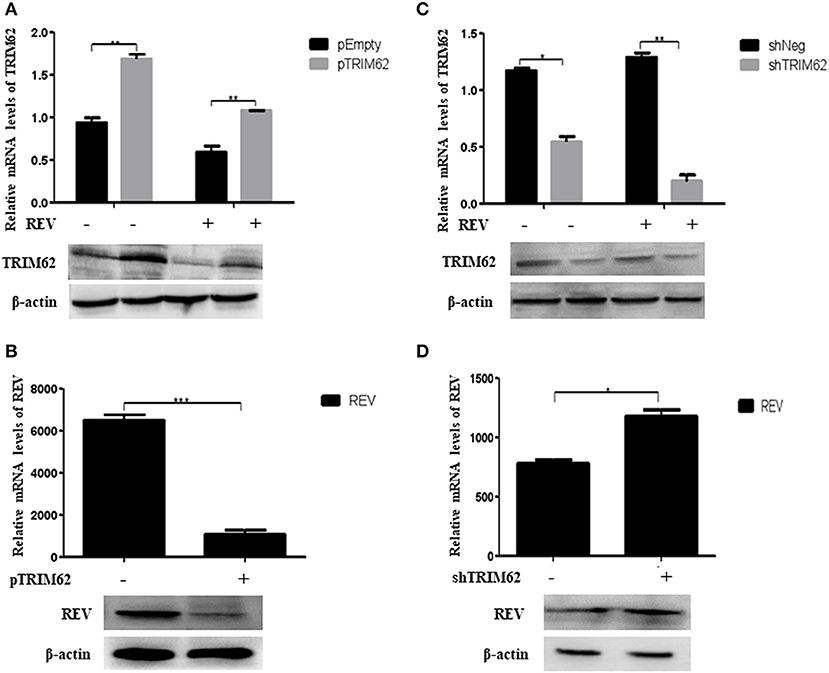
Figure 2. Restriction of REV replication in CEF cells induced by TRIM62. (A,B) CEF cells were transfected with pTRIM62 or empty vector before infection with REV. The expression levels of TRIM62 (A) and REV (B) were assessed by qRT-PCR and Western blotting. (C,D) CEF cells were transfected with shTRIM62 or a negative-control shRNA before infection with REV. The expression levels of TRIM62 (C) and REV (D) were determined by qPCR and Western blotting. *p < 0.05, **p < 0.01, and ***p < 0.001.
To further investigate the effect of domain on negative regulation of TRIM62, we explored the effect of TRIM62 domain deletion mutants on TRIM62 expression and REV replication. The mutants were prepared as previously described (14). Regardless of REV infection, the expression of TRIM62 was significantly higher in cells transfected with deletion of RING (Figure 3A) and B-box (Figure 3B) than empty vector (p > 0.1). When transfected with coiled-coil domain deletion mutant, viral infection decreased the expression of TRIM62 in cells (Figure 3C). These results indicated that the effect of REV infection on TRIM62 may associate with the coiled-coil domain. Compared with empty vector, there was no difference obtained of TRIM62 expression in cells transfected with deletion of SPRY domain mutant (Figure 3D). The qRT-PCR primers located in the SPRY domain may explain this result that deletion of SPRY domain did not result in difference of TRIM62 mRNA expression. To further confirm the results, we measured the TRIM62 protein expression with specific primary antibodies against TRIM62 (Figure 3E). The expression of TRIM62 in mutant-transfected cells is lower than that in complete TRIM62-transfected cells.
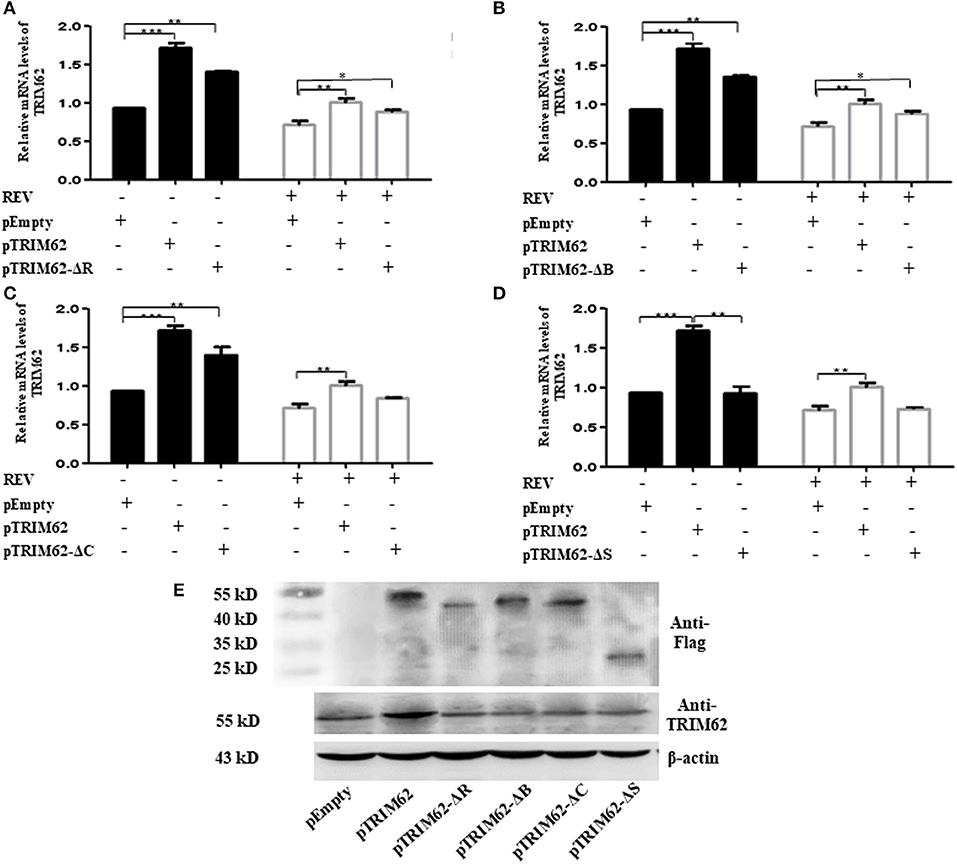
Figure 3. Effect of domain deletion on TRIM62 expression. The relative mRNA expression of TRIM62 in CEF cells transfected with RING (A), B-box (B), coiled-coil (C), or SPRY (D) domain deletion mutant with or without REV infection were measured by qRT-PCR using pEmpty and pTRIM62 as controls. (E) TRIM62 protein expression levels were detected with specific primary antibodies against Flag label and TRIM62 by Western blotting, respectively. pEmpty represent empty vector, pTRIM62 represent TRIM62 full length, pTRIM62-ΔR represent RING domain deletion, pTRIM62-ΔB represent B-box domain deletion, pTRIM62-ΔC represent coiled-coil domain deletion, and pTRIM62-ΔS represent SPRY domain deletion. *p < 0.05, **p < 0.01, and ***p < 0.001.
As compared with pEmpty vector-transfected cells, the level of REV mRNA expression was significant lower (p < 0.1) in CEF cells transfected by any of the TRIM62 domain deletion mutants (Figure 4A). The effect of coiled-coil domain deletion was the highest (77.9 ± 2.3%), and that of SPRY domain deletion was the lowest (22.8 ± 2.5%). Compared with complete TRIM62, the mRNA expression of REV was significantly higher in cells transfected with domain deletion mutants (Figure 4B). The viral expression in SPRY domain deletion-transfected cells was the highest (435.5 ± 6.2%), and that in coiled-coil domain deletion-transfected cells was the lowest (53.2% ± 6.4%). These results suggested that the deletion of domains partially abolished the antiviral activity of TRIM62.
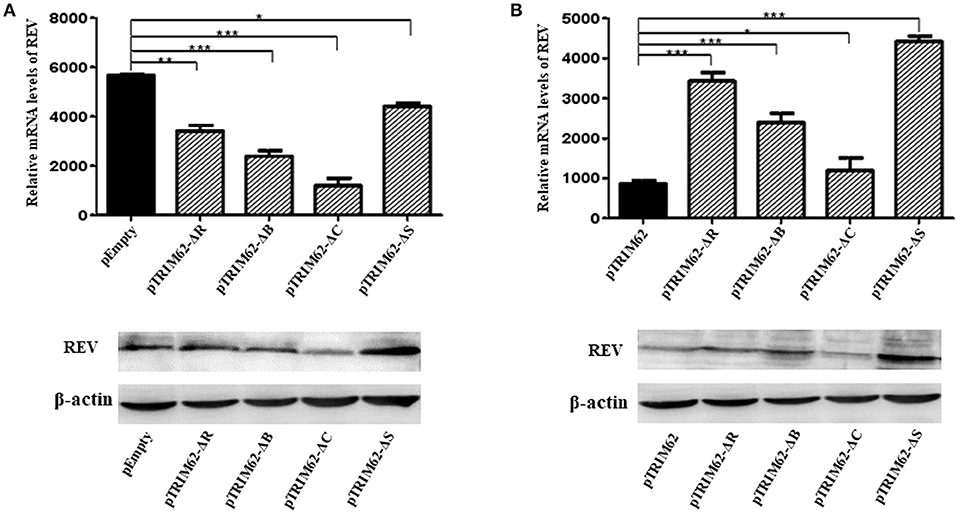
Figure 4. Inhibition of REV RNA expression in CEF cells by TRIM62 RING, B-box, coiled-coil, and SPRY domain deletion mutants. The relative mRNA expression levels of REV in CEF cells transfected with RING, B-box, coiled-coil, or SPRY domain deletion mutants were measured by qRT-PCR and Western blotting. pEmpty (A) and pTRIM62 (B) were used as controls. pEmpty represent empty vector, pTRIM62 represent TRIM62 full-length, pTRIM62-ΔR represent RING domain deletion, pTRIM62-ΔB represent B-box domain deletion, pTRIM62-ΔC represent coiled-coil domain deletion, and pTRIM62-ΔS represent SPRY domain deletion. *p < 0.05, **p < 0.01, and ***p < 0.001.
The REV infection affected the TRIM62 expression first upregulated and then downregulated in CEF cells. We hypothesized that other important interacting proteins besides the domains were involved in TRIM62's negative regulation of REV replication. To identify host cell proteins that interact with TRIM62, we used a tandem affinity purification approach coupled with mass spectrometry-based proteomics technology. These experiments identified 18 cell proteins that coimmunoprecipitated with transiently expressed Flag-tagged TRIM62 (Table 2). Decades of HIV research have testified to the integral role of the actin cytoskeleton in both establishing and spreading the infection (21). Of the TRIM62-interacting cell proteins identified, we focused on cytoskeletal proteins RAB5B and ARPC2. To verify the interaction between TRIM62 and RAB5B/ARPC2, CEF cells were transfected with a plasmid expression Flag-tagged RAB5B/APRC2 before REV infection, immunoprecipitated with anti-Flag antibody, and immunoblotted with anti-TRIM62 antibody. CEF cells were transfected with plasmid or empty vector uninfected with REV and transfected with empty vector infected with REV as control, respectively. Upon REV infection, anti-TRIM62 coprecipitated Flag-tag RAB5B (Figure 5A) and Flag-tag ARPC2 (Figure 5B).
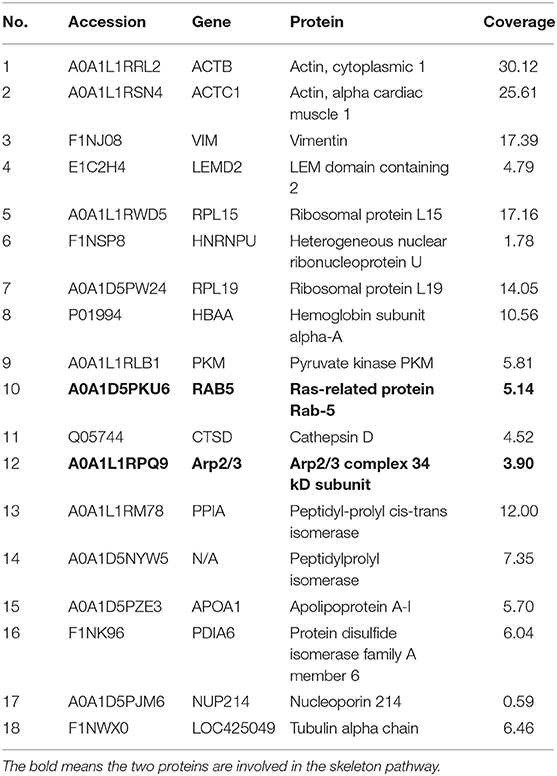
Table 2. Identified proteins interacting with TRIM62.
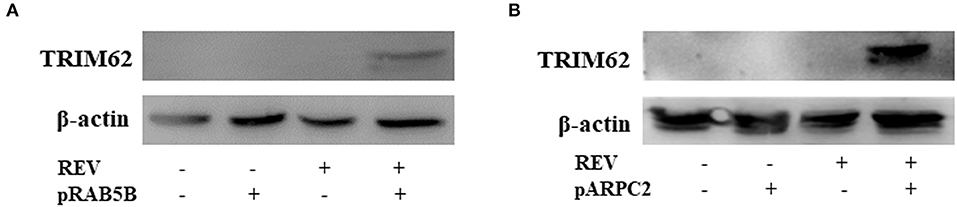
Figure 5. Validation of the interaction of TRIM62 with ARB5B and ARPC2. CEF cells were transfected with a plasmid expression Flag-tagged RAB5B (A) and APRC2 (B) before REV infection/uninfection and immunoprecipitated with anti-Flag antibody. The TRIM62 in immunoprecipitates was detected by Western blotting with anti-TRIM62 antibody.
Further, we found that TRIM62 affected the expression of RAB5B and ARPC2. As shown in Figures 6A,B, in CEF cells infected with REV, overexpression of TRIM62 exhibited a decrease of RAB5B expression, and silencing of TRIM62 enhanced the expression of RAB5B. On the contrary, in CEF cells infected with REV, the overexpression of TRIM62 resulted in an increase of ARPC2 expression, and silencing of TRM62 reduced the expression of ARPC2 (Figures 6C,D). These results indicated that RAB5B and ARPC2 play important roles in the negative regulation of TRIM62 on REV replication.
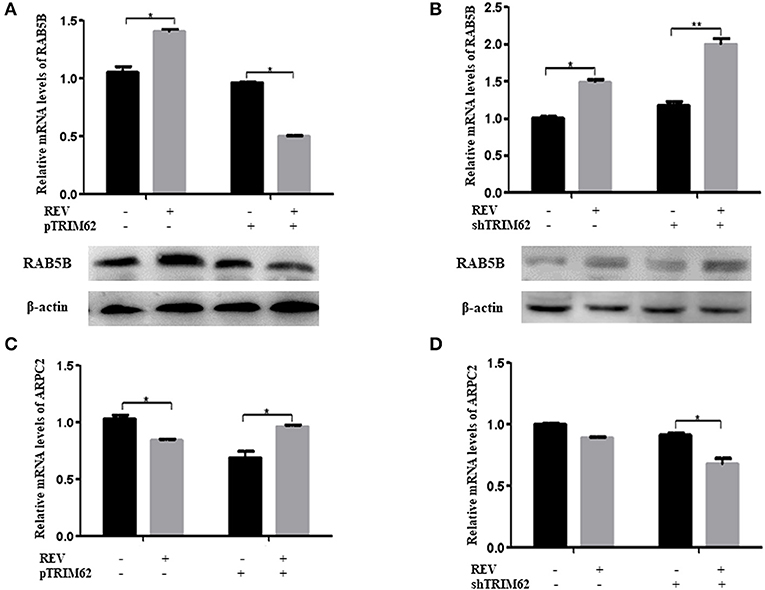
Figure 6. Effect of TRIM62 on the expression of RAB5B and ARPC2. (A,B) The relative mRNA and protein expression of RAB5B and in CEF cells were transfected with pTRIM62 (A) or shTRIM62 (B). (C,D) The relative mRNA expression of ARPC2 in CEF cells were transfected with pTRIM62 (C) or shTRIM62 (D). *p < 0.05, **p < 0.01.
The host innate immune system senses antigen and elicits local antiviral defense to control infection. However, the REV can evade surveillance by the immune system after infection. TRIM62 is an innate immune regulator (6). In this study, we found that the TRIM62 expression levels were first up and then down with the increase of REV replication in CEF cells. These results suggest that as an innate immune factor, TRIM62 was activated and induced a short periodic significant upregulation at the fast replication period of REV (p < 0.001), and REV may circumvent TRIM62's restrictive effects by reducing its expression levels. However, we did not obtain valid data of TRIM62 expression in supernatant because the RNA concentration was too low. The reason that TRIM62 expression was first increased and then decreased needed further confirmation.
In the current study, TRIM62 has been identified as a protein that negatively regulates REV replication. Gene silencing of TRIM62 enhanced REV infection, which indicated that TRIM62 contributes to the endogenous restriction of REV in CEF cells. The further characterization of mechanisms by which REV reduces TRIM62 expression is warranted.
Owing to the presence of similar domains, the TRIM proteins were involved in similar antiviral biological processes. However, the domains of TRIM proteins played complicated roles in the inhibition of different viruses. Deletion of the PRY-SPRY domain of TRIM62 from human did not compromise its signal transduction properties. Rather, in addition to the RING domain, both B-box and coiled-coil domains were required for NF-κB and AP-1 induction by TRIM62 (5). Thus, the B-box and/or coiled-coil domains that confer oligomerization/dimerization properties (22) are indispensable for TRIM62-mediated signaling (5). The SPRY domain mediated the affinity for the viral capsid and the function of RING domain to TRIM5α restriction of retrovirus infection was determined by the target virus (23). The antiviral activity of fish TRIM36 required both RING and SPRY domains (24). The RING and C-terminal tail were essential for TRIM56's antiviral activity against flaviviruses (25). Interestingly, the inhibitory effect of fish TRIM62 on interferon immune and inflammation response to negatively regulate virus replication was also dependent on RING and SPRY domains (7). In our study, this varied expression level of mutants (Figure 3) could be due to variation in transfection and expression efficiency of different plasmids in primary cells. Even though our results demonstrated that RING, B-box, coiled-coil, and SPRY domains contribute to the antiviral activity of chicken TRIM62, the effect of RING and SPRY was more significant than that of B-box and coiled-coil (Figures 4A,B). These results indicated that RING and SPRY domains are related to the antiviral activity of TRIM proteins.
Retroviruses are considered to use vesicular trafficking in infected cells (18, 26, 27). RAB5B is an isoform of RAB5, which is a member of the RAB family, a small GTPase family. RAB5B regulates fusion and motility of early endosomes, and is a marker of the early endosome compartment (28). RAB5B is a major regulator of hepatitis B virus (HBV) production (29). APRC2 is a component of Arp2/3 complex. The T cell-specific deletion of ARPC2 results in compromised peripheral T cell homeostasis (30). T cell survival and proliferation are mediated by complex homeostatic signals. Peripheral T cells are maintained at a constant cell number so that they can efficiently recognize foreign antigens and protect the host from pathogen invasion (31). Thus, of the TRIM62-interacting cell proteins identified, we focused on RAB5B and ARPC2. The interaction between RAB5B/ARPC2 and TRIM62 indicated that they played important roles in TRIM62 negative regulation on REV replication. A detailed understanding of host restriction may lead to antiviral therapies aimed at strengthening the innate immunity to retroviruses at the cellular and molecular level.
In addition, TRIM62 is a putative tumor suppressor and the TRIM62 levels represent an important prognostic marker in lung tumor (32) acute myeloid leukemia (AML) (33) and cervical cancer (34). ARPC2 promotes proliferation and metastasis (35, 36). REV is an oncogenic retrovirus. Further study is warranted to investigate the potential function of chicken TRIM62 and ARPC2 on REV infection inducing tumor formation.
We demonstrated that TRIM62 is a new suppressor that negatively regulates REV replication. The deletion of RING, B-box, coiled-coil, and SPRY domains partially affected TRIM62's antiviral activity, and the effects of RING and SPRY deletion were more significant than that of B-box and coiled-coil. The results suggested that all domains, RING, B-box, coiled-coil, and SPRY domains, are required for chicken TRIM62 to provide host defense against viral replication. Furthermore, both RAB5B and ARPC2 interacted with TRIM62, and maybe played important roles in TRIM62 negative regulation on REV replication. Our study provided a potential antiviral strategy targeting this novel regulator.
The datasets generated for this study are available on request to the corresponding author.
GW designed the experiments. LL, DN, and JY performed the experiments. GW, LL, and DN prepared the manuscript. JB, LZ, and ZC analyzed the data. All authors have read and approved the final version of the manuscript.
This work was supported by the National Natural Science Foundation of China (No. 31772703) and the Natural Science Foundation of Shandong Provincial (No. ZR2017MC011).
JB was employed by the company China Animal Husbandry Industry Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Lott ST, Chen N, Chandler DS, Yang Q, Wang L, Rodriguez M, et al. DEAR1 is a dominant regulator of acinar morphogenesis and an independent predictor of local recurrence-free survival in early-onset breast cancer. PLoS Med. (2009) 6:e1000068. doi: 10.1371/journal.pmed.1000068
2. Reymond A, Meroni G, Fantozzi A, Merla G, Cairo S, Luzi L, et al. The tripartite motif family identifies cell compartments. EMBO J. (2001) 20:2140–51. doi: 10.1093/emboj/20.9.2140
3. McNab FW, Rajsbaum R, Stoye JP, O'Garra A. Tripartite-motif proteins and innate immune regulation. Curr Opin Immunol. (2011) 23:46–56. doi: 10.1016/j.coi.2010.10.021
4. Napolitano LM, Meroni G. TRIM family: pleiotropy and diversification through homomultimer and heteromultimer formation. IUBMB Life. (2012) 64:64–71. doi: 10.1002/iub.580
5. Uchil PD, Hinz A, Siegel S, Coenen-Stass A, Pertel T, Luban J, et al. TRIM protein-mediated regulation of inflammatory and innate immune signaling and its association with antiretroviral activity. J Virol. (2013) 87:257–72. doi: 10.1128/JVI.01804-12
6. Uchil PD, Quinlan BD, Chan WT, Luna JM, Mothes W. TRIM E3 ligases interfere with early and late stages of the retroviral life cycle. PLoS Pathog. (2008) 4:e16. doi: 10.1371/journal.ppat.0040016
7. Yang Y, Huang Y, Yu Y, Zhou S, Wang S, Yang M, et al. Negative regulation of the innate antiviral immune response by TRIM62 from orange spotted grouper. Fish Shellfish Immunol. (2016) 57:68–78. doi: 10.1016/j.fsi.2016.08.035
8. Zavala G, Cheng S, Barbosa T, Haefele H. Enzootic reticuloendotheliosis in the endangered attwater's and greater prairie chickens. Avian Dis. (2006) 50:520–25. doi: 10.1637/7655-052806R.1
9. Barth CF, Ewert DL, Olson WC, Humphries EH. Reticuloendotheliosis virus REV-T(REV-A)-induced neoplasia: development of tumors within the T-lymphoid and myeloid lineages. J Virol. (1990) 64:6054–62. doi: 10.1128/JVI.64.12.6054-6062.1990
10. Mays JK, Silva RF, Kim T, Fadly A. Insertion of reticuloendotheliosis virus long terminal repeat into a bacterial artificial chromosome clone of a very virulent Marek's disease virus alters its pathogenicity. Avian Pathol. (2012) 41:259–65. doi: 10.1080/03079457.2012.675428
11. Awad AM, Abd El-Hamid HS, Abou Rawash AA, Ibrahim HH. Detection of reticuloendotheliosis virus as a contaminant of fowl pox vaccines. Poult Sci. (2010) 89:2389–95. doi: 10.3382/ps.2010-00899
12. Koo BS, Lee HR, Jeon EO, Jang HS, Han MS, Min KC, et al. An outbreak of lymphomas in a layer chicken flock previously infected with fowlpox virus containing integrated reticuloendotheliosis virus. Avian Dis. (2013) 57:812–17. doi: 10.1637/10551-041113-Case.R1
13. Wozniakowski G, Mamczur A, Samorek-Salamonowicz E. Common occurrence of Gallid herpesvirus-2 with reticuloendotheliosis virus in chickens caused by possible contamination of vaccine stocks. J Appl Microbiol. (2015) 118:803–8. doi: 10.1111/jam.12734
14. Li L, Feng W, Cheng Z, Yang J, Bi J, Wang XM, et al. TRIM62-mediated restriction of avian leukosis virus subgroup J replication is dependent on the SPRY domain. Poult Sci. (2019) 98:6019–25. doi: 10.3382/ps/pez408
15. Dai M, Feng M, Liu D, Cao W, Liao M. Development and application of SYBR Green I real-time PCR assay for the separate detection of subgroup J Avian leukosis virus and multiplex detection of avian leukosis virus subgroups A and B. Virol J. (2015) 12:52–62. doi: 10.1186/s12985-015-0291-7
16. Luan H, Wang Y, Li Y, Cui Z, Chang S, Zhao P. Development of a real-time quantitative RT-PCR to detect REV contamination in live vaccine. Poult Sci. (2016) 95:2023–9. doi: 10.3382/ps/pew147
17. Zhou D, Xue J, He S, Du X, Zhou J, Li C, et al. Reticuloendotheliosis virus and avian leukosis virus subgroup J synergistically increase the accumulation of exosomal miRNAs. Retrovirology. (2018) 15:45–56. doi: 10.1186/s12977-018-0427-0
18. Wang GH, Wang ZZ, Zhuang PP, Zhao XM, Cheng ZQ. Exosomes carring gag/env of ALV-J possess negative effect on immunocytes. Microb Pathogenesis. (2017) 112:142–7. doi: 10.1016/j.micpath.2017.09.013
19. Wang XM, Zhao YL, Li L, Cheng ZQ, Wang GH. Preparation and application of a monoclonal antibody against chicken TRIM62. Monoclon Antib Immunodiagn Immunother. (2018) 37:134–8. doi: 10.1089/mab.2017.0062
20. Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest. (2009) 119:1420–8. doi: 10.1172/JCI39104
21. Ospina Stella A, Turville S. All-round manipulation of the actin cytoskeleton by HIV. Viruses. (2018) 10:63–96. doi: 10.3390/v10020063
22. Diaz-Griffero F, Qin XR, Hayashi F, Kigawa T, Finzi A, Sarnak Z, et al. A B-Box 2 surface patch important for TRIM5α self-association, capsid binding avidity, and retrovirus restriction. J Virol. (2009) 83:10737–51. doi: 10.1128/JVI.01307-09
23. Li X, Kim J, Song B, Finzi A, Pacheco B, Sodroski J. Virus-specific effects of TRIM5α(rh) RING domain functions on restriction of retroviruses. J Virol. (2013) 87:7234–45. doi: 10.1128/JVI.00620-13
24. Chen B, Huo S, Liu W, Wang F, Lu Y, Xu Z, et al. Fish-specific finTRIM FTR36 triggers IFN pathway and mediates inhibition of viral replication. Fish Shellfish Immunol. (2019) 84:876–84. doi: 10.1016/j.fsi.2018.10.051
25. Liu BM, Li NL, Wang J, Shi PY, Wang TY, Miller MA, et al. Overlapping and distinct molecular determinants dictating the antiviral activities of TRIM56 against Flaviviruses and Coronavirus. J Virol. (2014) 88:13821–35. doi: 10.1128/JVI.02505-14
26. Booth AM, Fang Y, Fallon JK, Yang M Jr, Hildreth JEK, Gould SJ. Exosomes and HIV Gag bud from endosome-like domains of the T cell plasma membrane. J Cell Biol. (2006) 172:923–36. doi: 10.1083/jcb.200508014
27. Park IW, He JJ. HIV-1 is budded from CD4+ T lymphocytes independently of exosomes. Virol J. (2010) 7:234–39. doi: 10.1186/1743-422X-7-234
28. Stenmark H, Parton RG, Steele-Mortimer O, Lütcke A, Gruenberg J, Zerial M. Inhibition of rab5 GTPase activity stimulates membrane fusion in endocytosis. EMBO J. (1994) 13:1287–96. doi: 10.1002/j.1460-2075.1994.tb06381.x
29. Inoue J, Ninomiya M, Umetsu T, Nakamura T, Kogure T, Kakazu E, et al. Small interfering RNA screening for the small GTPase Rab proteins identifies Rab5B as a major regulator of Hepatitis B virus production. J Virol. (2019) 93:e00621–19. doi: 10.1128/JVI.00621-19
30. Li L, Zhuang PP, Wang XM, Zhou DF, Xue JW, Cheng ZQ, et al. Effects of miRNA-155 on the cytoskeletal pathways in REV and ALV-J Co-Infection. Chinese J Cell Biol. (2018) 40:1706–18. doi: 10.11844/cjcb.2018.10.0223
31. Takada K, Jameson SC. Naive T cell homeostasis: from awareness of space to a sense of place. Nat Rev Immunol. (2009) 9:823–32. doi: 10.1038/nri2657
32. Quintas-Cardama A, Post SM, Solis LM, Xiong S, Yang P, Chen N, et al. Loss of the novel tumour suppressor and polarity gene Trim62 (Dear1) synergizes with oncogenic Ras in invasive lung cancer. J Pathol. (2014) 234:108–19. doi: 10.1002/path.4385
33. Quintás-Cardama A, Zhang N, Qiu YH, Post S, Creighton CJ, Cortes J, et al. Loss of TRIM62 expression is an independent adverse prognostic factor in acute myeloid leukemia. Clin Lymphoma Myeloma Leuk. (2015) 15:115–27.e15. doi: 10.1016/j.clml.2014.07.011
34. Liu TY, Chen J, Shang CL, Shen HW, Huang JM, Liang YC, et al. Tripartite motif containing 62 is a novel prognostic marker and suppresses tumor metastasis via c-Jun/Slug signaling-mediated epithelial-mesenchymal transition in cervical cancer. J Exp Clin Cancer Res. (2016) 35:170–90. doi: 10.1186/s13046-016-0445-5
35. Cheng Z, Wei W, Wu Z, Wang J, Ding X, Sheng Y, et al. ARPC2 promotes breast cancer proliferation and metastasis. Oncol Rep. (2019) 41:3189–200. doi: 10.3892/or.2019.7113
Keywords: TRIM62, negative regulation, reticuloendotheliosis virus, domain deletion, RAB5B and ARPC2
Citation: Li L, Niu D, Yang J, Bi J, Zhang L, Cheng Z and Wang G (2020) TRIM62 From Chicken as a Negative Regulator of Reticuloendotheliosis Virus Replication. Front. Vet. Sci. 7:152. doi: 10.3389/fvets.2020.00152
Received: 20 January 2020; Accepted: 03 March 2020;
Published: 03 April 2020.
Edited by:
Anuwat Wiratsudakul, Mahidol University, ThailandReviewed by:
Baoming Liu, Johns Hopkins Medicine, United StatesCopyright © 2020 Li, Niu, Yang, Bi, Zhang, Cheng and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guihua Wang, d2d1aWh1YTExMjZAMTYzLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.