 Francesca Soutter
Francesca Soutter Dirk Werling
Dirk Werling Fiona M. Tomley
Fiona M. Tomley Damer P. Blake
Damer P. Blake
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Vet. Sci. , 26 February 2020
Sec. Veterinary Infectious Diseases
Volume 7 - 2020 | https://doi.org/10.3389/fvets.2020.00101
This article is part of the Research Topic Poultry Coccidiosis: Strategies to Understand and Control View all 9 articles
Eimeria infection impacts upon chicken welfare and economic productivity of the poultry sector. Live coccidiosis vaccines for chickens have been available for almost 70 years, but the requirement to formulate blends of oocysts from multiple Eimeria species makes vaccine production costly and logistically demanding. A multivalent vaccine that does not require chickens for its production and can induce protection against multiple Eimeria species is highly desirable. However, despite the identification and testing of many vaccine candidate antigens, no recombinant coccidiosis vaccine has been developed commercially. Currently, assessment of vaccine efficacy against Eimeria, and the disease coccidiosis, can be done only through in vivo vaccination and challenge experiments but the design of such studies has been highly variable. Lack of a “standard” protocol for assessing vaccine efficacy makes comparative evaluations very difficult, complicating vaccine development, and validation. The formulation and schedule of vaccination, the breed of chicken and choice of husbandry system, the species, strain, magnitude, and timing of delivery of the parasite challenge, and the parameters used to assess vaccine efficacy all influence the outcomes of experimental trials. In natural Eimeria infections, the induction of strong cell mediated immune responses are central to the development of protective immunity against coccidiosis. Antibodies are generally regarded to be of lesser importance. Unfortunately, there are no specific immunological assays that can accurately predict how well a vaccine will protect against coccidiosis (i.e., no “correlates of protection”). Thus, experimental vaccine studies rely on assessing a variety of post-challenge parameters, including assessment of pathognomonic lesions, measurements of parasite replication such as oocyst output or quantification of Eimeria genomes, and/or measurements of productivity such as body weight gain and feed conversion rates. Understanding immune responses to primary and secondary infection can inform on the most appropriate immunological assays. The discovery of new antigens for different Eimeria species and the development of new methods of vaccine antigen delivery necessitates a more considered approach to assessment of novel vaccines with robust, repeatable study design. Careful consideration of performance and welfare factors that are genuinely relevant to chicken producers and vaccine manufacturers is essential.
Poultry health and welfare is threatened by a number of pathogens and protozoan parasites of the genus Eimeria are among the most important. Infection with Eimeria, which invade and replicate within gut epithelial cells, can compromise chicken welfare and reduce productivity in both layer and broiler systems requiring relatively costly treatments. The cost of these interventions, combined with the losses caused by infection, are estimated to cost the global chicken industry in excess of £2 billion every year (1). Eimeria infection has also been demonstrated to exacerbate the outcome of exposure to other pathogens such as Clostridium perfringens, combining to cause necrotic enteritis (2).
Control of Eimeria in commercial chicken production relies on routine chemoprophylaxis (primarily broiler chickens) or vaccination using formulations of live virulent or attenuated Eimeria species (layer and breeder chickens). Mass administration of anticoccidial drugs has long been employed as a highly effective method of control, however drug resistance is widespread and public/legislative pressure demanding reduced use in livestock production is intensifying (3, 4). Classification of ionophores as antibiotics in markets such as the United States of America has reduced use further as demand for “no antibiotics, ever” food products grows, prompting more than 40% of producers to include vaccination in one or more production cycles per year (5). Unfortunately, existing vaccines are relatively expensive due to high production costs and are difficult to scale up, especially those that include attenuated parasite lines. In response, demand for cost-effective anticoccidial vaccines is greater than ever.
Seven species of Eimeria are recognized to infect chickens and these vary in their fecundity, pathogenicity and location of replication within the gut [reviewed in Chapman (6)]. Infection with each species induces a robust protective immune response against homologous challenge (7). However, the numbers of parasites and rounds of infection required to induce immunity that is sufficient to protect against disease varies depending on the infecting Eimeria species, dosing schedule and chicken breed (7–10). There is little or no immune protection against challenge infection with a heterologous species, and in some cases even infection with a different strain of the same species can escape immune protection (11, 12). Therefore, although immunization of chickens with live Eimeria oocysts is effective and has been the basis of live oral coccidiosis vaccines for almost 70 years, chickens must be vaccinated with oocysts from each Eimeria species in order to be fully protected. The requirement for a live vaccine to include controlled doses of oocysts for all pathogenic species of Eimeria, and in some cases multiple strains of Eimeria maxima, makes vaccine manufacture logistically demanding as all vaccine lines have to be propagated separately in chickens under stringent specific pathogen free conditions. Another important consideration is that fecal-oral recycling of vaccine parasites is required to generate levels of protective immunity that are sufficient to protect chickens against pathogenic challenge by most Eimeria species (13). A recombinant vaccine that is protective against multiple Eimeria species is therefore highly desirable and this should contain multiple antigens, derived from different parasite species and lifecycle stages.
Criteria to identify a “good” new vaccine are difficult to define and may depend on the stakeholder asked. Sterile immunity, defined as the ability to stop all oocyst production following challenge, might be regarded as the primary objective but current anticoccidial drugs and live vaccines do not meet this stringent requirement (14, 15). Indeed, achievement of such strong evolutionary pressure may select for vaccine resistance (13). Important measures of efficacy that should be considered include decreased parasite load or replication, lowering environmental oocyst occurrence and thus transmission, reduced clinical signs of disease, and improved performance. Important performance parameters include feed conversion efficiency, body growth rate, flock homogeneity, and/or egg production. A vaccine should also be easy to administer, and safe for the target- and overlapping populations. Stability and consistency between batches is of utmost importance. Onset and duration of immunity are also important although the relevance of these parameters vary depending on the target population; short lived broiler chickens require vaccines with rapid onset of immunity, but duration is of less importance, whilst the opposite is commonly true for layer chickens. In the absence of effective correlates of protection, assessment of novel vaccines to protect against coccidiosis can only be performed in-vivo (as discussed below), but the design of experimental vaccine challenge studies has been highly variable in the published literature. This lack of standardization results in difficulty evaluating new vaccines and makes comparison between studies challenging. As well as study design, the choice of parameters evaluated during a vaccine challenge is also important when evaluating novel vaccines, particularly for intracellular pathogens such as Eimeria where antibody response is not a reliable correlate of protection against disease (16). This review examines the challenges in design of vaccination studies for chicken coccidiosis and methods for evaluating vaccine efficacy.
The first vaccines against coccidia used live, wild-type, sporulated E. tenella oocysts and were initially marketed in the 1950's based on observations that administration of low doses of oocysts over a number of days induced protective immunity against homologous challenge. Over time these first generation vaccines were developed to incorporate further Eimeria species and have been widely utilized, particularly in North America (15). Methods for delivery of these live wild type vaccines have improved over the years; common methods include spraying live oocysts directly onto day of hatch chicks in a spray cabinet, although vaccines can also be delivered by spraying onto feed, in an edible gel or in drinking water (17). Nevertheless, uniform administration of oocysts is not always accomplished and recycling of live oocysts by reinfection from litter is required for the development of robust immunity within the flock (18). Recycling of oocysts is difficult in traditional layer cage setups, although the addition of floor coverings have been trialed and found to improve oocyst recycling (19). A significant risk in the use of these wild-type, and thus fully virulent, live vaccines is that large numbers of oocysts accumulate rapidly within chicken house litter resulting in high levels of exposure and clinical disease, even mortality, necessitating the use of anticoccidial drugs following vaccination (15).
Newer second generation anticoccidial vaccines frequently utilize oocysts derived from attenuated lines of Eimeria parasites, the majority of which have been selected for an abbreviated lifecycle termed “precocious development” [reviewed in Shirley and Bedrnik (20)]. Briefly, consistent selection during in vivo passage for those oocysts which are produced at the beginning of the patent period result in parasite lines with shorter prepatent periods, facilitated by fewer and/or smaller rounds of schizogony, and reduced total oocyst production (21). Precocious parasite lines exhibit reduced pathogenicity, but importantly retain their immunogenicity (22). The exception is an E. tenella line selected for growth in embryonic chicks (23, 24). As for wild type anticoccidial vaccines, attenuated vaccine formulations include a mixture of sporulated oocysts of different Eimeria species. Attenuated anticoccidial vaccines are also given orally, adopting strategies similar to those used for live wild-type vaccines.
Production of live wild-type and attenuated anticoccidial vaccines is limited by the necessity for in-vivo infection of chickens to produce oocysts as these cannot be produced in vitro. They are therefore costly and time-consuming to produce when compared to alternatives such as anticoccidial drugs, most notably for attenuated vaccine lines that are less productive than their wild-type equivalents. Attenuation of less fecund but highly pathogenic species such as E. brunetti and E. necatrix results in parasite lines with limited reproductive potential, making routine propagation challenging (25). Other limitations include the necessity for detailed quality control of each vaccine batch for efficacy that can only be achieved in vivo, as well as a short shelf life and the requirement for a cold chain without options for freezing or freeze drying. Such limitations in production of vaccinal parasite lines can restrict vaccine availability, encouraging demand for recombinant vaccines.
A number of Eimeria antigens have been identified as valid anticoccidial vaccine candidates, but no recombinant vaccines are commercially available (13). Comparison of results from vaccination and challenge experiments using different, or even the same, antigens is challenging because of high variability in the study design of published vaccination trials (Table 1). Common differences include magnitude and timing of the challenge dose, vaccine formulation (expression system, antigen-adjuvant combination, and quantity of antigen), vaccine dosing schedule, chicken breed or genetic line, husbandry set up and the choice of parameters measured to assess protection. Each factor can be confounding, which makes it difficult to compare efficacy of different antigens between studies. Selection of chicken breed and vaccine schedules might also make it difficult to assess whether vaccines could work in a commercial setting with large numbers of outbred birds, where cost of dose per chicken and improvements in production parameters are of utmost importance. The use of common standard chicken lines that are widely available, such as Cobb500 or Ross 308 broilers, or Hyline layers, will encourage comparison between studies. However, the hybrid nature of these chicken lines can result in elevated levels of experimental variation. Using specific pathogen free and/or inbred chicken lines can reduce non-treatment variation and improve statistical validity.
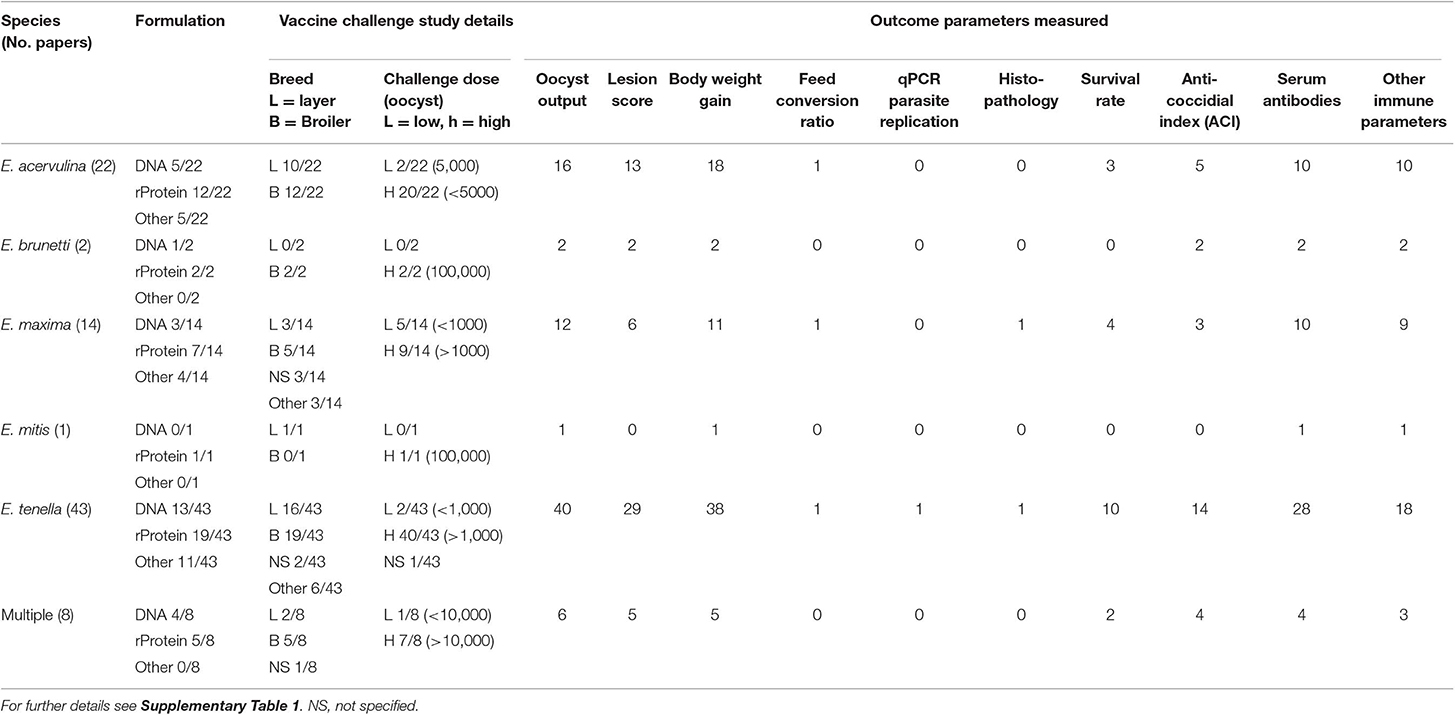
Table 1. A summary of key published anticoccidial vaccination studies.
All studies using live animals must be well-designed and have animal welfare at the forefront. The three Rs principles should be considered; replace, reduce and refine animal usage (26). Where possible consider replacing the use of animals; novel antigens could be examined in-vitro using existing cell lines before moving to in-vivo work, although scope is very limited given the inability of Eimeria to complete its lifecycle efficiently in-vitro. Reducing the number of chickens used in a study is important whilst balancing the need for adequate statistical power to examine the effect of any novel vaccine (27). Reducing variability in response to infection and vaccination by optimal chicken breed selection and careful study design could reduce the number of individuals required to achieve a statistically meaningful result. Refinement of studies through careful consideration of animal housing and procedures should maximize animal welfare and improve results; unnecessary stress during housing and handling is likely to impact on vaccine response. For example, housing chickens in larger cages for oocyst counting experiments can reduce the efficacy of oocyst recovery, increasing variation between groups and compromising statistical validity. Previous studies have suggested a minimum of between five and 10 chickens per treatment group is required for assessment of differences between lesion scores due to variability in response to infection with Eimeria. (28), although the magnitude of the anticipated response should be used to inform an appropriate estimate of statistical power (e.g., 2-sample t-test method). Similarly, when using quantitative PCR (qPCR) to assess parasite genome copy numbers in the caeca of chickens infected with E. tenella a group size of a minimum of six birds was required during validation of the technique at 5 days post infection (27).
The choice of chicken breed or line used in an Eimeria vaccine challenge study is likely to be determined based on a number of criteria including relevance to the target chicken population, experimental setup, outcomes to be measured, and genetic background. Use of inbred chicken lines can reduce host variation in response to vaccination and Eimeria infection, offering opportunities to minimize the number of chickens needed to assess vaccination outcomes and streamline experimental design. However, vaccine responses and the consequences of challenge are likely to differ from the hybrid commercial chicken lines that most anticoccidial vaccines will be intended for. If using commercial layer or broiler chickens the choice of line is also important and again could be influenced by the target market of the vaccine. One advantage of examining vaccine response in layer chickens is that cages can more readily be used in place of floor pens. Keeping chickens in cages can permit more flexible study design, also offering better control of oocyst recycling and opportunities for fecal collection for accurate enumeration of oocyst excretion. However, if measuring production outcomes such as weight gain is desirable then the use of broiler chickens is likely to be advantageous over layers.
It has been demonstrated that some inbred chicken lines are more susceptible to Eimeria infection than others (29). A potential association between MHC haplotype and Eimeria infection outcome has been identified, although results of individual studies have been inconsistent, possibly influenced by the choice of chicken haplotypes examined, the infection model used, chicken infection histories, and measures of infection used to assess susceptibility/resistance (30–32). It is also likely that additional genes are involved in disease susceptibility/tolerance/resistance and this has been supported by some studies (33, 34). As well as influencing the outcome of challenge, it is also likely that the genetic background of the chicken will influence response to any novel vaccine being assessed. For example, there is evidence that MHC B locus haplotype may influence protection after challenge; lesion scores after challenge infection with E. tenella were reduced in B5B5 chickens vaccinated with a recombinant protein derived from oocysts, but this response was not seen in B2B2 chickens (35). However, in another study B2B2 chickens were found to be more protected than other lines by vaccination following challenge with E. acervulina (36). The choice of chicken line and genetic background therefore must be considered when designing vaccine challenge studies, and it may be necessary to test the same vaccines in different geographically distributed chicken breeds.
Rodent models are sometimes used to examine responses to coccidial infection but are unlikely to replace the need for work in chickens as not only do different species of Eimeria infect each host, but the immune system of rodents is very different compared to avians (37, 38). While there are many similarities between Eimeria species that infect birds or mammals and most replicate within enterocytes, fundamental differences include antigenic profiles, schizont size and number of rounds of schizogony (39, 40), all of which may impact on vaccination and immunological outcomes. The availability of immunological tools and reagents for rodent species continue to offer value to vaccine development, but there has been a notable expansion of resources for work with chickens (see https://www.immunologicaltoolbox.co.uk/ for details). Furthermore, knockout mice with well-characterized phenotypes are readily available and can be utilized to better understand immune responses to infection. Although the limitations of these knockout mice is becoming more apparent (41), some experiments are still easier to perform in these mice compared to chicken, and may shed light on pathogenicity. For example, infection of mice with Eimeria vermiformis shows some similarities to E. maxima infection of chickens in terms of immunogenicity, pathogenicity and the area of gut infected, and have been utilized as an infection model (42). Nevertheless, mechanisms of pathogenicity or immunogenicity in rodent models are not necessarily reproducible in avian species.
The choice of Eimeria species used in vaccine development and validation is commonly based on regional priorities and identity of the candidate antigen(s) to be tested. The choice of Eimeria species influences study design. Variation in fecundity, pathogenicity, and immunogenicity among Eimeria species impacts on the outcome of vaccination and severity of disease, but also dictates the magnitude of challenge dose and study design. If a multivalent vaccine is developed to protect against multiple Eimeria species, efficacy against each species should initially be assessed in isolation with a separate challenge for each species to avoid confounding results. In some examples the choice of Eimeria strain/genotype is also important. Strain-specific antigenic variation has been noted to influence the outcome of vaccination for several species including E. maxima (11). The same strain or isolate used as template in production of a vaccine should also be employed as the challenge, ensuring antigenic as well as species homogeneity. Subsequent vaccine development should then include comparison with antigenically distinct strains to assess cross-protection. The choice of strain or isolate can vary between studies and regions, regularly prioritizing local parasites to promote local relevance. However, the use of multiple different strains, many of which may be antigenically distinct, precludes effective comparison between studies. Standardization using vaccines and challenges developed from reference strains such as Houghton (H) Eimeria strains (40) can improve opportunities for comparison.
Challenge doses should be determined empirically based on breed of chicken and species of Eimeria and should be tailored to the study design so that the desired outcome of vaccination can be measured reliably and at an appropriate time point post infection (e.g., reduction in lesion score or parasite replication). A lower challenge dose is more appropriate for measurements of parasite replication such as oocyst counts or quantitative PCR (qPCR) to measure Eimeria genome copies, as the linear dose dependent relationship between challenge dose and parasite replication eventually plateaus as the challenge dose is increased (27, 43). Any potential reduction in parasite replication as a result of your vaccination is thus difficult to interpret at higher challenge doses due to this biological phenomena. Higher challenge doses that induce at least moderate lesions and depress weight are more appropriate for assessment of reduction in pathogenicity by lesion scoring or for production measurements such as body weight gain. The age, quality and sporulation rate of oocysts used for challenge must also be considered; oocysts stored for a long time or inadequately (e.g., incorrect temperature) are likely to be less infectious. A reduction in pathogenicity in-vivo is usually observed when using oocysts older than 6 months of age and a reduction in replication can be detected from 3 months. Standardization of oocyst age at the time of dosing between experiments will promote comparison between studies, although it has been suggested that dose adjustment can be employed if using older cultures (44). Administering the challenge dose is usually performed by oral gavage of oocysts, either as a single dose or multiple doses to try to mimic natural infection. Infection from the environment can be initiated through re-use of litter or co-housing with infected individuals.
Elucidating the effector cells and molecules involved in protective immunity induced by natural Eimeria infection and then eliciting either a similar response by vaccination or a different one, if this is protective, is the desired outcome of many novel vaccines. However, it has to be stressed that the definition of an effective vaccine may differ widely between stakeholder groups. Indeed, this can range from the “best case” scenario, the induction of sterile immunity to more realistic responses, such as reduction in shedding and/or reduction of clinical signs. However, it is clear that understanding these immune responses can offer valuable insight into vaccine development and efficacy. It is apparent that Eimeria sporozoites interact with both phagocytic and non-phagocytic cells within the gut, but this interaction does not always result in clearance of the parasite. Early studies suggested that Eimeria sporozoites are transported from the gut lumen to the crypt epithelium, where most species develop further, via the lamina propria in intraepithelial lymphocytes (45). Another study using monoclonal antibodies to stain duodenal sections infected with E. acervulina found sporozoites to be within CD8+ cells, macrophages and, to a lesser extent, CD4+ cells (46). The same study confirmed that both live and dead sporozoites were present within macrophages cultured in-vitro, suggesting that uptake into the macrophage might not result in parasite killing (46). However, subsequent publications concluded that the percentage of E. tenella sporozoites found within intraepithelial lymphocytes was relatively low (47). In this study, most sporozoites were found within enterocytes although the number located within lymphocytes was higher in the lamina propria (47). It was suggested that in birds immune following natural infection sporozoites remain within intraepithelial lymphocytes and do not penetrate the crypt epithelial cells (45), whereas others suggested that sporozoites remain within the lamina propria and do not enter the crypt epithelium or are inhibited from further development within the crypt epithelium (47). It is important to note that effector cells and molecules might vary between primary and secondary immune responses, and this is certainly true of E. vermiformis infection in mice (42). Therefore, these potential differences must be considered during development of vaccines and protocols for vaccine characterization.
Eimeria specific antibodies are generated during Eimeria infection, but they do not appear to be involved in controlling infection following oocyst challenge (7), and antibody levels are not correlated with disease susceptibility (16). Early studies demonstrated that bursectomised chickens, unable to generate an antibody response, were resistant to a secondary challenge with E. tenella suggesting that a serological response is not required for protective immunity (48). However, it has been demonstrated that sera from chickens recovering from infection may inhibit sporozoite invasion of chick kidney cells in-vitro (49). Lysis of sporozoites by immune sera and changes suggestive of agglutination have been demonstrated in-vitro but surface changes to sporozoites and to a lesser extent merozoites, visible by electron microscopy, were not detected when immune sera was heat inactivated suggesting that this action could be mediated by complement (50). Passive protection of chickens against E. tenella infection with a monoclonal antibody has also been demonstrated in-vivo (51). The role of secretory IgA was initially considered important in protecting against parasite invasion (49). Caecal contents from immunized chickens were demonstrated to reduce sporozoite invasion of a chick kidney cell line however parasite development was not associated with immunoglobulin level or neutralization of sporozoites (52). Similarly, sporozoite numbers were reduced in the intestinal lumen of previously challenged infected chickens but these sporozoites were still infective and anti-sporozoite gall bladder IgA was not associated with immunity to reinfection (53). Antibodies therefore do not appear to play an important role in protective immunity against Eimeria infection in vivo and cell mediated responses appear to be of more importance.
The role of T-lymphocytes in inducing a protective immune response to Eimeria infection has long been recognized in rodent models devoid of T-lymphocytes, as these were unable to control infection [reviewed by Rose (54)]. An initial study in athymic nude rats infected with E. nieschulzi demonstrated that these rats did not become immune to a second infection. In contrast, heterozygous (nu/+) rats were very resistant to reinfection, suggesting a role for T lymphocytes (55). Similarly chickens treated with cyclosporin A, which prevents proliferation of T-lymphocytes, were not immune to secondary infection and had increased oocyst output compared with untreated controls (56). Transfer of cell mediated immunity (CMI) is also possible; adoptive transfer of peripheral blood lymphocytes and splenocytes from infected, immunized chickens resulted in the transfer of antibody producing cells as well as CMI when given intravenously to uninfected chickens, as shown by reduced oocyst output following E. maxima infection (57). Lymphoproliferative responses of peripheral blood lymphocytes to sporozoite antigen appear to correlate with resistance to E. maxima in different inbred chicken lines in primary and secondary infection, whilst antibody responses are inversely correlated to resistance to E. maxima and E. tenella (58). In the mouse CD4+ T-cells, and to a lesser extent CD8+ T cells, appear to be more important in controlling primary Eimeria infection, whilst CD8+ T cells appear to be more important in secondary infections (59). However in chickens, several studies have suggested there are differences in the cell populations involved in controlling infection by different Eimeria species, in single or mixed species infections (60, 61). A more recent study showed increased proportions of cytotoxic CD8+(TCRγδ−CD8β+) cells following primary infection but the proportion of TCRγδ−CD8β+ in the caeca was only maintained for 3 days following secondary E. tenella infection before gradually declining (62). The same study demonstrated induced CD107a+ cell surface mobilization on cytotoxic TCRγδ−CD8β+cells, a marker of recent degranulation, following primary and secondary infection with E. tenella (62). In secondary E. acervulina infection the percentage of CD8+ intraepithelial cells is increased, and one study has demonstrated a greater proportion of CD8+ intraepithelial lymphocytes in resistant chickens 10 days post infection (63). Variability in T-cell responses might also be seen between different chicken lines, presumably due to genetic variation (64). The role of CD8+ cells in mediating resistance to secondary infection has been proposed to be killing of infected epithelial cells (65).
In early studies, cytokines produced by T-lymphocytes were presumed to mediate immune responses to E. tenella and E. acervulina infection in chickens injected with T-cell supernatants (66). The role of interferon-gamma (IFN-γ) was initially examined in E. vermiformis infected BALB/c mice, where blocking of endogenous IFN-γ by an IFN-γ specific monoclonal antibody in-vivo resulted in increased susceptibility to primary but not secondary infection, and increased oocyst production during primary infection compared with mice not injected with the antibody (67). The role for cytokines in primary but not secondary infection has been supported by a qPCR study examining cytokine transcripts following E. maxima infection that demonstrated upregulation of both Th1 and Th2 cytokines in primary but not secondary infection (68). Studies suggest that antigen specific IFN-γ responses from infected chicken splenic cells do not peak until days 20–25 post infection (69, 70), or even up to 35 days after primary infection in one study (71). During E. tenella infection the specific IFN- γ response in peripheral blood lymphocytes appears to occur earlier, around 8 days post infection, and may coincide with an increase in CD8+ cells in the peripheral blood (72). The role of other cytokines such as tumor necrosis factor (TNF) and interleukin (IL)-10 in controlling Eimeria infection has also been explored. TNF production by macrophages is upregulated following Eimeria infection, with peak production coinciding with the incidence of intestinal lesions, leading the authors to be speculate that release of this pro-inflammatory cytokine may enhance disease pathology (73). Another study found it to be upregulated in primary but not secondary infection with E. tenella and found differences between levels of TNF produced by different inbred chicken lines (74). IL-10 mRNA transcripts were found to be upregulated in the spleen and small intestine of susceptible chickens after infection with E. maxima (75). Similarly, in E. tenella infection IL-10 mRNA was upregulated in the caeca post-infection (76), although there does not appear to be a clear relationship between IL-10 expression during infection and caecal pathology (33). Other cytokines might be linked to Eimeria-associated pathology (77), but further work is needed to define the underlying mechanisms. It is clear than although many cytokines are upregulated during Eimeria infection and the primary response in particular, no single cytokine is a clear biomarker for disease susceptibility or progression; although this could be because of the heterogenous cell populations/tissues assayed.
The role of macrophages in controlling Eimeria infection has also been investigated. Assessment of phagocytosis of Eimeria sporozoites by chicken peritoneal macrophages in-vitro showed low levels for E. tenella and, to a lesser extent, E. maxima in macrophages taken from non-infected chickens. Phagocytosis by peritoneal macrophages from infected chickens was increased but varied depending on the stage and number of infections, and was enhanced by immune serum. However, it has to be stressed that peritoneal macrophages have been described to be functionally very different to tissue-resident macrophages in the gut (78), and may have a more immunomodulatory role rather than an antigen-presenting function (79). Quantification of macrophages during caecal infection with E. tenella showed higher absolute numbers in immune birds at 8 h post infection compared with naïve birds, however there was a greater increase in macrophages in naïve birds compared with immune birds (80). The same study also demonstrated that sporozoites were more likely to be located within or next to macrophages in the caeca of naïve birds compared with immune birds, where sporozoites were also more often located within or next to CD8+ cells (80). The role of macrophages is therefore somewhat unclear, although they are likely to play an important role as antigen presenting cells to CD4+ T cells in primary infection and might also act as effector cells, potentially destroying parasites through the production of reactive oxygen species. Natural killer (NK) cells, mononuclear cells with cytotoxic activity, might also play a role in Eimeria infection although this is still unclear and may be mediated through IFN- γ (81).
It is clear that there is no single immune correlate of protection against Eimeria in the chicken. Circulating antibodies, IgY, IgM, and IgA, that are specific against Eimeria can be detected in infected chickens and ELISAs to measure serum antibody levels to Eimeria species in chickens have been developed (82, 83). Detection of antibodies raised against novel anticoccidial antigens post vaccination might be of some relevance in assessing antigenicity and can provide evidence of successful delivery. However, as serum antibody levels are not central to immune protection following Eimeria infection their relevance in assessment of vaccine efficacy is minimal. In natural immunity to Eimeria infection a cell mediated response is most important and presumably an efficacious vaccine will need to generate a similar response, ideally without inducing pathology. Measuring cell mediated responses to vaccination is more challenging than measuring serum antibodies, both technically and to interpret. Peripheral blood is the most easily acquired sample, although cell mediated responses in the periphery may differ compared with local responses in the gut. Examining cell mediated responses in organs such as the gut and spleen can only be achieved post mortem and, therefore, pre and post vaccine samples are not available for any individual chicken.
In the absence of clear identifiable immunological correlates of protection other relevant parameters must be used to assess vaccine efficacy. Reduction or absence of clinical disease can be assessed by observing individual chickens for signs of disease and more quantitatively by assessment of pathognomonic lesions in the gut. A standardized approach to scoring pathological lesions described by Johnson and Reid (28) is still widely utilized; a lesion scoring system from 0 to +4 was adopted for each Eimeria species with varying score criteria based on the infecting species and the nature of any lesions assessed during peak infection. Use of the lesion scoring system requires training from an experienced user or pathologist and can be subject to individual interpretation. It is therefore prudent to use the same individual or ideally multiple individuals to score all groups within each study and to consider that lesion scores might not be comparable between different studies examined by different individuals. Johnson and Reid (28) suggested that for some species such as E. maxima, lesion score does not appear to correlate with pathogenicity. Association between lesion scores and the clinical impact of coccidiosis appears more nuanced. For example, in one study chickens immunized with live oocysts and then challenged with a high E. tenella dose demonstrated high lesion scores but not the same reduction in body weight as unvaccinated challenged birds (84). Later studies confirmed that although some vaccinated chickens developed lesions, parasites did not appear to be associated with these lesions, possibly implicating the pro-inflammatory immune response (85).
Comparison of Eimeria replication through measurement of parasite load or output in vaccinated, mock vaccinated and/or unvaccinated groups is a common method for assessing vaccine efficacy. Traditional measures include microscopy to count oocysts excreted per gram (OPG) of feces or persisting in litter, or total oocysts excreted per unit of time (e.g., per day), with vaccine efficacy calculated as the difference between vaccinated and unvaccinated groups or individuals. Oocyst counts from total fecal material collected per day, or throughout the parasite's patent period, are the gold standard, removing variation incurred by differences in fecal consistency. For reliable oocyst counts from litter, sampling should ideally be randomized, collecting litter from multiple pre-determined points to avoid unconscious bias and ensure representation from different individuals within the group, although this is obviously difficult to achieve for large groups of chickens (86, 87). Sampling to obtain oocyst counts for multiple species of Eimeria should be undertaken with caution due to a variance in fecundity between species, with more fecund species potentially masking the presence of less fecund species.
Quantitative PCR (qPCR) is a viable alternative to oocyst counts as a method to quantify the number of Eimeria genomes in a sample. Assays have been developed and validated for quantification of all seven Eimeria species that infect chickens, using genomic DNA extracted from purified oocysts and fecal samples (88). For E. maxima, qPCR has been applied to total genomic DNA extracted from jejunum/ileum tissues adjacent to Meckel's diverticulum to define replication dynamics in chicken lines with varying susceptibility to infection (89). For E. tenella, total genomic DNA extracted from caecal tissue was used as template to quantify parasite genome numbers, highlighting a significant association between genome number and size of the oocyst dose used to initiate the infection. This method has the advantage of requiring lower group numbers due to within reduced intra-group variation between chickens given the same oocyst challenge dose compared with fecal oocyst counts (27). DNA extracted directly from feces can be unsuitable due to the presence of inhibitors which can impact qPCR efficiency and therefore quantification accuracy, although an internal qPCR control can be used for standardization (90). Extraction from tissues or oocysts is preferable given lower occurrence of PCR inhibitors. Variation in oocyst sporulation rates can influence quantification by qPCR given that an unsporulated oocyst contains two genomes, while a fully sporulated oocyst contains eight. Some studies have utilized non-sporulated oocysts for DNA extraction to minimize the impact of sporulation rate on quantification (91). qPCR has also been applied to validate and standardize next generation sequencing analyses, such as deep sequencing or Eimeria species 18S rDNA PCR amplicons (92).
While microscopic quantification of oocyst numbers and qPCR are reproducible and highly sensitive techniques, parasite replication can be confounded by several parasite and host effects. Parasite effects include the reproductive potential of the parasite species and strain. Species such as E. acervulina are highly fecund, producing many millions of progeny oocysts per chicken. In contrast, species such as E. maxima are far less productive (93). Host effects include inherent genetic resistance or susceptibility, nutritional status and immune status as a consequence of prior exposure history (94, 95). Parasite replication can also be influenced by the crowding effect, a phenomenon characterized by reducing fecundity in response to increasing level of parasite challenge, although the mechanism remains unclear (43). Co-infection with other Eimeria species or other infectious agents might also influence oocyst output (96, 97).
Production measures are of utmost importance to the poultry industry when assessing any novel vaccine but can be difficult to measure and/or interpret in an experimental setting. Body weight gain is of primary significance during broiler production, but might only be impacted during severe infection where weight gain can be up to 60% lower than in uninfected chickens (98). However, experimental models of severe infection can be unpredictable and are undesirable from a welfare perspective. Williams and Catchpole (99) advocated the use of body weight gain up to 7 days post infection and feed conversion efficiency as measures of vaccine efficacy, and demonstrated the use of these criteria for the live attenuated vaccine Paracox™.
The anticoccidial index (ACI) has been developed as a method to evaluate resistance to anticoccidial medication (100, 101). McManus et al. (101) incorporated percentage survival, percentage relative weight gain, lesion score and oocyst count, combining pathological, parasitological and production traits. Jeffers (100) used the difference in weight between day 0 and day 7 post-infection and then subtracted the average score of fecal abnormality, where 0 equated to normal feces and 4 equated to no normal feces visible, divided by 10. Chapman and Shirley (102) used a modified version of the former protocol and assigned cut-offs for drug sensitivity; >160 = sensitive, partially resistant = 120–160 and <120 = resistant. Some vaccine studies have used ACI as a method to evaluate vaccine efficacy and assigned different cut-off scores for efficacy with some variation between studies; an ACI of <160 is generally considered to be ineffective (103–107). However, whether this adapted scoring method is overly simplistic to evaluate the efficacy and usefulness of vaccines in the field has yet to be determined.
Combining the information derived from measurement of multiple parameters and interpreting this information can be difficult as correlation is commonly poor between the different measurements of vaccine efficacy (99). This could be in part because parameters such as weight gain and feed conversion efficiency need to be assessed over a longer period of time than the 5–6 days post infection which is optimal for assessment of lesion score and parasite replication. Even for vaccines which are currently on the market and considered efficacious there is variability in vaccine response with some chickens developing pathological lesions, although lesion score is not necessarily associated with weight gain in these vaccinated birds (85). Furthermore, the variability in measurements of each parameter will be influenced by the population selected with commercial breeds more likely to show variability in response to infection and vaccination than more inbred lines. Studies have tried to model outcome parameters following Eimeria infection and fit them to an “infection index,” however this too can be difficult with models working more efficiently at certain inoculation doses. The time point of measuring infection outcomes is also likely to impact on correlation between parameters (108).
Assessment of novel vaccines to protect against Eimeria infection in chickens is becoming more and more relevant with the discovery of new antigens for different Eimeria species and the development of new methods for delivery. The lack of a standardized approach to assessment of coccidial vaccine efficacy hampers comparison between studies, with different chicken breeds, challenge models, husbandry systems and outcomes being assessed. Ultimately, any new anticoccidial vaccine must be easily deliverable and relevant to the target market. While reductions in parasite replication and lesion score are important and might be considered optimal, producers in the majority broiler sector prioritize improvements in feed conversion efficiency and weight gain. As well as these considerations, future studies should adopt a methodical approach to animal study design and reporting, ideally conforming to the ARRIVE guidelines to maximize robustness, repeatability and transparency (109). Reporting of negative or inconclusive results must also be considered. A better understanding of response and variability to both infection and vaccination is imperative in informing future study design and minimizing animals used whilst maximizing outputs.
This manuscript has been assigned the reference PPS_01XXX by the RVC.
FS led preparation of this review, with input from DW, FT, and DB.
This study was funded by the Biotechnology and Biological Sciences Research Council (BBSRC) through the grant BB/P003931/1.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2020.00101/full#supplementary-material
1. Dalloul RA, Lillehoj HS. Poultry coccidiosis: recent advancements in control measures and vaccine development. Expert Rev Vaccines. (2006) 5:143–63. doi: 10.1586/14760584.5.1.143
2. Antonissen G, Eeckhaut V, Van Driessche K, Onrust L, Haesebrouck F, Ducatelle R, et al. Microbial shifts associated with necrotic enteritis. Avian Pathol. (2016) 45:308–12. doi: 10.1080/03079457.2016.1152625
3. Karavolias J, Salois MJ, Baker KT, Watkins K. Raised without antibiotics: impact on animal welfare and implications for food policy. Transl Anim Sci. (2018) 2:337–48. doi: 10.1093/tas/txy016
4. Peek HW, Landman WJ. Coccidiosis in poultry: anticoccidial products, vaccines and other prevention strategies. Vet Q. (2011) 31:143–61. doi: 10.1080/01652176.2011.605247
5. Chapman HD, Jeffers TK. Vaccination of chickens against coccidiosis ameliorates drug resistance in commercial poultry production. Int J Parasitol Drugs Drug Resist. (2014) 4:214–7. doi: 10.1016/j.ijpddr.2014.10.002
6. Chapman HD. Milestones in avian coccidiosis research: a review. Poult Sci. (2014) 93:501–11. doi: 10.3382/ps.2013-03634
7. Rose ME. Some aspects of immunity to Eimeria infections. Ann N Y Acad Sci. (1963) 113:383–99. doi: 10.1111/j.1749-6632.1963.tb40677.x
8. Blake DP, Hesketh P, Archer A, Carroll F, Shirley MW, Smith AL. The influence of immunizing dose size and schedule on immunity to subsequent challenge with antigenically distinct strains of Eimeria maxima. Avian Pathol. (2005) 34:489–94. doi: 10.1080/03079450500368292
9. Boulton K, Nolan MJ, Wu Z, Riggio V, Matika O, Harman K, et al. Dissecting the genomic architecture of resistance to Eimeria maxima parasitism in the chicken. Front Genet. (2018) 9:528. doi: 10.3389/fgene.2018.00528
10. Brake DA, Fedor CH, Werner BW, Miller TJ, Taylor RL Jr, Clare RA. Characterization of immune response to Eimeria tenella antigens in a natural immunity model with hosts which differ serologically at the B locus of the major histocompatibility complex. Infect Immun. (1997) 65:1204–10. doi: 10.1128/IAI.65.4.1204-1210.1997
11. Blake DP, Billington KJ, Copestake SL, Oakes RD, Quail MA, Wan KL, et al. Genetic mapping identifies novel highly protective antigens for an apicomplexan parasite. PLoS Pathog. (2011) 7:e1001279. doi: 10.1371/journal.ppat.1001279
12. Joyner LP. Immunological variation between two strains of Eimeria acervulina. Parasitology. (1969) 59:725–32. doi: 10.1017/S0031182000031243
13. Blake DP, Pastor-Fernandez I, Nolan MJ, Tomley FM. Recombinant anticoccidial vaccines - a cup half full? Infect Genet Evol. (2017) 55:358–65. doi: 10.1016/j.meegid.2017.10.009
14. Chapman HD. The effect of monensin on the immunity arising from repeated low-level infections with Eimeria maxima, E. brunetti and E tenella Avian Pathol. (1978) 7:269–77. doi: 10.1080/03079457808418278
15. Williams RB. Fifty years of anticoccidial vaccines for poultry (1952-2002). Avian Dis. (2002) 46:775–802. doi: 10.1637/0005-2086(2002)046[0775:FYOAVF]2.0.CO;2
16. Lillehoj HS, Ruff MD. Comparison of disease susceptibility and subclass-specific antibody response in SC and FP chickens experimentally inoculated with Eimeria tenella, E. acervulina, or E maxima Avian Dis. (1987) 31:112–9. doi: 10.2307/1590782
17. Jenkins MC, Parker C, O'Brien C, Persyn J, Barlow D, Miska K, et al. Protecting chickens against coccidiosis in floor pens by administering Eimeria oocysts using gel beads or spray vaccination. Avian Dis. (2013) 57:622–6. doi: 10.1637/10516-022213-Reg.1
18. Price K, Guerin M, Barta J. Success and failure: the role of relative humidity levels and environmental management in live Eimeria vaccination of cage-reared replacement layer pullets. J Appl Poult Res. (2014) 23:523–35. doi: 10.3382/japr.2014-00989
19. Price K, Hafeez M, Bulfon J, Barta J. Live Eimeria vaccination success in the face of artificial non-uniform vaccine administration in conventionally reared pullets. Avian Pathology. (2016) 45:82–93. doi: 10.1080/03079457.2015.1125442
20. Shirley MW, Bedrnik P. Live attenuated vaccines against avian coccidiosis: success with precocious and egg-adapted lines of Eimeria. Parasitol Today. (1997) 13:481–4. doi: 10.1016/S0169-4758(97)01153-8
21. McDonald V, Shirley MW. The endogenous development of virulent strains and attenuated precocious lines of Eimeria tenella and E. necatrix J Parasitol. (1987) 73:993–7. doi: 10.2307/3282523
22. McDonald V, Ballingall S. Further investigation of the pathogenicity, immunogenicity and stability of precocious Eimeria acervulina. Parasitology. (1983) 86(Pt 3):361–9. doi: 10.1017/S0031182000050551
23. Long PL. Development of Eimeria tenella in avian embryos. Nature. (1965) 208:509–10. doi: 10.1038/208509a0
24. Long PL, Millard BJ, Batty AF, da Vison C. Immunisation against coccidiosis in chickens: tests under simulated field conditions. Avian Pathol. (1982) 11:131–44. doi: 10.1080/03079458208436088
25. Shirley MW, McDonald V, Bellatti MA. Eimeria brunetti: selection and characteristics of a precocious (and attenuated) line. Avian Pathol. (1986) 15:705–17. doi: 10.1080/03079458608436333
26. Tannenbaum J, Bennett BT. Russell and Burch's 3Rs then and now: the need for clarity in definition and purpose. J Am Assoc Lab Anim Sci. (2015) 54:120–32.
27. Nolan MJ, Tomley FM, Kaiser P, Blake DP. Quantitative real-time PCR (qPCR) for Eimeria tenella replication - implications for experimental refinement and animal welfare. Parasitol Int. (2015) 64:464–70. doi: 10.1016/j.parint.2015.06.010
28. Johnson J, Reid WM. Anticoccidial drugs: lesion scoring techniques in battery and floor-pen experiments with chickens. Exp Parasitol. (1970) 28:30–6. doi: 10.1016/0014-4894(70)90063-9
29. Bumstead N, Millard BJ. Variation in susceptibility of inbred lines of chickens to seven species of Eimeria. Parasitology. (1992) 104(Pt 3):407–13. doi: 10.1017/S0031182000063654
30. Caron LA, Abplanalp H, Taylor RL Jr. Resistance, susceptibility, and immunity to Eimeria tenella in major histocompatibility (B) complex congenic lines. Poult Sci. (1997) 76:677–82. doi: 10.1093/ps/76.5.677
31. Pinard-Van Der Laan MH, Monvoisin JL, Pery P, Hamet N, Thomas M. Comparison of outbred lines of chickens for resistance to experimental infection with coccidiosis (Eimeria tenella). Poult Sci. (1998) 77:185–91. doi: 10.1093/ps/77.2.185
32. Smith AL, Hesketh P, Archer A, Shirley MW. Antigenic diversity in Eimeria maxima and the influence of host genetics and immunization schedule on cross-protective immunity. Infect Immun. (2002) 70:2472–9. doi: 10.1128/IAI.70.5.2472-2479.2002
33. Boulton K, Nolan MJ, Wu Z, Psifidi A, Riggio V, Harman K, et al. Phenotypic and genetic variation in the response of chickens to Eimeria tenella induced coccidiosis. Genet Sel Evol. (2018) 50:63. doi: 10.1186/s12711-018-0433-7
34. Lillehoj HS, Ruff MD, Bacon LD, Lamont SJ, Jeffers TK. Genetic control of immunity to Eimeria tenella. Interaction of MHC genes and non-MHC linked genes influences levels of disease susceptibility in chickens. Vet Immunol Immunopathol. (1989) 20:135–48. doi: 10.1016/0165-2427(89)90094-9
35. Clare RA, Danforth HD. Major histocompatibility complex control of immunity elicited by genetically engineered Eimeria tenella (Apicomplexa) antigen in chickens. Infect Immun. (1989) 57:701–5. doi: 10.1128/IAI.57.3.701-705.1989
36. Lillehoj HS, Jenkins MC, Bacon LD. Effects of major histocompatibility genes and antigen delivery on induction of protective mucosal immunity to E. acervulina following immunization with a recombinant merozoite antigen. Immunology. (1990) 71:127–32.
37. Nochi T, Jansen CA, Toyomizu M, van Eden W. The well-developed mucosal immune systems of birds and mammals allow for similar approaches of mucosal vaccination in both types of animals. Front Nutr. (2018) 5:60. doi: 10.3389/fnut.2018.00060
38. Zhou H, Chen S, Wang MS, Cheng AC. Interferons and their receptors in birds: a comparison of gene structure, phylogenetic analysis, and cross modulation. Int J Mol Sci. (2014) 15:21045–68. doi: 10.3390/ijms151121045
39. Heitlinger E, Spork S, Lucius R, Dieterich C. The genome of Eimeria falciformis–reduction and specialization in a single host apicomplexan parasite. BMC Genomics. (2014) 15:696. doi: 10.1186/1471-2164-15-696
40. Reid AJ, Blake DP, Ansari HR, Billington K, Browne HP, Bryant J, et al. Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Res. (2014) 24:1676–85. doi: 10.1101/gr.168955.113
41. Abolins S, King EC, Lazarou L, Weldon L, Hughes L, Drescher P, et al. The comparative immunology of wild and laboratory mice, Mus musculus domesticus. Nat Commun. (2017) 8:14811. doi: 10.1038/ncomms14811
42. Smith AL, Hayday AC. Genetic analysis of the essential components of the immunoprotective response to infection with Eimeria vermiformis. Int J Parasitol. (1998) 28:1061–9. doi: 10.1016/S0020-7519(98)00081-2
43. Williams RB. Quantification of the crowding effect during infections with the seven Eimeria species of the domesticated fowl: its importance for experimental designs and the production of oocyst stocks. Int J Parasitol. (2001) 31:1056–69. doi: 10.1016/S0020-7519(01)00235-1
44. Ruff MD, Doran DJ, Wilkins GC. Effect of aging on survival and pathogenicity of Eimeria acervulina and Eimeria tenella. Avian Dis. (1981) 25:595–9. doi: 10.2307/1589989
45. Lawn AM, Rose ME. Mucosal transport of Eimeria tenella in the cecum of the chicken. J Parasitol. (1982) 68:1117–23. doi: 10.2307/3281101
46. Trout JM, Lillehoj HS. Evidence of a role for intestinal CD8+ lymphocytes and macrophages in transport of Eimeria acervulina sporozoites. J Parasitol. (1993) 79:790–2. doi: 10.2307/3283625
47. Vervelde L, Vermeulen AN, Jeurissen SH. Eimeria tenella: sporozoites rarely enter leukocytes in the cecal epithelium of the chicken (Gallus domesticus). Exp Parasitol. (1995) 81:29–38. doi: 10.1006/expr.1995.1089
48. Long PL, Pierce AE. Role of cellular factors in the mediation of immunity to avian coccidiosis (Eimeria tenella). Nature. (1963) 200:426–7. doi: 10.1038/200426a0
49. Davis PJ, Parry SH, Porter P. The role of secretory IgA in anti-coccidial immunity in the chicken. Immunology. (1978) 34:879–88.
50. Witlock DR, Danforth HD. Surface changes induced by immune serum on Eimeria tenella sporozoites and merozoites. J Protozool. (1982) 29:441–5. doi: 10.1111/j.1550-7408.1982.tb05429.x
51. Crane MS, Murray PK, Gnozzio MJ, MacDonald TT. Passive protection of chickens against Eimeria tenella infection by monoclonal antibody. Infect Immun. (1988) 56:972–6. doi: 10.1128/IAI.56.4.972-976.1988
52. Davis PJ, Porter P. A mechanism for secretory IgA-mediated inhibition of the cell penetration and intracellular development of Eimeria tenella. Immunology. (1979) 36:471–7.
53. Rose ME, Hesketh P. Eimeria tenella: effects of immunity on sporozoites within the lumen of the small intestine. Exp Parasitol. (1987) 63:337–44. doi: 10.1016/0014-4894(87)90181-0
54. Rose ME. Immunity to Eimeria infections. Vet Immunol Immunopathol. (1987) 17:333–43. doi: 10.1016/0165-2427(87)90152-8
55. Rose ME, Hesketh P. Immunity to coccidiosis: T-lymphocyte- or B-lymphocyte-deficient animals. Infect Immun. (1979) 26:630–7. doi: 10.1128/IAI.26.2.630-637.1979
56. Lillehoj HS. Effects of immunosuppression on avian coccidiosis: cyclosporin A but not hormonal bursectomy abrogates host protective immunity. Infect Immun. (1987) 55:1616–21. doi: 10.1128/IAI.55.7.1616-1621.1987
57. Rose ME, Hesketh P. Immunity to coccidia in chickens: adoptive transfer with peripheral blood lymphocytes and spleen cells. Parasite Immunol. (1982) 4:171–185. doi: 10.1111/j.1365-3024.1982.tb00429.x
58. Bumstead JM, Bumstead N, Rothwell L, Tomley FM. Comparison of immune responses in inbred lines of chickens to Eimeria maxima and Eimeria tenella. Parasitology. (1995) 111(Pt 2):143–51. doi: 10.1017/S003118200006488X
59. Rose ME, Hesketh P, Wakelin D. Immune control of murine coccidiosis: CD4+ and CD8+ T lymphocytes contribute differentially in resistance to primary and secondary infections. Parasitology. (1992) 105(Pt 3):349–54. doi: 10.1017/S0031182000074515
60. Cornelissen JB, Swinkels WJ, Boersma WA, Rebel JM. Host response to simultaneous infections with Eimeria acervulina, maxima and tenella: a cumulation of single responses. Vet Parasitol. (2009) 162:58–66. doi: 10.1016/j.vetpar.2009.02.001
61. Trout JM, Lillehoj HS. T lymphocyte roles during Eimeria acervulina and Eimeria tenella infections. Vet Immunol Immunopathol. (1996) 53:163–72. doi: 10.1016/0165-2427(95)05544-4
62. Wattrang E, Thebo P, Lunden A, Dalgaard TS. Monitoring of local CD8beta-expressing cell populations during Eimeria tenella infection of naive and immune chickens. Parasite Immunol. (2016) 38:453–67. doi: 10.1111/pim.12331
63. Lillehoj HS. Analysis of Eimeria acervulina-induced changes in the intestinal T lymphocyte subpopulations in two chicken strains showing different levels of susceptibility to coccidiosis. Res Vet Sci. (1994) 56:1–7. doi: 10.1016/0034-5288(94)90188-0
64. Swinkels WJ, Post J, Cornelissen JB, Engel B, Boersma WJ, Rebel JM. Immune responses to an Eimeria acervulina infection in different broilers lines. Vet Immunol Immunopathol. (2007) 117:26–34. doi: 10.1016/j.vetimm.2007.01.020
65. Lillehoj HS, Trout JM. CD8+ T cell-coccidia interactions. Parasitol Today. (1994) 10:10–4. doi: 10.1016/0169-4758(94)90347-6
66. Lillehoj HS, Kang SY, Keller L, Sevoian M. Eimeria tenella and E. acervulina: lymphokines secreted by an avian T cell lymphoma or by sporozoite-stimulated immune T lymphocytes protect chickens against avian coccidiosis. Exp Parasitol. (1989) 69:54–64. doi: 10.1016/0014-4894(89)90171-9
67. Rose ME, Wakelin D, Hesketh P. Gamma interferon controls Eimeria vermiformis primary infection in BALB/c mice. Infect Immun. (1989) 57:1599–603. doi: 10.1128/IAI.57.5.1599-1603.1989
68. Hong YH, Lillehoj HS, Lillehoj EP, Lee SH. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet Immunol Immunopathol. (2006) 114:259–72. doi: 10.1016/j.vetimm.2006.08.006
69. Lillehoj HS. Immune response during coccidiosis in SC and FP chickens. I. In vitro assessment of T cell proliferation response to stage-specific parasite antigens. Vet Immunol Immunopathol. (1986) 13:321–30. doi: 10.1016/0165-2427(86)90025-5
70. Rose ME, Hesketh P. Infection with Eimeria tenella: modulation of lymphocyte blastogenesis by specific antigen, and evidence for immunodepression. J Protozool. (1984) 31:549–53. doi: 10.1111/j.1550-7408.1984.tb05500.x
71. Prowse SJ, Pallister J. Interferon release as a measure of the T-cell response to coccidial antigens in chickens. Avian Pathol. (1989) 18:619–30. doi: 10.1080/03079458908418637
72. Breed DG, Dorrestein J, Schetters TP, Waart LV, Rijke E, Vermeulen AN. Peripheral blood lymphocytes from Eimeria tenella infected chickens produce gamma-interferon after stimulation in vitro. Parasite Immunol. (1997) 19:127–35. doi: 10.1046/j.1365-3024.1997.d01-191.x
73. Byrnes S, Eaton R, Kogut M. In vitro interleukin-1 and tumor necrosis factor-alpha production by macrophages from chickens infected with either Eimeria maxima or Eimeria tenella. Int J Parasitol. (1993) 23:639–45. doi: 10.1016/0020-7519(93)90170-4
74. Zhang S, Lillehoj HS, Ruff MD. In vivo role of tumor necrosis-like factor in Eimeria tenella infection. Avian Dis. (1995) 39:859–66. doi: 10.2307/1592424
75. Rothwell L, Young JR, Zoorob R, Whittaker CA, Hesketh P, Archer A, et al. Cloning and characterization of chicken IL-10 and its role in the immune response to Eimeria maxima. J Immunol. (2004) 173:2675–82. doi: 10.4049/jimmunol.173.4.2675
76. Haritova AM, Stanilova SA. Enhanced expression of IL-10 in contrast to IL-12B mRNA in poultry with experimental coccidiosis. Exp Parasitol. (2012) 132:378–82. doi: 10.1016/j.exppara.2012.08.017
77. Zhang L, Liu R, Song M, Hu Y, Pan B, Cai J, et al. Eimeria tenella: interleukin 17 contributes to host immunopathology in the gut during experimental infection. Exp Parasitol. (2013) 133:121–30. doi: 10.1016/j.exppara.2012.11.009
78. Rose ME. Immune responses in infections with coccidia: macrophage activity. Infect Immun. (1974) 10:862–71. doi: 10.1128/IAI.10.4.862-871.1974
79. Cassado Ados A, D'Imperio Lima MR, Bortoluci KR. Revisiting mouse peritoneal macrophages: heterogeneity, development, and function. Front Immunol. (2015) 6:225. doi: 10.3389/fimmu.2015.00225
80. Vervelde L, Vermeulen AN, Jeurissen SH. In situ characterization of leucocyte subpopulations after infection with Eimeria tenella in chickens. Parasite Immunol. (1996) 18:247–56. doi: 10.1046/j.1365-3024.1996.d01-94.x
81. Ivanova DL, Denton SL, Fettel KD, Sondgeroth KS, Munoz Gutierrez J, Bangoura B, et al. Innate lymphoid cells in protection, pathology, and adaptive immunity during apicomplexan infection. Front Immunol. (2019) 10:196. doi: 10.3389/fimmu.2019.00196
82. Gilbert JM, Bhanushali JK, McDougald LR. An enzyme-linked immunosorbent assay for coccidiosis in chickens: correlation of antibody levels with prior exposure to coccidia in the laboratory and in the field. Avian Dis. (1988) 32:688–94. doi: 10.2307/1590986
83. Rose ME, Mockett AP. Antibodies to coccidia: detection by the enzyme-linked immunosorbent assay (ELISA). Parasite Immunol. (1983) 5:479–89. doi: 10.1111/j.1365-3024.1983.tb00762.x
84. Long PL, Johnson J, Wyatt RD. Eimeria tenella - clinical effects in partially immune and susceptible chickens. Poult Sci. (1980) 59:2221–4. doi: 10.3382/ps.0592221
85. Williams RB. Anticoccidial vaccination: the absence or reduction of numbers of endogenous parasites from gross lesions in immune chickens after virulent coccidial challenge. Avian Pathol. (2003) 32:535–43. doi: 10.1080/0307945031000154143
86. Hamzic E, Bed'Hom B, Juin H, Hawken R, Abrahamsen MS, Elsen JM, et al. Large-scale investigation of the parameters in response to Eimeria maxima challenge in broilers. J Anim Sci. (2015) 93:1830–40. doi: 10.2527/jas.2014-8592
87. Kumar S, Garg R, Moftah A, Clark EL, Macdonald SE, Chaudhry AS, et al. An optimised protocol for molecular identification of Eimeria from chickens. Vet Parasitol. (2014) 199:24–31. doi: 10.1016/j.vetpar.2013.09.026
88. Vrba V, Blake DP, Poplstein M. Quantitative real-time PCR assays for detection and quantification of all seven Eimeria species that infect the chicken. Vet Parasitol. (2010) 174:183–90. doi: 10.1016/j.vetpar.2010.09.006
89. Blake DP, Hesketh P, Archer A, Shirley MW, Smith AL. Eimeria maxima: the influence of host genotype on parasite reproduction as revealed by quantitative real-time PCR. Int J Parasitol. (2006) 36:97–105. doi: 10.1016/j.ijpara.2005.09.011
90. Raj GD, Aarthi S, Selvabharathi R, Raman M, Blake DP, Tomley FM. Real-time PCR-based quantification of Eimeria genomes: a method to outweigh underestimation of genome numbers due to PCR inhibition. Avian Pathology. (2013) 42:304–8. doi: 10.1080/03079457.2013.790531
91. Peek HW, ter Veen C, Dijkman R, Landman WJM. Validation of a quantitative Eimeria spp. PCR for fresh droppings of broiler chickens. Avian Pathology. (2017) 46:615–22. doi: 10.1080/03079457.2017.1337269
92. Hinsu AT, Thakkar JR, Koringa PG, Vrba V, Jakhesara SJ, Psifidi A, et al. Illumina next generation sequencing for the analysis of Eimeria populations in commercial broilers and indigenous chickens. Front Vet Sci. (2018) 5:176. doi: 10.3389/fvets.2018.00176
93. Long PL, Millard BJ, Joyner LP, Norton CC. A guide to laboratory techniques used in the study and diagnosis of avian coccidiosis. Folia Vet Lat. (1976) 6:201–17.
94. Brackett S, Bliznick A. The reproductive potential of five species of coccidia of the chicken as demonstrated by oocyst production. J Parasitol. (1952) 38:133–9. doi: 10.2307/3273831
95. Sakkas P, Oikeh I, Blake DP, Smith S, Kyriazakis I. Dietary vitamin D improves performance and bone mineralisation, but increases parasite replication and compromises gut health in Eimeria-infected broilers. Br J Nutr. (2019) 122:676–88. doi: 10.1017/S0007114519001375
96. Hein HE. Eimeria acervulina, E. brunetti, and E maxima: pathogenic effects of single or mixed infections with low doses of oocysts in chickens. Exp Parasitol. (1976) 39:415–21. doi: 10.1016/0014-4894(76)90045-X
97. Williams RB. The effect of Eimeria acervulina on the reproductive potentials of four other species of chicken coccidia during concurrent infections. Br Vet J. (1973) 129:xxix–xxxi. doi: 10.1016/S0007-1935(17)36498-9
98. Conway DP, McKenzie ME, Dayton AD. Relationship of coccidial lesion scores and weight gain in infections of Eimeria acervulina, E. maxima and E tenella in broilers. Avian Pathol. (1990) 19:489–96. doi: 10.1080/03079459008418702
99. Williams RB, Catchpole J. A new protocol for a challenge test to assess the efficacy of live anticoccidial vaccines for chickens. Vaccine. (2000) 18:1178–85. doi: 10.1016/S0264-410X(99)00387-4
100. Jeffers TK. Eimeria acervulina and E. maxima: incidence and anticoccidial drug resistance of isolants in major broiler-producing areas. Avian Dis. (1974) 18:331–42. doi: 10.2307/1589101
101. McManus EC, Campbell WC, Cuckler AC. Development of resistance to quinoline coccidiostats under field and laboratory conditions. J Parasitol. (1968) 54:1190–3. doi: 10.2307/3276989
102. Chapman HD, Shirley MW. Sensitivity of field isolates of Eimeria species to monensin and lasalocid in the chicken. Res Vet Sci. (1989) 46:114–7. doi: 10.1016/S0034-5288(18)31129-9
103. Chen P, Lv J, Zhang J, Sun H, Chen Z, Li H, et al. Evaluation of immune protective efficacies of Eimeria tenella EtMic1 polypeptides with different domain recombination displayed on yeast surface. Exp Parasitol. (2015) 155:1–7. doi: 10.1016/j.exppara.2015.04.020
104. Huang J, Zhang Z, Li M, Song X, Yan R, Xu L, et al. Immune protection of microneme 7 (EmMIC7) against Eimeria maxima challenge in chickens. Avian Pathol. (2015) 44:392–400. doi: 10.1080/03079457.2015.1071780
105. Zhai Q, Huang B, Dong H, Zhao Q, Zhu S, Liang S, et al. Molecular characterization and immune protection of a new conserved hypothetical protein of Eimeria tenella. PLoS ONE. (2016) 11:e0157678. doi: 10.1371/journal.pone.0157678
106. Zhang J, Chen P, Sun H, Liu Q, Wang L, Wang T, et al. Pichia pastoris expressed EtMic2 protein as a potential vaccine against chicken coccidiosis. Vet Parasitol. (2014) 205:62–9. doi: 10.1016/j.vetpar.2014.06.029
107. Zhang Z, Liu X, Yang X, Liu L, Wang S, Lu M, et al. The molecular characterization and immunity identification of microneme 3 of Eimeria acervulina. J Eukaryot Microbiol. (2016) 63:709–21. doi: 10.1111/jeu.12318
108. Zhu JJ, Lillehoj HS, Allen PC, Yun CH, Pollock D, Sadjadi M, et al. Analysis of disease resistance-associated parameters in broiler chickens challenged with Eimeria maxima. Poult Sci. (2000) 79:619–25. doi: 10.1093/ps/79.5.619
Keywords: chickens, Eimeria, coccidiosis, vaccine development, in vivo trials
Citation: Soutter F, Werling D, Tomley FM and Blake DP (2020) Poultry Coccidiosis: Design and Interpretation of Vaccine Studies. Front. Vet. Sci. 7:101. doi: 10.3389/fvets.2020.00101
Received: 30 November 2019; Accepted: 11 February 2020;
Published: 26 February 2020.
Edited by:
Michael Kogut, United States Department of Agriculture, United StatesReviewed by:
Ahmed Ali, Beni Suef University, EgyptCopyright © 2020 Soutter, Werling, Tomley and Blake. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Damer P. Blake, ZGJsYWtlQHJ2Yy5hYy51aw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.