 Shannon G. M. Kirejczyk1*†
Shannon G. M. Kirejczyk1*† Rachel E. Burns2†
Rachel E. Burns2† Michael W. Hyatt3†
Michael W. Hyatt3† Michael J. Yabsley4,5
Michael J. Yabsley4,5 Julia M. Ter Beest6Zoltan S. Gyimesi6
Julia M. Ter Beest6Zoltan S. Gyimesi6 Robert J. Ossiboff7†Amelia Waltman7†
Robert J. Ossiboff7†Amelia Waltman7† Tracie A. Seimon7Rita McManamon1,8
Tracie A. Seimon7Rita McManamon1,8- 1Department of Pathology, College of Veterinary Medicine, University of Georgia, Athens, GA, United States
- 2Connecticut Veterinary Medical Diagnostic Laboratory, Department of Pathobiology and Veterinary Science, College of Agriculture, Health and Natural Resources, University of Connecticut, Storrs, CT, United States
- 3Adventure Aquarium, Camden, NJ, United States
- 4Southeastern Cooperative Wildlife Disease Study, College of Veterinary Medicine, University of Georgia, Athens, GA, United States
- 5Warnell School of Forestry and Natural Resources, University of Georgia, Athens, GA, United States
- 6Louisville Zoological Garden, Louisville, KY, United States
- 7The Wildlife Conservation Society, Bronx, NY, United States
- 8Zoo and Exotic Animal Pathology Service and Infectious Diseases Laboratory, Department of Small Animal Medicine and Surgery, College of Veterinary Medicine, University of Georgia, Athens, GA, United States
Sarcocystis falcatula is a well-known cause of fatal pneumonia in some birds, particularly Old World psittacines. Here we describe fatal sarcosystosis due to S. falcatula in 3 penguins (Family Spheniscidae) under managed care, including one African penguin (Spheniscus demersus), and two Southern rockhopper penguins (Eudyptes chrysocome). Randomly distributed foci of necrosis, inflammatory cell infiltrates, edema, and variable numbers of round to elongated protozoal schizonts were observed in sections of lung. Protozoal organisms exhibited strong immunoreactivity for Sarcocystis sp. antigen by immunohistochemistry. Apicomplexan and Sarcocystis genus-specific PCR assays and sequence analysis confirmed S. falcatula as the etiologic agent. These cases of fatal pneumonia attributed to S. falcatula expand the list of aberrant intermediate avian hosts, with particular implications for penguins.
Background
Causes of protozoal pneumonia in birds include Sarcocystis falcatula, Toxoplasma gondii, Plasmodium spp., Haemoproteus spp., and Isospora (formerly Atoxoplasma) spp. Such infections are characterized by necrotizing interstitial pneumonia, with air capillaries containing tachyzoites (T. gondii) or meronts/schizonts (S. falcatula and Plasmodium spp.) within endothelial cells and/or macrophages. Malaria and toxoplasmosis are well-described causes of interstitial pneumonia in penguins (1, 2). Although molecular evidence of Sarcocystis spp. infection was recently reported in Magellenic penguins in Brazil (3), fatal disease due to pulmonary sarcocystosis has not previously been described in penguins. In this case series, we report the clinical, gross, microscopic, and ultrastructural features of fatal S. falcatula infections in 3 penguins under managed care.
Case Presentations
Case 1 was a 27-year-old, male African penguin (Spheniscus demersus) housed at an aquarium in the United States. The bird was at the end of a molting period and had been depressed and lethargic for a couple days before being found dead. Necropsy revealed congested lungs, and a complete set of formalin-fixed tissues was submitted to the Connecticut Veterinary Medical Diagnostic Laboratory for histopathology. Opossums had been seen on the property where this penguin was housed, but not inside the exhibit.
Case 2 was a 5-year-old, female, Southern rockhopper penguin (Eudyptes chrysocome) from a different zoological institution. Due to renovation of the birds' indoor exhibit, the penguin and its flock were temporarily housed outdoors for 6 weeks in a completely meshed enclosure with access to a temperature-regulated pool. Penguins were given oral itraconazole1 (15 mg, once daily) for aspergillosis prophylaxis for the duration of the relocation. Two days prior to its death, the bird's appetite declined and it was observed floating in water, rather than exhibiting normal swimming and diving behavior. Physical examination revealed severe dyspnea and generalized weakness. The bird was anesthetized for further diagnostics but died shortly after induction. Post-mortem radiographs revealed increased soft tissue opacity in the lungs. Necropsy revealed dark red, wet lungs that sank in formalin (Figure 1A), a friable spleen, and edematous pericardial sac. There was a well-demarcated, 1-cm-diameter, white to yellow, raised, coelomic plaque on the inner surface of the ribs and a moderate amount of green fecal staining around the cloacal orifice. Following the death of this bird and the onset of warmer spring temperatures, the remaining flock was moved to an indoor, chilled holding area.
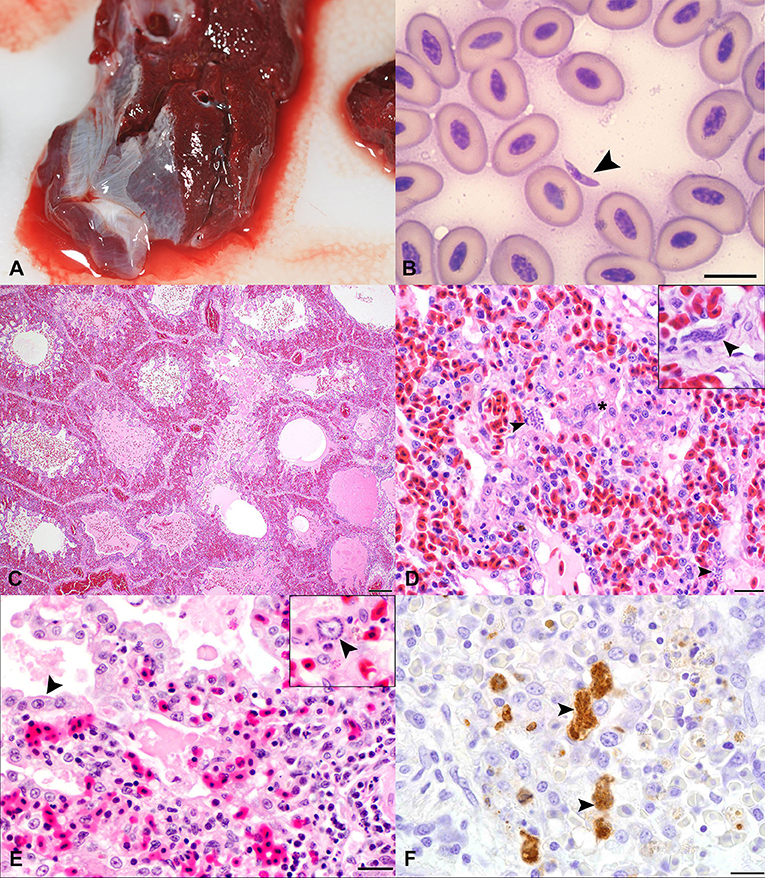
Figure 1. Gross, cytological, histopathologic, and immunohistochemical features of Sarcocystis falcatula infection in the lung of Southern rockhopper (Eudyptes chrysocome) and African (Spheniscus demersus) penguins. (A) Gross appearance of lung tissue from case 2 at necropsy. The lungs are dark red and edematous, with red-tinged fluid oozing along the periphery of the tissue. The lungs sank in formalin. (B) Cytology of tissue imprint from the lung of case 3 obtained at necropsy. The vast majority of cells are erythrocytes. A single, 2 × 8 micron, elongate to crescent-shaped tachyzoite with a small, round, slightly off-center nucleus is present at the center of the image (arrowhead). Bar = 10 μm. (C) Lung, from a 5-year-old, female, Southern rockhopper penguin (case 2), with S. falcatula pneumonia. Parabronchi and atria are frequently flooded by edema, hemorrhage and fibrin. H&E. Bar = 200 μm. (D) Lung, case 2. Air capillaries are multifocally obscured by foci of necrosis (*) containing protozoal schizonts (arrowheads), fibrin, hemorrhage, and low numbers of heterophils. The interstitium is hypercellular due to low numbers of lymphocytes and macrophages. H&E. Bar = 20 μm. Inset: High magnification image highlights the elongate to serpentine appearance of an intracellular schizont (case 3). (E) Lung, case 1. Pneumocytes lining the parabronchus in the top left of the image are hypertrophied (arrowhead) and the air space is filled with edema, fibrin, macrophages, and scant hemorrhage. The interstitium is expanded by inflammatory cell infiltrates, and multifocally obscured by necrosis, hemorrhage, and an accumulation of fibrin and edema. H&E. Bar = 20 μm. Inset: High magnification image with a schizont (arrowhead), which exhibits a “sunburst” pattern, with merozoites radiating around a clear zone. (F) Immunohistochemistry for S. neurona polyclonal antibody on lung tissue (case 2) demonstrates strongly immunoreactive, elongate to serpentine schizonts (arrowheads), free merozoites, and macrophages containing phagocytosed debris. DAB chromogen with hematoxylin counterstain. Bar = 10 μm.
Five days later, a 32-year-old, female, Southern rockhopper penguin (case 3) from the same institution began to exhibit weakness, anorexia and dyspnea, and auscultation revealed harsh lung sounds. The bird was treated with ponazuril2 (25 mg/kg PO, once), enrofloxacin3 (15 mg/kg SC, once), meloxicam4 (0.5 mg/kg IM, once), and furosemide5 (0.2 mg/kg IM, once) for presumed pulmonary edema, but died 1 h later. Necropsy examination revealed dark red, wet lungs, and an enlarged, nodular spleen. The caudal aspect of the left lung contained a small, focal, tan to brown, firm area.
Cytologic examination of lung tissue imprints from case 3 showed crescent-shaped, 2 × 4–8 μm protozoal zoites (Figure 1B), toxic heterophils and multinucleated giant cells. A complete set of tissues from cases 2 and 3 were fixed in 10% neutral-buffered formalin and submitted to the University of Georgia Zoo and Exotic Animal Pathology Service for histopathology. Fresh spleen from case 2 was submitted to Athens Veterinary Diagnostic Laboratory for aerobic and anaerobic cultures. Fresh lung and liver from Cases 2 and 3 were submitted to the Southeastern Cooperative Wildlife Disease Study (SCWDS, Athens, GA) for molecular testing.
Laboratory Investigations and Diagnostic Tests
Histopathology and Immunohistochemistry
Representative sections of all submitted tissues were routinely processed, embedded in paraffin wax, and 4–5-micron-thick sections were stained with Hematoxylin and Eosin (H&E) for light microscopy. For case 1, the only relevant immunohistochemical stain available at the Connecticut Veterinary Medical Diagnostic Laboratory was a rabbit polyclonal antibody against T. gondii (BioGenex, San Ramon, CA). Additional stains for Sarcocystis sp. (rabbit polyclonal antibody) and T. gondii (rabbit polyclonal antibody) were performed at the California Animal Health & Food Safety Laboratory System, Davis, CA as previously described (4). For cases 2 and 3, immunohistochemistry was performed at the University of Georgia College of Veterinary Medicine Histology Laboratory using antibodies for S. neurona (rabbit polyclonal antibody, 1:500 dilution for 60 min), Neospora caninum (goat polyclonal antibody6, 1:300 dilution for 30 min) and T. gondii (mouse monoclonal antibody6, 1:1,0000 dilution for 10 min).
The clinical, histologic, immunohistochemical, and molecular findings for three penguins with fatal S. falcatula infection are summarized in Table 1. All penguins had severe, necrotizing and lymphohistiocytic interstitial pneumonia. Parabronchi were flooded with hemorrhage, edema, and fibrin (Figure 1C). Air capillaries were obscured by foci of necrosis with fibrin exudation, and accumulations of heterophils, hemorrhage, and protozoal schizonts (Figure 1D). Air spaces were multifocally lined by hypertrophied epithelial cells (Figure 1E). In cases 2 and 3, schizonts were numerous and elongate, often conforming to the shape of capillaries (Figure 1D, inset), while schizonts were fewer and this classic serpentine morphology of S. falcatula was not observed in case 1. In all cases, schizonts occasionally exhibited a “sunburst” arrangement, in which merozoites radiated around a centralized clearing (Figure 1E, inset).
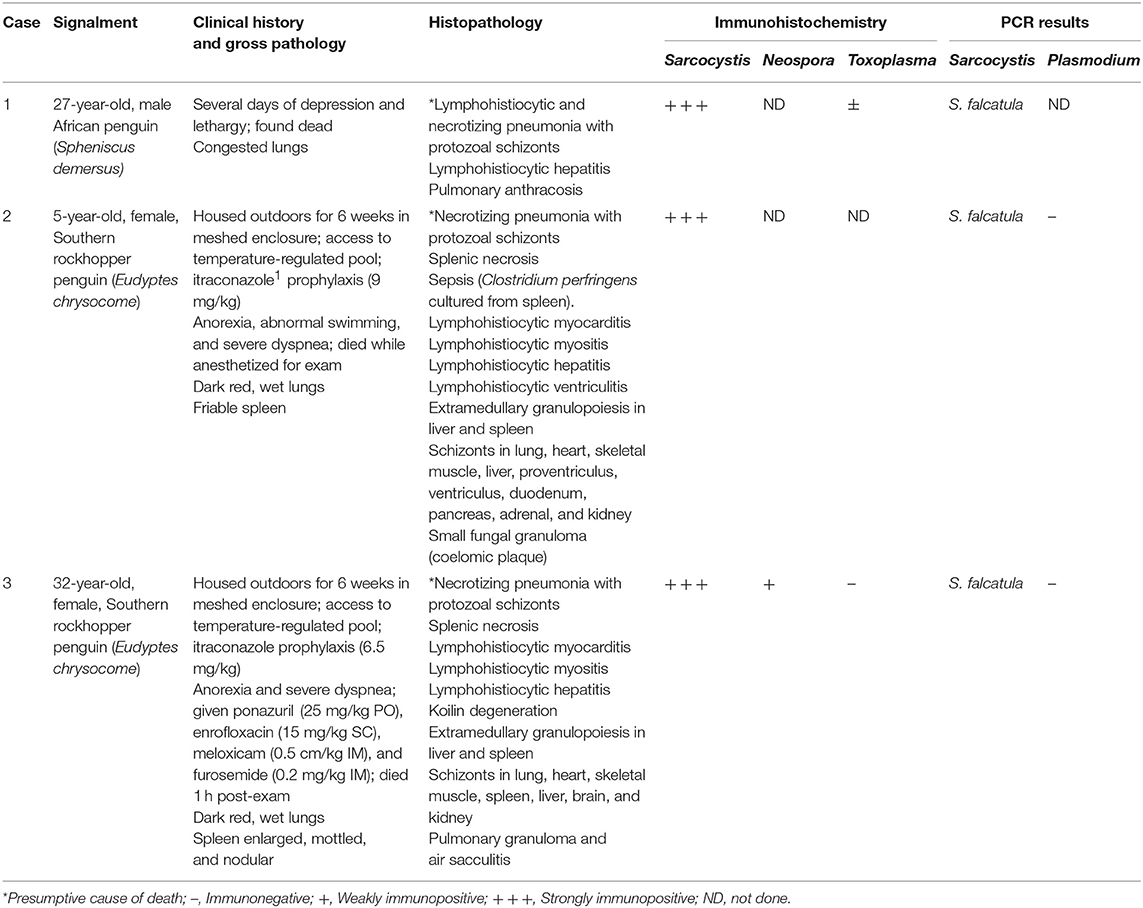
Table 1. Signalment, clinical history, gross pathology, histopathology, immunohistochemistry, and molecular findings for 3 penguins with S. falcatula infection.
A full set of tissues, including lung, liver, brain, skeletal muscle, and heart was examined for case 1. No extrapulmonary schizonts or sarcocysts were seen, and additional immunohistochemical stains were not pursued. Cases 2 and 3 exhibited mild lymphohistiocytic myocarditis and myositis and schizonts were observed in multiple tissues in these birds (Table 1). Protozoa stained variably PAS-positive on Periodic acid-Schiff reaction stains, and did not stain with Giemsa. On immunohistochemistry, protozoa exhibited strong immunoreactivity (Figure 1F) for polyclonal S. neurona antibodies and variable immunoreactivity for N. caninum and T. gondii antibodies (Table 1).
Cases 2 and 3 had markedly hypercellular spleens with foci of extramedullary granulopoiesis. There were also coalescing foci of coagulative to lytic necrosis, and case 3 had scattered, intraendothelial schizonts (Figure 2A). Under anaerobic conditions, Clostridium perfringens was cultured from the spleen of case 2. Splenic tissue from case 3 was not available for culture. Low numbers of schizonts were present in foci of splenic necrosis for case 3, so sarcosporidiosis is the most likely explanation for this lesion. However, given that low numbers of gram-positive bacilli in one section of skeletal muscle, septicemia cannot entirely be ruled out in case 3.
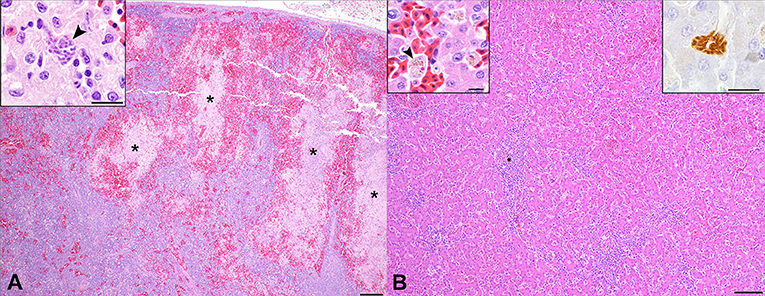
Figure 2. Hepatic and splenic pathology in Southern rockhopper penguins with fatal Sarcocystis falcatula infection. (A) Spleen, case 2. The splenic parenchyma is markedly hypercellular, with numerous hematopoietic cell precursors, lymphocytes, histiocytes, and plasma cells. There are multifocal to coalescing zones of coagulative to lytic necrosis (*) surrounded by a rim of hemorrhage. H&E. Bar = 200 μm. Inset: High magnification image of a S. falcatula schizont (arrowhead) within an area of necrosis in the spleen of case 3. Bar = 10 μm. (B) Liver, case 2. Portal regions are hypercellular, with moderate numbers of lymphocytes, plasma cells, macrophages and granulocytic precursor cells, which compress hepatocytes multifocally. Bar = 100 μm. Left insert: Sinusoids are congested, with macrophages containing phagocytosed cell debris, erythrocyte fragments and hemosiderin (arrowhead). H&E. Bar = 10 μm. Right inset: Immunohistochemistry for S. neurona highlights a schizont with radially arranged merozoites. Bar = 10 μm.
All cases had multifocal lymphohistiocytic portal hepatitis and extramedullary granulopoiesis (Figure 2B); cases 2 and 3 had low numbers of schizonts within sinusoids (Figure 2B, right inset). All cases also had moderate numbers of sinusoidal macrophages containing fragments of erythrocytes and intracytoplasmic hemosiderin (Figure 2B, left inset).
In addition to fulminant pulmonary sarcocystosis, case 2 had a small, coelomic fungal granuloma. Aspergillosis is suspected but fungal culture and/or PCR would be required for definitive diagnosis and further testing was not pursued. Case 3 had mild air sacculitis and a focal heterophilic granuloma at the caudal aspect of the left lung lobe. This lesion is believed to be related to a prior aspiration event, given the presence of foreign material and lack of microorganisms on special stains for fungi (Gomori Methenamine Silver), bacteria (modified Brown and Brenn Gram), and acid-fast bacilli (Ziehl-Neelsen).
Transmission Electron Microscopy
Formalin-fixed lung tissue from case 3 was trimmed into two, 2-mm-thick pieces and transferred to 2% paraformaldehyde, 2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.25. After overnight fixation, tissue was rinsed in 0.1 M phosphate buffer, post-fixed for 1 h with 1% buffered osmium tetroxide (OsO4), then rinsed in deionized water and dehydrated in an ascending ethanol series before infiltration with propylene oxide and Mollenhauer's Epon-Araldite resin mixture (5). Tissue samples were embedded in fresh resin mixture and allowed to polymerize in a 70°C oven. One-micron-thick sections were stained with Toluidine Blue O to select areas of interest before 60 nm sections were obtained and placed on grids. Grid sections were stained with uranyl acetate and lead citrate before examination with a JEOL JEM 1011 transmission electron microscope at 80 kV. Images were captured with an XR80M wide-angle multi-discipline mid-mount CCD camera7.
Transmission electron microscopy revealed protozoa with ultrastructural features compatible with Sarcocystis spp. (Figure 3) (6–8). Capillary endothelial cells occasionally contained an intracytoplasmic, serpentine schizont, which conformed to the cell shape. One schizont had a multilobulated nucleus with nuclear spindles arranged along the periphery (Figures 3A,B). Merozoites had a round, central nucleus, an anterior conoid and several tear-shaped, electron-dense micronemes at the posterior end (Figures 3C,D) (6).
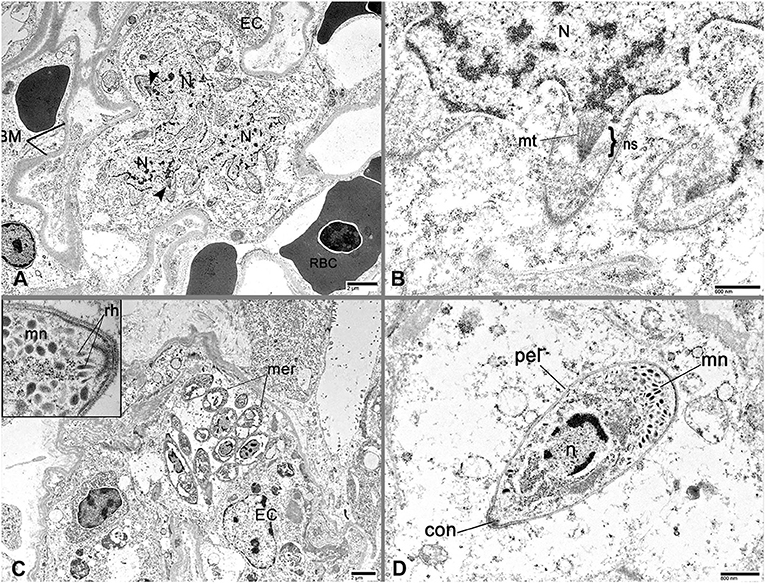
Figure 3. Ultrastructure of Sarcocystis falcatula asexual life stages by Transmission Electron Microscopy (TEM) in the lung of a 32-year-old, female, Southern rockhopper penguin (case 3). (A) TEM demonstrates an intraendothelial S. falcatula schizont. The schizont expands the endothelial cell (EC), is irregularly shaped and is separated from the EC cytoplasm by a thin membrane. Several nuclear spindles (arrowheads) project from the schizont nucleus (N). The endothelial cell abuts a capillary containing erythrocytes (RBC) and is supported by a basement membrane (BM). Bar = 2 μm. (B) Higher magnification TEM image of the S. falcatula schizont nucleus (N) from (A). Several conically arranged microtubules (mt) comprise the nuclear spindle (ns) of a developing merozoite. Bar = 600 nm. (C) TEM shows numerous, up to 2 × 6 μm merozoites clustered within an endothelial cell (EC). Inset: Higher magnification of a merozoite showing rhoptries (rh) and micronemes (mn). Bar = 2 μm. (D) A mature merozoite is present in a focus of necrotic lung tissue on TEM. The merozoite has a central, round nucleus (n), an anterior conoid (con), numerous micronemes (mn) at the posterior pole, and a double-membrane pellicle (pel). Bar = 800 nm.
Polymerase Chain Reaction (PCR) Analysis and Sequencing
For case 1, formalin-fixed, paraffin-embedded (FFPE) lung tissue was submitted to the Wildlife Conservation Society (Bronx, NY) and DNA was extracted using a QIAamp DNA FFPE Tissue Kit8. Extracts were tested by conventional PCR using both pan-apicomplexan and Sarcocystis-specific assays targeting the 18S rRNA and internal transcribed spacer 1 (ITS-1) regions, respectively. A small portion of the 18S rRNA gene was amplified as previously described (9). A portion of the ITS-1 region was amplified using primer P-ITSF and a degenerate primer (ShortITSR; 5′-GGGATTCARTKGYYGAAA-3′) designed based on publicly available Sarcocystis ITS-1 sequences (10). Amplicons were bi-directionally sequenced commercially, analyzed using Geneious R7 (Auckland, New Zealand) to generate a consensus sequence, trimmed of their primers, and analyzed by BLASTn.
The DNA sequence from the 18S gene was 100% identical to Sarcocystis falcatula in Genbank (MH626537, isolate Lorikeet ID #205850) but also identical to numerous other Sarcocystis isolates in GenBank. The DNA sequence from the ITS-1 region was 100% identical to Sarcocystis falcatula in GenBank (MH626538, isolate Lorikeet ID #205850) and Sarcocystis cf. falcatula in GenBank (AF389339) (11, 12). The next closest match for the ITS-1 region was Sarcocystis speeri and Sarcocystis neurona (98–99% identity).
For cases 2 and 3, DNA was extracted from fresh lung and liver using a commercial DNA extraction kit8 at SCWDS. The genus of the organism was determined by screening tissues with primers that amplify a short portion of the 18S rRNA gene of numerous apicomplexan parasites as previously described (13). Amplification products were visualized in 2% agarose gels stained with GelRed9. Amplicons were gel-purified using a kit8 and bi-directionally sequenced at the University of Georgia Genomics Facility (Athens, Georgia). Chromatograms were analyzed using Geneious R7 and the consensus sequence was compared to other sequences in GenBank.
To further characterize the Sarcocystis sp. in cases 2 and 3, partial cytochrome b gene was amplified with primers CYTB-F and CYTB-R, the ITS-1 region was amplified with primers ITS-234F19 and ITS-720R19, and the partial surface antigen 2 gene was amplified with primers SAG2-F1 and SAG2-R1 as previously described (14, 15). Amplicons were purified, sequenced, and analyzed as described above. To rule out the possibility of Plasmodium and/or Haemoproteus spp. infection in cases 2 and 3, a nested PCR targeting the mitochondrial cytochrome b (cytb) gene was conducted as previously described using primary primers HaemNFI and HaemNR3 and nested primers HaemF and HaemR2 (16).
For cases 2 and 3, liver and lung samples from both penguins were positive using the Tg18s58F and Tg18s348R PCR protocol and the resulting sequences (302 bp) were identical to each other and 100% similar to numerous Sarcocystis spp. The cytb gene sequences (580 bp) obtained from liver and spleen of cases 2 and 3 were identical and 100% similar to S. falcatula from captive bare-faced ibis (Phimosus infuscatus) from Brazil (KX265018) and Virginia opossum from California, USA (KP871704). The ITS-1 (287 bp) sequences were identical to each other and contained two polymorphic bases. These sequences were 98–99% similar to numerous S. falcatula strains from Brazil and the U.S.A. with some only differing at those two polymorphic bases. The four SAG2 sequences (402 bp) were identical and were 100% similar to the only S. falcatula (GQ851953) sequence available in GenBank. All four tissues were positive for the rpoB gene. The sequences were identical to each other and 100% similar to one S. falcatula (440 bp, KX265017) strain and >99% similar to several other S. falcatula sequences in GenBank (495 bp, e.g., AY164999).
Discussion
Death of these penguins was attributed to severe pneumonia caused by S. falcatula. The diagnosis was supported by IHC and confirmed by PCR. The differential diagnosis for apicomplexan pneumonia in penguins includes malaria, toxoplasmosis, and sarcocystosis. Fatal pulmonary infection with S. falcatula has been reported most often in psittacines (17–20), and the current report is believed to be the first in Spheniscidae. The histopathologic features of pneumonia in these penguins were similar to that described in other avian species infected with S. falcatula, namely necrosis, edema, fibrin deposition, congestion, hemorrhage, heterophilic, and mononuclear inflammatory infiltrates, endothelial cell lysis, and pneumocyte hyperplasia (7, 20–22).
The life cycle of Sarcocystis spp. involves definitive and intermediate hosts. In North America, the definitive host for S. falcatula and the closely related S. neurona is the Virginia opossum (Didelphis virginiana), which sheds infective sporocysts in feces (8). Infection of intermediate hosts typically occurs through ingestion of food contaminated with opossum feces, but insects can serve as mechanical vectors (17). In case 1, opossums were seen on the premises but not in the bird's enclosure, so ingestion of feces or sporozoite-containing fomite(s) is considered most likely. Contaminated water run-off or insect entry into the enclosure are additional possibilities.
In contrast to case 1 (African penguin), cases 2 and 3 (Southern rockhopper penguins) had disseminated infections with large numbers of schizonts. Potential factors that may have influenced the progression of disease in these animals include the dose of sporozoites ingested, time elapsed since infection, differences in host species susceptibility or pathogen virulence, and concomitant sepsis in case 2.
Acosta et al. (3) recently provided molecular evidence of Sarcocystis spp. infection in a cohort of Magellanic penguins (Spheniscus magellanicus) in Brazil. A Sarcocystis sp. closely related to S. falcatula was isolated from the pectoral muscle of 16 penguins, which were undergoing rehabilitation and died due to other causes. Given the tissue of origin (pectoral muscle), the authors postulated that the genetic material originated from tissue cysts, suggesting that the penguins survived an acute infection and formed sarcocysts. Sarcocysts were not identified in the penguins of the current report.
Cross-reactivity between cyst-forming apicomplexans has been reported for polyclonal antibodies targeting T. gondii, N. caninum, and Sarcocystis sp. (23–25). Although a previous study has shown a lack of cross-reactivity between a polyclonal S. neurona antibody and S. falcatula schizonts in budgerigars (Melopsittacus undulatus) (26), the S. neurona antibody used in all cases strongly reacted with S. falcatula in the tissue sections, which highlights the variability that can occur between IHC results based on different polyclonal antibodies.
This report expands the list of intermediate hosts for S. falcatula and underscores the importance of excluding opossums, their feces, and potential vectors from penguin enclosures. Additional studies are needed to determine individual and species-specific susceptibilities among penguins and whether or not infection and survival confers protective humoral immunity in individual birds. Prophylactic treatment for surviving penguins may be considered if sarcocystosis is diagnosed in a collection.
Data Availability Statement
All datasets generated for this study are included in the manuscript/supplementary files.
Ethics Statement
All animals included in this study were treated with the standard of care at each institution, as set forth by the Association of Zoos and Aquariums. An ethical review process was not required for this study, as the methods described herein were part of the routine diagnostic process to determine the cause of death of these penguins. All biological samples were obtained from penguins at postmortem exam, following the natural death of these animals.
Author Contributions
SK and RB drafted the manuscript. MH provided clinical care for case 1 and performed the post mortem exam. JT and ZG provided clinical care for cases 2 and 3 and performed the post mortem exams. RB performed the histopathology on case 1. SK and RM performed the histopathology on cases 2 and 3 and TEM on case 3. RO, TS, and AW designed and performed the PCR and sequencing and analyzed the sequencing data for case 1. MY designed and performed the PCR and sequencing and analyzed the sequencing data for cases 2 and 3. All authors assisted with writing and critically reviewing the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors wish to thank Denise Long and Ione Jackman from the CVMDL and Santiago Diab and Karen Sverlow from CAHFS-Davis for histology and immunohistochemistry of case 1. We also thank the staff of the UGA CVM Histology Laboratory for immunohistochemistry (cases 2 and 3), and Mary Ard at the Georgia Electron Microscopy Laboratory for assistance with Transmission Electron Microscopy.
Footnotes
1. ^Janssen Ortho LLC, Stateroad 933 Km 0 1 Street Statero, Gurabo, Puerto Rico 00778 USA. 100 mg capsules were opened and weighed out into 15 mg capsule doses by JT or ZG or by a compounding pharmacy (Taylors Pharmacy, 306 South Park Avenue, Winter Park, FL 32789 USA).
2. ^Compounded by Taylors Pharmacy, 306 South Park Avenue, Winter Park, FL 32789 USA. 50 mg/ml suspension.
3. ^Manufactured by Norbrook Laboratories Limited, Newry, BT35 6PU, Co. Down, Northern Ireland. 22.7 mg/ml.
4. ^Manufactured by Norbrook Laboratories Limited, Newry, BT35 6PU, Co. Down, Northern Ireland. 5 mg/ml.
5. ^Manufactured by Boehringer Ingelheim Vetmedica, Inc. 2621 North Belt Hwy, St. Joseph, MO 64506, USA.
6. ^Veterinary Medical Research & Development, 425 NW Albion Dr, Pullman, WA, 99163, U.S.A.
7. ^Advanced Microscopy Techniques, 242 West Cummings Park, Woburn, MA, 01801, U.S.A.
8. ^QIAGEN, Inc, 27220 Turnberry Lane 200, Valencia, CA 91355, U.S.A.
9. ^Biotium, Inc. 46117 Landing Pkwy, Fremont, CA 94538, U.S.A.
References
1. Ploeg M, Ultee T, Kik M. Disseminated toxoplasmosis in black-footed penguins (Spheniscus demersus). Avian Dis. (2011) 55:701–3. doi: 10.1637/9700-030411-Case.1
2. Vanstreels RE, da Silva-Filho RP, Kolesnikovas CK, Bhering RC, Ruoppolo V, Epiphanio S, et al. Epidemiology and pathology of avian malaria in penguins undergoing rehabilitation in Brazil. Vet Res. (2015) 46:30. doi: 10.1186/s13567-015-0160-9
3. Acosta ICL, Soares RM, Mayorga L, Alves BF, Soares HS, Gennari SM. Occurrence of tissue cyst forming coccidia in Magellanic penguins (Spheniscus magellanicus) rescued on the coast of Brazil. PLoS ONE. (2018) 13:e0209007. doi: 10.1371/journal.pone.0209007
4. Miller MA, Sverlow K, Crosbie PR, Barr BC, Lowenstine LJ, Gulland FM, et al. Isolation and characterization of two parasitic protozoa from a pacific harbor seal (Phoca vitulina richardsi) with meningoencephalomyelitis. J Parasitol. (2001) 87:816–22. doi: 10.1645/0022-3395(2001)087[0816:IACOTP]2.0.CO;2
5. Mollenhauer HH. Plastic embedding mixtures for use in electron microscopy. Stain Technol. (1964) 39:111–4.
6. Dubey JP. General biology. In: Dubey JPCS, and Fayer R, editors. Sarcocystosis of Animals and Man. 1st ed. Boca Raton, FL: CRC Press (1989). p. 1–81.
7. Smith JH, Meier JL, Neill PJG, Box ED. Pathogenesis of Sarcocystis falcatula in the budgerigar. II. Pulmonary pathology. Lab Invest. (1987) 56:72–84.
8. Smith JH, Meier JL, Neill PJG, Box ED. Pathogenesis of Sarcocystis falcatula in the budgerigar. I. Early pulmonary schizogony. Lab Invest. (1987) 56:60–71.
9. Sledge DG, Bolin SR, Lim A, Kaloustian LL, Heller RL, Carmona FM, et al. Outbreaks of severe enteric disease associated with Eimeria furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups. J Am Vet Med Assoc. (2011) 239:1584–8. doi: 10.2460/javma.239.12.1584
10. Kutkiene L, Prakas P, Sruoga A, Butkauskas D. The mallard duck (Anas platyrhynchos) as intermediate host for Sarcocystis wobeseri sp. nov. from the barnacle goose (Branta leucopsis). Parasitol Res. (2010) 107:879–88. doi: 10.1007/s00436-010-1945-4
11. Verma SK, Trupkiewicz JG, Georoff T, Dubey JP. Molecularly confirmed acute, fatal Sarcocystis falcatula infection in the rainbow lorikeets (Trichoglossus moluccanus) at the Philadelphia Zoo. J Parasitol. (2018) 104:710–2. doi: 10.1645/18-78
12. Dubey JP, Lindsay DS, Rosenthal BM, Kerber CE, Kasai N, Pena HFJ, et al. Isolates of Sarcocystis falcatula–like organisms from South American opossums Didelphis Marsupialis and Didelphis Albiventris from SÃo Paulo, Brazil. J Parasitol. (2001) 87:1449–53. doi: 10.1645/0022-3395(2001)087[1449:IOSFLO]2.0.CO;2
13. Su C, Shwab EK, Zhou P, Zhu XQ, Dubey JP. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology. (2010) 137:1–11. doi: 10.1017/S0031182009991065
14. Cesar MdO. Sarcocystis sp Eliminados por Didelphis aurita e Didelphis albiventris (Gambás) de vida livre no Estado de São Paulo: Infecção Experimental em Periquitos Australianos (Melopsittacus undulatus) e Camundongos Balb/c nude. São Paulo: Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo (2011).
15. Valadas SYOB. Caracterização Molecular de Isolados de SARCOCYSTIS spp. Obtidos de Marsupiais do Gênero Didelphis spp. Pela Análise de Gene Mitocondrial, Gene de Apicoplasto, Espaçador Interno Transcrito (ITS-1) e Genes Codificadores de Antígenos de Superfície (SAGs) [Doctoral]. São Paulo: Universidade de São Paulo. Faculdade de Medicina Veterinária e Zootecnia (2015).
16. Hellgren O, Waldenström J, Bensch S. A new PCR assay for simultaneous studies of leucocytozoon, plasmodium, and haemoproteus from avian blood. J Parasitol. (2004) 90:797–802. doi: 10.1645/GE-184R1
17. Clubb SL, Frenkel JK. Sarcocystis falcatula of opossums: transmission by cockroaches with fatal pulmonary disease in psittacine birds. J Parasitol. (1992) 78:116–24. doi: 10.2307/3283697
18. Godoy SN, De Paula CD, Cubas ZS, Matushima ER, Catao-Dias JL. Occurrence of Sarcocystis falcatula in captive psittacine birds in Brazil. J Avian Med Surg. (2009) 23:18–23. doi: 10.1647/2008-006R.1
19. Hillyer EV AM, Greiner EC, Atkinson CT, Frenkel JK. An Outbreak of Sarcocystis in a collection of psittacines. J Zoo Wildl Med. (1991) 22:434–45.
20. Villar D KM, Howard L, Hammond E, Cray C, Latimer K. Clinical presentation and pathology of sarcocystosis in psittaciform birds: 11 cases. Avian Dis. (2008) 52:187–94. doi: 10.1637/8104-090207-Case
21. Smith JH NP, Dillard EA, Box ED. Pathology of experimental Sarcocystis falcatula infections of canaries (Serinus canarius) and pigeons (Columba livia). J Parasitol. (1990) 76:59–68. doi: 10.2307/3282628
22. Smith JH, Craig TM, Dillard EA, Neill PJG, Jones LP. Naturally occurring apicomplexan acute interstitial pneumonitis in thick-billed parrots (Rhynchopsitta pachyrhyncha). J Parasitol. (1990) 76:285–8. doi: 10.2307/3283038
23. Barr BC, Conrad PA, Dubey JP, Anderson ML. Neospora-like encephalomyelitis in a calf: pathology, ultrastructure, and immunoreactivity. J Vet Diagn Invest. (1991) 3:39–46. doi: 10.1177/104063879100300109
24. Sundermann CA, Estridge BA, Branton MS, Bridgman CR, Lindsay DS. Immunohistochemical diagnosis of Toxoplasma gondii: potential for cross-reactivity with Neospora caninum. J Parasitol. (1997) 83:440–3. doi: 10.2307/3284407
25. Uzeda RS, Schares G, Ortega-Mora LM, Madruga CR, Aguado-Martinez A, Corbellini LG, et al. Combination of monoclonal antibodies improves immunohistochemical diagnosis of Neospora caninum. Vet Parasitol. (2013) 197(3–4):477–86. doi: 10.1016/j.vetpar.2013.07.008
Keywords: penguin, protozoal pneumonia, Sarcocystis falcatula, apicomplexa, Spheniscus demersus, Eudyptes chrysocome
Citation: Kirejczyk SGM, Burns RE, Hyatt MW, Yabsley MJ, Ter Beest JM, Gyimesi ZS, Ossiboff RJ, Waltman A, Seimon TA and McManamon R (2019) Fatal Sarcocystis falcatula Infection in Three Penguins. Front. Vet. Sci. 6:340. doi: 10.3389/fvets.2019.00340
Received: 16 July 2019; Accepted: 20 September 2019;
Published: 10 October 2019.
Edited by:
Amy Louise Warren, University of Calgary, CanadaReviewed by:
Frederique Nguyen, Agroalimentaire et de l'alimentation de Nantes-Atlantique, FranceFrancisco Ruben Carvallo Chaigneau, Virginia Tech, United States
Copyright © 2019 Kirejczyk, Burns, Hyatt, Yabsley, Ter Beest, Gyimesi, Ossiboff, Waltman, Seimon and McManamon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shannon G. M. Kirejczyk, c2tpcmVqY0BlbW9yeS5lZHU=
†Present address: Shannon G. M. Kirejczyk, Division of Pathology, Yerkes National Primate Research Center, Emory University, Atlanta, GA, United States
Rachel E. Burns, Disease Investigations, Institute for Conservation Research, San Diego Zoo Global, San Diego, CA, United States
Michael W. Hyatt, Wildlife Conservation Society, New York Aquarium, Brooklyn, NY, United States
Robert Ossiboff, Aquatic, Amphibian, and Reptile Pathology Program, College of Veterinary Medicine, University of Florida, Gainesville, FL, United States
Amelia Waltman, Pritzker School of Medicine, University of Chicago, Chicago, IL, United States