 Maryne Jaÿ1†‡
Maryne Jaÿ1†‡ Guillaume Girault1‡Ludivine Perrot1Benoit Taunay1Thomas Vuilmet1Frédérique Rossignol2Pierre-Hugues Pitel3Elodie Picard4
Guillaume Girault1‡Ludivine Perrot1Benoit Taunay1Thomas Vuilmet1Frédérique Rossignol2Pierre-Hugues Pitel3Elodie Picard4 Claire Ponsart1
Claire Ponsart1 Virginie Mick1*
Virginie Mick1*- 1ANSES/Paris-Est University, EU/OIE/FAO and National Reference Laboratory for Brucellosis, Animal Health Laboratory, Maisons-Alfort, France
- 2DDPP de la Drôme, Valence, France
- 3LABEO, Caen, France
- 4LABEO-Orne, Alençon, France
Several Brucella isolates have been described in wild-caught and “exotic” amphibians from various continents and identified as B. inopinata-like strains. On the basis of epidemiological investigations conducted in June 2017 in France in a farm producing domestic frogs (Pelophylax ridibundus) for human consumption of frog's legs, potentially pathogenic bacteria were isolated from adults showing lesions (joint and subcutaneous abscesses). The bacteria were initially misidentified as Ochrobactrum anthropi using a commercial identification system, prior to being identified as Brucella spp. by MALDI-TOF assay. Classical phenotypic identification confirmed the Brucella genus, but did not make it possible to conclude unequivocally on species determination. Conventional and innovative bacteriological and molecular methods concluded that the investigated strain was very close to B. microti species, and not B. inopinata-like strains, as expected. The methods included growth kinetic, antimicrobial susceptibility testing, RT-PCR, Bruce-Ladder, Suis-Ladder, RFLP-PCR, AMOS-ERY, MLVA-16, the ectoine system, 16S rRNA and recA sequence analyses, the LPS pattern, in silico MLST-21, comparative whole-genome analyses (including average nucleotide identity ANI and whole-genome SNP analysis) and HRM-PCR assays. Minor polyphasic discrepancies, especially phage lysis and A-dominant agglutination patterns, as well as, small molecular divergences suggest the investigated strain should be considered a B. microti-like strain, raising concerns about its environmental persistence and unknown animal pathogenic and zoonotic potential as for other B. microti strains described to date.
Introduction
Based on bacteriological features, host preference and pathogenicity, the taxonomy of the Brucella genus (http://www.bacterio.net/brucella.html) currently identifies 12 species split into (i) “core” Brucella species, including the six “classical” species (Brucella melitensis, B. abortus, B. suis, B. canis, B. ovis, B. neotomae; http://www.oie.int/fr/normes/code-terrestre/acces-en-ligne/ < underline)>, B. ceti and B. pinnipedialis isolated from marine mammals (1, 2), and the recently described B. papionis from baboons (3), and ii) the emerging atypical Brucella species (4–6). The atypical Brucella species include fast-growing B. microti initially isolated from common voles (7) and reported from soil (8) and red fox (9), B. inopinata BO1 isolated from a breast implant (10), B. vulpis from red fox (11), as well as, unclassified isolates: BO2 isolated from a patient with chronic destructive pneumonia (12), probably representing a novel lineage of B. inopinata, and novel Australian rodent isolates (13). Interestingly, the atypical Brucella isolates are phenotypically close to Ochrobactrum spp., a soil-associated facultative human pathogen (14), but genetically close to the Brucella genus. Molecular data show that Australian rodent isolates are related to B. inopinata and strain BO2, although B. microti is genetically close to the core phylogenetic clade of Brucella, especially to B. suis 1330 (15).
Brucella infections have been described in wild-caught and captive-bred anuran species native to Africa, South and Central America, and Australia, from animals showing systemic or localized infections (16–22), as well as, from other apparently healthy individuals (23). These exotic frog strains are affiliated with the atypical Brucella group, genetically close to B. inopinata (24), (18).
Although human infections due to B. inopinata have been reported (10, 12), its zoonotic potential remains unclear. Likewise, the pathogenicity of atypical Brucella bacteria and their transmission among amphibians are unknown (25).
This study presents the isolation and phenotypic identification of a new Brucella field isolate from Pelophylax ridibundus, a domestic frog on a breeding farm, as well as, its in-depth genomic characterization.
Results and Discussion
Detection of a Presumptive Brucella Field Isolate From the Domestic Frog P. ridibundus
In June 2017, epidemiological investigations were conducted for research purposes on a frog farm in France breeding the first domesticated strain of P. ridibundus Rivan92®, selected by the French National Institute for Agricultural Research (INRA) for human consumption (frog's legs). Animals were sampled randomly from the farm, based on development stages and ponds (3 batches of tadpoles, 1 batch of 20 small frogs and 2 batches of 8 adults) for pan-pathogen examination. All the selected batches were apparently healthy except for one batch of adults that showed lesions: swollen joint (n = 1) and subcutaneous edema (n = 2), confirmed at necropsy. After necropsy, bacteriological analyses were performed on 6 pools of individuals (whole animal for early stages [20 g] and internal organs for adults), and on visible lesions. A number of regular, brownish colonies, reaching 2 mm after 48 h, were isolated from the only adult batch showing lesions. Testing using the commercial API20-NE identification system (Biomérieux, France) pointed to Ochrobactrum anthropi. MALDI-TOF assay (Bruker Daltonics, France) run on a spot of pure culture overlaid with 1 μL of HCCA matrix indicated Brucella spp. using the Biotyper Security-Related (SR) database (26). Brucella misidentification using commercial biochemical tests is frequently reported (27); (28), and can result in laboratory-acquired infections (29, 30). Isolates were subsequently sent to the national reference laboratory for reliable identification and refined characterization.
Phenotypic Identification
Standard phenotypic identification (31) confirmed the Brucella genus (Table 1), without concluding unequivocally on species determination. Interestingly, strain biotyping traits were not strictly consistent with the B. inopinata-like profile previously described in anurans, in particular due to phage lysis. Surprisingly, phenotypic features (Table 1) were closer to the B. microti reference strain CCM 4915, except for the A-dominant agglutination pattern, already described for one B. microti fox isolate (32).
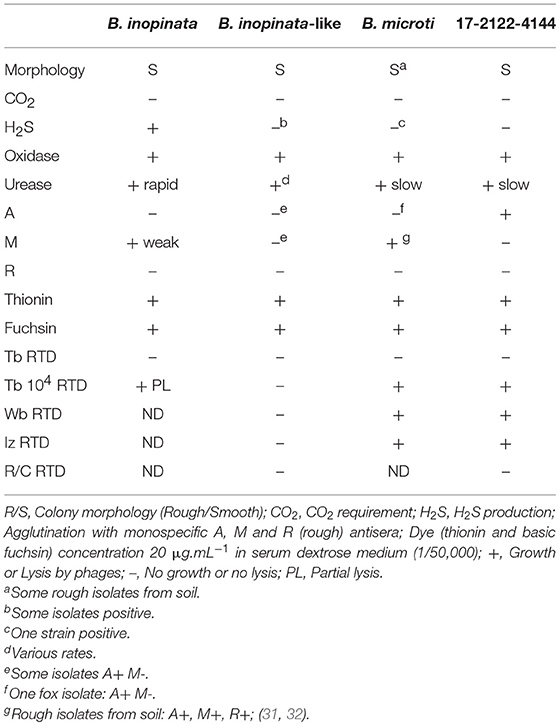
Table 1. Classical phenotypic characterization of the frog isolate investigated in this study vs. B. inopinata, B. inopinata-like strains isolated from exotic frogs, and B. microti field/reference strains.
Growth kinetics in nutritive tryptic soy and M9 minimal broths confirmed faster growth than classical fastidious Brucella for the investigated frog strain, named 17-2122-4144, with a generation time identical to B. microti CCM 4915 (i.e., approximately 4 h in our growth conditions).
Moreover, antimicrobial susceptibility testing (AST) performed by thedisk and E-test methods highlighted an identical pattern of susceptibility to the main anti-Brucella antibiotics of veterinary and human interest: doxycycline (DX), rifampicin (RIF), streptomycin (STM), ofloxacin (OFX), and sulfamethoxazole/trimethoprim (SMX/TMP) for the strain 17-2122-4144 vs B. microti CCM 4915.
Molecular Analysis
Conventional genus- and species-specific PCR methods (33) were performed (Table 2). The real-time PCR assays confirmed that the investigated strain belongs to the Brucella genus. The obtained Bruce-Ladder pattern was shared with the B. microti and B. suis biovar 2 reference strains and was distinct from other Brucella reference and vaccine strains. The Suis-Ladder method split the biovars of B. suis, B. canis and B. microti as previously described (32), and concluded that there was a single pattern between B. microti and the investigated strain.
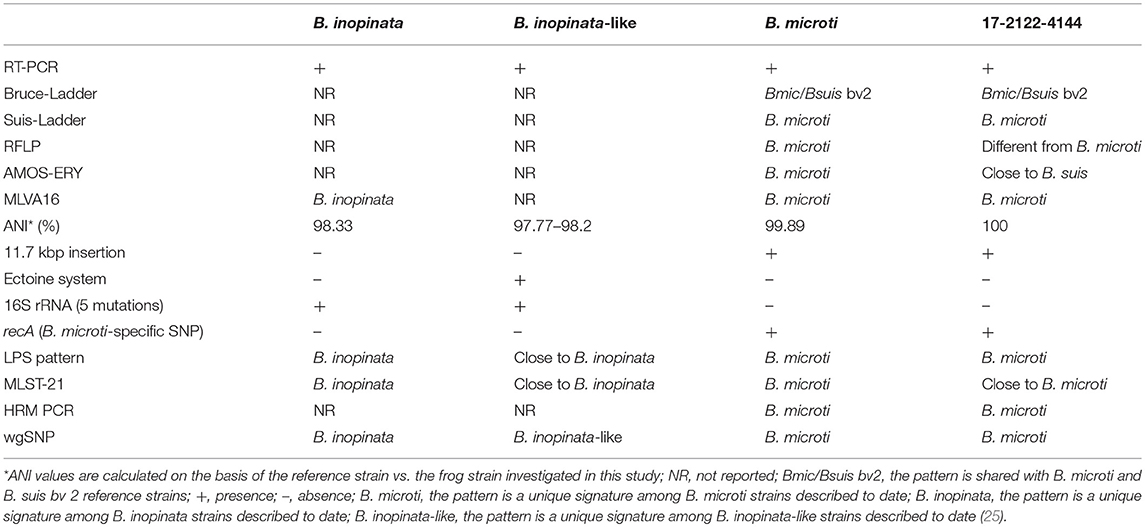
Table 2. Molecular characterization of the frog isolate investigated in this study vs. B. inopinata, B. inopinata-like strains isolated from exotic frogs, and B. microti field/reference strains.
Although most conventional molecular techniques did not make it possible to differentiate between CCM 4915 and 17-2122-4144, minor differences were observed regarding RFLP results (34): the restriction profile of the omp2b target digested by EcoRI for 17-2122-4144 was distinct from the CCM 4915 profile, but similar to the B. pinnipedialis reference strain B2/94. Interestingly, the AMOS-ERY profile of the studied strain (2 fragments of 1.3 kbp and 1.2 kbp) was divergent from classical Brucella spp. profiles, as well as, from the atypical B. microti (one single 1.3 kbp fragment), but close to B. suis reference strains (1.3 and 1.2 kbp).
In addition to classical molecular approaches, phylogenomic methods were used (Table 2). Unsurprisingly, MLVA-16 results showed that 17-2122-4144 clustered within B. microti reference strains CCM 4915 and CCM 4916 and together with the 10 field strains reported to date (Figure 1; Supplementary Figure 1), close to the B. neotomae reference strain 5K33 (32).
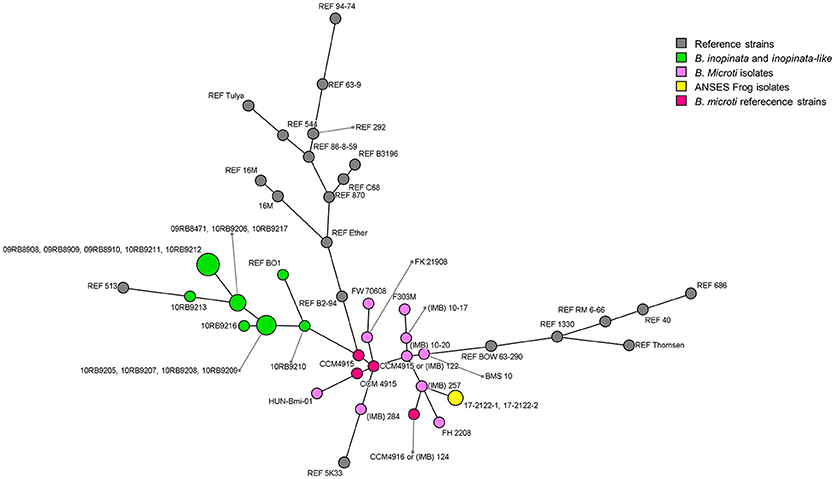
Figure 1. Minimum Spanning Tree of MLVA-16 genotypes of the frog strain investigated in this study, B. microti isolates published to date, and all Brucella reference strains. B. microti isolates are distinguished by different colors: yellow for the frog strain investigated in this study; pink for previously published isolates (32); red for B. microti reference strains; other reference strains are colored in gray.
De novo assembly showed a genome with a total length of 3,335,258 bp, vs 3.37 Mbp for B. inopinata BO1 and 3.34 Mbp for B. microti CCM 4915. Moreover, the total number of predicted genes per genome (evaluated by QUAST) for 17-2122-4144 (3,141 genes) is very similar to CCM 4915 (3,145 genes), closer than for BO1 (3,220 genes). ANI exhibited maximum identity with B. microti CCM 4915 (99.89%); 98.33% identity with B. inopinata and 97.77–98.2% with 3 frog Brucella genomes from the NCBI database (24). Similarly, a bacteriophage-related 11,742 bp insertion, previously described as present only in B. microti isolates (15), was also found within the investigated genome. Further analyses using Roary and Scoary to compare gene presence or absence did not underline any gene signature specific to the investigated field isolate vs. B. inopinata BO1, B. microti CCM 4915 and B. melitensis bv1 16M. Moreover, the ectoine system, conferring salt and temperature resistance, described in atypical Brucella (24), was absent in 17-2122-4144, as well as, BO1 and CCM 4915. Similarly, 16S rRNA and recA comparative analyses (27) confirmed that 17-2122-4144 was closely related to B. microti, with absence of 5 rrs mutations in 17-2122-4144 and CCM 4915, systematically present in B. inopinata and B. inopinata-like strains, and presence of a single recA B. microti-specific SNP in 17-2122-4144 ( 24, 25, 32).
In line with previous studies (4, 5, 25, 35), we assessed in silico the LPS profile of the investigated isolate, especially focusing on the genes essential for LPS synthesis: the wbk region, wboA and wboB genes, the manBCA region, as well as, the tagH and rfbD genes. Regions of the investigated isolate were strikingly similar to B. microti. In addition, our analysis concluded presence of the wboA, wboB and manBCA genes (unlike bullfrog strains, BO2 and B13-0095) and absence of the rmlACBD region and tagH gene found in BO2 and B13-0095 in the investigated genome. Interestingly, the rfbD gene was present in 17-2122-4144, but disrupted by numerous stop codons, as in B. microti CCM 4915. Our results show that the LPS profile of the novel isolate matches that described in B. microti.
In silico MultiLocus Sequence Typing-21 (MLST-21) confirmed this genetic proximity of 17-2122-4144 with B. microti CCM 4915. Except for the mutL gene involved in DNA mismatch repair, which harbored a point mutation at position 1149 (E383V), the MLST-21 pattern was strictly identical between the novel frog isolate and B. microti.
In parallel, B. microti and B. inopinata-specific High Resolution Melting (HRM) PCR assays were designed and performed against 17-2122-4144, emphasizing a profile similar to B. microti and divergent from B. inopinata. Phylogenetic comparative whole-genome SNP analysis showed that 17-2122-4144 is very close to B. microti CCM 4915 (323 SNPs without filtering, 73 SNPs in an overall phylogeny context) among the classical Brucella group (Figure 2; Supplementary Table 2), unlike strains previously isolated from frogs that clustered with B. inopinata in the “early-diverging” Brucella group (25).
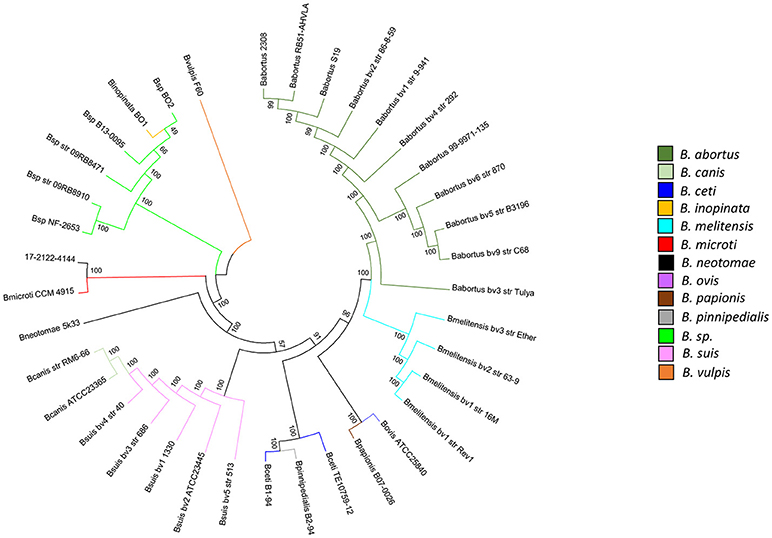
Figure 2. Phylogenetic comparative whole-genome SNP analysis of the frog strain investigated in this study and all Brucella reference strains. The dendrogram was constructed using the maximum likelihood method with 200 bootstrap repetitions (36,590 SNPs). Species are distinguished by different colors. A log scale is used in this tree, allowing a better distinction between isolates.
Taxonomic Conclusions
The investigated frog strain is very close to B. microti species, and not to B. inopinata-like strains, as might be expected given the current taxonomy of strains isolated from frogs. Despite minor polyphasic discrepancies, 17-2122-4144 is qualified as a B. microti-like strain.
B. microti has been isolated from wild animals, such as the common vole Microtus arvalis (36), (7), wild boars (37), and red foxes (9), and is described as persistent over a long period in soil (8), suggesting the existence of environmental reservoirs. Interestingly, although B. microti is suspected to induce epizootic mortality in the common vole (36), isolated cases from other described hosts seem to be asymptomatic, with no associated clinical signs (9, 37), suggesting asymptomatic carriage. In addition, the replication ability of B. microti was demonstrated in mouse macrophages (25, 38) and its pathogenic potential was shown to cause death in murine models (38–40) and lesions in chicken embryo models (41).
Anthropogenic interference has previously been reported to impact brucellosis prevalence in wildlife (42), raising questions on the influence of natural selection and selective breeding on B. microti fitness. Long-term environmental persistence outside the host and the putative ubiquitous nature of the B. microti-like strain investigated in this study, as well as, its unknown—but suspected—animal pathogenic and zoonotic potential, raise possible concerns for animal and public health. Further epidemiological investigations in wild frogs, as well as, in the natural environment might be required to offer new insights regarding bacterial carriage and possible clinical expression, depending on housing conditions. Moreover, in vitro cell infection experiments, as well as, in vivo infections will be required to determine the pathogenic potential of the B. microti-like isolates from frogs, in accordance with previous approaches applied to amphibian strains (25).
This study is the first isolation of B. microti-like bacteria from P. ridibundus on a domestic frog farm in France.
Materials and Methods
Bacterial Strains and Genomes
Strains and/or genomes used in this study are listed in Supplementary Table 1.
Phenotypic Identification
Isolates were characterized using standard procedures (31) in BSL-3 facilities. AST was performed by the disk (Thermo Scientific - Oxoid) and E-test (Biomerieux) diffusion methods on Mueller-Hinton agar plates, supplemented with 5% sheep blood (DX, RIF, STM, OFX, SMX/TMP), following the recommendations of the Clinical and Laboratory Standards Institute (43). Growth kinetics were performed in nutritive tryptic soy and on M9 minimal broths (44). Stationary phase cultures were diluted to an OD600 of 0.03 and grown in 75 cm2 cell culture flasks at 37°C. OD600 was measured every hour, and each point was serially diluted and plated on Brucella agar to determine colony-forming units. Each strain was assayed in triplicate.
Molecular Analysis
Genomic DNA was extracted using the High Pure PCR template preparation kit (Roche Diagnostics, France), according to the manufacturer's instructions.
Real-Time PCR (45), Bruce-Ladder (46), Suis-Ladder (47), RFLP-PCR (34), AMOS-ERY (48) and MLVA-16 (49) assays were performed as previously described. All tests have been carried out in duplicate (i.e., from 2 independent isolates). Clustering analysis was performed by using a minimum spanning tree (MST) and the cophenetic correlation coefficient with the UPGMA algorithm from MLVA data (Bionumerics v7.6.2; Applied Maths, Belgium), as well as, a maximum likelihood tree based on the Jukes Cantor model (with 200 repetitions for bootstrap) from WGS data (Bionumerics 7.6.2 and MEGA software v. 6).
Whole-genome sequencing (Illumina HiSeq2500 platform, 100X) was performed. De novo assembly was performed using the SPAdes v3.9 algorithm. QUAST 4.6.3 was used to assess assembly robustness by gathering extensive assembly statistics. Nucleotide sequences of contigs from this work were deposited in the European Nucleotide Archive (EMBL-EBI) –Bioproject: http://www.ebi.ac.uk/ena/data/view/PRJEB26927; Accession Number: ERZ654921–. Average Nucleotide Identity (ANI) values were calculated using Jspecies (50). Phylogenetic SNP distances were determined using the Bionumerics v7.6.2 wgSNP-module. Roary v3.6.1 and Scoary were used to generate and compare matrices of gene presence/absence. Polymorphism of 16S rRNA (27), recA (27), the ectoine system (24), the LPS pattern (4, 5, 25, 35) as well as, the presence of a bacteriophage-related 11,742 bp insertion (15) were studied as previously described, using Bionumerics v7.6.2 for multiple sequence alignments. The 21 locus scheme (MLST-21) was determined in silico as previously described (33, 51).
HRM PCR assays were carried out as previously described (52) using the Bmic_1F (5′-AACTGCCGGATGTGAAAAAG-3′) and Bmic_1R (5′-AAGGATCGAGGCGTCATAAA-3′) primers.
Author Contributions
MJ, GG, and VM designed the study and wrote the paper. FR, P-HP, and EP carried out preliminary identification studies. MJ and BT performed standard bacteriology. LP and GG performed growth kinetics and antimicrobial susceptibility tests. LP, BT, TV, and GG contributed to molecular studies. GG and VM performed bioinformatics analyses. GG, MJ, CP, and VM performed data interpretation. All authors read and approved the manuscript content.
Funding
The first steps of investigation were supported by a grant from the Conseil Regional de Normandie. This work was supported by the European Union Reference Laboratory for Brucellosis.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Vincent Sester and Madeleine Carle for their technical support and all the staff at local laboratory LVD61, in particular Kathy Rocton, and local and central veterinary services DDPP26, especially Françoise Jacquet.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2018.00283/full#supplementary-material
Supplementary Figure 1. MLVA-16 analysis of the frog strain investigated in this study, B. microti isolates published to date and all Brucella reference strains. The dendrogram was constructed with the cophenetic correlation coefficient and UPGMA algorithm. B. microti isolates are distinguished by different colors: yellow for the frog strain investigated in this study; pink for previously published isolates (32); red for B. microti reference strains; other reference strains are colored in gray.
Supplementary Table 1. Brucella strains used in this study.
Supplementary Table 2. List of identified SNPs among the novel frog isolate investigated in this study and Brucella strains, as well as, location of mutations found within 17-2122-4144 vs. CCM4915.
References
1. Cloeckaert A, Verger J-M, Grayon M, Paquet J-Y, Garin-Bastuji B, Foster G, et al. Classification of Brucella spp. isolated from marine mammals by DNA polymorphism at the omp2 locus. Microbes Infect Institut Pasteur. (2001) 3:729–38. doi: 10.1016/S1286-4579(01)01427-7
2. Foster G, Osterman BS, Godfroid J, Jacques I, Cloeckaert A. Brucella ceti sp. nov and Brucella pinnipedialis sp nov for Brucella strains with cetaceans and seals as their preferred hosts. Int J Syst Evol Microbiol. (2007) 57:2688–93. doi: 10.1099/ijs.0.65269-0
3. Whatmore AM, Davison N, Cloeckaert A, Al Dahouk S, Zygmunt MS, Brew SD, et al. Brucella papionis sp. nov, isolated from baboons (Papio spp.). Int J Syst Evol Microbiol. (2014) 64:4120–8. doi: 10.1099/ijs.0.065482-0
4. Wattam AR, Inzana TJ, Williams KP, Mane SP, Shukla M, Almeida NF, et al. Comparative genomics of early-diverging Brucella strains reveals a novel lipopolysaccharide biosynthesis pathway. mBio (2012) 3:e00246–12. doi: 10.1128/mBio.00246-12
5. Zygmunt MS, Jacques I, Bernardet N, Cloeckaert A. Lipopolysaccharide heterogeneity in the atypical group of novel emerging Brucella species. Clin Vaccine Immunol. (2012) 19:1370–3. doi: 10.1128/CVI.00300-12
6. Scholz HC, Vergnaud G. Molecular characterisation of Brucella species. Rev Sci Tech Off Int Epiz. (2013) 32:149–62. doi: 10.20506/rst.32.1.2189
7. Scholz HC, Hubalek Z, Sedlacek I, Vergnaud G, Tomaso H, Al Dahouk S, et al. Brucella microti sp. nov, isolated from the common vole Microtus arvalis. Int J Syst Evol Microbiol. (2008) 58:375–82. doi: 10.1099/ijs.0.65356-0
8. Scholz HC, Hubalek Z, Nesvadbova J, Tomaso H, Vergnaud G, Le Fleche P, et al. Isolation of Brucella microti from soil. Emerg Infect Dis. (2008) 14:1316–7. doi: 10.3201/eid1408.080286
9. Scholz HC, Hofer E, Vergnaud G, Le Fleche P, Whatmore AM, Al Dahouk S, et al. Isolation of Brucella microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in lower Austria. Vector Borne Zoonotic Dis. (2009) 9:153–6. doi: 10.1089/vbz.2008.0036
10. Scholz HC, Nockler K, Gollner C, Bahn P, Vergnaud G, Tomaso H, et al. Brucella inopinata sp. nov, isolated from a breast implant infection. Int J Syst Evol Microbiol. (2010) 60:801–8. doi: 10.1099/ijs.0.011148-0
11. Scholz HC, Revilla-Fernandez S, Al Dahouk S, Hammerl JA, Zygmunt MS, Cloeckaert A, et al. Brucella vulpis sp. nov, a novel Brucella species isolated from mandibular lymph nodes of red foxes (Vulpes vulpes) in Austria. Int J Syst Evol Microbiol. (2016) 66:2090–8. doi: 10.1099/ijsem.0.000998
12. Tiller R, Gee J, Lonsway D, Gribble S, Bell S, Jennison A, et al. Identification of an unusual Brucella strain (BO2) from a lung biopsy in a 52 year-old patient with chronic destructive pneumonia. BMC Microbiol. (2010) 10:23. doi: 10.1186/1471-2180-10-23
13. Tiller RV, Gee JE, Frace MA, Taylor TK, Setubal JC, Hoffmaster AR, et al. Characterization of novel Brucella strains originating from wild native rodent species in North Queensland, Australia. Appl Environ Microbiol. (2010) 76:5837–45. doi: 10.1128/AEM.00620-10
14. Al Dahouk S, Scholz HC, Tomaso H, Bahn P, Gollner C, Karges W, et al. Differential phenotyping of Brucella species using a newly developed semi-automated metabolic system. BMC Microbiol. (2010) 10:269. doi: 10.1186/1471-2180-10-269
15. Audic S, Lescot M, Claverie JM, Scholz HC. Brucella microti: the genome sequence of an emerging pathogen. BMC Genomics (2012) 10:352. doi: 10.1186/1471-2164-10-352
16. Fischer D, Lorenz N, Heuser W, Kampfer P, Scholz HC, Lierz M. Abscesses associated with a Brucella inopinata-like bacterium in a big-eyed tree frog (Leptopelis vermiculatus). J Zoo Wildl Med. (2012) 43:625–8. doi: 10.1638/2011-0005R2.1
17. Whatmore AM, Dale EJ, Stubberfield E, Muchowski J, Koylass M, Dawson C, et al. Isolation of Brucella from a White's tree frog (Litoria caerulea). JMM Case Rep. (2015) 2:e17. doi: 10.1099/jmmcr.0.000017
18. Muhldorfer K, Wibbelt G, Szentiks CA, Fischer D, Scholz HC, Zschock M, et al. The role of 'atypical' Brucella in amphibians: are we facing novel emerging pathogens? J Appl Microbiol. (2017) 122:40–53. doi: 10.1111/jam.13326
19. Eisenberg T, Risse K, Schauerte N, Geiger C, Blom J, Scholz HC. Isolation of a novel 'atypical' Brucella strain from a bluespotted ribbontail ray (Taeniura lymma). Antonie Van Leeuwenhoek (2017) 110:221–34. doi: 10.1007/s10482-016-0792-4
20. Eisenberg T, Hamann HP, Kaim U, Schlez K, Seeger H, Schauerte N, et al. Isolation of potentially novel Brucella spp. from frogs. Appl Environ Microbiol. (2012) 78:3753–5. doi: 10.1128/AEM.07509-11
21. Helmick KE, Garner MM, Rhyan J, Bradway D. Clinicopathologic features of infection with novel Brucella organisms in captive waxy tree frogs (Phyllomedusa sauvagii) and Colorado River Toads (Incilius Alvarius). J Zoo Wildl Med. (2018) 49:153–61. doi: 10.1638/2017-0026R1.1
22. Soler-Llorens PF, Quance CR, Lawhon SD, Stuber TP, Edwards JF, Ficht TA, et al. A Brucella spp. isolate from a pac-man frog (Ceratophrys ornata) reveals characteristics departing from classical Brucellae. Front Cell Infect Microbiol. (2016) 6:116. doi: 10.3389/fcimb.2016.00116
23. Kimura M, Une Y, Suzuki M, Park ES, Imaoka K, Morikawa S. Isolation of Brucella inopinata-like bacteria from White's and Denny's tree frogs. Vector Borne Zoonotic Dis. (2017) 17:297–302. doi: 10.1089/vbz.2016.2027
24. Scholz HC, Muhldorfer K, Shilton C, Benedict S, Whatmore AM, Blom J, et al. The change of a medically important genus: worldwide occurrence of genetically diverse novel Brucella species in exotic frogs. PLoS ONE (2016) 11:e0168872. doi: 10.1371/journal.pone.0168872
25. Al Dahouk S, Kohler S, Occhialini A, Jimenez de Bagues MP, Hammerl JA, Eisenberg T, et al. Brucella spp. of amphibians comprise genomically diverse motile strains competent for replication in macrophages and survival in mammalian hosts. Sci Rep. (2017) 7:44420. doi: 10.1038/srep44420
26. Ferreira L, Vega Castano S, Sanchez-Juanes F, Gonzalez-Cabrero S, Menegotto F, Orduna-Domingo A, et al. Identification of Brucella by MALDI-TOF mass spectrometry. Fast and reliable identification from agar plates and blood cultures. PLoS ONE (2010) 5:e14235. doi: 10.1371/journal.pone.0014235
27. Scholz HC, Pfeffer M, Witte A, Neubauer H, Al Dahouk S, Wernery U, et al. Specific detection and differentiation of Ochrobactrum anthropi, Ochrobactrum intermedium and Brucella spp. by a multi-primer PCR that targets the recA gene. J Med Microbiol. (2008) 57:64–71. doi: 10.1099/jmm.0.47507-0
28. Poonawala H, Marrs Conner T, Peaper DR. The brief case: misidentification of brucella melitensis as Ochrobactrum anthropi by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS). J Clin Microbiol. (2018) 56:e00914–17. doi: 10.1128/JCM.00914-17
29. Traxler RM, Lehman MW, Bosserman EA, Guerra MA, Smith TL. A literature review of laboratory-acquired brucellosis. J Clin Microbiol. (2013) 51:3055–62. doi: 10.1128/JCM.00135-13
30. Yang J, Ren X, Xue W, Yu X. Misidentification of Brucella and a review of the literature. Rev Med Microbiol. (2015) 26:85–7. doi: 10.1097/MRM.0000000000000038
31. OIE. Brucellosis (Brucella abortus, B. melitensis and B. suis) (Infection with B. abortus, B. melitensis and B. suis). Paris: OIE Terrestrial Manual (2016).
32. Al Dahouk S, Hofer E, Tomaso H, Vergnaud G, Le Fleche P, Cloeckaert A, et al. Intraspecies biodiversity of the genetically homologous species Brucella microti. Appl Environ Microbiol. (2012) 78:1534–43. doi: 10.1128/AEM.06351-11
33. Whatmore AM. Current understanding of the genetic diversity of Brucella, an expanding genus of zoonotic pathogens. Infect Genet Evol. (2009) 9:1168–84. doi: 10.1016/j.meegid.2009.07.001
34. Al Dahouk S, Tomaso H, Prenger-Berninghoff E, Splettstoesser WD, Scholz HC, Neubauer H. Identification of Brucella species and biotypes using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP). Crit Rev Microbiol. (2005) 31:191–6. doi: 10.1080/10408410500304041
35. Cardoso PG, Macedo GC, Azevedo V, Oliveira SC. Brucella spp noncanonical LPS: structure, biosynthesis, and interaction with host immune system. Microb Cell Fact. (2006) 5:13. doi: 10.1186/1475-2859-5-13
36. Hubalek Z, Scholz HC, Sedlacek I, Melzer F, Sanogo YO, Nesvadbova J. Brucellosis of the common vole (Microtus arvalis). Vector Borne Zoonotic Dis. (2007) 7:679–87. doi: 10.1089/vbz.2007.0143
37. Ronai Z, Kreizinger Z, Dan A, Drees K, Foster JT, Banyai K, et al. First isolation and characterization of Brucella microti from wild boar. BMC Vet Res. (2015) 11:147. doi: 10.1186/s12917-015-0456-z
38. Jimenez de Bagues MP, Ouahrani-Bettache S, Quintana JF, Mitjana O, Hanna N, Bessoles S, et al. The new species Brucella microti replicates in macrophages and causes death in murine models of infection. J Infect Dis. (2010) 202:3–10. doi: 10.1086/653084
39. Jimenez de Bagues MP, de Martino A, Quintana JF, Alcaraz A, Pardo J. Course of infection with the emergent pathogen Brucella microti in immunocompromised mice. Infect Immun. (2011) 79:3934–9. doi: 10.1128/IAI.05542-11
40. Jimenez de Bagues MP, Iturralde M, Arias MA, Pardo J, Cloeckaert A, Zygmunt MS. The new strains Brucella inopinata BO1 and Brucella species 83–210 behave biologically like classic infectious Brucella species and cause death in murine models of infection. J Infect Dis. (2014) 210:467–72. doi: 10.1093/infdis/jiu102
41. Wareth G, Bottcher D, Melzer F, Shehata AA, Roesler U, Neubauer H, et al. Experimental infection of chicken embryos with recently described Brucella microti: pathogenicity and pathological findings. Comp Immunol Microbiol Infect Dis. (2015) 41:28–34. doi: 10.1016/j.cimid.2015.06.002
42. Mick V, Le Carrou G, Corde Y, Game Y, Jay M, Garin-Bastuji B. Brucella melitensis in France: persistence in wildlife and probable spillover from alpine ibex to domestic animals. PLoS ONE (2014) 9:e94168. doi: 10.1371/journal.pone.0094168
43. CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. Wayne: Clinical and Laboratory Standards Institute (2015).
44. Sambrook J, Fritsch E, Maniatis T. Molecular Cloning: A Laboratory Manual. Melbourne: C. S. H. Laboratory (1989).
45. Bounaadja L, Albert D, Chenais B, Henault S, Zygmunt MS, Poliak S, et al. Real-time PCR for identification of Brucella spp.: a comparative study of IS711, bcsp31 and per target genes. Vet Microbiol. (2009) 137:156–64. doi: 10.1016/j.vetmic.2008.12.023
46. Lopez-Goni I, Garcia-Yoldi D, Marin CM, de Miguel MJ, Munoz PM, Blasco JM, et al. Evaluation of a multiplex PCR assay (Bruce-ladder) for molecular typing of all Brucella species, including the vaccine strains. J Clin Microbiol. (2008) 46:3484–7. doi: 10.1128/JCM.00837-08
47. Lopez-Goni I, Garcia-Yoldi D, Marin CM, de Miguel MJ, Barquero-Calvo E, Guzman-Verri C, et al. New Bruce-ladder multiplex PCR assay for the biovar typing of Brucella suis and the discrimination of Brucella suis and Brucella canis. Vet Microbiol. (2011) 154:152–5. doi: 10.1016/j.vetmic.2011.06.035
48. Ocampo-Sosa AA, Aguero-Balbin J, Garcia-Lobo JM. Development of a new PCR assay to identify Brucella abortus biovars 5, 6 and 9 and the new subgroup 3b of biovar 3. Vet Microbiol. (2005) 110:41–51. doi: 10.1016/j.vetmic.2005.06.007
49. Le Fleche P, Jacques I, Grayon M, Al Dahouk S, Bouchon P, Denoeud F, et al. Evaluation and selection of tandem repeat loci for a Brucella MLVA typing assay. BMC Microbiol. (2006) 6:9. doi: 10.1186/1471-2180-6-9
50. Richter M, Rossello-Mora R. Shifting the genomic gold standard for the prokaryotic species definition. PNAS (2009) 106:19126–31. doi: 10.1073/pnas.0906412106
51. Whatmore AM, Koylass MS, Muchowski J, Edwards-Smallbone J, Gopaul KK, Perrett LL. Extended multilocus sequence analysis to describe the global population structure of the genus Brucella: phylogeography and relationship to biovars. Front Microbiol. (2016) 7:2049. doi: 10.3389/fmicb.2016.02049
Keywords: Brucellosis, Brucella microti, domestic frog, Pelophylax ridibundus, Europe
Citation: Jaÿ M, Girault G, Perrot L, Taunay B, Vuilmet T, Rossignol F, Pitel P-H, Picard E, Ponsart C and Mick V (2018) Phenotypic and Molecular Characterization of Brucella microti-Like Bacteria From a Domestic Marsh Frog (Pelophylax ridibundus). Front. Vet. Sci. 5:283. doi: 10.3389/fvets.2018.00283
Received: 29 June 2018; Accepted: 24 October 2018;
Published: 15 November 2018.
Edited by:
Holger C. Scholz, Institut für Mikrobiologie der Bundeswehr, GermanyReviewed by:
Yuehua Ke, Centers for Disease Control and Prevention (CDC), United StatesAlice Rebecca Wattam, Virginia Tech, United States
Alessandra Occhialini, Université de Montpellier, France
Adrian Whatmore, Animal and Plant Health Agency, United Kingdom
Copyright © 2018 Jaÿ, Girault, Perrot, Taunay, Vuilmet, Rossignol, Pitel, Picard, Ponsart and Mick. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Virginie Mick, dmlyZ2luaWUubWlja0BhbnNlcy5mcg==
‡Present Address: Maryne Jaÿ, ANSES, Laboratoire de Lyon, UMR Mycoplasmoses des Ruminants, Lyon, France Université de Lyon, VetAgro Sup, UMR Mycoplasmoses des Ruminants, Marcy L'Etoile, France
‡These authors have contributed equally to this work