
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Trop. Dis., 14 April 2025
Sec. Emerging Tropical Diseases
Volume 6 - 2025 | https://doi.org/10.3389/fitd.2025.1565116
 Chloé Bohers1
Chloé Bohers1 Marie Vazeille1Luidji Pascalin2Kevin Meignan2Anaïs Karch2Xavier De Lamballerie3,4
Marie Vazeille1Luidji Pascalin2Kevin Meignan2Anaïs Karch2Xavier De Lamballerie3,4 Anna-Bella Failloux1*
Anna-Bella Failloux1*We report that Aedes albopictus Paris collected in June 2023 were experimentally able to transmit at least three dengue virus (DENV) serotypes, from 14 days post-infection (dpi) for DENV-3 and DENV-4, and from 21 dpi for DENV-2. Given the growing number of dengue autochthonous cases reported in France, episodes of active transmission in Paris are expected as it was reported in 2023 in the greater Paris area.
The last dengue outbreak in Europe was reported in 1927–1928 in Greece, causing ~ 1 million cases and ~ 1,000 deaths. Aedes aegypti was the main vector (1). Dengue virus (DENV; Flavivirus, Flaviviridae) disappeared from continental Europe following the successful control of Ae. aegypti in 1953 (2). In 1979, the tiger mosquito Ae. albopictus, native to Asia, arrived in Europe in Albania (3); it is now present in 20 European countries (4). In the absence of Ae. aegypti, Ae. albopictus was the vector of the first dengue autochthonous cases in 2010, in France (5) and Croatia (6).
In France, Ae. albopictus was first detected in the Southeast region close to the Italian border in 2004 (7) and is now established in 78 of the 96 departments in mainland France including Paris (8). Since 2010, dengue autochthonous cases were reported each year with a peak of 45 cases in 2023 (9) and 83 cases in 2024 (10). While most autochthonous dengue cases were reported in the south of France, three local cases were detected in the greater Paris in September 2023 (11, 12); dengue 2 serotype has been identified, probably imported via infected travelers from Martinique and Guadeloupe (13). Here, we assess the vector competence of Ae. albopictus Paris to the four DENV serotypes and discuss the epidemiological consequences and vector control measures to be implemented.
Animals were housed in the Institut Pasteur animal facilities (Paris) accredited by the French Ministry of Agriculture for performing experiments on live rodents. Work on animals was performed in compliance with French and European regulations on care and protection of laboratory animals (EC Directive 2010/63, French Law 2013–118, February 6th, 2013). All experiments were approved by the Ethics Committee #89 and registered under the reference APAFIS#6573-201606l412077987 v2.
From May to November each year, a batch of 50 ovitraps was usually set in Paris (department n°75) and replaced on a monthly basis. In 2023, from 96 traps containing eggs, 20 larvae were identified as Ae. albopictus and obtained adults were used to generate the colony. Neonate larvae were placed in pans containing 1 liter of dechlorinated water and a yeast tablet renewed every 2 days. Pupae were collected in bowls placed in cages where adults emerged. Adults were fed with a 10% sucrose solution and kept at 28 ± 1°C with a 16L:8D cycle and 80% relative humidity. Three times a week, adults were fed on an anesthetized 4-8-week-old OF1 mice to obtain eggs. We used the generations F2-F3 of Ae. albopictus 75 for experimental infections.
We infected mosquitoes with four DENV strains: (i) DENV-1 2010-1806P France (GenBank accession number: MG518567.1 (5), (ii) DENV-2 Bangkok 1974 (GenBank accession number: MK268692 (14), (iii) DENV-3 Martinique 2020 (kindly provided by the National Reference Centre of Arboviruses, Marseille, France), and (iv) DENV-4 CAY.09 (15). Viral stocks were produced on Aedes albopictus C6/36 cells and stored at -80°C until use.
Seven-to-ten-day-old mosquitoes were sorted on ice and females placed by 60 in plastic boxes, 24 hours before experimental infection. The infectious blood meal composed of 2/3 rabbit red blood cells, 1/3 viral suspension and ATP (a phagostimulant) was at a titer of 107 foci-forming units (FFU)/mL. Mosquitoes were allowed to feed for 15-20 min through a piece of pig intestine covering a capsule containing the infectious blood heated at 37°C using the Hemotek® system. Engorged females were sorted on ice and placed in cardboard boxes. Mosquitoes were maintained at 28 ± 1°C, relative humidity of 80%, 12L:12D cycle and supplied with 10% sugar solution. We analyzed mosquitoes at 14 and 21 days post-infection (dpi) to estimate three parameters describing the vector competence (16): (i) infection rate (IR) corresponding to the percentage of mosquitoes with an infected abdomen (a proxy of the midgut) among analyzed mosquitoes, (ii) stepwise dissemination rate (SDR) representing the percentage of mosquitoes with infected thorax and head among mosquitoes having an infected abdomen; it measures the ability of the virus to cross the midgut barrier and to disseminate into the mosquito’s general cavity, and (iii) stepwise transmission rate (STR) referring to the percentage of mosquitoes with virus in saliva among mosquitoes with infected head; it reflects the ability of the virus to invade the salivary glands, replicate and be excreted with the saliva produced by the mosquito when it bites. Values of SDR and STR assign the relative importance of, respectively, the midgut and salivary glands as barriers to the progression of viral infection in the mosquito; the higher the value, the less effective the barrier will be.
Mosquitoes were cold anesthetized on ice. Then legs and wings were removed and the proboscis was inserted into a pipette tip containing 5 μL of fetal bovine serum (FBS). After 30 min, the tip content was transferred in 45 μL of L15 medium. Abdomen was then separated from the thorax and head. These two samples were separately ground in 300 μL of Leibovitz L15 medium (Invitrogen, CA, USA) supplemented with 2% FBS, and centrifuged at 10,000×g for 5 min at +4°C.
Infection status of samples were determined by titration on Ae. albopictus C6/36 cells (17). After 5 days of an incubation period at 28˚C, cells were stained using the primary antibody, Ms X Dengue complex MAB 8705 (Millipore, MA, USA) and then, the secondary antibody, Alexa Fluor 488 goat anti-mouse IgG (Life Technologies, CA, USA). Foci were counted under a fluorescent microscope and titers expressed as FFU/mL.
IR, SDR and STR were compared using Fisher’s exact test and viral loads using Mann-Whitney test. Statistical analyses were conducted using the Stata software (StataCorp LP, Texas, USA). p-values < 0.05 were considered significant.
At 14 dpi, 8.3% (IR; 2/24) of mosquitoes had an infected midgut and among them, 50% (SDR; 1/2) were able to disseminate the virus in the mosquito general cavity but none of them (STR; 0/1) excreted infectious saliva (Figure 1A; Supplementary Table S1). This profile was also observed at 21 dpi (Figure 1E; Supplementary Table S1) suggesting that Ae. albopictus Paris was not able to transmit DENV-1 before 21 dpi, the critical step being midgut infection.
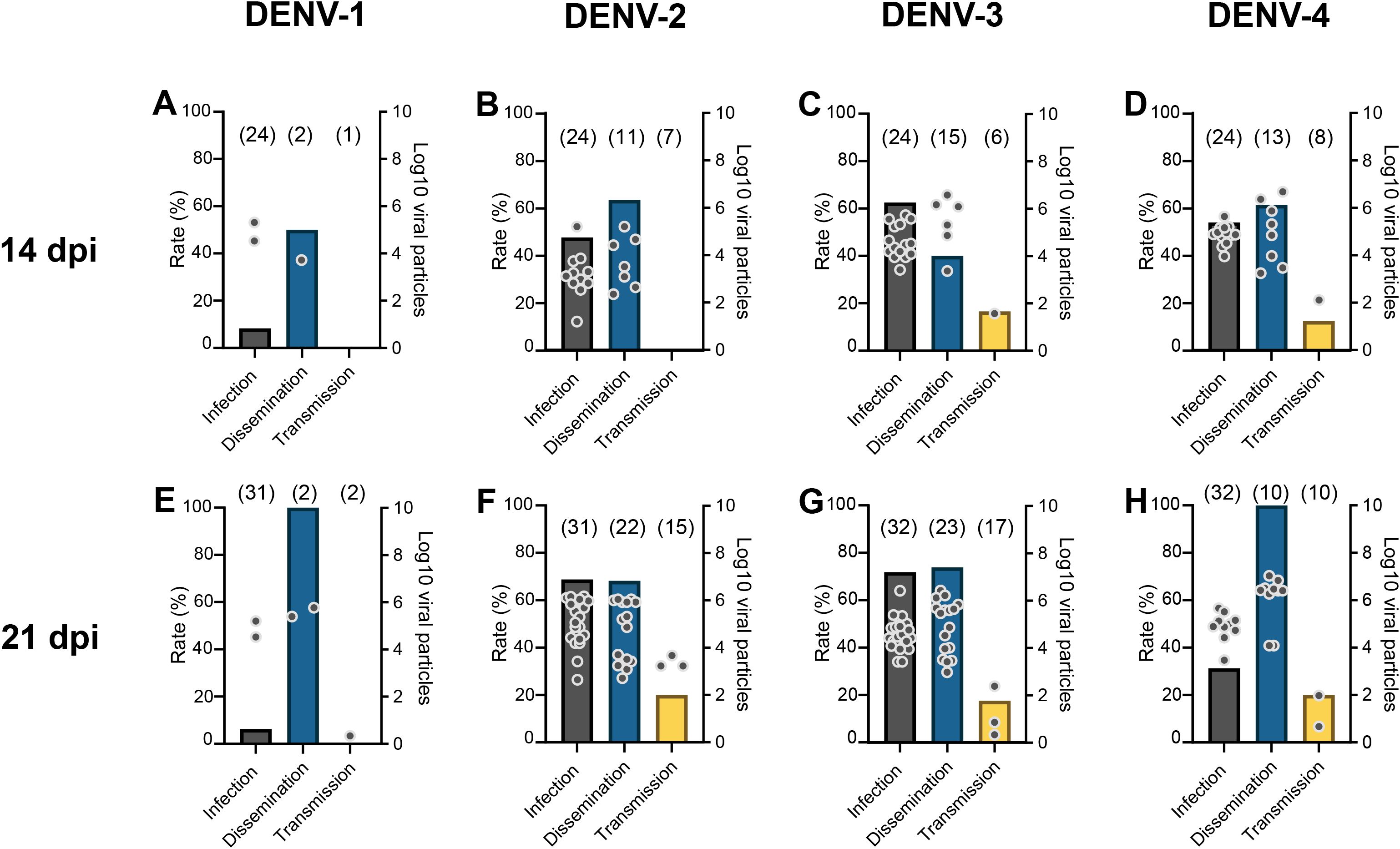
Figure 1. Infection, stepwise dissemination, stepwise transmission rates, and number of viral particles in abdomen, head&thorax, saliva of Aedes albopictus Paris examined at 14 (A-D) and 21 (E-H) days post-infection with a blood meal containing DENV at a titer of 107 FFU/mL. Mosquitoes were examined to measure: (i) infection rate (IR) corresponding to the percentage of mosquitoes with an infected abdomen (a proxi of the midgut) among analyzed mosquitoes, (ii) stepwise dissemination rate (SDR) representing the percentage of mosquitoes with infected head&thorax among mosquitoes having an infected abdomen and thorax, and (iii) stepwise transmission rate (STR) referring to the percentage of mosquitoes with virus in saliva among mosquitoes with infected head&thorax. Samples were processed for viral titration on mosquito cells. Bars correspond to rates (infection, dissemination, and transmission) and dots to numbers of viral particles. In brackets are the numbers of mosquitoes examined.
At 14 dpi, 45.8% (IR; 11/24) of mosquitoes were infected, and among them, 63.6% (SDR; 7/11) were able to disseminate the virus in the general cavity but none (STR; 0/7) could excrete the virus (Figure 1B; Supplementary Table S1). At 21 dpi, a similar profile was obtained with IR=68.7% (22/32) and SDR=68.2% (15/22), but 20% of mosquitoes were able to excrete infectious saliva (STR; 3/15) (Figure 1F; Supplementary Table S1). These results suggest that the salivary glands played a more significant role than the midgut as barrier to the DENV-2 progression in the mosquito but the virus overcomes this ultimate obstacle at 21 dpi.
For DENV-3, at 14 dpi, 62.5% (IR; 15/24) of mosquitoes were infected, among which, 40% (SDR; 6/15) were able to disseminate the virus, and among them, 16.7% (STR; 1/6) were able to transmit DENV-3 (Figure 1C; Supplementary Table S1). At 21 dpi, IR (71.9%; 23/32) and SDR (73.9%; 17/23) significantly increased while STR remained steady (17.6%; 3/17) suggesting that the salivary glands regulate DENV-3 transmission (Figure 1G; Supplementary Table S1).
For DENV-4, at 14 dpi, IR, SDR and STR were respectively, 54.2% (13/24), 61.5% (8/13) and 12.5% (1/8) indicating that the salivary glands are a strong barrier to DENV-4 transmission (Figure 1D; Supplementary Table S1). At 21 dpi, while only 31.2% of mosquitoes (IR; 10/32) were infected, 100% of mosquitoes (SDR; 10/10) were able to disseminate the virus and 20% (STR; 2/10) had infectious saliva (Figure 1H; Supplementary Table S1).
DENV-3 and DENV-4 were transmitted by Ae. albopictus Paris from 14 dpi.
We counted the number of viral particles in abdomen (proxy of midgut, infection), head (dissemination), and saliva (transmission) of all mosquitoes (including all four DENV and the two dpi). The mean number of viral particles in midgut was significantly higher in mosquitoes that could disseminate the virus (mean ± SD; 66987 ± 107741, N=66), as compared to mosquitoes unable to disseminate the virus (19959 ± 33566, N=32) (p=0.003) (Figure 2A; Supplementary Table S2). Similarly, the mean number of viral particles in midgut was significantly higher in mosquitoes able to transmit the virus (174960 ± 194034, N=10), as compared to mosquitoes unable to transmit (37616 ± 61484, N=88) (p=0.0093) (Figure 2B; Supplementary Table S2).
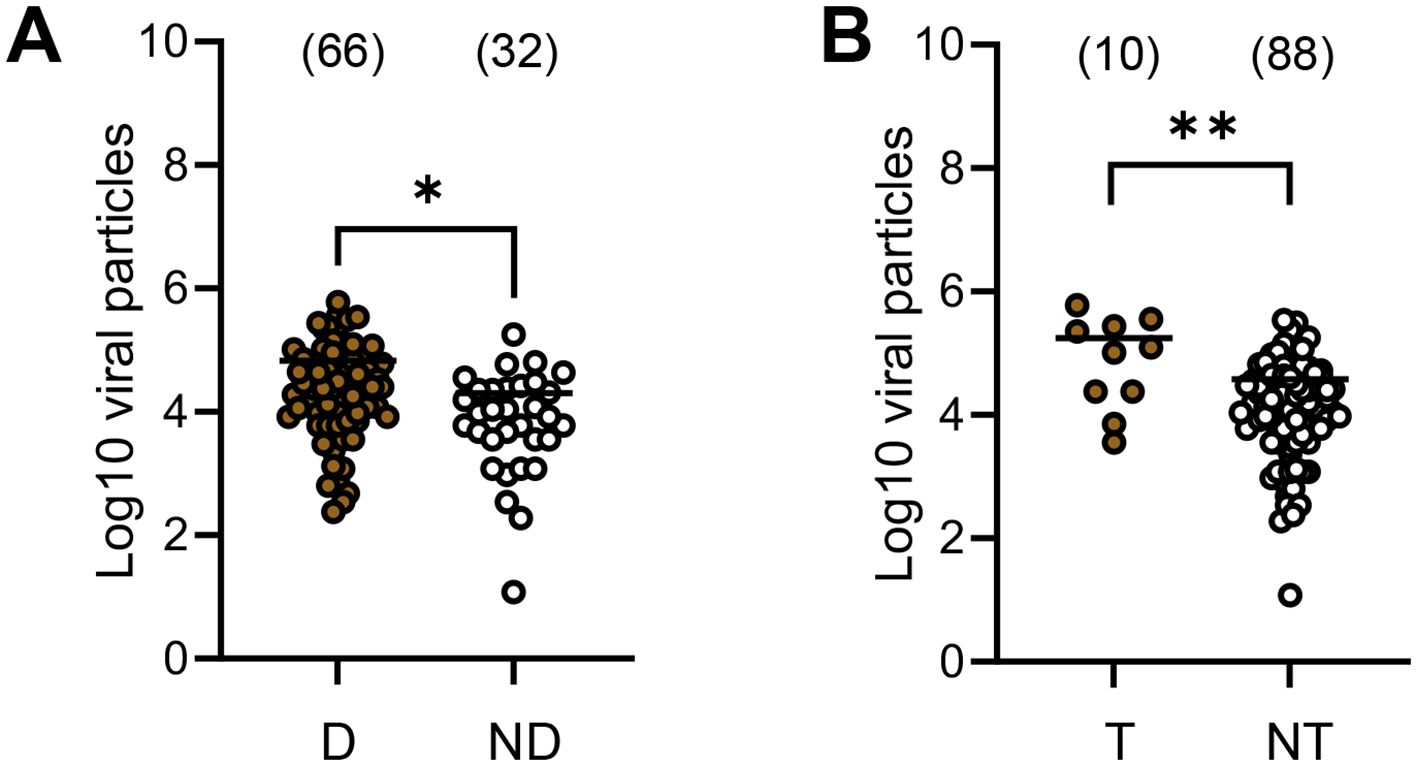
Figure 2. Number of virus particles (in Log10) detected in the midgut of Aedes albopictus Paris according to mosquito status: with/without dissemination (A) and with/without transmission (B). We examined all mosquitoes regardless of the DENV serotype and the day post-infection. The number of virus particles in abdomen (a proxi of the midgut), head&thorax (dissemination) and saliva (transmission) were estimated by titration on mosquito cells. D, mosquitoes able to disseminate the virus in the general cavity. ND, mosquitoes unable to disseminate the virus in the general cavity (undetected virus in the head&thorax). T, mosquitoes able to transmit the virus. NT, mosquitoes unable to transmit the virus (undetected virus in saliva). *0.01 ≤ p < 0.05, **0.001 ≤ p < 0.01 by Kruskall-Wallis non-parametric test (one-sided). In brackets are the numbers of mosquitoes tested.
On examination of the DENV serotype, the mean viral load of viral particles in midgut was not significantly different between mosquitoes able to disseminate versus mosquitoes unable to disseminate (p>0.05) except for DENV-3 where mosquitoes able to disseminate had a mean of 59,739 ± 124,114 versus 12,720 ± 15,924 for mosquitoes unable to disseminate (p=0.033) (Figure 3A; Supplementary Table S3). When considering transmission, the mean number of viral particles in midgut was not significantly different between mosquitoes able to transmit compared to mosquitoes unable to transmit except for DENV-2 with 288,000 ± 66,813 versus 55,353 ± 94,898 (p=0.012) (Figure 3B; Supplementary Table S3).
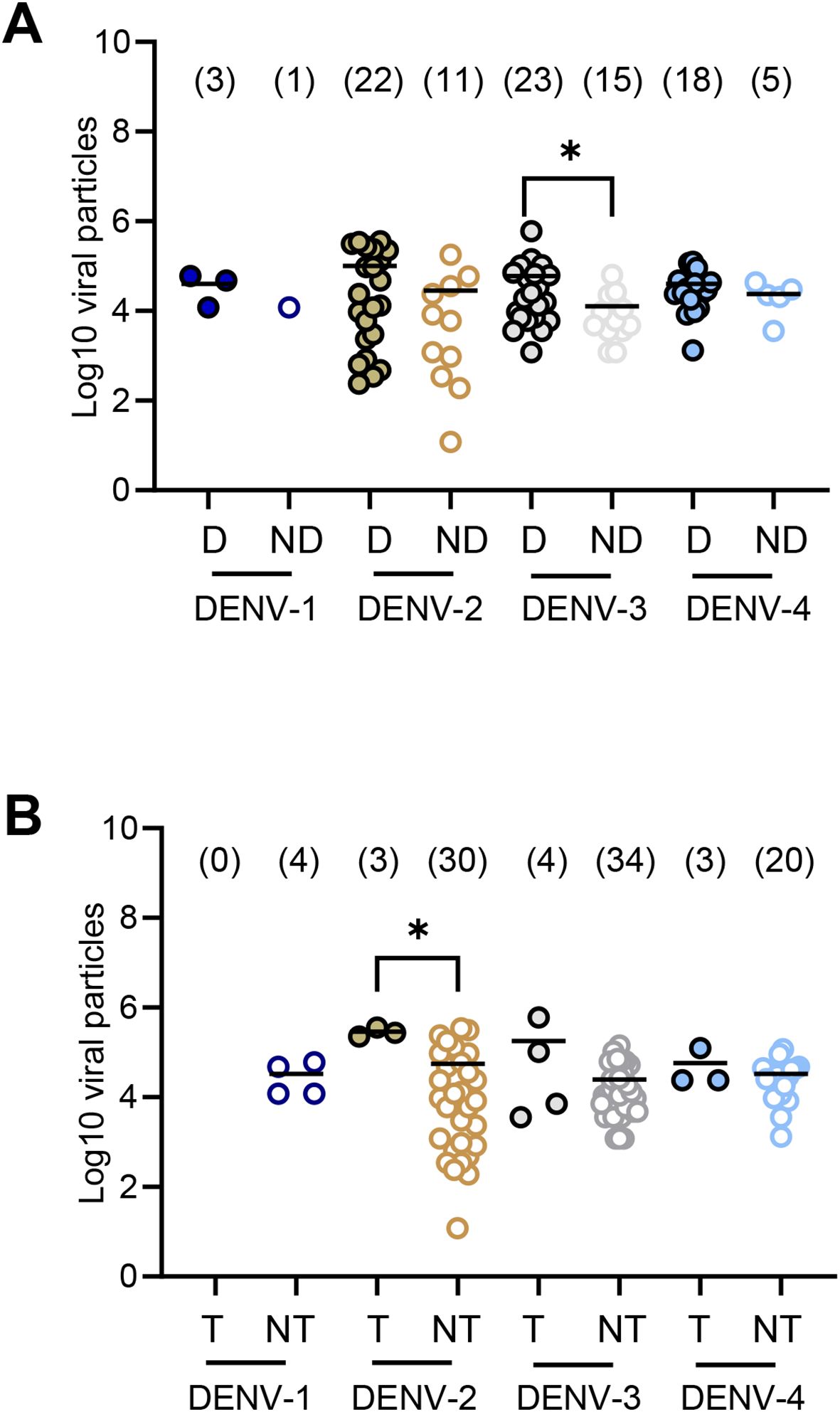
Figure 3. Number of virus particles (in Log10) detected in the midgut of Aedes albopictus Paris according to the DENV (1–4) and the mosquito status: with/without dissemination (A) and with/without transmission (B). We examined mosquitoes for each DENV serotype and each day post-infection. The number of virus particles in abdomen (a proxi of the midgut), head&thorax (dissemination) and saliva (transmission) were estimated by titration on mosquito cells. D, mosquitoes able to disseminate the virus in the general cavity. ND, mosquitoes unable to disseminate the virus in the general cavity (undetected virus in the head&thorax). T, mosquitoes able to transmit the virus. NT, mosquitoes unable to transmit the virus (undetected virus in saliva). *0.01 ≤ p < 0.05, by Kruskall-Wallis non-parametric test (one-sided). In brackets are the numbers of mosquitoes tested.
Therefore, the viral load required to allow viral dissemination and transmission in mosquitoes depended on the DENV serotype. A high viral load in the midgut did not lead necessarily to successful dissemination and transmission.
Aedes albopictus is usually a less competent vector for DENV (18) but participates in dengue outbreaks in areas where Ae. aegypti is absent (19). In September 2023, the arbovirus surveillance system detected the first autochthonous cases of dengue in the greater Paris. We showed that Ae. albopictus Paris was competent for at least three DENV serotypes (-2, -3, and -4) with transmission from day 21 post-infection.
While most human infections are asymptomatic or develop minor symptoms, ~25% are symptomatic and detected by the national surveillance system implemented each year from May to November (20). When an autochthonous case is identified, an investigation of infected people is implemented in the area to determine the extent of local transmission. The « Agence Régionale de Démoustication » (ARD), responsible for vector control, set up entomological and vector control measures by eliminating larval breeding sites and using insecticides against immature stages with Bacillus thuringiensis and adults with a pyrethroid, Deltamethrin. People with inapparent DENV infections are, against all expectations, more infectious to mosquitoes than people with symptomatic infection (21). These DENV viremic people without clinical symptoms are more likely exposed to mosquito bites as infection does not disrupt their daily activities, contributing to virus dissemination.
For Ae. aegypti, it has been demonstrated that the level of viremia required to infect 50% of mosquitoes was lower for DENV-1 and DENV-2 than for DENV-3 and DENV-4, suggesting a lower infectiousness of DENV-3 and DENV-4 (22). However, we demonstrated that Ae. albopictus Paris transmitted DENV-3 and DENV-4 earlier at day 14 post-infection than DENV-2 when offering a blood meal at a titer of 107 FFU/mL. Coincidentally, virus surveillance in mainland France indicates that the largest source of autochthonous transmission (i.e. 34 infection cases) was with DENV-3 in Southeast France (Saint Jeannet, Gattières, Alpes-Maritimes) (23). Nevertheless, multiple other factors may determine the prevalence of DENV serotypes in epidemic areas including the mosquito life span (24); to transmit, the mosquito’s life span should be longer than the time necessary for a mosquito to become infectious. Beyond the DENV serotype, a high viral load in mosquito midgut can lead to an efficient viral transmission. We showed that mosquitoes able to transmit DENV-2 presented 3 times more viral particles in midgut than mosquitoes unable to transmit, 288,000 and 55,353, respectively. This was not the case for the other three DENV serotypes. To note, in natural conditions, mosquitoes can acquire a second non-infectious blood meal and this may enhance viral dissemination and transmission via blood meal-induced micro-perforations in the basal lamina surrounding the midgut epithelium (25).
Growing international travels with dengue endemic regions and the establishment of Ae. albopictus since 2015 in the greater Paris will intensify the risk for autochthonous dengue transmission in the region. This unusual episode of local dengue transmission emphasizes the threat of dengue for northern European countries where Ae. albopictus is well establish as in Belgium or Germany. Eliminating sources of larval breeding in artificial containers or natural stagnant water must be conducted on a routine basis. The use of insecticides should be implemented only when autochthonous cases are detected. As long as the arsenal of vector control methods is limited to the use of a few insecticides, these should be used sparingly to delay the development of resistance as long as possible (26).
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The animal study was approved by Ethics Committee #89 registered under the reference APAFIS#6573-201606l412077987 v2. The study was conducted in accordance with the local legislation and institutional requirements.
CB: Investigation, Writing – review & editing. MV: Methodology, Writing – review & editing. LP: Methodology, Writing – review & editing. KM: Methodology, Writing – review & editing. AK: Methodology, Writing – review & editing. XD: Conceptualization, Writing – review & editing. AF: Conceptualization, Data curation, Funding acquisition, Supervision, Validation, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the project funded by the Credit Mutuel.
We would like to thank Patricia Smiley for editing.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fitd.2025.1565116/full#supplementary-material
2. Curtin TJ. Status of Aedes aEgypti in the Eastern Mediterranean. J Med Entomol. (1967) 4:48–50. doi: 10.1093/jmedent/4.1.48
3. Adhami Jm N. The presence of the mosquito Aedes albopictus in Albania Rev Mjekësore. Revistë Mjekësore. (1987) 1:13–6.
4. Authority. ECfDPaCaEFS. Mosquito maps(2024). Available online at: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/mosquito-maps (Accessed June 11, 2024).
5. La Ruche G, Souares Y, Armengaud A, Peloux-Petiot F, Delaunay P, Despres P, et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Euro Surveill. (2010) 15:19676. doi: 10.2807/ese.15.39.19676-en
6. Gjenero-Margan I, Aleraj B, Krajcar D, Lesnikar V, Klobucar A, Pem-Novosel I, et al. Autochthonous dengue fever in Croatia, August-September 2010. Euro Surveill. (2011) 16:pii: 19805. doi: 10.2807/ese.16.09.19805-en
7. Scholte EJ, Schaffner F. Waiting for the tiger: establishment and spread of the Asian tiger mosquito in Europe. In: Takken W, Knols BGJ, editors. Emerging pests and vector-borne diseases in Europe. Wageningen Academic Publishers, Wageningen, The Netherlands (2007). p. 241–60.
8. Ministère du Travail dlSedS. Cartes de présence du moustique tigre (Aedes albopictus) en France métropolitaine. Point au 21 avril 2023(2023). Available online at: https://sante.gouv.fr/sante-et-environnement/risques-microbiologiques-physiques-et-chimiques/especes-nuisibles-et-parasites/article/cartes-de-presence-du-moustique-tigre-aedes-albopictus-en-France-metropolitaine (Accessed August 02, 2024).
9. Fournier L, Calba C, Cochet A, Fournet N, Brottet É, Grard G, et al. Bilan de la surveillance de la dengue, du chikungunya et du Zika en France hexagonale en 2023. Bull Épidémiologique Hebdomadaire. (2024) 13:260–6.
10. France SP. Chikungunya, dengue et zika - Données de la surveillance renforcée en France hexagonale 2024(2024). Available online at: https://www.santepubliqueFrance.fr/maladies-et-traumatismes/maladies-a-transmission-vectorielle/chikungunya/articles/donnees-en-France-metropolitaine/chikungunya-dengue-et-zika-donnees-de-la-surveillance-renforcee-en-France-hexagonale-2024 (Accessed December 24, 2024).
11. Zatta M, Brichler S, Vindrios W, Melica G, Gallien S. Autochthonous dengue outbreak, Paris region, France, September-October 2023. Emerg Infect Dis. (2023) 29:2538–40. doi: 10.3201/eid2912.231472
12. Fournet N, Voiry N, Rozenberg J, Bassi C, Cassonnet C, Karch A, et al. A cluster of autochthonous dengue transmission in the Paris region - detection, epidemiology and control measures, France, October 2023. Euro Surveill. (2023) 28:1–5. doi: 10.2807/1560-7917.ES.2023.28.49.2300641
13. France SP. Recrudescence de cas importés de dengue en France hexagonale: appel à la vigilance à l’approche de la saison d’activité du moustique tigre(2023). Available online at: https://www.santepubliqueFrance.fr/les-actualites/2024/recrudescence-de-cas-importes-de-dengue-en-France-hexagonale-appel-a-la-vigilance-a-l-approche-de-la-saison-d-activite-du-moustique-tigre (Accessed April 23, 2024).
14. Vazeille-Falcoz M, Mousson L, Rodhain F, Chungue E, Failloux A-B. Variation in oral susceptibility to dengue type 2 virus of populations of Aedes aEgypti from the islands of Tahiti and Moorea, French Polynesia. Am J Trop Med Hyg. (1999) 60:292–9. doi: 10.4269/ajtmh.1999.60.292
15. Vazeille M, Gaborit P, Mousson L, Girod R, Failloux AB. Competitive advantage of a dengue 4 virus when co-infecting the mosquito Aedes aEgypti with a dengue 1 virus. BMC Infect Dis. (2016) 16:318. doi: 10.1186/s12879-016-1666-0
16. Bohers C, Vazeille M, Bernaoui L, Pascalin L, Meignan K, Mousson L, et al. Aedes albopictus is a competent vector of five arboviruses affecting human health, greater Paris, France, 2023. Euro Surveill. (2024) 29:1–7. doi: 10.2807/1560-7917.ES.2024.29.20.2400271
17. Bohers C, Mousson L, Madec Y, Vazeille M, Rhim A, M’Ghirbi Y, et al. The recently introduced Aedes albopictus in Tunisia has the potential to transmit chikungunya, dengue and Zika viruses. PloS Negl Trop Dis. (2020) 14:e0008475. doi: 10.1371/journal.pntd.0008475
18. Vazeille M, Rosen L, Mousson L, Failloux AB. Low oral receptivity for dengue type 2 viruses of Aedes albopictus from Southeast Asia compared with that of Aedes aEgypti. Am J Trop Med Hyg. (2003) 68:203–8. doi: 10.4269/ajtmh.2003.68.203
19. Lambrechts L, Scott TW, Gubler DJ. Consequences of the expanding global distribution of Aedes albopictus for dengue virus transmission. PloS Negl Trop Dis. (2010) 4:e646. doi: 10.1371/journal.pntd.0000646
20. Ministère du Travail dlEedlS. Guide relatif aux modalités de mise en œuvre du plan anti-dissémination du chikungunya et de la dengue en métropole(2015). Available online at: https://sante.gouv.fr/IMG/pdf/instruction_et_guide_chik_dengue_16_avril_2015.pdf (Accessed April 16, 2015).
21. Duong V, Lambrechts L, Paul RE, Ly S, Lay RS, Long KC, et al. Asymptomatic humans transmit dengue virus to mosquitoes. Proc Natl Acad Sci U S A. (2015) 112:14688–93. doi: 10.1073/pnas.1508114112
22. Nguyet MN, Duong TH, Trung VT, Nguyen TH, Tran CN, Long VT, et al. Host and viral features of human dengue cases shape the population of infected and infectious Aedes aEgypti mosquitoes. Proc Natl Acad Sci U S A. (2013) 110:9072–7. doi: 10.1073/pnas.1303395110
23. Cochet A, Calba C, Jourdain F, Grard G, Durand GA, Guinard A, et al. Autochthonous dengue in mainland France, 2022: geographical extension and incidence increase. Euro Surveill. (2022) 27:1–5. doi: 10.2807/1560-7917.ES.2022.27.44.2200818
24. Brady OJ, Johansson MA, Guerra CA, Bhatt S, Golding N, Pigott DM, et al. Modelling adult Aedes aEgypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasit Vectors. (2013) 6:351. doi: 10.1186/1756-3305-6-351
25. Armstrong PM, Ehrlich HY, Magalhaes T, Miller MR, Conway PJ, Bransfield A, et al. Successive blood meals enhance virus dissemination within mosquitoes and increase transmission potential. Nat Microbiol. (2020) 5:239–47. doi: 10.1038/s41564-019-0619-y
Keywords: Aedes albopictus, vector competence, dengue virus, France, risk assessment
Citation: Bohers C, Vazeille M, Pascalin L, Meignan K, Karch A, De Lamballerie X and Failloux A-B (2025) Aedes albopictus of Paris (France) is competent to transmit at least three of the four dengue virus serotypes. Front. Trop. Dis. 6:1565116. doi: 10.3389/fitd.2025.1565116
Received: 22 January 2025; Accepted: 06 March 2025;
Published: 14 April 2025.
Edited by:
Yenddy Carrero, Technical University of Ambato, EcuadorReviewed by:
Frank William Avila, University of Antioquia, ColombiaCopyright © 2025 Bohers, Vazeille, Pascalin, Meignan, Karch, De Lamballerie and Failloux. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anna-Bella Failloux, YW5uYS1iZWxsYS5mYWlsbG91eEBwYXN0ZXVyLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.