Abstract
American cutaneous leishmaniasis (ACL) presents a wide spectrum of clinical and immunopathological manifestations. In Brazil, Leishmania (L.) amazonensis[La] and Leishmania(V.)braziliensis[Lb] show the highest pathogenic potential for humans causing different clinical forms: localized cutaneous leishmaniasis (LCL : Lb/La), anergic diffuse cutaneous leishmaniasis (ADCL : La) and mucocutaneous leishmaniasis (MCL : Lb). ADCL and MCL are the most severe forms and infection leads to a cellular immune response at the hyposensitivity and hypersensitivity poles. Th17-cells are involved in the ACL pathogenesis, are derived from naïve TCD4+ cells regulated by RORγt, differentiate in presence of IL-6, TGF-β, IL- 1β, IL-23 and express IL-17. Aim of this study was to characterize the cellular immune response mediated by Th17-profile cells through in situ determination of the expression of RORγt, IL-17, IL-6, TGF-β, IL-1β, and IL-23 in the ACL clinical-immunopathological spectrum caused by L.(L.)amazonensis and L.(V.)braziliensis. Biopsies of skin and mucosal lesions from forty patients including ADCL(n=8), LCL[La](n=17), LCL[Lb](n=9) and MCL(n=6), were examined by immunohistochemistry. The immunostained cells density (cells/mm2) was determined in image analysis system using AxionVision 4.8 software (Zeiss). As the disease evolution time (DET) was different among ACL patients, the effect of DET on the expression of immunological markers was evaluated in different clinical forms and histopathological changes, using ANCOVA. Our results showed significantly increased expression of RORγt, IL-17, IL-6, IL-1β and IL-23 in patients with ACL polar forms (ADCL and MCL); higher TGF-β expression was found in ADCL. DET influenced the expression of RORγt and IL-6 in: clinical forms of ACL and in categories of parasitism. DET also affected the production of RORγt, IL-17, IL-6, TGF-β and IL-1β in types of inflammatory infiltrate, evidencing that DET had effect on the expression of Th17 profile cytokines in ACL. Together, the expression of immunological mediators of Th17 profile in the ACL spectrum, as well as the DET effect, demonstrate the participation of this cell lineage in the immunopathogenesis of ACL, mainly in the polar and more severe forms of ACL spectrum. The dubious role played by Th17-cells may favors immune response suppression and parasitic persistence in ADCL, while in MCL it contributes to an exacerbated immune response and parasite scarcity.
Introduction
American cutaneous leishmaniasis (ACL) is a noncontagious protozoan disease that has a broad clinical and immunopathological spectrum and is caused by parasites of the subfamily Leishmaniinae (1). It is present in 18 America’s countries, with an average of 55,317 new cases/year (2). Seven species that cause ACL are recognized in Brazil: six of the subgenus Viannia and one of the subgenus Leishmania. The two species of greatest medical interest are Leishmania (V.) braziliensis and Leishmania (L) amazonensis (3–5). ACL may have a broad spectrum of clinical and immunological manifestations, depending on the immunological profile of the host and the species of the parasite. The mildest ACL is localized cutaneous leishmaniasis (LCL), caused by L. (L.) amazonensis and L. (V.) braziliensis, characterized by a well-balanced cellular immune response profile, with moderate hypersensitivity (positive Montenegro Intradermal Reaction/MIR+). It may also present mucosal cutaneous leishmaniasis (MCL) in the hyperreactive cellular immune pole, with strong hypersensitivity (MIR++++) and many lymphocytes (mainly CD4+), dense and diffuse infiltrate and mucosal tissue necrosis (4, 5), as well as a high expression of proinflammatory mediators of the Th1 profile, such as tumour necrosis factor (TNF)-α and interferon (IFN)-γ (4, 5). On the other hand, it can present as the hyporeactive form called anergic diffuse cutaneous leishmaniasis (ADCL), with weak or absent hypersensitivity, characterized by an infiltrate of intensely parasitized vacuolated macrophages, and discrete lymphoplasmacytic infiltrate, whose clinical presentation is nodular skin lesions rich in amastigote forms of the parasite disseminated over the entire body, characterized by the predominance of the Th2-type cellular immune response, with high production of mediators such as interleukin (IL)-10 and transforming growth factor (TGF)-β (4, 6, 7).
Depending on the cytokines present in the inflammatory microenvironment, naïve TCD4+ cells can be activated via major histocompatibility complex type II and differentiate into subpopulations of effector cells. In addition to Th1 and Th2 lymphocytes, other cell groups have been described as being involved in the pathogenesis of ACL, such as Th17 cells and regulatory T (Treg) cells (8–10).
Th17 cells are derived from naïve TCD4+ cells that express the transcription factor retinoic acid receptor–related orphan receptor (ROR)γt (a nuclear gamma receptor associated with retinoic acid). Their classic differentiation is stimulated by IL-6, TGF-β and IL-1β and they express IL-17 and IL-22 (11). RORγt is an isoform of RORγbelonging to the retinoic acid receptor–related orphan nuclear receptor family, which includes RORα and RORγ) (12–14). This factor is expressed in immune response cells, such as thymocytes and Th17 cells. The expression of these receptors is stimulated by the activation of the (signal transducer and activator of transcription (STAT)3 protein in the presence of IL-6 and TGF-β, leading to the differentiation of classic Th17 cells, which can produce IL-10 and have regulatory characteristics (15). IL-6 and TGF-β induce naïve CD4+ T cells to produce IL-21, which helps maintain differentiation in an autocrine manner, stimulating the production of IL-1 and IL-23 receptors (IL-1R1 and IL-23R, respectively). In the absence of TGF-β, the alternative Th17 cells which can produce proinflammatory molecules such as IFN-γ, may expand (15–17).
Finally, IL-23 and IL-1β are associated with the maintenance and expansion of Th17 cells. Once expressed, RORγt and RORα induce the differentiation of Th17 cells and the expression of cytokines such as IL-17 and IL-22 by these cells. Thus, the differentiation and maintenance of cells of this profile are intrinsically linked to the expression of ROR transcription factors (12, 13).
Th17 cells act in the production of proinflammatory cytokines and in the cellular recruitment of leukocytes and neutrophils by the expression of IL-17, which induces the production of other proinflammatory cytokines, such as IL-6 and TNF-α (11, 18).
In the presence of TGF-β, IL-6, IL-1β and IL-23 occur the “classic” Th17 cells differentiation, as mentioned above, that can be associated with a regulatory role and suppression of the host response. Nevertheless, Th17 cells can develop in the absence of TGF-β, but in the presence of IL-1β and IL-23, the differentiation of “alternative” Th17 cells, wich are associated with the production of IFN-γ and elimination of parasites and are more pathogenic than “classic” cells (11, 15, 17).
The expression cytokine profiles including IL-17 may be strongly influenced by the infecting species, along with other factors. Flow cytometry analysis of serum samples from 33 patients from the state of Amazonas, Brazil, infected with different species of the subgenera Viannia and Leishmania showed different levels of IL-2, TNF-α, IFN-γ, IL-4, IL-6, IL-10 and IL-17, and IL-17 had higher levels in the sera of patients with ACL caused by L. (V.) guyanensis than in samples from patients infected with L. (L.) amazonensis (19). Bacellar et al. (20) stimulated peripheral blood mononuclear cell of patients diagnosed with LCL and MCL caused by L. (V) braziliensis. When analyzing the expression of cytokines such as IL-17, IFN-γ, and TNF-α, they found high concentrations of IL-17 in patients with different clinical forms of ACL compared to the concentration in the control group and a positive correlation between IL-17 and TNF-α in the MCL clinical form, suggesting that IL-17 may contribute to tissue damage and disease severity. Martins et al. (2020) stimulated in vitro macrophages from C57BL/6 mice with IL-17 and then infected them with L. major promastigotes. The results showed that IL-17 favors parasite survival in host cells and it was associated with the development of lesions (21). By stimulating macrophages with IL-17 and infecting them with L. (L.) amazonensis, Sousa et al. (2014) demonstrated that this cytokine led to an increase in arginase activity and parasite load (22). These data corroborate studies that showed that IL-17 may induce polarization of type M2 macrophages, which have immunoregulatory and tissue repair functions, as described by Shen et al. (2018), who demonstrated that IL-17 induces the differentiation of these cells by the activation of nuclear factor kappa B (NF-κB) (23).
Understanding the pathogenesis of ACL, especially with regard to the development of these different clinical forms, represents a major challenge to better understanding this protozoan disease due to the complex interaction between the various Leishmania species that cause the disease and immune response.
Given the above and the need for a better understanding of the participation of Th17 profile cells in the immunopathogenesis of the ACL, the present study aimed to characterize the cellular immune response mediated by Th17-profile cells by determining in situ expression of immunological mediators involved in the differentiation and maintenance of these cells, thus demonstrating the participation of this cell group in the clinical and immunopathological spectrum of ACL.
Our findings showed the participation of Th17 cells response especially in the polar and more severe forms of the ACL caused by L. (L.) amazonensis and L. (V.) braziliensis. The dubious role played by Th17-cells possibly favors an immune response suppression and parasitic persistence in ADCL; while in MCL it contributes to an exacerbated immune response and parasite scarcity.
Materials and methods
Patients
Biopsies of active lesions were taken from forty patients with different clinical forms of ACL; including eight with ADCL [L. (L.) amazonensis]; 17 with LCL [L. (L.) amazonensis]; nine with LCL [L. (V.) braziliensis]; and six with MCL [L. (V.) braziliensis], seen at the Evandro Chagas Institute Outpatient Clinic (Leishmaniasis Laboratory “Prof. Dr. Ralph Lainson”) in Pará state, Brazil.
Patients who agreed to participate in the study filled out a clinical-epidemiological form, with data regarding age, sex, race/ethnicity, probable site of infection and time of disease evolution. Patients were diagnosed for ACL through the Montenegro intradermal reaction, visualization of amastigotes in lesion smears and polymerase chain reaction for detection and identification of parasite species. To molecular identification of Leishmania species was permormed by PCR-RFLP having as target the coding sequences for the heat shock protein of 70kD (hsp70) as described by Graça et al., 2012 (24).
All patients with a confirmed diagnosis of ACL received specific treatment. Skin samples were also collected from six uninfected individuals as controls, obtained in Laboratory of Microsurgery and Plastic Surgery (LIM-04), Division of Plastic Surgery, Faculty of Medicine of the University of São Paulo (FMUSP).
Histopathological analysis
A slide with histological sections of skin fragments from each patient affected with ACL was processed and stained with haematoxylin–eosin. The slides were observed under an Axioskop 2 Plus Zeiss optical microscope, and a descriptive histopathological analysis of histological changes in the epidermis and dermis was performed.
Immunohistochemistry
In order to analyze the cellular immune response in lesions of patients in the four clinical forms of ACL, an immunohistochemical reaction by immunoperoxidase was performed in the Laboratory of Pathology of Infectious Diseases (LIM - 50 - HCFMUSP). Antibody reactions were performed against rettinoid-acid-receptor-related orphan nuclear receptor gamma t (RORγt), interleukin-17 (IL-17), interleukin-6 (IL-6), transforming growth factor-β (TGF-β), interleukin-1β (IL-1β), and interleukin-23 (IL-23). The immunohistochemical reaction was performed as follows: The slides with the sections were placed in a xylene bath at room temperature for 15 minutes for deparaffinization. Hydration was performed in alcohol baths with decreasing concentration. Endogenous peroxidase was blocked using a hydrogen peroxide solution (3%), and antigenic recovery was performed using 10 mM pH 6 citrate buffer in a water bath for IL-6, IL-23, and IL- 1β; 10 mM pH 6 citrate in a pressure cookerfor RORγt; 1 mM pH 8 EDTA buffer in a water bath for IL-17; and Tris-EDTAfor TGF-β. After recovery, nonspecific sites were blocked using 6% Molico® skim milk with 10% foetal bovine serum. After these steps, the primary antibodies previously diluted in bovine serum albumin were added to the tissue samples: anti-RORγt (polyclonal, ab78007) diluited 1:250, anti-IL-17 (polyclonal, SC-7927) diluited 1:200, anti-IL-6 (monoclonal SC-130326) diluited 1:200, anti-TGF-β (polyclonal SC-146) diluted 1:100, anti-IL-1β (polyclonal, ab2105) diluited 1:800, and anti-IL-23 (monoclonal C-3, SC-271279) diluited 1:500. The sections were incubated in a humid chamber overnight at 4°C.
The following day, the amplification and development system NOVOLINK ™ polymer detection systems kit (RE7280-K, Leica Biosystems, Newcastle, UK) was used for all markers, and the slides were incubated with the secondary antibody and polymer. To reveal the reaction, the chromogenic substrate diaminobenzidine with hydrogen peroxide (K3468, Dako Cytomation) was added for 5 minutes. Next, the samples were counterstained with Harris haematoxylin, or eosin in the case of RORγt. The slides were dehydrated in serial baths with increasing concentrations of alcohol and xylene and mounted with coverslips and synthetic resin.
Quantitative morphometric analysis of immunolabelled cells
The evaluation of the immune response was based on the quantitative morphometric analysis of immunostained cells in the immunohistochemical reaction. The material processed in the immunohistochemistry reaction was analysed in an image analysis system composed of an Axioskop 2 Plus Zeiss optical microscope at 400× magnification coupled to a microcomputer containing AxioVision 4.8 software. Eight to ten fields of each histological section were photographed. The immunolabelled cells were quantified using ImageJ software. To determine the density of immunostained cells, the mean count of 10 fields divided by the area of the photo (0.03536412 mm 2) was used.
Data analysis
The results of the immunohistochemical evaluations were stored in a Microsoft Access database and were analysed using Prism 5 software (GraphPad Software, San Diego, CA, USA). The results were subjected to statistical analysis and are presented as measures of central tendency (mean) and dispersion (standard deviation, maximum and minimum values) and was transformed into z-score and subjected to the Kolmogorov-Sminorv normality test. Parametric tests were used to calculate differences between quantitative results when the variables had a normal distribution. For those that did not show normal distribution, z-scores were calculated. The significance level adopted for the tests was p ≤ 0.05. To effect of disease evolution time (DET – considered at the time of the first signs/symptoms referred by the patients in the first medical care) on the expression and production of immunological markers in the different clinical forms of ACL, analysis of covariance (ANCOVA) was run in SPSS software, adopting as a dependent variable the production of the immunological markers of the Th17 profile in the different clinical forms and, as a covariable, the time of disease progression, considering the densities of immunostained cells per photographed field previously transformed into z scores and subjected to the Kolmogorov−Smirnov normality tests. The mean densities of immunolabelled cells transformed into z scores are presented in the error bar graph, which shows the 95% confidence intervals (CI).
From the covariation analysis, the effect size of disease progression time was given in partial eta squared (η2), whose size ranges from 0 to 1, in which effects greater than or equal to 0.3 are considered of reasoned magnitude. Due to the rarity of cases of ADCL [L.(L.) amazonensis] and MCL [L. (V.) braziliensis], all η values with p = 0.000 were considered impacting.
Finally, the post hoc Bonferroni test was run after each ANCOVA to determine which clinical forms showed statistically significant differences in the densities of cells that expressed the immunological markers of the Th17 profile.
Results
Histopathology
The histopathological analysis of the lesion of ADCL patients due to [La] showed an intense mononuclear inflammatory infiltrate in the dermis, consisting basically of vacuolated macrophages, with many forms suggestive of Leishmania sp. amastigotes, rare lymphocytes, and plasma cells (Figure 1A). In patients with LCL [La], a discrete to moderate mononuclear inflammatory infiltrate was observed in the dermis, predominantly macrophages, with vacuolated macrophages and moderate parasitism, and in rare cases an outline of granuloma was observed (Figure 1B). On the other hand, in the histopathological analysis of the lesions of patients with clinical forms caused by L. (V.) braziliensis, a localized cutaneous form of lymphoplasmacytic inflammatory infiltrate rich in plasmocytes and containing lymphocytes and macrophages in variable numbers was observed. Low parasitism was detected (Figure 1C). A granulomatous outline or well-formed epithelioid granulomas were also observed, often with the presence of giant cells. Epidermal thickening and irregular acanthosis were detected in the epidermis. In patients with MCL, histopathology was characterized by an intense diffuse lymphoplasmacytic infiltrate containing rare macrophages and few parasites. A granulomatous outline or well-formed epithelioid granuloma was identified in most cases (Figure 1D).
Figure 1

Histological section of the lesions of patients with diferente clinical forms of Cutaneous Leishmaniasis caused by L. (L.) amazonensis and L. (V.) braziliensis, showing the histopathological changes of the dermal lesion. (A) ADCL [La]: Anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis, showing an Intense mononuclear inflammatory infiltrate predominantly of vacuolated macrophages and with parasites inside. (B) LCL [La]: Localized Cutaneous Leishmaniasis caused by L. (L.) amazonensis, showing a moderate mononuclear infiltrate, formed predominantly by parasitized macrophages and lymphocytes. (C) LCL [Lb]: Localized Cutaneous Leishmaniasis caused by L. (V.) braziliensis, showing intense inflammatory infiltrate with plasma cells, lymphocytes and macrophages, and presence of granuloma containing giant cell. parasitic scarcity. (D) MCL [Lb]: Mucocutaneous Leishmaniasis caused by L. (V.) braziliensis, demonstrate intense lymphoplasmacytic inflammatory infiltrate and parasitic scarcity. P-parasite; cg- giant cell (stained by Hematoxilin-Eosin, 200x).
Immunopathological analysis of lesions
All analyzed cases of the ACL spectrum showed immunostained cells for the six antibodies. The comparative analysis of the densities of immunostained cells (cells/mm2) for the different markers showed significantly higher proportions of RORγt+, IL-17+, IL- 6+, IL-1β+, and IL-23+ cells in the lesions of patients with the polar forms, as well as a higher frequency of TGF-β+ cells in ADCL (p ≤ 0.05). Analyses were also performed on histological sections of healthy skin (control) for each immunological marker, all of which had significantly lower mean density values than ACL tissues (Table 1, Figure 2).
Table 1
| Expression of immunological markers of the Th17 profile in the different clinical forms of ATL | |||||
|---|---|---|---|---|---|
| Immunological marker | ADCL [La] (n=8) | LCL [La] (n=16) | LCL [Lb] (n=10) |
MCL [Lb] (n=6) | Control (n=6) |
| RORγt | 908.02±36.85 | 610.55±29.74 | 628.13±26.69 | 802.76±29.34 | 1.13±1.06 |
| IL-17 | 611.09±24.40 | 431.86±17.63 | 494.38±8.61 | 700.98±32.34 | 12.59±3.23 |
| IL-6 | 418.51±35.22 | 260.66±16.17 | 300.22±15.18 | 466.54±32.16 | 5.42±2.24 |
| TGF-β | 806.22±22.59 | 358.23±19.25 | 317.97±18.65 | 234.45±20.10 | 2.26±1.53 |
| IL-1β | 1135.35±60.93 | 565.11±19.91 | 528.68±21.21 | 1290.51±87.68 | 2.26±1.53 |
| IL-23 | 846.22±12.41 | 404.28±21.53 | 361.50±28.21 | 599.31±50.38 | 2.26±1.53 |
Mean densities of RORγt+, IL-17+, IL-6+, TGF-β+, IL-1β+, and IL-23+ cells in the different clinical forms of the American cutaneous leishmaniasis spectrum and in skin samples without lesions.
ADCL, anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La], localized cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [Lb], localized cutaneous leishmaniasis caused by L. (V.) braziliensis; MCL, mucocutaneousleishmaniasis caused by L. (V.) braziliensis; Control, healthy individual; n, number of patients per clinical form.
Figure 2
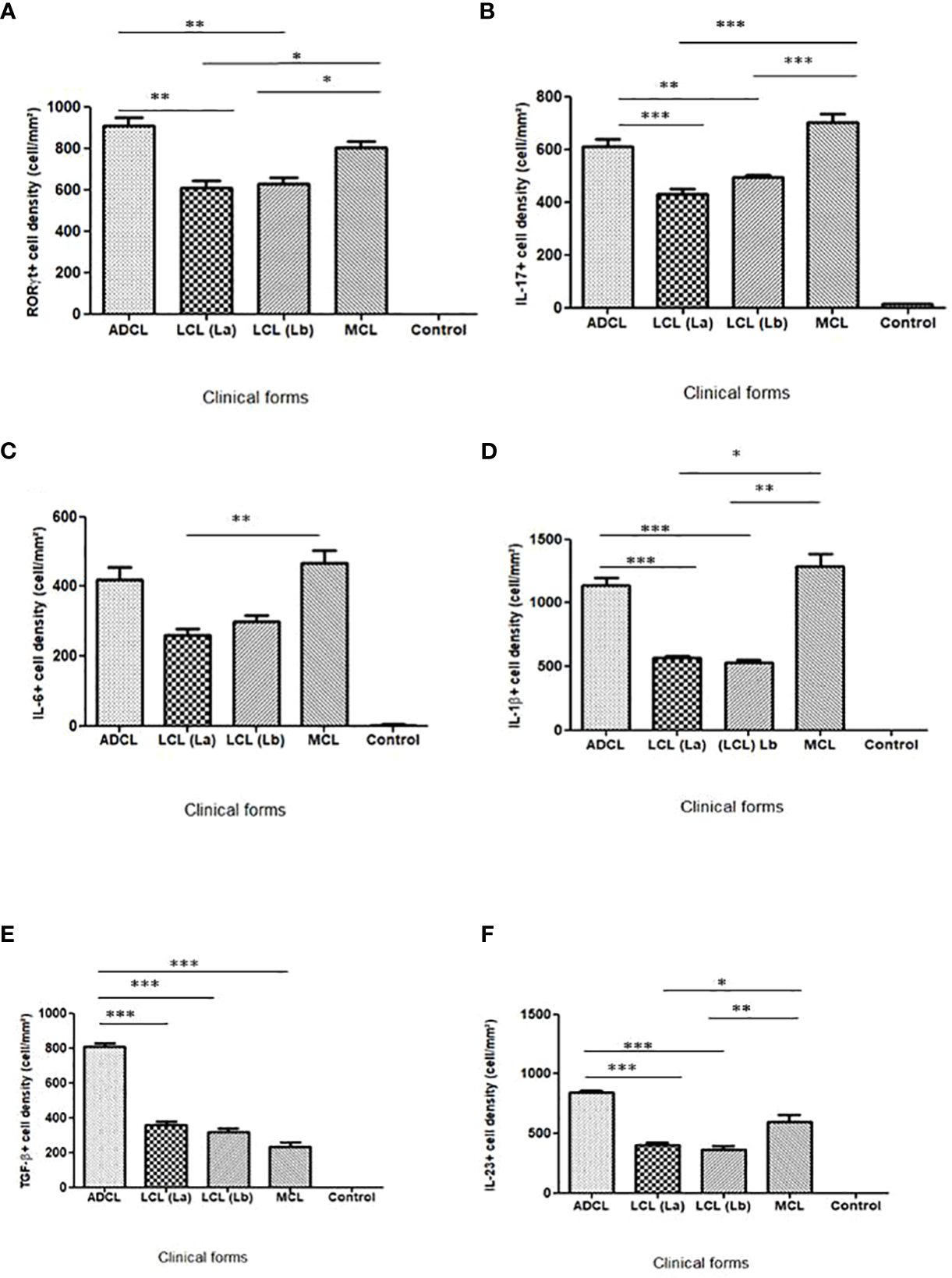
Bar graphs showing the average densities of cells that expressed RORγt, IL-17, IL-6, IL-1β, TGF-β and IL-23 (average+standard error) in histological sections of lesions from patients affected with the different forms clinics of American Cutaneous Leishmaniasis caused by L. (L.) amazonensis and L. (V.) braziliensis and in skin samples from healthy individual. *=p<0.05; **=p<0.001 and ***=p<0.0001. ADCL [La], Anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La], Localized Cutaneous Leishmaniasis caused by L. (L.) amazonensis; LCL [Lb], Localized Cutaneous Leishmaniasis caused by L. (V.) braziliensis; MCL [Lb], Mucocutaneous Leishmaniasis caused by L. (V.) braziliensis.
As seen in the graphs of Figure 2, the expression of the immunological markers of the Th17 profile varied across the ACL spectrum. The immunostaining of the histological sections is shown in Figure 3.
Figure 3

Histological section of the lesions of patients with diferente clinical forms of Cutaneous Leishmaniasis caused by por L. (L.) amazonensis and L. (V.) braziliensis showing immunostaining of cells: RORγt+ (brown staining in the nucleus); and IL-17+, IL-6+, IL-1β+, TGF-β+ and IL-23+ (brown staining in the cell cytoplasm) in the clinical forms of ACL. ADCL [La]: Anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La]: Localized Cutaneous Leishmaniasis caused by L. (L.) amazonensis; LCL [Lb]: Localized Cutaneous Leishmaniasis caused by L. (V.) braziliensis; MCL [Lb]: Mucocutaneous Leishmaniasis caused by L. (V.) braziliensis (stained by immunohistochemistry, 400x).
Analysis of the effect of disease evolution time on immunological mediators of histopathological changes
Considering that the disease evolution time (DET) was very different in the polar forms (ADCL: 12 – 420 months and MCL:1-120 months) compared to the central forms of the spectrum: LCL[La]:1-9 months and LCL[Lb]: 2-18 months (Supplementary Material-Table S1), it was necessary to verify whether this factor was a possible confounding variable when evaluating differences in the expression of cytokines in the ACL spectrum and the correlation between these cytokines. For this purpose, ANCOVA was used to assess whether the clinical form of ACL alone influenced the expression of immunological markers when it was a covariate for DET and the effect of DET on the expression of immunological markers.
In ANCOVA, the density of immunostained cells per field photographed per patient was adopted as the unit of analysis. In total, 302 photographed fields were used: 62 from ADCL, 119 from LCL [La], 82 from LCL [Lb], and 39 from MCL. The mean densities transformed into z scores are listed in Table 2 and are presented as error bar graphs with 95% CIs (Figure 4).
Table 2
| Descriptive statistics | ||||||
|---|---|---|---|---|---|---|
| Clinical forms/ n amostral | Means (z-score) of immunological markers | |||||
| RORγt | IL-17 | IL-6 | TGF-β | IL-1β | IL-23 | |
| ADCL [La] n=62 | 1.11±0.84 | 0.62±0.83 | 0.62±0.97 | 1.63±0.68 | 1.00±0.64 | 0.62±0.83 |
| LCL [La] n=119 | -0.50±0.85 | -0.61±0.83 | -0.624±0.65 | -0.28±0.56 | -0.59±0.31 | -0.61±0.83 |
| LCL [Lb] n=82 | -0.33±0.67 | -0.6±0.68 | -0.07±0.74 | -0.48±0.47 | -0.51±0.69 | -0.6±0.68 |
| MCL [Lb] n=39 | 0.46±0.53 | 1.00±0.93 | 1.04±0.99 | -0.734±0.35 | 1.28±0.99 | 1.00±40.93 |
Descriptive statistics of means (standardized in z-score), standard deviation and sample n (fields photographed and quantified) of cell densities immunostained with RORγt, IL-17, IL-6, TGF-β, IL-1β, and IL-23, according to clinical forms of the AL spectrum.
ADCL [La], anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La], localized cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [Lb], localized cutaneous leishmaniasis caused by L. (V.) braziliensis; MCL [Lb], mucocutaneous leishmaniasis caused by L. (V.) braziliensis. n, number of fields photographed with immunostaining per clinical form.
Figure 4
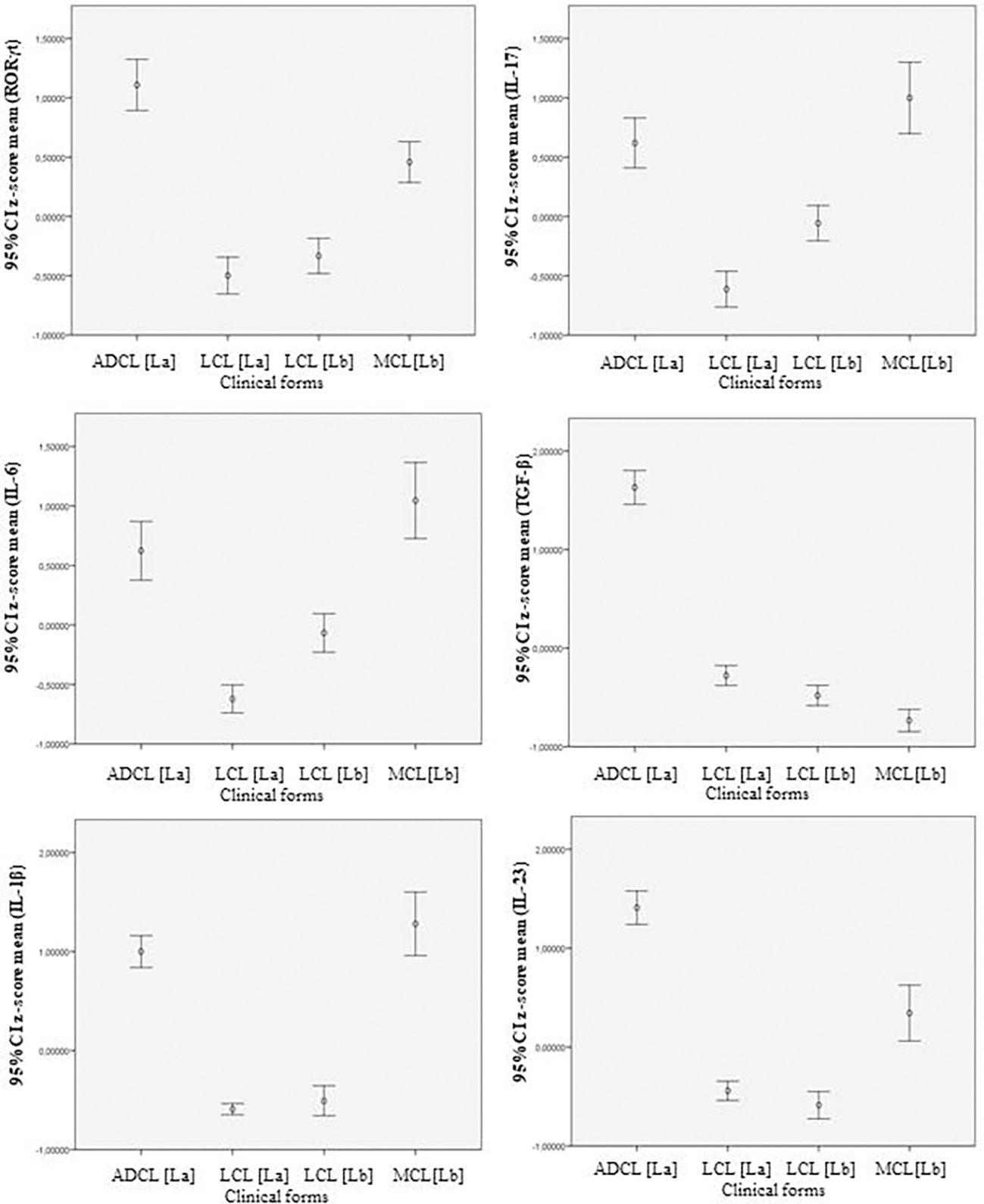
Error bar graphs showing average z-scores of immunostained cells and the 95% confidence intervals between the production of immunological markers in the different clinical forms of Cutaneous Leishmaniasis caused by por L. (L.) amazonensis and L. (V.) braziliensis. ADCL [La]: Anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La]: Localized Cutaneous Leishmaniasis caused by L. (L.) amazonensis; LCL [Lb]: Localized Cutaneous Leishmaniasis caused by L. (V.) braziliensis; MCL [Lb]: Mucocutaneous Leishmaniasis caused by L. (V.) braziliensis. CI: Confidence interval.
The statistically significant difference observed in the expression of the immunological markers of the Th17 profile between some clinical forms, from the analysis of the error bar graph (95% CI) in Figure 4, showed a profile similar to that observed in the mean cell density table of immunostaining in the ACL spectrum, confirming a higher expression of the immune mediators of the Th17 profile in the polar and more severe forms of this endemic disease (Table 1). Next, the sample values of positive cell densities (n = 302) for the different immunological markers of the Th17 profile (six markers) in the four clinical forms of ACL were subjected to ANCOVA to evaluate the differences between the covariate clinical forms with different DETs. The size of the effect of the covariate “time of disease progression” was given in partial η2, and the clinical forms were compared by adopting, randomly, the measurements obtained for the MCL form as a parameter.
The results obtained in ANCOVA were subjected to the post hoc Bonferroni test, which was used to verify possible statistically significant differences between the expression of immunological mediators in the different clinical forms, and it was found that DET influenced the greater expression of RORγt (η=0.018; p=0.023) among the clinical forms ADCL and MCL (p=0.690) and in the expression of IL-6 (η=0.082; p=0.000) among the clinical forms: LCL [La] and LCL [Lb] (p=1.000); ADCL and MCL (p=0.026). Nevertheless, in the production of IL-6, the post hoc test did not show a statistically significant difference between ADCL and LCL caused by L. (V.) braziliensis (p=1.000), previously observed in the error bar graph (Figure 4), showing that the previously observed differences were under the influence of DET (Supplementary Material-Table S2).
We also found the DET influenced the expression of RORγt (η=0.145; p=0,000) among the categories of parasitism (confirmed after the post hoc Bonferroni test): negative and intense (p=1.000); moderate and intense (p=1.000). The effect of DET are observed on production of IL-6 (η=0.198; p=0.000) among: negative and intense (p=0.065); discreet and moderate (p=0.319); moderate and intense (p=0.102); intense and discreet (p=1.000) (Supplementary Material-Table S2).
When evaluating the effect of DET on the types of inflammatory infiltrate, it was found that DET influenced the expression of RORγt (η=0.187; p=0.000) between: discreet and moderate (p=0.371); discreet and intense (1.000). On the production of IL-17, were found effect of DET among: discreet and moderate (p=1.000); discreet and intense (p0.397). Were found effect of DET on production of IL-6 between: discreet and moderate (p0.287); discreet and intense (p=1.000). Were found too effect of DET on production of TGF-β between: moderate and intense (p=0.162). Were found influence of DET on production of IL-1β between discreet and moderate (p=0.286) Supplementary Material-Table S2).
Thus, according to our data, it is possible to conclude that the element that generated the differences in the expression of Th17 profile cytokines previously observed among these histopathological findings is not a peculiar characteristic to each clinical form but the time of disease progression that interferes with the inflammatory response pattern.
The other differences found from the Bonferroni post hoc test reproduced the pattern observed in the error bar graph before the covariation (Figure 4), showing that these differences do not depend on the DET covariate but on some other peculiar characteristic of the different clinical forms of ACL spectrum.
Discussion
In Leishmania infection, in addition to the TCD4+ Th1 and Th2 cell profiles, the participation of other cell types such as Treg and Th17 cells has also been studied to better understand the role of cytokines in the anti-Leishmania immune response and in the search for markers of risk/severity of ACL, in addition to the development of new therapeutic strategies for the disease. Th17 cells (CD4+ IL17+) are the main source of IL- 17 production in the host and can also produce IL-21 and IL-22. Previous findings from our group showed a predominance of cells that expressed IL-17 in situ in the most severe forms of ACL caused by parasites of the subgenera Viannia and Leishmania (25–28).
The present study aimed to characterize, in situ, Th17 cells and their immune mediators and their participation in the cellular immune response of the different clinical forms of ACL, aiming to evaluate the involvement of these cells in the immunopathogenesis of the disease caused by L. (L.) amazonensis and L. (V.) braziliensis. In this direction, it is important to emphasize that the differentiation of TCD4+ RORγt+ cells into classic Th17 cells is stimulated in the presence of IL-6 and TGF-β. Activation of the transcription factor RORγt stimulates the expression of IL-1 and IL-23 receptors (IL-23R and IL-1R1, respectively) and IL-21 production. In addition, cytokines such as IL-23 and IL-1β are involved in the completion and maintenance of the Th17-cell differentiation process (15, 17, 29). However, more recent studies have shown that the differentiation of Th17 cells in the absence of TGF-β, but in the presence of IL- 1β and IL-23, resulted in the “alternative” Th17 underlying, which is considered more pathogenic than the classic Th17. These Th17-cell sub lines have different effector characteristics: while alternative Th17 cells produce IFN-γ and are associated with parasite elimination, classic Th17 cells produce IL-10 and are associated with regulatory and/or suppression of immune response, probably working in synergy with TregFoxp3 cells, whose expression is a strong feature of the ADCL clinical form, thus contributing to the persistence of infection and anergy (15, 17, 29).
Data obtained by our group (unpublished data) show a significantly higher in situ expression of IFN-γ in lesions of patients with the polar MCL form when compared to ADCL and LCL forms, corroborating the participation of this cytokine, along with “alternative” Th17 cells, in the parasite elimination.
The transcription factor RORγt belongs to the family of orphan receptors related to retinoic acid and is directly associated with the differentiation of Th17 cells, acting in the control of the gene expressions such as IL-17 (12). In the present study, higher mean densities of cells expressing RORγt and IL-17 were detected in the polar and severe forms (ADCL and MCL) of ACL caused by L. (L.) amazonensis and L. (V.) braziliensis.
The expression of cytokines related to the Th17 profile in patients with MCL caused by L. (V.) braziliensis from the state of Bahia, Brazil, showed a positive correlation between IL-17 and RORγt mRNA expression, as well as a positive correlation between RORγt and IL-23 mRNA expression, as observed by real-time PCR, showing an association between this transcription factor and cytokines involved in the differentiation and maintenance of Th17 profile cells (29). Gonzalez et al. (2020) evaluated the expression of RORγt+ and IL-17+ in lesions of patients infected with L. (V.) panamensis by immunohistochemistry, finding a positive correlation between these immunological mediators, showing the participation of Th17 profile cells in the pathogenesis of ACL caused by parasites of the subgenus Viannia. Nevertheless, in the same study a negative correlation was seen between the density of cells that expressed the transcription factor RORγt and the presence of amastigotes, suggesting that Th17 cells may contribute to the elimination of parasites; though their contribution to the maintenance of the inflammatory process should not be ruled out (27). Concerning to the subgenus Leishmania, positive correlations between RORγt+ cells and TCD4+ lymphocytes and between TCD4+ and IL- 17+ lymphocytes were demonstrated in the investigation of the skin biopsies from patients with atypical cutaneous leishmaniasis caused by L. (L.) infantum chagasi (26).
Regarding the participation of the other immunological mediators of the Th17 profile evaluated in the present study, we found a high production of IL-6 in the polar and severe forms caused by L. (L.) amazonensis and L. (V.) braziliensis, indicating an association with disease severity, which was also demonstrated by other authors who found a correlation between the in situ expression of IL-6 and Th17 cells in patients infected with L. (L.) infantum chagasi (28), suggesting that IL-6 may play as a marker of ACL severity as occurs in visceral leishmaniasis (28, 30).
A significantly higher density of TGF-β+ cells was also observed in patients with ADCL compared to the other clinical forms, besides the demonstration of the abundant presence of parasitized vacuolated macrophages, suggesting that TGF-β inhibits the activation of these cells and favors an inefficient immune response in these patients, which would lead to the development of nodular lesions replete of parasites, typical of ADCL. Some studies suggest that in the presence of TGF-β, classic Th17 cells are generated, which together with Th2 and TregFoxp3 cells produce regulatory cytokines, such as IL-10, and may be associated with M2 macrophages that have a regulatory role and low expression of antigen and arginase, resulting in parasite survival (15, 31, 32).
IL-17 may contribute to the polarization of M2-type macrophages by inducing the activation of NF-κB, thus showing a broad role of this cytokine (23). These data, corroborates finding of our group that a higher density of M2 macrophages in ADCL contributes to explaining the parasite persistence and severity of infection in this clinical form (31).
In addition, recent studies of our group found a high expression of TregFoxp3+ and IL-10+ cells in ADCL form, compared to MCL form, which may be contributing, along with classic Th17 cells, in suppressing the effector actions of Th1 cells, leading to the anergy pole and persistence of the parasite in the lesion (3, 25).
When analyzing the production of IL-1β, our results showed a significantly higher expression of IL-1β+ cells in patients with the polar and more severe forms of the clinical spectrum caused by L. (V.) braziliensis and L. (L.) amazonensis, in consonance with the expression of this marker in L. (L.) mexicana infection and disease severity (33, 34).
IL-23 expression was increased in the polar forms (ADCL and MCL) caused by L. (L.) amazonensis and L. (V.) braziliensis. These data suggest an important role of this cytokine in the immune response to ACL, since IL-23 is associated with the maintenance and expansion of Th17 cells. This cytokine may contribute to the persistence of inflammation and may be associated with the chronicity of the disease.
Our findings suggest that the immune response related to the Th17 profile favors the disease severity. The increased expression of RORγt, IL-17, IL-6, IL-1β, and IL-23 and the low density of TGF-β+ cells in the MCL suggest that the “alternative” Th17 cells has a pivotal role in MCL. It is also worth mentioning that TGF-β showed high expression in ADCL when compared to LCLs and MCL, which facilitate intracellular Leishmania replication and parasite persistence in this ACL polar form, considering their potent down-regulating effects on macrophages (35).
Studies on the effect of DET in ACL are rare (36), which reinforces the importance of our results. When analyzing the effect of the DET covariate on the expression of the immune mediators of the Th17 profile, we found that DET influenced the expression of RORγt between the clinical forms ADCL and MCL also IL-6 production among ADCL and LCL [Lb]; ADCL and MCL; LCL [La] and LCL [Lb]. Thus, DET represented an important independent variable for the higher expression of these immune mediators of the Th17 profile in the different clinical forms of ACL spectrum, exerting a significant influence in some cases where the disease is severe and more chronic. In addition, RORγt and IL-6 may represent severity factors similar to what occurs in visceral leishmaniasis (30), since the higher density of cells that expressed RORγt and IL-6 under the effect of DET occurred mainly in the polar forms, MCL and ADCL.
When the dependent variable parasitism was analyzed at different intensities (negative, discreet, moderate and intense), DET also influenced on expression of RORγt and IL-6. These findings may be related to the participation of the “alternative” Th17 cells in the clinical forms of the hyperreactivity pole, a process caused by the cellular host response to specific antigens of L. (V.) braziliensis.
Although much remains to be learned about the phenotype, function and regulation of Th17 cells, their participation in the cellular immune response in cutaneous lesions of ACL due to L. (L.) amazonensis and L. (V.) braziliensis seems to begin to clarify. In this way, taken together, our findings show that Th17-profile cells seem to have significant role in the immunopathogenesis of ACL caused by L. (L.) amazonensis and L. (V.) braziliensis, displaying a double role, that is, on the one hand exerting an anti- inflammatory role through the classic action of Th17 cells and favoring immune response suppression and parasitic persistence in ADCL by L. (L.) amazonensis, while on the other hand exerting a clearly inflammatory role through the “alternative” action of Th17 cells and contributing to an exacerbated immune response and parasitic scarcity and tissue damage in MCL by L. (V.) braziliensis.
Statements
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving human participants were reviewed and approved by Ethics Research Committee of the Faculty of Medicine of the University of São Paulo, Brazil, Protocol number CAAE 25714814.0.0000.0065 e CAAE:03952418.3.0000.0065. The patients/participants provided their written informed consent to participate in this study.
Author contributions
GR, LA, JB, FS and CG contributed to the acquisition of the data and drafted the manuscript. CG and FS equally contributed to the present work. GR, MC, CHCM, VM, FS and CG critically helped to interpret the data and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the Sao Paulo Research Foundation (FAPESP) grants #2014/50315-0, #2019/17957-2 and GFR and LSA Scholarship from CAPES (Social Demand: #888.377397/2019-01 and #88887.649146/2021-00) and Laboratório de Patologia de Moléstias Infecciosas (LIM - 50 HC-FMUSP) and Laboratório de Microcirurgia e Cirurgia Plástica, Hospital das Clínicas, Faculdade de Medicina, Universidade de São Paulo. (Dr. Cristina Pires Camargo). Weacknowledge the people who helped in a great way the first steps of this study: Thayse Tomokane and Dr. Kadir Gonzalez.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fitd.2023.1067595/full#supplementary-material
Supplementary Table 1Clinical and histopathological data of patients affected with the different forms of ACL caused by L. (L.) amazonensis and L. (V.) braziliensis. ADCL [La]: anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La]: localized cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [Lb]: localized cutaneous leishmaniasis caused by L. (V.) braziliensis; MCL [Lb]: mucocutaneous leishmaniasis caused by L. (V.) braziliensis. MIR: Montenegro Intradermal Reaction. Parasitism: (-): negative; (+) discreet; (++): moderate; (+++): intense. DET: Disease Evolution Time (months).
Supplementary Table 2Analysis of the effect of the variable DET on the different histopathological findings and production of Th17 profile immunological markers. Main effects of DET on histopathological findings and production of Th17 profile immunological markers. Clinical forms of ACL: ADCL [La]: Anergic diffuse cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [La]: Localized cutaneous leishmaniasis caused by L. (L.) amazonensis; LCL [Lb]: Localized cutaneous leishmaniasis caused by L. (V.) braziliensis; MCL [Lb]: Mucocutaneous leishmaniasis caused by L. (V.) braziliensis. η= effect size; p ≤0,05.
References
1
Espinosa OA Serrano MG Camargo EP Teixeira MMG Shaw JJ . An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as leishmania and Endotrypanum. Parasitology (2018) 145(4):430–42. doi: 10.1017/S0031182016002092
2
Sisleish-Opas/Oms . Dados notificados pelos programas nacionais de Leishmanioses/Serviços de vigilância. Disponível Em (2022) 5:2.
3
Campos MB De Castro Gomes CM de Souza AA Lainson R Corbett CE Silveira FT . In vitro infectivity of species of Leishmania (Viannia) responsible for American cutaneous leishmaniasis. Parasitol Res (2008) 103(4):771–6. doi: 10.1007/s00436-008-1039-8
4
Silveira FT Lainson R Corbett CE . Clinical and immunopathological spectrum of American cutaneous leishmaniasis with special reference to the disease in Amazonian brasil – a review. Mem Inst Oswaldo Cruz (2004) 99(3):239–51. doi: 10.1590/s0074-02762004000300001
5
Silveira FT. What makes mucosal and anergic diffuse cutaneous leishmaniases clinically and immunopathogically different? A review in Brazil. Trans R Soc Trop Med Hyg (2019) 29:trz037. doi: 10.1093/trstmh/trz037
6
Costa JML et al . Leishmaniose cutânea difusa (Lcd) no brasil após 60 anos de sua primeira descrição. Gazeta Médica Da Bahia (2009) 79(3).
7
Silveira FT Müller SR De Souza AAA Lainson R. Gomes CMC Laurenti MD et al . Revisão sobre a patogenia da leishmaniose tegumentar Americana Na amazônia, com Ênfase À doença causada por Leishmania (V.) braziliensis e L. (L.) Amazonensis. Rev Paraense Medicina Pará (2008) 22:9–20.
8
Sacks D Noben-Trauth N . The immunology of susceptibility and resistance to leishmania major in mice. Nat Rev Immunol (2002) 2:845–58. doi: 10.1038/nri933
9
Maspi N Abdoli A Ghaffarifar F . Pro and anti-inflammatory cytokines in cutaneous leishmaniasis: A review. Pathog Glob Health (2016) 110(6):247–60.
10
Gonzalez Carrion Ka. Caracterização das alterações histopatológicas e da resposta imune celular em lesões de Pele de pacientes com leishmaniose cutânea no panamá [Tese]. Faculdade De Medicina Da Universidade De São Paulo (2020).
11
Gonçalves-De-Albuquerque S Pessoa-E-Silva R Trajano-Silva L De Goes Tc De Morais R Da C Oliveira Cn et al . The equivocal role of Th17 cells and neutrophils on immunopathogenesis of leishmaniasis. Front Immunol. (2017) 8:1437. doi: 10.3389/fimmu.2017.01437
12
Castro G Liu X Ngo K De Leon-Tabaldo A Zhao S Luna-Roman R et al . Rorγt and rorα signature genes in human Th17 cells. PloS One (2017) 12(8):e0181868. doi: 10.1371/journal.pone.0181868
13
Dong C . Th17 cells in development: An updated view of their molecular identity and genetic programming. Nat Rev Immunol (2008) 8(5):337–48. doi: 10.1038/nri2295
14
Dong C . Differentiation and function of pro-inflammatory Th17 cells. Microbes Infect (2009) 11(5):584–8. doi: 10.1016/j.micinf.2009.04.001
15
Peters A Lee Y Kuchroo Vk. The many faces of Th17 cells. Curr Opin Immunol (2011) 23(6):702–6. doi: 10.1016/j.coi.2011.08.007
16
Veldhoen M Hocking Rj Atkins Cj Locksley Rm Stockinger B . Tgf beta in the context of an inflammatory cytokine milieu supports De Novo differentiation of il-17-Producing T cells. Immunity. (2006) 24(2):179–89. doi: 10.1016/j.immuni.2006.01.001
17
Ghoreschi K Laurence A Yang XP Tato CM McGeachy MJ Konkel JE et al . Generation of pathogenic Th17 cells in the absence of TGF-β signalling. Nature (2010) 467(7318):967–71. doi: 10.1038/nature09447
18
Jin W Dong C . Il-17 cytokines in immunity and inflammation. Emerg Microbes Infect (2013) 2(9):E60. doi: 10.1038/emi.2013.58
19
Espir Tt Figueira LP Naiff MF Da Costa Ag Ramalho-Ortigão M Malheiro A et al . Role of inflammatory, anti-inflammatory, and regulatory cytokines in patients infected with cutaneous leishmaniasis in Amazonas state, Brazil. The J Immunol Res (2014) 2014:481750. doi: 10.1155/2014/481750
20
Bacellar O Faria D Nascimento M Cardoso Tm Gollob Kj Dutra Wo et al . Interleukin 17 production among patients with American cutaneous leishmaniasis. J Infect Dis (2009) 200(1):75–8. doi: 10.1086/599380
21
Martins Vd Silva Fc Caixeta F Carneiro Mb Go-es Gr Torres L et al . Obesity impairs resistance to leishmania major infection in C57bl/6 mice. PloS Negl Trop Dis (2020) 14(1):E0006596. doi: 10.1371/journal.pntd.0006596
22
Sousa Lm Carneiro Mb Resende Me Martins Ls Dos Santos Lm Vaz Lg et al . Neutrophils have a protective role during early stages of Leishmania amazonensis infection in Balb/C mice. Parasite Immunol (2014) 36(1):13–31. doi: 10.1111/pim.12078
23
Shen J Sun X Pan B Cao S Cao J Che D et al . Il-17 induces macrophages to M2-like phenotype Via nf-κb. Cancer Manag Res (2018) 10:4217–28. doi: 10.2147/cmar.s174899
24
Graça Gc Volpini Ac Romero Ga Oliveira Neto Mp Hueb M Porrozzi R et al . Development and validation of pcr-based assays for diagnosis of American cutaneous leishmaniasis and identification of the parasite species. Mem Inst Oswaldo Cruz. (2012) 107(5):664–74. doi: 10.1590/s0074-02762012000500014
25
Menezes J . Expressão de Foxp3, il-17 e il-23 Na leishmaniose tegumentar Americana causada por Leishmania (Leishmania) amazonensis e Leishmania (Viannia) braziliensis. Dissertação (Mestrado) (2013).
26
Flores G Sandoval Pacheco Cm Sosa Ochoa Wh Gomes C Zúniga C Corbett Cp et al . Th17 lymphocytes in atypical cutaneous leishmaniasis caused by Leishmania L. Infantum Chagasi in central America. Parasite Immunol (2020) 42(11):E12772. doi: 10.1111/pim.12772
27
Gonzalez K Calzada Je Corbett C Saldaña A Laurenti Md. Involvement of the inflammasome and Th17 cells in skin lesions of human cutaneous leishmaniasis caused by Leishmania (Viannia) panamensis. Mediators Inflamm (2020) 27:9278931. doi: 10.1155/2020/9278931
28
Laurenti Md Sosa-Ochoa W Araujo Flores Gv Sandoval Pacheco Cm Tomokane Ty Oliveira L et al . Evaluation of systemic immunity in atypical cutaneous leishmaniasis caused by Leishmania (L.) Infantum Chagasi. Parasite Immunol (2022) 44(1-2):E12896. doi: 10.1111/Pim.12896
29
Boaventura Vs Santos Cs Cardoso Cr De Andrade J Dos Santos Wl Clarêncio J et al . Human mucosal leishmaniasis: Neutrophils infiltrate areas of tissue damage that express high levels of Th17-related cytokines. Eur J Immunol (2010) 40(10):2830–6. doi: 10.1002/Eji.200940115
30
Costa Dl Rocha Rl Carvalho Rm Lima-Neto As Harhay Mo Costa Ch et al . Serum cytokines associated with severity and complications of kala- azar. Pathog Glob Health (2013) 107(2):78–87. doi: 10.1179/2047773213y.0000000078
31
Sandoval Pacheco Cm Araujo Flores Gv Gonzalez K De Castro Gomes Cm Passero L Tomokane Ty et al . Macrophage polarization in the skin lesion caused by Neotropical species of Leishmania sp. J Immunol Res (2021) 2021:5596876. doi: 10.1155/2021/5596876
32
Carneiro Fp De Magalhães Av De Jesus Abreu Almeida Couto M Bocca Al Muniz-Junqueira Mi Ribeiro Sampaio Rn. Foxp3 expression in lesions of the different clinical forms of American tegumentary leishmaniasis. Parasite Immunol (2009) 31(10):646–51. doi: 10.1111/J.1365-3024.2009.01148.X
33
Fernandéz-Figueroa Ea. Rangel-Escareño C Espinosa-Mateos V Carrillo-Sánchez K Salaiza-Suazo N Carrada-Figueroa G et al . Disease severity in patients infected with Leishmania mexicana relates to IL-1β. PloS Negl Trop Dos (2012) 6(5):e1533. doi: 10.1371/Jornal.Pntd.0001533
34
Novais Fo Carvalho Am Clark Ml Carvalho Lp Beiting Dp Brodsky Ie et al . Cd8+ T cell cytotoxicity mediates pathology in the skin by inflammasome activation and il-1β production. PloS Pathog (2017) 13(2):E1006196. doi: 10.1371/Journal.Ppat.1006196
35
Saha A Souravi R Ukil A . Cytokines and signaling networks regulating disease outcomes in leishmaniasis. Infect Immun (2022) 90(8):e0024822. doi: 10.1128/Iai.00248-22
36
Gomes AHS Martines RB Kanamura CT Barbo M Iglezias SD Lindoso JAL et al . American Cutaneous leishmaniasis: In situ imune response of patients with recent and late lesions. Parasite Immunol (2017) 39(4). doi: 10.1111/Pim.12423
Summary
Keywords
American cutaneous leishmaniasis, leishmania(L.)amazonensis, leishmania (V.) braziliensis, cellular immunity, Th17-cells, immunohistochemistry
Citation
Rodrigues GF, Alcântara LS, Barros JPB, de Lima ACS, Campos MB, Moraes C, Ferreira AF, Matta VLR, Laurenti MD, Corbett CEP, Silveira FT and Gomes CMC (2023) In situ expression of Th17 immunologic mediators in American cutaneous leishmaniasis caused by Leishmania (V.) braziliensis and Leishmania (L.) amazonensis in the Brazilian Amazon. Front. Trop. Dis 4:1067595. doi: 10.3389/fitd.2023.1067595
Received
12 October 2022
Accepted
13 January 2023
Published
10 February 2023
Volume
4 - 2023
Edited by
Gisely Melo, Fundação de Medicina Tropical Doutor Heitor Vieira Dourado (FMT-HVD), Brazil
Reviewed by
Rajendranath Ramasawmy, Universidade Nilton Lins, Brazil; Fátima Ribeiro-Dias, Universidade Federal de Goiás, Brazil
Updates
Copyright
© 2023 Rodrigues, Alcântara, Barros, de Lima, Campos, Moraes, Ferreira, Matta, Laurenti, Corbett, Silveira and Gomes.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fernando T. Silveira, fernandotobias@iec.gov.br; Claudia Maria Castro Gomes, gomescla@usp.br
This article was submitted to Neglected Tropical Diseases, a section of the journal Frontiers in Tropical Diseases
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.