
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Trop. Dis. , 18 March 2022
Sec. Major Tropical Diseases
Volume 3 - 2022 | https://doi.org/10.3389/fitd.2022.845451
This article is part of the Research Topic Insights In Tropical Diseases: 2021 View all 6 articles
 Jaiana L. M. Cardoso1,2
Jaiana L. M. Cardoso1,2 Yanka E. A. R. Salazar1Anne C. G. Almeida1,2Laila R. A. Barbosa1,2Emanuelle L. Silva1
Yanka E. A. R. Salazar1Anne C. G. Almeida1,2Laila R. A. Barbosa1,2Emanuelle L. Silva1 Maria Gabriela Almeida Rodrigues1,2Fernanda Rodrigues-Soares3
Maria Gabriela Almeida Rodrigues1,2Fernanda Rodrigues-Soares3 Vanderson S. Sampaio1,2
Vanderson S. Sampaio1,2 André M. Siqueira4
André M. Siqueira4 Marcus V. G. Lacerda1,2,5
Marcus V. G. Lacerda1,2,5 Wuelton M. Monteiro1,2
Wuelton M. Monteiro1,2 Gisely C. Melo1,2*
Gisely C. Melo1,2*Background: The influence of the CYPs (cytochrome P-450) in the success of antimalarial therapy remains uncertain. In this study, the association of CYP2D6, CYP2C19 and CYP3A4 polymorphisms and predicted phenotypes with malaria recurrence was investigated.
Methods: After diagnosis of vivax malaria, individuals treated at a reference center in Manaus were followed up for 180 days. Patients were separated into two groups: a recurrence group and a non-recurrence group. Genotyping of CYP2D6, CYP2C19 and CYP3A4 was performed using a TaqMan™ assay and real-time PCR.
Findings: The frequencies of decreased-function and normal-function alleles and phenotypes for all CYPs were similar between the groups, except for the CYP2D6*2xN allele (p=0.047) and the CYP2D6 gUM phenotype (p=0.057), which were more frequent in individuals without recurrence. Despite this, the CYP2D6, CYP2C19 and CYP3A4 genotypes had no association with an increased risk of recurrence. CYPs polymorphisms also had no influence in parasite clearance, neither in the time nor the number of recurrence episodes. MAIN
Conclusion: This prospective cohort study demonstrated that CYP2D6, CYP2C19 and CYP3A4 polymorphisms have no influence on malaria recurrence. Nonetheless, our findings suggest that the CYP2D6 predicted ultrarapid phenotype was less susceptible to recurrence, and that patients with the CYP2D6 gUM phenotype are less susceptible to primaquine failure. Additional investigation of pharmacogenetics and pharmacokinetics are needed before implementing CYP analysis to better orientate individualized radical treatment of vivax malaria in reference centers that treat patients with multiple recurrences.
Malaria is a globally distributed infectious disease and, in 2020, 241 million cases and 627 thousand deaths from malaria were reported worldwide (1). In Brazil, for the same period, approximately 145,188 cases and 42 deaths were recorded, of these, 82.5% were caused by Plasmodium vivax (1). P. vivax has certain characteristics, such as its ability to remain latent in the liver in the form of hypnozoites that can cause relapses when reactivated, that are an obstacle for its elimination (2, 3).
Primaquine (PQ), a hypnozoiticidal drug, needs to be biotransformed into an active metabolite in order to exert its antimalarial effect (4). Biotransformation occurs through two main pathways: cytochrome P-450 (CYPs) and monoamine oxidase (MAO-A) (5). Biotransformation mediated by CYPs is attributed to the enzymes CYP2C19, CYP2D6 and CYP3A4 (6). The MAO-A pathway generates aldehyde derivatives, such as carboxyprimaquine, a predominant but inert metabolite (6, 7).
The contribution of the human host’s genetics to the results of antimalarial treatment has been demonstrated in recent studies (8–11). Bennet et al. (8) reported therapeutic failure of PQ in the treatment of vivax malaria, which was attributed to the presence of polymorphism in CYP2D6, thus resulting in decreased biotransformation of the drug and, consequently, low levels of the active metabolites. Baird et al. (9) showed that decreased CYP2D6 activity was associated with an increased risk of therapeutic failure, which suggests a relation between CYP2D6 with PQ biotransformation and an increased risk of relapse. The relation between the therapeutic failure of PQ and the presence of polymorphism in CYP2D6 has also been reported in Brazil (10–12).
Pybus et al. (6) demonstrated that MAO-A and CYP2D6 are responsible for 93% of PQ metabolites and that only traces levels of oxidated and dimethylated metabolites are generated by CYP3A4 and CYP2C19. Ariffin et al. (13) associated the presence of the CYP2C19*2 allele with the decrease in PQ biotransformation into carboxyprimaquine (13). This null activity allele has also been related to the decrease in biotransformation of other antimalarial drugs (7, 14, 15). Furthermore, CYP3A4 has a role in the biotransformation of PQ, but no clear genotype-phenotype association for this enzyme, and the effect of its variants remains controversial (6, 16–18).
Polymorphic variants of CYP2C19, CYP2D6 and CYP3A4 have been reported in populations in Brazilian Amazon, as well as incidences of P. vivax malaria infection and recurrent infections by the parasite. Genetic variability of these CYPs is likely to have an impact on the clinical response of patients, and the detection of genotypes related to PQ metabolism could guide individualized treatment protocols in the future in patients with multiple recurrences. As such, this study aimed to investigate the frequency of genotypes and phenotypes of CYP2D6, CYP3A4 and CYP2C19 and their association with recurrence of P. vivax in patients from the Brazilian Amazon.
This study was approved by the Ethics Review Board of Fundação de Medicina Tropical Dr Heitor Vieira Dourado (FMT-HVD) (CAAE 0002.0.114.000-11 and 44605015.4.0000.0005). The individuals invited to participate were informed about the objectives of the study and signed an informed consent form. In the case of individuals under 18 years old, an assent form was signed by the parents or a legal representative.
This study was conducted at FMT-HVD, a reference center for infeccious diseases in Manaus, Brazil, in the periods from 2012 to 2014 and 2016 to 2017. The study included individuals of either gender, aged 6 months or older, with a bodyweight of greater than 5 kg, blood parasite density from 250 to 100,000 parasites/mL and axillary temperature of 37.5°C or history of fever in the last 48 hours. Use of antimalarial drugs in the previous 30 days, refusal to be followed up, pregnancy, or any clinical complication were considered to be non-inclusion criteria.
All individuals were treated with 25 mg/kg of chloroquine phosphate (CQ) for 3 days (10 mg/kg on day 1 and 7.5 mg/kg on days 2 and 3). A dose of 0.5 mg/kg/day of PQ was administered for 7 days, together with CQ or on the 42nd day. Clinical and laboratory tests were performed, and interviews and sample collection were done on D1, D2, D3, D4, D7, D14, D28 and D42 of follow-up. If there were any extra days of follow-up, the same sample collection procedures were performed.
This study was carried out using convenience sampling from other previous follow-ups. The individuals were alocated in two groups: a recurrence group (patients with at least one episode of recurrence in the last 180 days) and a non-recurrence group (those who had no reports of malaria episode in the last 180 days). The dates of the recurrence episodes were obtained during the follow-up or by passive detection via the national SIVEP-Malaria system (Malaria Epidemiological Surveillance System), which is the official malaria epidemiological surveillance system in Brazil.
Asexual parasitemia and gametocytemia, as well as clearance of parasitemia, were determined using optical microscopy. All diagnostics were performed by an experienced microscopist using parasite counts per 500 leukocytes.
Genomic DNA was purifed from whole blood samples using the QIAmp® Blood Mini kit (Qiagen, Hilden, Germany). The single-nucleotide polymorphisms (SNPs) that were genotyped were chosen according to their functional importance and their frequency in the Brazilian population. We selected eleven polymorphisms in CYP2D6 (2549delA [rs35742686], 100C>T [rs1065852], 1846G>A [rs3892097], 4180G>C [rs1135840], 2988G>A [rs28371725], 3183G>A [rs59421388], 1584C>G [rs1080985], 1023C>t [rs28371706], 2615_2617delAAG [rs5030656], 31G>A [rs769258], 2850C>T [rs16947]); one polymorphism in CYP2C19 (681G>A [rs4244285]); and one polymorphism in CYP3A4 (-392A>G [rs2740574]). The choice of CYP2D6, CYP2C19 and CYP3A4 genes and polymorphisms was based on data from previous work with populations from the Amazon region, which dealt with the influence of CYP450 gene polymorphisms on the response to antimalarials (11, 19, 20), as well as the frequencies of the alleles already described in the Brazilian population (21).
The analysis was performed via an allelic discrimination assay using TaqMan™ probes and a real-time PCR System (7500 Fast, Applied Biosystems Foster City, CA). CYP2D6 haplotypes were inferred using the HaploStats package (version 1.7.7) implemented in the R platform (www.r-project.org). The haplotypes identified were compared to the Human Cytochrome P450 (CYP) Allele Nomenclature Database, for star (*) allele designation. The wild-type allele (*1) was determined when there was no mutated allele (13).
The determination of the predicted CYP2D6 phenotype was performed according to the activity score system (AS). Alleles were grouped according to their perceived functionality: zero, null-function alleles (*4,*4xN,*5); 0.5, decreased-function alleles (*9, *17, *29, *41) and 0.25 (*10); 1, normal-function alleles (*1, *2, *35, *39); and 2 or more, increased-function alleles (*1xN, *2xN), depending on the number of copies (CNV). The predicted phenotype was obtained from the sum of diplotype activity scores. For heterozygous individuals with variations in the number of copies, this was accounted for in the activity score calculation, even when it was not possible to attribute the multiplication with complete certainty to one of the alleles of these individuals. Individuals with AS = 0 and AS >2.5 were designated as genetically poor and ultrarapid metabolizers (gPM and gUM), respectively. On the other hand, individuals with AS = 0.25-1 and AS = 1.25-2.25 were designated as genetically intermediate and normal metabolizers (gIM and gNM), respectively (22).
The predicted CYP2C19 phenotype was classified as a normal metabolizer (*1/*1), intermediate metabolizer (*1/*2) or poor metabolizer (*2/*2) (23). For CYP3A4, the presence or absence of the CYP3A4*1B was analyzed, without the inference of the phenotype, since there are no guidelines for its prediction.
Difference in frequency values of the star alleles of CYP2D6, CYP2C19 and CYP3A4 were assessed using the chi-square test. To evaluate the influence of the genotype in the recurrence and clearance time, the survival analysis method was applied through Kaplan-Meier curves and the Wilcoxon-Breslow-Gehan test of equality. Poisson regression was used to assess the effect of genotype in the number of recurrence episodes. The association between the genetic polymorphisms of CYP2D6, CYP3A4 and CYP2C19 and the clinical response of patients who were treated with PQ was assessed using a multiple log-binomial generalized linear regression model. Analyses were performed using the software Stata v. 13.
A total of 311 patients diagnosed with P. vivax malaria were selected for the study, of which 102 presented recurrence and 209 had no recurrence. Of all the 311 patients recruited for the study, 256 samples were genotyped for CYP2D6, 309 samples for CYP3A4 and 303 samples for CYP2C19. Genotyping of the 3 CYPs was achieved for 249 samples (Figure 1).
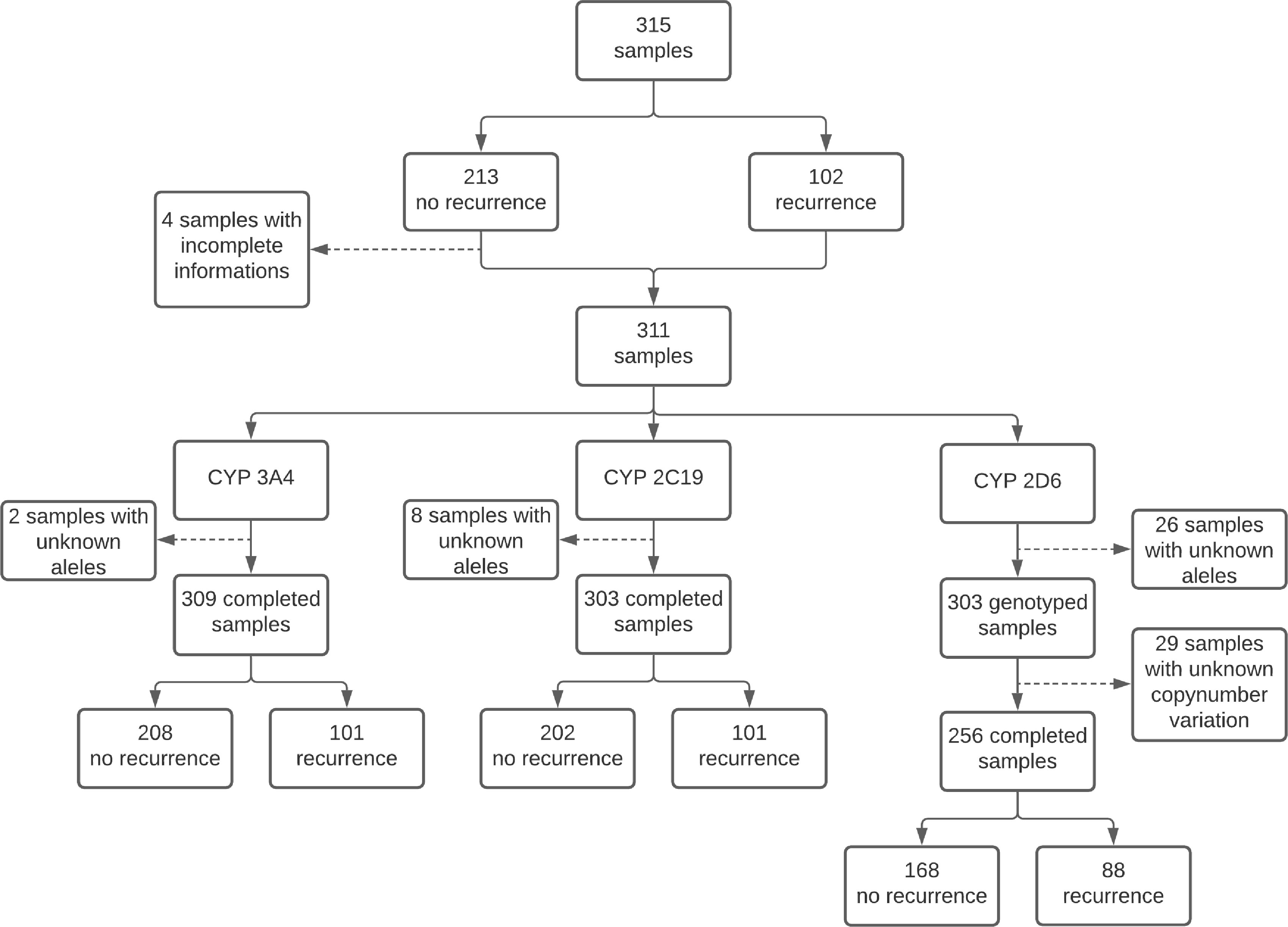
Figure 1 Flowchart of processed samples.
The baseline characteristics of the participants are shown in Table 1. Between the two groups, gender distribution was similar. The mean age for patients with recurrence and without recurrence was 40 and 36.7 years, respectively (p=0.036). For malaria recurrence episodes, 80.4% had only one recurrence episode, which occurred between 61-120 days (56.9%) after the initial malaria episode, with a mean time of 85.5 days (95%CI 78.4-92.6).
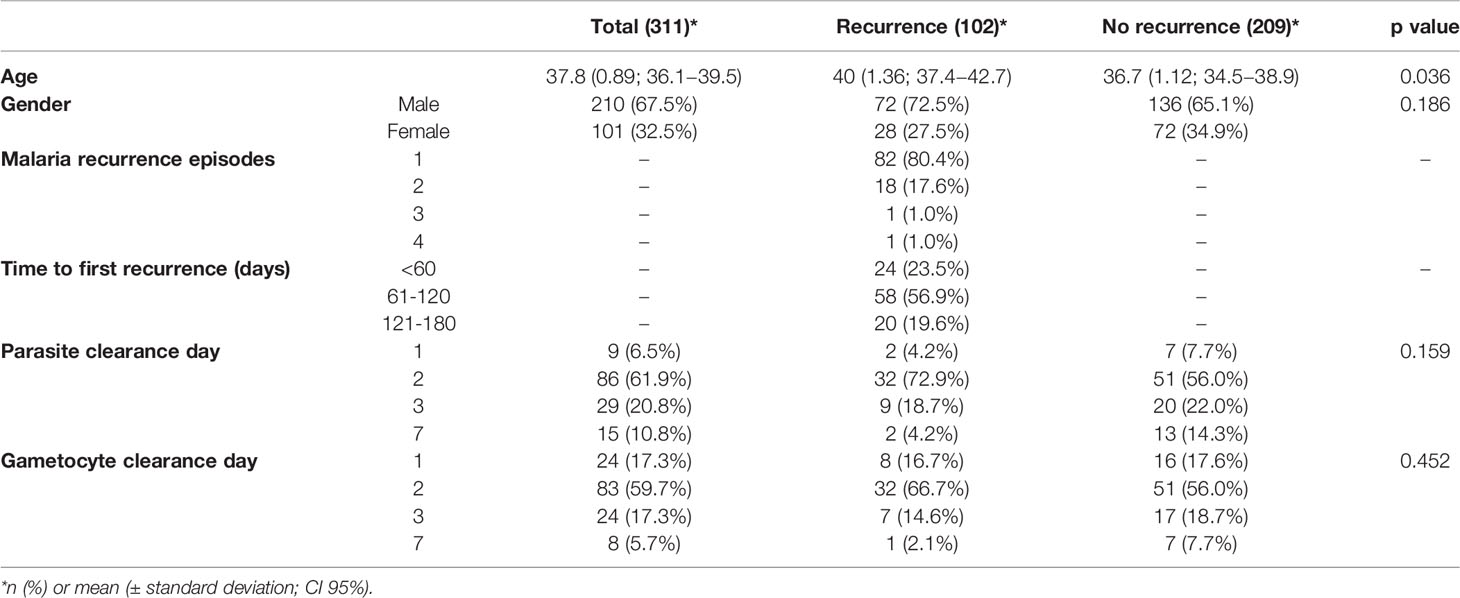
Table 1 Baseline characteristics of individuals involved in this study.
The asexual parasite clearance and gametocyte clearance analysis was performed with 147 individuals that took PQ on day 1 of their treatment. The clearance of asexual parasitemia on D1, D3 and D7 was similar between patients with and without recurrence (p>0.05). On D2, the clearance occurred in 72.9% of patients with recurrence and 56.0% of patients without recurrence (p=0.05). For gametocyte clearance, there was no significant difference between the groups according the clearance day (p>0.05).
The allele frequency distribution of CYP2D6, CYP2C19 and CYP3A4 and of predicted CYP2D6 and CYP2C19 phenotypes are presented in Table 2. The star allele frequencies of CYP2D6, CYP2C19 and CYP3A4 were similar between the groups (p>0.05). The CYP2C19*2 null function allele was present in 9.8% and 10.5% of the patients with and without recurrence, respectively (p=0.924). The CYP3A4 *1B allele had similar frequency in both groups (p=0.841) (Table 2). The proportions of CYP2D6 null function alleles (*4 and *5), decreased function alleles (*9, *10, *17, *29 and *41) and normal function alleles (*1, *2, *35 and *39) were similar between the groups (p>0.05). After the analysis of CNV, it was found that 6.3% of individuals genotyped for CYP2D6 had multiplications and, of the 32 alleles of these individuals, it was possible to attribute the multiplications to only 11 alleles of homozygous individuals. CYP2D6 ultrarapid alleles (*1xN and *2xN) were observed only in patients without recurrence (p=0.018).
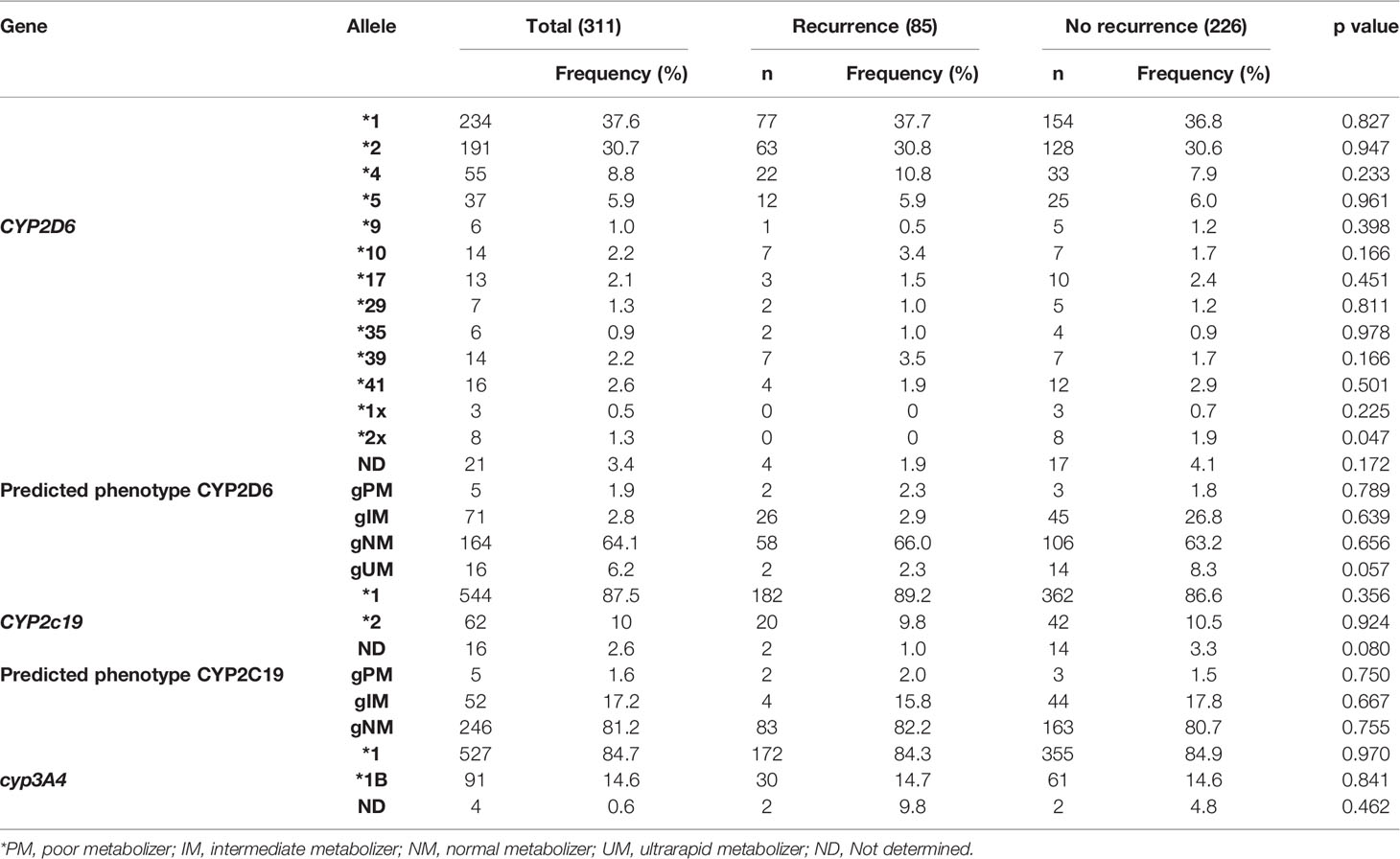
Table 2 Allele frequency of CYP2D6, CYP2C19 and CYP3A4 and predicted CYP2D6 and CYP2C19 phenotypes.
The frequency of the predicted CYP2D6 phenotype was not different between the groups (p=0.372). The most frequent phenotype in both groups was the normal metabolizer phenotype (p=0.656). Although the ultrarapid alleles were found only in individuals without recurrence, the calculation of the activity score revealed the occurrence of the ultrarapid phenotype in patients with recurrence and without recurrence, 2.3% and 8.3% respectively (p=0.057) (Table 2).
The frequency of the predicted CYP2C19 phenotype was similar in both groups (p=0.873). The frequency of gPM was 2.0% and 1.5% in individuals with recurrence and without recurrence, respectively (p=0.750). The frequency of gIM was 15.8% in the recurrent group and 17.8% in non-recurrent group (p=0.667).
The predicted CYP2D6 phenotype did not differ in frequency between the days of asexual parasite clearance (p=0.066). However, even with no statistical significance, it was observed that asexual parasitemia clearance occurred earlier in individuals with the ultrarapid metabolizer phenotype (p=0.108). There was no difference in the asexual parasitemia clearance according to the predicted CYP2C19 phenotype and CYP3A4*1B (p>0.05) (Table 3).
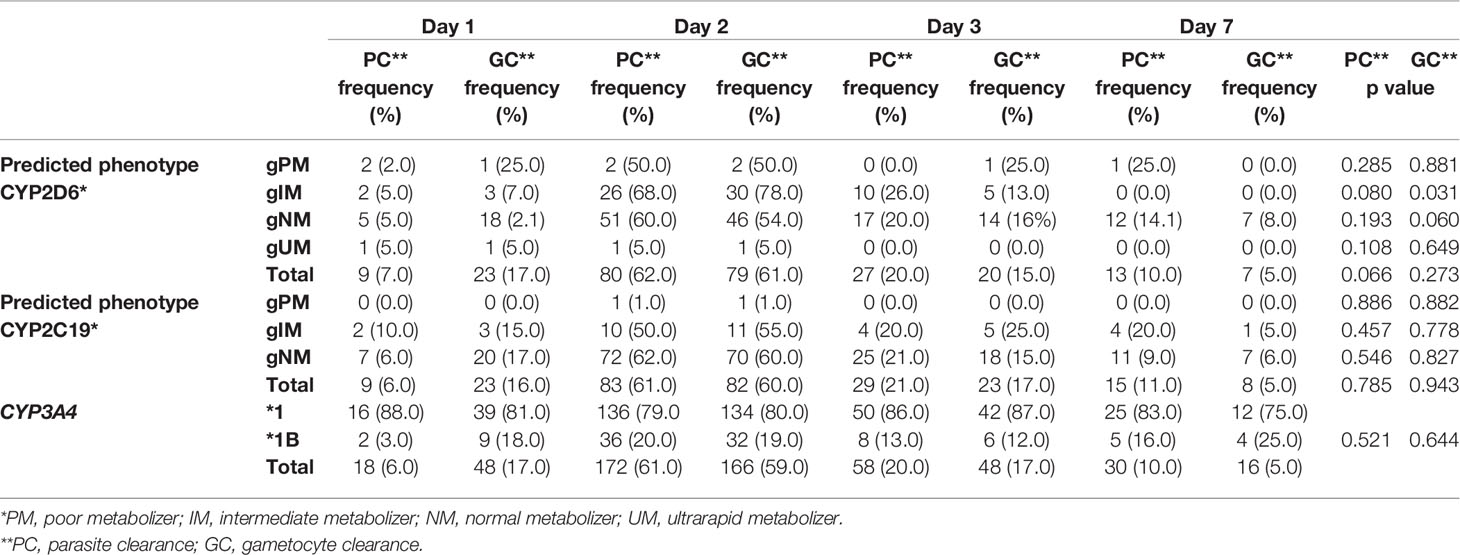
Table 3 Predicted phenotype of CYP2D6, CYP2C19 and CYP3A4 allele x parasite clearance day.
The Kaplan-Meier analysis of the time to asexual parasitemia clearance was not significant for the predicted CYP2D6 and CYP2C19 phenotype and the presence CYP3A4*1B (p>0.05) (Figure 2).

Figure 2 Malaria parasite clearance between individuals with different CYP2D6 and CYP2C19 phenotypes. Kaplan–Meier curve shows malaria parasite clearance time between individuals with normal and reduced predicted CYP2D6 and CYP2C19 activity phenotype and normal and mutated CYP3A4 allele. (A) CYP2D6 phenotype; (B) CYP2C19 phenotype; (C) CYP3A4 allele; PM, poor metabolizer; IM, intermediate metabolizer; NM, normal metabolizer; UM, ultrarapid metabolizer. The symbol * refers to the star allele.
No association was found between gametocytes clearance and predicted CYP2D6 and CYP2C19 phenotype and the presence of CYP3A4*1B (p=0.576, p=0.676 and p=0.535, respectively) (Figure 3 and Table 3).
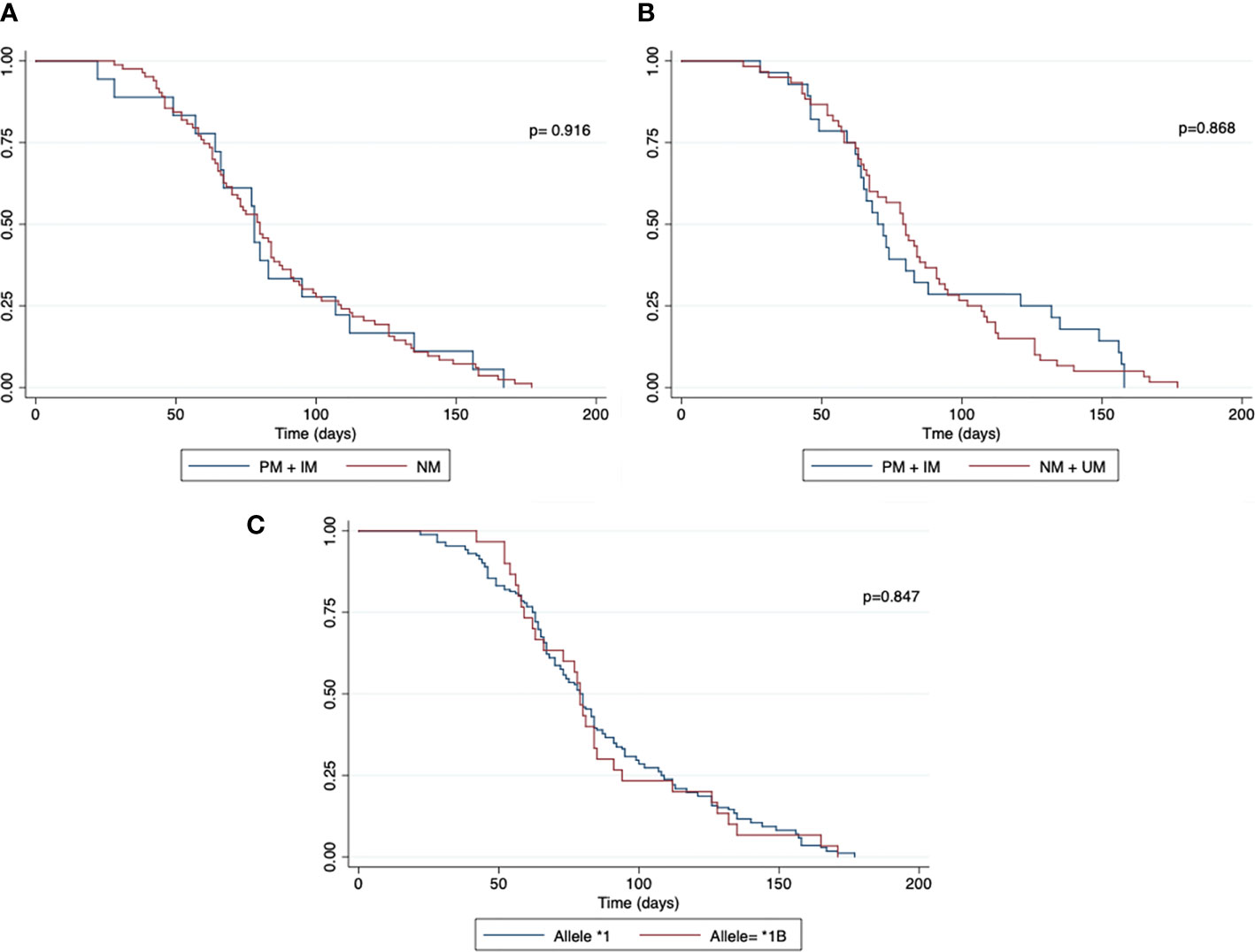
Figure 3 Malaria gametocyte clearance between individuals with different CYP2D6 and CYP2C19 phenotypes. Kaplan–Meier curve shows malaria gametocyte clearance time between individuals with normal and reduced CYP2D6 and CYP2C19 activity phenotype and normal and mutated CYP3A4 allele. (A) CYP2D6 phenotype; (B) CYP2C19 phenotype; (C) CYP3A4 allele; PM, poor metabolizer; IM, intermediate metabolizer; NM, normal metabolizer; UM, ultrarapid metabolizer. The symbol * refers to the star allele.
The number of malaria recurrence episodes did not differ for the predicted phenotype of the CYP2D6, CYP2C19 and CYP3A4 allele (p>0.05). Additionally, there were no significant differences in the occurrence of one or more malaria episodes (CYP2D6 p=0.455, CYP2C19 p=0.832 and CYP3A4 p=0.437).
For CYP2D6 and CYP2C19, the time until the first recurrence episode was similar between the two groups (poor metabolizer + intermediate metabolizer x normal metabolizer) (p=0.868 and p=0.916). For CYP3A4, the length of time did not differ with the presence of CYP3A4*1B (p=0.847) (Figure 4).
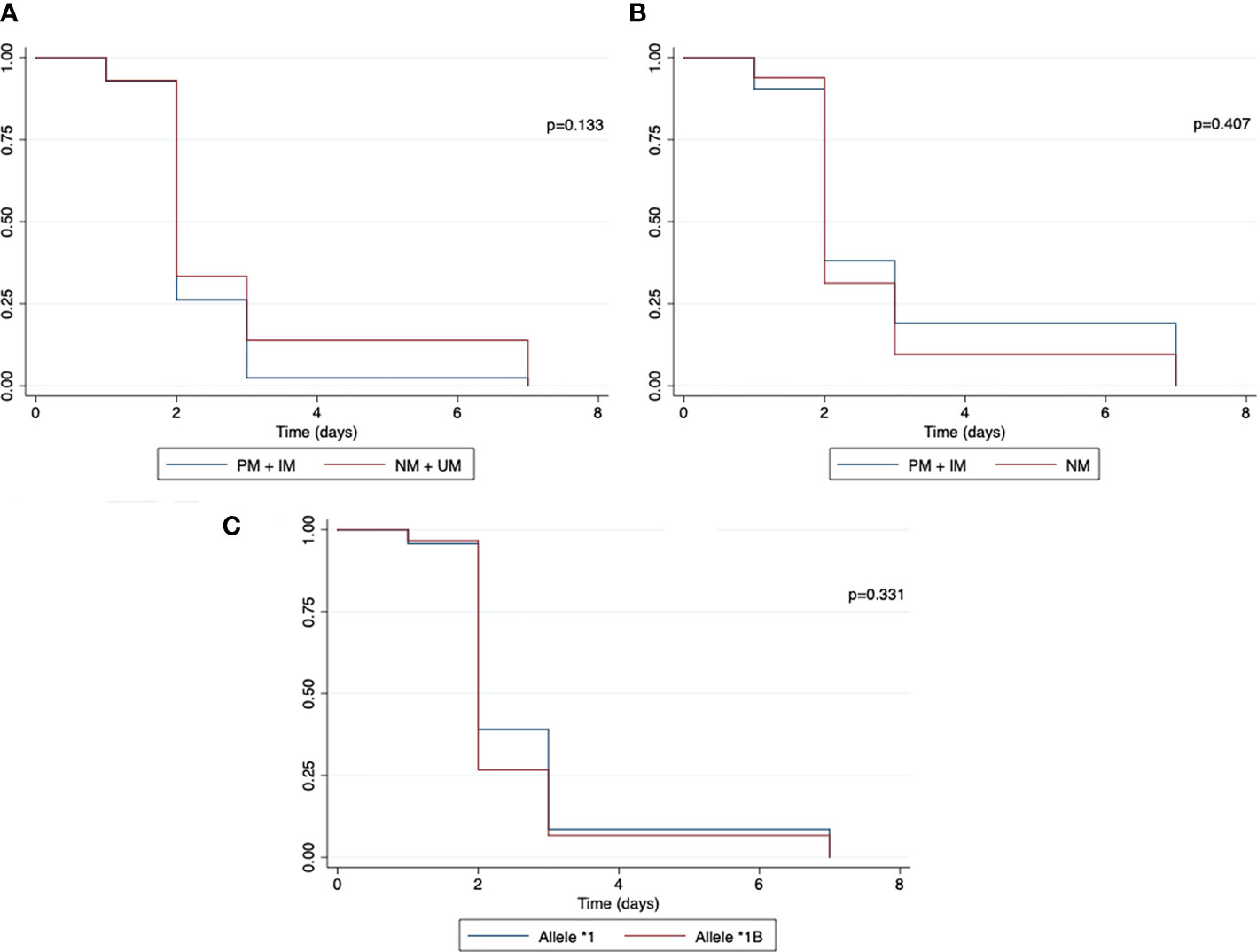
Figure 4 Kaplan–Meier curve for malaria recurrence time between individuals with normal and reduced CYP2D6 and CYP2C19 activity phenotype and normal and mutated CYP3A4 allele. (A) CYP2D6 phenotype; (B) CYP2C19 phenotype; (C) CYP3A4 allele; PM, poor metabolizer; IM, intermediate metabolizer; NM, normal metabolizer. The symbol * refers to the star allele.
The relative risk (RR) was measured to evaluate the influence of predicted CYP2C19 and CYP2D6 phenotype, CYP3A4*1B and combined CYP2C19+CYP2D6 in the clinical response of patients treated with PQ (Table 4). The RR of recurrence with classification CYP2C19 gPM or gIM was 0.93 (95%CI, 0.61-1.42; p=0.758) when compared with a classification of normal metabolizer. For CYP2D6, the RR with decreased predicted phenotype (gPM+gIM) was 1.10 (95%CI, 0.77-1.58; p=0.585) in comparison with normal and ultrarapid metabolizers. There was no association between recurrence and CYP3A4*1B (95%CI, 0.73-1.38; p=0.951).

Table 4 Relative risk for recurrence associated with CYP2C19 and CYP2D6 phenotype, CYP3A4 genotype and combined CYP2C19+CYP2D6.
Finally, the association between CYP2C19+CYP2D6 and recurrence was also measured. The presence of one or two gPM or gIM alleles in any of the CYPs was not associated with risk of recurrence (p=0.972).
This study is the first to simultaneously evaluate the influence of CYP2D6, CYP2C19 and CYP3A4 genetic polymorphisms in relation to the recurrence of P. vivax in the Brazilian Amazon. Since some patients followed up in this study come from urban areas, where the vector mosquito is absent, the cause of these recurrences could be attributed to P. vivax relapses (24). This causes problems in the elimination of malaria, since these individuals can directly contribute to the maintenance of local transmission. In addition, hypnozoites are difficult to eradicate, as they can only be eliminated by treatment with primaquine, which may have its effectiveness reduced due to altered metabolism of CYP2D6 (6, 8) With the constant decline in malaria transmission in some areas of the Brazilian Amazon (25, 26), protocols for personalized therapy could become more viable and their implementation could be possible in referral centers.
In this study, after genotyping of 11 SNPs, no significant difference was found between CYP2D6 allele frequencies and patients with or without recurrence (p=0.646). In addition, increased function alleles were only observed in patients without recurrence (p=0.018). In general, the frequency of these alleles is low and since the majority of the studies are conducted to evaluate the association of impaired genotypes and PQ failure, this is the first study to report a high frequency of CYP2D6 multiplication in individuals without recurrence.
Furthermore, the gUM phenotype presented a higher frequency in those without recurrence (p=0.057). Some studies have shown that the presence of the gUM phenotype is related to the high risk of toxicity of the prodrug codeine, and a high risk of failure of tricyclic antidepressants (27, 28). However, for individuals with the gUM and gNM phenotype, it is expected that a standard dose of tamoxifen should achieve the therapeutic effect in breast cancer treatment (29). For PQ, it is supposed that individuals with the gUM phenotype would exhibit the fastest metabolism through the CYP2D6 pathway. Gonçalves et al. (30) evaluated the effect of the CYP2D6 activity score (AS) on PQ plasma concentration over time after a single dose of PQ, and showed that the prodrug concentration decreased faster in children with AS=3 than those with AS ≤ 2. Moreover, the clinical effect of the gUM phenotype is still unknown (5). In our study, the frequencies of the CYP2D6 gPM, gIM and gNM phenotypes were similar between those with or without recurrence, similar to what has previously been observed in the Brazilian Amazon region (11).
The influence of decreased CYP2D6 phenotypes (PM+IM) in the recurrence was not associated with high risk of recurrence. No significant effect of predicted CYP2D6 phenotype in recurrence episodes was reported in Australia and Thailand (31, 32). However, the effect of the CYP2D6 impaired phenotype and the increased risk of PQ failure has been demonstrated in other studies, thus suggesting that PQ metabolism is dependent on CYP2D6 (8, 9, 11, 33).
The Brazilian Amazon is a region that is endemic for malaria; therefore, there is a possibility that part of the unidentified recurrences could be reinfections and relapses due to primaquine failure. This could be the cause of the disagreements between the results of this study and other previously published studies (8–11). Furthermore, there are differences regarding the system for CYP2D6 phenotype prediction from the genotype; specifically, regarding the activity index and the cut-off points used to predict CYP2D6 activity levels (22, 34). Some studies have already reported inhibition of CYP2D6 with the use of chloroquine, which would affect the actual phenotype of the enzyme and the response to primaquine (35, 36).
In this study, we did not find any association between the CYP2D6 predicted phenotype and gametocytemia or asexual parasitemia clearance. A recent study in the Amazon region also did not show any association between the presence of polymorphisms in CYP genes and early asexual clearance of P. vivax (37). However, Pett et al. (38) demonstrated the effect of the CYP2D6 gPM/gIM phenotype in P. falciparum gametocyte clearance, and it was also demonstrated that the biotransformation of PQ is not a condition for the eradication of the blood stages (asexual and sexual) of P. berghei (39). The CYP2D6 predicted phenotype was also not associated with the number of recurrence episodes and neither with the time to the first episode of recurrence, similar to previously observed (11, 33).
In our study, the frequency of this allele is in agreement with what is already known for northern Brazilians (40, 41) (Table 2). Even though the PM phenotype was more often observed in individuals with recurrence, the frequencies of PM and IM phenotypes were similar between the groups. Furthermore, this study demonstrated that there is no association between the CYP2C19 polymorphism and the recurrence episodes neither with asexual parasitemia clearance nor gametocyte clearance. Pybus et al. (6) showed that CYP2C19 plays a role in PQ metabolism, although the influence of this CYP in the production of PQ’s hemolytic and/or therapeutic metabolite is not clear.
For CYP3A4*1B, no statistical difference was found in the allele frequency between the individuals with or without recurrence (p=0.762). Several studies have been conducted to explain the importance of polymorphisms in the CYP3A4 genotype–phenotype relationship, but the functional effect of the polymorphism remains contentious (15, 42). Although CYP3A4 is known to have a role in PQ biotransformation (6), we could not show the effect of CYP3A4*1B in the recurrence episodes in individuals treated with PQ, neither in the asexual parasitemia clearance nor in gametocyte clearance. A previous study conducted in the Amazon also showed no association between this allele and the recurrence episodes nor between the presence of CYP3A4*1B and the clearance time (37).
Because PQ is not metabolized by only one enzyme (6, 12), a multigenic analysis might be more suitable for determining the effect of the CYP450 polymorphism in PQ failure. In our study, we found a single individual representing the CYP2D6 gPM (*4/*4) + CYP2C19 gIM (*1/*2) that had had 2 recurrence episodes. Despite this, a CYP2D6 + CYP2C19 analysis was not associated with a higher risk of recurrence.
This genotype-based prediction assumes that the genotyping precisely corresponds the metabolic activity (42, 43). However, the prediction of phenotype from the genotype has not been correlated with complete certainty, since the gene splicing, SNPs, epigenetics, microRNA, transcription regulation and multiple gene copies are factors that can modify the enzymatic activity (44–47). Furthermore, phenoconversion may occur, whereby a genotypic gNM can be converted into a transient phenotypic IM or PM, which is mainly caused by extrinsic factors, such as drug-drug interactions and some pro-inflammatory cytokines in the inflammatory process (43, 44, 48).
This study had some limitations. The phenotype was predicted by the genotype. Recently, Baird et al. (9) showed a highly significant correlation between the genotype-determined activity score and the measured phenotypes when using dextromethorphan as a probe drug. Primaquine metabolites were not assessed to confirm the impact of CYP2D6 in the pharmacokinetics of the drug. Drug-drug interactions and the presence of other inflammatory conditions were not evaluated. It was not possible to certify factors related to the quality of the CQ and PQ drugs administered to patients and PQ administration was not supervised. The low sample size for individuals with recurrence found here is in agreement with other studies (9, 11). Moreover, data regarding recurrence episodes were accessed using the SIVEP-Malaria platform, which is susceptible to underreporting issues.
In this study, it was not possible to establish an association between CYP2D6, CYP3A4, and CYP2C19 decreased metabolizing genotypes and recurrence of P. vivax. The investigation of other causes of recurrence for these individuals is necessary since it is a region where there is a high risk of re-infection and many asymptomatic infections (49).
This prospective cohort study demonstrated no influence of CYP2D6, CYP2C19 and CYP3A4 polymorphisms on malaria recurrence. Despite this, our findings suggest that the CYP2D6 predicted ultrarapid phenotype was less susceptible to recurrence (p=0.057). Future studies are warranted in order to understand the association of PQ and CYP450 considering factors such as the metabolite concentration, the drug–drug interactions and the presence of some inflammatory conditions, and these may guide the individualized radical treatment of vivax malaria in reference centers that treat patients with multiple recurrences.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
The studies involving human participants were reviewed and approved by Fundacao de Medicina Tropical Dr. Heitor Vieira Dourado. The patients/participants provided their written informed consent to participate in this study.
Conceived and designed the experiments: GM, WM, ML, VS, and JC. Sample processing: JC and YS. Performed the experiments: JC, AA and LB. Data entry and analyses: JC, MR, VS, and AA. Wrote the paper: JC, GM, YS, LB, VS, FR-S, WM, and ML. All authors read and approved the final manuscript.
This research was funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) via the Universal (MCTIC/CNPq 2018), and Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM) (Pró-Estado Program—#002/2008, POSGRAD 2021 Program and PCTI-EMERGESAÚDE Program—#005/2020).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank all the participants who collaborated with this study, including Dr Monica Costa, head of the Malaria Department at FMT-HVD for technical support, CNPq for funding our research and CNPq and FAPEAM for awarding grants. WM and ML are CNPq fellows. We would also like to thank FAPEAM for providing financial support for publication via the PAPAC Call N° 005/2019.
1. World Health Organization. World Malaria Report 2021 (2021). Available at: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2021 (Accessed Jan 27, 2022).
2. White MT, Shirreff G, Karl S, Ghani AC, Mueller I. Variation in Relapse Frequency and the Transmission Potential of Plasmodium Vivax Malaria. Proc R Soc B Biol Sci (2016) 283:1827. doi: 10.1098/rspb.2016.0048
3. Zanghi G, Vaughan AM. Plasmodium Vivax Pre-Erythrocytic Stages and the Latent Hypnozoite. Parasitol Int (2021) 85:102447. doi: 10.1016/j.parint.2021.102447
4. Puaprasert K, Chu C, Saralamba N, Day NPJ, Nosten F, White NJ, et al. Real Time PCR Detection of Common CYP2D6 Genetic Variants and its Application in a Karen Population Study. Malar J (2018) 17:427. doi: 10.1186/s12936-018-2579-8
5. Marcsisin SR, Reichard G, Pybus BS. Primaquine Pharmacology in the Context of CYP 2D6 Pharmacogenomics: Current State of the Art. Pharmacol Ther (2016) 161:1–10. doi: 10.1016/j.pharmthera.2016.03.011
6. Pybus BS, Sousa JC, Jin X, Ferguson JA, Christian RE, Barnhart R, et al. CYP450 Phenotyping and Accurate Mass Identification of Metabolites of the 8-Aminoquinoline, Anti-Malarial Drug Primaquine. Malar J (2012) 11:1–9. doi: 10.1186/1475-2875-11-259
7. World Health Organization. Guidelines for the Treatment of Malaria ([amp]]lrm;2015). Available at: https://apps.who.int/iris/handle/10665/162441 (Accessed August 18, 2021).
8. Bennett JW, Pybus BS, Yadava A, Tosh D, Sousa JC, McCarthy WF, et al. Primaquine Failure and Cytochrome P-450 2D6 in Plasmodium Vivax Malaria. N Engl J Med (2013) 369(14):1381–2. doi: 10.1056/NEJMc1301936
9. Baird JK, Louisa M, Noviyanti R, Ekawati L, Elyazar I, Subekti D, et al. Association of Impaired Cytochrome P450 2d6 Activity Genotype and Phenotype With Therapeutic Efficacy of Primaquine Treatment for Latent Plasmodium Vivax Malaria. JAMA Netw Open (2018) 1(4):e181449. doi: 10.1001/jamanetworkopen.2018.1449
10. Silvino ACR, Costa GL, de Araujo FCF, Ascher DB, Pires DEV, Fontes CJF, et al. Variation in Human Cytochrome P-450 Drug-Metabolism Genes: A Gateway to the Understanding of Plasmodium Vivax Relapses. PloS One (2016) 11(7):e0192534. doi: 10.1371/journal.pone.0192534
11. Brasil LW, Rodrigues-Soares F, Santoro AB, Almeida ACG, Kühn A, Ramasawmy R, et al. CYP2D6 Activity and the Risk of Recurrence of Plasmodium Vivax Malaria in the Brazilian Amazon: A Prospective Cohort Study. Malar J (2018) 17(1):57. doi: 10.1186/s12936-017-2139-7
12. Suarez-Kurtz G. Impact of CYP2D6 Genetic Variation on Radical Cure of Plasmodium Vivax Malaria. Clin Pharmacol Ther (2021) 10(3):595–8. doi: 10.1002/cpt.2313
13. Ariffin NM, Islahudin F, Kumolosasi E, Makmor-Bakry M. Effects of MAO-A and CYP450 on Primaquine Metabolism in Healthy Volunteers. Parasitol Res (2019) 118(3):1011–8. doi: 10.1007/s00436-019-06210-3
14. Elewa H, Wilby KJ. A Review of Pharmacogenetics of Antimalarials and Associated Clinical Implications. Eur J Drug Metab Pharmacokinet (2017) 42(5):745–56. doi: 10.1007/s13318-016-0399-1
15. Thapar MM, Ashton M, Lindegårdh N, Bergqvist Y, Nivelius S, Johansson I, et al. Time-Dependent Pharmacokinetics and Drug Metabolism of Atovaquone Plus Proguanil (Malarone) When Taken as Chemoprophylaxis. Eur J Clin Pharmacol (2002) 58(1):19–27. doi: 10.1007/s00228-002-0426-9
16. Zanger UM, Turpeinen M, Klein K, Schwab M. Functional Pharmacogenetics/Genomics of Human Cytochromes P450 Involved in Drug Biotransformation. Anal Bioanal Chem (2008) 392(6):1093–108. doi: 10.1007/s00216-008-2291-6
17. Pharmacogene Variation Consortium - PharmVar (2021). Available at: https://www.pharmvar.org/ (Accessed August 18, 2021).
18. Lamba JK, Lin YS, Schuetz EG, Thummel KE. Genetic Contribution to Variable Human CYP3A-Mediated Metabolism. Adv Drug Delivery Rev (2002) 54(10):1271–94. doi: 10.1016/s0169-409x(02)00066-2
19. Sortica VA, Lindenau JD, Cunha MG, Ohnishi MD, Ventura AMR, Ribeiro Dos-Santos ÂK, et al. The Effect of SNPs in CYP450 in Chloroquine/Primaquine Plasmodium Vivax Malaria Treatment. Pharmacogenomics (2016) 17:1903–11. doi: 10.2217/pgs-2016-0131
20. Almeida AC, Elias ABR, Marques MP, de Melo GC, da Costa AG, Figueiredo EFG, et al. Impact of Plasmodium Vivax Malaria and Antimalarial Treatment on Cytochrome P450 Activity in Brazilian Patients. Br J Clin Pharmacol (2021) 87(4):1859–68. doi: 10.1111/bcp.14574
21. Friedrich DC, Genro JP, Sortica VA, Suarez-Kurtz G, de Moraes ME, Pena SD, et al. Distribution of CYP2D6 Alleles and Phenotypes in the Brazilian Population. PloS One (2014) 9(10):e110691. doi: 10.1371/journal.pone.0110691
22. Caudle KE, Sangkuhl K, Whirl-Carrillo M, Swen JJ, Haidar CE, Klein TE, et al. Standardizing CYP 2d6 Genotype to Phenotype Translation: Consensus Recommendations From the Clinical Pharmacogenetics Implementation Consortium and Dutch Pharmacogenetics Working Group. Clin Transl Sci (2020) 13(1):116–24. doi: 10.1111/cts.12692
23. Scott SA, Sangkuhl K, Stein CM, Hulot JS, Mega JL, Roden DM, et al. Clinical Pharmacogenetics Implementation Consortium Guidelines for CYP2C19 Genotype and Clopidogrel Therapy: 2013 Update. Clin Pharmacol Ther (2013) 94(3):317–23. doi: 10.1038/clpt.2013.105
24. Boulos M, Amato Neto V, Dutra AP, Di Santi SM, Shiroma M. Análise Da Freqüência De Recaídas De Malária Por Plasmodium Vivax Em Região Não Endêmica. Rev Inst Med Trop Sao Paulo (1991) 33:143–6. doi: 10.1590/S0036-46651991000200009
25. Vítor Silva S, Siqueira AM, Sampaio VS, Guinovart C, Reyes Lecca RC, Melo GC, et al. Declining Malaria Transmission in Rural Amazon: Changing Epidemiology and Challenges to Achieve Elimination. Malar J (2016) 15:266. doi: 10.1186/s12936-016-1326-2
26. Sampaio VS, Siqueira AM, Alecrim M das GC, Mourão MPG, Marchesini PB, Albuquerque BC, et al. Malaria in the State of Amazonas: A Typical Brazilian Tropical Disease Influenced by Waves of Economic Development. Rev Soc Bras Med Trop (2015) 48(Suppl 1):4–11. doi: 10.1590/0037-8682-0275-2014
27. Hicks JK, Sangkuhl K, Swen JJ, Ellingrod VL, Müller DJ, Shimoda K, et al. Clinical Pharmacogenetics Implementation Consortium Guideline (CPIC) for CYP2D6 and CYP2C19 Genotypes and Dosing of Tricyclicantidepressants: 2016 Update. Clin Pharmacol Ther (2017) 102(1):37–44. doi: 10.1002/cpt.597
28. Crews KR, Gaedigk A, Dunnenberger HM, Leeder JS, Klein TE, Caudle KE, et al. Clinical Pharmacogenetics Implementation Consortium Guidelines for Cytochrome P450 2D6 Genotype and Codeine Therapy: 2014 Update. Clin Pharmacol Ther (2014) 95(4):376–82. doi: 10.1038/clpt.2013.254
29. Goetz MP, Sangkuhl K, Guchelaar H-J, Schwab M, Province M, Whirl-Carrillo M, et al. Clinical Pharmacogenetics Implementation Consortium (CPIC) Guideline for CYP2D6 and Tamoxifen Therapy. Clin Pharmacol Ther (2018) 103(5):770–7. doi: 10.1002/cpt.1007
30. Gonçalves BP, Pett H, Tiono AB, Murry D, Sirima SB, Niemi M, et al. Age, Weight, and CYP2D6 Genotype Are Major Determinants of Primaquine Pharmacokinetics in African Children. Antimicrob Agents Chemother (2017) 61(5):e02590–16. doi: 10.1128/AAC.02590-16
31. Chamnanphon M, Gaedigk A, Puangpetch A, Pasomsub E, Chantratita W, Longley RJ, et al. Pharmacogene Variation in Thai Plasmodium Vivax Relapse Patients Treated With a Combination of Primaquine and Chloroquine. Pharmgenomics Pers Med (2020) 13:1–12. doi: 10.2147/PGPM.S201007
32. Chen N, Dowd S, Gatton ML, Auliff A, Edstein MD, Cheng Q. Cytochrome P450 2D6 Profiles and Their Relationship With Outcomes of Primaquine Anti-Relapse Therapy in Australian Defence Force Personnel Deployed to Papua New Guinea and East Timor. Malar J (2019) 18(1):140. doi: 10.1186/s12936-019-2774-2
33. Silvino ACR, Kano FS, Costa MA, Fontes CJF, Soares IS, de Brito CFA, et al. Novel Insights Into Plasmodium Vivax Therapeutic Failure: CYP2D6 Activity and Time of Exposure to Malaria Modulate the Risk of Recurrence. Antimicrob Agents Chemother (2020) 64(5):e02056–19. doi: 10.1128/AAC.02056-19
34. Bank PCD, Caudle KE, Swen JJ, Gammal RS, Whirl-Carrillo M, Klein TE, et al. Comparison of the Guidelines of the Clinical Pharmacogenetics Implementation Consortium and the Dutch Pharmacogenetics Working Group. Clin Pharmacol Ther (2018) 103(4):599–618. doi: 10.1002/cpt.762
35. Pukrittayakamee S, Tarning J, Jittamala P, Charunwatthana P, Lawpoolsri S, Lee SJ, et al. Pharmacokinetic Interactions Between Primaquine and Chloroquine. Antimicrob Agents Chemother (2014) 58(6):3354–9. doi: 10.1128/AAC.02794-13
36. Fasinu PS, Tekwani BL, Avula B, Chaurasiya ND, Dhammika Nanayakkara NP, Wang YH, et al. Pathway-Specific Inhibition of Primaquine Metabolism by Chloroquine/Quinine. Malar J (2016) 15:466. doi: 10.1186/s12936-016-1509-x
37. Almeida ACG, Puça MCB, Figueiredo EFG, Barbosa LR, Salazar YEAR, Silva EL, et al. Influence of CYP2C8, CYP3A4 and CYP3A5 Host Genotypes on Early Recurrence of Plasmodium Vivax. Antimicrob Agents Chemother (2020) 64(7):e02125-19. doi: 10.1128/AAC.02125-19
38. Pett H, Bradley J, Okebe J, Dicko A, Tiono AB, Gonçalves BP, et al. CYP2D6 Polymorphisms and the Safety and Gametocytocidal Activity of Single-Dose Primaquine for Plasmodium Falciparum. Antimicrob Agents Chemother (2019) 63(10):e00538–19. doi: 10.1128/AAC.00538-19
39. Milner EE, Berman J, Caridha D, Dickson SP, Hickman M, Lee PJ, et al. Cytochrome P450 2D-Mediated Metabolism is Not Necessary for Tafenoquine and Primaquine to Eradicate the Erythrocytic Stages of Plasmodium Berghei. Malar J (2016) 15(1):588. doi: 10.1186/s12936-016-1632-8
40. Rede Nacional De Farmacogenética - Refargen (2021). Available at: https://www.refargen.org.br/rubrique.php3?id_rubrique=36&recalcul=oui (Accessed August 18, 2021).
41. Suarez-Kurtz G, Genro JP, de Moraes MO, Ojopi EB, Pena SDJ, Perini JA, et al. Global Pharmacogenomics: Impact of Population Diversity on the Distribution of Polymorphisms in the CYP2C Cluster Among Brazilians. Pharmacogenomics J (2012) 12(3):267–76. doi: 10.1038/tpj.2010.89
42. Klein K, Zanger UM. Pharmacogenomics of Cytochrome P450 3a4: Recent Progress Toward the “Missing Heritability” Problem. Front Genet (2013) 4:12. doi: 10.3389/fgene.2013.00012
43. Shah RR. Pharmacogenetics and Precision Medicine: Is Inflammation a Covert Threat to Effective Genotype-Based Therapy? Ther Adv Drug Saf (2017) 8(9):267–72. doi: 10.1177/2042098617712657
44. Shah RR, Smith RL. Inflammation-Induced Phenoconversion of Polymorphic Drug Metabolizing Enzymes: Hypothesis With Implications for Personalized Medicine. Drug Metab Dispos (2015) 43(3):400–10. doi: 10.1124/dmd.114.061093
45. Waring RH. Cytochrome P450: Genotype to Phenotype. Xenobiotica (2020) 50(1):9–18. doi: 10.1080/00498254.2019.1648911
46. de Andrés F, Sosa-Macías M, Ramos BPL, Naranjo M-EG, LLerena A. CYP450 Genotype/Phenotype Concordance in Mexican Amerindian Indigenous Populations-Where to From Here for Global Precision Medicine? OMICS (2017) 21(9):509–19. doi: 10.1089/omi.2017.0101
47. He Z-X, Chen X-W, Zhou Z-W, Zhou S-F. Impact of Physiological, Pathological and Environmental Factors on the Expression and Activity of Human Cytochrome P450 2D6 and Implications in Precision Medicine. Drug Metab Rev (2015) 47(4):470–519. doi: 10.3109/03602532.2015.1101131
48. Gaedigk A, Dinh JC, Jeong H, Prasad B, Leeder JS. Ten Years’ Experience With the CYP2D6 Activity Score: A Perspective on Future Investigations to Improve Clinical Predictions for Precision Therapeutics. J Pers Med (2018) 8(2):15. doi: 10.3390/jpm8020015
Keywords: Plasmodium vivax, malaria, primaquine, recurrence, CYP450
Citation: Cardoso JLM, Salazar YEAR, Almeida ACG, Barbosa LRA, Silva EL, Rodrigues MGA, Rodrigues-Soares F, Sampaio VS, Siqueira AM, Lacerda MVG, Monteiro WM and Melo GC (2022) Influence of CYP2D6, CYP3A4 and CYP2C19 Genotypes on Recurrence of Plasmodium vivax. Front. Trop. Dis 3:845451. doi: 10.3389/fitd.2022.845451
Received: 29 December 2021; Accepted: 02 February 2022;
Published: 18 March 2022.
Edited by:
Esaki M. Shankar, Central University of Tamil Nadu, IndiaReviewed by:
Natarajan Gopalan, Central University of Tamil Nadu, IndiaCopyright © 2022 Cardoso, Salazar, Almeida, Barbosa, Silva, Rodrigues, Rodrigues-Soares, Sampaio, Siqueira, Lacerda, Monteiro and Melo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gisely C. Melo, Y2FyZG9zb2dpc2VseUBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.