 Belkisyole Alarcón de Noya
Belkisyole Alarcón de Noya Zoraida Díaz-Bello
Zoraida Díaz-Bello Raiza Ruiz-Guevara
Raiza Ruiz-Guevara Oscar Noya
Oscar Noya- 1Instituto de Medicina Tropical, Facultad de Medicina Universidad Central de Venezuela, Caracas, Venezuela
- 2Escuela de Medicina “Luis Razetti”, Facultad de Medicina Universidad Central de Venezuela (UCV), Caracas, Venezuela
- 3Centro para Estudios sobre Malaria, Instituto de Altos Estudios “Dr. Arnoldo Gabaldón”, Ministerio para el Poder Popular de la Salud (MPPS), Caracas, Venezuela
The infection with the hemoflagellate parasite Trypanosoma cruzi originates from America where the wildlife cycle remains to alternate between mammals and hematophagous triatomines. Transmission through contamination of the bite site by vector feces containing highly infectious forms of parasite or direct ingestion of T. cruzi-infected triatomines appear to be the dominant transmission mechanisms. Man joins the transmission when he enters this wild environment or takes the leaves of palms carrying vectors to build houses. Rural Chagas disease develops associated with populations of low economic resources, with infection and reinfection of vector bites since childhood, and the consequent evolution toward chronic cases in adults, when there is little therapeutic benefit to infected people. The progressive migration of people from rural to urban areas and the adaptation of vectors to the peripheries of cities due to displacement caused by deforestation or urbanization that has favored the presence of enzootic cycles with Panstrongylus geniculatus as the most widely distributed species and mammals (synanthropic and domestic) allow vector transmission by ingestion of food contaminated with excrements containing infectious trypomastigotes as the dominant transmission mechanism in the urban environment. Human-to-human transmissions through vertical mother–child infection, transfusions, organ transplants, and the possibility of sexual transmission, transform the epidemiology and the clinical evolution of Chagas disease in the urban environment. Vectors of American trypanosomiasis are no longer restricted to the endemic area, but its presence has been demonstrated in nonendemic areas of the United States, Asia, and other latitudes. The worldwide plague of bedbugs (Cimex lectularius) threatens the possibility of expansion of transmission since they are vectors susceptible to infection, transmission to mammals, trans-stadial penetration, and not being affected by T. cruzi infection at least experimentally. These factors, added to the presence of an unknown number of migrating Latin American asymptomatic carriers together with the presence of triatomines in other continents, have initiated the globalization of a pathology originating in the American continent. Only with an integrative approach, based on new and better tolerated and efficient drugs, vaccines and residual action insecticides, all of them included in an epidemiological surveillance program.
1 Introduction
Chagas disease (ChD), caused by the parasite Trypanosoma cruzi, is endemic in 21 countries in the Americas from the southern region of the United States to the northern part of Argentina. Approximately 6–7 million people are being affected by the disease with an estimated annually death rate of 12,000. People at risk of infection is estimated to be around 75 million due to the high number of people who remained undiagnosed or untreated, combined with the areas with remaining active transmission (1, 2). The houses with mud walls and palm roofs, the sign of Romaña, the heart disease in the elderly, the infected marsupial, the chicks in the chicken coop, and the bites of children by large nocturnal insects, all characterized rural ChD (3). In the Christmas of 2007, the largest outbreak of acute cases of oral transmission of the T. cruzi infection described to date happened in a middle-class neighborhood in the heart of the capital city of Venezuela. It was to be expected since the presence of the entire evolutionary cycle of the T. cruzi parasite had been previously described in Caracas. Since then, oral transmission has been described as the most frequent cause of acute cases of ChD in Venezuela, with four oral outbreaks reported in the capital city and two in cities in the west of the country. The other eight episodes of oral transmission have occurred in small towns (4, 5). Since 1967, numerous outbreaks of acute cases had been reported in Brazil where the main ChD transmission mechanism seems to be oral transmission as well (6). Subsequently, Colombia (7–10), Bolivia (11), and Guyana (12) are added in the list of countries with numerous outbreaks of oral transmission.
ChD has become an emblematic example of the urbanization of a metaxenic infectious disease that is difficult to imagine, since the existence of the infected vector seems essential for the transmission of ChD. In this article, we will review the epidemiological and clinical characteristics of T. cruzi infection in humans from their rural environment to the changing epidemiological characteristics of transmission in Latin American cities, as well as the issue on the export of the infection and the danger of globalization.
2 Rural Chagas Disease
Chagas disease is caused after an infection with the flagellate protozoan T. cruzi which circulates among vector insects of the Triatominae subfamily (Hemiptera: Reduviidae) and some 120 species of mammals, including humans, reflecting its zoonotic nature, representing various complex transmission cycles in domestic and wild environments.
2.1 Wild and Peridomiciliary Cycle
The epidemiology of the transmission of T. cruzi differs in each ecosystem, as each host and vector species have peculiarities (behaviors, immune response, genetic variants, etc.) that led to the adaptations in the ways in which the infection is maintained.
The wild cycle has existed for millions of years and is maintained among mammals with the common opossum (Didelphis marsupialis) (13) and wild triatomines of the genera Rhodnius, Triatoma, and Panstrongylus being the most important vectors. Wild animals become infected not only by ingesting the infected insect but also by contamination of the bite site with parasite containing feces left behind by the infective vectors. In turn, insects become infected by feeding on infected mammals. Didelphis marsupialis is the most important wild reservoir on the continent due to its wide geographical distribution, high rate of natural infection by T. cruzi, and the presence of infective metacyclic trypomastigotes in its perianal glands, the same stages of the parasite present in the rectal ampulla of the vector (14). This stage of the fluids expelled from these glands is highly infectious through subcutaneous, intraperitoneal, oral, and ocular routes (15).
In large areas of South America, transmission was originally limited to the wild cycle in the tropical forest environments, where triatomines fed on small mammals, without human intervention. The presence of T. cruzi does not appear to greatly affect naturally infected triatomines or mammals, suggesting equilibrium between species as a result of long periods of adaptation.
Drought, floods, deforestation, urbanization, and other events facilitate the migration of mammals, resulting in the progressive reduction of food sources for triatomines and extending the radius of the flight in search for food. Adult triatomines attracted to light, heat, or odors can find human dwellings (16). In addition, hungry insects can be passively transported from jungle areas when moving building materials or harvesting and reaching human households. Inside the house, with people and domestic animals, they find the possibility of a blood intake, and therefore the probability of transmission of T. cruzi to people (17).
The natural link between the wild and domiciliary cycles is basically made up of breeding domestic mammals and some wild mammals that occasionally arrive in the human habitat and reside in the peridomicile. Dogs, cats, and other mammals such as goats, sheep, rabbits, pigs, horses, and cattle, are food sources of T. cruzi-infected triatomines, all becoming reservoirs for the parasite. Synanthropic murids, weasels, skunks, ferrets, bats, and man himself can be added when coming in contact with vectors while resting in sheds. Birds such as chickens, although they do not allow the development of T. cruzi, they are triatomine source of food and contribute to the maintenance of the parasitic cycle (18). In certain regions, dogs, opossums, and rodents are probably the most important reservoir hosts in the peridomiciliary cycle (19).
Enzootics in human populations that frequently involve man and his domestic animals constitute zooanthroponotic foci (17) where the peridomicile cycle is a good example (20). The role of humans in the dynamic of transmission is very important by the fact that the life expectancy of a human being is usually over 70 years, and parasitemia can remain for more than 40 years, giving it a particular function as host-reservoir for T. cruzi (21).
The proportion of reservoirs infected with T. cruzi varies according to geographic location, local epidemiology, density of triatomines, and their rate of infection with T. cruzi. The circulation of the parasite in the domestic cycle is dynamic, and the reservoirs become infected at an early age by contact with infected triatomines (22). Triatomines from the wild environment must “adapt” to the domestic environment to colonize and transmit the parasite to the inhabitants of the house. The domestication of triatomines is a complex process, with important genotypic and phenotypic consequences (23). The transition from the wild to the domestic environment is mainly due to the stability of the habitat, with the latter being more stable than the wild due to the ease of continuous reproduction without predation and the permanent source of food (people and domestic animals) (22).
The possibilities of interaction of the elements of the biological cycle among animals and man could constitute the following:
- Wild T. cruzi-infected triatomines infect man or domestic carnivores (dogs and cats) by contamination or by ingestion in the wild environment itself or after invading human dwellings.
- Wild mammals infected by T. cruzi contaminate man when he skins them and uses their blood or raw viscera as food.
- Wild mammals parasitized by T. cruzi penetrate the dwellings or the peridomicile, being a source of infection of domestic triatomines, which later contaminate man and domestic animals.
- Domestic carnivores become contaminated by feeding on wild mammals parasitized by T. cruzi, which they capture from the wild or have invaded the home (24).
- Domestic mammals and man become infected by ingesting feces of infected triatomines or the infected insects themselves contaminating food (22)
- Domestic mammals and man become infected by ingesting food contaminated by the secretions and excretions of the odoriferous glands of marsupials that enter the house contaminating food and beverages (14)
The existence of human ChD is purely accidental. The concentration of parasites is diluted in the environment between vectors and the multitude of reservoirs (25). As man came into contact with natural sources and caused ecological imbalances, he forced the infected triatomines to occupy human dwellings, allowing the latter to find shelter and enough food in domestic animals and humans. Through this, man becomes an active part of the epidemiological chain of American Trypanosomiasis.
Trypanosoma cruzi cycles (wild, peridomiciliary, and domiciliary to which the urban one is added, in areas without triatomines) have a tendency to immortalize themselves; they are not closed, even being isolated from the others. There is an accommodation of the epidemiological chain that seems to be in accordance with the biological determination of parasitosis itself, but to other causes that make it more complex and worsen its endemicity, depending on the ecological, economic, political, and cultural practices of the society (26).
2.2 Epidemiologic and Clinical Characteristics of Chagas Disease by Cutaneous Vector Transmission
In rural areas, contamination of the point of the bite (skin or mucosa) with excrements of the triatomine infected with T. cruzi is the most frequent vector transmission. The vectors Triatoma infestans and Rhodnius prolixus, depending on the geographic location, are mostly responsible for domiciliation in low-income homes with palm roofs or mud walls. Inhabitants are subjected to frequent vector bites and may have knowledge about the existence of the vector in their homes and its association with ChD.
Acute cases are isolated and generally present in children with evidence of entry point such as the Romaña sign (bipalpebral edema and preauricular adenopathy) and the skin inoculation called Chagoma characterized by a painful nodule caused by the bite. These acute cases can occur with all the symptomatic procession of fever, prostration, and myocarditis and are generally left unnoticed, undiagnosed or confused with other pathologies (arbovirus diseases, malaria). The inoculum of an infecting triatomine is lower in this transmission as it refers to the content of metacyclic trypomastigotes in the rectal ampulla of the hematophagus that manages to penetrate through the site of the bite.
The detection of chronic cases of ChD is the frequent situation in rural areas among exposed people who do not know that they are infected. Patients are diagnosed late either fortuitously by some medical control or a serology in blood banks in the case of having a voluntary donor situation or by establishment of symptoms of incipient or advanced heart failure such as cardiomegaly, edema, arrhythmia, and other electrocardiogram alterations.
The serological diagnosis with determination of specific IgG by two diagnostic methods is the guideline to follow since the isolation of the parasite is very difficult at this stage of the disease. Antiparasitic treatment is selective as it depends on the physical conditions of the patients and is not recommended if they are already in the advanced stages of the disease (27).
Control in rural endemic areas depends on the efficiency of the control programs of each country, since it depends on the availability of national resources, distance and access facilities of health institutions and they are highly variable in Latin America.
3 Urban Chagas Disease
Much of the rural exodus has settled on the outskirts of the main cities. People transfer rural living conditions such as crops and infected domestic animals, which favor the maintenance of the periurban cycle. The maintenance of poultry attracts Triatoma maculata, and the association of rodents around the houses coexisting with Rhodnius prolixus are conditions already demonstrated in urban neighborhoods (24, 28–30). The biological cycle has moved from the wild environment to the peridomiciliary and domestic environment, demanding new control strategies such as vaccine production and urban planning to avoid the domiciliation of primary and secondary vectors in urban areas (31). The dilution factor of the parasite plays an important role in the transmission dynamics (32, 33). This concept can be extrapolated to the infection of vectors and reservoirs by T. cruzi (34). In rural areas, the population density of T. cruzi decreases in the environment, either in its various vectors or in the diversity of reservoirs. In contrast, in urban settings, the parasite is concentrated in few reservoirs including man and in vectors, practically restricted to a single species such as Panstrongylus geniculatus, favoring exposure and infection of blood-sucking triatomines.
Chagas disease has other more marked expressions of transmission in cities. Congenital transmission is not exclusive to the urban environment, but it is in the cities where the diagnosis is made. Cases derived from transfusions and transplants of tissues and organs are more appreciated in the cities where the blood banks and the transplant institutions operate. For these reasons, we include these two transmission mechanisms in the urban environment.
3.1 Evidence of the Presence of the Enzootic Cycle in Latin American Cities
The presence of some species of reservoirs and vectors has been noted in modern Latin American cities. Triatoma dimidiata is present in the capitals of Central America and in Guayaquil (Ecuador); Triatoma infestans in the cities of Uruguay; Triatoma rubrofasciata in the cities of Recife, Belem, and Salvador (Brazil); Panstrongylus megistus in El Salvador; P. geniculatus in the metropolitan area of Caracas (16, 35–38), and T. maculata colonizing houses in an urban area of Maracaibo in Venezuela (39). In the cities of Bolivia and southwestern Peru, guinea pigs behave as the main reservoirs when raised as food sources for man, while in Panama and Costa Rica, Rattus rattus seems to be the main domiciliary reservoir (20).
The study on the presence of T. cruzi foci in the Caracas valley and of its reservoirs and potential and/or real vectors dates from the investigations of Quintini (40), who found a morphologically different reduvid to R. prolixus in the gardens of the Central University of Venezuela. Benain Pinto in 1949 (41) recorded an infection rate of 30% for R. prolixus and 100% for P. geniculatus inside homes in the Petare region of Caracas, where he observed the colonization of homes by R. prolixus. By xenodiagnosis, he found 20% of people positive for T. cruzi, doubting the autochthonous origin of the cases, due to the high migratory potential of people from rural areas to the city and the proximity of endemic areas to the Caracas valley.
Pifano in 1986 (42) reported the existence of 39% infection with T. cruzi and 0.5% with Trypanosoma rangeli (a species of trypanosomatid from the continent that shares the same ecological niche as T. cruzi) in P. geniculatus in highly urbanized areas of the Caracas valley. Studies of the intestinal content of these triatomines revealed that there was frequent feeding on humans. Additionally, they found an infection rate of 47% for T. cruzi in D. marsupialis and natural infection in Calluromys phillander (weasel) and Sciurus granatensis (squirrel).
The genus Panstrongylus has shown a great adaptive plasticity, and species considered wild such as P. geniculatus (16, 43), P. megistus (44), and P. rufotuberculatus (45) have shown adaptation to human houses. P. geniculatus, widely distributed in America, is found from southern Mexico to northern Argentina (46). It is of great epidemiological importance since this triatomine has frequently been found to be infected with T. cruzi (16, 43). The invasion of the human houses by P. geniculatus has been documented in Venezuela (47, 48). The existences of enzootic foci of T. cruzi in the Caracas city, in which P. geniculatus and D. marsupialis participate, have already been demonstrated (49, 50). Herrera and Urdaneta-Morales (36) reported a 17% T. cruzi infection in Rattus rattus in the periphery of Caracas with abundant invasion of the heart and moderate tropism towards the skeletal and smooth muscles of the intestine. They found low parasitemias in naturally infected rodents, which contrasted with the high mortality produced in laboratory animals, suggesting the no adaptation and selection of the isolates (14). In 2016, the presence of a zoonotic focus T. cruzi—rodents—P. geniculatus was confirmed in the vicinity of a house where a fruit juice was prepared (37), facilitating its accidental contamination and causing the appearance of acute ChD cases about 6.7 km away in a school of the same city, which gives rise to the largest reported outbreak of oral transmission recorded to date (51).
The study of autochthonous cases of ChD in the city of Rio de Janeiro has been conducted since in 1943 vectors and reservoirs infected with T. cruzi were detected. Although the State of Rio de Janeiro is not classified as an endemic area, occurrence of domestic invasion by Triatoma vitticeps at various locations in the state of RJ has been described (52).
3.2 Human T. cruzi Infection by Oral Vector Transmission
The most striking thing due to the epidemiological change is the oral transmission of infection by T. cruzi. However, there are other types of transmission which have contributed to the urbanization of ChD and are related to human-to-human transmission.
3.2.1 Epidemiological and Clinical Characteristics of Oral Vector-Borne Chagas Disease
Oral transmission of T. cruzi is known from experimental studies carried out in mammals fed with infected vectors, whether these are metaxenic (triatomines) or mechanical (flies) (53). It is difficult to understand that the skin of a “cachicamo” (Dasypus novemcinctus) or the fur of a D. marsupialis or an anteater (Myrmecophaga tridactyla) allows the penetration of the prosbocide of a hematophagus insect. The coexistence of these insects in the caves of mammals, allowing their ingestion, is more feasible (16).
Oral transmission of T. cruzi to man becomes important after the emergence of outbreaks in Brazil in 1967. Since then, records appeareing in other countries and have greatly increased (4, 8, 10, 54). It is very possible that in rural areas, cutaneous and oral vector transmission occurred at the same time since human contact with the vectors is daily due to coexistence in infected houses (55). It is also possible that isolated cases of oral transmission to man have always occurred, which go unnoticed due to our ignorance in medical practice. Hence, recognizing this entity in order to identify both, outbreaks and isolated cases, is of utmost importance. Causal agent T. cruzi can contaminate drinks or food by accidental incorporation of infected vectors or by contamination with urine from marsupials that roam open kitchens, these vehicles can be shared by several people. These factors determine the following more highlights epidemiological and clinical characteristics of the oral transmitted ChD (4):
- Outbreaks occur in groups in families (6), friends, schools (51, 56), and restaurants (9)
- There are no signs of entry of the parasite through the skin (Romaña or Chagoma of inoculation)
- The inoculum of parasites is considerably high since the entire contents of the digestive tract of one or more vectors with very infective metacyclic trypomastigotes are incorporated. The intestine of a T. infestans can harbor 684,000 trypomastigotes (57)
- The group’s symptoms are simultaneous and characterized by fever, prostration, and edema of the face and extremities
- The medical examination detects frequent cardiomegaly due to pericardial effusion. The cases are usually of moderate and severe intensity and often require hospitalization (58, 59)
- Mortality is around 5% (5)
- The occurrence of transmission can occur at a distance from the site where the food was contaminated (60)
- People have total ignorance of the clinical entity with the vectors. This includes patients, doctors, and even health authorities
- The initial diagnosis must be based on serology with determination of IgM and IgG, specifying all persons exposed to the infection, whether or not they are symptomatic. Ideally for all those exposed, but if the group is large, take blood samples to isolate the parasite and determine the parasitic DNA in moderate or severe cases (61)
- Massive treatment of all positives even if they have only one positive test. Benznidazole for 60 days or nifurtimox for 90 days with periodic evaluations of baseline parameters (62)
- Generally, the sanitary workers who handle the outbreak are exposed to interviews by the media, and uniform criteria must be used to avoid confusion and misunderstandings in the messages to the public.
3.3 Transmission of T. cruzi Infection From Human to Human
One of the great pitfalls in the Control of Chagas Disease is due to the multiple transmission mechanisms and that vector transmission, although it is the most important, is not the only modality. There is man-to-man transmission, which is carried out from mother to child or transplacental transmission, transmission by transfusions of blood products and organ transplants, and the possibility of sexual transmission is also being investigated. Laboratory accident due to the mishandling of biological material or infected animals is another transmission mechanism and the reactivation of a latent infection in immunosuppressed patients. Migrations of people from areas where there is vector transmission of T. cruzi to urban centers in Latin America and other countries around the world have modified the epidemiological panorama of ChD, favoring transmission in which the vector is not involved.
3.3.1 Congenital Transmission
Congenital infection with Trypanosoma cruzi is a global problem, occurring on average in 5% of children born from chronically infected mothers in endemic areas, with variations depending on the region, with Bolivia being the country with the highest frequency of congenital ChD infection (63). In countries where vector transmission is controlled, such as Uruguay, the only way to acquire ChD is by congenital transmission (at least until 2015), which has a prevalence of 4% (64). The congenital ChD is a neglected public health problem in Brazil, since there is no systematic search for T. cruzi in pregnant women from endemic areas; the prevalence of ChD in pregnant women ranged from 0.1% to 8.5%, with congenital transmission rates from 0% to 5.2% in Brazil (65). The estimated incidence of congenital ChD is approximately 10,000 cases per year (1). Screening for T. cruzi infection during pregnancy is very important because although antiparasitic treatment is contraindicated at this stage, the doctor needs to be prepared to receive a child with a probable transplacental infection who should be treated immediately. The greatest risk is constituted by those mothers with high parasite loads, since if the pregnancy is not interrupted due to fetal death, the possibility of a child with ChD with severe symptoms is high. The few cases of congenital transmission described in Venezuela in the last decades have been two cases of pregnant women infected orally with T. cruzi (62, 66) in whom parasitic infection results in massive fetal invasion. ChD control programs in endemic countries generally do not include screening for infection by serology in pregnant women, this measure being highly beneficial in the prevention of transplacental infection (67).
3.3.2 Blood Transfusions
Currently, blood transfusion is the second way of acquiring the infection after vector transmission in various regions of America. Between 2006 and 2011, it was found that 2.55%–3.29% of donors in Paraguay had positive serology for ChD (68). The transmission of ChD by transfusion of blood products depends on the donor’s parasitaemia, the recipient’s immune status, and possibly the T. cruzi genotype involved, among other factors (69). T. cruzi can survive in whole blood and blood products for long periods of time (70). Investigating the origin of donors, as well as the active search for individuals with ChD through the detection of antibodies against T. cruzi in the comprehensive evaluation of blood donors are actions in blood banks to rule out the risk of transmission to others persons. Díaz-Bello et al. (71) found that in 254 blood donors sent from different blood banks for presenting anti-T. cruzi antibodies in screening tests in a reference center in Caracas Venezuela, 50.79% had IgG antibodies detected by ELISA-IgG or Indirect Hemaglutination and Complement Fixation Test; the “artificial xenodiagnosis” was positive in 8.5% of the people with positive serology (71). In Chile, screening for T. cruzi is the main source of identification of people with ChD (72). In the United States, there is a legislation for the use of serological tests to reduce the risk of transmission of T. cruzi infection in the blood (Guidance for Industry. US Department of Health and Human Services Food and Drug Administration Center for Biologics Evaluation and Research December 2017; Chagas Biovigilance Network (http://www.aabb.org). Through the active search of specific T. cruzi IgG in blood banks, some autochthonous human cases have been found in the United States (73). In several communities in Spain, the detection of anti-T. cruzi is already a standard for Latin American blood donors (74).
3.3.3 Organ Transplant Transmission
Transmission of ChD by organ transplantation from an infected donor to a recipient without ChD is rare, but it can occur with solid organ transplants such as kidney (0%–18.7%), liver (29%), heart (75%) and even bone marrow (75). More than transmission by transplantation per se, the greatest problem in these patients is the reactivation of latent infection, a condition that must be taken into consideration in all centers where transplants are performed, even in nonendemic countries.
In the case of transmission by organ transplantation, PAHO recommends taking into account the following (75):
a. If the recipient has ChD, it is necessary to consider the use of drugs such as benznidazole or nirurtimox in the event that parasitaemia reappears, regardless of whether there are symptoms or not. It is evident that in these patients, very sensitive diagnostic methods must be used that allow the discovery of T. cruzi trypomastigotes in peripheral blood or the real-time polymerase chain reaction (RT-PCR).
b. If the recipient does not have ChD, but the donor does, consider the possibility of indicating etiological treatment for the donor. It is not appropriate for a donor to be in the acute phase of ChD, or to have chronic cardiomyopathy or intestinal forms, or to donate the heart or any other organ.
c. As with blood transfusions, it is essential to find out if the patient or recipient of transplanted organs comes from areas endemic for ChD and to carry out the appropriate laboratory screening (75).
In transplant patients with fever or other clinical manifestations such as hepatomegaly, splenomegaly, lymphadenopathy, myocarditis, or meningoencephalitis, ChD should be suspected when it comes from endemic areas of Latin America (76).
3.3.4 Sexual Transmission
Human infections produced by T. cruzi could be sexually transmitted, according to several investigations that provide epidemiological, clinical, parasitological, immunological, and nucleic acid data of the parasite (77–79). Experimental studies in laboratory animals revealed the presence of amastigote nests of T. cruzi in the seminiferous tubes, epididymis, and vas deferens, as well as in the uterus, tubes, and ovary of acute infected mice (80). Interestingly, the life cycle of T. cruzi was observed in the ejaculation of one infected mouse. Sexual transmission of parasites was observed from mice infected with T. cruzi to negative receptors (81). Furthermore, amplification of parasite DNA from germ line cells of infected human hosts and their descendants suggests that T. cruzi infections can be transmitted sexually (82). The endemicity of the infection in urban areas of Latin America, and its recognition to some extent in Africa, Asia, Europe, and Oceania, in absence of insect vectors and other modes of infection can be explained by sexual transmission of T. cruzi (83–85). The analysis of four families with acute ChD with fever, headache, malaise, generalized muscle pain, prostration, and occasional signs of heart problems revealed the sexual transmission of T. cruzi infection in humans (78).
Sexual transmission of T. cruzi infection is a potential threat to public health throughout the world, and further research is required on this mode of transmission.
4 The Transition From Rural Trypanosomiasis to Urban Tripanosomiasis
Chagas disease, once thought as a rural problem, has become common in cities (86, 87). The conditioning factors for the mobility of ChD from rural areas to cities are numerous and range from biotic to abiotic factors, which include parasites, human behavior, migrations, land cultivation, periphery human settlements, urbanization, climate change, and others of lesser weight (34). In recent decades, the world’s croplands, grasslands, plantations, and urban areas have grown considerably (88). Anthropogenic changes in the natural habitats of vectors and even hosts may contribute to directional selection pressures on sylvatic populations (89), which in turn may favor variation in terms of morphology and assemblages of species in urban, rural, and jungle environments (90). The modification of rural and urban environments can exert selection pressures on Triatominae populations in domiciliary habitats (91).
Multivariate modeling of Triatominae speciation could be related to a rapid process driven mainly by localized ecological factors (92). Climate change studies document that human behavior contributes significantly to global warming, the fragmentation of natural environments, and the loss of ecosystem processes and ecological resilience. Global warming has contributed to significant changes in insect morphology, as well as in physiological processes, such as development rates (93). There is increasing evidence that habitat disturbance resulting from anthropogenic land-use change can lead to increased vector abundance and, in many cases, increased risk of disease transmission (94, 95). The disturbance of the forest landscape can increase human contact with T. cruzi by increasing the invasion and reinvasion of vectors to homes from jungle foci (96). An extensive meta-analysis of Rhodnius species suggests that forest habitat loss may increase contact between Rhodnius and humans (97).
Infection by T. cruzi is balanced in the jungle environment among very ancient animal species. Progressively, the parasite has moved to other ecosystems with other mammals and vectors getting closer to man, to the point of dispensing with the hematophagous insects giving way to man-to-man transmission.
In Table 1, we compare the evolution of the most frequent vectorial transmission events that take place in rural areas with the most frequent vector transmission that occurs in urban areas in the understanding that these two different ecoepidemiological systems can be interchanged and overlapped.
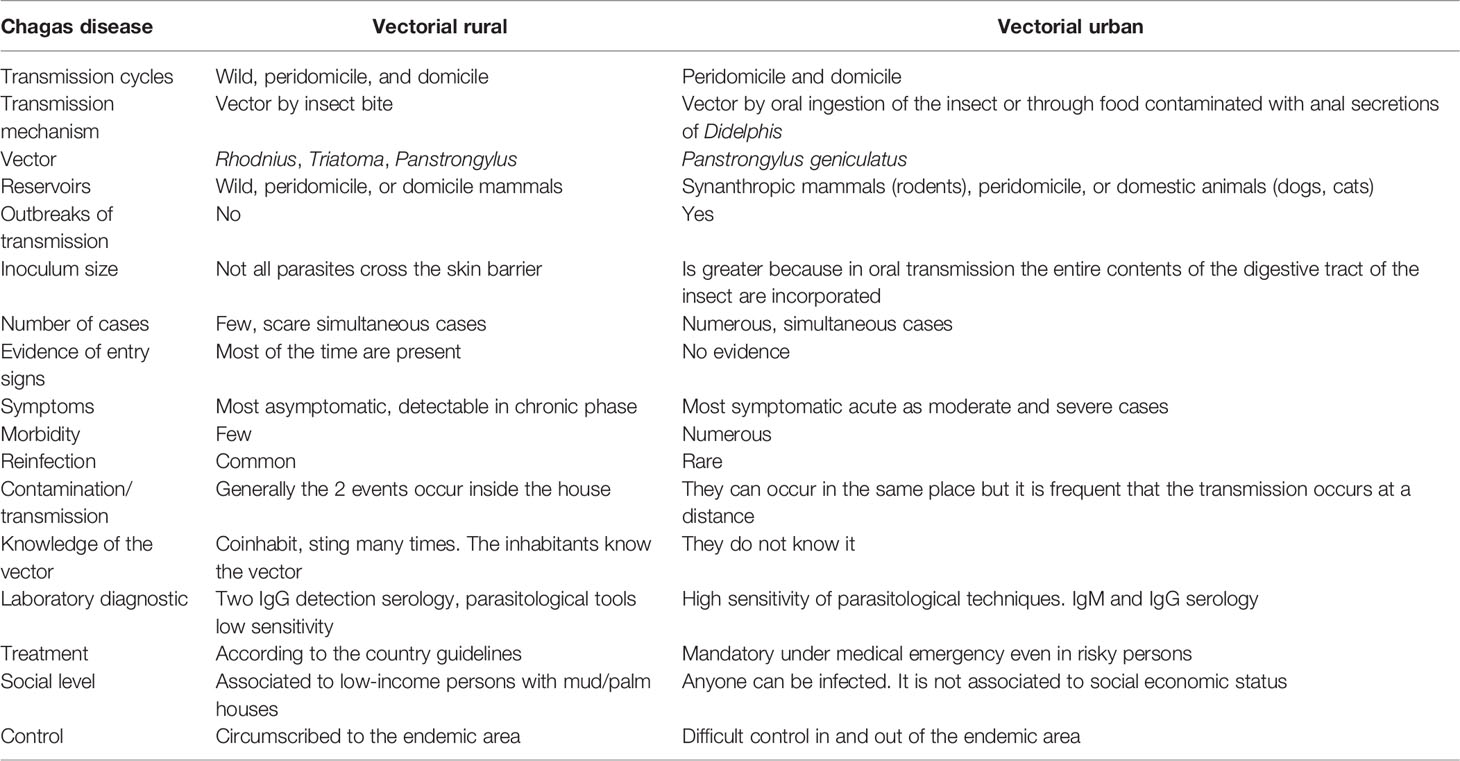
Table 1 Comparison between rural and urban vector-transmitted Chagas disease.
These features are not exclusive to the rural or urban condition.
The biological aspects related to cycles, vectors, reservoirs, and mechanisms of infection, as well as clinical characteristics and ecological conditions that were associated with the progressive transition towards urbanization of ChD are summarized in Figure 1. As this is a multifactorial pathology and includes many areas of knowledge, this figure does not encompass the socioeconomic, cultural, and political determinants that have contributed to the transition of ChD towards urbanization. Written informed consent was obtained from the parents (ON and TG) or legal guardians of the minor for the publication of images or potentially identifiable data included in this article.
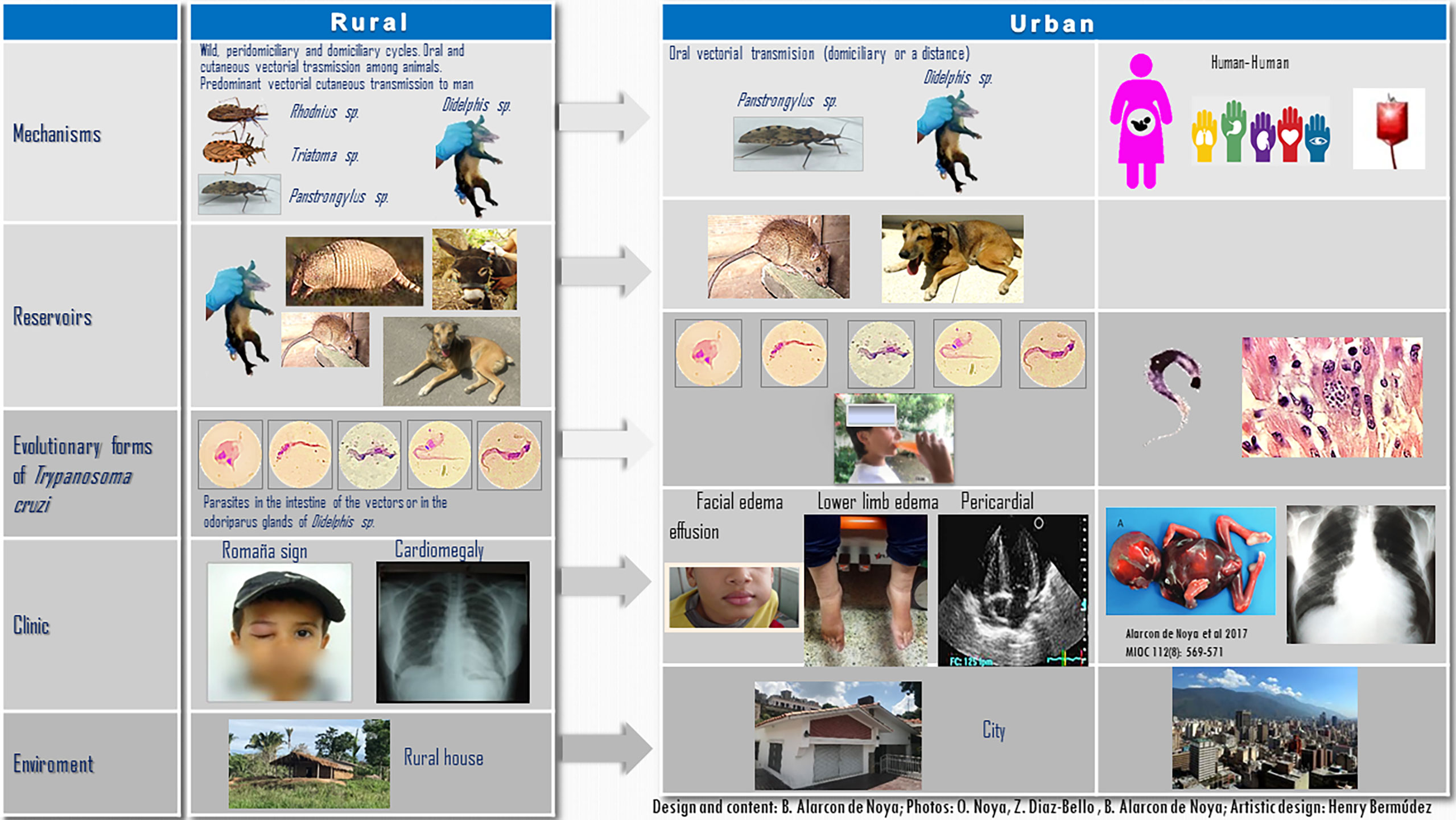
Figure 1 Differences in the ecoepidemiological and clinical patterns of Chagas disease in rural and urban ecosystems. The five evolutionary stages of Trypanosoma cruzi were visualized in a single microscopy field obtained from the intestinal content of the natural infected Panstrongylus geniculatus shown herein. Written informed consent was obtained from minor the legal guardian andnext of kin for the publication of any potentially identifiable images or data included in this article.
5 Exporting Chagas Disease
The estimated number of infected people in Europe is 68,000 to 123,000 (98) and from 200 to 300,000 in the United States (99) while the number is more limited in Japan, Canada, Australia, and New Zealand with a range of 140 in Australia to 12,000 in England (100). In the same proportion are the receiving countries of the great migration of Latin Americans to the world, where Spain stands out as the country that houses the largest number of Latin Americans in Europe (84, 85). Of 2,090,695 Latin Americans living in Spain, it is considered that 47,738 to 67,423 could have ChD (100). Hence, in 2007, the World Health Organization and the Pan American Health Organization convened a meeting entitled “Revisiting Chagas disease: from a Latin American health perspective to a global health perspective,” from which the “Non-endemic Countries Initiative” emerged to highlight the presence of ChD in nonendemic areas of the world (84).
The presence of Latin Americans in the world absorbed by the labor market, not necessarily rural, increases the probabilities of human-to-human transmission outside the American Continent. These infected people can become the source of infection of local hematophagous triatomines with vector potentiality. For health campaigns to recruit infected people in other latitudes, such as those carried out in Spain, and whose objective is to diagnose and administer treatment to serologically positive people, it is important to point out that although medical treatment considerably reduces parasitaemia and the values from serological tests, supervised antiparasitic treatment even in the acute phase in children does not guarantee the cure of this parasitosis (101)
The dispersal of the parasitic seed such as the presence of people infected with the T. cruzi parasite is the ingredient that guarantees the export of ChD and its globalization, from a neglected disease due to its affectation in human populations of low socioeconomic strata to a global public health problem.
Thus, the demonstration of potential vectors transmitting the infection in the two modalities described, the cutaneous vector and the oral vector, the presence and dispersal of reservoirs susceptible to natural infection plus the parasite, envision the emergence of enzootic cycles in nonendemic periurban areas and the Urban Chagas disease.
5.1 Vectors
5.1.1 Evidence of the Presence of Vectors Outside the Traditional Endemic Area
The distribution of ChD has traditionally been circumscribed to the geographic extent of American Trypanosomiasis in America. This parasitic infection originates from the American Continent and its extension depends in turn on the presence of hematophagous vectors infected with T. cruzi of the Rhodnius, Triatoma, and Panstrongylus genera. The vectors spread first to South America, then to Central America and North America, and finally to other continents. Triatoma infestans originating from Bolivia spread from northeastern Brazil to southern Argentina becoming the main vector in South America. Rhodnius prolixus originating from Colombia and Venezuela spread to the Andean region, the Guyana, and Central America (102). In endemic countries, Triatoma infestans is considered to be the main vector domiciled in the countries from the southern cone of America, Rhodnius prolixus and Triatoma dimidiata in the Andean and Central American countries, and Triatoma barberi in Mexico (31).
Natural infection by T. cruzi has been documented in the United States in vectors, wild animals, and in domestic dogs (103–108). Indigenous human cases have been reported in the southern United States (73, 109, 110).
The discrete typing units (DTU) related to the TcII and TcVI group members have been found in insects in California (111). The analysis of 1,510 specimens of triatomines from the southern United States, especially from Texas, yielded these findings: 54.4% T. cruzi infection prevalence of 6 species of triatomines. This prevalence is distributed as follows: T. gerstaeckeri (63.3%; n = 897) and T. lecticularia (66.7%; n = 66) greater than in T. sanguisuga (47.6%; n = 315), T. indictiva (47.8% n = 67), T. rubida (14.1%; n = 64), and T. protracta (10.5%; n = 19). On the other hand, T. gerstaeckeri (n = 548) were more frequently infected with TcI (53.9%) than TcIV (34.4%), and 11.9% showed mixed TcI/TcIV infections. In contrast, T. sanguisuga (n = 135) were more frequently infected with TcIV (79.3%) than TcI (15.6%), and 5.2% showed mixed infections (112). These findings require surveillance in the control of T. cruzi infection based on vector control and frequent monitoring of the triatomine population in other areas of the United States where findings of naturally infected triatomines have been reported (113).
5.1.2 Role of Other Insects as Efficient Vectors of Chagas Disease
The potential role of other vectors susceptible to infection by T. cruzi is a further concern. It has shown that bedbugs, Cimex lectularius, an insect with worldwide distribution, are able to infect experimental animals not only through the skin but also by oral route (114). This important finding could mean that T. cruzi can be transmitted through this biological vector in any of the continents outside the traditionally endemic areas.
Bed bugs (Cimex lectularius) are capable of harboring and transmitting T. cruzi for up to 97 days (115). The pathogen can persist in the insect even through molting processes, between one nymphal stage and the next. This discovery can change the status of bed bugs from a nuisance pest to a public health problem. As ChD is considered a neglected tropical disease and if the parasite that causes it really begins to spread also through bed bugs, it would cause a setback on decades of progress in the control of the disease. The proliferation of these insects is not always associated with lack of hygiene, but rather comes through contamination, feeds on our own blood, and begins to proliferate and spread very quickly, which have recently experienced explosive growth, spreading in rural areas and big cities around the world. Bed bugs share many important traits with triatomine insects. More research is needed to judge whether transmission of bed bug disease to humans can happen in a real-world environment.
5.2 Reservoirs
Since American trypanosomiasis is a widely distributed zooanthroponosis in America, it is essential to know the role played by wild and urban reservoirs in the transmission dynamics of this highly relevant pathology in human public health. As a wild enzootia, this parasitosis is present from latitude 42° N (North Carolina and Maryland, USA) to parallel 49° S (southern areas of Argentina and Chile), including the Caribbean islands, such as Curaçao (25, 116).
Natural infection with T cruzi has been demonstrated in a total of 180 species of mammals belonging to the following orders: Didelphidomorphia, Lagomorpha, Chiroptera, Rodentia, Pilosa, Cingulata, Carnivora, Primata, and Perisodactyla (25). In the amazonian region of Brazil, Jansen etal. (117) examined a total of 6,587 wild mammal specimens during the period 1992–2017. They found that 17% of mammals were seropositive and 8% of all animals displayed positive hemocultures, mainly Philander spp. and Didelphis spp. The coati Nasua nasua, the capuchin monkey Sapajus libidinosus, and the golden lion tamarin Leontopithecus rosalia are the mammals with higher rates of positive hemocultures. The DTU TcII has been found in small rodents from Louisiana in the United States (118). TcI is frequently found in opossums and TcIV in dogs and raccoons (106, 119).
Moving to the peridomiciliary areas, some studies arise, which explore the role of canines as reservoirs in rural areas of the western part of Venezuela, showing a general seropositivity of 67.6% (382/565) to T. cruzi (120).
In an area with important risk of transmission of T. cruzi to humans in western Mexico, the importance of rats as reservoirs of T. cruzi was searched. From 312 Rattus norvegicus examined, 71 (22.7%) were positive for T. cruzi by indirect hemagglutination, which was confirmed by xenodiagnoses. These results and their association with a high prevalence (77.9%) of triatomines (Meccus phyllosomus longipennis) infected with T. cruzi allowed them to conclude that rats seem to constitute an important domiciliary and peridomiciliary reservoir for T. cruzi, furthering the risk of infection for human beings (121).
In the founder focus in Caracas, where the largest oral ChD known outbreak originated, 2 out of three domestic dogs were serologically positive and 4/10 rats captured around the juice-prepared house were parasitologically and molecularly positive to T. cruzi (101).
Undoubtedly, the diversity of the fauna of potential reservoirs in rural and wild areas is much greater than in cities; however, the few species of important reservoirs in urban and periurban areas are of great importance in maintaining transmission. Rats, mice, dogs, cats, and occasionally opossums are the species responsible for maintaining transmission in this ecosystem. In a city as Caracas, with an estimated population of 3.5 million inhabitants (122), there should be at least 875,000 rats, according to very conservative calculations with respect to cities like New York where it is estimated that the rat population is 25% of the human population (123). In 1989, it was calculated that with an estimated population of 3,575,989 inhabitants in Caracas, there should be 286,079 dogs (8% of the human population) of which 14,304 would be strays (124). The presence of high densities of reservoirs susceptible to infection by T. cruzi, (rats and dogs) in cities maintains the danger of urban cycles. As the complete T. cruzi cycle has been demonstrated in Caracas, its natural consequence are oral ChD outbreaks and in fact, four urban outbreaks have occurred in this city (5).
The process of globalization of ChD from America initially began with the migration of patients to other continents, and then it was followed by the dispersion of triatomines to Southeast Asia and Africa (125).
The only thing missing is the dispersal of infected domestic or wild mammals, and in that sense, the majority of pets are dogs and cats. However, migration of wild animals that act as reservoirs for ChD has occurred. This is the case of raccoons, which is an important reservoir in the United States (126). It was introduced in Europe, initially in Germany in 1923 where it is already considered a pest, since they have reached one million specimens. In Spain, raccoon is a plague in the Community of Madrid, but it has spread mostly to the entire country (127). Epidemiological surveillance on this potential host in Europe is a necessary control measure.
6 Final Remarks
Trypanosoma cruzi remains in the wild in many endemic regions of Latin America where the persistence of the parasite is successful among blood-sucking vectors and natural reservoirs. Once the passage to man has been made, the same transmission mechanisms are carried out as among mammals that coexist with vectors, such as cutaneous and oral vector transmission, and congenital transmission, then other forms of transmission such as transfusion, organ transplantation, or accidental mechanisms are introduced to the epidemiological scene as part of the technologies of man. New settlements in the Amazon region associated to agricultural activities especially the açai mass cultivation where the triatomines live, have increased the oral transmission outbreaks in Latin America (128). First at all, migration has occurred from the rural areas to the main Latin American cities as Caracas and Rio de Janeiro where the population size of expansive slums make it difficult to applied sanitary measures (129). In addition, human migration has been moved from the endemic to nonendemic countries. Added to these aspects are the incomplete measures of ChD control looking for cases in infants and young ladies, lack of access to treatment, and toxicity of the old antiparasitic drugs and their limited efficacy even when administered in children, in acute, supervised phases, are all circumstances under which the drugs should be more effective (101). Improving the active search for infected people at different levels (schools, medical certificates, control of pregnant women, military, blood banks, transplantation units, etc.), as well as the medical care for chagasic patients, community participation under the citizen science approach, rats and dogs surveillance, are aspects to be incorporated into the local Programs of Chagas Disease Control. Although traditional vector control and active search for cases have been proven to be insufficient, they are basic control measure for epidemiological surveillance.
Human-to-human transmission can be a public health problem in the transmission of T. cruzi from already infected people to the healthy population. It is probable that these events will gradually decrease, and their penetration in the population would theoretically become less and less. However, since there are vectors and reservoirs susceptible to infection by T cruzi, the only thing missing is the seed of the parasite to close natural transmission cycles in nature in other latitudes different from the American continent. This step is already guaranteed with the growing volume of Latin American migrants to other regions of the planet. All these mentioned factors lead us to the conclusion that once transmission is established in a certain place, the different transmission mechanisms of T. cruzi guarantee the perpetuity of ChD. A discussion is required at various decision levels to increase investment in vaccines and new antiparasitic drugs as the main strategies added to modern vector control. Including the spatial structure of city streets, for example, can more accurately describe the spatial characteristic associations of insect infestations and help to develop effective public health interventions to reduce transmission in densely populated areas (130). Geographic Information Systems coupled with rigorous spatial analysis and remote sensing data (spatially continuous and satellite-derived proxies of environmental factors) cannot only provide information on the factors and mechanisms responsible for heterogeneous contacts between vertebrate hosts and vectors but also help guide science-based disease surveillance, control, and prevention programs (131, 132).
Author Contributions
This manuscript has been written, read and approved by all its authors. There is an agreement between the authors on the order in which they should appear.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors are very grateful to Henry Bermudez, chemist and graphic artist, for his participation in the elaboration of the Figure and to Cecilia Colmenares for helping with the references. Thanks to Dr. Julio Castro Mendez for his personal support. Special thanks also to the patients who agreed to share their photos for scientific purposes and to whom we owe all of our learning.
References
1. WHO. Chagas Disease in Latin America: An Epidemiological Update Based on 2010 Estimates. World Health Organ Weekly Epidemiol Record) (2015) 90(6):33–44.
2. WHO. American Trypanosoniasis (2020). Available at: https://www.who.int/news-room/fact-sheets/detail/chagas-disease.
3. Alarcón de Noya B, Jackson Y. Chagas Disease Epidemiology: From Latin America to the World. In: Pinazo Delgado MJ, Gascón J, editors. Chagas Disease. Cham: Springer (2020). doi: 10.1007/978-3-030-44054-1_2.
4. Alarcón de Noya B, Díaz-Bello Z, Colmenares C, Ruiz-Guevara R, Mauriello L, Muñoz-Calderón A, et al. Update on Oral Chagas Disease Outbreaks in Venezuela: Epidemiological, Clinical and Diagnostic Approaches. Mem Inst Oswaldo Cruz (2015) 110(3):377–86. doi: 10.1590/0074-02760140285
5. Ruiz-Guevara R, Muñoz-Calderón A, Alarcón de Noya B, Redondo C, Beitia Y, Pulido L, et al. Brote Familiar De Enfermedad De Chagas Por Transmisión Oral En Yaguapita, Estado Miranda, Venezuela. Bol Mal Salud Amb LVII (2017) 2:1–16.
6. Santos EF, Silva ÂAO, Leony LM, Freitas NEM, Daltro RT, Regis-Silva CG, et al. Acute Chagas Disease in Brazil From 2001 to 2018: A Nationwide Spatiotemporal Analysis. PloS Negl Trop Dis (2020) 14(8):e0008445. doi: 10.1371/journal.pntd.0008445
7. Cáceres D, Nicholls RS, Corredor A, Gualdrón L, Slait E, Dib J, et al. Investigación De Un Brote De Síndrome Febril Con Miocarditis Aguda En Guamal, Magdalena, 7 a 11 De Junio De 1999. Inf Quinc Epidemiol Nac (1999) 4:170–8.
8. Hernández C, Vera MJ, Cucunubá Z, Flórez C, Cantillo O, Buitrago LS, et al. High-Resolution Molecular Typing of Trypanosoma Cruzi in 2 Large Outbreaks of Acute Chagas Disease in Colombia. J Infect Dis (2016) 214:1252–5. doi: 10.1093/infdis/jiw360
9. Zuleta-Dueñas LP, López-Quiroga AJ, Torres-Torres F, Castañeda-Porras O. Posible Transmisión Oral De La Enfermedad De Chagas En Trabajadores Del Sector De Los Hidrocarburos En Casanare, Colombia 2014. Biomédica (2017) 37(2):218–32. doi: 10.7705/biomedica.v37i3.3153
10. Rincon-Acevedo CY, Parada-Garcia AS, Olivera MJ, Torres-Torres F, Zuleta-Dueñas LP, Hernandez C, et al. Clinical and Epidemiological Characterization of Acute Chagas Disease in Casanare, Eastern Colombi–2020. Front Med (2021) 8:681635. doi: 10.3389/fmed.2021.681635
11. Santalla-Vargas J, Oporto Carrasco P, Espinoza E, Rios T, Brutus L. Primer Brote Reportado De La Enfermedad De Chagas En La Amazonía Boliviana: Reporte De 14 Casos Agudos Por Transmisión Oral De Trypanosoma cruzi En Guayaramerín, Beni-Bolivia. BIOFARBO (2011) 19(1):52–8.
12. Blanchet D, Brenière SF, Schijman AG, Bisio M, Simon S, Véron V, et al. First Report of a Family Outbreak of Chagas Disease in French Guiana and Posttreatment Follow-Up. Infect Genet Evol (2014) 28:245–50. doi: 10.1016/j.meegid.2014.10.004
13. Coura JR, Dias JC. Epidemiology, Control and Surveillance of Chagas Disease: 100 Years After its Discovery. Mem Inst Oswaldo Cruz 104 (2009) 1):31–40. doi: 10.1590/s0074-02762009000900006
14. Deane MP, Lenzi HL, Jansen A. Trypanosoma Cruzi: Vertebrate and Invertebrate Cycles in the Same Mammal Host, the Opossum Didelphis Marsupialis. Mem Inst Oswaldo Cruz (1984) 79(4):513–5. doi: 10.1590/s0074-02761984000400021
15. Urdaneta-Morales S, Nironi I. Trypanosoma cruzi in the Anal Glands of Urban Opossums. Isolation and Experimental Infections. Mem Inst Oswaldo Cruz (1996) 91:399–403. doi: 10.1590/S0074-02761996000400002
16. Reyes-Lugo M. Panstrongylus Geniculatus Latreille 1811 (Hemiptera: Reduviidae: Triatominae), Vector De La Enfermedad De Chagas En El Ambiente Domiciliario Del Centro-Norte De Venezuela. Rev Biomédica (2009) 20(3):180–205. doi: 10.32776/revbiomed.v20i3.136
17. Barretto MP. Epidemiologia. In: Brener Z, Andrade Z, editors. Trypanosoma Cruzi E Doença De Chagas. Guanabara-Koogan, Brasil (1979). p. 89 –151.
18. Carpintero DJ. El Problema De La Infección Chagásica En Las Grandes Urbes. Consultor Salud 26 Agosto Buenos Aires N° (1994) 99:10.
19. WHO. Informe De Un Comité De Expertos. Control De La Enfermedad De Chagas. Serie De Informes Técnicos 811. Ginebra (1991). p. 24.
20. Zeledón R. Epidemiology, Modes of Transmission and Reservoir Hosts of Chagas' Disease. In: n: Trypanosomiasis and Leishmaniasis With Special Reference to Chagas Disease, Ciba Foundation Symposium 20 (New Series). Pitman, London: Elsevier Publishing Co (1974). p. 51–77.
21. Pinto Dias JC. Epidemiologia. In: Brener Z, Andrade Z, Barral-Netto M, editors. Trypanosoma Cruzi E Doença De Chagas. Rio de Janeiro: Guanabara-Koogan (2000). p. 48–74.
22. WHO. World Health Organization Expert Committee on the Control of Chagas Disease Brasilia, Brazil) & World Health Organization. In: Control of Chagas Disease: Second Report of the WHO Expert Committee. Geneva: World Health Organization (2002). Available at: https://apps.who.int/iris/handle/10665/42443.
23. Schofield CJ, Diotaiuti L, Dujardin JP. The Process of Domestication in Triatominae. Mem Inst Oswaldo Cruz (1999) 94(Suppl 1):375–8. doi: 10.1590/s0074-02761999000700073
24. Herrera L. Trypanosoma Cruzi, the Causal Agent of Chagas Disease: Boundaries Between Wild and Domestic Cycles in Venezuela. Front Public Health (2014) 2:259. doi: 10.3389/fpubh.2014.00259
25. Herrera L. Una Revisión Sobre Reservorios De Trypanosoma (Schizotrypanum) Cruzi (Chagas 1909), Agente Etiológico De La Enfermedad De Chagas. Bol Mal Salud Amb (2010) 50(1):3-15.
26. Briceño-León R. La Enfermedad De Chagas En Las Américas: UnaPerspectiva De Ecosalud. Cad Saúde Pública Rio Janeiro (2009) 25(1):S71–82. doi: 10.1590/S0102-311X2009001300007
27. OPS/OMS. Guía Para El Diagnóstico Y Tratamiento De La Enfermedad De Chagas. Washington: Organizacion Panamericana de la Salud (2018). p. 160. Available at: http://iris.paho.org.
28. Garcia-Alzate R, Lozano-Arias D, Reyes-Lugo RM, Morocoima A, Herrera L, Mendoza-Leon A. Triatoma Maculata, the Vector of Trypanosoma Cruzi in Venezuela. Phenotypic and Genotypic Variability as Potential Indicator of Vector Displacement Into the Domestic Habitat. Front Pub Health (2014) 2:1–9. doi: 10.3389/fpubh.2014.00170
29. Arias-Giraldo LM, Muñoz M, Hernández C, Herrera G, Velásquez-Ortiz N, Cantillo-Barraza O, et al. Identification of Blood-Feeding Sources in Panstrongylus, Psammolestes, Rhodnius and Triatoma Using Amplicon-Based Next-Generation Sequencing. Parasites Vectors (2020) 13:434. doi: 10.1186/s13071-020-04310-z
30. Hernández C, Salazar C, Brochero H, Teheran A, Buitrago LS, Vera M, et al. Untangling the Transmission Dynamics of Primary and Secondary Vectors of Trypanosoma Cruzi in Colombia: Parasite Infection, Feeding Sources and Discrete Typing Units. Parasites Vectors (2016) 9:620. doi: 10.1186/s13071-016-1907-5
32. Keesing F, Belden L, Daszak P, Dobson A, Harvell CD, Holt RD, et al. Impacts of Biodiversity on the Emergence and Transmission of Infectious Diseases. Nature (2010) 468:647–52. doi: 10.1038/nature09575
33. Schmidt K, Ostfeld R. Biodiversity and the Dilution Effect in Disease Ecology. Ecology (2001) 82(3):609–19. doi: 10.1890/0012-9658(2001)082[0609:BATDEI]2.0.CO;2
34. Alarcón de Noya B, Noya O. An Ecological Overview on the Factors That Drives to Trypanosoma Cruzi Oral Transmission. Acta Tropica (2015) 151:94–102. doi: 10.1016/j.actatropica.2015.06.004
35. Herrera L, Urdaneta-Morales S. Didelphis Marsupialis: A Primary Reservoir of Trypanosoma Cruzi in Urban Areas of Caracas, Venezuela. Ann Trop Med Parasitol (1992) 86(6):607–12. doi: 10.1080/00034983.1992.11812716
36. Herrera L, Urdaneta-Morales S. Synantropic Rodent Reservoirs of Trypanosoma (Schizotrypanum) Cruzi in the Valley of Caracas, Venezuela. Rev Inst MedTrop Sao Paulo (1997) 39:279–82. doi: 10.1590/S0036-46651997000500006
37. Díaz-Bello Z, Zavala-Jaspe R, Reyes-Lugo M, Colmenares C, Noya-Alarcón O, Noya O, et al. Urban Trypanosoma Cruzi Oral Transmission From a Zoonotic Founder Focus to the Largest Microepidemic of Chagas Disease. SOJ Microbiol Infect Dis (2016) 4(1):1–9. doi: 10.15226/sojmid/4/1/00147
38. Carrasco HJ, Torrellas A, García C, Segovia M, Feliciangeli MD. Risk of Trypanosoma Cruzi I (Kinetoplastida: Trypanosomatidae) Transmission by Panstrongylus Geniculatus (Hemiptera: Reduviidae) in Caracas (Metropolitan District) and Neighboring States, Venezuela. Int J Parasitol (2005) 35(13):1379–84. doi: 10.1016/j.ijpara.2005.05.003
39. Torres RA. Sobre Un Foco Urbano De Trypanosoma Cruzi (Chagas 1909) En La Ciudad De Maracaibo Vol. 10. . Venezuela: Kasmera (1982) p. 57–1. Available at: https://produccioncientificaluz.org/index.php/kasmera/article/view/4455.
40. Quintini J. Nota Sobre Un Nuevo Conorrhinus Capturado En Caracas. Gac Med Caracas (1920) 27:171–2.
41. Benaim Pinto H. Resultados Preliminares De Una Encuesta De Reconocimiento Sobre Enfermedad De Chagas Y Tripanosomiasis Rangeli En Un Sector Del Valle De Caracas (Petare, Edo. Miranda). Rev Polic Caracas (1949) 18:103–17.
42. Pifano F. El Potencial Enzoótico Silvestre Del Complejo Ecológico Schizotrypanum Cruzi (T. cruzi)-Didelphis marsupiales-Panstrongylus geniculatus Y Sus Incursiones a La Vivienda Humana Del Valle De Caracas, Venezuela. Bol Acad Cienc Fis Mat Nat (1986) XLVI:143–4.
43. Wolff M, Castillo D. Evidencias De Domesticación Y Aspectos Biológicos De Panstrogylus geniculatus (Letreill) (Hemiptera: Rediviidae). Acta Entomol Chil (2000) 24:77–83.
44. Forattini OP, Rocha-e-Silva EO, Rabello EX, Andrade JC, Rodríguez VL. Ecological Aspects of South American Trypanosomiasis. XIII. Domestic Enzootic Potential in an Area of Ocurrence of Panstrongylus Megistus, Under Epidemiological Surveillance. Rev Saúde Pública (1978) 2:417–24. doi: 10.1590/S0034-89101978000400002
45. Noireau F, Vargas F, Bosseno MF, Brenniére SF. Apparent Trend to Domesticity Observed in Panstrongylus Rufotuberculatus (Hemiptera: Reduviidae) in Bolivia. Res Rev Parasitol (1994) 54:249–50.
46. Patterson JS, Barbosa SE, Feliciangeli DM. On the Genus Panstrongylus Berg 1879: Evolution, Ecology and Epidemiological Significance. Acta Trop (2009) 110:187–99. doi: 10.1016/j.actatropica.2008.09.008
47. Reyes-Lugo M, Rodriguez-Acosta A. Domiciliation of the Sylvatic Chagas Disease Vector Panstrongylus Geniculatus Latreill(Triatominae: Reduviidae) in Venezuela. Trans R Soc Trop Med Hyg (2000) 94(5):508. doi: 10.1016/s0035-9203(00)90068-3
48. Feliciangeli MD, Carrasco H, Patterson J, Suárez B, Martínez C, Medina M. Mixed Domestic Infestation by Rhodnius Prolixus Stal 1859 and Panstrongylus Geniculatus Latreille 1811, Vector Incrimination, and Seroprevalence for Trypanosoma Cruzi Among Inhabitants in El Guamito, Lara State, Venezuela. Am J Trop Med Hyg (2004) 71:501–5. doi: 10.4269/ajtmh.2004.71.501
49. Sampson-Ward L, Urdaneta-Morales S. Urban Trypanosoma cruzi: Biological Characterization of Isolates From Panstrongylus geniculatus. Ann SocBelg Med Trop (1988) 68:95–106.
50. Scorza C, Urdaneta-Morales S, Sampson-Ward L. Urban Trypanosoma (Schizotrypanum) cruzi: Pathology in White Mice of Isolates From Panstrongylus geniculatus. Ann Soc Belg Med Trop (1989) 69:283–9.
51. Alarcón de Noya B, Díaz-Bello Z, Colmenares C, Ruiz-Guevara R, Mauriello L, Zavala-Jaspe R, et al. Large Urban Outbreak of Orally-Acquired Acute Chagas Disease, at a School in Caracas, Venezuela. J Infect Dis (2010) 201(9):1308–15. doi: 10.1086/651608
52. Conde Sangenis LH, De Sousa AS, Sperandio Da Silva GM, Xavier SS, Machado CR, Brasil P, et al. First Report of Acute’Chagas Disease by Vector Transmission in Rio De Janeiro State, Brazil. Revista do Instituto De Medicina Tropical De Sao Paulo. (2015) 57: (4):361–4. doi: 10.1590/S0036-46652015000400017
53. Diaz-Ungría C. La Mosca Doméstica Como Transmisora Del Trypanosoma cruzi. Rev La Universidad Del Zulia (1969) 44:114–9.
54. Nery-Guimarães F, Silva NN, Calusell DT, Mello AL, Rapone T, Snell T, et al. Um Surto Epidêmico De Doença De Chagas De Provável Transmissão Digestiva Ocorrido Em Teutônia (Estrela, Rio Grande do Sul). Hospital (1968) 73:1767–804.
55. Briceño-León R. La Casa Enferma: Sociología De La Enfermedad De Chagas. Fondo Editorial Acta Científica De Venezuela Y Consorcio De Ediciones Capriles C. A. Caracas, Venezuela: Consorcio de Ediciones Capriles (1990). p. 149.
56. Alarcón de Noya B, Colmenares C, Díaz-Bello Z, Ruiz-Guevara R, Medina K, Muñoz-Calderón A, et al. Orally-Transmitted Chagas Disease: Epidemiological, Clinical, Serological and Molecular Outcomes of a School Microepidemic in Chichiriviche De La Costa, Venezuela. Parasite Epidemiol Control (2016) 1(2):188–98. doi: 10.1016/j.parepi.2016.02.005
57. Shaub GA. Trypanosoma Cruzi: Quantitative Studies of Development of Two Strains in Small Intestine and Rectum of the Vector Triatoma Infestans. Exp Parasitol (1989) 68:260–73. doi: 10.1016/0014-4894(89)90108-2
58. Alarcón de Noya B, Veas J, Ruiz-Guevara R, Martín A, Rojas C, Machado I, et al. Evaluación Clínica Y De Laboratorio De Pacientes Hospitalizados Durante El Primer Brote Urbano De Enfermedad De Chagas De Transmisión Oral En Venezuela. Rev Patol Trop (2013) 42:177–86. doi: 10.5216/rpt.v42i2.25523
59. Alarcón de Noya B, Noya-González O. Clinical Aspects in Foodborne Chagas Disease. In: Alarcón de Noya B, Noya-González O, Robertson LJ, editors. Trypanosoma Cruzi as a Foodborne Pathogen, 1st edition, vol. 2015a . New York, USA: Springer (2015). p. 33–40.
60. Xavier SCC, Roque ALR, Bilac D, Araújo VAL, Neto SFC, Lorosa ES, et al. Distantiae Transmission of Trypanosoma Cruzi: A New Epidemiological Feature of Acute Chagas Disease in Brazil. PloS Negl Trop Dis (2014) 8(5):e2878. doi: 10.1371/journal.pntd.0002878
61. Alarcón de Noya B, Díaz-Bello Z, Colmenares C, Zavala-Jaspe R, Abate T, Contreras R, et al. The Performance of Laboratory Tests in the Management of a Large Outbreak of Orally Transmitted Chagas Disease. Mem Inst Oswaldo Cruz (2012) 107(7):893–8. doi: 10.1590/S0074-02762012000700009
62. Alarcón de Noya B, Ruiz-Guevara R, Noya O, Castro J, Ossenkopp J, Díaz-Bello Z, et al. Long-Term Comparative Pharmacovigilance of Orally Transmitted Chagas Disease: First Report. Expert Rev Anti-Infective Ther (2017) 15(3):319–25. doi: 10.1080/14787210.2017.1286979
63. Carlier Y, Sosa-Estani S, Luquetti AO, Buekens P. Congenital Chagas Disease: An Update. Mem Inst Oswaldo Cruz (2015) 110(3):363–8. doi: 10.1590/0074-02760140405
64. Barrios P, Más M, Giachetto G, Basjmadjián Y, Rodríguez M, Viera A, et al. Enfermedad De Chagas: Transmisión Vertical. Descripción De Casos Clínicos. Rev Méd Uruguay (2015) 31(3):209–13.
65. Martins-Melo FR, Ramos AN, Alencar CH, Heukelbach J. Prevalence of Chagas Disease in Brazil: A Systematic Review and Meta-Analysis. Acta Trop (2014) 130:167–74. doi: 10.1016/j.actatropica.2013.10.002
66. Suárez J, de Suárez CB, Alarcón de Noya B, Espinosa R, Chiurillo MA, Villaroel A, et al. Enfermedad De Chagas Sistémica En Fase Aguda Por Transmisión Oral: Diagnóstico Integral De Un Caso Autopsiado. Gac Med Caracas (2010) 118(3):212–22.
67. Sosa-Estani S, Cura E, Velázquez E, Yampotis C, Segura E. Etiological Treatment of Young Women Infected With Trypanosoma Cruzi and Prevention of Congenital Transmission. Rev Soc Bras Med Trop (2009) 42:484–7. doi: 10.1590/S0037-86822009000500002
68. Márquez-Roa NM, Zelada ML, Molas A. Frecuencia Serológica De Infección Por Trypanosoma cruzi En Donantes De Sangre En El Paraguay Entre Los Años 2006 Y 2011. Memorias Del Instituto Investigaciones En Cienc La Salud (2013) 11(2):26-31.
69. Schmunis GA. Prevention of Transfusional Trypanosoma cruzi Infection in Latin America. Mem Inst Oswaldo Cruz (1999) 94(I):93–101. doi: 10.1590/S0074-02761999000700010
70. Wendel S, Pinto Días JC. Transfusion Trasmited Chagas Disease. In: Wendel S, Brener Z, Camargo ME, editors. Chagas Disease (American Tripanosomiasis): Its Impact on Transfusion and Clinical Medicine. Sao Paulo: International Society of Blood Transfusion (1992). p. 103–33.
71. Diaz-Bello Z, Zavala-Jaspe R, Diaz-Villalobos M, Mauriello L, Maekelt A, Alarcon de Noya B. Diagnóstico Confirmatorio De Anticuerpos Anti-Trypanosoma cruzi En Donantes Referidos Por Bancos De Sangre En Venezuela Investigacion Clinica. Invest Clin (2008) 49: (2):141–50.
72. Apt WB, Heitmann I, Jercic MI, Jofré L, Muñoz P, Noermí I, et al. Guías Clínicas De La Enfermedad De Chagas: Parte II. Enfermedad De Chagas En El Adulto, La Infancia Y Adolescencia. Rev Chil Infectol (2008) 25(3):194–9. doi: 10.4067/S0716-10182008000300009
73. Cantey PT, Stramer SL, Townsend RL, Kamel H, Ofafa K, Todd CW, et al. The United States Trypanosoma Cruzi Infection Study: Evidence for Vector-Borne Transmission of the Parasite That Causes Chagas Disease Among United States Blood Donors. Transfusion (2012) 52(9):1922–30. doi: 10.1111/j.1537-2995.2012.03581.x
74. Benjamin RJ, Stramer SL, Leiby DA, Dodd RY, Fearon M Castro E. Trypanosoma Cruzi Infection in North America and Spain: Evidence in Support of Transfusion Transmission. Transfusion (2012) 52:1913–21. doi: 10.1111/j.1537-2995.2011.03554.x
75. PAHO. Oficina Regional Para Las Américas De La Organización Mundial De La Salud. © Organización Panamericana De La Salud). In: Enfermedad De Chagas Transmitida Por Trasplante De Órganos. Decálogo Para Prevenir La Transmisión. Washinton DC: EEUU. Organizacion Panamericana de la Salud. (2021). Available at: www.paho.org/es/documentos/enfermedad-chagas-transmitida-por-trasplante-organos-decalogo-para-prevenir-transmision.
76. Yuste JR, Poz JL, Quetglás EG, Azanza JR. Infecciones Más Comunes En El Paciente Trasplantado. Anales Del Sistema Sanitario Navarra (2006) 29(Supl. 2):175–205. doi: 10.4321/S1137-66272006000400016
77. Gomes C, Almeida AB, Rosa AC, Araujo PF, Teixeira A. American Trypanosomiasis and Chagas Disease: Sexual Transmission. Int J Infect Dis (2019) 81:81–4. doi: 10.1016/j.ijid.2019.01.021
78. Araujo PF, Almeida AB, Pimentel CF, Silva AR, Sousa A, Valente SA, et al. Sexual Transmission of American Trypanosomiasis in Humans: A New Potential Pandemic Route for Chagas Parasites. Mem Inst Oswaldo Cruz (2017) 112(6):437–46. doi: 10.1590/0074-02760160538
79. Herrera L, Urdaneta-Morales S. Experimental Transmission of Trypanosoma Cruzi Through the Genitalia of Albino Mice. Mem Inst Oswaldo Cruz (2001) 96:713–7. doi: 10.1590/S0074-02762001000500024
80. Carvalho LO, Abreu-Silva AL, Hardoim D, Tedesco RC, Mendes VG, Costa D, et al. Trypanosoma Cruzi and Myoid Cells From Seminiferous Tubules: Interaction and Relation With Fibrous Components of Extracellular Matrix in Experimental Chagas' Disease. Int J Exp Pathol (2009) 90(1):52–7. doi: 10.1111/j.1365-2613.2008.00592.x
81. Alencar AA, Brito CMM, Azevedo BA. Studies on the Infectivity of Semen From Trypanosoma cruzi-Infected Mice With Destruction of Neuronal Structures. Rev Bras Neurol (1991) 27:51–6.
82. Hecht M, Nitz N, Araujo P, Sousa A, Cássia A, Gomes DA, et al. Inheritance of DNA Transferred From American Trypanosomes to Human Hosts. PloS One (2010) 5:e9181. doi: 10.1371/journal.pone.0009181
83. Schmunis GA, Yadon ZE. Chagas Disease: A Latin American Health Problem Becoming a World Health Problem. Acta Trop (2010) 115(1-2):14–21. doi: 10.1016/j.actatropica.2009.11.003
84. Albajar-Viñas P, Jannin J. The Hidden Chagas Disease Burden in Europe. Euro Surveill (2011) 16(38):pii=19975. doi: 10.2807/ese.16.38.19975-en
85. Pérez-Molina JA, Norman F, López-Vélez R. Chagas Disease in non-Endemic Countries: Epidemiology, Clinical Presentation and Treatment. Curr Infect Dis Rep (2012) 14(3):263–74. doi: 10.1007/s11908-012-0259-3
86. LaDeau SL, Allan BF, Leisnham PT, Levy MZ. The Ecological Foundations of Transmission Potential and Vector-Borne Disease in Urban Landscapes. Funct Ecol (2015) 29(7):889–901. doi: 10.1111/1365-2435.12487
87. Delgado S, Ernst KC, Pumahuanca MLH, Yool SR, Comrie AC, Sterling CR, et al. A Country Bug in the City: Urban Infestation by the Chagas Disease Vector Triatoma Infestans in Arequipa, Peru. Int J Health Geographics (2013) 12(1):1. doi: 10.1186/1476-072X-12-48
88. Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, et al. Global Consequences of Land Use. Science (2005) 309:570–4. doi: 10.1126/science.1111772
89. Chevin LM, Lande R, Mace GM. Adaptation, Plasticity, and Extinction in a Changing Environment: Towards a Predictive Theory. PloS Biol (2010) 8:e1000357. doi: 10.1371/journal.pbio.1000357
90. Blair RB, Launer AE. Butterfly Diversity and Human Land Use: Species Assemblages Along an Urban Gradient. Biol Conserv (1997) 80:113–25. doi: 10.1016/S0006-3207(96)00056-0
91. Carmona-Galindo V, Marín Recinos M, Gámez Hidalgo S, Recinos Paredes G, Posada Vaquerano E, Romero-Magaña A, et al. Morphological Variability and Ecological Characterization of the Chagas Disease Vector Triatoma Dimidiate (Hemiptera: Reduviidae) in El Salvador. Acta Trop (2020) 205:105392. doi: 10.1016/j.actatropica.2020.105392
92. Dujardin JP, Panzera P, Schofield CJ. Triatominae as a Model of Morphological Plasticity Under Ecological Pressure. Mem Inst Oswaldo Cruz (1999) 94:223–8. doi: 10.1590/S0074-02761999000700036
93. Robinet C, Roque A. Direct Impacts of Recent Climate Warming on Insects. Integr Zool (2010) 5:132–42. doi: 10.1111/j.1749-4877.2010.00196.x
94. Yasuoka J, Levins R. Impact of Deforestation and Agricultural Development on Anopheline Ecology and Malaria Epidemiology. Am J Trop Med Hyg (2007) 76:450–60. doi: 10.4269/ajtmh.2007.76.450
95. Vora N. Impact of Anthropogenic Environmental Alterations on Vector-Borne Diseases. Medscape J Med (2008) 10:238.
96. Calzada JE, Pineda V, Montalvo E, Alvarez D, Santamaria AM, Samudio F, et al. Human Trypanosome Infection and the Presence of Intradomicile Rhodnius Pallescens in the Western Border of the Panama Canal, Panama. Am J Trop Med Hyg (2006) 74:762–5. doi: 10.4269/ajtmh.2006.74.762
97. Abad-Franch F, Monteiro FA, Jaramillo ON, Gurgel-Goncalves R, Dias FB, Diotaiuti L. Ecology, Evolution, and the Long Term Surveillance of Vector-Borne Chagas Disease: A Multi-Scale Appraisal of the Tribe Rhodniini Triatominae). Acta Trop (2009) 110:159–77. doi: 10.1016/j.actatropica.2008.06.005
98. Basile L, Jansa JM, Carlier Y, Salamanca DD, Angheben A, Bartoloni A, et al. Chagas Disease in European Countries: The Challenge of a Surveillance System. Euro Surveill (2011) 16(37):pii=19968. doi: 10.2807/ese.16.37.19968-en
99. Manne-Goehler J, Umeh CA, Montgomery SP, Wirtz VJ. Estimating the Burden of Chagas Disease in the United States. PloS Negl Trop Dis (2016) 10(11):e0005033. doi: 10.1371/journal.pntd.0005033
100. Pinazo MJ, Gascon J. The Importance of the Multidisciplinary Approach to Deal With the New Epidemiological Scenario of Chagas Disease (Global Health). Acta Tropica (2015) 151:16–20. doi: 10.1016/j.actatropica.2015.06.013
101. Díaz-Bello Z, Alarcón de Noya B, Muñoz-Calderón A, Ruiz-Guevara R, Mauriello L, Colmenares C, et al. Ten-Year Follow-Up of the Largest Oral Chagas Disease Outbreak. Lab Biomarkers Infect Indic Ther Failure Acta Trop (2021) 222:106034. doi: 10.1016/j.actatropica.2021.106034
102. Coura JR. The Main Sceneries of Chagas Disease Transmission. The Vectors, Blood and Oral Transmissions a Comprehensive Review. Mem Inst Oswaldo Cruz (2015) 110(3):277–82. doi: 10.1590/0074-0276140362
103. Brown EL, Roellig DM, Gompper ME, Monello RJ, Wenning KM, Gabriel MW, et al. Seroprevalence of Trypanosoma Cruzi Among Eleven Potential Reservoir Species From Six States Across the Southern United States. Vector Borne Zoonotic Dis (Larchmont NY) (2010) 10(8):757–63. doi: 10.1089/vbz.2009.0009
104. Burkholder JE, Allison TC, Kelly VP. Trypanosoma Cruzi (Chagas) in Invertebrate Reservoir and Human Host of the Lower Rio Grande Valley of Texas. J Parasitol (1980) 66:305–11. doi: 10.2307/3280824
105. Curtis-Robles R, Wozniak EJ, Auckland LD, Hamer GL, Hamer SA. Combining Public Health Education and Disease Ecology Research: Using Citizen Science to Assess Chagas Disease Entomological Risk in Texas. PloS Neglected Trop Dis (2015) 9(12):e0004235. doi: 10.1371/journal.pntd.0004235
106. Curtis-Robles R, Lewis BC, Hamer SA. High Trypanosoma Cruzi Infection Prevalence Associated With Minimal Cardiac Pathology Among Wild Carnivores in Central Texas. Int J Parasitol Parasites Wild (2016) 5(2):117–23. doi: 10.1016/j.ijppaw.2016.04.001
107. Kjos SA, Snowden KF, Craig TM, Lewis B, Ronald N, Olson JK. Distribution and Characterization of Canine Chagas Disease in Texas. Vet Parasitol (2008) 152(3-4):249–56. doi: 10.1016/j.vetpar.2007.12.021
108. Kjos SA, Snowden KF, Olson JK. Biogeography and Trypanosoma Cruzi Infection Prevalence of Chagas Disease Vectors in Texas, USA. Vector Borne Zoonotic Dis (2009) 9(1):41–50. doi: 10.1089/vbz.2008.0026
109. Dorn PL, Perniciaro L, Yabsley MJ, Roellig DM, Balsamo G, Diaz J, et al. Autochthonous Transmission of Trypanosoma Cruzi, Louisiana. Emerg Infect Dis (2007) 13(4):605–7. doi: 10.3201/eid1304.061002
110. Garcia MN, Aguilar D, Gorchakov R, Rossmann SN, Montgomery SP, Rivera H, et al. Evidence of Autochthonous Chagas Disease in Southeastern Texas. Am J Trop Med Hyg (2015) 92(2):325–30. doi: 10.4269/ajtmh.14-0238
111. Hwang WS, Zhang G, Maslov D, Weirauch C. Infection Rates of Triatoma Protracta (Uhler) With Trypanosoma Cruzi in Southern California and Molecular Identification of Trypanosomes. Am J Trop Med Hyg (2010) 83(5):1020–2. doi: 10.4269/ajtmh.10-0167
112. Curtis-Robles R, Auckland L, Snowden K, Hamer G, Hamer S. Analysis of Over 1500 Triatomine Vectors From Across the US, Predominantly Texas, for Trypanosoma Cruzi Infection and Discrete Typing Units. Infect Genet Evol (2018) 58:171–80. doi: 10.1016/j.meegid.2017.12.016
113. Schofield C, Grijalva M, Diotaluti L. Distribución De Los Vectores De La Enfermedad De Chagas En Países No Endémicos: La Posibilidad De Transmisión Vectorial Fuera De América Latina. Enf Emerg (2009) 11(Supl 1):20–7.
114. Salazar R, Castillo-Neyra R, Tustin AW, Borrini-Mayorí K, Náquira C, Levy MZ. Bed Bugs (Cimex Lectularius) as Vectors of Trypanosoma Cruzi. Am J Trop Med Hyg (2015) 92(2):331–5. doi: 10.4269/ajtmh.14-0483
115. Peterson JK, Salazar R, Castillo-Neyra R, Borrini-Mayori K, Condori C, Bartow-McKenney C, et al. Trypanosoma Cruzi Infection Does Not Decrease Survival or Reproduction of the Common Bed Bug, Cimex Lectularius. Am J Trop Med Hyg (2018) 98(3):724–34. doi: 10.4269/ajtmh.17-0593
116. Dias JCP. Epidemiologia. In: Brener Z, Andrade Z, Barral-Netto M, editors. Trypanosoma Cruzi E Doença De Chagas. Rio de Janeiro, Brazil: Guanabara Koogan Rio de Janeiro (2000). p. 48–74.
117. Jansen AM, Xavier S, Roque A. Trypanosoma Cruzi Transmission in the Wild and its Most Important Reservoir Hosts in Brazil. Parasites Vectors (2018) 11(1):502. doi: 10.1186/s13071-018-3067-2
118. Herrera CP, Licon MH, Nation CS, Jameson SB, Wesson DM. Genotype Diversity of Trypanosoma Cruzi in Small Rodents and Triatoma Sanguisuga From a Rural Area in New Orleans, Louisiana. Parasites Vectors (2015) 8:123. doi: 10.1186/s13071-015-0730-8
119. Roellig DM, Brown EL, Barnabé C, Tibayrenc M, Steurer FJ, Yabsley MJ. Molecular Typing ofIsolates, United States. Emerg Infect Dis (2008) 14(7):1123–5. doi: 10.3201/eid1407.080175
120. Crisante G, Rojas A, Teixeira MM, Añez N. Infected Dogs as a Risk Factor in the Transmission of Human Trypanosoma Cruzi Infection in Western Venezuela. Acta Trop (2006) 98:247–54. doi: 10.1016/j.actatropica.2006.05.006
121. Rosal GG, Nogueda-Torres B, Villagrán ME, de Diego-Cabrera JA, Montañez-Valdez OD, Martínez-Ibarra JA. Chagas Disease: Importance of Rats as Reservoir Hosts of Trypanosoma Cruzi (Chaga) in Western Mexico. J Infect Public Health (2018) 11(2):230–3. doi: 10.1016/j.jiph.2017.07.017
122. CAF. Banco De Desarrollo De America Latina. Archivo De Datos De Las Ciudades De La Red. Observatorio De Movilidad Urbana. Tabla Especial Sobre El Area Metropolitana Caracas Y Sus Municipalidades (2010) p. 116.
123. Auerbach J. Does New York City Really Have as Many Rats as People? Significance (2014) 11(4):22-7. doi: 10.1111/j.1740-9713.2014.00764.x
124. Pifano F, Orihuela AR, Delgado O, Cortez R, Abdul Hadi S, Dale de Ortíz M, et al. La Toxocariasis Humana En Venezuela, Especialmente En El Valle De Caracas. Gac Méd Caracas (1989) 97: 31-41.
125. Dujardin JP, Lam TX, Khoa PT, Schofield CJ. The Rising Importance of Triatoma Rubrofasciata. Mem Inst Oswaldo Cruz (2015) 110(3):319–23. doi: 10.1590/0074-02760140446
126. Majeau A, Pronovost H, Sanford A, Cloherty AE, Anderson N, Balsamo G, et al. Raccoons as an Important Reservoir for Trypanosoma Cruzi: A Prevalence Study From Two Metropolitan Areas in Louisiana. Vector-Borne Zoonotic Dis (2020) 20(7):1–6. doi: 10.1089/vbz.2019.2559
127. Beltrán-Beck B, García FJ, Christian Gortázar C. Raccoons in Europe: Disease Hazards Due to the Establishment of an Invasive Species. Eur J Wildl Res (2012) 58:5–15. doi: 10.1007/s10344-011-0600-4
128. Alarcón de Noya B, Noya-González O, Robertson LJ. Future Challenges and Final Remarks. In: Alarcón de Noya B, Noya-González O, Robertson LJ, editors. Trypanosoma Cruzi as a Foodborne Pathogen, 1st edition. New York, USA: Springer (2015). p. 89–92.
129. Ferreira A. Favelas No Rio De Janeiro: Nascimento, Expansao, Remoao E, Agora, Exclusao a Través De Muros (2009). Available at: http://www.ub.es/geocrit/b3w-828.
130. Weaver SC. Urbanization and Geographic Expansion of Zoonotic Arboviral Diseases: Mechanisms and Potential Strategies for Prevention. Trends Microbiol (2013) 21(8):360–3. doi: 10.1016/j.tim.2013.03.003
131. Kitron U. Landscape Ecology and Epidemiology of Vector-Borne Diseases: Tools for Spatial Analysis. J Med Entomol (1998) 35:435– 445. doi: 10.1093/jmedent/35.4.435
Keywords: Chagas disease, vector, reservoirs, transmission, rural, urban
Citation: Alarcón de Noya B, Díaz-Bello Z, Ruiz-Guevara R and Noya O (2022) Chagas Disease Expands Its Epidemiological Frontiers From Rural to Urban Areas. Front. Trop. Dis 3:799009. doi: 10.3389/fitd.2022.799009
Received: 20 October 2021; Accepted: 09 February 2022;
Published: 25 April 2022.
Edited by:
Jorge Enrique Gómez Marín, University of Quindío, ColombiaReviewed by:
Diana Carolina Hernández Castro, Rosario University, ColombiaGabriel Parra-Henao, Instituto Nacional de Salud, Colombia
Copyright © 2022 Alarcón de Noya, Díaz-Bello, Ruiz-Guevara and Noya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Belkisyole Alarcón de Noya, YmVsa2lzdW9sZUBnbWFpbC5jb20=