
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Trop. Dis., 08 November 2021
Sec. Neglected Tropical Diseases
Volume 2 - 2021 | https://doi.org/10.3389/fitd.2021.778724
This article is part of the Research TopicNovel Therapeutics for Parasitic NTDsView all 5 articles
 Samuel Wanji1,2*
Samuel Wanji1,2* Valerine Chawa Chunda1,2Fanny Fri Fombad2,3
Valerine Chawa Chunda1,2Fanny Fri Fombad2,3 Abdel Jélil Njouendou2,4Narcisse Victor T. Gandjui1,2
Abdel Jélil Njouendou2,4Narcisse Victor T. Gandjui1,2 Manuel Ritter5Peter A. Enyong1,2Charles Mackenzie6
Manuel Ritter5Peter A. Enyong1,2Charles Mackenzie6 Mark J. Taylor7Achim Hoerauf5,8
Mark J. Taylor7Achim Hoerauf5,8 Joseph D. Turner7*
Joseph D. Turner7*The tropical disease, loiasis, caused by the filarial parasite, Loa loa, has gained prominence in global public health as a cause of excess mortality and a barrier to the elimination of the related prioritized neglected tropical diseases (NTDs), lymphatic filariasis and onchocerciasis, within Central Africa. There are no effective drug cures or vaccines available to treat loiasis safely. Here we review recent advances in loiasis preclinical platform technologies, including novel in vitro culturing systems, animal models and innovations in experimental infections of the L. loa vector, Chrysops, that have facilitated access to all L. loa filarial life-cycle stages. We detail applications of these new model systems in anti-filarial drug screening, diagnostic development, immunology, and pathophysiology research. Finally, we provide an overview of how loiasis preclinical platforms may be further utilized in translational medicine applications to support the development of much needed new interventions against filarial NTDs.
Loiasis is a tropical parasitic infection caused by the filarial nematode, Loa loa, endemic in forested areas of Central and West Africa. Infective larvae (L3) are transmitted by diurnal bites of tabanid deer flies of the genus, Chrysops. L3 infect subcutaneous tissues and undergo morphogenesis via two cuticle moults to develop into immature adults, at approximately 50 days post-infection (1). From five months post-infection, male and female adults mature and sexually reproduce, releasing first-stage microscopic larvae, microfilariae (mf), which migrate into the bloodstream (2). Among individuals infected with L. loa, clinically distinct sub-groups can be identified with manifestations ranging from asymptomatic microfilaremia to a hyper-responsive state characterized by more frequent episodes of local angioedema (Calabar swellings) and pronounced eosinophilia in the absence of mf (3). Despite these symptoms, and that over 14 million people currently reside in high-risk areas, loiasis has often been mis-classified as a benign disease (2). Contrary to other filarial infections such as onchocerciasis and lymphatic filariasis (LF), loiasis has not yet been prioritized in the World Health Organization’s list of neglected tropical diseases (NTDs) (4). However, chronic infections can lead to renal, cardiac, pulmonary and neurological pathologies and loaisis has recently been identified as a cause of excess death, calling for re-appraisal of its status as a medically important filarial disease (5). Of additional public health relevance, L. loa hyper-microfilaraemias are a significant risk factor for the development of severe adverse events (SAEs) in individuals treated with ivermectin during mass drug administration (MDA) filariasis elimination campaigns in sub-Saharan Africa (6–9). A microfilarial load >30,000 mf/ml is a significant risk factor for SAE development and recent modelling predicts incidence of SAEs will occur in 10% of individuals with parasitaemias of 50,000 mf/ml (10). Non-neurological inflammatory AEs can develop with increased frequency below this level, which still have the potential to incapacitate individuals for several days.
Ivermectin-induced acute pathology presents initially as headache, fever and/or haemorrhaging of the palpebral conjunctiva (11) which can progress to more pronounced neurological manifestations, encephalopathy, coma and death. Whilst the aetiology of this pathology was, until recently, ill-defined due to a paucity of tractable animal models, it has been speculated that drug-mediated paralysis and entrapment of mf in brain tissue capillary beds initiates blockage, haemorrhage (as indicated by ocular haemorrhage) and ultimately breakdown of the blood brain barrier (3). In support of this, L. loa mf have been evidenced in cerebral-spinal fluid of individuals suffering SAEs (10). A host inflammatory component is associated with adverse events and may exacerbate neurological pathophysiology. Following microfilaricidal treatment, elevation of allergic mediators such as interleukin-5 and eosinophil degranulation proteins are apparent in the circulation of loiasis patients (12, 13). A distinct mechanism hypothesised by Geary and colleagues suggests that rare mutations in CNS drug efflux pumps (multi-drug resistance 1 gene; mdr-1) may result in an ivermectin toxicity syndrome (caused by activation of, for instance GABA neuroreceptors present in mammalian CNS), as is apparent in veterinary filariasis treatment (14).
The clinical association between loiasis microfilaraemia and ivermectin SAEs was established more than 20 years ago. For elimination of onchocerciasis, the subsequent impact has been to avoid ivermectin MDA in loiasis co-endemic health districts where levels of Onchocerca volvulus are hypo-endemic (less than 20% prevalence). MDA campaigns have ensued in meso- and hyper-endemic onchocerciasis regions but with increased surveillance of post-treatment SAE and protocols for clinical care established in the local health system. Whilst a ‘test-and-not-treat’ strategy based on a novel point-of-care L. loa diagnostic is one potential strategy to tackle elimination of hypo-endemic onchocerciasis in co-endemic regions (15), the cost estimate per test (between $4-8 USD) may be prohibitive to rollout in annual MDA campaigns (16). Further, social science investigations have identified the perceived risk of loiasis adverse events (whether neurological or non-neurological yet interfering with economic activities) as a major factor in persistent non-participation in ivermectin MDA where onchocerciasis remains meso- to hyper-endemic (17). Because ivermectin MDA must be delivered annually, at a coverage of 80%, for periods of 15 years or more to prevent onchocerciasis transmission, elimination may not be feasible with the current strategy in loiasis-endemic Central African foci. Recent modelling suggests onchocerciasis will persist beyond 2045 in L. loa co-endemic countries with annual ivermectin MDA (18). Similarly, L. loa endemicity would represent a major barrier to future rollout of ivermectin MDA as an endectocide for vector control of malaria (19).
A distinct complication of L. loa co-endemicity, disrupting the elimination of LF in Central Africa, is the occurrence of cross-reactivity to current point-of-care rapid diagnostic tests (RDTs) used in mapping of LF endemic regions and decision making to stop MDA (20, 21).
For these reasons, development of new tools to address the treatment of filarial disease in Central Africa are urgently required. Small scale trials have indicated that various extended or intermittent dose regimens of the human anthelmintic, albendazole, may be either partially macrofilaricidal and/or mediate disruption of mf production in loiasis patients (22, 23). An ideal novel short-course therapeutic product profile would include a high degree of selective efficacy against adult stage L. loa and/or O. volvulus (ideally both), without inducing the ivermectin-like acute microfilaricidal activities which put hyper-microfilaraemic loiasis individuals at risk of SAE. Identification of affordable adjunctive treatments which limit inflammatory adverse events following ivermectin treatment may also improve attitudes to community participation in current MDA. A scalable serological point-of-care rapid diagnostic test for determining L. loa clinical status may also become a pre-requisite as part of a test-and-treat algorithm for filariasis elimination in Central Africa.
Research and development of new therapeutics and diagnostics for loiasis have been hampered by a long-standing lack of investment and the inertia created by a paucity of tractable tools to enable facile preclinical research to be undertaken. Recently, renewed investment in filariasis selective macrofilaricide development has been initiated by funders such as The Bill and Melinda Gate Foundation. This has concomitantly spurred research to improve and innovate L. loa preclinical platform technologies. In this review, we focus on these most recent advances and applications in loiasis pathophysiology, drug and diagnostic translational medicine.
In order to manipulate L. loa in vitro accurately (e.g. to examine drug responses) it is first necessary to define culture conditions which support parasite longevity outside of the body. Optimum culture conditions enabling the successful long-term maintenance of L. loa mf or developing larvae has been recently elucidated. Standard mammalian culture media, such as Dulbecco’s Modified Eagles Medium (DMEM), supplemented with calf serum (5-15%) is sufficient to maintain L. loa mf purified from the venous blood of hyper-microfilaraemic non-human primates (NHP) for periods of between 6-12 days (24) with a fully motile phenotype. Similarly, calf serum supplementation supports full motility of infectious stage L3 larvae in mammalian culture for up to eight days after isolation from the mouthparts of Chrysops flies. Completion of the L3-4 moulting process occurs in approximately 20% in these basic cultures. Addition of an immortalised monkey kidney epithelial cell monolayer to serum-supplemented cultures (LLC-MK2 cell line) extends full motility of mf to 22 days. LLC-MK2 co-cultures also increases L3/L4 motility for periods as long as 17 days and concomitantly boosts the moulting success rate to as much as 60%.
The L. loa NHP model is one of the best-studied animal models of human filarial infections (25). Indeed, a sylvatic L. loa life cycle is evident in monkey species in forested Central Africa (26) indicating the cross-species adaptation of this filaria. Much of the biology of L. loa has been elucidated via parasitological experiments using NHP. The human strain L. loa can be experimentally transmitted to Mandrillus leucophaeus (drill), M. sphinx (mandrill), Papio anubis (baboon), Erythrocebus patas (patas monkey) and Macaca mulatta (rhesus macaque) (26–29). Chrysops naturally infected with L. loa have been collected by baited sweep traps in high transmission areas or, alternatively, human landing catch on loiasis individuals to provide a source of infectious stage L3 larvae. Experimental subcutaneous inoculations of 600 L3 are typically utilised to infect NHPs (30–33). The pre-patent interval after experimental infection of susceptible monkeys is about 150 days, irrespective of the species of experimental host involved. In most monkey species, once the infection has become patent, the microfilarial densities increase sharply, reach a peak and then fall within several weeks to very low levels, which then persist throughout the infection (30). The spleen plays a major role in the clearance of mf from the peripheral blood (30–33) Splenectomised monkeys develop very high L. loa microfilaraemias which can persist for many months (27, 30). Although drills and monkeys are excellent laboratory hosts for L. loa, ethically, drills are under strict restrictions according to the convention on international trade in endangered species (CITES) classification of primates. As such, this species is no longer used for biomedical research. As an alternative, the baboon offers potential to be used as an experimental NHP model for L. loa as the parasite behaves in this primate in essentially the same way as it does in the drill (28). The pre-patent interval after experimental infection of splenectomised baboons is 5 months, with microfilaremias accruing for periods up to 18 months (32). Importantly for the study of pathophysiology of microfilaricidal adverse reactions, hyper-microfilaremia (>30,000 mf/mL) can be achieved in the majority (approximately 70%) of infected splenectomised baboons and all infected animals develop eosinophilia significantly exceeding the normal range (32).
Whilst closely emulating the life cycle of human loiasis, throughput of the baboon NHP model is severely constraining for anti-filarial drug research and to identify potential targets for adjunct therapies to limit ivermectin adverse reactions. Availability of L. loa susceptible laboratory rodent models, particularly mice, would be a step-change improvement, both because they are a convenient, standardised model with tractable genetic and immunological tools available but also as a less sentient animal substitute to reduce or replace usage of NHP. L. loa does not undergo full development in laboratory ‘wild-type’ immunocompetent mice. L. loa infective larvae (inoculations of between 50-200 L3s) administered sub-cutaneously survive only for a week in BALB/c mice, an inbred laboratory strain that is conversely permissive or semi-permissive to serous cavity infections with related filariae, Litomosoides sigmodontis and Brugia spp. respectively (34–36). When BALB/c mice are immuno-suppressed with hydrocortisone, L. loa survival is extended for up to 3 weeks (37, 38). Control of infection in BALB/c mice is associated with a ‘type-2’ cellular immune response of splenocytes when re-stimulated with L. loa L3 antigen, notably with elevated production of interleukins-4, -9 and -13 (39). Confirming a role for both IL-4/IL-13 signalling and IL-5 in the early adaptive immune control of L. loa in mice, BALB/c IL-4 receptor and IL-5 combination deficient mice are susceptible to pre-patent adult L. loa infections (40). (CCR)-3 knockout mice, deficient in recruitment of eosinophils via eotaxins and other eosinophil chemokines, demonstrate extended survival of L. loa developing larvae, linking type-2 immune responses with tissue eosinophil recruitment as a mediator of early immunity to loiasis (41). Since the minimum pre-patent period prior to the release of mf in blood is 5 months, Pionnier et al, investigated the long-term parasitological success of L. loa infection in a panel of ‘severe-combined’ lymphopenic immunodeficient mice lacking all adaptive immunity and facets of innate immune responses (42). Moderate levels of pre-patent adult L. loa infection were evident in CB.17 SCID mice (a BALB/c congenic background strain) at 3 months post-infection, meanwhile, Non-Obese Diabetic (NOD) SCID mice and BALB/c RAG2-/- mice had cleared the infection at the same time point. Fecund adult L. loa infections in the natural parasitic niche were reproducibly evident at 5 months in compound immunodeficient, lymphopenic mouse strains: NOD SCID γc−/− and BALB/c RAG2−/−γc−/−, which lack both lymphocytes and the common gamma chain (γc) cytokine signalling pathway. At this time point, in both compound immunodeficient mouse strains, most worms were found in the natural tissue niches of L. loa adult stages, namely the subcutaneous and muscle fascia tissues, while some adults were recovered from cardiopulmonary tissues as well as the pleural and peritoneal cavities. Parasitism of L. loa within these organs suggests that larvae might have migrated via the lymphatics through the thoracic duct, corroborating a theory that filariae have a unified lymphatic larval phase (43). By Implantation of a defined number of male and female L. loa adults from these strains under the skin of either BALB/c RAG2-/- or RAG2-/-γc-/- recipients, it was possible to establish microfilaraemic mice one-month post-implant with high rates of adult worm survival retained in subcutaneous tissues (42).
Due to the long pre-patent period following L3 infection, a more facile approach of infusing purified L. loa mf derived from baboon NHP directly into venous blood of mice was evaluated (42). This approach was based on the success of establishing long-term microfilaraemias in a variety of inbred mouse strains and genetic knockouts using the human LF parasite, B. malayi (44, 45). After infusion with inoculates of 4x104 purified mf, persistent microfilaraemias could be established in either BALB/c or CB.17 SCID mice over a period of at least eight days. The majority of mf were sequestered in cardiopulmonary circulation (approximately 10% of the inoculate) with a scant peripheral microfilaraemia evident. Both peripheral and cardio-pulmonary microfilaraemias were improved slightly with splenectomy in BALB/c mice although non-splenectomised SCID mice supported the highest parasitaemias, indicative of adaptive immunity regulating density of L. loa microfilaraemia. Increasing the L. loa inoculate to 1x105 mf per mouse lead to significantly increased cardiopulmonary microfilaraemias in both SCID and BALB/c mice toward a hyper-microfilaraemic density associated with risk of SAE in humans (Figure 1A). The bias of L. loa mf accumulation in the cardiopulmonary circulation may reflect the anatomical differences between murine and human microvasculature. In addition, because in humans L. loa exhibits a diurnal periodicity (46), physiological cues for peripheral circulatory migration versus cardiopulmonary sequestration may vary between mice and humans. Human sub-periodic B. malayi also demonstrate a tropism for cardiopulmonary circulation when infused into mice (47, 48). In follow on experiments, mf purified from venous blood samples from human volunteers was found to be comparable to those isolated from baboons and was used as a more abundant and ethical source to avoid NHP usage in onward applications of the mouse models (Figure 1B).
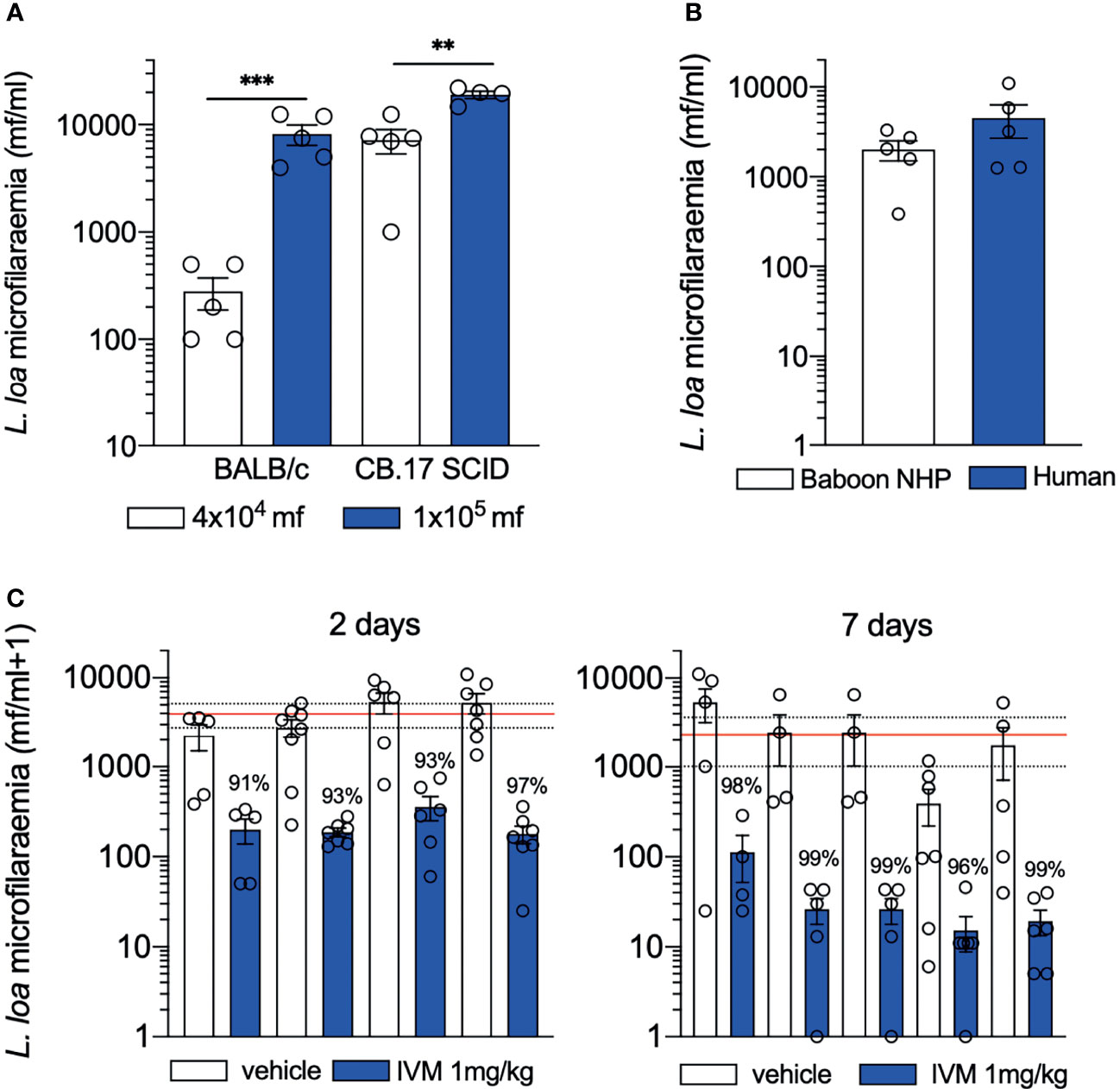
Figure 1 Performance of loiasis microfilaraemic mouse models. (A) microfilaraemias ≥10000 mf/ml can be achieved in both BALB/c (immunocompetent) and CB.17 SCID mice by increasing unit of inoculation (B) L. loa mf ethically sourced from human volunteers establishes similar microfilaraemias to those derived from NHP (C) consistent >90% depletions in L. loa microfilaraemias are mediated following single oral treatment with ivermectin 2 days (left) or 7 days (right) post-treatment in CB.17 SCID mice in multiple independent experiments. Data plotted is mean ± SEM. Lines are global averages of vehicle treated mice (red) and 95% confidence intervals (dashed). Significant differences are indicated ***P < 0.001 and **P < 0.01. Data is previously unpublished (A, B) and combination of published (42) and previously unpublished (C).
The successful development of a range of loiasis in vivo models increases opportunity for translational science applications. However, the generation of infectious stage larvae has been limited to dissections of wild-caught Chrysops in baited traps or human landing catch, which is laborious and hinders throughput. Intrathoracic injection of mf into blackfly vectors have been successful with Onchocerca species (49–51). Recently, it was demonstrated that L. loa mf purified from experimentally infected baboons and intrathoracically injected in to wild caught Chrysops developed to infective larvae after 14 days of fly rearing with high resultant yields of L3 (52). Validation experiments with these experimentally reared L. loa L3 demonstrated they could be cultured to undergo L3-L4 moulting in vitro and developed to adult stages in C57BL/6 RAG2-/-IL-2γc-/-mice.
The microfilaricidal activities of anti-malarial drugs (mefloquine, amodiaquine, artesunate, chloroquine and quinine), anthelmintics (praziquantel, flubendazole and its metabolites), trypanocidal agents (fexinidazole and Scynexis-7158) and the anti-cancer drug, imatinib, have been evaluated against L. loa mf in dose titration assays (53). Imatinib is of interest as it has established in vitro adulticidal activity against B. malayi (54). Further, ‘counter-screening’ experiments have been run to scrutinise potential off-target effects of the onchocerciasis anti-Wolbachia macrofilaricide clinical candidate, ABBV-4083 and lead-optimised quinazoline anti-Wolbachial compounds (55, 56). Rapid inhibition of L. loa motility was noted with mefloquine and amodiaquine. These antimalarial compounds achieved 50% inhibition concentrations (IC50) <5 µg/mL within the first 24 hours of exposure. Scynexis-7158 also induced a concentration-dependent reduction in mf motility but more gradually, with an IC50 of 10μg/mL after 5 days, whereas imatinib only had minor reductions in mf motility (<50%) over five days in culture, in a concentration-dependent manner. Contrastingly, praziquantel and fexinidazole were completely inactive. Flubendazole and its metabolites as well as the anti-Wolbachials ABBV-4083, CBR417 and CBR490 were also inactive until doses far exceeded the physiological range (55, 56). In a distinct study, 10μM flubendazole or its active metabolite was confirmed inactive against L. loa mf after 72 hours exposure (57). The direct activities of anti-malarial or anti-trypanosomidal drugs on L. loa mf in vitro highlights a potential opportunity for re-purposing development but also flags a possibility of AE risk during the treatment of other tropical diseases in loiasis microfilaraemic individuals. However, in a single open-label randomised trial, standard dose amodiaquine did not impact on L. loa mf in circulation up to 90 days post-treatment (58).
In comparison to inactivity against L. loa mf in vitro, when tested against developing L3 in culture, flubendazole and its metabolites achieved gradual IC50 threshold reductions in larval motility after 15 days, in a dose-dependent manner (41). This benzimidazole anthelmintic also completely blocked the L3-L4 moult at all doses tested (as low as 50 ng/mL). These pilot in vitro L. loa studies illustrate the potential of new L. loa culture systems as pharmacological screens to identify selective efficacies of repurposing or novel small molecule drugs before triaging into animal testing.
Because CB.17 SCID mice sustained the highest yields of L. loa microfilaraemias, this model was validated as a drug screen. CB.17 SCID mice were infused with 4x104 L. loa mf and treated with single-dose oral ivermectin at 1mg/kg, aligning to human standard dose systemic exposures (42). Multiple experiments demonstrated a consistent, rapid >90% clearance of mf two days after single oral dose ivermectin which further increased to approximately 99% mf clearance after seven days (Figure 1C). This model has been utilised to evaluate five direct-acting and nine anti-Wolbachia Onchocerca macrofilaricide candidates. Oxfendazole, a veterinary benzimidazole anthelmintic with macrofilaricidal activity in Litomosoides sigmodontis rodent infection models (59), has undergone phase I trials as a repurposed treatment for human helminthiases (60). Encouragingly, this candidate showed no direct activity against L. loa mf (42) and is currently undergoing further clinical development for filarial indications (61). Similarly, an oral-bioavailable formulation of flubendazole developed by Janssen Pharmaceutica as a potential Onchocerca macrofilaricide (62) was tested in the L. loa microfilaraemic SCID mouse model and found to be inactive (42).
When baboons with L. loa mf densities >8000 mf/ml of blood are treated with ivermectin at a standard dose of 150μg/kg, a significant decrease of microfilaremia in all treated animals, up to 98.4% reduction, is achieved by day 7 (33). Clinical manifestations are evident 2 days post-treatment mirroring early symptomology observed in human loiasis (7, 8). Body temperature, respiratory rates and pulse rates all increase above the normal range. Other clinical manifestations observed are body rashes and itches, pinkish ears, swollen face, conjunctival haemorrhages, loss of appetite, and diarrhoea. Animals display subdued behaviour 48 hours after ivermectin treatment with fatal SAE apparent in baboons >100,000 mf/ml (33). Autopsy findings in this study revealed a widespread vasculopathy. Petechial haemorrhages were seen in the CNS, the lungs, the conjunctiva, the cardiac tissues, the peritoneum and the omentum. Histopathological examination identified mf in varying degrees of degeneration in small vessels, associated with deposition of fibrin, endothelial changes including damage of blood vessels and the presence of extravascular erythrocytes. There was an increased presence of extravascular eosinophils and mononuclear cells, often in large numbers and associated with mf destruction. Highly vascularized organs like the brain, heart, lungs, and kidneys were observed to have more mf in tissue sections and mf were also present within the peritoneal cavity indicating extravasation following ivermectin treatment (33).
Exploiting the establishment of L. loa microfilariaemias in BALB/c immune-intact mice, the inflammatory response post-ivermectin was evaluated. Immune priming of BALB/c mice with subcutaneous inoculations of heat-killed L. loa mf 2-weeks prior to infusion with 4x104 L. loa mf and coincident treatment augmented the microfilaricidal treatment response from 90% in naive to 97% in immune-challenged mice, providing evidence that prior exposure to L. loa mf antigens in immune-competent mice can bolster the already substantial rapid efficacy of ivermectin (42). Elevated systemic inflammatory responses were noted in antigen-experienced mice after receiving ivermectin. Whilst initially both type-1 (IFNγ) type-2 (IL-4, IL-5, CCL11) and regulatory type (IL-10) inflammatory mediators were upregulated post-treatment, by day 7, a switch to a predominant type-2 inflammatory signature was apparent, characterised by maintenance of IL-4 and IL-5, downregulation of IFNγ and IL-10 and significant increases of the eosinophil chemokine, eotaxin (CCL11). Increased eosinophilia was apparent in peripheral circulation, in secondary lymphoid tissue and in the peritoneal cavity of microfilaraemic mice post-ivermectin treatment.
Following clinical evidence that W. bancrofti antigen-based immunodiagnostic tests cross-react with L. loa patients with high microfilaraemias (63), laboratory investigations were initiated to understand the molecular basis of cross-reactivity. Experimental studies using whole blood or sera derived from microfilaraemic baboons or the supernatants of short-term (6h) cultures of L. loa mf or L3, determined consistent, strong cross-reactivity with the W. bancrofti immuno-chromatographic test (ICT) (64). Follow on culture studies utilising pre-patent or patent male or female 24h cultures derived from RAG2-/-γc-/- mice demonstrated cross-reactivity was evident to the filarial test strip (FTS, a next generation ICT test) (42). Further, whole blood from lymphopenic mice chronically infected with adult stage L. loa were FTS positive both before and after the onset of mf production (42). By using a quantitative immunoassay (ELISA), Og4C3, which utilises distinct monoclonal antibodies to those employed in the ICT/FTS rapid diagnostic, cross-reactivity in frozen plasma derived from patently infected RAG2-/-γc-/- mice was also evident. These studies illustrate that a common or several distinct secreted antigen/s from L. loa mf, L3, immature adults, mature males and mature females cross-react with current monoclonal antibodies used in available antigen tests for LF mapping and elimination surveillance. Proteomic pull-down experiments have identified a specific ‘Av33-like’ secreted L. loa protein antigen derived from immature adult L. loa cultures and from loiasis patient serum that is recognised by the monoclonal antibody, AD12, used in LF ICT/FTS rapid diagnostic tests (65).
In the past decade, a substantial advance has been made in preclinical technologies for the tropical neglected disease, loiasis (Figure 2). These have been motivated to a large extent by the pressing public health barrier to filarial NTD elimination within Central Africa caused by this infection, namely: ivermectin SAEs in the treatment of onchocerciasis and cross-reactivity to current LF diagnostics. Important outcomes of applying loiasis preclinical tools have been to confirm that three onchocerciasis macrofilaricide clinical candidates positioned in phase I or II trials (two anti-Wolbachia drugs, ABBV-4083 & AWZ1066S, one re-purposed benzimidazole, oxfendazole) (61, 66) do not emulate ivermectin rapid microfilaricidal efficacies in vitro or in vivo. Application of a baboon NHP model has confirmed experimentally that ivermectin neurological SAEs are associated with pre-treatment hyper-microfilaraemias, multi-organ vasculopathy and CNS inflammation. Finally, use of L. loa animal models have elucidated the molecular basis for cross-reactivity with current LF immunodiagnostics.
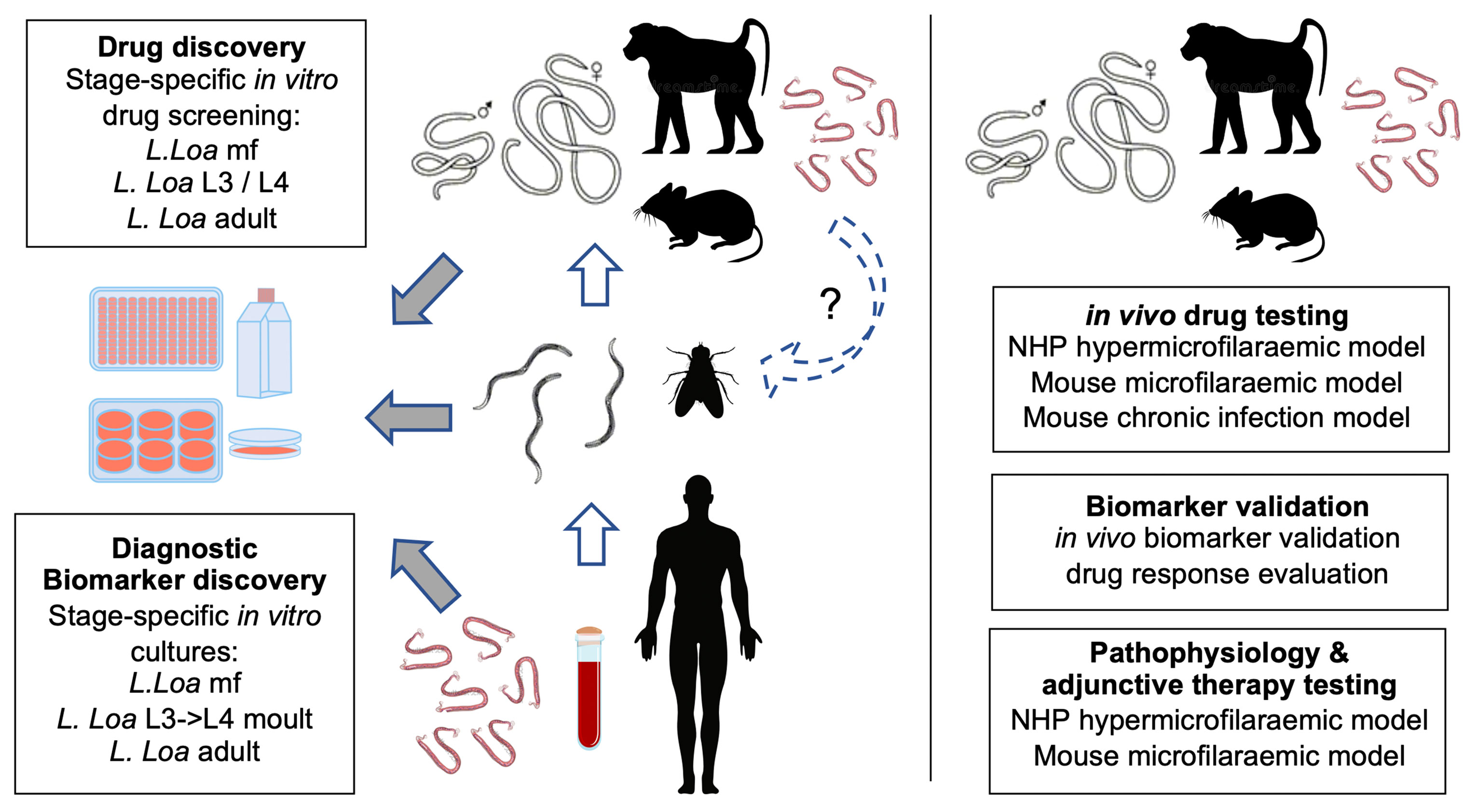
Figure 2 Summary of loiasis preclinical platforms and applications in translational research. Left: generation of all life cycle stages is achievable by blood sampling human patients, purification of L. loa mf, experimental injections in Chrysops, isolation of infectious stage larvae and chronic infections of either compound lymphopenic mice or splenectomised baboons. Derived L. loa mf, L3 or adults can be utilised in long-term in vitro cultures for drug or diagnostic biomarker discovery. It is proposed that mf from chronic mouse infections can be used to passage into Chrysops to establish a full laboratory life cycle (dashed loop). Right: in vivo infection models can be utilised to determine selective macrofilaricidal efficacy, validate candidate biomarkers and to research the aetiology of ivermectin adverse events including targetable pathways for adjunct therapeutics.
Current limitations of loiasis preclinical platforms are the requirements for wild-caught Chrysops for infectious larvae generation. Should a method be established to maintain the Chrysops life-cycle in the laboratory, this would resolve a final barrier in establishing a full laboratory life-cycle. In the interim, it may be possible to cryopreserve and ship large batches of L. loa L3 to expand experimental usage, including in vitro screening and experimental infections of mice, as has been founded for O. volvulus (67).
Now that tools for loiasis preclinical research are established and scalable, at least within the context of specialist laboratories within Central Africa (proximal to sources of natural infection and vectors), it is opportune to consider how these platforms may be applied in the future to advance filariasis therapeutics and diagnostics research (Box 1). The success in establishing a chronic mouse model of loiasis offers a new tool to define the true extent of albendazole efficacy with chronic dose regimens, utilising human bio-equivalent 400-800mg/day dosages defined in mice (68). Comparatively, this model can also be utilised to determine the minimum sufficient drug exposures of clinical Onchocerca macrofilaricide candidates, oxfendazole and emodepside, mediating significant curative activity against L. loa. By modelling pharmacokinetic (PK) metrics of drug exposures achieved in mice aligned with tolerated doses in humans (derived from available phase I trial data) an effective dose prediction can be determined which will be an important decision-making tool for proceeding into clinical testing for loiasis-specific indications (Figure 3). Simultaneously, establishing a complete curative drug regimen in the loiasis mouse model will be important to evaluate the utility of LF cross-reactive immunodiagnostics as serological biomarkers of adult living L. loa and prognostic determinants of macrofilaricidal activity. Biobanking of serum and urine from drug-cured and vehicle control mice could similarly be exploited in future for omics-based discovery of living adult worm biomarkers. For emodepside, a macrofilaricidal regimen with selectivity over acute killing of mf in circulation remains to be resolved for the treatment loiasis or onchocerciasis in L. loa co-endemic regions. For this, the SCID microfilaraemic mouse model can be employed to address whether human predicted efficacious doses mediate ivermectin-like rapid microfilaricidal activity.
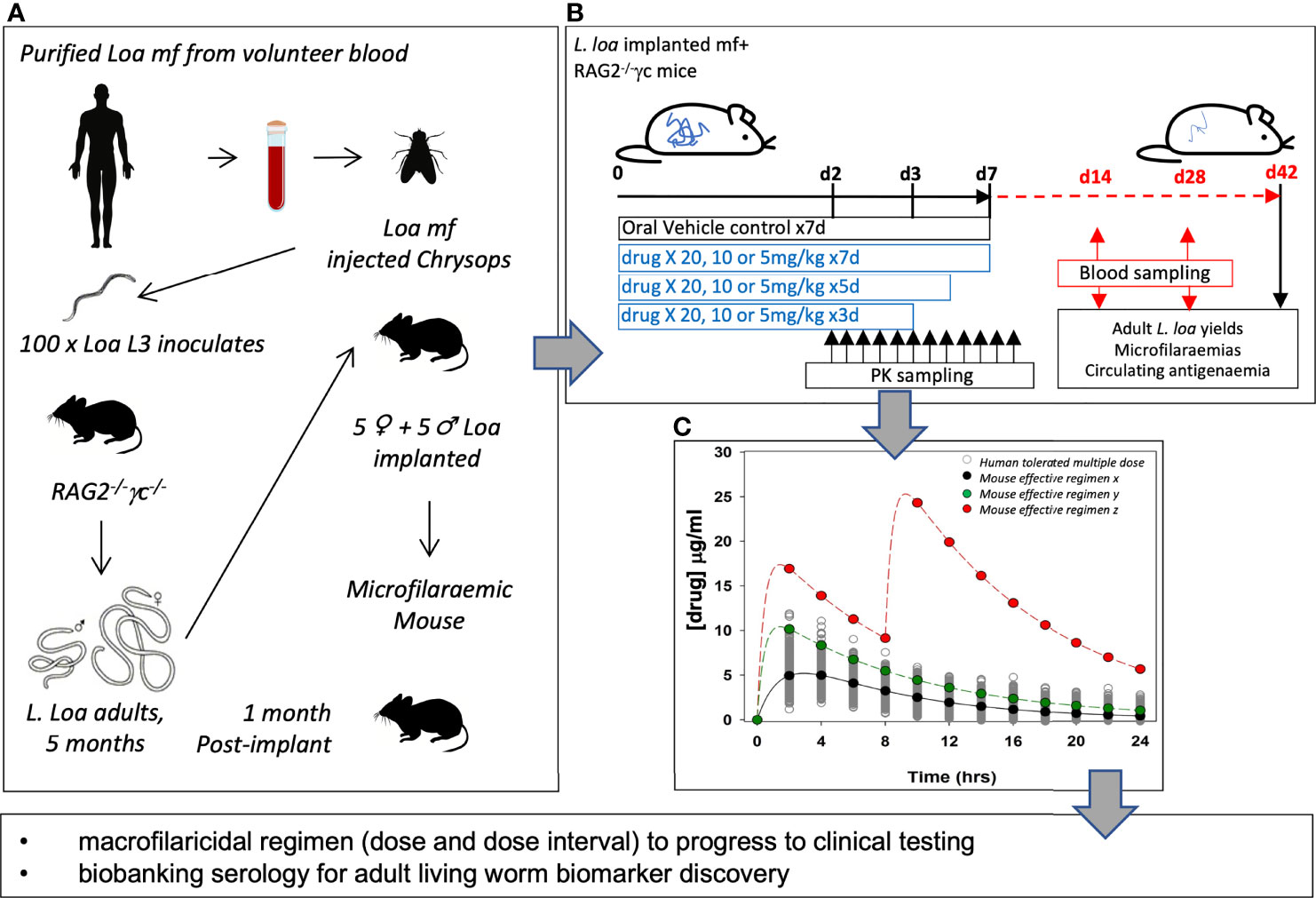
Figure 3 Application of a chronic loiasis mouse model in candidate macrofilaricide drug evaluation. (A) strategy for developing microfilaraemic mice with defined implanted number of male and female L. loa. (B) schematic of drug screen to comprehensively evaluate macrofilaricidal activity of candidate drugs. (C) PK modelling of effective macrofilaricidal drug exposures to predict human efficacious dose.
Box 1. Future directions in loiasis preclinical research.
drug development
● dose optimization of albendazole as a loiasis indication
● dose prediction of oxfendazole and emodepside as loiasis curative indications
● assessment of emodepside selectivity as a loiasis macrofilaricide vs microfilaricide
diagnostics development
● can LF immunodiagnostic cross-reactivity be exploited to measure macrofilaricidal efficacy?
● novel loiasis-specific biomarker discovery
aetiology of ivermectin AE and adjunctive therapeutic development
● role of host inflammatory responses in development of ivermectin adverse events
● assessment of pharmacological and biological interventions that modify host inflammation
Building on initial findings in both NHP and mouse microfilaraemic mouse models that ivermectin triggers type-2 inflammation, vasculopathy and a myeloid, eosinophil-rich cell recruitment, the availability of murine reagents and genetic modifications to manipulate facets of this inflammatory pathway can be exploited to define causal inflammatory components driving adverse reactions. Central to this research will be the development of quantitative clinical measures of ivermectin adverse events in mice, such as heart rate, oxygen saturation, core temperature and gross motor activity changes, which can be simultaneously captured with remote telemetry devices. A detailed knowledge of underlying inflammatory circuits in loiasis microfilaraemic mice following ivermectin treatment can then be considered for pharmacological or biological interventions aimed at disrupting these pathways and thus ameliorating febrile/allergic-type inflammatory adverse events. Selective secondary testing of any highly promising adjunctive intervention in the hypermicrofilaraemic NHP model would evaluate whether it might be possible to mitigate against SAE development or prolongment. Affordable, effective adjunctive treatments may possibly increase acceptability of ivermectin-based MDA campaigns if clinically validated. In lieu of safe macrofilaridical treatments for onchocerciasis and/or loaisis, this strategy might be operationalised to achieve filariasis elimination targets set for 2030.
SW: writing, reviewing, and funding. VC: data collection and writing. FF: writing and reviewing. AN: writing and reviewing. NG: writing and reviewing. MR: reviewing and funding. PE: reviewing. CM: reviewing and funding. MT: reviewing and funding. AH: reviewing and funding. JT: data analysis, writing, reviewing, and funding. All authors contributed to the article and approved the submitted version.
This work was supported by: Bill and Melinda Gates Foundation Grand Challenges Explorations Grant (OPP1119043) to JT, SW, and MT; Wellcome Trust Institutional Translational Partnership Award to JT and SW (219576/Z/19/Z); Merck & Co via the Mectizan Donation Programme (MDP) to SW and CM; German Research Foundation (DFG) (Grant DFG HO 2009/10-1, HO 2009/14-1, RI 3036/1-1) within the “German-African Cooperation Projects in infectiology” (grants awarded to AH, MR, and SW). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Eberhard ML, Orihel TC. Development and Larval Morphology of Loa Loa in Experimental Primate Hosts. J Parasitol (1981) 67(4):556–64. doi: 10.2307/3280490
2. Zouré HGM, Wanji S, Noma M, Amazigo UV, Diggle PJ, Tekle AH, et al. The Geographic Distribution of Loa Loa in Africa: Results of Large-Scale Implementation of the Rapid Assessment Procedure for Loiasis (RAPLOA). PloS Negl Trop Dis (2011) 5(6):e1210. doi: 10.1371/journal.pntd.0001210
3. Boussinesq M, Kamgno J, Pion SD, Gardon J. What Are the Mechanisms Associated With Post-Ivermectin Serious Adverse Events? Trends Parasitol (2006) 22(6):244–6. doi: 10.1016/j.pt.2006.04.006
4. Whittaker C, Walker M, Pion SDS, Chesnais CB, Boussinesq M, Basáñez MG. The Population Biology and Transmission Dynamics of Loa Loa. Trends Parasitol (2018) 34(4):335–50. doi: 10.1016/j.pt.2017.12.003
5. Chesnais CB, Takougang I, Paguélé M, Pion SD, Boussinesq M. Excess Mortality Associated With Loiasis: A Retrospective Population-Based Cohort Study. Lancet Infect Dis (2017) 17(1):108–16. doi: 10.1016/S1473-3099(16)30405-4
6. Ducorps M, Gardon-Wendel N, Ranque S, Ndong W, Boussinesq M, Gardon J, et al. Secondary Effects of the Treatment of Hypermicrofilaremic Loiasis Using Ivermectin. Bull Soc Pathol Exot (1995) 88(3):105–12.
7. Boussinesq M, Gardon J, Gardon-Wendel N, Kamgno J, Ngoumou P, Chippaux JP. Three Probable Cases of Loa Loa Encephalopathy Following Ivermectin Treatment for Onchocerciasis. Am J Trop Med Hyg (1998) 58(4):461–9. doi: 10.4269/ajtmh.1998.58.461
8. Gardon J, Kamgno J, Folefack G, Gardon-Wendel N, Bouchité B, Boussinesq M. Marked Decrease in Loa Loa Microfilaraemia Six and Twelve Months After a Single Dose of Ivermectin. Trans R Soc Trop Med Hyg (1997) 91(5):593–4. doi: 10.1016/S0035-9203(97)90041-9
9. Chippaux JP, Boussinesq M, Gardon J, Gardon-Wendel N, Ernould JC. Severe Adverse Reaction Risks During Mass Treatment With Ivermectin in Loiasis-Endemic Areas. Parasitol Today (1996) 12(11):448–50. doi: 10.1016/0169-4758(96)40006-0
10. Gardon J, Gardon-Wendel N, Demanga-Ngangue, Kamgno J, Chippaux JP, Boussinesq M. Serious Reactions After Mass Treatment of Onchocerciasis With Ivermectin in an Area Endemic for Loa Loa Infection. Lancet (1997) 350(9070):18–22. doi: 10.1016/S0140-6736(96)11094-1
11. Fobi G, Gardon J, Santiago M, Ngangue D, Gardon-Wendel N, Boussinesq M. Ocular Findings After Ivermectin Treatment of Patients With High Loa Loa Microfilaremia. Ophthalmic Epidemiol (2000) 7(1):27–39. doi: 10.1076/0928-6586(200003)7:1;1-2;ft027
12. Herrick JA, Makiya MA, Holland-Thomas N, Klion AD, Nutman TB. Infection-Associated Immune Perturbations Resolve 1 Year Following Treatment for Loa Loa. Clin Infect Dis (2021) 72(5):789–96. doi: 10.1093/cid/ciaa137
13. Legrand F, Herrick J, Makiya M, Ramanathan R, Thompson R, Rampertaap S, et al. A Randomized, Placebo-Controlled, Double-Blind Pilot Study of Single-Dose Humanized Anti-IL5 Antibody (Reslizumab) for the Reduction of Eosinophilia Following Diethylcarbamazine Treatment of Loa Loa Infection. Clin Infect Dis (2020) 73(7):e1624–31. doi: 10.1093/cid/ciaa1365
14. Mackenzie C, Geary T, Prichard R, Boussinesq M. Where Next With Loa Loa Encephalopathy? Data Are Badly Needed. Trends Parasitol (2007) 23(6):237–8. doi: 10.1016/j.pt.2007.04.007
15. D’Ambrosio MV, Bakalar M, Bennuru S, Reber C, Skandarajah A, Nilsson L, et al. Point-Of-Care Quantification of Blood-Borne Filarial Parasites With a Mobile Phone Microscope. Sci Transl Med (2015) 7(286):286re4. doi: 10.1126/scitranslmed.aaa3480
16. Lenk EJ, Moungui HC, Boussinesq M, Kamgno J, Nana-Djeunga HC, Fitzpatrick C, et al. A Test-And-Not-Treat Strategy for Onchocerciasis Elimination in Loa Loa-Coendemic Areas: Cost Analysis of a Pilot in the Soa Health District, Cameroon. Clin Infect Dis (2020) 70(8):1628–35. doi: 10.1093/cid/ciz461
17. Forrer A, Wanji S, Obie ED, Nji TM, Hamill L, Ozano K, et al. Why Onchocerciasis Transmission Persists After 15 Annual Ivermectin Mass Drug Administrations in South-West Cameroon. BMJ Glob Heal (2021) 6(1):e003248. doi: 10.1136/bmjgh-2020-003248
18. Kim YE, Remme JHF, Steinmann P, Stolk WA, Roungou JB, Tediosi F. Correction to: Control, Elimination, and Eradication of River Blindness: Scenarios, Timelines, and Ivermectin Treatment Needs in Africa. PloS Negl Trop Dis (2015) 9(5):e0003777. doi: 10.1371/journal.pntd.0003777
19. Billingsley P, Binka F, Chaccour C, Foy BD, Gold S, Gonzalez-Silva M, et al. A Roadmap for the Development of Ivermectin as a Complementary Malaria Vector Control Tool. Am J Trop Med Hyg (2020) 102:3–24. doi: 10.4269/ajtmh.19-0620
20. Beng AA, Esum ME, Deribe K, Njouendou AJ, Ndongmo PWC, Abong RA, et al. Mapping Lymphatic Filariasis in Loa Loa Endemic Health Districts Naïve for Ivermectin Mass Administration and Situated in the Forested Zone of Cameroon. BMC Infect Dis (2020) 20(1):284. doi: 10.1186/s12879-020-05009-3
21. Wanji S, Esum ME, Njouendou AJ, Mbeng AA, Chounna Ndongmo PW, Abong RA, et al. Mapping of Lymphatic Filariasis in Loiasis Areas: A New Strategy Shows No Evidence for Wuchereria Bancrofti Endemicity in Cameroon. PloS Negl Trop Dis (2018) 13(3):e0007192. doi: 10.1371/journal.pntd.0007192
22. Kamgno J, Nguipdop-Djomo P, Gounoue R, Téjiokem M, Kuesel AC. Effect of Two or Six Doses 800 Mg of Albendazole Every Two Months on Loa Loa Microfilaraemia: A Double Blind, Randomized, Placebo-Controlled Trial. PloS Negl Trop Dis (2016) 10(3):e0004492. doi: 10.1371/journal.pntd.0004492
23. Klion AD, Massougbodji A, Horton J, Ekoué S, Lanmasso T, Ahouissou NL, et al. Albendazole in Human Loiasis: Results of a Double-Blind, Placebo-Controlled Trial. J Infect Dis (1993) 168(1):202–6. doi: 10.1093/infdis/168.1.202
24. Zofou D, Fombad FF, Gandjui NVT, Njouendou AJ, Kengne-Ouafo AJ, Chounna Ndongmo PW, et al. Evaluation of In Vitro Culture Systems for the Maintenance of Microfilariae and Infective Larvae of Loa Loa. Parasit Vectors (2018) 11(1):275. doi: 10.1186/s13071-018-2852-2
25. Wahl G, Georges AJ. Current Knowledge on the Epidemiology, Diagnosis, Immunology, and Treatment of Loiasis. Trop Med Parasitol (1995) 46(4):287–91.
26. Duke BO, Wijers DJ. Studies on Loiasis in Monkeys. I. The Relationship Between Human and Simian Loa in the Rain-Forest Zone of the British Cameroons. Ann Trop Med Parasitol England; (1958) 52(2):158–75. doi: 10.1080/00034983.1958.11685857
27. Orihel TC, Eberhard ML. Loa Loa: Development and Course of Patency in Experimentally-Infected Primates. Trop Med Parasitol (1985) 36(4):215–24.
28. Orihel TC, Moore PJ. Loa Loa: Experimental Infection in Two Species of African Primates. Am J Trop Med Hyg (1975) 24(4):606–9. doi: 10.4269/ajtmh.1975.24.606
29. Pinder M, Everaere S, Roelants GE. Loa Loa: Immunological Responses During Experimental Infections in Mandrills (Mandrillus Sphinx). Exp Parasitol (1994) 79(2):126–36. doi: 10.1006/expr.1994.1072
30. DUKE BO. Studies on Loiasis in Monkeys. II.–The Population Dynamics of the Microfilariae of Loa in Experimentally Infected Drills (Mandrillus Leucophaeus). Ann Trop Med Parasitol Engl (1960) 54:15–31. doi: 10.1080/00034983.1960.11685953
31. DUKE BO. Studies on Loiasis in Monkeys. III. The Pathology of the Spleen in Drills (Mandrillus Leucophaeus) Infected With Loa. Ann Trop Med Parasitol England; (1960) 54:141–6. doi: 10.1080/00034983.1960.11685969
32. Wanji S, Eyong EE, Tendongfor N, Ngwa C, Esuka E, Kengne-Ouafo A, et al. Parasitological, Hematological and Biochemical Characteristics of a Model of Hyper-Microfilariaemic Loiasis (Loa Loa) in the Baboon (Papio Anubis). PloS Negl Trop Dis (2015) 9(11):e0004202. doi: 10.1371/journal.pntd.0004202
33. Wanji S, Eyong EEJ, Tendongfor N, Ngwa CJ, Esuka EN, Kengne-Ouafo AJ, et al. Ivermectin Treatment of Loa Loa Hyper-Microfilaraemic Baboons (Papio Anubis): Assessment of Microfilarial Load Reduction, Haematological and Biochemical Parameters and Histopathological Changes Following Treatment. PloS Negl Trop Dis (2017) 11(7):e0005576. doi: 10.1371/journal.pntd.0005576
34. Turner JD, Pionnier N, Furlong-Silva J, Sjoberg H, Cross S, Halliday A, et al. Interleukin-4 Activated Macrophages Mediate Immunity to Filarial Helminth Infection by Sustaining CCR3-Dependent Eosinophilia. PloS Pathog (2018) 14(3):e1006949. doi: 10.1371/journal.ppat.1006949
35. Howells RE, Devaney E, Smith G, Hedges T. The Susceptibility of BALB/C and Other Inbred Mouse Strains to Brugia Pahangi. Acta Trop (1983) 40(4):341–50. doi: 10.5169/seals-313141
36. Petit G, Diagne M, Maréchal P, Owen D, Taylor D, Bain O. Maturation of the Filaria Litomosoides Sigmodontis in BALB/c Mice; Comparative Susceptibility of Nine Other Inbred Strains. Ann Parasitol Hum Comparée (1992) 67(5):144–50. doi: 10.1051/parasite/1992675144
37. Bain O, Wanji S, Enyong P, Petit G, Noireau F, Eberhard MI, et al. New Features on the Moults and Morphogenesis of the Human Filaria Loa Loa by Using Rodent Hosts. Consequences. Parasite (1998) 5(1):37–46. doi: 10.1051/parasite/1998051037
38. Wanji S, Tendongfor N, Vuong PN, Enyong P, Bain O. The Migration and Localization of Loa Loa Infective and Fourth-Stage Larvae in Normal and Immunosuppressed Rodents. Ann Trop Med Parasitol (2002) 96(8):823–30. doi: 10.1179/000349802125002220
39. Chunda VC, Ritter M, Bate A, Gandjui NVT, Esum ME, Fombad FF, et al. Comparison of Immune Responses to Loa Loa Stage-Specific Antigen Extracts in Loa Loa-Exposed BALB/c Mice Upon Clearance of Infection. Parasit Vectors (2020) 13(1):51. doi: 10.1186/s13071-020-3921-x
40. Tendongfor N, Wanji S, Ngwa JC, Esum ME, Specht S, Enyong P, et al. The Human Parasite Loa Loa in Cytokine and Cytokine Receptor Gene Knock Out BALB/c Mice: Survival, Development and Localization. Parasit Vectors (2012) 5(1):43. doi: 10.1186/1756-3305-5-43
41. Fombad FF, Njouendou AJ, Ndongmo PC, Ritter M, Chunda VC, Metuge HM, et al. Effect of Flubendazole on Developing Stages of Loa Loa In Vitro and In Vivo: A New Approach for Screening Filaricidal Agents. Parasit Vectors (2019) 12(1):14. doi: 10.1186/s13071-018-3282-x
42. Pionnier NP, Sjoberg H, Chunda VC, Fombad FF, Chounna PW, Njouendou AJ, et al. Mouse Models of Loa Loa. Nat Commun (2019) 10(1):1429. doi: 10.1038/s41467-019-09442-0
43. Kilarski WW, Martin C, Pisano M, Bain O, Babayan SA, Swartz MA. Inherent Biomechanical Traits Enable Infective Filariae to Disseminate Through Collecting Lymphatic Vessels. Nat Commun (2019) 10(1):2895. doi: 10.1038/s41467-019-10675-2
44. Cadman ET, Thysse KA, Bearder S, Cheung AYN, Johnston AC, Lee JJ, et al. Eosinophils Are Important for Protection, Immunoregulation and Pathology During Infection With Nematode Microfilariae. PloS Pathog (2014) 10(3):e1003988. doi: 10.1371/journal.ppat.1003988
45. Pionnier N, Sjoberg H, Furlong-Silva J, Marriott A, Halliday A, Archer J, et al. Eosinophil-Mediated Immune Control of Adult Filarial Nematode Infection Can Proceed in the Absence of IL-4 Receptor Signaling. J Immunol (2020) 205(3):731–40. doi: 10.4049/jimmunol.1901244
46. Hawking F. The Chemotherapy of Filarial Infections. Pharmacol Rev United States; (1955) 7(2):279–99.
47. Halliday A, Guimaraes AF, Tyrer, Metuge HM, Patrick CNW, Arnaud KOJ, et al. A Murine Macrofilaricide Pre-Clinical Screening Model for Onchocerciasis and Lymphatic Filariasis. Parasit Vectors (2014) 7:472. doi: 10.1186/s13071-014-0472-z
48. Sjoberg HT, Pionnier N, Aljayyoussi G, Metuge HM, Njouendou AJ, Chunda VC, et al. Short-Course, Oral Flubendazole Does Not Mediate Significant Efficacy Against Onchocerca Adult Male Worms or Brugia Microfilariae in Murine Infection Models. PloS Negl Trop Dis (2019) 13(1):e0006356. doi: 10.1371/journal.pntd.0006356
49. Bianco AE, Ham PJ, Townson S, Mustafa MB, Nelson GS. A Semi-Automated System of Intrathoracic Injection for the Large-Scale Production of Onchocerca Lienalis Infective Larvae. Trop Med Parasitol (1989) 40(1):57–64.
50. Dohnal J, Blinn J, Wahl G, Schulz-key H. Distribution of Microfilariae of Onchocerca Lienalis and Onchocerca Gutturosa in the Skin of Cattle in Germany and Their Development in Simulium Ornatum and Culicoides Nubeculosus Following Artificial Infestation. Vet Parasitol (1990) 36(3–4):325–32. doi: 10.1016/0304-4017(90)90044-C
51. Fukuda M, Takaoka H, Uni S, Bain O. Infective Larvae of Five Onchocerca Species From Experimentally Infected Simulium Species in an Area of Zoonotic Onchocerciasis in Japan. Parasite (2008) 15(2):111–9. doi: 10.1051/parasite/2008152111
52. Ndzeshang LB, Fombad FF, Njouendou AJ, Chunda VC, Gandjui NVT, Akumtoh DN, et al. Generation of Loa Loa Infective Larvae by Experimental Infection of the Vector, Chrysops Silacea. PloS Negl Trop Dis (2020) 14(8):e0008415. doi: 10.1371/journal.pntd.0008415
53. Njouendou AJ, Fombad FF, O’Neill M, Zofou D, Nutting C, Ndongmo PC, et al. Heterogeneity in the In Vitro Susceptibility of Loa Loa Microfilariae to Drugs Commonly Used in Parasitological Infections. Parasit Vectors (2018) 11(1):223. doi: 10.1186/s13071-018-2799-3
54. O’Connell EM, Bennuru S, Steel C, Dolan MA, Nutman TB. Targeting Filarial Abl-Like Kinases: Orally Available, Food and Drug Administration-Approved Tyrosine Kinase Inhibitors Are Microfilaricidal and Macrofilaricidal. J Infect Dis (2015) 212(5):684–93. doi: 10.1093/infdis/jiv065
55. Bakowski MA, Shiroodi RK, Liu R, Olejniczak J, Yang B, Gagaring K, et al. Discovery of Short-Course Antiwolbachial Quinazolines for Elimination of Filarial Worm Infections. Sci Transl Med (2019) 11(491):eaav3523. doi: 10.1126/scitranslmed.aav3523
56. Taylor MJ, Von Geldern TW, Ford L, Hübner MP, Marsh K, Johnston KL, et al. Preclinical Development of an Oral Anti-Wolbachia Macrolide Drug for the Treatment of Lymphatic Filariasis and Onchocerciasis. Sci Transl Med (2019) 11(483):eaau2086. doi: 10.1126/scitranslmed.aau2086
57. O’Neill M, Njouendou JA, Dzimianski M, Burkman E, Ndongmo PC, Kengne-Ouafo JA, et al. Potential Role for Flubendazole in Limiting Filariasis Transmission: Observations of Microfilarial Sensitivity. Am J Trop Med Hyg (2018) 98(1):21–6. doi: 10.4269/ajtmh.17-0390
58. Kamgno J, Djomo PN, Pion SD, Thylefors B, Boussinesq M. A Controlled Trial to Assess the Effect of Quinine, Chloroquine, Amodiaquine, and Artesunate on Loa Loa Microfilaremia. Am J Trop Med Hyg (2010) 82(3):379–85. doi: 10.4269/ajtmh.2010.09-0573
59. Hübner MP, Martin C, Specht S, Koschel M, Dubben B, Frohberger SJ, et al. Oxfendazole Mediates Macrofilaricidal Efficacy Against the Filarial Nematode Litomosoides Sigmodontis In Vivo and Inhibits Onchocerca Spec. Motility In Vitro. PloS Negl Trop Dis (2020) 14(7):e0008427. doi: 10.1371/journal.pntd.0008427
60. Bach T, Galbiati S, Kennedy JK, Deye G, Nomicos EYH, Codd EE, et al. Pharmacokinetics, Safety, and Tolerability of Oxfendazole in Healthy Adults in an Open-Label Phase 1 Multiple Ascending Dose and Food Effect Study. Antimicrob Agents Chemother (2020) 64(11):e01018–20. doi: 10.1128/AAC.01018-20
61. Drugs for Neglected Diseases initiative (DNDi). Oxfendazole | DNDi . Available at: https://dndi.org/research-development/portfolio/oxfendazole/ (Accessed Accessed: 6th June 2021).
62. Lachau-Durand S, Lammens L, van der Leede BJ, Van Gompel J, Bailey G, Engelen M, et al. Preclinical Toxicity and Pharmacokinetics of a New Orally Bioavailable Flubendazole Formulation and the Impact for Clinical Trials and Risk/Benefit to Patients. PloS Negl Trop Dis (2019) 13(1):e0007026. doi: 10.1371/journal.pntd.0007026
63. Pion SD, Montavon C, Chesnais CB, Kamgno J, Wanji S, Klion AD, et al. Positivity of Antigen Tests Used for Diagnosis of Lymphatic Filariasis in Individuals Without Wuchereria Bancrofti Infection But With High Loa Loa Microfilaremia. Am J Trop Med Hyg (2016) 95(6):1417–23. doi: 10.4269/ajtmh.16-0547
64. Wanji S, Amvongo-Adjia N, Njouendou AJ, Kengne-Ouafo JA, Ndongmo WPC, Fombad FF, et al. Further Evidence of the Cross-Reactivity of the Binax NOW® Filariasis ICT Cards to non-Wuchereria Bancrofti Filariae: Experimental Studies With Loa Loa and Onchocerca Ochengi. Parasit Vectors (2016) 9(1):267. doi: 10.1186/s13071-016-1556-8
65. Hertz MI, Nana-Djeunga H, Kamgno J, Jelil Njouendou A, Chawa Chunda V, Wanji S, et al. Identification and Characterization of Loa Loa Antigens Responsible for Cross-Reactivity With Rapid Diagnostic Tests for Lymphatic Filariasis. PloS Negl Trop Dis (2018) 12(11):e0006963. doi: 10.1371/journal.pntd.0006963
66. Johnston KL, Hong WD, Turner JD, O’Neill PM, Ward SA, Taylor MJ. Anti-Wolbachia Drugs for Filariasis. Trends Parasitol (2021) S1471-4922(21):00143–4. doi: 10.1016/j.pt.2021.06.004
67. Abraham D, Lange AM, Yutanawiboonchai W, Trpis M, Dickerson JW, Swenson B, et al. Survival and Development of Larval Onchocerca Volvulus in Diffusion Chambers Implanted in Primate and Rodent Hosts. Parasitol (1993) 79(4):571–82. doi: 10.2307/3283385
Keywords: Loa, loiasis, filariasis, onchocerciasis, lymphatic filariasis, neglected tropical disease
Citation: Wanji S, Chunda VC, Fombad FF, Njouendou AJ, Gandjui NVT, Ritter M, Enyong PA, Mackenzie C, Taylor MJ, Hoerauf A and Turner JD (2021) Advances in Preclinical Platforms of Loa loa for Filarial Neglected Tropical Disease Drug and Diagnostics Research. Front. Trop. Dis 2:778724. doi: 10.3389/fitd.2021.778724
Received: 17 September 2021; Accepted: 19 October 2021;
Published: 08 November 2021.
Edited by:
Jeremy Foster, New England Biolabs, United StatesReviewed by:
Edward Mitre, Uniformed Services University of the Health Sciences, United StatesCopyright © 2021 Wanji, Chunda, Fombad, Njouendou, Gandjui, Ritter, Enyong, Mackenzie, Taylor, Hoerauf and Turner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Samuel Wanji, c3dhbmppQHlhaG9vLmZy; Joseph D. Turner, Sm9zZXBoLlR1cm5lckBsc3RtZWQuYWMudWs=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.