 Mariana V. N. Arena
Mariana V. N. Arena Rogério H. Toppa2
Rogério H. Toppa2 Isabel Alves-dos-Santos
Isabel Alves-dos-Santos- 1Programa de Pós-Graduação em Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, SP, Brazil
- 2Departamento de Ciências Ambientais, Núcleo de Estudos em Ecologia da Paisagem e Conservação, Universidade Federal de São Carlos, Sorocaba, SP, Brazil
- 3Departamento de Geografia, Turismo e Humanidades, Núcleo de Estudos em Ecologia da Paisagem e Conservação, Universidade Federal de São Carlos, Sorocaba, SP, Brazil
- 4Departamento de Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, SP, Brazil
Contemporary urban ecology views cities as heterogeneous and dynamic ecosystems, composed of biotic, physical, social, and man-made systems. In this context, urban beekeeping stands out as a growing practice that demands further studies. Many species of stingless bees (tribe Meliponini) have adapted to urban areas, while others are more sensitive. The composition of the surroundings where the beehive is located is important for the health of the nests and quality of the bee products. This work used bee capture and release techniques to evaluate the use of this methodology for releasing and monitoring native bees in urban areas and to identify whether proportion of green and gray urban nature areas affect the flying activities of stingless bees in an urban landscape. We used nests of Melipona quadrifasciata (mandaçaia), a species of stingless bee native to the Atlantic Rainforest, from three meliponaries located in different parts of the city of São Paulo. The travel time of bees in the landscape were related to the green urban areas up to 900 m in diameter from the meliponaries. Although we did not find a relationship between green areas and bee return times and numbers, it was noticeable that there was variation between the study areas, indicating that the release and monitoring methodology can be used in urban areas following the recommended modifications.
1. Introduction
Within the contemporary scope of the urban ecology, cities are studied as heterogeneous and dynamic ecosystems, composed of biotic, physical, social and built complexes (Cadenasso and Pickett, 2008). Wachsmuth and Angelo (2018) defined two representations of urban nature that can characterize urban sustainability policy: green urban nature and gray urban nature. Green urban nature encompasses all vegetation forms that can be found in urban areas (e.g., street trees, gardens, plazas, grass), while gray urban nature includes other sustainable urban spaces, such as dense urban cores and buildings (Wachsmuth and Angelo, 2018). About gray urban nature features, urban areas have a high percentage of impermeable surfaces and buildings, which influence the local microclimate not only by replacing natural forest areas and altering the natural flow of water systems, but also by intensely absorbing solar radiation, creating heat islands (Yuan and Bauer, 2007; Hamblin et al., 2018). A road, for example, can act as a barrier to flying insects, and the composition of these organisms can differ between the two sides of the road (Andersson et al., 2017).
Regarding green urban nature features, urban areas not only contain remnants of patches of vegetation (Pickett and Cadenasso, 2012), but also feature resources not native to the region, distributed in gardens and green areas (Kaluza et al., 2016; Silva and Kleinert, 2020), and resource heterogeneity distinctly influence both bee dynamics and diet (Banaszak-Cibicka et al., 2016; Dylewski et al., 2019). Urban gardens, for example, can act as providers of floral resources and refuge for bees and other pollinators when resources are scarce (Langellotto et al., 2018), and urban green areas can provide habitat for several species of nesting bees pre-existing cavities (Rocha-Filho et al., 2020).
In this context, meliponiculture, the breeding and management of stingless bees (Meliponini tribe), is an increasingly common modern practice in urban areas in the west hemisphere that demands further studies (Cortopassi-Laurino et al., 2006; Venturieri et al., 2013). Brazil has a great diversity of stingless bee species which increasingly attract the interest of society, whether to produce specialty honeys and marketable products, for conservation and educational purposes or as a leisure activity (Koser et al., 2020). Recent updates in federal (Brasil, 2020) and state (São Paulo, 2021) legislation have helped regulate the practice of rearing these native bees, which have advantages over honeybees (Apis mellifera Linnaeus, 1758) because they are native, have special honeys and are not harmful to humans.
Some species such as “yellow jataí” (Tetragonisca angustula Latreille, 1811) and “arapuá” (Trigona spinipes Fabricius, 1793) have adapted to urban areas and live in anthropic habitats (Sousa et al., 2002), being able to nest both in tree trunks and in artificial structures, such as hollow walls (Silveira et al., 2002). Other species, however, such as those of the genus Melipona, are more sensitive and have not adapted so well to these new environments (Pioker-Hara et al., 2014). Melipona bees can suffer adverse effects of deforestation and can be considered indicators of landscape change (Brown and Albretcht, 2001), mainly due to the necessity of finding large cavities for nesting, such as large trees, which are rare in urban areas (Pioker-Hara et al., 2014). Thus, their breeding requires experience of the stingless beekeeper and additional handling of nest, as well as greater availability and diversity of plant species in the area where they are located (Oliveira-Abreu et al., 2014; Andrade et al., 2019), in addition to beekeeping programs that aim to maintain regional biodiversity (Brown and Albretcht, 2001). However, the potential of Melipona bees for beekeeping is excellent, even in urban areas, not only for the creation and commercialization of unique bee products, but also to contribute to the conservation of this endangered species, combining the need to increase the number of these individuals with the conservation of forest remnants and their plant-pollinator interactions.
Although beekeepers provide their hives with energy and protein artificial supplements, it is essential for the maintenance of the nests that the bees forage and seek natural sources of nectar and pollen to meet all their nutritional needs that maintain the health of the colony. Thus, it is known that the composition and proportion of the landscape directly affect the richness of resources (pollen) collected by stingless bees (Machado et al., 2020). The arrangement and distribution of trees and shrubs and the presence of tall structures such as buildings (Forman, 2016), for example, are green and gray interferences that affect organisms in urban areas. In addition, stingless bees have diversified flight ranges, which define how far a bee can go to reach best quality floral resources. Maximum flight distances can reach more than 2,000 m for Melipona bees (Roubik and Aluja, 1983). However, in methodologies involving capture and release, bees typically do not return to the colonies when they are released at distances greater than 1,000 m, and only stingless bees of the genera Melipona and Trigona return from these distances (Araújo et al., 2004; Greenleaf et al., 2007). Still, the real distances that bees travel for foraging depend on the attractiveness of the resources to compensate for their distance from the nest and on the availability of alternative resources nearby, so that they usually do not exceed 500 m (Heard, 1999). Nevertheless, none of the mentioned studies were carried out in urban areas, so nothing is known about the flight ranges of these bees in this landscape context.
In this context, the bee flying activities on urban landscapes need further studies to better understand their foraging and pollination dynamics. In this work we used bee capture and release techniques to identify whether green infrastructure affect the activity of stingless bees and to evaluate the effectiveness of this method in an urban landscape. Therefore, we aimed to answer: can the methodology for releasing and monitoring native bees be used in urban areas? What is the role of green and gray infrastructure in the capacity and return time of bees to the hive?
2. Materials and methods
2.1. Study area and species
The study was carried out in the city of São Paulo, state of São Paulo, Brazil, and the target species chosen was Melipona quadrifasciata Lepeletier 1836 (typically called mandaçaia). São Paulo is a large metropolis, being the most populous city in Brazil and the fifth most populated in the world, with more than 20 million inhabitants (World Population Review, 2022). Of the total area of the municipality, 735.99 km2 (48.18%) are composed of vegetal cover, including areas of natural cover, regeneration, reforestation, etc., being 21% of natural Atlantic Forest vegetal cover (São Paulo, 2020). The municipality's rural area represents 31.78% of the territory, from which 79.37% are vegetation cover, while the urban area represents 68.22% of the territory, from which only 33.65% are of vegetation cover (São Paulo, 2020).
Mandaçaia is an important species native to the Atlantic Forest (Nogueira-Neto, 1997; Michener, 2007; Camargo et al., 2013) and is commonly breed in meliponaries in the state of São Paulo. This species is typical from forest environments (Silveira et al., 2002), so it is very sensitive to the loss of habitat, that is the reality of urban areas. Mandaçaia was chosen because it is native to the region, it has a valuable potential for the beekeeper, and it is one of the favorites among breeders, mainly in urban areas.
2.2. Data collection
Three landscapes were selected to compose a gradient of green-gray urban nature, where meliponaries used to collect bees for the experiment were located (Figure 1). To ensure that all hives of mandaçaia were in similar condition, nests were monitored during 6 months (autumn and winter) and fed with syrup (sugar solution: 50% water; 50% organic sugar). The experiments took place in early spring (September 2022). For each study area (meliponary) the bees were randomly divided into three groups of 15 individuals (n = 45), and bees were marked with a water-based colored pen (POSCA PC-5M), so that each group was assigned to a color. Bees were placed in plastic pots (100 ml) according to their groups, and immediately transported in thermic boxes with ice. Considering that bees typically do not return to the colonies when they are released at distances greater than 1,000 m (Araújo et al., 2004; Greenleaf et al., 2007), we established a maximum release distance of 900 m. For each study area, groups of bees were released at three distances from the nest: 300 m (subbuffer 1), 600 m (subbuffer 2), and 900 m (subbuffer 3), and the direction in which the releases were made was randomized (Figure 1).
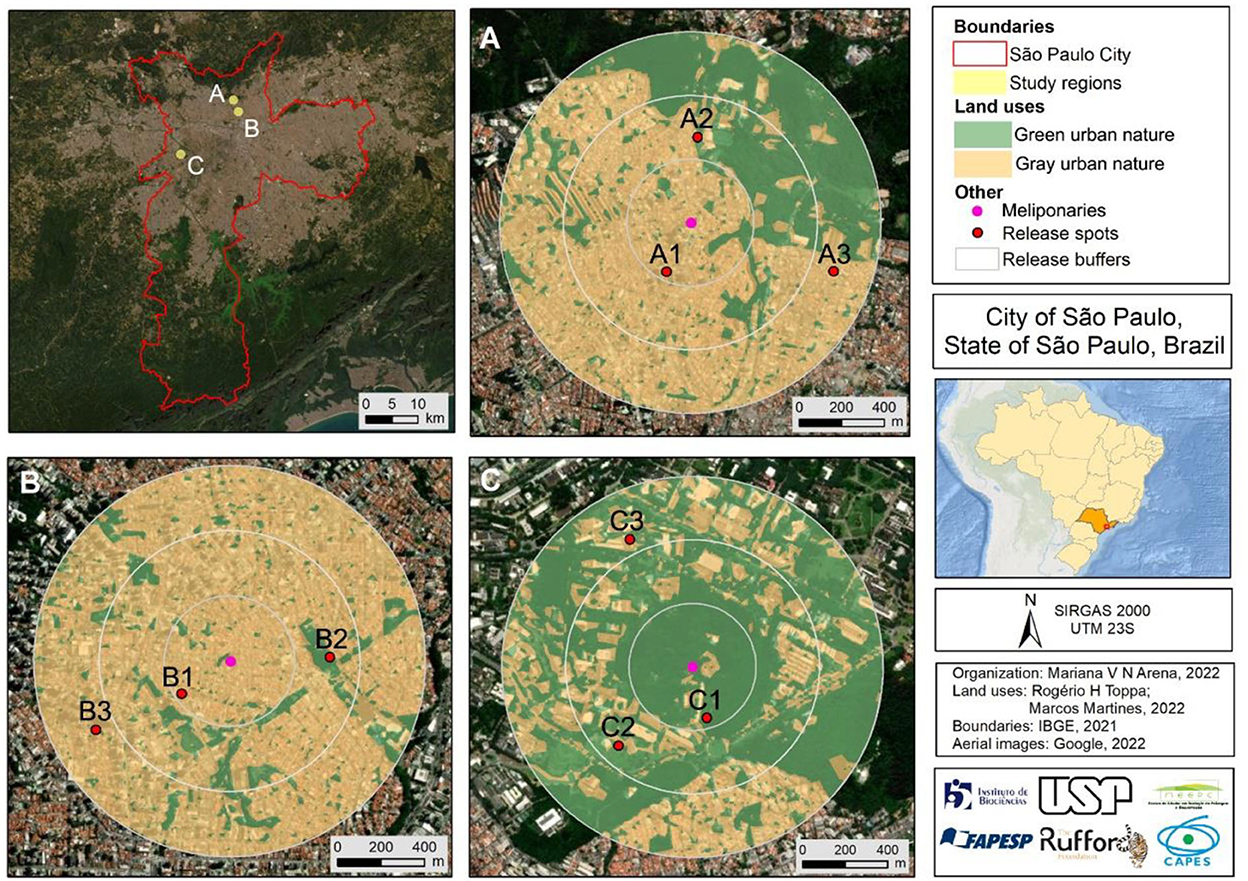
Figure 1. Location of study areas (A–C) (meliponaries) in the city of São Paulo, state of São Paulo, Brazil, showing coverage regions, urban green and gray nature areas, and bees' release spots and coverage regions.
The releases of bees from each nest were carried out on the same day of collection, on sunny days, between 8:00 am and 9:00 am. The releases started from the closest point (300 m) to the farthest (900 m), and the exact time of each release was noted. A video camera monitoring system was used to check how many individuals returned to the nests and how long they took to return. A webcam was installed at the entrance of the nest and connected to a notebook to record the bees arriving to the hive, for 3 h after the last bee was released. The images were analyzed to obtain the arrival times of the bees of each release group (visually identified by color) to the nest.
2.3. Spatial analysis
The land cover for each study area was classified as “green urban nature” and “gray urban nature” (Figure 1). The classification was performed with Sentinel satellite images, using as a filter the temporal space of 1 year (08/30/21 to 22), to obtain the median with the most characteristic classification of the spectral response of that image. Having the meliponaries as a central point, 900 m radius buffers were generated to demarcate the coverage regions around each study area. Filters were created based on the NDVI and EVI bands, in order to obtain specific responses regarding the presence of vegetation cover. The automatic classification of land uses was performed using the Earth Engine platform using the “random forest” classifier, and the training was performed 1,000 times to verify the best classification. Finally, to confirm the accuracy of the classification, confusion matrices were generated.
2.4. Data analysis
For each bee individual that returns to the nest, the difference between the time of release and the time of record was calculated to obtain the number of individuals that returned per nest and the return times (in minutes). Generalized linear models (GLMs) with gamma and poisson distributions were created to verify the relationship between the response variables and the predictor variables, in order to identify which parameters influenced the return time intervals (minutes) of bees. Data were organized in Excel and statistical analysis were performed using the Jamovi 2.2.5 software.
We estimated if there were differences in the return times between subbuffers within buffers, assuming subbuffers categories as predictor variables. To find if there were differences due to the distances of the release points (subbuffers) within coverage regions (buffers), we generated GLMs for buffer A considering subbuffers A1 (300 m), A2 (600 m), and A3 (900 m); for buffer B considering subbuffers B1 (300 m), B2 (600 m), and B3 (900 m); and for buffer C considering subbuffers C1 (300 m), C2 (600 m), and C3 (900 m).
Additionally, we answered if there were differences in return time between the cover regions, assuming buffers as predictor variables. Finally, we wanted to know if there was a relationship between the vegetation amount and the return time. Therefore, data was organized considering the total vegetation area (Km2) and number of patches (NP) as predictor variables.
3. Results
Exploratory analysis showed that in study area C bees took longer to return from the release points to the nest, followed by the bees from the areas B and A, respectively (Figure 2). Study area A presented the higher number of returning bees (31), followed by areas C (24) and B (21) (Figures 2–4).
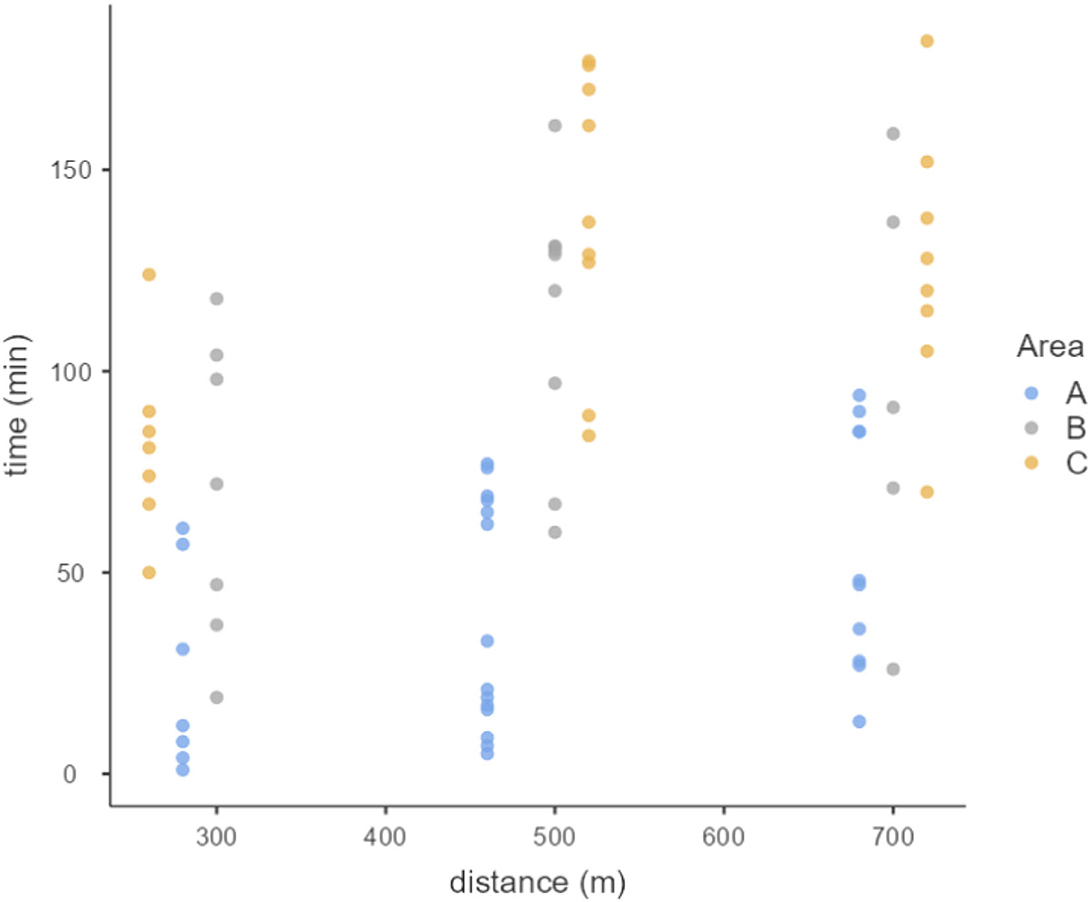
Figure 2. Dispersion graph showing the return time (minutes) of each bee according to the release point distance for each study area (A, B, C).
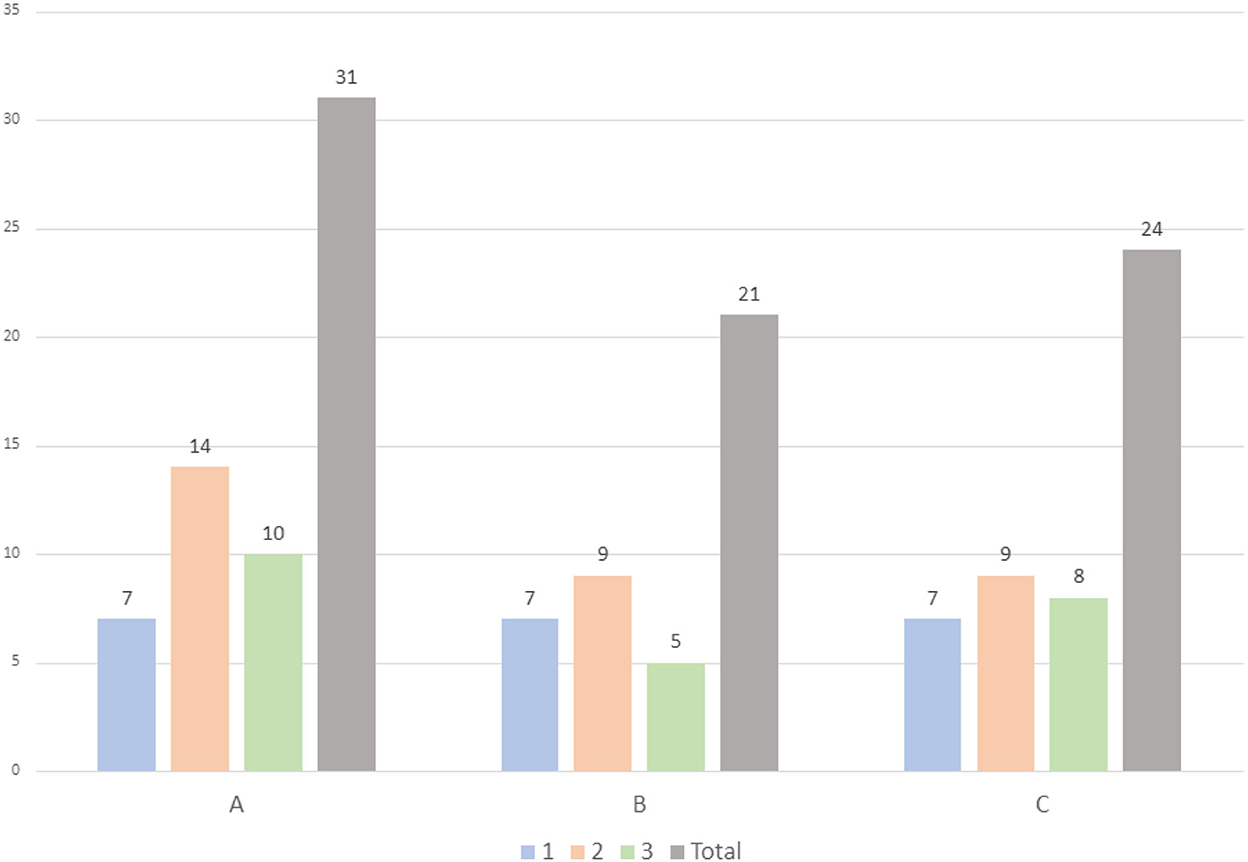
Figure 3. Total number of returning bees per study cover regions (A, B, C) and subbuffers (1, 2, 3, and total).
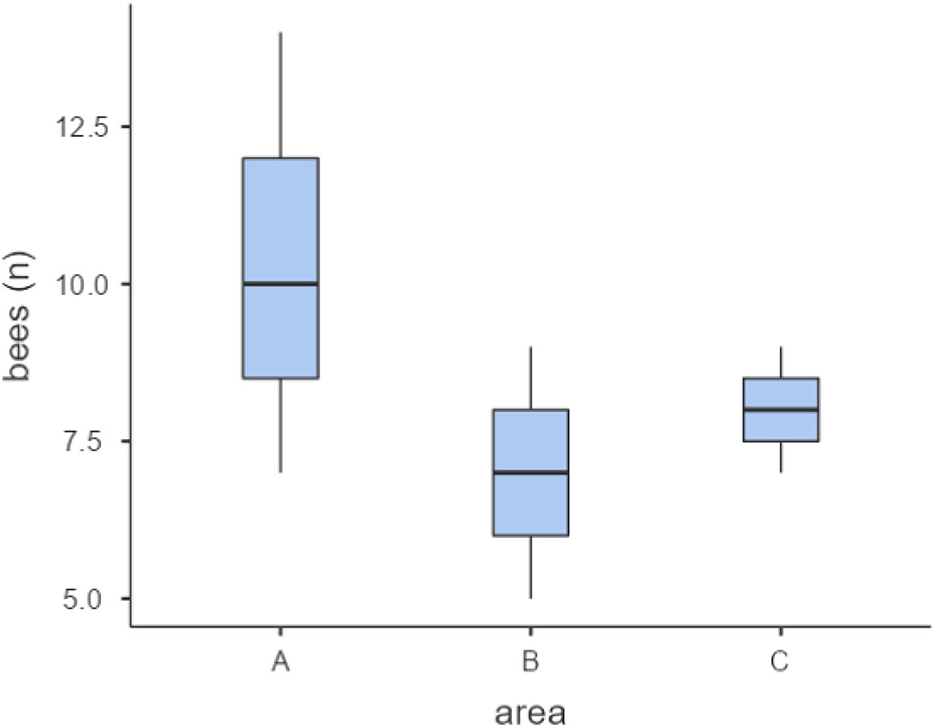
Figure 4. Plots accounting the number of bees that returned to the nest after release for each study area (A,B,C).
Models regarding return time comparisons between subbuffers within coverage buffers (Table 1; Figure 5) showed no significant differences, except for subbuffers C1 and C3 (p < 0.001). Differences in return time between buffers showed significant differences between study areas A and B and between areas A and C (Table 2; Figure 6). We did not find statistically significant differences between the vegetation area and the return time or between the number of patches and the return time (Table 3). Detailed results of the generated models are available in the Supplementary material.
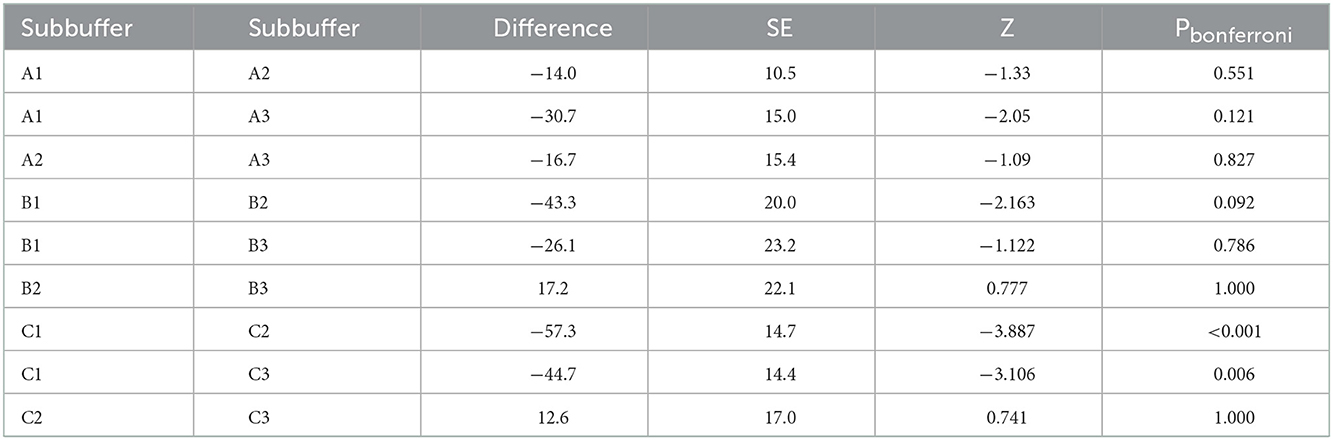
Table 1. Post-hoc comparison tests for the generalized linear models regarding differences between subbuffers.
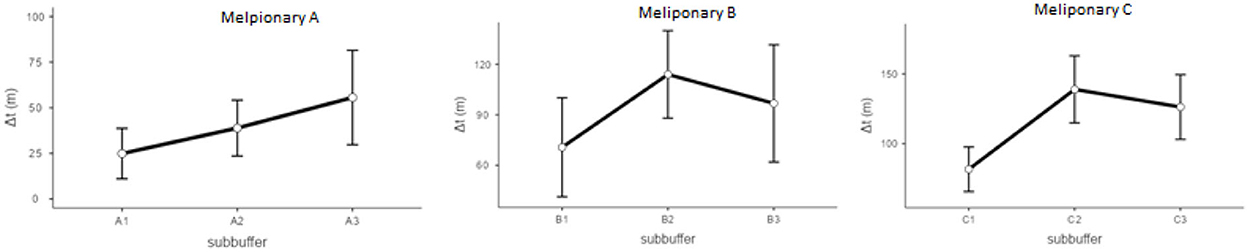
Figure 5. Plot results for the generalized linear models regarding differences between subbuffers.
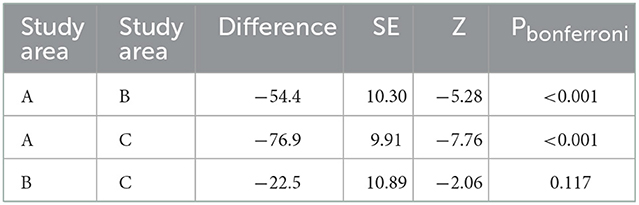
Table 2. Post-hoc comparison tests for the generalized linear models regarding differences in return time between buffers.
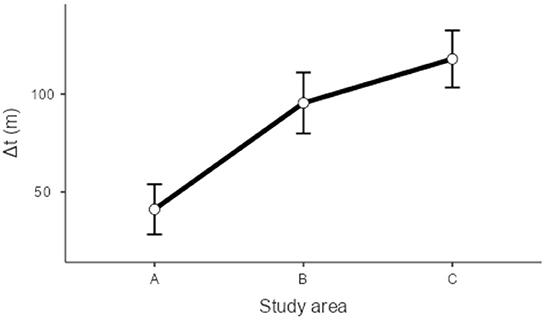
Figure 6. Plot results for the generalized linear models regarding differences in return time between buffers.
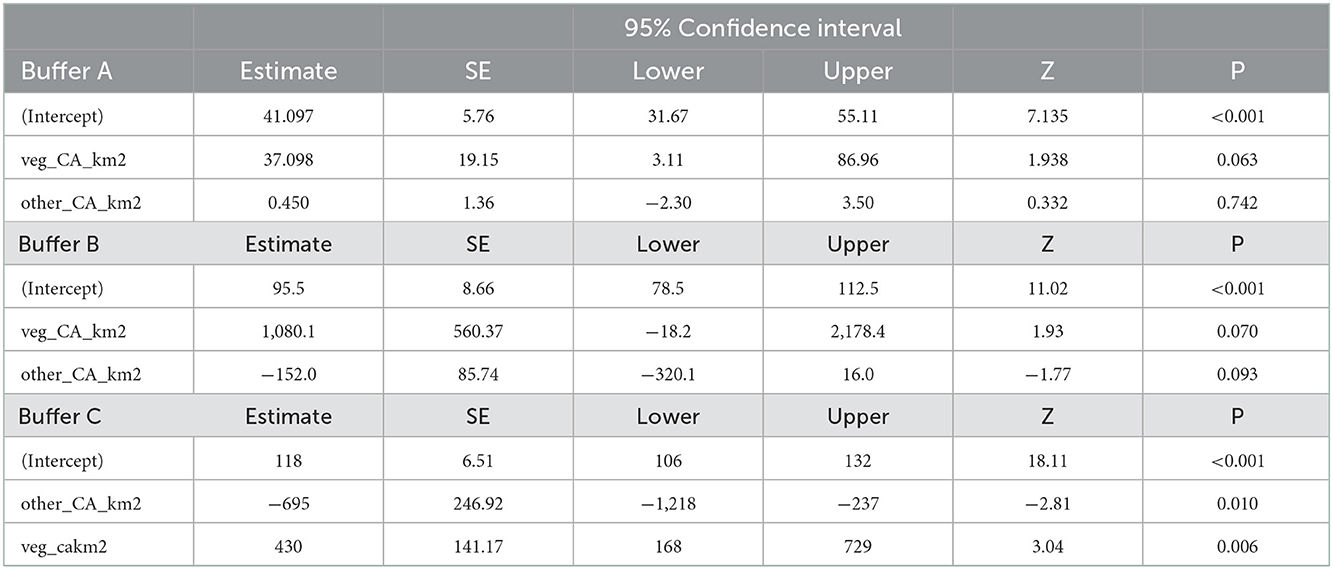
Table 3. Parameter estimates for the generalized linear models regarding differences between the vegetation area and the return time for buffers A, B, and C.
4. Discussion
Although we did not find a relationship between vegetation and bee return times and numbers, it was noticeable that there was variation between the study regions, showing that the release and monitoring methodology can be used in urban areas. We obtained satisfactory bee return rates of 46.6% (21 out of 45 bees in area B), 53.3% (24 out of 45 bees in area C), and 68.8% (31 out of 45 bees in area A). Marking techniques have been widely applied in studies with several conservation purposes and with a variety of permanent and impermanent methods (see Briggs et al., 2022). Our study proved that the method of marking and releasing bees can also be successful on different green-gray urban nature gradients. Additionally, we used a very wide spatial scale in this work in order to cover the green-gray gradient. To efficiently develop conservation strategies for local bees in urban landscapes, it is necessary to consider different spatial scales (Zanette et al., 2005). Thus, our findings report a new context for studies applied to the ecology of the movement of pollinating organisms in urban environments. Accordingly, it would be interesting to carry out a new study that uses greater spatial detail to investigate the influences of different elements of the urban landscape, such as street trees, paved roads or vacant lots.
In order to be able to infer whether variations between study regions occurred due to the landscape around the nests and not to other factors (such as intercolonial variations), it would be suitable to repeat the experiment with a greater number of study areas. We suggest, for a next experiment, that more releases be carried out, at different distances, in order to cover a better representation of the landscape around the meliponaries. Bees are organisms that carry out recursive movement, defined as repeated visits to the same locations in a systematic way, between series of known points (“traplines”), in search of renewable resources, such as pollen and nectar (Berger-Tal and Bar-David, 2015). Thus, understanding how bees move and interact with anthropic landscapes, both in search of resources and in pollination, is extremely important both for the conservation of bees and the resources they need, and for good agricultural productivity (Heard, 1999; Machado et al., 2020; Silva and Kleinert, 2020).
Unfortunately, the technique used is limited to identify the routes of the bees returning to the hive. For this, there would be necessary to use more modern but expensive technologies for monitoring bee activity, such as harmonic radar. Although transmitter sizes have recently become small enough to allow tracking of insects under natural field conditions, they are still too big for most bee species and are still too expensive (Kissling et al., 2014; Nunes-Silva et al., 2019). It is also important to highlight that no marked bees were recorded returning with floral resources, demonstrating that during the return journey of the bees to the hive they did not carry out collection activities, focusing only on their return. Thus, there is no need to assess which resources are available in the vegetation to carry out this type of experiment, which allows the focus of the analyzes to be concentrated only on the type of urban element present in the landscape.
Bee flight capacities are an important factor that determines the area that a colony can exploit (Costa et al., 2021). Tetragonisca angustula and T. spinipes, for example, stingless bees commonly found in urban areas, can reach distances up to 1,000 m (Van Nieuwstadt and Iraheta, 1996; Araújo et al., 2004; Greenleaf et al., 2007). Typically, M. quadrifasciata can reach a flight distance of 2,100 m (Roubik and Aluja, 1983). However, recent studies using RFID tracking technology have revealed that Melipona bees can reach much greater flight ranges, with foraging flight distances from 1,000 to 2,000 m, and a maximum homing distance from 5 to 10 km (Nunes-Silva et al., 2019; Costa et al., 2021). Therefore, it is fundamental to study in detail the movement of bees and their relationship with the landscape elements in a radius of 1,000 m around the colony in order to assign the best conservation and management strategies of the meliponary and its surroundings. It is also important to highlight that, in both mentioned studies, the releases were carried out in forest areas, emphasizing the originality and importance of our survey and other studies that evaluate the movement of native bees in urban areas.
Although some studies about the flight ranges of native bees have been successfully developed in natural areas, there is a lack of studies about the dynamics of these organisms in urban landscapes. Research that considers the new anthropic scenario to which native bees are submitted is of unquestionable importance for the development of public and conservation policies that concern biodiversity and urban ecology. Thus, our study presents a relevant suggestion for new research within this modern context. We conclude that, although some modifications are necessary, studies involving the marking, release and monitoring of bees can be of great value for the development of urban ecology studies, in order to better understand how these organisms interact in these landscapes and, thus, develop better conservation and maintenance strategies for native bees in urban areas.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical review and approval was not required for the animal study because no authorization is required for ex-situ collection and transport of invertebrates. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
MA, RT, and IA-d-S contributed to conception and design of the study. MA and RT organized the database. MA, RT, and MM performed the statistical analysis. MA wrote the first draft of the manuscript and sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Acknowledgments
MA thanks CAPES and FAPESP (2021/10875-0) for the doctoral scholarship. We thank the Rufford Foundation and FUSP (3867 - RUFFORD/IB) for the grant funding and the stingless beekeepers for the partnership during the survey. We would also like to thank the editor and the reviewers of Frontiers for all the suggestions made to the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frsc.2022.1103835/full#supplementary-material
References
Andersson, P., Koffman, A., Sjödin, N. E., and Johansson, V. (2017). Roads may act as barriers to flying insects: species composition of bees and wasps differs on two sides of a large highway. Nat. Conserv. 18, 47–59. doi: 10.3897/natureconservation.18.12314
Andrade, B. R., Nascimento, A. S., Franco, E. L., Santos, D. R., Alves, R. M. O., Costa, M. A. P. C., et al. (2019). Pollen spectrum and trophic niche width of Melipona scutellaris Latreille, 1811 (Hymenoptera: Apidae) in highly urbanized and industrialized sites. Sociobiology 66, 279–286. doi: 10.13102/sociobiology.v66i2.3427
Araújo, E. D., Costa, M., Chaud-Netto, J., and Fowler, H. G. (2004). Body size and flight distance in stingless bees (Hymenoptera: Meliponini): inference of flight range and possible ecological implications. Braz. J. Biol. 64, 589–596. doi: 10.1590/S1519-69842004000400003
Banaszak-Cibicka, W., Ratyńska, H., and Dylewski, Ł. (2016). Features of urban green space favourable for large and diverse bee populations (Hymenoptera: Apoidea: Apiformes). Urban For Urban Green 20, 448–452. doi: 10.1016/j.ufug.2016.10.015
Berger-Tal, O., and Bar-David, S. (2015). Recursive movement patterns: review and synthesis across species. Ecosphere 6, 1–12. doi: 10.1890/ES15-00106.1
Brasil (2020). Resolução n° 496, de 19 de agosto de 2020. Disciplina o uso e o manejo sustentáveis das abelhas-nativas sem-ferrão em meliponicultura. Available online at: https://www.in.gov.br/en/web/dou/-/resolucao-n-496-de-19-de-agosto-de-2020-273217120 (accessed August 17, 2021).
Briggs, E. L., Baranski, C., Münzer Schaetz, O., Garrison, G., Collazo, J. A., Youngsteadt, E., et al. (2022). Estimating bee abundance: can mark-recapture methods validate common sampling protocols? Apidologie 53, 1–24. doi: 10.1007/s13592-022-00919-4
Brown, J. C., and Albretcht, C. (2001). The effect of tropical deforestation on stingless bees of the genus Melipona (Insecta: Hymenoptera: Apidae: Meliponini) in central Rondonia. Brazil. J. Biog. 28, 623–634. doi: 10.1046/j.1365-2699.2001.00583.x
Cadenasso, M. L., and Pickett, S. T. A. (2008). Urban principles for ecological landscape design and management: scientific fundamentals. Cities Environ. 1, 1–16. doi: 10.15365/cate.1242008
Camargo, J. M. F., Pedro, S. E. M., and Melo, G. A. R. (2013). Meliponini Lepeletier, 1836. In Moure JS, Urban D, Melo GAR (2021) Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region. Available online at: http://www.moure.cria.org.br/catalogue (accessed July 07, 2021).
Cortopassi-Laurino, M., Imperatriz-Fonseca, V. L., Roubik, D. W., Dollin, A., Heard, T., Aguilar, I., et al. (2006). Global meliponiculture: challenges and opportunities. Apidologie 37, 275–292. doi: 10.1051/apido:2006027
Costa, L., Nunes-Silva, P., Galaschi-Teixeira, J. S., Arruda, H., Veiga, J. C., Pessin, G., et al. (2021). RFID-tagged amazonian stingless bees confirm that landscape configuration and nest re-establishment time affect homing ability. Insectes Sociaux 68, 101–108. doi: 10.1007/s00040-020-00802-4
Dylewski, Ł., Maćkowiak, Ł., and Banaszak-Cibicka, W. (2019). Are all urban green spaces a favourable habitat for pollinator communities? Bees, butterflies and hoverflies in different urban green areas. Ecol. Entomol. 44, 678–689. doi: 10.1111/een.12744
Forman, R. T. T. (2016). Urban ecology principles: are urban ecology and natural area ecology really different? Landsc. Ecol. 31, 1653–1662. doi: 10.1007/s10980-016-0424-4
Greenleaf, S. S., Williams, N. M., Winfree, R., and Kremen, C. (2007). Bee foraging ranges and their relationship to body size. Oecologia 153, 589–596. doi: 10.1007/s00442-007-0752-9
Hamblin, A. L., Youngsteadt, E., and Frank, S. D. (2018). Wild bee abundance declines with urban warming, regardless of floral density. Urban Ecosyst. 21, 419–428. doi: 10.1007/s11252-018-0731-4
Heard, T. A. (1999). The role of stingless bees in crop pollination. Annu. Revi. of Entomol. 44, 183–206. doi: 10.1146/annurev.ento.44.1.183
Kaluza, B. F., Wallace, H., Heard, T. A., Klein, A. M., and Leonhardt, S. D. (2016). Urban gardens promote bee foraging over natural habitats and plantations. Ecol. Evol. 6, 1304–1316. doi: 10.1002/ece3.1941
Kissling, W. D., Pattemore, D. E., and Hagen, M. (2014). Challenges and prospects in the telemetry of insects. Biol. Rev. 89, 511–530. doi: 10.1111/brv.12065
Koser, R. J., Barbiéri, C., and Francoy, T. M. (2020). Legislation on meliponiculture in Brazil: social and environmental demand. Sustentabilidade em Debate 11, 164–194. doi: 10.18472/SustDeb.v11n1.2020.30319
Langellotto, G. A., Melathopoulos, A., Messer, I., Anderson, A., McClintock, N., Costner, C., et al. (2018). Garden pollinators and the potential for ecosystem service flow to urban and peri-urban agriculture. Sustain 10, 12–14. doi: 10.3390/su10062047
Machado, T., Viana, B. F., da Silva, C. I., and Boscolo, D. (2020). How landscape composition affects pollen collection by stingless bees? Landscape Ecol. 35, 747–759. doi: 10.1007/s10980-020-00977-y
Nogueira-Neto, P. (1997). Vida e criação de abelhas indígenas sem ferrão. São Paulo: Nogueirapis, 445.
Nunes-Silva, P., Costa, L., Campbell, A. J., Contrera, F. A. L., Teixeira, J. S. G., Gomes, R. L. C., et al. (2019). Radio Frequency Identification (RFID) reveals long distance flight and homing abilities of the stingless bee Melipona fasciculata. Apidologie. 51, 240–253. doi: 10.1007/s13592-019-00706-8
Oliveira-Abreu, C., Hilário, S. D., Luz, C. F. P., and Alves-dos-Santos, I. (2014). Pollen and nectar foraging by Melipona quadrifasciata anthidioides Lepeletier (Hymenoptera: Apidae: Meliponini) in natural habitat. Sociobiology 61, 441–448. doi: 10.13102/sociobiology.v61i4.441-448
Pickett, S. T. A., and Cadenasso, M. L. (2012). “Urban ecology,” in Ecological Systems: Selected Entries From the Encyclopedia of Sustainability Science and Technology, ed R. Lee- mans (New York: Springer), 273–301. doi: 10.1007/978-1-4419-0851-3_325
Pioker-Hara, F. C., Drummond, M. S., and Kleinert, A. M. P. (2014). The influence of the loss of Brazilian Savanna vegetation on the occurrence of stingless bees nests (Hymenoptera: Apidae: Meliponini). Sociobiology 61, 393–400. doi: 10.13102/sociobiology.v61i4.393-400
Rocha-Filho, L. C., Montagnana, P. C., Boscolo, D., and Garófalo, C. A. (2020). Green patches among a grey patchwork: the importance of preserving natural habitats to harbour cavity-nesting bees and wasps (Hymenoptera) and their natural enemies in urban areas. Biodivers. Conserv. 29, 2487–2514. doi: 10.1007/s10531-020-01985-9
Roubik, D. W., and Aluja, M. (1983). Flight ranges of Melipona and Trigona in tropical forest. J. Kansas Entomol. Soc. 56, 217–222. Available online at: http://www.jstor.org/stable/25084398
São Paulo (2020). Secretaria Municipal do Verde e do Meio Ambiente - Coordenação de Planejamento Ambiental. Mapeamento Digital da Cobertura Vegetal do Município de São Paulo: Relatório Final. São Paulo, SVMA. Available online at: https://www.prefeitura.sp.gov.br/cidade/secretarias/upload/meio_ambiente/RelCobVeg2020_vFINAL_compressed(1).pdf (accessed November 20, 2022).
São Paulo (2021). Resolução SIMA n° 11, de 03 de fevereiro de 2021. Cria a categoria de empreendimento de fauna silvestre “Meliponário” e dispõe sobre os procedimentos autorizativos para o uso e manejo de abelhas-nativas- sem-ferrão no Estado de São Paulo. Available online at: https://www.infraestruturameioambiente.sp.gov.br/legislacao/2021/02/resolucao-sima-no-11-2021 (accessed August 17, 2021).
Silva, C. I., and Kleinert, A. M. P. (2020). Plantas e pólen em áreas urbanas: uso no paisagismo amigável aos polinizadores. Rio Claro, Brasil: CISE, 40. Available online at: www.researchgate.net/publication/341408297_Plantas_e_polen_em_areas_urbanas_uso_no_paisagismo_amigavel_aos_polinizadores (accessed July 31, 2020).
Silveira, F. A., Melo, G. A. R., and Almeida, E. A. B. (2002). Abelhas brasileiras: sistemática e identificação. Fundação Araucária, Belo Horizonte.
Sousa, L. M., Pereira, T. O., Prezoto, F., and Faria-Mucci, F. M. (2002). Nest foundation and diversity of Meliponini (Hymenoptera: Apidae) in an urban area of the municipality of Juiz de Fora, MG, Brazil. Biosci. J. 18, 59–65. Available online at: https://seer.ufu.br/index.php/biosciencejournal/article/view/6420
Van Nieuwstadt, M. G. L., and Iraheta, C. E. R. (1996). Relation between size and foraging range in stingless bees (Apidae, Meliponinae). Apidologie 27, 219–228. doi: 10.1051/apido:19960404
Venturieri, G. C., Alves, D. D. A., Villas-Bôas, J. K., Carvalho, C. A. L. D., Menezes, C., Vollet-Neto, A., et al. (2013). “Meliponicultura no Brasil: Situação Atual e Perspectivas Futuras para o Uso na Polinização Agrícola,” in Polinizadores no Brasil - contribuição e perspectivas para a biodiversidade, uso sustentável, conservação e serviços ambientais, ed Imperatriz Fonseca et al. (Org.), São Paulo: Editora da Universidade de São Paulo. 213–236.
Wachsmuth, D., and Angelo, H. (2018). Green and gray: new ideologies of nature in urban sustainability policy. Ann. Am. Assoc. Geogr. 0, 1–19. doi: 10.1080/24694452.2017.1417819
World Population Review (2022). World City Populations 2022. Available online at: https://worldpopulationreview.com/world-cities (acessed November 20, 2022).
Yuan, F., and Bauer, M. E. (2007). Comparison of impervious surface area and normalized difference vegetation index as indicators of surface urban heat island effects in Landsat imagery. Remote Sens. Environ. 106, 375–386. doi: 10.1016/j.rse.2006.09.003
Keywords: urban ecology, urban biodiversity, urban green nature, urban gray nature, urban meliponaries, stingless beekeeping, Melipona quadrifasciata, mandaçaia
Citation: Arena MVN, Toppa RH, Martines M and Alves-dos-Santos I (2023) Release experiments as an indicator of flying activities of stingless bees in urban areas. Front. Sustain. Cities 4:1103835. doi: 10.3389/frsc.2022.1103835
Received: 21 November 2022; Accepted: 15 December 2022;
Published: 06 January 2023.
Edited by:
Léo Correia da Rocha-Filho, Federal University of Uberlandia, BrazilReviewed by:
Leopoldo Alvarez, Universidad Nacional de La Plata, ArgentinaWilliam De Jesús May Itzá, Universidad Autónoma de Yucatán, Mexico
Copyright © 2023 Arena, Toppa, Martines and Alves-dos-Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana V. N. Arena,  bWFyaS5hcmVuYUB1c3AuYnI=
bWFyaS5hcmVuYUB1c3AuYnI=