
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Soil Sci., 03 June 2024
Sec. Soil Organic Matter Dynamics and Carbon Sequestration
Volume 4 - 2024 | https://doi.org/10.3389/fsoil.2024.1369971
This article is part of the Research TopicEffects of agricultural management on soil carbon sequestrationView all 3 articles
 Keiji Jindo1*
Keiji Jindo1* Omar El Aroussi2
Omar El Aroussi2 Joris de Vente3
Joris de Vente3 Jorge López Carratalá4
Jorge López Carratalá4 Felipe Bastida3
Felipe Bastida3 Carlos Garcia Izquierdo3Yoshito Sawada5
Carlos Garcia Izquierdo3Yoshito Sawada5 Travis L. Goron6
Travis L. Goron6 Gonzalo G. Barberá3
Gonzalo G. Barberá3Soil organic carbon (SOC) is essential in semi-arid agricultural land for enhancing soil health, particularly through the promotion of microbial activities. This study assessed the impact of different agronomic practices on soil properties, microbial communities, and SOC levels in semi-arid Moroccan wheat fields. Three treatments were investigated: eucalyptus (Eucalyptus spp.) companion planting (EU), and fallowing with harvest residue mulching (FA), with the latter involving both short (3 months; FAS) and long (15 months; FAL) fallow periods. The study revealed significant variation in soil characteristics and microbial communities between these agronomic management regimes. Notably, soils managed with FAL contained elevated SOC levels (1.2%) compared to other treatments (FAS and EU) which show lower SOC range (0.62–0.86%). Both labile C (water-soluble carbon) and recalcitrant C (humic substances) were increased by FAL. Additionally, soil microbial biomass and dehydrogenase activity were observed to be high in FAL-managed soils, along with increased levels of extracellular enzymes related to nutrient cycling (β-glucosidase, alkaline phosphatase, and urease). Phospholipid fatty acid (PLFA) analysis indicated positive correlation between carbon content in soils and microbial populations. In contrast, soils managed with EU had significantly lower SOC levels, possibly due to differences in carbon fractionation. FAL increased soil enzymatic activities and enriched the microbial community when compared to EU management. In conclusion, this study indicated the importance of fallowing and fallowing period for conservation of SOC, and potential to mitigate negative effects of biophysical constraints on agricultural productivity in semi-arid soils of Northwest Africa.
Cereal species, especially durum wheat (Triticum durum Desf.), are among the main crops of Morocco, and account for 62% of the nation’s production acreage (1, 2). However, much of the country is semi-arid with annual precipitation between 350 and 450 mm which often limits durum wheat yield (3, 4). Additionally, intensive cultivation has promoted soil erosion and degradation, evidenced by loss of soil structure and organic carbon content (5). Increasingly frequent extreme weather events exacerbated by climate change (e.g. drought, heat waves) can further deteriorate soil structure, negatively impacting cereal production in Morocco (6, 7). As a result, fields affected by the above processes are often low in soil organic matter (SOM). Such constraints are expected to become more common in future decades and hinder production in small and mid-sized farms, notably those in regions already struggling with a crop grain deficit (8).
SOM contributes to long-term agricultural sustainability by enhancing crop productivity and soil fertility (9). Increasing soil organic carbon (SOC, a major component of SOM) to optimal levels can improve yields through i) increased available water capacity, ii) increased plant-available nutrients, iii) improved soil structure, and iv) reduced soil erosion (10 and 11). However, semi-arid soils are often low in SOM. Additionally, inputs of SOM through animal manure on wheat fields among smallholder farmers in Morocco is uncommon – resources are often prioritized for fruits and other high-value crops (12). Soil erosion and salinization is also frequent, resulting in high salinity and electro-conductivity (EC), and low carbon content.
Land management practices exist in Morocco to prevent soil degradation, and aid in the restoration of soil fertility. For example, establishing Eucalyptus (Eucalyptus camaldulensis and Eucalyptus eomphocephala) trees as crop-companion plantations in semi-arid Moroccan fields is a well-known strategy to preserve important soil qualities (13), first promoted by the government in the 1950’s (14). In contrast, fallowing followed by mulching has had longer use in traditional, local farming systems. Fallowing and mulching can increase water content in coarse-textured soils (15), and reduce evapotranspiration, surface run-off, and weed proliferation (12, 16). Both practices can contribute to increased soil water, carbon, and nitrogen, which in turn may increase soil fertility by enhancing biomass production and SOM. However, different fallowing durations can result in altered SOM content depending on local biophysical conditions such as climate, soil type, and crop type (17, 18).
Carbon, and plant-available nitrogen released through decomposition of organic sources can strongly influence the soil microbiome, as microbial community structure is dependent on C:N ratio (19). Water-soluble carbon (WSC) which is mainly composed of labile and easily degradable compound may serve as an index of organic C mineralization (mainly composed of labile and easily-degradable compounds) (20, 21), while recalcitrant carbon contains humic substances (an essential part of OM) which contributes to soil fertility and health (22, 23). Measurement of these different nutrient pools can reveal effects of different land use management on the larger cropping system.
Soil microbial biomass and enzymatic activities are often correlated with nutrient availability (e.g. mineralization and immobilization) - soil biochemical analysis may therefore be used to examine effects of land use and management changes on soil and ecosystem quality (24). For instance, soil microbial biomass (SMB) and dehydrogenase can serve as evaluators for potential soil microbial activity (25–27). Other enzymatic indicators include β-glucosidase, involved in cellulose decomposition yielding glucose as a reaction product, a potential energy resource for microorganisms (28). Urease plays an essential role in the N-cycle through hydrolysis of urea, yielding ammonia and CO2 (29). Phosphatase converts unavailable organic P to mineral forms, which can then be taken up by plant roots (30). Phospholipid fatty acid (PLFA) analysis can also describe microbial community composition (27), and has been used for over two decades to characterize the soil microbiome in arid and semi-arid regions (31–34).
The aim of this study was to examine effects of two different management practices (eucalyptus companion cropping, EU; and fallowing with harvest residue mulching, FA) on soil chemical/biochemical properties and microbial diversity, measured through analysis of soil carbon fractionation, enzymatic activity, and microbial diversity. It was hypothesized that i) Fallowing/mulching and establishment of Eucalyptus plantations will have different effects on soil carbon fractionation, soil enzymatic activities, and microbial community; and ii) soil enzyme activities and microbial populations will be elevated in soils enriched with SOC by these different farm management practices.
Seven soil samples were collected from different durum wheat (Triticum turgidum var. L. durum) fields near Fez, Morocco (Latitude (dd): 34.03715, Longitude (dd): 4.998). To obtain a baseline of local soil degradation, a previously farmed area (now abandoned and uncultivated due to erosion and land degradation) in Blad Sadar was also sampled (DS, Table 1). Regional mean annual temperature and potential evapotranspiration between 1970 and 2011 at the Fez-Sais station was 16.9°C and 863 mm per year, respectively (35). Clay-loam is the dominant soil texture. Despite relatively poor moisture retention, light reddish siliceous soil (locally named “Hamri”) is typically used for wheat production in the area (36). Wheat fields in the region often contain large amounts of stones and gravel (37). Soil degradation is widespread due to dry conditions and intensive land use, accompanied by frequent ploughing and overgrazing. Average annual rain-fed wheat yield in the Fez region between 1989–2014 was 1.27 t ha-1 (https://www.yieldgap.org/accessed on 19 November 2022), with 180 growing days for wheat cultivation between December and January (https://www.yieldgap.org/accessed on 19 November, 2022). The rainy season begins in October and lasts until April, with most precipitation during December and February (38).
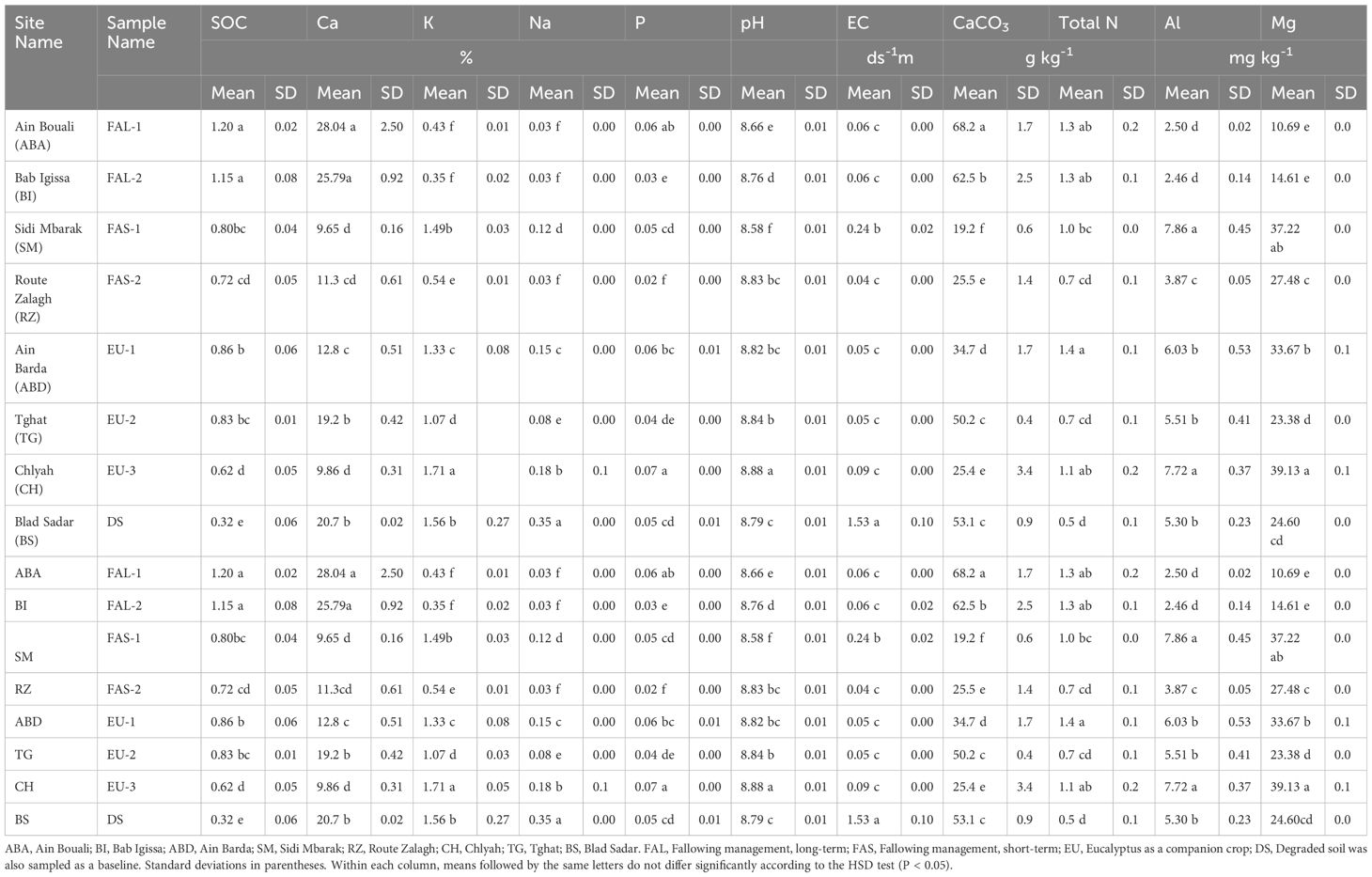
Table 1 Properties of soil sampled from durum wheat fields under differing agricultural management near Fez, Morocco.
Sampling was performed in July after durum wheat (Triticum turgidum var. L. durum) harvest. Soils were sampled between 5–30 cm. Soil samples were collected from within 2.5 km of Fez, at altitudes ranging from 60 to 300 m. Three different types of wheat field were sampled: i) those planted with Eucalyptus trees as a companion crop (EU); and those that were kept fallow after harvest residue mulching for either ii) long-term (15 months, FAL); or iii) short-term (3 months: FAS). The cropping system of all sampled fields was a rotation of wheat and barley. Three different EU locations were sampled, and two different FAL and FAS locations. Sample locations and names are described in the map of Fez region within the supplemental information. Six subsamples per sampling site were randomly collected from different points of the field, and then pooled. After removing plant and other debris, soil samples were air-dried for two days, sieved with a 2 mm screen, and stored at 4°C prior to laboratory analysis.
Electrical conductivity and pH were measured in a 1:10 (w/v) aqueous solution (39). Moisture content was obtained by weighing before and after drying at 105°C for 12 h. Organic matter (OM) concentration was determined by the loss on ignition method at 430°C for 24 h (20). Total carbon, and N were determined by an automatic C/N/S analyser (NA1500, Carlo Erba, Emmendingen, Germany). Macro and micro-nutrients, and heavy metal content was determined by inductively coupled plasma optical emission spectroscopy (ICP-OES; Thermo Scientific iCAP 6500, Waltham, MA). Water-soluble carbon (WSC) was determined in a 1:10 extract by a TOC analyser for liquid samples (Shimazdu 5050A, Kyoto, Japan) after shaking for 2 h and filtering through ashless filter paper (Albet 145 110). Water-soluble carbohydrates were assayed with a colorimetric method (40) from the same extract with 4 ml of 0.2% anthrone solution in concentrated sulphuric acid. Absorbance was measured in a spectrophotometer (Termo Electron Corporation Hexiosα). Soil humic fractions were quantified by measurement of the 0.1M Na4P2O7-extractable organic contents, via oxidation with K2Cr2O7 and spectrophotometric determination of Cr+3 at 590 nm (20).
Microbial biomass carbon was determined with the fumigation–extraction method, with extraction of organic C by K2SO4 (41), followed by measurement of extract C content with a TOC analyser (Shimadzu TOC-5050A). β-glucosidase activity was determined by colorimetric estimation of p-nitrophenol (PNP) formed by hydrolysis of p-nitorophenyl-β-d-glucopyranoside (PNG), as described by Eivazi and Tabatabai (42). The same method was used for the determination of alkaline phosphatase activity, with the substitution of PNG by p-nitrophenyl phosphatase as suggested by Tabatabai and Bremner (43). Urease activity was determined by the method of Kandeler and Gerber (44), estimated from ammonia release following incubation with urea in an alkaline buffer.
Phospholipids were extracted from 6 g of soil with a chloroform-methanol solution based on Bligh and Dyer (45), then fractionated and quantified using the procedure described by Frostegard et al. (46) and Bardgett et al. (47). Phospholipids were transformed into fatty acid methyl esters (FAMEs) by alkaline methanolysis, then quantified by gas chromatography (Trace GC Ultra, Thermo Scientific) through a 30 m capillary column (Thermo TR-FAME 30 m x 0.25 mm ID x 0.25 µm film) with helium as the carrier gas. Temperature was initially 150 °C for 0.5 min, then increased by 2 °C min-1 to 180 °C, and then by 4 °C min-1 to 240 °C. The fatty acids 10:0, 11:0, 12:0, 13:0, 14:0, i15:0, a15:0, 15:0, i16:0, i17:0, C18:3n3, cy17:0, and cy19:0 were selected to represent bacterial biomass (46–48), and 18:2ω6 was measured to indicate fungal biomass (49, 50). The ratio of bacterial to fungal PLFAs represents the ratio between bacterial and fungal biomass (47). The Gram+ specific fatty acids i15:0, a15:0, i16:0, i17:0, and C18:3n3, and Gram- specific fatty acids 14:1, cy17:0 and cy19:0 were recorded as a measure of the ratio between Gram+ and Gram- bacterial biomass. The ratio of monounsaturated PLFAs to saturated PLFAs is expressed as mono/sat. All results are given in nmol g-1.
All results of soil chemical properties, soil enzymatic activities, and PLFAs were reported as means from one-way ANOVA, followed by Tukey’s HSD test (HSD, honestly significant difference at the 95% confidence interval). Error bars represent standard deviation. Physico-chemical parameters, carbon fractions, biomass indicators, enzymatic activities, and the relative abundances of all identified FAMEs of sampled soils were subjected to correlation analysis to assess if measured soil properties differed between different management practices. All statistical analyses were conducted in R (51). The authors have defined the p level for each statistical test for scientific clarity.
Soil characteristics are recorded in Table 1. All sampled soils were basic in pH (8.5- 8.9) with elevated EC. Calcium was the dominant cation element in all samples. Soils from fields under long-duration fallowing management (FAL-1 and FAL-2) contained significantly higher SOC than the other sampled soils (Table 1). Lower total N was observed in soils under short-term fallowing management (FAS-1 and FAS-2) than long-term fallowing management (FAL-1 and FAL-s2) (Table 1). One of fields with Eucalyptus (EU-2) has the lowest total N (0.7 g kg-1) together with FAS-2. Degraded soil (DS) from abandoned areas were sampled as a baseline, and contained the lowest SOC and total N, while EC and Na+ were the highest of all sampled soils (Table 1).
Carbon fractions were significantly influenced by agricultural management (Figure 1). Long-term fallow management (FAL-1 and FAL-2) soils were high in labile carbon parameters (1750–2325 mg kg-1 for water-soluble carbon and 110 mg kg-1 for carbohydrate). On the contrary, soils from fields with Eucalyptus (EU-3) had the lowest contents (748.8 mg kg-1 for water-soluble carbon and 27.9 mg kg-1 for carbohydrate). Both FAL-1 and FAL-2 contained higher organic carbon content (1.15–1.2%) than the other sites (Table 1), and high amounts of recalcitrant humic C (Figure 1). The FAL-1 soil had the highest humic C content (2.76 mg kg-1)(Figure 1), and total organic carbon content of the different soil samples (Table 1).
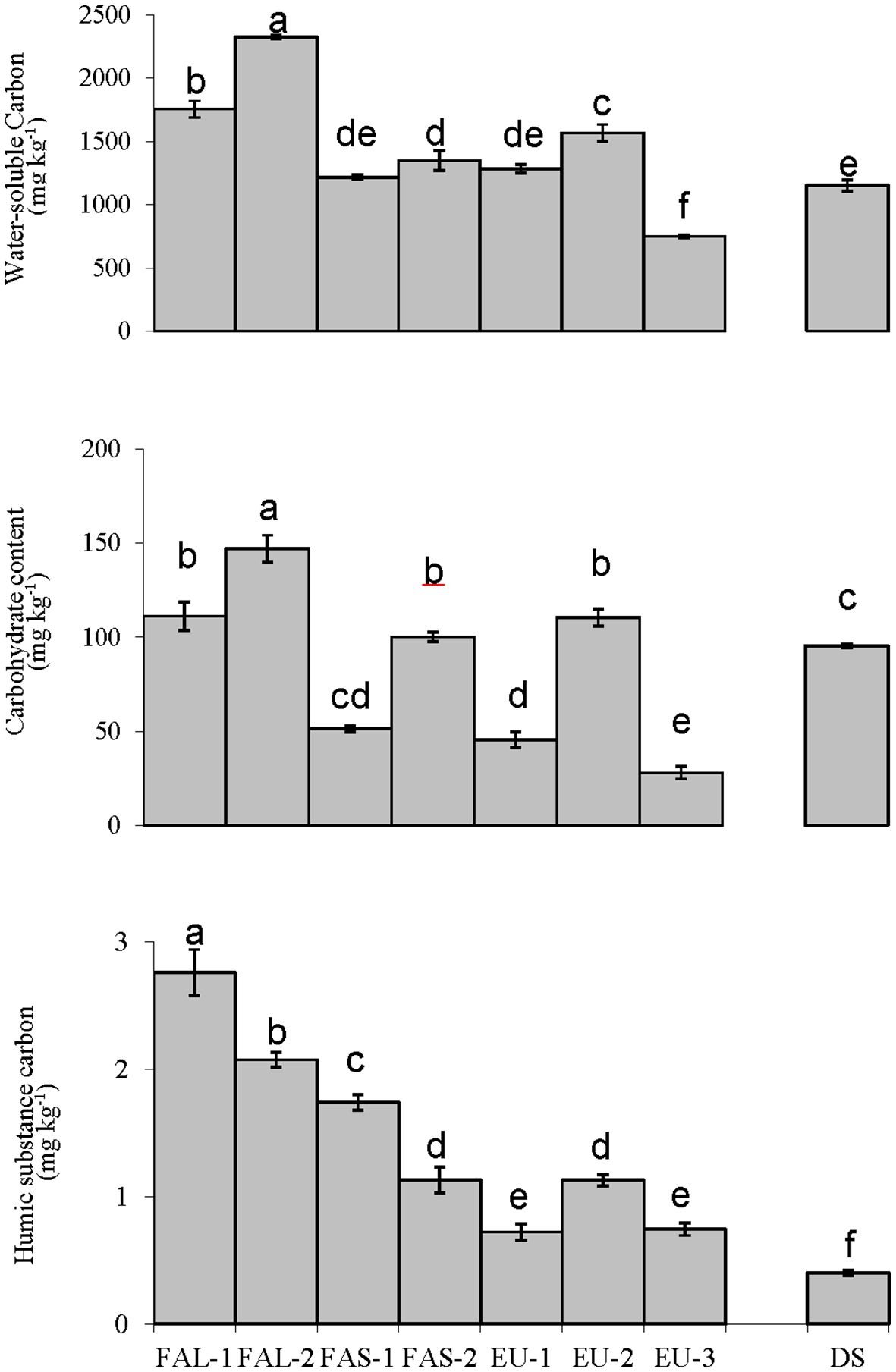
Figure 1 Water-soluble carbon, carbohydrate, and humic substance carbon in soils sampled from wheat fields treated with fallowing for a long-term (15 months; FAL), short-term (3 months; FAS), or companion- cropped with Eucalyptus (EU). A degraded, abandoned field was sampled as a baseline control (DS). Different letters represent significant difference determined by Tukey's HSD test, at a 95% confidence interval. Bars represent standard deviation.
Concerning general microbial parameters (Figure 2), the highest value of dehydrogenase (2.2 μg INTF g-1h-1) was recorded in FAL-2, followed by EU-2 and FAL-1 (1.76 and 1.45 μg INTF g-1h-1, respectively). Microbial biomass carbon was also greatest (237.26 mg kg-1) in FAL-2, followed by FAL-1 and EU-1 (228.60 and 183.87 mg kg-1). Regarding extracellular enzymatic activities, Soils of FAL-2 and EU-1 showed high β-glucosidase activity (0.874 and 0.818 μg PNP g-1h-1, respectively). The highest alkaline phosphatase activity was recorded in EU-1 (2.53 μg PNP g-1h-1). The greatest urease activity was observed in FAL-2 (3.52 μg NH+4-N g-1h-1) in Figure 3. The lowest activities of β-glucosidase (0.02 μg PNP g-1h-1) and alkaline phosphatase (0.987 μg PNP g-1h-1) are seen in FAS-2, while the lowest urease activity (0.26 NH+4-N g-1h-1) is observed in EU-3. And those two soils have relatively low carbon content (Table 1). The shortage of OC in the degraded soil baseline (DS) was congruent with lowest activities of all three specific enzymes (β-glucosidase: 0.13 μg PNP g-1h-1, alkaline phosphatase: 0.10 μg PNP g-1h-1; urease: 0.02 NH4-N g-1h-1) as well as markers of general microbial activities (SBC and dehydrogenase).
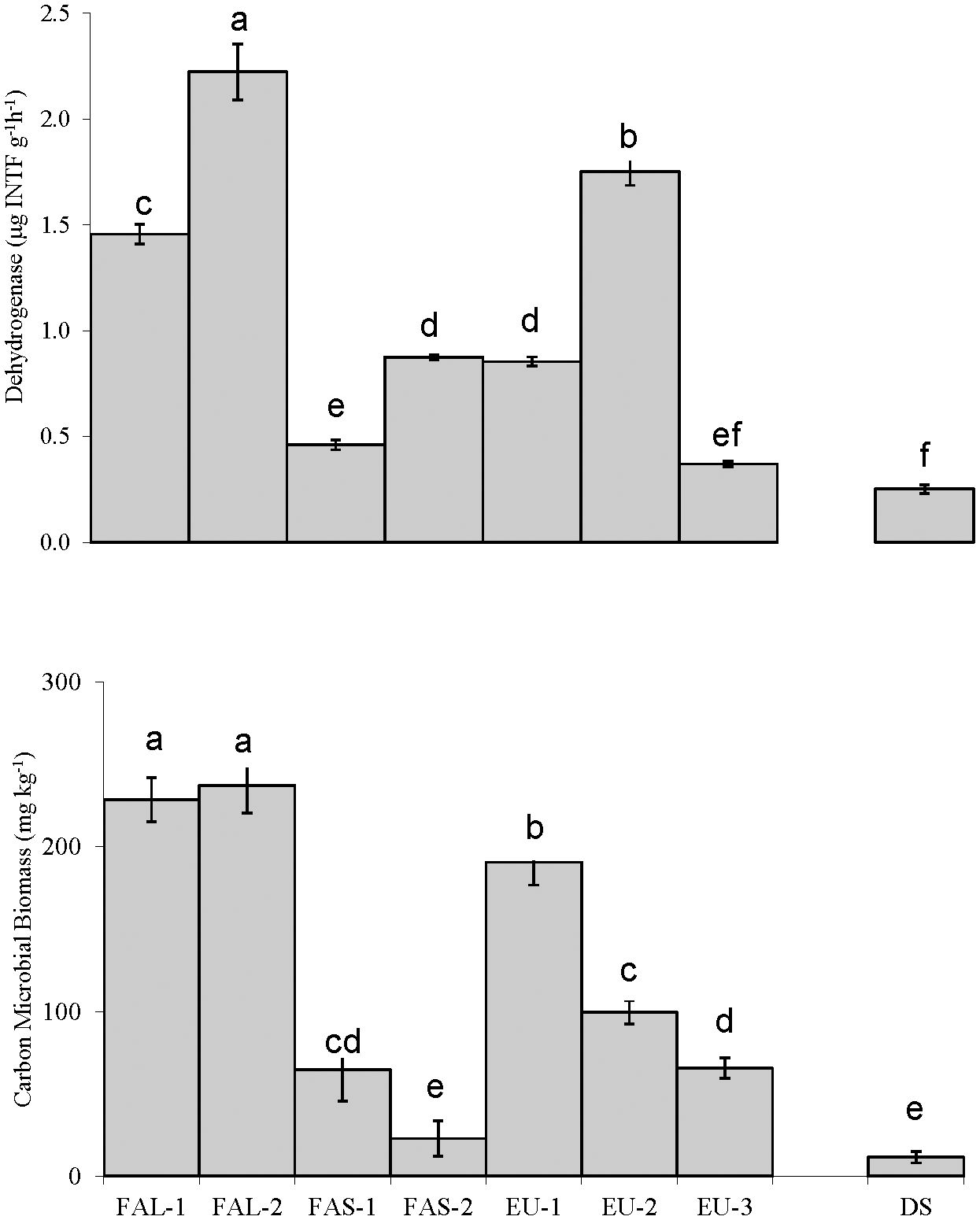
Figure 2 Microbial biomass carbon and dehydrogenase activity in soils taken from wheat fields treated with fallowing for a long-term (15 months; FAL), short-term (3 months; FAS), or companion-cropped with Eucalyptus (EU). A degraded, abandoned field was sampled as a baseline control (DS). A degraded, abandoned field was sampled as a baseline control (DS). Different letters represent significant difference determined by Tukey's HSD test, at a 95% confidence interval. Bars represent standard deviation.
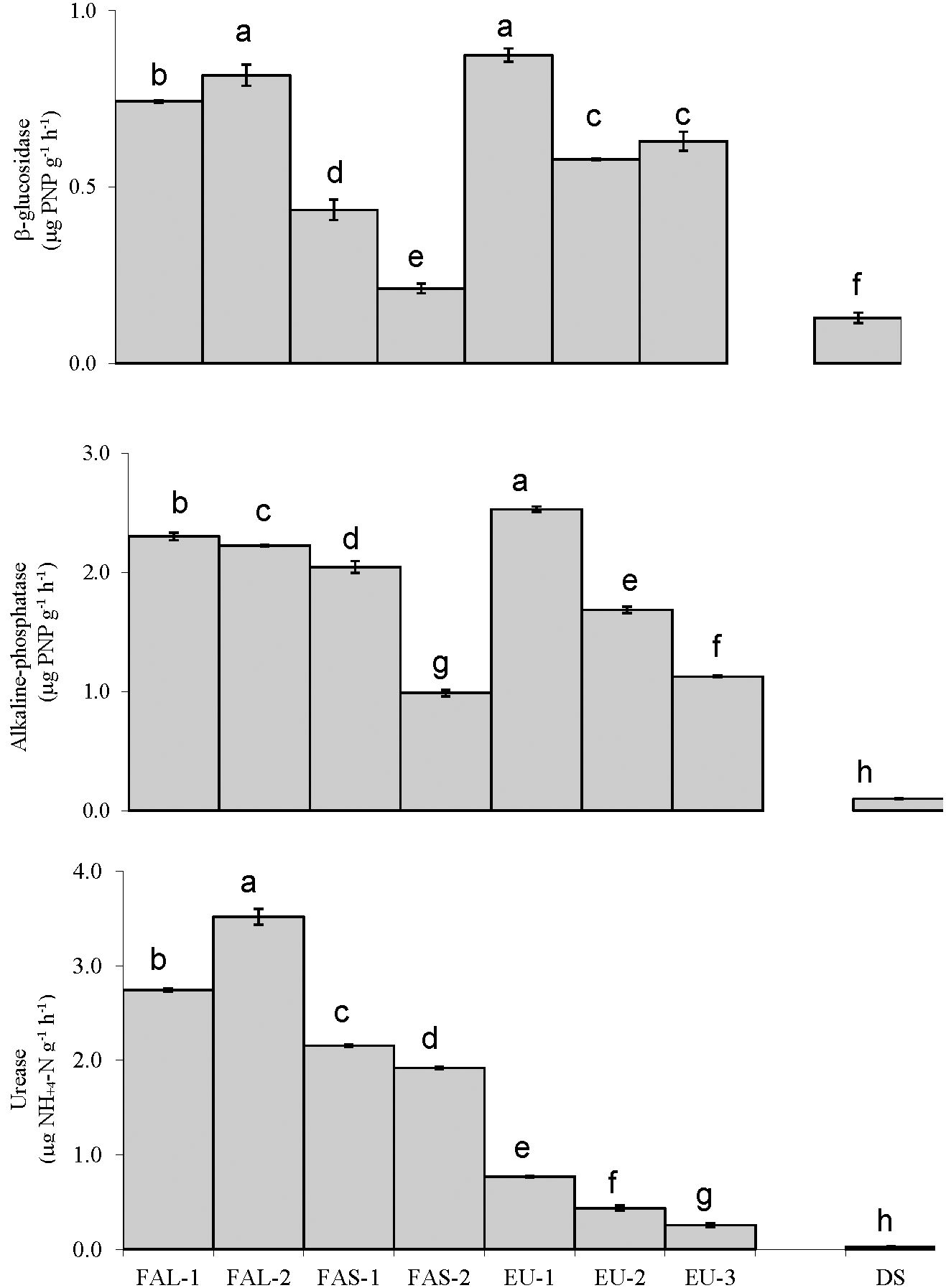
Figure 3 Urease activity, alkaline phosphatase activity, and B-glucosidase activity of soils sampled from wheat fields treated with fallowing for long-term (FAL), short-term (FAS), and Eucalyptus plantation (EU). A degraded, abandoned field was sampled as a baseline control (DS). Different letters represent significant difference determined by Tukey's HSD test, at a 95% confidence interval. Bars represent standard deviation.
Different management resulted in significantly different microbial biomass (Table 2). Generally, soils with higher carbon content (SOC > 1 g 100 g-1) such as FAL-1 and FAL-2, maintained high abundance of bacteria, fungi, and total microbes. These two soils also displayed higher total nitrogen content (1.3 g kg-1). In contrast, soils with low organic carbon (SOC < 0.75 g 100 g-1) such as FAS-1, FAS-2 and EU-3 had lower microbial abundances (Bacteria: 15.3–19.6 nmol g soil-1, Fungi: 0.5–0.8 nmol g soil-1). Bacteria were more abundant than fungi in all sampled soils. Lower populations of Gram- bacteria (1.1–1.2 nmol g soil-1) were observed in soils with low SOC (FAS-2, EU-1 and EU-3). The highest ratio of monounsaturated to saturated fatty acids (0.250) was observed in FAL-2.
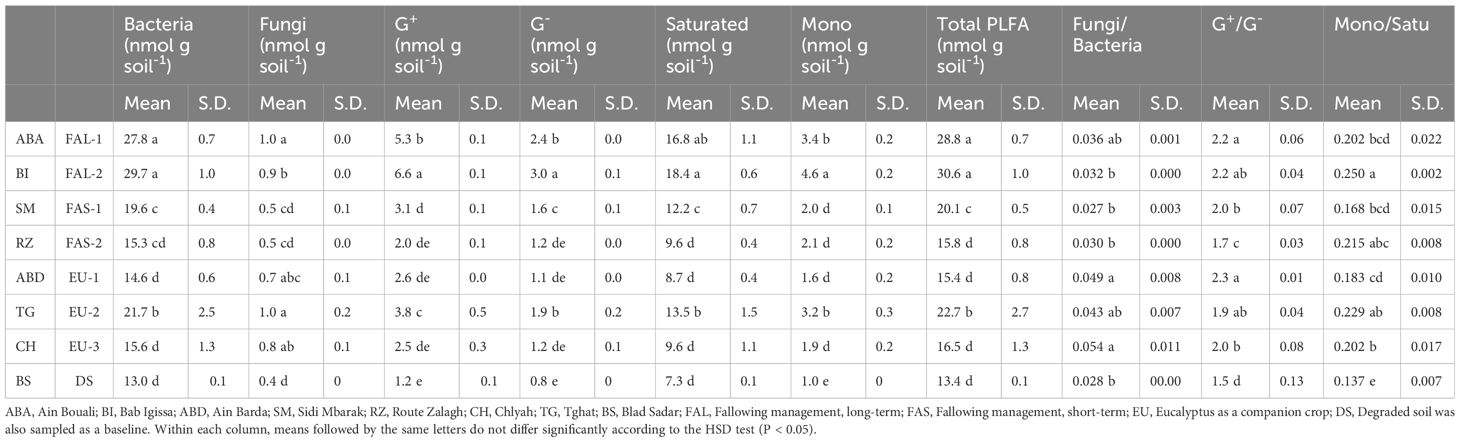
Table 2 Concentrations of bacteria, fungi, Gram positive bacteria (G+), Gram negative bacteria (G-); saturated, monounsaturated, total fatty acids; fungi/bacteria, G+/G-, and saturated/monounsaturated ratios within soils sampled from wheat fields near Fez, Morocco.
The degraded baseline soil (DS) contained the lowest levels of fatty acids (13.4 nmol g soil-1)), and the lowest abundance of all microbe classifications (Bacteria:13.0 nmol g soil-1, Fungi:0.4 nmol g soil-1. Gram+: 1.2 nmol g soil-1, Gram-: 0.8 nmol g soil-1, Saturated: 7.3 nmol g soil-1; Monosaturated: 1.0 nmol g soil-1). Furthermore, the lowest ratios of monounsaturated: saturated fatty acids (0.137), and Gram+/Gram- ratio (1.5) were observed in the DS soil sample.
A correlation matrix allows summaries of relationships between nutrient content and microbial communities across different land uses. Within sampled soils, SOC was positively correlated (Pearson correlation efficient (r.) =0.819) with total N (Figure 4). General microbial markers (SMB and dehydrogenase activity) were negatively correlated with EC [(r. =-0.483 and -0.492, respectively) (Figure 4), indicating a stressful circumstance for microbial growth. Negative correlation (p <0.05) was observed between EC and different parameters of microbial communities (e.g. fungal biomass (r. =-0.574)), enzymatic activities (e.g. β-glucosidase (r. = -0.646), alkaline phosphatase (r.=-0.727)) and carbon fractions (e.g. humic substances (r.=-0.554). In contrast, SOC was found to be positively correlated with all other variables, except for the ratio of fungi and bacteria. Nitrogen content was also positively correlated with high abundance of different microbial communities and enzymatic activities.
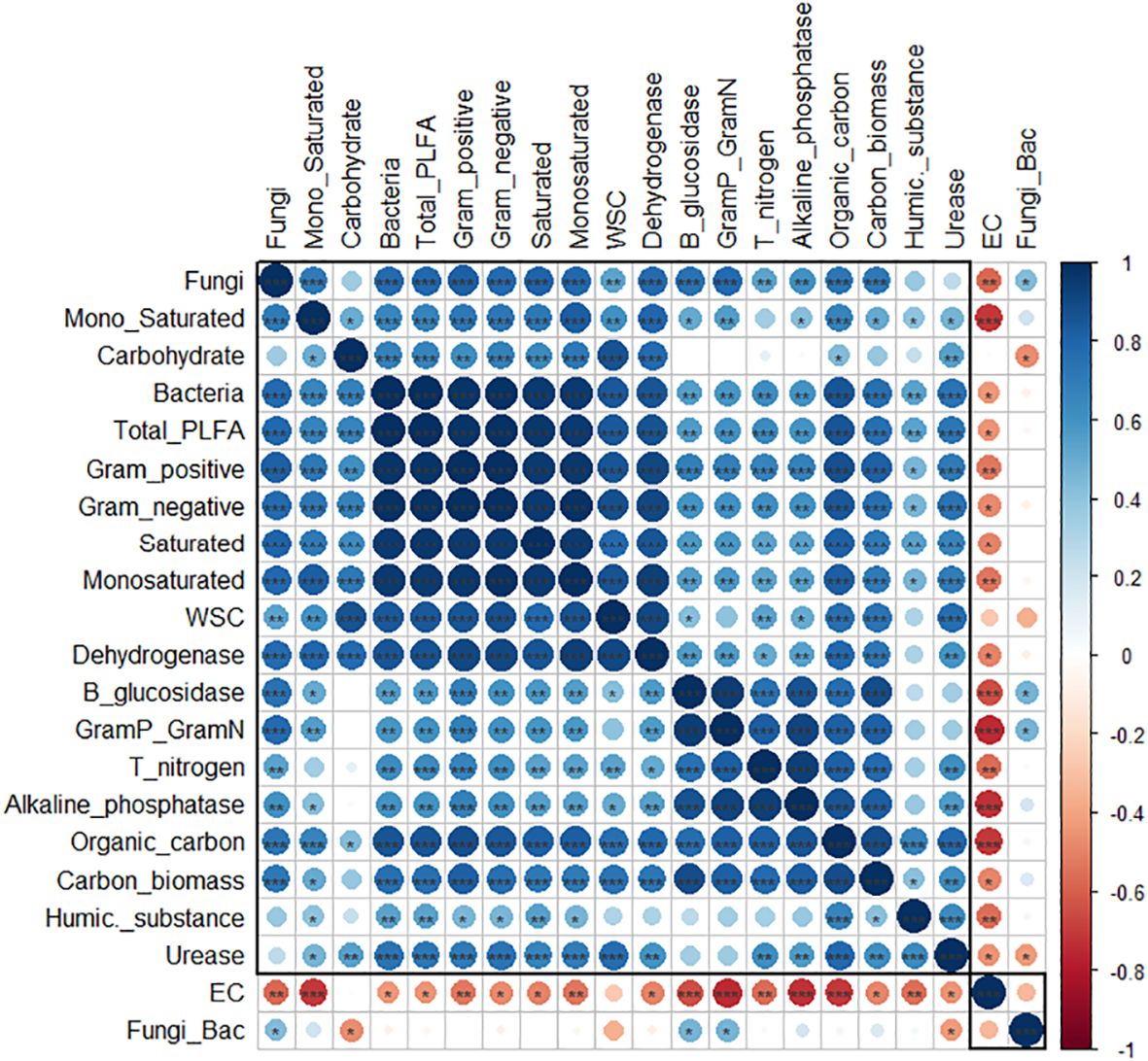
Figure 4 Pearson correlation heat-map matrix of soil properties in sampled wheat fields. Blue and red circles represent significant positive and negative correlation, respectively (p<0.05). Blank cells represent no significant correlation. Asterisks indicate the significance in the correlations (*, **, ***P<0.05, 0.01, 0.0001, respectively). Rectangles around the plot of correlation matrix are considered as clusters based on the results of hierarchical clustering. Organic carbon (Organic_carbon); Total nitrogen (T_nitrogen); Water-soluble carbon (WSC); Total amount of Phospholipid-derived fatty acids (Total PLFA); Ratio of fungi and bacteria (Fung_Bac); Ratio of monounsaturated fatty acids/saturated fatty acids (Mono_Saturated); Ratio of Gram positive/Gram negative bacteria (GramP_GramN).
Eighty percent of agricultural land in Morocco is rain-fed (52), and sustainable yet productive cultivation practices are needed to allow reliable food production for an increasing population (8). Increased resiliency against drought, soil degradation (53), and extreme weather events is also required. Previous work with crop simulation modelling (54) has indicated that improved soil moisture will be needed in Morocco to reduce the gap of 5.35 t/ha between potential and realized yields. Different crop rotation management and agricultural practices can be utilized to improve soil fertility, water holding capacity, and crop yield (54). Several studies report implementation of techniques within Morocco such as reduced tillage, soil cover and crop rotation (12, 55), conducive for conservation agriculture. According to El-Shater and Yigezu (12), retention of harvest residue in the field could increase wheat yields by as much as 30% through increased rain infiltration and suppression of weeds.
This study reports how different land management practices in wheat fields affect biologically-available soil nutrients (C and N), microbial communities, and enzymatic activities. Higher SOC (SOC > 1.1%) was observed in the long-term fallow management treatment (FAL-1 and FAL-2), and these soils also conserved abundance of different microbial communities despite semi-arid conditions, due to the availability of different microbial substrates. Total water-soluble carbon (WSC) is considered an index of organic C mineralization and primarily composed of labile and easily degradable compounds (20), while carbohydrates represent energy readily available for microorganisms (56, 57). The labile SOC, such as the WSC and carbohydrate fractions correlated with increased active microbial populations as observed by dehydrogenase activity (58). Carbon and nitrogen content can greatly influence microbial dynamics in semi-arid and arid regions, as the emergence of a new community structure is dependent on soil C to N ratio (19, 59).
Bacterial abundance was higher than that of fungi in all samples, perhaps due to lower fungal tolerance against regional stress including high temperatures and alkaline soils (60). Gram- bacteria are less resistant against severe environmental conditions (61 and 62) perhaps due to structural differences, including the absence of a peptidoglycan layer. In this study, low abundances of Gram– bacteria were detected in soils with low SOC (FAS-2 and EU-3). The lowest Gram+/Gram- ratio in the DS field may be due to low nutrient (C, N) availability (63). The low-SOC soils also displayed low enzymatic activities and low soil microbial abundance, similar to the high-salinity DS soil. The lowest Gram+/Gram- ratio in DS (Table 2) is probably due to low nutrient availability (63). The lowest monounsaturated to saturated fatty acid ratio was observed in DS (Table 2), strongly corresponding to limited nutrients and high salinization as a consequence of soil degradation. This ratio may serve as a stress indicator (64, 65), and lower ratios are often recorded in microbial communities inhabiting environments where SOC and/or nutrients are limiting (66). Due to previous field management practices, the DS soil may have become degraded over time resulting in soils unsuitable for crop production (67).
Land use and management can strongly affect soil microbial biomass (9, 68 and 69) and enzymatic activities (27). Concerning hydrolytic activities, β-glucosidase interacts with plant and microbial cellulose decomposition processes. Glucose is a reaction product and can serve as an energy source for microorganisms (28, 70). Urease plays an essential role in the N-cycle, hydrolyzing urea to yield ammonia and CO2 (29). Phosphatase is important for the mineralization of organic P (30, 71). Phosphatase release (induction) by soil microbes and the capacity to solubilize unavailable phosphate may be agriculturally advantageous, as a global phosphate shortage is predicted within the next 50–100 years (72). These extracellular enzyme activities can be altered by several factors, including land management practices and the physico-chemical parameters of the soil-plant interface within the microbial habitat (57).
In this study, soils managed with long-term fallowing (FAL-1 and FAL-2) were enriched with SOC and displayed high activities of the three enzymes described above (Figure 3). In contrast, soils with low carbon content (such as FAS-2) displayed low activities. As all three activities are correlated positively with SOC and TN (Figure 4), this result may be a response to high or low amounts of substrate within the soil. In comparison, the critical shortage of OC in the degraded soil (DS) resulted in lowest activities of all three enzymes, as well as lowest general microbial activity (SBC and dehydrogenase). There was also negative correlation between markers of general microbial activities (SMB and dehydrogenase activity) and EC (Figure 4). This may indicate a severely limited environment for microbial growth, as high EC and low SOC can trigger osmotic stress (70), and is typically problematic in semi-arid and arid agricultural conditions.
Fallowing contributes not only to the accumulation of SOC,but can also increase levels of other nutrients (73) and water conservation by increasing soil aggregation, which in turn enhances water-holding capacity and reduces water run-off (15, 18, 74). Fallowing of longer durations is preferable for the improvement of soil fertility (18). However, long-term fallowing is not always feasible under certain circumstances (e.g., increasing human population pressure and land scarcity).
In Morocco, a harsh climate and population expansion require crop yields which may exceed the potential of current systems, and necessitate improvement of soil fertility (53). According to previous work comparing different fallowing lengths (17, 75, 76), durations of two years still can result in significant gains. However, optimizing the fallow length for soil properties and crop production should be considered based on various biophysical factors such as soil types, crop types, and climate conditions (77–81) as well as farmer’s livelihood (76).
In the case of the current study, soil treated by a year-long fallowing period (FAL-2) had the highest humic C fraction (Figure 1), in accordance with the highest organic carbon content of the different soils sampled (Table 1, statistically equivalent to FAL-1). Fallow vegetation and harvest residue require time to decompose, simultaneously replenishing a portion of the nutrients removed by the crop and protecting the soil against erosion.
Compared to soils managed with fallowing, soils taken from wheat fields mixed with Eucalyptus trees generally contained lower amounts of carbon, enzymatic activities, and microbial communities. The Moroccan Forest Service began to encourage the establishment of Eucalyptus plantations in 1949 - Eucalyptus species now represent 40% of the nation’s total reforested area (14). Mixing farming with trees and crops is common in Morocco (82). Integrating a crop with Eucalyptus trees can modify the microclimate (e.g. reduced air temperature and net radiation distribution), resulting in conditions more favorable to wheat growth by reducing stress during the post-anthesis period (83, 84). This practice can also contribute to soil multi-functionality for primary production, water regulation, climate regulation, soil biodiversity, and nutrient cycling (24). However, local agroecological knowledge about tree-based diversification was scarce among farmers in the study area (82). Furthermore, farmers are mostly not interested in woody species locally classified as “wild trees” including Eucalyptus, as production requires costly permits from the forest authority (82). Additional disadvantages of integrating crops and Eucalyptus include possible reduction of soil moisture and nutrients due to competition (85).
The results of our study indicate that EU management was not associated with elevated soil organic carbon and soil microbial communities. Future studies should additionally assess other metrics including crop yield, use efficiency of resources (e.g. nutrient, light and water), and cost-benefit ratio for calculation of return-on-investment.
Fields managed with long-term fallowing resulted in high SOC with conserved abundance of microbial communities, due to increased availability of nutritional substrates (C and N). In contrast, lower SOC was found in soils managed with short-term fallowing, which displayed low enzymatic activities and microbial populations. Severe biophysical stresses such as low rainfall and high salinization can reduce availability of soil microbial nutrients and hence soil microbial populations. It is essential to identify optimum farming practices for enhancing the productivity in semi-arid soils, and for protection of these soils against degradation. Besides long-term fallowing and Eucalyptus companion planting, agronomic practices such as no-tillage, crop rotation, and compost amendments should also be investigated, which may reinforce SOC, and microbial populations.
Access to the dataset is restricted to authorized individuals or organizations with a legitimate research purpose. Requests to access the datasets should be directed to keiji.jindo@wur.nl.
KJ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. OE: Data curation, Writing – review & editing. JV: Writing – review & editing. JL: Writing – review & editing. FB: Data curation, Writing – review & editing. CG: Writing – review & editing. YS: Writing – review & editing. TG: Writing – review & editing. GG: Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. KJ wishes to acknowledge financial support (3710473400).
We thank Juan Manuel Quiñonero Rubio for the data collection and revision.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsoil.2024.1369971/full#supplementary-material
1. Karrou M, Oweis T. Assessment of the severity and impact of drought spells on rainfed cereals in Morocco. Afr J Agric Res. (2014) 9:3519–30. doi: 10.5897/2014.9130
2. Bartali EH, Boutfirass M, Yigezu YA, Niane AA, Boughlala M, Belmakki M, et al. Estimates of food losses and wastes at each node of the wheat value chain in Morocco: Implications on food and energy security, natural resources, and greenhouse gas emissions. Sustainability. (2022) 14:16561. doi: 10.3390/su142416561
3. Aker J, Lemtouni A. A framework for assessing food security in the face of globalization: The case of Morocco. Agroalimentaria. (1999) 8:13–26.
4. Bregaglio S, Frasso N, Pagani V, Stella T, Francone C, Cappelli G, et al. New multi-model approach gives good estimations of wheat yield under semi-arid climate in Morocco. Agron Sustain Dev. (2015) 35:157–67. doi: 10.1007/s13593–014-0225–6
5. Chikhaoui M, Bonn F, Bokoye AI, Merzouk A. A spectral index for land degradation mapping using ASTER data: application to a semi-arid Mediterranean catchment. Int J Appl Earth Observ Geoinfo. (2005) 7:140–53. doi: 10.1016/j.jag.2005.01.002
6. Lehmann J, Kretschmer M, Schauberger B, Wechsung F. Potential for early forecast of Moroccan wheat yields based on climatic drivers. Geophysical Res Lett. (2020) 47. doi: 10.1029/2020GL087516
7. Kessabi R, Hanchane M, Krakauer NY, Aboubi I, El Kassioui J, El Khazzan B. Annual, seasonal, and monthly rainfall trend analysis through non-parametric tests in the Sebou River Basin (SRB), Northern Morocco. Climate. (2022) 10:170. doi: 10.3390/cli10110170
8. Amiri N, Lahlali R, Amiri S, El Jarroudi M, Khebiza MY, Messouli M. Development of an integrated model to assess the impact of agricultural practices and land use on agricultural production in Morocco under climate stress over the next twenty years. Sustainability. (2021) 13:11943. doi: 10.3390/su132111943
9. Obalum SE, Chibuike GU, Peth S, Ouyang Y. Soil organic matter as sole indicator of soil degradation. Environ Monit Assess. (2017) 189. doi: 10.1007/s10661-017-5881-y
10. Lal R. Managing soils and ecosystems for mitigating anthropogenic carbon emissions and advancing global food security. Bio Sci. (2010) 60:708–21. doi: 10.1525/bio.2010.60.9.8
11. Lal R. Sequestering carbon in soils of agro-ecosystems. Food Policy. (2011) 36:33–9. doi: 10.1016/j.foodpol.2010.12.001
12. El-Shater T, Yigezu YA. Can retention of crop residues on the field be justified on socioeconomic grounds? A case study from the mixed crop-livestock production systems of the Moroccan drylands. Agronomy. (2021) 11:1465. doi: 10.3390/agronomy11081465
13. Boulmane M, Oubrahim H, Halim M, Bakker MR, Augusto L. The potential of Eucalyptus plantations to restore degraded soils in semi-arid Morocco (NW Africa). Ann For Sci. (2017) 74:57. doi: 10.1007/s13595-017-0652-z
14. Khattabi A. Socio-economic importance of eucalyptus plantations in Morocco. In: Yoshimoto A, Yukutake K, editors. Global concerns for forest resource utilization, vol. 62 . Springer, Dordrecht (1999). doi: 10.1007/978–94-017–6397-4_7
15. Adil M, Zhang S, Wang J, Shah AN, Tanveer M, Fiaz S. Effects of fallow management practices on soil water, crop yield and water use efficiency in winter wheat monoculture System: A Meta-Analysis. Front Plant Sci. (2022) 13:825309. doi: 10.3389/fpls.2022.825309
16. Aboudrare A, Debaeke P, Bouaziz A, Chekli H. Effects of soil tillage and fallow management on soil water storage and sunflower production in a semi-arid Mediterranean climate. Agric Water Manage. (2006) 83. doi: 10.1016/j.agwat.2005.12.00183–196
17. Tian G, Kang BT, Kolawole GO, Idinoba P, Salako FK. Long-term effects of fallow systems and lengths on crop production and soil fertility maintenance in West Africa. Nutr Cycl Agroecosyst. (2005) 71:139–50. doi: 10.1007/s10705-004-1927-y
18. Partey ST, Zougmoré RB, Ouédraogo M, Thevathasan NV. Why promote improved fallows as a climate-smart agroforestry technology in Sub-Saharan Africa? Sustainability. (2017) 9:1887. doi: 10.3390/su9111887
19. Ford DJ, Cookson WR, Adams MA, Grierson PF. Role of soil drying in nitrogen mineralization and microbial community function in semi-arid grasslands of north-west Australia. Soil Biol Biochem. (2007) 39:1557–69. doi: 10.1016/j.soilbio.2007.01.014
20. García C, Hernández T, Costa F, Ceccanti B. Biochemical parameters in soils regenerated by the addition of organic wastes. Waste Manage Res. (1994) 12:457–66. doi: 10.1177/0734242X9401200602
21. Surey R, Schimpf CM, Sauheitl L, Mueller CW, Rummel PS, Dittert K, et al. Potential denitrification stimulated by water-soluble organic carbon from plant residues during initial decomposition. Soil Biol Biochem. (2020) 147:107841. doi: 10.1016/j.soilbio.2020.107841
22. Stevenson F,J. Humus chemistry: Genesis, composition, reactions. 2nd ed. New York: John Wiley and Sons (1994).
23. Schaeffer A, Nannipieri P, Kästner M, Schmidt B, Botterweck J. From humic substances to soil organic matter–microbial contributions. In honour of Konrad Haider and James P. Martin for their outstanding research contribution to soil science. J Soils Sediments. (2015) 15:1865–81. doi: 10.1007/s11368–015-1177–4
24. Zwetsloot MJ, van Leeuwen J, Hemerik L, Martens H, Simó Josa I, Van de Broek M, et al. Soil multifunctionality: Synergies and trade-offs across European climatic zones and land uses. Eur J Soil Sci. (2020) 72:1640–54. doi: 10.1111/ejss.13051
25. Beyer L, Wachendorf C, Elsner CD, Knabe R. Suitability of dehydrogenase activity assay as an indexo f soil biolocigical activity. Biol Fertil. Soils. (1993) 16:52–6. doi: 10.1007/BF00336515
26. Garcia C, Hernandez T, Costa F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun Soil Sci Plant Anal. (1997) 28:123–34. doi: 10.1080/00103629709369777
27. van Leeuwen JP, Djukic I, Bloem J, Lehtinen T, Hemerik L, Ruiter PC, et al. Effects of land use on soil microbial biomass, activity and community structure at different soil depths in the Danube floodplain. Eur J Soil Biol. (2017) 79:14–20. doi: 10.1016/j.ejsobi.2017.02.001
28. Tabatabai MA. Soil enzymes. In: Dick WA, editor. Methods of soil analysis. Part 2. Microbiological and biochemical properties, vol. 5 United States: ACSESS (1994). p. 775–833. Soil Sci. Soc. Am. doi: 10.2136/sssabookser5.2
29. Benitez E, Melgar R, Nogales R. Estimating soil resilience to a toxic organic waste by measuring enzyme activities. Soil Biol Biochem. (2004) 36:1615–23. doi: 10.1016/j.soilbio.2004.07.014
30. Nannipieri P, Pedrazzini F, Arcara PG, Piovanelli C. Changes in amino acids, enzymatic activities and biomasses during soil microbial growth. Soil Sci. (1979) 127:26–34. doi: 10.1016/0038–0717(83)90032–9
31. Steinberger Y, Zelles L, Bai QY, Lützow M, Munch JC. Phospholipid fatty acid profiles as indicators for microbial community structure and biodiversity in soils along a climatic transect in the Judean desert. Biol Fertil. Soils. (1999) 28:292–300. doi: 10.1007/s003740050496
32. Bessam F, Mrabet R. Long-term changes in soil organic matter under conventional tillage and no-tillage systems in semiarid Morocco. Soil Use Manage. (2003) 19:139–43. doi: 10.1079/SUM2003181
33. Sadiki A, Faleh A, Navas A, Bouhlassa S. Assessing soil erosion and control factors by the radiometric technique in the Boussouab catchment, Eastern Rif, Morocco. Catena. (2007) 71:13–20. doi: 10.1016/j.catena.2006.10.003
34. Diedhiou S, Dossab EL, Badianec AN, Diedhioud I, Sènee M, Dickf RP. Decomposition and spatial microbial heterogeneity associated with native shrubs in soils of agroecosystems in semi-arid Senegal. Pedobiologia. (2009) 52:273–86. doi: 10.1016/j.pedobi.2008.11.002
35. Bouizrou I, Chahinian N, Perrin JL, Müller R, Rais N. Network representation in hydrological modelling on urban catchments in data-scarce contexts: A case study on the Oued Fez catchment (Morocco). J Hydrology: Regional Stud. (2021) 34:100800. doi: 10.1016/j.ejrh.2021.100800
36. El Amri N, Errachidi F, Bour A, Bouhaddaoui S, Chabir R. Morphological and nutritional properties of Moroccan Capparis spinosa Seeds. Sci World J. (2019) 2019:8594820. doi: 10.1155/2019/8594820
37. El Allaoui N, Yahyaoui H, Douira A, Benbouazza A, Ferrahi M, Achbani EH, et al. Assessment of the impacts of plant growth-promoting micro-organisms on potato farming in different climatic conditions in Morocco. Microbiol Res. (2023) 14:2090–104. doi: 10.3390/microbiolres14040141
38. Achli S, Epule TE, Dhiba D, Chehbouni A, Er-Raki S. Vulnerability of barley, maize, and wheat yields to variations in growing season precipitation in Morocco. Appl Sci. (2022) 12:3407. doi: 10.3390/app1207340
39. Herrero J, Weindorf DC, Castañeda C. Two fixed ratio dilutions for soil salinity monitoring in hypersaline wetlands. PLoS One. (2015) 10(5):e0126493. doi: 10.1371/journal.pone.0126493
40. Brink RH, Dubar P, Lindch DL. Measurement of carbohydrates in soil hydrolysates with anthrone. Soil Sci. (1960) 89:157–66. doi: 10.1097/00010694-196003000-00006
41. Vance ED, Brookes PC, Jenkinson DS. An extraction method for measuring soil microbial biomass C. Soil Biol Biochem. (1987) 19:703–7. doi: 10.1016/0038–0717(87)90052–6
42. Eivazi F, Tabatabai MA. Glucosidases and galactosidases in soils. Soil Biol Biochem. (1988) 20:601–6. doi: 10.1016/0038–0717(88)90141–1
43. Tabatabai MA, Bremner JM. Use of p-nitrophenil phosphate for assay of soil phosphatase activity. Biol Fertil. Soil. (1969) 1:301–7. doi: 10.1016/0038–0717(69)90012–1
44. Kandeler E, Gerber H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol Fertil. Soil. (1988) 6:68–72. doi: 10.1007/BF00257924
45. Blig EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem And Physiol. (1959) 37:911–7. doi: 10.1139/o59–099
46. Frostegard A, Tunlid A, Baath E. Phospholipid fatty-acid composition, biomass, and activity of microbial communities from 2 soils types experimentally exposed to different heavy-metals. Appl Environ Microbio. (1993) 59:3606–17. doi: 10.1128/aem.59.11.3605–3617.1993
47. Bardgett RD, Hobbs PJ, Frostegård A. Changes in soil fungal:bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol Fertil. Soils. (1996) 22:261–4. doi: 10.1007/BF00382522
48. Keinänen MM, Martikainen PJ, Korhonen LK, Suutari MH. Microbial communities in drinking water systems analysed by lipid biomarkers. Water Sci Technol. (2003) 47:143–7.
49. Federle TW, Dobbins DC, Thornton-Manning JR, Jones DD. Microbial biomass, activity, and community structure in subsurface soils. Ground Water. (1986) 24:365–74. doi: 10.1111/j.1745-6584.1986.tb01013.x
50. Rinnan R, Baath E. Differential utilization of carbon substrates by bacteria and fungi in tundra soil. Appl Environ Microb. (2009) 75:3611–20. doi: 10.1128/AEM.02865–08
51. R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing (2020). Available at: http://www.R-project.org/, ISBN: 3–900051–07-0.
52. Devkota M, Devkota KP, Kumar S. Conservation agriculture improves agronomic, economic, and soil fertility indicators for a clay soil in a rainfed Mediterranean climate in Morocco. Agric Syst. (2022) 201:103470. doi: 10.1016/j.agsy.2022.103470
53. Lamane H, Moussadek R, Baghdad B, Mouhir L, Briak H, Laghlimi M, et al. Soil water erosion assessment in Morocco through modeling and fingerprinting applications: A review. Heliyon. (2022) 8:e10209. doi: 10.1016/j.heliyon.2022.e10209
54. Dewenam LEF, Er-Raki S, Ezzahar J, Chehbouni A. Performance evaluation of the WOFOST model for estimating evapotranspiration, soil water content, grain yield and total above-ground biomass of winter wheat in Tensift Al Haouz (Morocco): Application to yield gap estimation. Agronomy. (2021) 11:2480. doi: 10.3390/agronomy11122480
55. Diop M, Chirinda N, Beniaich A, El Gharous M, El Mejahed K. Soil and water conservation in Africa: State of play and potential role in tackling soil degradation and building soil health in agricultural lands. Sustainability. (2022) 14:13425. doi: 10.3390/su142013425
56. Saviozzi A, Biasci A, Riffaldi R, Levi-Minzi R. Long-term effects of farmyard manure and sewage sludge on some soil biochemical characteristics. Biol Fertil. Soils. (1999) 30:100–6. doi: 10.1007/s003740050594
57. Rakhsh F, Golchin A. Carbohydrate concentrations and enzyme activities as influenced by exchangeable cations, mineralogy and clay content. Appl Clay Sci. (2018) 163:214–26. doi: 10.1016/j.clay.2018.07.031
58. Bastida F, Kandeler E, Moreno JL, Ros M, García C, Hernández. T. Application of fresh and composted organic wastes modifies structure, size and activity of soil microbial community under semiarid climate. Appl Soil Ecol. (2008) 40:318–29. doi: 10.1016/j.apsoil.2008.05.007
59. Hao Z, Zhao Y, Wang X, Wu J, Sun Y. Thresholds in aridity and soil carbon-to-nitrogen ratio govern the accumulation of soil microbial residues. Commun Earth Environ. (2021) 2:236. doi: 10.1038/s43247-021-00306-4
60. Bárcenas-Moreno G, García-Orenes F, Mataix-Solera J, Mataix-Beneyto J, Bååth E. Soil microbial recolonisation after a fire in a Mediterranean forest. Biol Fertil. Soils. (2011) 47:261. doi: 10.1007/s00374–010-0532–2
61. Beffa T, Blanc M, Aragno M. Obligately and facultatively autotrophic, sulfur- and hydrogen-oxidizing thermophilic bacteria isolated from hot composts. Arch Microbiol. (1996) 165:34–40. doi: 10.1007/s002030050293
62. Beffa T, Blanc M, Lyon PF, Vogt G, Marchiani M, Fischer JL, et al. Isolation of Thermus strains from hot composts (60 to 80°C). Appl Environ Microbiol. (1996) 62:1723–7. doi: 10.1128/aem.62.5.1723-1727.1996
63. Zhang QC, Wang GH, Yao HY. Phospholipid fatty acid patterns of microbial communities in paddy soil under different fertilizer treatments. J Environ Sci. (2007) 19:55–9. doi: 10.1016/s1001–0742(07)60009–6
64. Bossio DA, Scow KM. Impact of carbon and flooding on the metabolic diversity of microbial communities in soils. Appl Environ Microbio. (1995) 61:4043–50. doi: 10.1128/aem.61.11.4043–4050.1995
65. Carrasco L, Gattinger A, Fließbach A, Roldán A, Schloter M, Caravaca F. Estimation by PLFA of Microbial Community Structure Associated with the Rhizosphere of Lygeum spartum and Piptatherum miliaceum Growing in Semiarid Mine Tailings. Microbial Ecol. (2010) 60:265–71. doi: 10.1007/s00248-009-9619-4
66. Gómez-Brandón M, Aira M, Lores M, Domínguez J. Changes in microbial community structure and function during vermicomposting of pig slurry. Bioresour. Technol. (2011) 102:4171–8. doi: 10.1016/j.biortech.2010.12.057
67. Azedou A, Lahssini S, Khattabi A, Meliho M, Rifai NA. Methodological comparison of three models for gully erosion susceptibility mapping in the rural municipality of El Faid (Morocco). Sustainability. (2021) 13:682. doi: 10.3390/su13020682
68. Oraegbunam CJ, Obalum SE, Watanabe T, Madegwa YM, Uchida Y. Differences in carbon and nitrogen retention and bacterial diversity in sandy soil in response to application methods of charred organic materials. Appl Soil Ecol. (2022) 170:104284. doi: 10.1016/j.apsoil.2021.104284
69. Oraegbunam CJ, Kimura A, Yamamoto T, Madegwa YM, Obalum SE, Tatsumi C, et al. Bacterial communities and soil properties influencing dung decomposition and gas emissions among Japanese dairy farms. J Soil Sci Plant Nutr. (2023) 23:3343–8. doi: 10.1007/s42729–023-01250–2
70. Chowdhury N, Marschner P, Burns RG. Soil microbial activity and community composition: impact of changes in matric and osmotic potential. Soil Biol Biochem. (2011) 43:1229–36. doi: 10.1016/j.soilbio.2011.02.012
71. Li J, Xie T, Zhu H, Zhou J, Li C, Xu L, et al. Alkaline phosphatase activity mediates soil organic phosphorus mineralization in a subalpine forest ecosystem. Geoderma. (2021) 404:115376. doi: 10.1016/j.geoderma.2021.115376
72. Lin VS, Rosnow JJ, McGrady MY, Smercina DN, Nuñez JR, Renslow RS, et al. Non-destructive spatial analysis of phosphatase activity and total protein distribution in the rhizosphere using a root blotting method. Soil Biol Biochem. (2020) 146:107820. doi: 10.1016/j.soilbio.2020.107820
73. Zheng H, Zhou L, Wei J, Tang Q, Zou Y, Tang J, et al. Cover crops and chicken grazing in a winter fallow field improve soil carbon and nitrogen contents and decrease methane emissions. Sci Rep. (2020) 10:12607. doi: 10.1038/s41598-020-69407-y
74. Verburg K, Bond WJ, Hunt JR. Fallow management in dryland agriculture: Explaining soil water accumulation using a pulse paradigm. Field Crops Res. (2012) 130:68–79. doi: 10.1016/j.fcr.2012.02.016
75. Kolawole GO, Tian G, Tijani-Eniola H. Effects of fallow duration on soil phosphorus fractions and crop P uptake under natural regrowth and planted Pueraria phaseoloides fallow systems in Southwestern Nigeria. Exp Agric. (2005) 41:51–68. doi: 10.1017/S0014479704002327
76. Wood SL, Rhemtulla JM, Coomes OT. Cropping history trumps fallow duration in long-term soil and vegetation dynamics of shifting cultivation systems. Ecol applications: Publ Ecol Soc America. (2017) 27:519–31. doi: 10.1002/eap.1462
77. Bacq-Labreuil A, Neal AL, Crawford JW, Mooney SJ, Akkari E, Zhang X, et al. Significant structural evolution of a long-term fallow soil in response to agricultural management practices requires at least 10 years after conversion. Eur J Soil Sci. (2020) 1–13. doi: 10.1111/ejss.13037
78. Deressa A, Yli-Halla M, Mohamed M. Soil organic carbon stock and retention rate among land uses along Didessa toposequence in humid Western Ethiopia. Environ Syst Res. (2020) 9:34. doi: 10.1186/s40068-020-00199-w
79. Adekiya AO, Aremu C, Agbede TM, Olayanju A, Ejue WS, Adegbite KA, et al. Soil productivity improvement under different fallow types on Alfisol of a derived savanna ecology of Nigeria. Heliyon. (2021) 7:e06759. doi: 10.1016/j.heliyon.2021.e06759
80. Laskar SY, Sileshi GW, Pathak K, Debnath N, Nath AJ, Laskar KY, et al. Variations in soil organic carbon content with chronosequence, soil depth and aggregate size under shifting cultivation. Sci Total Environ. (2021) 762:143114. doi: 10.1016/j.scitotenv.2020.143114
81. Ruis SJ, Stepanovic S, Blanco-Canqui H. Intensifying a crop– fallow system: impacts on soil properties, crop yields, and economics. Renewable Agric Food Syst. (2023) 38:e42, 1–10. doi: 10.1017/S1742170523000133
82. Kmoch L, Pagella T, Palm M, Sinclair F. Using local agroecological knowledge in climate change adaptation: A study of tree-based options in Northern Morocco. Sustainability. (2018) 10:3719. doi: 10.3390/su10103719
83. Kohli A, Saini BC. Microclimate modification and response of wheat planted under trees in a fan design in northern India. Agroforestry Syst. (2003) 58:109–17. doi: 10.1023/A:1026090918747
84. Soni ML, Subbulakshmi V, Sheetal KR, Yadava ND, Dagar JC. Agroforestry for increasing farm productivity in water-stressed ecologies. In: Dagar J, Tewari V, editors. Agroforestry. Springer, Singapore (2017). doi: 10.1007/978–981-10–7650-3_15
Keywords: soil organic carbon, PLFA, enzymatic activity, semi-arid, soil microbes, fallowing, Eucalyptus
Citation: Jindo K, El Aroussi O, de Vente J, López Carratalá J, Bastida F, Izquierdo CG, Sawada Y, Goron TL and Barberá GG (2024) Effects of local farming practices on soil organic carbon content, enzymatic activities, and microbial community structure in semi-arid soils of Morocco. Front. Soil Sci. 4:1369971. doi: 10.3389/fsoil.2024.1369971
Received: 13 January 2024; Accepted: 16 May 2024;
Published: 03 June 2024.
Edited by:
Riccardo Spaccini, University of Naples Federico II, ItalyReviewed by:
Heba Elbasiouny, Al-Azhar University, EgyptCopyright © 2024 Jindo, El Aroussi, de Vente, López Carratalá, Bastida, Izquierdo, Sawada, Goron and Barberá. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Keiji Jindo, a2VpamkuamluZG9Ad3VyLm5s
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.