 Jacopo Talamini1
Jacopo Talamini1 Eric Medvet
Eric Medvet Stefano Nichele
Stefano Nichele- 1Evolutionary Robotics and Artificial Life Lab, Department of Engineering and Architecture, University of Trieste, Trieste, Italy
- 2Department of Computer Science, Artificial Intelligence Lab, Oslo Metropolitan University, Oslo, Norway
- 3Department of Holistic Systems, Simula Metropolitan Center for Digital Engineering, Oslo, Norway
The paradigm of voxel-based soft robots has allowed to shift the complexity from the control algorithm to the robot morphology itself. The bodies of voxel-based soft robots are extremely versatile and more adaptable than the one of traditional robots, since they consist of many simple components that can be freely assembled. Nonetheless, it is still not clear which are the factors responsible for the adaptability of the morphology, which we define as the ability to cope with tasks requiring different skills. In this work, we propose a task-agnostic approach for automatically designing adaptable soft robotic morphologies in simulation, based on the concept of criticality. Criticality is a property belonging to dynamical systems close to a phase transition between the ordered and the chaotic regime. Our hypotheses are that 1) morphologies can be optimized for exhibiting critical dynamics and 2) robots with those morphologies are not worse, on a set of different tasks, than robots with handcrafted morphologies. We introduce a measure of criticality in the context of voxel-based soft robots which is based on the concept of avalanche analysis, often used to assess criticality in biological and artificial neural networks. We let the robot morphologies evolve toward criticality by measuring how close is their avalanche distribution to a power law distribution. We then validate the impact of this approach on the actual adaptability by measuring the resulting robots performance on three different tasks designed to require different skills. The validation results confirm that criticality is indeed a good indicator for the adaptability of a soft robotic morphology, and therefore a promising approach for guiding the design of more adaptive voxel-based soft robots.
1 Introduction
Traditionally, engineers have designed robotic systems modeled by connected joints made of rigid materials. These rigid-body robots can be programmed to efficiently perform a single task in a predictable way, but often with limited adaptability (Rus and Tolley, 2015).
Soft robots, on the contrary, are designed using soft materials, in order to mimic nature in the way they interact with the environment. Since they are made of soft materials, soft robots are provided with almost infinite degrees of freedom and thus are capable of more natural movements. This allows a variety of different behaviors that were not possible with traditional robots and many new opportunities for robotics, enabled by their greater adaptability (Lipson, 2014). Soft robotic bodies are able to bend and twist with high curvatures, through deforming part of their body in a continuous way (Mazzolai et al., 2012), and thus can move in confined spaces. Since they can adapt their body shape to the environment, soft robots are thus able to manipulate objects, move on rough terrain, and execute rapid and agile manoeuvres underwater. However, due to the intrinsic complexity of their bodies, the design and control of soft robots is a challenging task, which suggests that traditional robotics techniques might not be effective.
Among the existing categories of soft robots, Voxel-based Soft Robots (VSRs) are made of many elastic blocks called voxels, defined by mechanical properties similar to those of biological tissues, which allow them to expand or contract when controlled by an external signal. A VSR is defined by a body (or morphology), which is simply an aggregate of voxels, and a brain, which is the control algorithm responsible for actuating its body. Biological inspired meta-heuristics such as Evolutionary Computation (EC) have been extensively applied to the domain of VSRs (Hiller and Lipson, 2011; Cheney et al., 2013; Cheney et al., 2015; Talamini et al., 2019; Medvet et al., 2020a; Ferigo et al., 2021) and have been shown to be a promising approach for both the design and control of VSRs. However the optimization of these robots is often oriented toward a specific task, e.g., a locomotion task, which provides no guarantee on the effectiveness of the resulting designs when subjected to a different task.
In this work, we explore the possibility of automatically designing adaptable soft-robotic bodies by means of EC, such that the resulting bodies are able to successfully accomplish tasks requiring different motor skills. A simple method to find such bodies might consist in evaluating each candidate solution on all the relevant tasks, and optimizing the body toward the maximization of the overall performance. However, this approach does not necessarily scale well, since as more and more tasks are to be achieved the computation required to evaluate the robot performances on all tasks may become not practical. In addition, not all the possible tasks are necessarily known in advance, and the definition of a new task would make the results of the previous optimization process outdated.
A better approach may be to identify measures for soft-robot bodies that may evaluate the potential richness of robot dynamics (the potentially available robot behaviors), such that different controllers may be able to successfully operate the robot on different tasks. We propose here a task-agnostic approach for automatically designing adaptable bodies without requiring any information on the tasks, and instead based on the definition of criticality (Bak et al., 1988).
A system in the critical state operates at the edge between two qualitatively different types of behavior. Below the critical state a system is said to be in a subcritical state, where the behavior is highly ordered (static or oscillating between very few distinct states). On the other hand, a system in a supercritical state shows chaotic behavior (unpredictable). At a phase transition between such regimes, a systems is typically able to efficiently respond to a wide range of inputs. Langton (1990) has shown that Cellular Automata at the transition phase, i.e., edge of chaos, are able to efficiently transmit, store, and modify information, and therefore are able to perform complex computations. Biological neural networks and the brain are examples of systems that operate at criticality (Shew and Plenz, 2013; Hesse and Gross, 2014; Heiney et al., 2019). Criticality is typically measured by looking at how activity propagates through the system, such as spike propagation in neural networks. When a certain activity propagates through the system, this is referred to as avalanche. Avalanches may have different size and duration. When the avalanche distribution follows a power law, the system is said to be at criticality. A recent review on criticality and connectivity in relation to neural computation is available at (Heiney et al., 2021).
A popular machine learning approach which relies on dynamical systems computation is Reservoir Computing (RC) (Jaeger, 2001; Maass et al., 2002). In RC, the dynamical system is able to represent input signals in a high-dimensional temporal state, which can be easily distinguished by a linear trained readout. While RC has been implemented in several physical dynamical systems (see Konkoli et al. (2018) and Tanaka et al. (2019) for a recent review), including non-modular soft robots (Li et al., 2012; Nakajima et al., 2013) and origami robots (Bhovad and Li, 2021), it has been shown that the reservoir performs best if the dynamic regime is in a critical state (Schrauwen et al., 2007; Legenstein and Maass, 2007). A VSR body is a dynamical system and may therefore be considered a reservoir computer.
Our hypothesis is that VSR bodies engineered or optimized to perform robustly for a given task may show sub-critical behavior due to their specific topology, i.e., a fairly static or oscillating behavior that works particularly well for one single task. In other words, the required computation is strongly embedded in the body morphology. However, such robust behavior and topology is not particularly adaptive to other tasks requiring very different motor skills and consequently a radically different dynamical behavior. On the other hand, a topology supporting a critical behavior, i.e., a wide range of controllable dynamics, may achieve sub-optimal performances on a wide variety of tasks requiring different motor skills, while not being explicitly optimized for any of them.
In this work we provide a criticality score value, based on the fitting of the empirical avalanches distribution (Bak et al., 1988), coming from the assessment of a body, with a target distribution. Given this score, we evolve VSR morphologies that are optimized for criticality, rather than for a single or a few tasks. The morphology evolution is hence task-agnostic.
To validate the adaptability of the morphologies resulting from the evolution, we design three reasonably different tasks: a locomotion task on a plain ground, a jump task, and a task of escape from a narrow cave-like environment, and we test the bodies on these tasks by optimizing their controller on each task. Figure 1 shows an overview of our approach and its experimental evaluation. While our focus is on the adaptability of the VSR morphologies to different tasks based on their criticality, we also investigate whether morphologies evolved with high criticality show adaptability to different controller types, e.g., phase controllers and neural network controllers. Brain adaptability for robotics is a rather consolidated researched topic where adaptive mechanisms may act on short time scales, i.e., through learning. Mechanisms of body adaptation are less understood both for robots and biological organisms. In addition to the bodies evolved toward criticality, we have considered several other bodies for the validation process, some of them inspired by previous works such as Talamini et al. (2019), while others automatically generated thorough algorithms based on randomness. We compare all the robots on the different tasks, and we draw an overall ranking that shows that criticality is indeed a good predictor of the adaptability of a robot morphology. All the experimental work is performed in simulation.
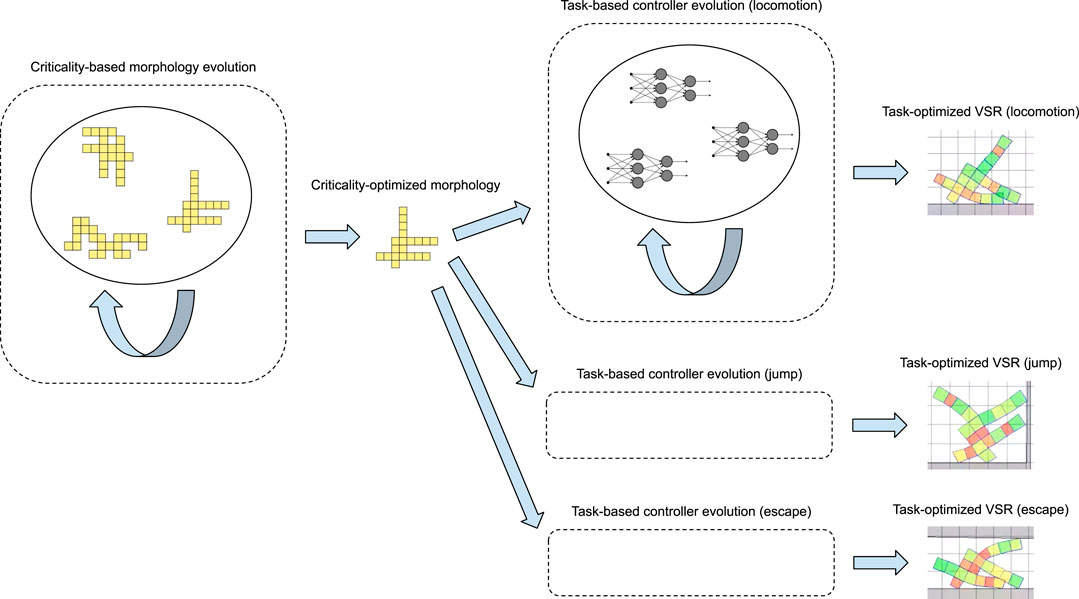
FIGURE 1. Schematic view of our approach and its experimental evaluation. We first evolve a morphology driving the optimization with a measure of its criticality: during this evolution (left), candidate morphologies are not evaluated on any specific task. Then, we take a criticality-optimized morphology and test its adaptability, i.e., whether a VSR with a task-optimized controller working with this morphology obtains reasonable performance over a set of different tasks. For this evaluation, we evolve a controller (e.g., a neural network) for the criticality-optimized morphology: during each one of these evolutions (right), candidate controllers are evaluated, embedded in a robot with the criticality-optimized morphology, on a specific task.
In summary, we experimentally tested the following hypothesis, here formulated as research questions:
RQ1. Do morphologies evolved for criticality obtain a higher criticality value than handcrafted morphologies?
RQ2. Do VSRs with morphologies evolved for criticality obtain an average performance that is not inferior (on a set of different tasks) than that of VSRs with handcrafted morphologies? In both cases, the controllers are evolved for each specific task.
RQ3. Given a pseudo-randomized method for generating morphologies that resemble the ones evolved for criticality, do VSRs with those pseudo-randomized morphologies obtain an average performance that is not inferior (on a set of different tasks) than that of VSRs with handcrafted morphologies? In both cases, the controllers are evolved for each specific task.
Our main findings are the following. 1) Criticality is a good predictor of the potential adaptability of a robot morphology. 2) Criticality as task-agnostic fitness score for the evolution of robot bodies result in the design of robots that are among the most adaptable. 3) Bodies evolved for a specific tasks exhibit almost no adaptability and have indeed particularly low levels of criticality. 4) Morphologies evolved toward criticality show a rather randomized structure that can be mimicked in a random generation procedure for morphologies. Such procedure generate morphologies that are sometimes adaptable and with criticality values that are in a range between those evolved for criticality and those evolved for a specific task. The random generation procedure is computationally fast as compared to EC. 5) The morphologies evolved for criticality may be controlled by different controller types, i.e., phase controller and neural network controller. This suggests that morphologies supporting critical behavior are agnostic to the type of controller being used.
2 Background and Related Work
In this section, we first describe voxel-based soft robots. Subsequently, we introduce the concepts related to reservoir computing, a method that relies on dynamical systems for efficient computation. We then provide links between VSRs and RC and finally introduce a beneficial property for dynamical systems and their computational power, namely criticality.
2.1 Voxel-Based Soft Robots
A Voxel-based Soft Robots (VSR) is an aggregation of soft cubic blocks, voxels, that can vary their volume in response to a control signal. Hiller and Lipson (2012) first introduced VSRs along with a procedure to physically build them using a deformable foam: the control signal corresponded to the atmospheric pressure and voxels were not controllable individually. Later approaches to the realization of VSRs explored different paths as, e.g., silicone cubes actuated by injected air pressure (Kriegman et al., 2020b; Sui et al., 2020) or living matter (Kriegman et al., 2020a). Regardless of the way they are built, VSRs are often designed automatically by means of optimization and simulation: many different designs are simulated and their ability to perform a given task is measured.
In this work, we consider a 2-D variant of simulated VSRs that has been introduced by Medvet et al. (2020b) together with a time-discrete simulation engine that facilitates their optimization. A detailed description of the mechanical model of the VSRs and how they are simulated is given in the cited work and in (Medvet et al., 2020c): we here summarize the salient concepts.
A VSR is defined by its morphology and its controller. The former describes how the voxels are arranged. The controller determines how the area of each voxel varies over the time, possibly based on the readings of the sensors the VSR is equipped with. The ability of sensing both itself and the environment makes VSRs potentially more effective in performing tasks where perception is beneficial as, e.g., locomotion on uneven terrains (Talamini et al., 2019).
2.1.1 Morphology
A morphology is a 2-D grid of voxels: adjacent voxels are rigidly connected at their vertices. In the simulation, the voxel is modeled as a compound of spring-dampers systems, masses, and distance constraints. By varying the parameters of those components, different materials for the voxels can be simulated: in this work, however, we use the same default values for each voxel and hence assume that the voxels of a VSR are composed of the same material.
2.1.2 Controller
During the simulation, voxels vary their area in response to a control signal imposed by the controller and to the external forces deriving from the contact with other voxels and with the ground. The control signal is a value in
The value of the control signal is set for each voxel at each time-step by the VSR controller. Different controllers have been used in previous works, ranging from simple ones where the control signal for each voxel depends only on the time (Kriegman et al., 2018), to others where a recurrent neural network processes the VSR sensor readings to produce the control signals (Medvet et al., 2020a). With the aim of evaluating the adaptability also with respect to different forms of controller, in this study we consider two controllers.
In the first and simplest one, that we call phase controller, the control signal of the ith voxel at time
That is, all the voxels are controlled with a sinusoidal signal with the same amplitude and a frequency of 1 Hz, but different phases
The second controller, that we call neural controller, is based on a fully connected feedforward artificial neural network (NN) and resembles that proposed by Talamini et al. (2019). At each time step, the controller collects the vector
When using the neural controller, we equipped each voxel of the VSR with four sensors, resulting in an overall number of sensors
2.2 Adaptability
One of the main challenges and opportunities of VSRs is their potential adaptability. Robots made of soft materials may interact with their environment in a more natural way, inspired by how biological systems interact with their environments (Trimmer, 2013).
According to Rus and Tolley (2015), these robotic systems are more adaptable and versatile than traditional ones made of rigid joints. However, this aspect has never been completely explored, since most of the research in the literature has dealt with the automatic design of robots for a very specific task. Kriegman et al. (2018) and Cheney et al. (2013) considered different aspects of soft robots such as the lifetime development and an effective representation and the results are evaluated on their locomotion performance. Sadeghi et al. (2017) engineered a soft robot inspired by the plants for growing. Finally, Shen et al. (2017) took inspiration from cephalopod molluscs for developing a robot that exhibited interesting underwater propulsion and maneuvering mechanisms.
Auerbach and Bongard (2014) showed that the morphological complexity is a result of the selection pressure provided by increased environmental complexity. Recently, Miras et al. (2020) wrote about (Auerbach and Bongard, 2014) that: The authors demonstrated that increasing the complexity of the environmental conditions might result in an increase to the morphological complexity of the creatures. However, measuring complexity does not provide clear insights concerning properties of intelligible morphological traits, for instance, the number of limbs a robot has. Importantly, two environments could be equally complex, but induce the emergence of different phenotypic and behavioral traits. Chvykov et al. (2021) attempted self-organization away from equilibrium through the use of shape-changing robotic active matter, and outlined a methodology for controlling collective behavior. Corucci et al. (2018) investigated the evolution of walking and swimming soft robots in different environments to explore the effect of major environmental transitions during evolution. Horibe et al. (2021) focused on the regeneration of soft robot bodies using growing neural cellular automata when damages are induced to the morphology.
An approach for task-agnostic morphology evolution (TAME) is presented by Hejna et al. (2021), where an information theoretic objective is used instead of the robot performance in a specific task. While the used morphologies are not voxel-based (as in our work) but are encoded as a tree of limbs and joints, the underlying assumption is similar to ours in that morphologies that can achieve a large amount of states in a controllable way may perform well in different tasks. Their approach is based on a graph neural network as predictor of the actions taken by each joint and mutual information-based fitness to maximize the number of possible actions.
Very recently, Bhovad and Li (2021) have proposed soft robots with origami capabilities, which result in a dynamical and high-dimensional system that can be harnessed as physical substrate for RC. The framework of RC in the context of soft robots may provide a novel view on soft robot bodies as dynamical systems and foster the investigation about dynamical behaviors that may be controllable and adaptive.
2.3 Reservoir Computing
Reservoir Computing (RC) is a computational framework derived from echo state networks (Jaeger, 2002) and liquid state machines (Fernando and Sojakka, 2003), which leverages on the high-dimensionality of a dynamical system, i.e., its substrate, to produce training-efficient linear readout mechanisms as an alternative to expensive recurrent neural network training. In practice, dynamical systems that possess the echo state property (previous inputs “echo” through the network for a certain amount of time and thereby slowly disappear without being amplified) may be used.
Recent works on RC, such as (Li et al., 2012; Nakajima et al., 2013; Tanaka et al., 2019), suggest that soft robotic systems, thanks to the intrinsically high-dimensionality, non-linearity, and elasticity property of soft materials, which lead to overall highly complex and time-varying dynamics under actuation, are perticularly well suited substrates for RC. Specifically, Li et al. (2012) and Nakajima et al. (2013) showed that the structure of an octopus arm inspired soft robot can be exploited as a computational resource.
Indeed, it has been shown that the rich dynamics used by reservoir computers are often present in systems that show complex behaviors, typically at a critical point between ordered and chaotic dynamics (Schrauwen et al., 2007).
2.4 Criticality
The brain criticality hypothesis states that the brain may operate in critical state between ordered and disordered dynamics (Wilting and Priesemann, 2019). Criticality is considered one of the mechanisms by which complexity arises in nature, suggesting the possibility that criticality has been selected by evolution as a beneficial trait (Hesse and Gross, 2014). Theoretical and experimental work has shown that systems at criticality allow for optimal information processing and computing properties (Zimmern, 2020). In practice, in order to achieve criticality, a system parameter has to be precisely tuned. However, some dynamical systems such as the brain have the ability to self-tune through local processes, i.e., self-organized criticality (SOC). SOC is a property typically observed in slowly driven non-equilibrium systems with many degrees of freedom and strongly non-linear dynamics (Bak et al., 1988), which naturally evolve toward a critical point of phase transition between chaotic and non-chaotic regimes (Bertschinger and Natschläger, 2004).
Being close to this phase transition allows systems in which complex computation is possible, as presented by Langton (1990), who investigated the conditions that allow computation in Cellular Automata (CA). Criticality is also inherently connected to the separation property, used by Gibbons (2010) for the assessment of neural reservoirs. Brodeur and Rouat (2012) and Heiney et al. (2019) demonstrated, for spiking neural networks, that the regulation of a RC substrate toward criticality may allow to obtain a more powerful reservoir.
A standard metric for estimation of criticality is to measure how close is the empirical avalanches distribution in the system (Bak et al., 1988) to a target power law distribution. Such metric exploits a convenient representation for the distributions based on the method presented by Clauset et al. (2009), which is also used in this work. A graphical explanation of avalanche quantification is depicted in Figure 2. An example of using such criticality metric as fitness function in EC is provided in (Pontes-Filho et al., 2020a; Pontes-Filho et al., 2020b), where the goal was to evolve CAs toward SOC.
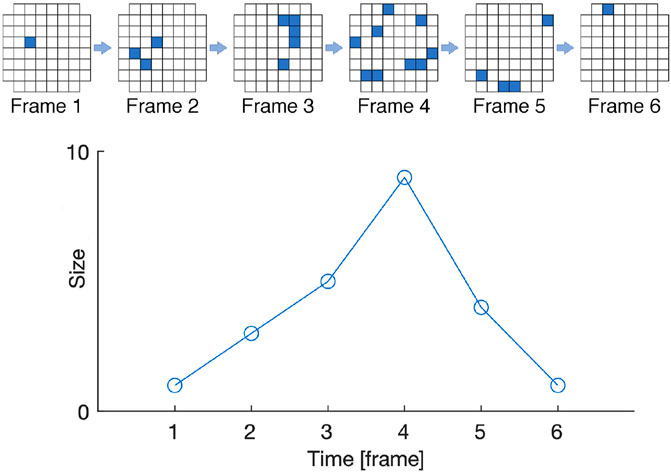
FIGURE 2. Graphical representation of avalanches in a system composed by 60 cells arranged in a grid. The top row shows an initial activity from a single cell at Frame 1 that propagates to three cells at Frame 2, eventually ending at Frame 6. The bottom row shows the avalanche size and duration. Image adapted from (Heiney et al., 2021) in the context of biological neural networks on Micro-Electrode Arrays.
3 Optimizing Voxel-Based Soft Robot Morphology for Criticality
The central hypothesis of this study is that a VSR morphology that exhibits the property of criticality is adaptable; that is, VSRs with that morphology, but possibly different controllers, are good in a diverse set of tasks. As a consequence, we speculate that optimizing for criticality results in optimizing for adaptability. In order to verify this hypothesis, we need a way for measuring the degree to which a VSR morphology exhibits the criticality property and a way for optimizing a VSR for criticality. In the following sections, we describe in detail our approaches for both things.
3.1 Criticality of a Voxel-Based Soft Robot Morphology
We build our definition of criticality of a VSR morphology based on the concept of avalanche. Intuitively, given a dynamical system in which an initial input stimulus can be injected, the avalanche is the resulting outcome and its extension can be measured in time (how long it lasts) and space (how broadly it involves the system). A system that is chaotic will often produce extended avalanches, regardless of the initial input stimulus. Conversely, a system that is predictable and prone to reach equilibrium will in general produce large avalanches only for few input stimuli. Based on this intuitive observation, Bak et al. (1988) showed that systems that lie on the boundary between chaotic and non-chaotic regimes (the edge of chaos), i.e., that exhibit criticality, produce avalanches whose extension distribution resembles the power law distribution. Summarizing, the closer the avalanche extension distribution to the power law distribution, the more the system exhibits criticality.
In order to apply this intuitive definition of criticality to the morphology of a VSR, we need to define what is an input stimulus and how to measure the avalanche extension, provided that the morphology itself, i.e., a 2-D grid of voxels, constitutes the dynamical system.
We define as input stimulus the application of a control signal
where
In order to minimize the impact of the effect of the gravity and to make our definition of criticality agnostic with respect to the morphology rotation, we perform the simulation in absence of gravity.
Based on the above definition of avalanche extension and on the previous studies on avalanche distribution in systems exhibiting criticality, we measure the criticality as follows. Given a morphology b composed of n voxels, we obtain n measures of avalanche extensions
with
3.1.1 Choice of the Threshold τ for the Relative Area Variation
In practice, a key role in the quality of the measure of criticality is played by the parameters
Concerning
We consider the square morphologies of
Then, we consider 10 values for τ evenly distributed in
This way, we obtain for each one of 10 robots of sizes ranging from
Figure 3 shows the fitted model and the values for
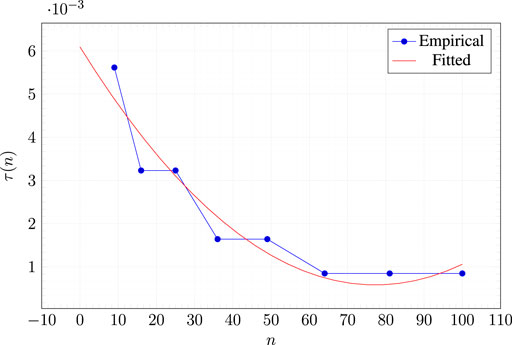
FIGURE 3. Experimental values for the optimal threshold
3.2 Evolutionary Optimization of Criticality
We aim at optimizing a VSR morphology for criticality, i.e., we want to search in the space of VSR morphologies for the one that corresponds to the largest criticality, as defined in Eq. 3.
We tackle this optimization problem with EC: this large family of optimization methods has been showed to be effective for uncommon search spaces (here, the space of VSR morphologies) and objective functions that are not well characterized. Indeed, EC has been used extensively for the optimization of VSRs: e.g., the morphology in (Cheney et al., 2013), the controller in (Talamini et al., 2019), the sensory apparatus in (Ferigo et al., 2021).
Two key components of any optimization performed by means of EC are the solution representation and the Evolutionary Algorithm (EA). Concerning the former, we represent VSR morphologies as numerical vectors in
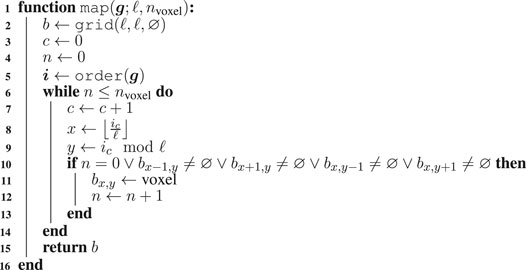
Algorithm 1: The algorithm for mapping a genotype
For the evolutionary optimization of the morphology, we use the simple EA of Algorithm 2. We iteratively evolve a population of

Algorithm 2: The EA for the optimization of the morphology.
We initialize the population by randomly sampling
We remark that we do not optimize the morphology of the VSR for a specific task. Instead, we optimize for criticality, on the assumption that great criticality corresponds to a morphology that is adaptable to different tasks.
4 Experiments and Results
In this section, we describe the experimental work carried out to verify our hypotheses. First, we investigate whether it is possible to evolve VSR morphologies that are optimized for criticality, and therefore task-agnostic. In order to quantify the level of criticality of VSR morphologies, we measure the avalanche distribution produced by each morphology as described in Section 3.1.
Second, we test the criticality-evolved morphologies on three different tasks. The research hypothesis here is that the morphologies evolved for criticality perform reasonably well on all tasks, therefore showing adaptation. In this step, we also test some handcrafted morphologies that are expected to be very well suited for specific tasks. While the handcrafted morphologies may be better than the criticality-evolved ones on specific tasks, we expect the overall ability to solve different tasks to be inferior than the criticality-evolved morphologies. In order to test the criticality-evolved morphologies as well as the handcrafted morphologies on the different tasks, we optimize the phase controller (see Section 2.1.2) of several VSRs for each one of the morphologies. We do the optimization with a state-of-the-art EA, detailed below.
Third, we explore whether there are cheaper ways to optimize task-agnostic morphologies with similar dynamical properties as those evolved for criticality. To do that, we start with the observation that VSRs that show higher degree of criticality seem to have morphologies that are less structured, with some degree of randomness in the components stretching out of the bodies. We introduce a pseudo-random generation method that, starting from a voxel, adds a new voxel in one of the neighboring positions. Also, we introduce a random method that adds voxels in random locations until a connected component of a target size is achieved. We measure the criticality of both randomized methods and we test the resulting VSRs on the same three tasks used in the previous experiments (for comparison) by optimizing their phase controllers.
Finally, we investigate whether the criticality-evolved task-agnostic morphologies retain their adaptability also when coupled with a neural controller (see Section 2.1.2), rather than a phase controller. Again, we optimize the controllers for each task and each morphology using a state-of-the-art EA.
We performed all the experiments using publicly available software tools: JGEA1 for the evolutionary optimization and 2D-VSR-Sim2 for the VSR simulations.
4.1 Task-Agnostic Evolution of Voxel-Based Soft Robots Morphology
In this first experiment, we aim at evolving VSR morphologies using a fitness function that is purely based on the criticality measure introduced in Section 3.1. Therefore, the aim is to evolve robot morphologies that are agnostic to the task they may be tested for. Our assumption is that greater criticality corresponds to a morphology that is adaptable, once associated with a controller, to different tasks.
We executed the EA of Section 3.2 ten times with different random seeds. We set the parameters related to the representation to
Figure 4 shows the criticality of the best morphology during the evolution (median value and interquartile range across the 10 runs). It can be seen that there is an improvement of the criticality: the largest portion of the improvement is obtained in the first stage of the evolution. The figure also shows that there is still an intrinsic boundary which prevents the evolution to find a morphology with maximum theoretical criticality, i.e.,
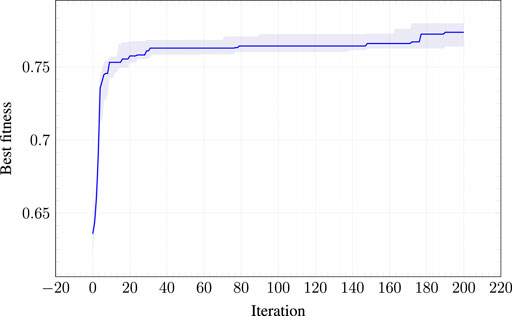
FIGURE 4. Criticality of the best morphology at each iteration of the EA used for optimizing the morphologies. The solid lines indicates the median across the 10 runs; the shaded area corresponds to the interquartile range (from 25-th to 75-percentile).
Figure 5 shows the final 10 optimized morphologies, i.e., for each run, the morphology of the final population with the largest value for the criticality. The evolved morphologies have several offshoots that are stretching out from the “central core” (i.e., the central portion of the morphology). Apparently, there is not a recurrent pattern in the placement, orientation, and length of those offshoots. In addition, the largest central cores are fairly small, ranging from rectangles of size
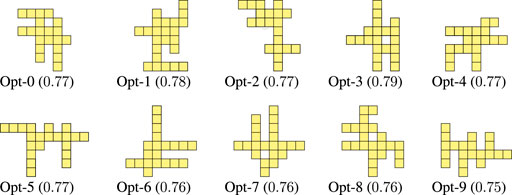
FIGURE 5. Morphologies evolved for criticality together with the corresponding value of criticality.
4.2 Validating the Adaptation to Different Tasks
In order to test how adaptable the task-agnostic morphologies evolved in the previous experiment are, we consider three tasks. For each task and each morphology, we evolve a phase controller of a VSR with that morphology; we drive the evolution with a measure of how well the VSR is performing the task. The more adaptable a task-agnostic morphology, the better the performance on all tasks of VSRs with evolved task-specific controllers and that morphology.
4.2.1 Tasks
We considered the tasks described below. In each task, we simulate the VSR under evaluation for a fixed time in a predefined, task-specific environment (with gravity); upon the simulation, we take a quantitative measure f of the degree of task achievement.
In the locomotion task, the VSR has to run along a flat terrain the farthest possible. The simulation lasts 20 s (simulated time) and we measure the average velocity of the VSR along the positive x-axis, i.e., we measure the difference between the x-coordinate of the center of mass of the VSR at
In the jump task, the VSR has to jump the highest possible. The simulation lasts 20 s and we measure the height of the highest jump done in that interval, i.e., the difference between the maximum observed value of the y-coordinate of the center of mass and its value at
Finally, in the escape task, the VSR has to exit from a cave-like environment. Here the VSR initially starts in an environment with a ceiling, a wall on the left, and a narrow aperture on the right. An example of a cave-like environment is shown in Figure 6, where the aperture has an initial height of 75% of the VSR largest dimension, and decreases slowly. The simulation lasts 40% and we measure the distance “walked” by the VSR along the positive x-axis, as for the locomotion task. Differently from the latter, however, the VSR here does not only need to exhibit a gait that allows to move effectively, but it also needs to “squeeze” itself in order to actually pass through the aperture.
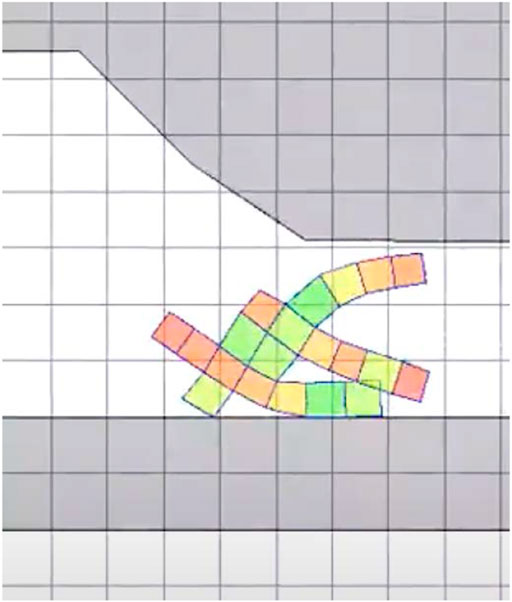
FIGURE 6. Example of cave-like environment used in the escape task.
4.2.2 Controller Evolution
Given a morphology and a task, we evolve a phase controller using Covariance Matrix Adaptation Evolutionary Strategies (CMA-ES) (Hansen et al., 2003). CMA-ES is widely considered as a state-of-the-art EA for continuous optimization, i.e., for optimization in the space
CMA-ES iteratively optimizes a numerical vector in the form of a multivariate normal distribution. At each iteration, it samples the distribution obtaining a number λ of solutions and then updates the parameters of the distribution by recomputing them based on the best
We used CMA-ES with the default parameters suggested in (Hansen, 2016): we set the initial step size
4.2.3 Comparison Baselines: Handcrafted Morphologies
To benchmark the adaptability of the criticality-evolved morphologies, we consider also four manually designed morphologies that we show in Figure 7.
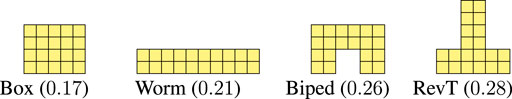
FIGURE 7. Handcrafted morphologies, designed based on domain knowledge, together with the corresponding value of criticality.
For the Worm and the Biped morphologies, we got inspiration by the results of (Talamini et al., 2019; Medvet et al., 2020b; Medvet et al., 2020c), where they have been thoroughly assessed in the task of locomotion. The Box and RevT are some of the simplest morphologies that can be manually designed. All these morphologies are made of exactly
4.2.4 Results and Discussion
For each task and each morphology, we executed CMA-ES 10 times with different random seeds. We hence performed
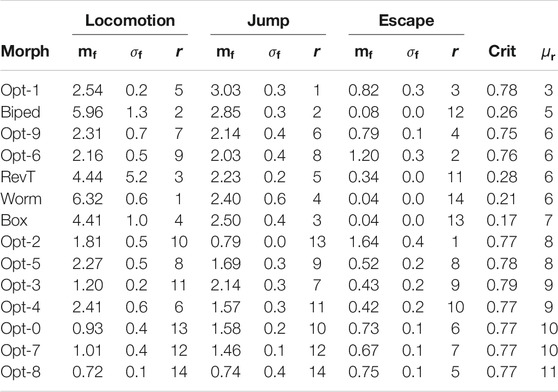
TABLE 1. Results for the optimized and handcrafted morphologies coupled with the phase controller. For each task and each morphology, the table shows the median
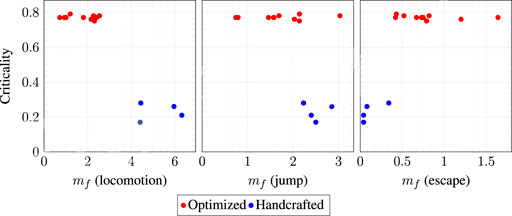
FIGURE 8. Scatter plots of the criticality vs. the median task achievement degree
Table 1 shows all the salient indices for the 10 optimized morphologies (denoted by Opt-n, with
A more detailed analysis of the results can be done by observing the rankings task by task. Considering only the results on the locomotion task, it is clear that the handcrafted morphologies, namely Worm, Biped, RevT, and Box, are actually the most effective ones. However, it can also be seen from Table 1 that some of the handcrafted morphologies, while being often extremely effective on a specific task, perform poorly on other tasks. For instance, the Worm is the most effective in locomotion and the least effective in escape.
The above considerations are corroborated by Figure 8. The figure shows, for each task, a scatter plot of the median task achievement degree
We performed an analysis of the statistical significance of the results of this experiment. Precisely, we aimed at testing two null hypotheses. First, we tested, for each task, the null hypothesis that morphologies optimized for criticality obtain the same median task achievement degree
4.3 A Cheap Proxy for Task-Agnostic Evolution
In the previous experiments, we observed that the criticality appears to be a good predictor of the adaptability: VSRs based on morphologies evolved for criticality using a task-agnostic optimization showed to be good, on average, on different tasks. However, evolution toward criticality is a resources intensive process. We hence explored the possibility of designing a generative procedure able to build, from scratch and in a one-shot fashion, a morphology that exhibits good criticality.
For designing that procedure, we observed the features of the optimized morphologies (see Figure 5). We also considered the raw results of the experiments of Section 4.1. The solutions in the initial populations of the runs of the criticality optimizations exhibited criticality values larger than those of the handcrafted morphologies (see Figure 4): that is, the joint result of the genotype initialization and the solution representation produced alone, without evolutionary pressure, morphologies with larger criticality than the handcrafted ones. Based on these considerations, we devised a non-deterministic procedure for generating morphologies with good criticality, as follows.
Given an unlimited empty grid, we start by putting a voxel in a randomly chosen position in the grid. Then, we iteratively add one voxel at a position adjacent to the last added voxel until
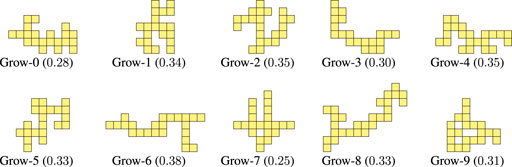
FIGURE 9. Morphologies generated with the Grow procedure together with the corresponding value of criticality.
As a comparison baseline, we also devised another non-deterministic procedure that still generates morphologies randomly, but does not attempt to mimic the shape of the optimized morphologies, namely their offshoots. In this case, we simply add voxels at random positions of an initially empty
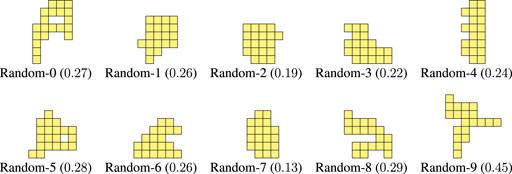
FIGURE 10. Morphologies generated with the Random procedure together with the corresponding value of criticality.
By looking at Figure LABEL:fig:bodies-grow,fig:bodies-random it can be seen that the Grow morphologies are visually more similar to the optimized ones (see Figure 5) than the Random morphologies. Moreover, the criticality of the Grow morphologies is in general larger than the criticality of the Random morphologies. We remark that both procedures are task-agnostic, as the criticality-driven evolution of the morphology.
4.3.1 Adaptability of Grow Morphologies
As a further validation of the fact that the Grow procedure can be a cheap proxy for the criticality-driven evolution of morphologies, we repeated experiments of Section 4.2 for the 10 Grow morphologies of Figure 9. We also included the 10 Random morphologies of Figure 10. We hence performed other
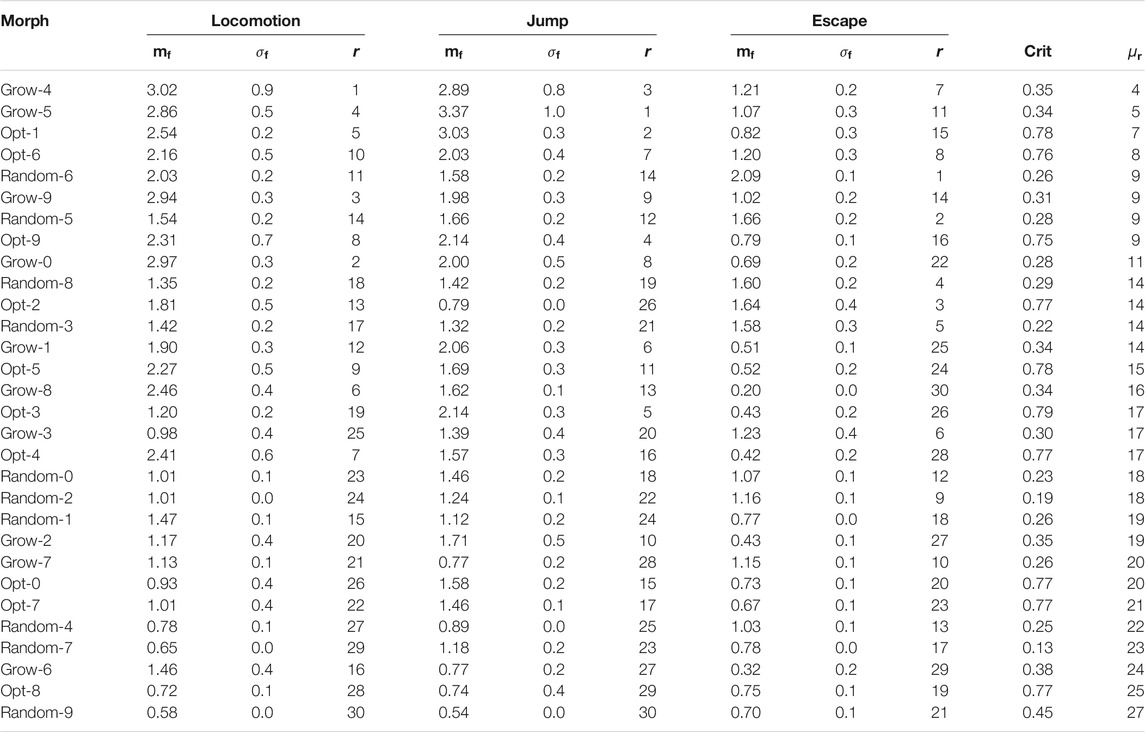
TABLE 2. Results for the optimized and randomly generated morphologies coupled with the phase controller. For each task and each morphology, the table shows the median
It can be seen that the Grow procedure, i.e., the one inspired by the results of the criticality-driven evolution results, generates morphologies that exhibit, on average, a greater adaptability (i.e., lower
We performed the same statistical significance analysis of Section 4.2.4. Considering
A few videos of evolved VSRs with the morphologies obtained with the procedures above in the three considered tasks are available online: locomotion (https://www.youtube.com/watch?v=rRPI-WvezY0), jump (https://www.youtube.com/watch?v=5JYwQPptVZE), escape (https://www.youtube.com/watch?v=1G8eO5DCRLM).
4.4 Validating the Adaptation to Different Controllers
We further investigated the adaptability of the morphology generated with the task-agnostic procedures described above—namely, the criticality-driven evolution, the Grow, and the Random procedures, by repeating the same validation process of the previous sections with a different controller. Here, we used the neural controller described in Section 2.1.2. This controller is more complex than the phase one for two reasons: 1) it is based on a neural network, and can hence generate a richer dynamics in the control signals and 2) it collects and processes the readings of the sensors the robot is equipped to.
We speculate that the ability to sense itself and the environment might be ever more beneficial with irregular morphologies.
We performed the
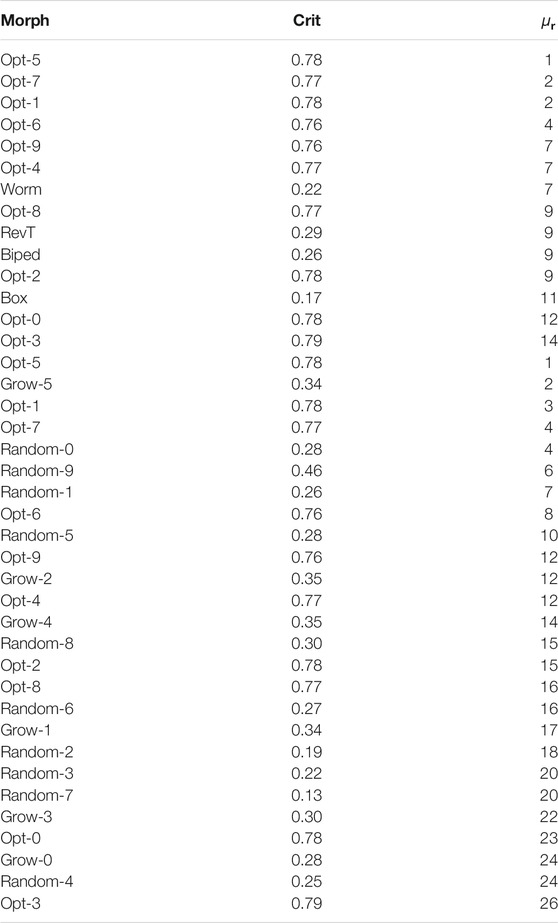
TABLE 3. Results for the optimized and randomly generated morphologies coupled with the neural controller. For each morphology, the table shows the average rank
Looking at the results presented in Table 3, our thesis is further corroborated. On one hand, the number of optimized morphologies that exhibit greater adaptability (i.e., lower
5 Conclusion
In this work, we have introduced a task-agnostic approach for measuring and predicting the adaptability potential of a soft robotic body based on the definition of criticality, a property of dynamical systems that operate at a phase transition between order and disorder. To this extent, we have proposed an algorithm for guiding the automatic design of adaptable soft robotic morphologies by means of evolutionary computation that, without the need to define a specific target task, relies entirely on the resulting dynamics of the robots morphology measured as the avalanche distributions, i.e., how close they are to a power law distribution. The morphologies evolved with our method obtain criticality values far greater than those handcrafted with domain knowledge to perform specific tasks.
We have then validated the evolved morphologies on three tasks requiring different motor skills, against some manually designed morphologies inspired by previous works where domain knowledge is considered, using a simple non-sensing control algorithm based on periodic control signals. The results show that our criticality-based approach results into more adaptable robots, i.e., performing reasonably well in the different tasks, w.r.t. the manually crafted ones. On the other hand, the robots optimized for specific tasks perform poorly on other tasks.
Motivated by the computational cost of the criticality optimization, we have then considered other design algorithms based on randomness, which on the contrary are not computationally expensive. Such randomized algorithms may produce morphologies that are visually similar to those evolved for criticality, but with criticality values that are inferior to the evolved ones (while being higher that the handcrafted robots). In addition, such morphologies generated with randomized methods show sometimes promising adaptability.
Finally, we have repeated the validation process to investigate whether a sensing controller based on neural networks would also be able to control the criticality evolved robots, and thus show adaptability to the different tasks. Neural network sensing controllers allow the robots with optimized morphologies to leverage their body complexity more than non sensing controllers. Provided with distributed sensing, the criticality evolved morphologies reach the highest overall ranking when compared to any baseline. This promising result suggests that morphologies supporting critical behavior are more adaptive independently of the type of controller being used, and independently of the target task.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
EM and SN devised the research idea and supervised the work. JT carried out most of the experimental work. All authors contributed to paper writing, data analysis, and interpretation of results.
Funding
This work has been partly financed by Project CA4VSR, grant agreement 312537, Forskermobilitet, Research Council of Norway, and Project DeepCA, grant agreement 286558, Young Research Talent, Research Council of Norway.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The experimental work has been executed partly on the high performance computing infrastructure “Experimental Infrastructure for Exploration of Exascale Computing” (eX3) at Simula Research Laboratory (Oslo, Norway) and partly on CINECA HPC cluster within the CINECA-University of Trieste agreement.
Footnotes
1https://github.com/ericmedvet/jgea.
2https://github.com/ericmedvet/2dhmsr.
References
Auerbach, J. E., and Bongard, J. C. (2014). Environmental Influence on the Evolution of Morphological Complexity in Machines. Plos Comput. Biol. 10, e1003399. doi:10.1371/journal.pcbi.1003399
Bak, P., Tang, C., and Wiesenfeld, K. (1988). Self-organized Criticality. Phys. Rev. A. 38, 364–374. doi:10.1103/physreva.38.364
Bertschinger, N., and Natschläger, T. (2004). Real-time Computation at the Edge of Chaos in Recurrent Neural Networks. Neural Comput. 16, 1413–1436. doi:10.1162/089976604323057443
Bhovad, P., and Li, S. (2021). Physical Reservoir Computing with Origami and its Application to Robotic Crawling. arXiv preprint arXiv:2101.08348.
Brodeur, S., and Rouat, J. (2012). “Regulation toward Self-Organized Criticality in a Recurrent Spiking Neural Reservoir,” in International Conference on Artificial Neural Networks (Springer), 547–554. doi:10.1007/978-3-642-33269-2_69
Cheney, N., Bongard, J., and Lipson, H. (2015). “Evolving Soft Robots in Tight Spaces,” in Proceedings of the 2015 annual conference on Genetic and Evolutionary Computation, 935–942.
Cheney, N., MacCurdy, R., Clune, J., and Lipson, H. (2013). “Unshackling Evolution: Evolving Soft Robots with Multiple Materials and a Powerful Generative Encoding,” in Proceedings of the 15th annual conference on Genetic and evolutionary computation, 167–174.
Chvykov, P., Berrueta, T. A., Vardhan, A., Savoie, W., Samland, A., Murphey, T. D., et al. (2021). Low Rattling: A Predictive Principle for Self-Organization in Active Collectives. Science 371, 90–95. doi:10.1126/science.abc6182
Clauset, A., Shalizi, C. R., and Newman, M. E. J. (2009). Power-law Distributions in Empirical Data. SIAM Rev. 51, 661–703. doi:10.1137/070710111
Corucci, F., Cheney, N., Giorgio-Serchi, F., Bongard, J., and Laschi, C. (2018). Evolving Soft Locomotion in Aquatic and Terrestrial Environments: Effects of Material Properties and Environmental Transitions. Soft robotics 5, 475–495. doi:10.1089/soro.2017.0055
Drew, J. H., Glen, A. G., and Leemis, L. M. (2000). Computing the Cumulative Distribution Function of the Kolmogorov-Smirnov Statistic. Comput. Stat. Data Anal. 34, 1–15. doi:10.1016/s0167-9473(99)00069-9
Ferigo, A., Iacca, G., and Medvet, E. (2021). “Beyond Body Shape and Brain: Evolving the Sensory Apparatus of Voxel-Based Soft Robots,” in International Conference on the Applications of Evolutionary Computation (Part of EvoStar) (Springer).
Fernando, C., and Sojakka, S. (2003). “Pattern Recognition in a Bucket,” in European conference on artificial life (Springer), 588–597. doi:10.1007/978-3-540-39432-7_63
Gibbons, T. E. (2010). “Unifying Quality Metrics for Reservoir Networks,” in The 2010 International Joint Conference on Neural Networks (IJCNN) (IEEE), 1–7.
Hansen, N., Müller, S. D., and Koumoutsakos, P. (2003). Reducing the Time Complexity of the Derandomized Evolution Strategy with Covariance Matrix Adaptation (Cma-es). Evol. Comput. 11, 1–18. doi:10.1162/106365603321828970
Heiney, K., Huse Ramstad, O., Fiskum, V., Christiansen, N., Sandvig, A., Nichele, S., et al. (2021). Criticality, Connectivity, and Neural Disorder: A Multifaceted Approach to Neural Computation. Front. Comput. Neurosci. 15, 7. doi:10.3389/fncom.2021.611183
Heiney, K., Ramstad, O. H., Sandvig, I., Sandvig, A., and Nichele, S. (2019). “Assessment and Manipulation of the Computational Capacity of In Vitro Neuronal Networks through Criticality in Neuronal Avalanches,” in 2019 IEEE Symposium Series on Computational Intelligence (SSCI) (IEEE), 247–254.
Hejna, D. J., Abbeel, P., and Pinto, L. (2021). Task-agnostic Morphology Evolution. arXiv preprint arXiv:2102.13100.
Hesse, J., and Gross, T. (2014). Self-organized Criticality as a Fundamental Property of Neural Systems. Front. Syst. Neurosci. 8, 166. doi:10.3389/fnsys.2014.00166
Hiller, J., and Lipson, H. (2011). Automatic Design and Manufacture of Soft Robots. IEEE Trans. Robotics 28, 457–466.
Hiller, J., and Lipson, H. (2012). Automatic Design and Manufacture of Soft Robots. IEEE Trans. Robot. 28, 457–466. doi:10.1109/tro.2011.2172702
Horibe, K., Walker, K., and Risi, S. (2021). Regenerating Soft Robots through Neural Cellular Automata. arXiv preprint arXiv:2102.02579.
Jaeger, H. (2002). Adaptive Nonlinear System Identification with echo State Networks. Adv. Neural Inf. Process. Syst. 15, 609–616.
Jaeger, H. (2001). The “echo State” Approach to Analysing and Training Recurrent Neural Networks-With an Erratum Note. Bonn, Germany: German National Research Center for Information Technology GMD Technical Report 148, 13.
Konkoli, Z., Nichele, S., Dale, M., and Stepney, S. (2018). “Reservoir Computing with Computational Matter,” in Computational Matter (Springer), 269–293. doi:10.1007/978-3-319-65826-1_14
Kriegman, S., Cheney, N., and Bongard, J. (2018). How Morphological Development Can Guide Evolution. Sci. Rep. 8, 13934–13943. doi:10.1038/s41598-018-31868-7
Kriegman, S., Blackiston, D., Levin, M., and Bongard, J. (2020a). A Scalable Pipeline for Designing Reconfigurable Organisms. Proc. Natl. Acad. Sci. USA 117, 1853–1859. doi:10.1073/pnas.1910837117
Kriegman, S., Nasab, A. M., Shah, D., Steele, H., Branin, G., Levin, M., et al. (2020b). “Scalable Sim-To-Real Transfer of Soft Robot Designs,” in 2020 3rd IEEE International Conference on Soft Robotics (RoboSoft) (IEEE), 359–366.
Langton, C. (1990). Computation at the Edge of Chaos: Phase Transitions and Emergent Computation. Physica D: Nonlinear Phenomena 42, 12. doi:10.1016/0167-2789(90)90064-V
Legenstein, R., and Maass, W. (2007). Edge of Chaos and Prediction of Computational Performance for Neural Circuit Models. Neural networks 20, 323–334. doi:10.1016/j.neunet.2007.04.017
Li, T., Nakajima, K., Cianchetti, M., Laschi, C., and Pfeifer, R. (2012). “Behavior Switching Using Reservoir Computing for a Soft Robotic Arm,” in 2012 IEEE International Conference on Robotics and Automation (IEEE), 4918–4924.
Lipson, H. (2014). Challenges and Opportunities for Design, Simulation, and Fabrication of Soft Robots. Soft Robotics 1, 21. doi:10.1089/soro.2013.0007
Maass, W., Legenstein, R., and Markram, H. (2002). “A New Approach towards Vision Suggested by Biologically Realistic Neural Microcircuit Models,” in International Workshop on Biologically Motivated Computer Vision (Springer), 282–293. doi:10.1007/3-540-36181-2_28
Mazzolai, B., Margheri, L., Cianchetti, M., Dario, P., and Laschi, C. (2012). Soft-robotic Arm Inspired by the octopus: Ii. From Artificial Requirements to Innovative Technological Solutions. Bioinspir. Biomim. 7, 025005. doi:10.1088/1748-3182/7/2/025005
Medvet, E., Bartoli, A., De Lorenzo, A., and Fidel, G. (2020a). “Evolution of Distributed Neural Controllers for Voxel-Based Soft Robots,” in Proceedings of the 2020 Genetic and Evolutionary Computation Conference, 112–120.
Medvet, E., Bartoli, A., De Lorenzo, A., and Seriani, S. (2020b). 2d-vsr-sim: A Simulation Tool for the Optimization of 2-d Voxel-Based Soft Robots. SoftwareX 12, 100573. doi:10.1016/j.softx.2020.100573
Medvet, E., Bartoli, A., De Lorenzo, A., and Seriani, S. (2020c). Design, Validation, and Case Studies of 2d-Vsr-Sim, an Optimization-Friendly Simulator of 2-d Voxel-Based Soft Robots. arXiv, arXiv–2001.
Medvet, E., and Bartoli, A. (2021). “GraphEA: a Versatile Representation and Evolutionary Algorithm for Graphs,” in Workshop on Evolutionary and Population-based Optimization (Part of AIxIA).
Miras, K., Ferrante, E., and Eiben, A. E. (2020). Environmental Influences on Evolvable Robots. PloS one 15, e0233848. doi:10.1371/journal.pone.0233848
Nagelkerke, N. J. D., et al. (1991). A Note on a General Definition of the Coefficient of Determination. Biometrika 78, 691–692. doi:10.1093/biomet/78.3.691
Nakajima, K., Hauser, H., Kang, R., Guglielmino, E., Caldwell, D. G., and Pfeifer, R. (2013). A Soft Body as a Reservoir: Case Studies in a Dynamic Model of Octopus-Inspired Soft Robotic Arm. Front. Comput. Neurosci. 7, 91. doi:10.3389/fncom.2013.00091
Pontes-Filho, S., Lind, P., Yazidi, A., Zhang, J., Hammer, H., Mello, G. B. M., et al. (2020b). A Neuro-Inspired General Framework for the Evolution of Stochastic Dynamical Systems: Cellular Automata, Random Boolean Networks and echo State Networks towards Criticality. Cogn. Neurodyn. 14, 657–674. doi:10.1007/s11571-020-09600-x
Pontes-Filho, S., Lind, P., Yazidi, A., Zhang, J., Hammer, H., Mello, G. B. M., Sandvig, I., Tufte, G., and Nichele, S. (2020a). “Evodynamic: A Framework for the Evolution of Generally Represented Dynamical Systems and its Application to Criticality,” in International Conference on the Applications of Evolutionary Computation (Part of EvoStar) (Springer), 133–148. doi:10.1007/978-3-030-43722-0_9
Rus, D., and Tolley, M. T. (2015). Design, Fabrication and Control of Soft Robots. Nature 521, 467–475. doi:10.1038/nature14543
Sadeghi, A., Mondini, A., and Mazzolai, B. (2017). Toward Self-Growing Soft Robots Inspired by Plant Roots and Based on Additive Manufacturing Technologies. Soft robotics 4, 211–223. doi:10.1089/soro.2016.0080
Schrauwen, B., Defour, J., Verstraeten, D., and Van Campenhout, J. (2007). “The Introduction of Time-Scales in Reservoir Computing, Applied to Isolated Digits Recognition,” in Proceedings of the 15th european symposium on artificial neural networks, 471–479. doi:10.1007/978-3-540-74690-4_48
Shen, Z., Na, J., and Wang, Z. (2017). A Biomimetic Underwater Soft Robot Inspired by Cephalopod Mollusc. IEEE Robot. Autom. Lett. 2, 2217–2223. doi:10.1109/lra.2017.2724760
Shew, W. L., and Plenz, D. (2013). The Functional Benefits of Criticality in the Cortex. Neuroscientist 19, 88–100. doi:10.1177/1073858412445487
Sui, X., Cai, H., Bie, D., Zhang, Y., Zhao, J., and Zhu, Y. (2020). Automatic Generation of Locomotion Patterns for Soft Modular Reconfigurable Robots. Appl. Sci. 10, 294.
Talamini, J., Medvet, E., Bartoli, A., and De Lorenzo, A. (2019). “Evolutionary Synthesis of Sensing Controllers for Voxel-Based Soft Robots,” in Artificial Life Conference Proceedings (MIT Press), 574–581.
Tanaka, G., Yamane, T., Héroux, J. B., Nakane, R., Kanazawa, N., Takeda, S., et al. (2019). Recent Advances in Physical Reservoir Computing: A Review. Neural Networks 115, 100–123. doi:10.1016/j.neunet.2019.03.005
Wilting, J., and Priesemann, V. (2019). 25 Years of Criticality in Neuroscience - Established Results, Open Controversies, Novel Concepts. Curr. Opin. Neurobiol. 58, 105–111. doi:10.1016/j.conb.2019.08.002
Keywords: reservoir computing, voxel-based soft robot, evolutionary robotics, criticality, adaptability
Citation: Talamini J, Medvet E and Nichele S (2021) Criticality-Driven Evolution of Adaptable Morphologies of Voxel-Based Soft-Robots. Front. Robot. AI 8:673156. doi: 10.3389/frobt.2021.673156
Received: 26 February 2021; Accepted: 27 May 2021;
Published: 17 June 2021.
Edited by:
David Howard, Commonwealth Scientific and Industrial Research Organization (CSIRO), AustraliaReviewed by:
A. E. Eiben, Vrije Universiteit Amsterdam, NetherlandsSam Kriegman, University of Vermont, United States
John Rieffel, Union College, United States
Copyright © 2021 Talamini, Medvet and Nichele. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eric Medvet, ZW1lZHZldEB1bml0cy5pdA==