 Manuel Vieira1,2*
Manuel Vieira1,2* Noelia Ríos3
Noelia Ríos3 Sebastian Muñoz-Duque1,4
Sebastian Muñoz-Duque1,4 Jodanne Pereira1,4
Jodanne Pereira1,4 Rita Carriço5
Rita Carriço5 Marc Fernandez6,7
Marc Fernandez6,7 João G. Monteiro6,7
João G. Monteiro6,7 Miguel Pessanha Pais1,2
Miguel Pessanha Pais1,2 Bernardo R. Quintella1,2
Bernardo R. Quintella1,2 Gonçalo Silva3
Gonçalo Silva3 Rodrigo P. Silva6,7
Rodrigo P. Silva6,7 Paulo J. Fonseca1,8M. Clara P. Amorim1,2
Paulo J. Fonseca1,8M. Clara P. Amorim1,2- 1Departamento de Biologia Animal, Faculdade de Ciências, Universidade de Lisboa, Lisboa, Portugal
- 2MARE - Marine and Environmental Sciences Centre, ARNET - Aquatic Research Network, Universidade de Lisboa, Lisbon, Portugal
- 3MARE - Marine and Environmental Sciences Centre, ARNET - Aquatic Research Network, ISPA, Instituto Universitário, Lisbon, Portugal
- 4IMBRSEA Master Programme, Ghent University, Ghent, Belgium
- 5Direção Regional de Políticas Marítimas da Secretaria Regional do Mar e Pescas, Governo dos Açores, Azores, Portugal
- 6MARE - Marine and Environmental Sciences Centre, ARNET - Aquatic Research Network, Agência Regional para o Desenvolvimento da Investigação Tecnologia e Inovação (ARDITI/Uma), Madeira, Portugal
- 7Faculdade de Ciências da Vida, Universidade da Madeira, Maderia, Portugal
- 8cE3c - Centre for Ecology, Evolution and Environmental Changes & CHANGE - Global Change and Sustainability Institute, Lisbon, Portugal
There is growing evidence that studying aquatic acoustic communities can provide ecologically relevant information. Understanding these communities may offer unique insights into species behaviour and ecology, while consolidating passive acoustic monitoring as a tool for mapping the presence of target species or estimating changes in aquatic biodiversity. Fish can be significant soundscape contributors, but most soniferous fish species are yet to be identified. Here, we crossed information of three key fish acoustic communities in the Lusitanian Province of the Temperate Northern Atlantic (the Madeira archipelago, the Azores archipelago and Arrábida in mainland Portugal) to unveil potential sources of unidentified fish sounds. We found that the three communities shared various sound types and we were able to narrow down the list of possible fish sound sources. Several sound types were suggested to be produced by species of the Pomacentridae, Scorpaenidae and Serranidae families. We also observed that the sound type /kwa/, associated with Scorpaena spp., exhibited more variations in the geographic area where more species of this genus are known to be present. This study showcases that, as databases of unidentified fish sounds continue to grow, future comparisons of multiple acoustic communities may provide insights into unknown fish sound sources and sound types.
1 Introduction
Protecting marine biodiversity is crucial but challenging due to the limitations of traditional survey methods in marine and coastal environments. These methods are costly, weather-dependent, restricted to daytime, and mainly suitable for shallow areas. To improve management and regulatory policies, new methodologies are required to autonomously gather timely and accurate data on complex marine ecosystems (European Commission, 2008; Shephard et al., 2015; Cardoso-Andrade et al., 2022).
Acoustic communities can provide ecologically relevant information about an ecosystem (Farina and James, 2016). Monitoring biological sounds offers complementary insights into species behaviour and ecology, since active fish sounds are produced in ecologically relevant contexts like feeding, breeding, school cohesion, and territorial defence (Amorim, 2006), providing a means to map the presence of target species or to monitor biodiversity changes. While there is a growing amount of data on underwater sounds and their patterns in natural habitats, most sounds in marine environments still lack an identified source (Mouy et al., 2018; Carriço et al., 2019; Bolgan et al., 2022; Looby et al., 2022; Parsons et al., 2022; Raick et al., 2023a). Also, although passive acoustic monitoring (PAM) tools can provide crucial information on marine biophony, their success will be greatly enhanced by the identification of the soniferous species behind each sound type and by the ability to automatically extract relevant information from the complex polyphony of acoustic recordings. This will contribute to drastically reduce data processing times (Muoy et al., 2018; Parsons et al., 2022; Hawkins et al., 2023).
Families with soniferous fish species make up almost two-thirds of actinopterygian species (Rice et al., 2022), often being a dominant component of acoustic communities (e.g., Di Iorio et al., 2018; Vieira et al., 2021). However, it is estimated that only 4% of soniferous fish species have been reported so far (Lobel et al., 2022), mostly based on recordings from captive fishes, and sometimes during manipulation by the researcher (e.g., Fish and Mowbray, 1970; Pereira et al., 2020). These approaches are highly time-consuming, known to potentially change species’ acoustic behaviour (Bolgan et al., 2019), and to underrepresent fishes’ acoustic repertoires. Identifying fish sounds and their sources in nature can be achieved by combining acoustic and video underwater recordings but it is both logistically and time-demanding (Tricas and Boyle, 2014; Muoy et al., 2018; Puebla-Aparicio et al., 2024). Despite the challenges in assigning sounds to fish species, numerous studies are documenting unidentified sounds, given their potential use for marine environmental management and conservation (Raick et al., 2023b; Hawkins et al., 2023).
Using PAM, researchers have described fish acoustic communities by cataloguing a wide range of fish sound types of unknown sources and identifying soniferous and potentially soniferous fish species (e.g., Carriço et al., 2019; Parmentier et al., 2021; Bolgan et al., 2022; Raick et al., 2023a; Puebla-Aparicio et al., 2024). While understanding unidentified sounds can offer insights into biodiversity, ecosystem dynamics, and ecosystem health, there are clear benefits to identifying their sources, including the ability to monitor target species, enhance our understanding of aquatic acoustic diversity, deepen our knowledge of taxonomic biodiversity and ecology, and enable a more comprehensive assessment of biodiversity using PAM (Parsons et al., 2022). The use of PAM in the marine environment is still relatively new and, as a result, we lack the ability to effectively use this data. However, these recordings can provide different types of information depending on the methodology used to analyse the complex data that captures the soundscape of a specific location. Here, we cross-referenced catalogues of fish acoustic communities in three regions within the Lusitanian Province of the Temperate Northern Atlantic (Spalding et al., 2007): the Azores archipelago, the Madeira archipelago and coastal waters of mainland Portugal. The aim was to compare species (soniferous and potentially soniferous fishes) and sound types occurrences, to narrow down possible sound producing candidates.
2 Materials and methods
2.1 Study areas
2.1.1 The Madeira archipelago
The Madeira archipelago (Portugal) is of volcanic origin and belongs to the Macaronesian region, (Azores, Canaries and Madeira Ecoregion; sensu Spalding et al., 2007; Figure 1). This archipelago has two populated islands, Madeira and Porto Santo, and two uninhabited sub-archipelagos, the Desertas Islands and the Selvagens Islands. The Madeira Archipelago has a unique biogeography which favours the presence of fauna by long-distance dispersal from adjacent continental areas (Whittaker and Fernández-Palacios, 2007) or, in certain cases, adjacent archipelagos (Domingues et al., 2008). The recordings considered for the purposes of the present study were obtained, as reported by Muñoz-Duque et al. (2024), in the Garajau Partial Nature Reserve on Madeira Island (32° 38.195′N 16° 51.204′ W, depth = 28 m in a rocky and sandy habitat) and in the Desertas Islands Nature Reserve (32° 30.718′ N 16° 30.500′ W; depth = 16 m in a rocky and sandy habitat) between June and September 2021. Garajau Partial Nature Reserve was created in 1996 and is located on the south-east coast of Madeira Island (Alves et al., 2019; IFCN, 2022b). The Desertas Islands have been legally protected since 1990, and became a Nature Reserve in 1995 (IFCN, 2022a).
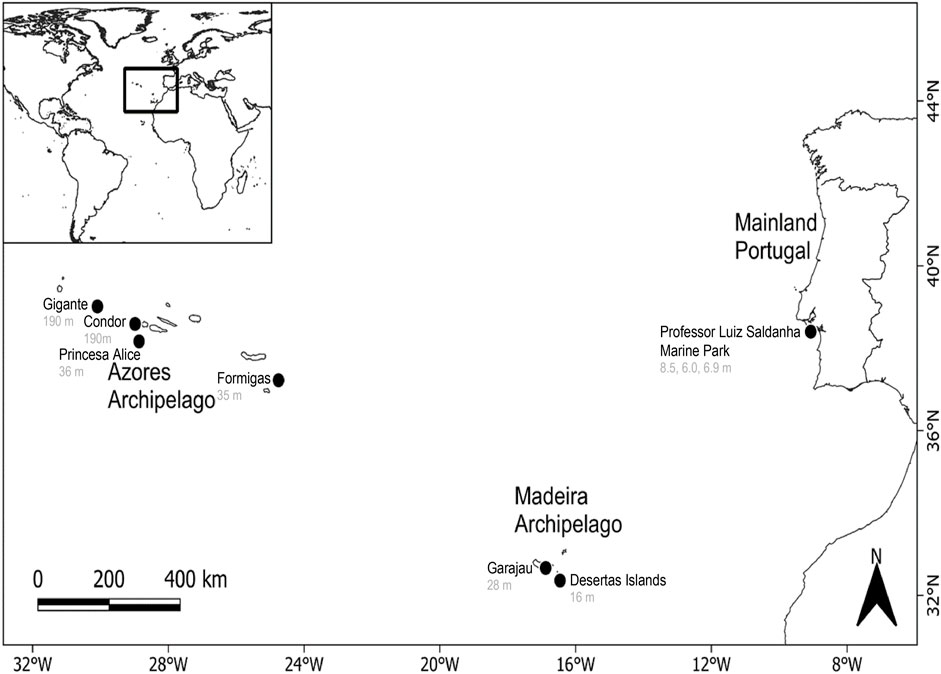
Figure 1. The three main regions of the fish acoustic communities used in this study: Azores and Madeira archipelagos and the Professor Luiz Saldanha Marine Park (Arrábida, mainland Portugal). Recording sites are marked as black circles.
2.1.2 The Azores archipelago
The Macaronesian Azorean archipelago is located in the Temperate Northern Atlantic and comprises nine islands and several seamounts of volcanic origin (Figure 1). It is considered a hotspot of marine life, harbouring many resident and migratory species of cephalopods, fish, marine turtles, sea birds and marine mammals (Morato et al., 2008). The recordings used for this study are from four seamounts distributed across the archipelago (Carriço et al., 2019; Carriço et al., 2020b; Carriço, 2022): Princesa Alice (38° 00.290′ N, 29° 17.917′ W, depth = 36 m), Formigas (37 15.341 N 24 45. 604 W, depth = 35 m), Condor (38° 32.407′ N, 29° 02.627′ W, depth = 190 m), and Gigante (38° 59.252′ N, 29° 52.830’ W, depth = 190 m) (details in Supplementary Table S1). Note that seamounts are considered biodiversity hotspots, with habitats hosting both migratory and resident species (Pitcher et al., 2007).
2.1.3 The Professor Luiz Saldanha Marine Park (Arrábida, mainland Portugal)
The Professor Luiz Saldanha Marine Park, sometimes also referred as Arrábida Marine Park (hereafter named Arrábida), on the west coast of mainland Portugal, is an important biodiversity hotspot with diverse habitats ranging from sandy bottoms to complex rocky shore habitats (Henriques et al., 2007; Figure 1). It is located in a biogeographic transition zone and, as a result, species of Mediterranean and subtropical areas as well as species of northern temperate waters coexist (Henriques et al., 2007; Cunha et al., 2014; Pita et al., 2020). Furthermore, its proximity to the highly productive Sado and Tagus estuaries and submarine canyons contributes to its status as a biodiversity hotspot (Cabeçadas et al., 2004). Recordings were collected on three sites between June 2021 and September 2022 (Pereira, 2023) with increasing levels of protection: one in the complementary protection area (38°25.701′ N, 09°09.907′ W, depth = 8.5 m in a rocky habitat), another in the partially protected area (38°26.146′ N, 009°03.817′ W, depth = 6.9 m in a rocky habitat) and the other in the fully protected area (38°26.787′ N, 09°02.432’ W, depth = 6 m at an interface of a sandy and rocky habitat).
2.2 Cross-referencing of soniferous and potentially soniferous fish species from all locations
The cross-referencing of soniferous and potentially soniferous fish species from all regions was based on the lists reported by Muñoz-Duque et al. (2024; Madeira), Carriço et al. (2019; Azores) and Pereira (2023; Arrábida). The list originally reported in Carriço et al. (2019) was updated with new reports about sound producing species (Looby et al., 2023). The above lists were in turn based on Santos et al. (1997) and Wirtz et al. (2008), for Azores and Madeira respectively. For Arrábida, in mainland Portugal, the list of soniferous species was based on the fish database available on the marine park’s geoportal https://arrabidaparquemarinho.ualg.pt/. Given that chondrichthyan fishes are generally not considered soniferous (Ladich and Winkler, 2017), they were not included. Species with reported sounds were considered soniferous and species belonging to families with soniferous species (but with no reported sounds) were considered potentially soniferous species (Carriço et al., 2019; Parmentier et al., 2021).
The list of soniferous and potentially soniferous fishes of the three geographic regions were compared to find overlapping species. Families with species occurring in more than one region were selected for further analysis. However, sound production can vary substantially according to species. Some families like Pomacentridae, Gobiidae, Sciaenidae, and Batrachoididae are known to have soniferous species (Colleye and Parmentier, 2012; Horvatic et al., 2021; Vieira et al., 2022; Han et al., 2023) while others have been rarely reported (e.g., Kyphosidae and Sphyraenidae, only described in Fish and Mowbray, 1970). Furthermore, some families like the Gobiidae, Blenniidae and Syngnathidae are known to produce sounds with a low source level which makes it challenging to hear them even at a few centimeters away and limits the possibility of being detected using PAM (discussed in Carriço et al., 2020a). In this regard, families with doubtful records of soniferous fish species or species producing low source level sounds were not taken into account. The aim was to only consider the soniferous families most probable to be detected by PAM recordings on the three regions. The depth range and habitat of each species was also considered as it can provide information to exclude or confirm potential sound sources. Figure 2 shows a schematic overview of the pipeline for cross-referencing unidentified fish sound data sets.
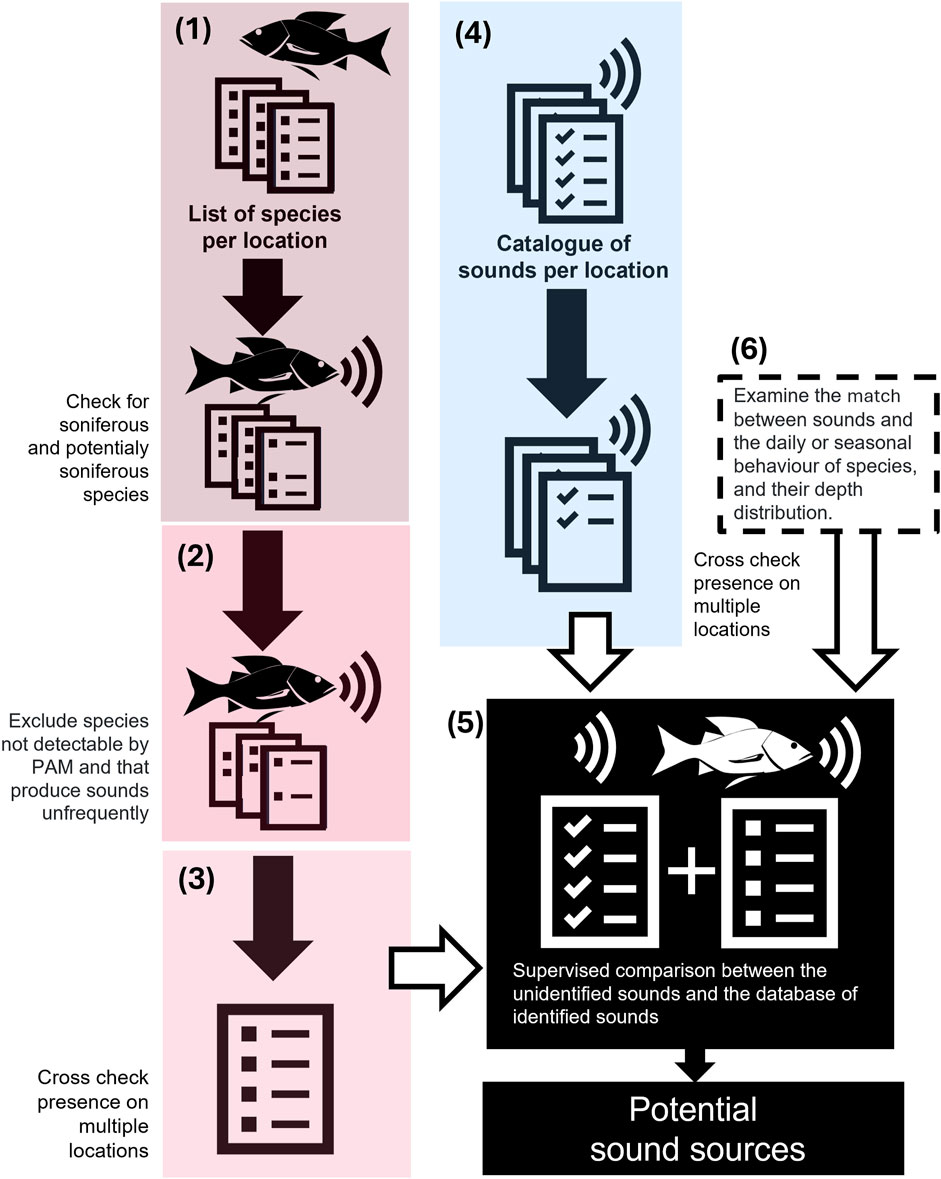
Figure 2. Schematic overview of the pipeline for cross-referencing unidentified fish sound data sets. The pipeline for cross-referencing unidentified fish sound data sets to identify sound sources involves several key steps: (1) Checking previously reported lists of species at each location to identify vocal soniferous and potentially vocal soniferous species. (2) Exclusion of families with doubtful records of soniferous fish species or species producing low source level sounds. (3) Comparison of species lists to narrow down possible sound-producing candidates for sounds occurring at multiple locations. (4) Comparison of sound catalogues from different locations to identify common similar sounds. (5) Comparison of narrowed-down list of sounds with the narrowed list of possible sound sources to find potential sound sources. The focus of the analysis on a smaller number of possibilities should optimize the results obtained. (6) Additionally, matching information about temporal patterns, behavior, or depth between the sounds and the species will strengthen the results. By following this intuitive pipeline, researchers can now optimize the identification of sound sources in putative fish sound data sets. Future tools should enable us to compare new databases of unidentified sounds (e.g., using new machine learning tools to automate the process), match the information on temporal and spatial patterns, and compare the obtained results with known sounds from the list of relevant soniferous fish species.
2.3 Putative fish sound types
2.3.1 Acoustic recordings
Recordings were made using autonomous acoustic loggers at the three regions described in section 2.1. The sites were mainly characterized by rocky and sandy sea floors, with a prevalence of rocky reef fish communities. Loggers were positioned at the seafloor, except for the loggers at the two deeper sites in the Azores that were in the water column, about 8–10 m above the mooring weight. In Madeira and Arrábida, the acoustic loggers were custom-made and had low-cost data loggers (Audiomoth 1.2.0; Hill et al., 2018) connected to custom-made hydrophones (Piezo tubes PTZ-P5 with 24 × 20 × 20 mm, with a signal pre-amplification of 50×). The piezoelectric sensor has a measured response sensitivity of ca. −184.5 dB re 1 V/μPa at 1 kHz, and a frequency response within ±3 dB in the range 150 Hz to 10 kHz. The acoustic loggers recorded WAV audio files at a sampling rate of 48 kHz. In Azores, autonomous Ecological Acoustic Recorders (EARs) were used (Lammers et al., 2008) at a sampling rate of 40 or 50 kHz. EARs use Sensor Technology SQ26-01 hydrophones with a response sensitivity of −193.14/−194.17 dB re 1 V/μPa (varying between deployments) with a reported flat frequency response (±1.5 dB) from 1 Hz to 28 kHz, and a total gain of 47.5 dB. All the recordings were downsampled to 8 kHz, allowing spectrogram inspections for fish sounds up to 4 kHz. The sampling cycle varied between locations (details of the deployments in Supplementary Table S1).
2.3.2 Catalogues of the acoustic fish communities
As mentioned above, this work used annotations and catalogues created for different studies (Carriço et al., 2019; 2020b; Carriço, 2022; Pereira, 2023; Muñoz-Duque et al., 2024). As such, the sampling scheme used differed between locations. Madeira included the analysis of 32 h of acoustic recording (2 h per day × 4 days per month × 2 months × 2 sites), Azores included 48 h (2.4 h per day × 5 days per month × 1 months × 4 sites) and Arrábida included 96 h (2 h per day × 4 days per month × 4 months × 3 sites) (details in Supplementry Table S1). The considered recordings covered different times of day (dawn, day, dusk and night periods). Note that for Azores, however, we updated the catalogue reported by Carriço et al. (2019) using additional data from Carriço et al. (2020b) and Carriço (2022), and reconsidering the sound type units of the originally reported sequences of sounds. These three studies classified putative fish sounds by aural and visual analysis of the recordings (software: Raven Pro 1.6; Adobe Audition 3.0). It is worth noticing that the fish sounds of each catalogue were grouped using dichotomous classification keys to facilitate the organization of the sound types (Desiderà et al., 2019; Raick et al., 2023; Puebla-Aparicio et al., 2024).
2.3.3 Cross-referencing sound types between acoustic communities
By leveraging the expertise of multiple bioacousticians, a qualitative classification and comparison of sound types from different regions was conducted to identify common sound types. All putative fish sound types from the three regions were aurally compared and their spectrograms visually inspected to detect similarities. This analysis was conducted by five researchers who were previosly trained to annotate recordings of these soundscapes. The four researchers responsible for creating each of the catalogues and their annotations (N.R and J.P. for Arrábida; S.M.D. for Madeira and R.C. for Azores) were asked to review the sound types from the other regions and classify them based on their own catalogue. Finally, a discussion led by M.V. was conducted to compile a list of similar and potentially common sound types across regions. A final inspection was done using the following parameters on Raven Pro 1.6: FFT 256 points, Hann window, sampling rate of 8 kHz, temporal range up to 2 s and frequency range up to 1 kHz. Narrowing down the list of sound types and species aimed to optimize the identification of sound sources for the selected fish sounds (see Figure 2).
3 Results
3.1 Cross-referencing lists of soniferous and potentially soniferous species
A total of 212 soniferous and potentially soniferous fish species were identified to occur in the three regions (Supplementary Table S2). The Madeira archipelago, the Azores archipelago and Arrábida, presented 137, 96 and 99 soniferous and potentially soniferous species, respectively. Among these, 14 soniferous and 27 potentially soniferous species occur in all three regions (Supplementary Table S2). As stated in the methods, 37 families were disregarded as probable sources of the detected sound types due to having only a few and doubtful records, producing sounds with low source levels or for only occurring in one region (see Supplementary Table S3). Also, knowing the relative abundance of a species or family can give us an insight into the probability of detecting their sounds. Hence, we only considered families that have a clear presence in the considered regions. By applying these criteria, we can consider the following families as the most probable sources of the sounds detected in more than one region: Balistidae, Haemulidae, Labridae, Lotidae, Mullidae, Ophidiidae, Pomacentridae, Scorpaenidae, Serranidae, Triglidae and Zeidae (Table 1). Table 1 summarises the presence of species from these families for each region. Regarding these 11 selected families, some key differences between fish communities were identified that could be relevant in the identification of sound sources, namely 1) several species of the Labridae family are considered common but none is reported as soniferous and in general there are few descriptions of sounds in this family; 2) Ophidiidae are known to be vocal species, but otherwise cryptic, so their presence and distribution may require additional clarification (Parmentier et al., 2022). 3) within the Pomacentridae family, Chromis limbata and Similiparma lurida are usually the most commonly observed species in both archipelagos (only at shallower waters, depth up to 45 m; and associated to rocky reefs habitats), albeit not present in Arrábida; 4) Azores has a high diversity of Scorpaenidae species and the most common species in both archipelagos is Scorpaena maderensis, while in Arrábida this species is rare and the most common species are Scorpaena notata and Scorpaena porcus; 5) within Serranidae, the dusky grouper Epinephelus marginatus is common in both archipelagos and considered rare in Arrábida, while the island grouper Mycteroperca fusca only occurs in the archipelagos. In all three regions it is easy to encounter species of the genus Serranus - Serranus atricauda is more common in the archipelagos, and Serranus cabrilla is more common in Arrábida; 6) Triglidae species are only common in Arrábida, namely Chelidonichthys lastoviza and Chelidonichthys lucernus, but there are species of this family in all three regions. Note that most species occur in environments with rocky substrate (Table 1).

Table 1. Summary of the soniferous families with species known to produce sounds easily detected with PAM and that are present on at least two of the three analysed fish communities.
3.2 Known characteristics of the sounds produced by species in the selected families
Table 1 summarises some of the major known characteristics that might help suggest sound sources for each sound type. Pomacentridae, Scorpaenidae and Serranidae have some well-characterised sound types, which should be easier to distinguish from sounds produced by other families. Although there may produce other sound types, including sounds yet to be described, Pomacentridae typically produce low frequency fast pulse trains down to single pulse sounds (Santiago and Castro, 1997; Picciulin et al., 2002; Parmentier et al., 2010, see Table 1). Serranidae have several descriptions of low frequency tonal sounds and of short /boom/ sounds (Bertucci et al., 2014b; Wilson et al., 2020). Scorpaenidae emit, for example, a characteristic pulse train with short pulse periods (ca. 13 ms) and a peak frequency around 800 Hz, referred to as /kwa/. (Di Iorio et al., 2018; Bolgan et al., 2019). Furthermore, the presence of several species from each of these families suggests that each soundscape might contain several similar sound types. Note that species of Pomacentridae and Serranidae are also known to produce several sound types like pulse series or downsweeps (e.g., Parmentier et al., 2010: Bertucci et al., 2014b).
3.3 Cross-referencing sound types between acoustic communities
A total of 153 putative fish sound types were identified in the Azores archipelago, the Madeira archipelago and Arrábida in mainland Portugal. Seven groups of similar sounds were identified among regions (Figure 3; Supplementary Table S4 describes each sound type). Note that some sound types are so similar among regions that they could be considered the same (e.g., sounds “1” in Azores and Arrábida as shown in Figure 2A), while others are similar sounds but with significant variations (e.g., sounds “a,” “b,” “c” and “d” in Figure 3C differ in frequency range but are similar otherwise).
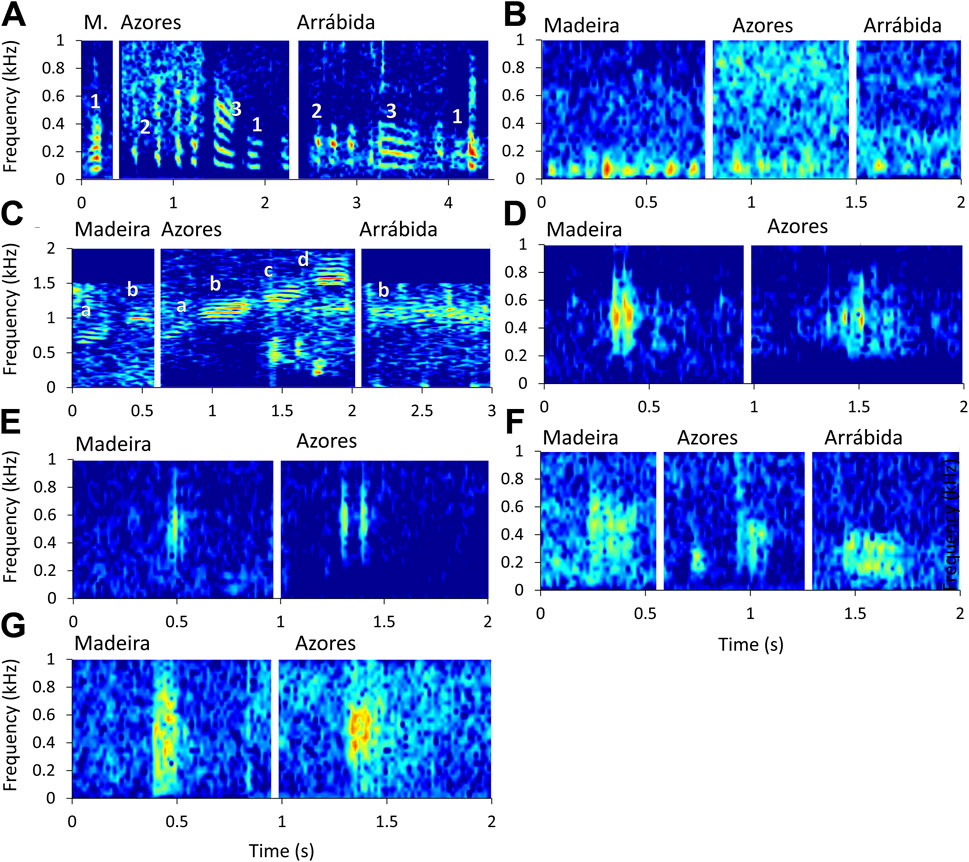
Figure 3. Similar sound types detected in multiple soundscapes. (A) Three tonal sound types detected mostly during the night, similar to sounds reported for subfamily Epinephelinae; (B) Low frequency sound type consistent with the reported serial /boom/ sound produced by Epinephelus marginatus; (C) Four high frequency /kwa/ sound types, similar to sounds reported for the genus Scorpaena; (D, E) Short pulsed sound types consistent with sounds reported for Similiparma lurida; (F) Grunt-like similar sound types encountered in all three regions; (G) two similar “noisy” low frequency sound types encountered in Madeira and Azores.
Figure 3A shows a short tonal sound type that usually occurred in a train (annotated as “2”) and a longer downsweep sound type (annotated as “3”). These sound types occurred in both Azores (in all considered seamounts) and Arrábida. Furthermore, these sound types sometimes occurred as a sequence possibly produced by a single individual. In both regions, these sound types were more common during dusk and night (Carriço et al., 2020a; Pereira, 2023) and are similar to the sounds produced by E. marginatus (listed on Table 1) and other species of the subfamily Epinephelidae, reported by Bertucci et al. (2014a) and Wilson et al. (2020). Additionally, another sound similar to sound type “3” was also detected (annotated as “1” in Figure 3A). This sound type was shorter, had lower frequency components, and could be a variation of sound type “3.” Nevertheless, similar sound types denominated as “1” occur in Madeira, Azores, and Arrábida. Altogether, the sounds characteristics point to these sound types being produced by grouper species of the Epinephelidae subfamily. However, it is currently estimated that very few individuals of E. marginatus and none M. fusca occur in Arrábida (down to a depth of 10 m), which raises the possibility of other species of Serranidae (listed on Table 1) being the source of these sound types. Taking into account that these sounds occurred down to 190 m deep, the other species of this family that occur at those depths are Anthias anthias and S. cabrilla (Table 1). Therefore, these species are candidates for sound “1.”
Figure 3B shows serial /boom/ sounds that were detected in Madeira, Azores, and Arrábida. Similar sound types with frequencies below 100 Hz were previously reported in E. marginatus (Bertucci et al., 2014a). These sounds were rare in all three soundscapes and were also rarely detected by Bertucci et al. (2014a). However, Bertucci et al. (2014a) reports these sounds with dominant frequencies still below the ones detected in these three regions (see Supplementary Table S4), so further investigation is needed to understand if these sounds are produced by this species. Bertucci et al. (2014a) also reports single /boom/s, which are not reported in these three catalogues. However, these single /boom/s can easily be disregarded as abiotic noise, especially when the signal-to-noise ratio is low. Therefore, future work analysing these soundscapes should be aware of these types of sounds that could easily be disregarded.
Figure 3C displays several sounds similar to /kwa/ sound type (Di Iorio et al., 2018; Bolgan et al., 2019; Table 1). It is possible to observe substantial differences in the frequency range, as represented by the examples labelled as “a,” “b,” “c,” and “d” in Figure 3C. In a previous study, Bolgan et al. (2019) used Scorpaena scrofa and S. porcus in a controlled experiment to identify the source of /kwa/ s similar to the ones presented as “a” (with a peak frequency around 800 Hz). Later, Bolgan et al. (2022) observed the presence of /kwa/ s with higher peak frequencies in the Eastern Mediterranean region, which the authors discuss to be due to the higher abundance of S. maderensis. This difference was also observed in a study that compared /kwa/ sounds from Corsica (France) and Sicily (Italy), with peak frequency at ca. 800 Hz and 1,000 Hz, respectively (Raick et al., unpublished data). Moreover, Raick et al. (2023a) suggested that two /kwa/ -like sound types (differing in peak frequency) in French Polynesia are produced by different species of Scorpaenidae. In the current study, we observed /kwa/ s with peak frequencies around 800 Hz, 1,000–1,100 Hz, 1,265 Hz, and 1,570 Hz. The region with the most diversity of /kwa/ s was the Azores (only on the shallower seamounts; depth at ca. 35 m), and Arrábida (depth between 6 and 8 m) presented only a few examples of a /kwa/ with a peak frequency of around 1,000 Hz. Note that in Arrábida this sound type was rare and only detected with a low signal to noise ratio.
In the Azores and Madeira, several short, pulsed sounds occur mostly during the day, and they are the most similar to reported Pomacentridae sound types (Amorim, 1996; Picciulin et al., 2002; Table 1). The sounds represented in Figure 3E are the most similar and might be produced by the same species. These pulsed sounds are one of the most common sound types reported during the day in both regions (Carriço et al., 2020b; Muñoz-Duque et al., 2024). Moreover, these sounds were not detected in deeper water (Carriço et al., 2020b), which is in agreement with the depth range of this family (Table 1). This information, along with the report on S. lurida (Santiago and Castro, 1997), makes this species a plausible candidate for the source of this sound type. Figure 3D also shows sounds consistent with sounds reported by Pomacentridae. Other pulse trains with similarities to those of Pomacentridae were also found in each soundscape, but they had different frequency ranges, dominant frequencies, and temporal patterns, making them distinct from each other and we lack information to suggest plausible sources.
Figure 3F shows three similar sound types but with different duration and frequency ranges. All these sounds are grunt-like sounds. As different families/species produce these types of sounds it is difficult to propose a possible candidate.
Figure 3G shows two similar ‘noisy’ sound types with high entropy and not well-defined pulses. Both have a frequency range of up to approximately 1,000 Hz. For example, Balistidae, Haemulidae and Lotidae species have sound descriptions of toothy grunts and thumps that could be similar and have species on the range of depths where these sounds were recorded (e.g., Fish and Mowbray, 1970; Rountree and Juanes, 2010; Raick et al., 2018; Table 1).
4 Discussion
As databases of passive acoustic recordings and unidentified fish sound types continue to expand, new opportunities arise (Parsons et al., 2022). Comparing multiple fish acoustic communities in the future may offer insights into the sound-producing fish and provide new perspectives on using these data. In the current study, recently catalogued fish acoustic communities from the Temperate Northern Atlantic (Carriço et al., 2019; 2020b; Carriço, 2022; Pereira, 2023; Muñoz-Duque et al., 2024) were compared to potentially uncover sound sources of unidentified putative fish sounds. In this pursuit, several sound types were identified in recordings collected in regions spaced by more than 1,000 km, including sounds similar to those reported for species of the Serranidae, Scorpaenidae and Pomacentridae families.
4.1 Is it possible to find similarities to previously reported fish sounds?
To better understand unidentified sound types an important step is to compare them with documented fish sounds. It is important to gather more reports on fish sound production to determine if certain sounds are unique to specific species and to link sounds with specific contexts, such as reproduction. Currently, only a low percentage of species are reported as soniferous (Lobel et al., 2022). Many of the reported soniferous species were not studied after the 1970s (e.g., Fish and Mowbray, 1970) and although the rate of studies examining fish species for sound production has been steadily increasing it cannot accompany the pace of unknown sound recordings (Parson et al., 2022). Nevertheless, with the limited information available, it is already possible to propose with reasonable confidence potential sources for the sound types encountered in the three fish acoustic communities assessed in the present study.
4.2 How to compare sounds from different soundscapes?
A total of 26 potential fish sound types, from a pool of 153 identified sounds, were categorized into seven groups. However, to identify sound types is a time-consuming approach, that can be subjective even if performed by trained experts, and lacks quantitative characterization of the similarities between sounds. Quantifying these similarities could potentially increase our ability to compare the fish communities and characterize their differences. A common approach to compare sound types is the use of data-reduction statistical techniques, such as principal component analysis (PCA), using traditional spectral and temporal features (Carriço et al., 2019; Odom et al., 2021; Puebla-Aparicio et al., 2024). However, these features may have limitations in their ability to properly represent the properties that a human can use to classify a sound through visual and aural inspection (Carriço et al., 2019; Puebla-Aparicio et al., 2024). Recent advances in machine learning and artificial intelligence, such as deep learning techniques using neural embedding representations, may provide more robust features to discriminate sounds, either based on supervised or self-supervised models (Julião et al., 2022; Stowell, 2022; Williams et al., 2024). The use of these neural embeddings has been successful in representing “fingerprints” of sounds from terrestrial and marine environments (Sethi et al., 2020; Ozanich et al., 2021; Best et al., 2023; Williams et al., 2024), suggesting this methodology as a promising approach. However, developing neural network models for underwater fish sounds faces challenges due to the need for large annotated training sets and difficulties with recordings of varying characteristics (Hamer et al., 2023), especially the typical low signal-to-noise ratio sounds detected with PAM. Accurately defining and grouping unidentified sounds as putative fish sound types remains an open challenge. Maintaining global sound databases, with an increased number of recordings from different species and soundscapes can help increase our knowledge while also improving the performance of these models for a wide range of applications (Parsons et al., 2022). It is thus a critical research priority to develop and test robust methods to compare and quantify similarities between sound types, and even acoustic communities as a whole, from different recordings and soundscapes.
4.3 Which soniferous families should be considered?
More than 200 soniferous and potentially soniferous fish species were identified to occur in the three considered soundscapes. However, not all species have the same potential to be detected in field passive acoustic recordings. Carriço et al. (2020a) highlighted that some species might be detected at greater distances than others. For example, most known Gobiidae and Blenniidae species can only be recorded up to a few centimeters from the hydrophone (de Jong et al., 2007; Lugli and Fine, 2007; Amorim et al., 2018), thus having a low probability of being detected with PAM. But other species, like the Pomacentridae or Scorpaenidae, produce sounds that can be detected several metres away (Carriço et al., 2020a; Bolgan et al., 2022). Furthermore, not every species might produce sounds, at least regularly. Some species of the families Pomacentridae, Sciaenidae or Batrachoididae are known to produce sounds at a substantial calling rate, or even to produce choruses during the breeding season (Mann and Lobel, 1997; Carriço et al., 2020b; Vieira et al., 2021), but other families have been rarely reported to produce sounds (Rice et al., 2022). It is also important to note that some species produce significantly fewer sounds outside the breeding season although they can be very active acoustically during the reproductive period (e.g., Vieira et al., 2021). As such, some families are more likely to be detected than others, and in different times of the year. In this study, the following families were considered the most likely sound sources: Balistidae, Haemulidae, Labridae, Lotidae, Mullidae, Ophidiidae, Pomacentridae, Scorpaenidae, Serranidae, Triglidae and Zeidae. Note that, for example, Scorpaena sp. can now be referred to as highly soniferous but was only identified as soniferous in a recent field study (Bolgan et al., 2019), or that only a few reports exist on deeper environments so far (e.g., Mann and Jarvis, 2004; Carriço et al., 2020a). As such, the exclusion of some families could be revised with the publication of more studies.
4.4 Can we suggest sound sources by crossing information from several soundscapes?
Now-a-days, one of the most challenging tasks in the study of fish acoustic communities is identifying the species responsible for producing the numerous sounds detected using PAM (Lobby et al., 2022; Parsons et al., 2022). It is almost like playing a “Who’s Who?” game, considering not only the sounds described elsewhere but also the presence/absence information about the types of sounds and the list of species responsible for them. However, comparing sound types from different acoustic communities may provide a broader perspective on common sound types and their respective sources. In this study, we compared the list of soniferous fishes and the catalogue of putative fish sounds for three different soundscapes and discussed the observed similarities. We were able to identify certain groups of species as potential sources of several sound types, with some sounds likely produced by Serranidae, Scorpaenidae, and Pomacentridae.
The sounds suggested to be produced by Serranidae (Figures 3A,B) are similar to sounds reported for species of the subfamily Epinephelidae and are mostly produced at night, which is typical of these species (Bertucci et al., 2014a; Wilson et al., 2020). Further studies should focus on validating this possibility. Note that despite the depth variability of the recorded sites among regions some sounds were present both in shallow and deeper seamounts in Azores (Carriço et al., 2020b), highlighting the ubiquitousness of some sound types.
The distinctive /kwa/ sounds reported for species of the Scorpaenidae family (Di Iorio et al., 2018; Bolgan et al., 2022) were detected. Scorpaena spp. are very common in the recording regions of Madeira (recording depths between 16–28 m) and Azores (present on the recordings made at ca. 35 m). In Arrábida (<10 m), although the species are common, they do not appear to be usually present at the sites where the recordings were made. This seems to be reflected in the likelihood to detect /kwa/ sounds, as more of these sounds were detected in Azores and Madeira than in Arrábida. Furthermore, in the Azores, seven species of the genus Scorpaena have been described, which might explain the higher diversity of /kwa/ s encountered. It is worth noting that the most common species in Madeira and Azores is S. maderensis and that the most common /kwa/ in both regions had a peak frequency of around 1,000–1,100 Hz. This is consistent with the findings of Bolgan et al. (2022), who noted an increased prevalence of /kwa/ with a peak frequency of around 1,000–1,100 Hz in a location with a higher abundance of S. maderensis. We are confident that these sounds might be produced by Scorpaena species, but further studies should be conducted to understand if the observed differences are related to the differences between species of this genus.
Two Pomacentridae species are common in the Azores and Madeira (Chromis limbata and S. lurida), but are only found down to 45 m deep (Table 1). Sound types similar to those described for this family are prominent contributors to the soundscapes of the shallow waters of Azores and Madeira, but are not present in deeper waters of Azores (Carriço et al., 2020b; Muñoz-Duque et al., 2024). Double pulsed sound types with a peak frequency of around 500 Hz were found to be similar in both soundscapes. The sound types reported in Figures 3D,F might be produced by the dominant species in Madeira and Azores, C. limbata and S. lurida. In fact, the sound reported in Figure 3D is similar to that reported by Santiago and Castro (1997) for S. lurida. To confirm C. limbata as a sound source, the sounds produced by this species still need to be described (see Table 1). Note that several factors, including temperature, size, and background noise, can influence acoustic parameters, thus complicating comparisons (Parmentier et al., 2016).
4.5 Implications for monitoring programmes
Fish sound types could be used to track the temporal and spatial dynamics of target species, but also to survey their respective communities or habitats. For passive acoustics, specific sounds have been used for detecting and tracking natural fish populations (Luczkovich et al., 1999; Stratoudakis et al., 2024). However, these studies are typically limited to specific regions (Parmentier et al., 2018). An important criterion for using fish sounds as effective indicators of distinct species occurrence is that it should be identifiable or measurable over large geographical scales (Di Iorio et al., 2018; Parmentier et al., 2018). Parmentier et al. (2018) observed that reproductive sounds from a commercial sciaenid species could be unequivocally identified on recordings spanning 17 years and two regions within the Mediterranean Sea, highlighting that PAM can be confidently used to monitor this population and contribute towards its management. Another example of the use of PAM to monitor target fish species comes from studies on groupers. The sounds of groupers have been used to track spawning aggregations and courtship behaviour (Bertucci et al., 2015; Wilson et al., 2020; Desiderà et al., 2022). Since groupers play an important ecological role as top predators, their presence and abundance might provide ecologically relevant information about their communities (Shertzer and Williams, 2008; Hackradt et al., 2014). Also, fish sounds have been proposed as environmental proxies for habitat monitoring (Di Iorio et al., 2018). Within the Mediterranean Sea, Di Iorio et al. (2018) reported /kwa/ sound, produced by Scorpaena species (Bolgan et al., 2019), as an ubiquitous sound type that could be detected on locations covering a distance of more than 200 km. In addition to its wide geographic range and easily identifiable spectral features, it can be detected throughout seasons, is minimally affected by noise interference, and exhibits acoustic diversity. As the sound is associated with seagrass meadows (Posidonia oceanica), these authors proposed that the /kwa/ sound could be tested as an ecological indicator for habitat monitoring, and contribute towards the management of this critical Mediterranean habitat. Further work should be done to validate the source and use of these sound types as reliable indicators to assess the condition of the environment or to diagnose the cause of environmental change (Niemi and McDonald, 2004). Moreover, note that the possibility of tracking a well-characterised group of sound types is a major advantage, as it offers the possibility to use supervised machine learning to automatically annotate them in different soundscapes with lower efforts and higher precision (Barroso et al., 2023).
5 Conclusion
Unidentified underwater biological sounds can provide valuable information about the local acoustic community, serving as a proxy for biodiversity and ecosystem health, even if the source is unknown (Desiderà et al., 2019; Mooney et al., 2020; Raick et al., 2023). As databases of passive acoustic recordings and unidentified fish sound types continue to grow, comparing multiple fish acoustic communities may offer insights into the sound sources and provide new perspectives for research. Moreover, such comparisons should take into account ecological parameters that may drive/restrict the presence/acoustic activity of soniferous species, such as depth of the recording sites and time of day. This study demonstrates how comparing fish acoustic communities from different regions within the Temperate Northern Atlantic can provide a better understanding of possible sources for sounds that occur in more than one region and supports the need for identified and unidentified fish sound libraries (Parsons et al., 2022).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the study reports results using Passive Acoustic Monitoring in marine waters. No animals were handled or harmed.
Author contributions
MV: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. NR: Data curation, Formal Analysis, Investigation, Resources, Writing–review and editing. SM-D: Data curation, Formal Analysis, Investigation, Writing–review and editing. JP: Data curation, Formal Analysis, Investigation, Writing–review and editing. RC: Data curation, Formal Analysis, Investigation, Writing–review and editing. MF: Resources, Writing–review and editing, Funding acquisition. JGM: Resources, Writing–review and editing. MP: Resources, Writing–review and editing, Supervision. BRQ: Resources, Writing–review and editing. GS: Resources, Writing–review and editing, Supervision. RPS: Resources, Writing–review and editing. PJF: Resources, Supervision, Writing–review and editing. MCPA: Conceptualization, Methodology, Resources, Supervision, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was funded by the Science and Technology Foundation, I.P. (FCT), Portugal: strategic projects UID/BIA/00329/2020 granted to CE3C, UIDP/04292/2020 (https://doi.org/10.54499/UIDP/04292/2020) and UIDB/04292/2020 (https://doi.org/10.54499/UIDB/04292/2020) to MARE and LA/P/0069/2020 (https://doi.org/10.54499/LA/P/0069/2020) to the Associate Laboratory ARNET; FCT doctoral scholarship UI/BD/150958/2021 for NR. Likewise, we acknowledge the FCT Scientific Employment Stimulus - Institutional Call - (CEECINST/00037/2021) for funding JGM. and to MARE for funding MF. Similarly, we acknowledge the EU Horizon Europe project CLIMAREST: Coastal Climate Resilience and Marine Restoration Tools for the Arctic Atlantic basin, with grant agreement 101093865 for RS. MPP is funded through FCT and FCUL with researcher contract DL57/2016/CP1479/CT0020. Collection of the Azores archipelago’s dataset was supported by the FCT, Azores 2020 Operational Programme and the Azorean Regional Fund to Science and Technology (FRCT), through research projects TRACE (PTDC/MAR/74071/2006), MAPCET (M2.1.2/F/012/2011), FCT-Exploratory (IF/00943/2013/CP1199/CT0001), WATCH IT (Acores-01-0145-FEDER-000057), and MISTIC SEAS II (GA11.0661/2017/750679/SUB/ENV.C2), through funds from FEDER, COMPETE, QREN, POPH, ESF, the Portuguese Ministry for Science and Education, and EU-DG/ENV. The Azores 2020 Operational Programme is funded by the community structural funds ERDF and ESF. The original catalogue for Azores was also funded by the FRCT in the scope of RC’s doctoral grant (M3.1.a/F/070/2015). Furthermore, this work also received national funds through the FCT under the project UIDB/05634/2020 and UIDP/05634/2020 and through the Regional Government of the Azores through the project M1.1.A/FUNC.UI&D/003/2021-2024. Collection of the Madeira archipelago’s dataset was conducted under MARE Mini Grants 2020 funded through the FCT strategic project UIDP/04292/2020 (https://doi.org/10.54499/UIDP/04292/2020) awarded to MARE, with the partial support of the projects MIMAR+ (MAC2/4.6.d/249) and PLASMAR+ (MAC2/1.1a/347) in the framework of the INTERREG MAC 2014–2020 Program. This work was also conducted in the framework of CoastNet Research Infrastructure, funded by FCT and the European Regional Development Fund (FEDER), through LISBOA2020 and ALENTEJO2020 regional operational programs (PINFRA/22128/2016). Collection of the Professor Luiz Saldanha Marine Park’s dataset was conducted with the support of the Project LIFE06NAT/P/192 (BIOMARES), funded by the European Union LIFE Program and co-funded by the cement company SECIL, Companhia de Cal e Cimentos S.A., Portugal (0000192-INFORBIOMARES POSEUR-03-2215-FC-000047), and the logistic support of the Institute for Nature Conservation and Forests (ICNF) for making the boat available for field trips. For Arrábida in mainland Portugal, the fauna reported by BIOMARES-BIOMARES 2.0 LIFE 06 NAT/P/0000192-INFORBIOMARES POSEUR-03-2215-FC-000047 project was used.
Acknowledgments
We thank Mónica A. Silva, Pedro Afonso and Gui M. Menezes for their help and support to the creation of Azores catalogue. We also thank Inês Avó Nobre, Carlos “Lobo-do-mar,” Mariana Coxey, Maria José Jimenez, Mario Rolim, Sylvie Dias, Friederike Peiffer for all the help with Arrábida’s recordings, and acknowledge the company “Madeira SeaEmotions” for their logistic support in the collection of Madeira’s recordings. Recordings used on this study were carried out in marine protected areas with the permission of IFCN—Instituto das Florestas e ConservaFção da Natureza and ICNF—Instituto da Conservação da Natureza e das Florestas. We also thank the reviewers for providing helpful comments that greatly improved the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frsen.2024.1377206/full#supplementary-material
References
Afonso, P. (2016). Underwater fish visual census in the Azores from 1997 to 2015 Portugal: Institute of Marine Research (IMAR - Azores), Portugal; Department of Oceanography and Fisheries (DOP) - UAc. doi:10.14284/210
Almada, V. C., Amorim, M. C. P., Pereira, E., Almada, F., Matos, R., and Godinho, R. (1996). Agonistic behaviour and sound production in Gaidropsarus mediterraneus,(Gadidae). J. Fish Biol. 49 (2), 363–366. doi:10.1111/j.1095-8649.1996.tb00031.x
Alves, F., Canning-Clode, J., Ribeiro, C., Gestoso, I., and Kaufmann, M. (2019). Local benthic assemblages in shallow rocky reefs find refuge in a marine protected area at Madeira Island. J. Coast. Conservation 23 (2), 373–383. doi:10.1007/s11852-018-0669-y
Amorim, M. C. P. (1996). Acoustic communication in triglids and other fishes. Aberdeen, Scotland, United Kingdom: Ph.D. Thesis, University of Aberdeen. Ph.D. Thesis.
Amorim, M. C. P. (2006). “Diversity of sound production in fish,” in Communication in fishes. Editors F. Ladich, S. P. Collin, P. Moller, and B. G. Kapoor (Enfield: Science Publishers), Vol. 1, 71–104.
Amorim, M. C. P., and Hawkins, A. D. (2000). Growling for food: acoustic emissions during competitive feeding of the streaked gurnard. J. Fish Biol. 57 (4), 895–907. doi:10.1006/jfbi.2000.1356
Amorim, M. C. P., Simões, J. M., and Fonseca, P. J. (2008b). Acoustic communication in the Lusitanian toadfish, Halobatrachus didactylus: evidence for an unusual large vocal repertoire. J. Mar. Biol. Assoc. U. K. 88 (5), 1069–1073. doi:10.1017/S0025315408001677
Amorim, M. C. P., Simões, J. M., Fonseca, P. J., and Turner, G. F. (2008a). Species differences in courtship acoustic signals among five Lake Malawi cichlid species (Pseudotropheus spp.). J. Fish Biol. 72 (6), 1355–1368. doi:10.1111/j.1095-8649.2008.01802.x
Amorim, M. C. P., Stratoudakis, Y., and Hawkins, A. D. (2004). Sound production during competitive feeding in the grey gurnard. J. Fish Biol. 65 (1), 182–194. doi:10.1111/j.0022-1112.2004.00443.x
Amorim, M. C. P., Vasconcelos, R. O., Bolgan, M., Pedroso, S. S., and Fonseca, P. J. (2018). Acoustic communication in marine shallow waters: testing the acoustic adaptive hypothesis in sand gobies. J. Exp. Biol. 221 (22), jeb183681. doi:10.1242/jeb.183681
Assis, J. (2022). Biodiversity of Luiz Saldanha Marine Park. Version 1.7 (Centre of Marine Sciences). doi:10.15468/fkwxdcaccessedviaGBIF.orgon2023-11-29
Barroso, V. R., Xavier, F. C., and Ferreira, C. E. L. (2023). Applications of machine learning to identify and characterize the sounds produced by fish. ICES J. Mar. Sci. 80 (7), 1854–1867. doi:10.1093/icesjms/fsad126
Bertucci, F., Lejeune, P., Payrot, J., and Parmentier, E. (2014a). Sound production by dusky grouper Epinephelus marginatus at spawning aggregation sites. J. Fish Biol. 87 (2), 400–421. doi:10.1111/jfb.12733
Bertucci, F., Ruppé, L., Van Wassenbergh, S., Compére, P., and Parmentier, E. (2014b). New insights into the role of the pharyngeal jaw apparatus in the sound-producing mechanism of Haemulon flavolineatum (Haemulidae). J. Exp. Biol. 217 (21), 3862–3869. doi:10.1242/jeb.109025
Best, P., Paris, S., Glotin, H., and Marxer, R. (2023). Deep audio embeddings for vocalisation clustering. Plos one 18 (7), e0283396. doi:10.1371/journal.pone.0283396
Bolgan, M., Di Iorio, L., Dailianis, T., Catalan, I. A., Lejeune, P., Picciulin, M., et al. (2022). Fish acoustic community structure in Neptune seagrass meadows across the Mediterranean basin. Aquatic Conservation Mar. Freshw. Ecosyst. 32 (2), 329–347. doi:10.1002/aqc.3764
Bolgan, M., Soulard, J., Di Iorio, L., Gervaise, C., Lejeune, P., Gobert, S., et al. (2019). Sea chordophones make the mysterious/Kwa/sound: identification of the emitter of the dominant fish sound in Mediterranean seagrass meadows. J. Exp. Biol. 222 (11), jeb196931. doi:10.1242/jeb.196931
Boyle, K. S., and Cox, T. E. (2009). Courtship and spawning sounds in bird wrasse Gomphosus varius and saddle wrasse Thalassoma duperrey. J. Fish Biol. 75 (10), 2670–2681. doi:10.1111/j.1095-8649.2009.02459.x
Cabeçadas, G., Monteiro, M. T., Brogueira, M. J., Guerra, M., Gaudêncio, M. J., Passos, M., et al. (2004). Caracterização ambiental da zona costeira adjacente aos estuários do Tejo e do Sado. Relatórios Científicos e Técnicos IPIMAR. Série Digit. 20, 40.
Cardoso-Andrade, M., Queiroga, H., Rangel, M., Sousa, I., Belackova, A., Bentes, L., et al. (2022). Setting performance indicators for coastal marine protected areas: an expert-based methodology. Front. Mar. Sci. 9, 848039. doi:10.3389/fmars.2022.848039
Carriço, R., Silva, M. A., Menezes, G. M., Fonseca, P. J., and Amorim, M. C. P. (2019). Characterization of the acoustic community of vocal fishes in the Azores. PeerJ 7, e7772. doi:10.7717/peerj.7772
Carriço, R., Silva, M. A., Menezes, G. M., Vieira, M., Bolgan, M., Fonseca, P. J., et al. (2020a). Temporal dynamics in diversity patterns of fish sound production in the Condor seamount (Azores, NE Atlantic). Deep Sea Res. Part I Oceanogr. Res. Pap. 164, 103357. doi:10.1016/j.dsr.2020.103357
Carriço, R., Silva, M. A., Vieira, M., Afonso, P., Menezes, G. M., Fonseca, P. J., et al. (2020b). The use of soundscapes to monitor fish communities: meaningful graphical representations differ with acoustic environment. Acoustics 2 (2), 382–398. doi:10.3390/acoustics2020022
Carriço, R. A. O. (2022). “Passive acoustic monitoring as a tool to monitor habitat health and vocal fish species of commercial or conservation interest,” in PhD’s thesis, doctoral degree in marine sciences - university of the Azores, Portugal.
Colleye, O., and Parmentier, E. (2012). Overview on the diversity of sounds produced by clownfishes (Pomacentridae): importance of acoustic signals in their peculiar way of life. PLoS One 7 (11), e49179. doi:10.1371/journal.pone.0049179
Cunha, A. H., Erzini, K., Serrão, E. A., Gonçalves, E., Borges, R., Henriques, M., et al. (2014). Biomares, a LIFE project to restore and manage the biodiversity of prof. Luiz Saldanha marine park. J. Coast. Conservation 18 (6), 643–655. doi:10.1007/s11852-014-0336-x
De Jong, K., Bouton, N., and Slabbekoorn, H. (2007). Azorean rock-pool blennies produce size-dependent calls in a courtship context. Anim. Behav. 74 (5), 1285–1292. doi:10.1016/j.anbehav.2007.02.023
Desiderà, E., Guidetti, P., Panzalis, P., Navone, A., Valentini-Poirrier, C. A., Boissery, P., et al. (2019). Acoustic fish communities: sound diversity of rocky habitats reflects fish species diversity. Mar. Ecol. Prog. Ser. 608, 183–197. doi:10.3354/meps12812
Desiderà, E., Mazzoldi, C., Navone, A., Panzalis, P., Gervaise, C., Guidetti, P., et al. (2022). Reproductive behaviours and potentially associated sounds of the mottled grouper Mycteroperca rubra: implications for conservation. Diversity 14 (5), 318. doi:10.3390/d14050318
Di Iorio, L., Raick, X., Parmentier, E., Boissery, P., Valentini-Poirier, C. A., and Gervaise, C. (2018). Posidonia meadows calling’: a ubiquitous fish sound with monitoring potential. Remote Sens. Ecol. Conservation 4 (3), 248–263. doi:10.1002/rse2.72
Domingues, V. S., Alexandrou, M., Almada, V. C., Robertson, D. R., Brito, A., Santos, R. S., et al. (2008). Tropical fishes in a temperate sea: evolution of the wrasse Thalassoma pavo and the parrotfish Sparisoma cretense in the Mediterranean and the adjacent Macaronesian and Cape Verde Archipelagos. Mar. Biol. 154, 465–474. doi:10.1007/s00227-008-0941-z
European Commission (2008). Directive 2008/56/EC of the European parliament and of the council of 17 June 2008 establishing a framework for community actions in the field of marine environmental policy (marine strategy framework directive - MSFD). Official J. Eur. Communities L164/19.
Farina, A., and James, P. (2016). The acoustic communities: definition, description and ecological role. Biosystems 147, 11–20. doi:10.1016/j.biosystems.2016.05.011
Fish, M. P., and Mowbray, W. H. (1970). Sounds of western north atlantic fishes, A reference file of biological underwater sounds. Baltimore, MD: The Johns Hopkins Press.
Hackradt, C. W., García-Charton, J. A., Harmelin-Vivien, M., Perez-Ruzafa, A., Le Direach, L., Bayle-Sempere, J., et al. (2014). Response of rocky reef top predators (Serranidae: Epinephelinae) in and around marine protected areas in the Western Mediterranean Sea. PLoS One 9 (6), e98206. doi:10.1371/journal.pone.0098206
Hamer, J., Triantafillou, E., van Merrienboer, B., Kahl, S., Klinck, H., Denton, T., et al. (2023). BIRB: a generalization benchmark for information retrieval in bioacoustics. arXiv preprint arXiv:2312.07439.
Han, S. M., Land, B. R., Bass, A. H., and Rice, A. N. (2023). Sound production biomechanics in three-spined toadfish and potential functional consequences of swim bladder morphology in the Batrachoididae. J. Acoust. Soc. Am. 154 (5), 3466–3478. doi:10.1121/10.0022386
Hawkins, L. A., Saunders, B. J., Landero Figueroa, M. M., McCauley, R. D., Parnum, I. M., Parsons, M. J., et al. (2023). Habitat type drives the spatial distribution of Australian fish chorus diversity. J. Acoust. Soc. Am. 154 (4), 2305–2320. doi:10.1121/10.0021330
Henriques, M., Gonçalves, E., and Almada, V. (2007). Rapid shifts in a marine fish assemblage follow fluctuations in winter sea conditions. Mar. Ecol. Prog. Ser. 340, 259–270. doi:10.3354/meps340259
Hill, A. P., Prince, P., Piña Covarrubias, E., Doncaster, C. P., Snaddon, J. L., and Rogers, A. (2018). AudioMoth: evaluation of a smart open acoustic device for monitoring biodiversity and the environment. Methods Ecol. Evol. 9 (5), 1199–1211. doi:10.1111/2041-210X.12955
Horvatić, S., Malavasi, S., Vukić, J., Šanda, R., Marčić, Z., Ćaleta, M., et al. (2021). Correlation between acoustic divergence and phylogenetic distance in soniferous European gobiids (Gobiidae; Gobius lineage). Plos one 16 (12), e0260810. doi:10.1371/journal.pone.0260810
IFCN (2022a). Reserva Natural das Ilhas Desertas. Available at: https://ifcn.madeira.gov.pt/areas-protegidas/ilhas-desertas.html.
IFCN (2022b). Reserva natural parcial do Garajau. Available at: https://ifcn.madeira.gov.pt/areas-protegidas/garajau.html.
Julião, M., Abad, A., and Moniz, H. (2022). Can prosody transfer embeddings be used for prosody assessment? Proc. Speech Prosody 2022, 292–296. doi:10.21437/SpeechProsody.2022-60
Ladich, F., and Winkler, H. (2017). Acoustic communication in terrestrial and aquatic vertebrates. J. Exp. Biol. 220 (13), 2306–2317. doi:10.1242/jeb.132944
Lammers, M., Brainard, R., Whitlow, W., Mooney, T., and Wong, K. (2008). An Ecological Acoustic Recorder (EAR) for long-term monitoring of biological and anthropogenic sounds on coral reefs and other marine habitats. J. Acoust. Soc. Am. 123, 1720–1728. doi:10.1121/1.2836780
Looby, A., Cox, K., Bravo, S., Rountree, R., Juanes, F., Reynolds, L. K., et al. (2022). A quantitative inventory of global soniferous fish diversity. Rev. Fish Biol. Fish. 32 (2), 581–595. doi:10.1007/s11160-022-09702-1
Looby, A., Vela, S., Cox, K., Riera, A., Bravo, S., Davies, H. L., et al. (2023). FishSounds Version 1.0: a website for the compilation of fish sound production information and recordings. Ecol. Inf. 74, 101953. doi:10.1016/j.ecoinf.2022.101953
Luczkovich, J. J., Sprague, M. W., Johnson, S. E., and Pullinger, R. C. (1999). Delimiting spawning areas of weakfish, Cynoscion regalis (family Sciaenidae) in Pamlico Sound, North Carolina using passive hydroacoustic surveys. Bioacoustics 10, 143–160. doi:10.1080/09524622.1999.9753427
Lugli, M., and Fine, M. L. (2007). Stream ambient noise, spectrum and propagation of sounds in the goby Padogobius martensii: sound pressure and particle velocity. J. Acoust. Soc. Am. 122 (5), 2881–2892. doi:10.1121/1.2783113
Mann, D. A., and Jarvis, S. M. (2004). Potential sound production by a deep-sea fish. J. Acoust. Soc. Am. 115 (5), 2331–2333. doi:10.1121/1.1694992
Mann, D. A., and Lobel, P. S. (1997). Propagation of damselfish (Pomacentridae) courtship sounds. J. Acoust. Soc. Am. 101 (6), 3783–3791. doi:10.1121/1.418425
Mooney, T. A., Di Iorio, L., Lammers, M., Lin, T. H., Nedelec, S. L., Parsons, M., et al. (2020). Listening forward: approaching marine biodiversity assessments using acoustic methods. R. Soc. open Sci. 7 (8), 201287. doi:10.1098/rsos.201287
Morato, T., Varkey, D. A., Dâmaso, C., Machete, M., Santos, M., Prieto, R., et al. (2008). Evidence of a seamount effect on aggregating visitors. Mar. Ecol. Prog. Ser. 357, 23–32. doi:10.3354/meps07269
Mouy, X., Rountree, R., Juanes, F., and Dosso, S. E. (2018). Cataloging fish sounds in the wild using combined acoustic and video recordings. J. Acoust. Soc. Am. 143 (5), EL333–EL339. doi:10.1121/1.5037359
Muñoz-Duque, S. E., Fonseca, P., Quintella, B., Monteiro, J. G., Fernandez, M., Silva, R., et al. (2024). Acoustic fish community in the Madeira Archipelago (North Atlantic Ocean): characterization of sound diversity and daily patterns. Mar. Environ. Res. 10660. doi:10.1016/j.marenvres.2024.106600
Niemi, G. J., and McDonald, M. E. (2004). Application of ecological indicators. Annu. Rev. Ecol. Evol. Syst. 35, 89–111. doi:10.1146/annurev.ecolsys.35.112202.130132
Odom, K. J., Araya-Salas, M., Morano, J. L., Ligon, R. A., Leighton, G. M., Taff, C. C., et al. (2021). Comparative bioacoustics: a roadmap for quantifying and comparing animal sounds across diverse taxa. Biol. Rev. 96 (4), 1135–1159. doi:10.1111/brv.12695
Ozanich, E., Thode, A., Gerstoft, P., Freeman, L. A., and Freeman, S. (2021). Deep embedded clustering of coral reef bioacoustics. J. Acoust. Soc. Am. 149 (4), 2587–2601. doi:10.1121/10.0004221
Parmentier, E., Bertucci, F., Bolgan, M., and Lecchini, D. (2021). How many fish could be vocal? An estimation from a coral reef (Moorea Island). Belg. J. Zoology 151. doi:10.26496/bjz.2021.82
Parmentier, E., Bouillac, G., Dragičević, B., Dulčić, J., and Fine, M. (2010). Call properties and morphology of the sound-producing organ in Ophidion rochei (Ophidiidae). J. Exp. Biol. 213 (18), 3230–3236. doi:10.1242/jeb.044701
Parmentier, E., Di Iorio, L., Picciulin, M., Malavasi, S., Lagardère, J. P., and Bertucci, F. (2018). Consistency of spatiotemporal sound features supports the use of passive acoustics for long-term monitoring. Anim. Conserv. 21 (3), 211–220. doi:10.1111/acv.12362
Parmentier, E., Gaëlle, S., Renaud, B., Fine, M. L., Loïc, K., Iorio, L. D., et al. (2022). Sound production and mechanism in the cryptic cusk-eel Parophidion vassali. J. Anat. 241 (3), 581–600. doi:10.1111/joa.13691
Parmentier, E., Kéver, L., Casadevall, M., and Lecchini, D. (2010). Diversity and complexity in the acoustic behaviour of Dacyllus flavicaudus (Pomacentridae). Mar. Biol. 157, 2317–2327. doi:10.1007/s00227-010-1498-1
Parmentier, E., Lecchini, D., and Mann, D. A. (2016). “Sound production in damselfishes,” in Biology of damselfishes (Boca Raton: CRC Press), 204–228.
Parsons, M. J., Lin, T. H., Mooney, T. A., Erbe, C., Juanes, F., Lammers, M., et al. (2022). Sounding the call for a global library of underwater biological sounds. Front. Ecol. Evol. 10, 39. doi:10.3389/fevo.2022.810156
Pereira, B. P., Vieira, M., Pousão-Ferreira, P., Candeias-Mendes, A., Barata, M., Fonseca, P. J., et al. (2020). Sound production in the Meagre, Argyrosomus regius (Asso, 1801): intraspecific variability associated with size, sex and context. PeerJ 8, e8559. doi:10.7717/peerj.8559
Pereira, J. C. L. (2023). “Spatial and temporal patterns of fish acoustics in a marine protected area: the professor luis Saldanha marine park,” in International master of science in marine biological Resources – IMBRSea (Belgium: Ghent University).
Picciulin, M., Costantini, M., Hawkins, A. D., and Ferrero, E. A. (2002). Sound emissions of the mediterranean damselfish Chromis chromis (Pomacentridae). Bioacoustics 12 (2-3), 236–238. doi:10.1080/09524622.2002.9753707
Pita, C., Horta e Costa, B., Franco, G., Coelho, R., Sousa, I., Gonçalves, E. J., et al. (2020). Fisher’s perceptions about a marine protected area over time. Aquac. Fish. 5 (5), 273–281. doi:10.1016/j.aaf.2020.01.005
Pitcher, T., Morato, T., Hart, P., Clark, M., Haggan, N., and Santos, R. (2007). Seamounts: ecology, fisheries and conservation, fish and aquatic resource series, 12. Oxford: Blackwell.
Puebla-Aparicio, M., Ascencio-Elizondo, C., Vieira, M., Amorim, M. C. P., Duarte, R., and Fonseca, P. J. (2024). Characterization of the fish acoustic communities in a Mozambican tropical coral reef. Mar. Ecol. Prog. Ser., 14450. doi:10.3354/meps14450
Radford, C. A., Putland, R. L., and Mensinger, A. F. (2018). Barking mad: the vocalisation of the john dory, zeus faber. PloS one 13 (10), e0204647. doi:10.1371/journal.pone.0204647
Raick, X., Collet, P., Lecchini, D., Bertucci, F., and Parmentier, E.Under The Pole Consortium (2023b). Diel cycle of two recurrent fish sounds from mesophotic coral reefs. Sci. Mar. 87 (4), e078. doi:10.3989/scimar.05395.078
Raick, X., Di Iorio, L., Lecchini, D., Gervaise, C., Hédouin, L., Under The, P. C., et al. (2023a). Fish sounds of photic and mesophotic coral reefs: variation with depth and type of island. Coral Reefs 42 (2), 285–297. doi:10.1007/s00338-022-02343-7
Raick, X., Lecchini, D., Kéver, L., Colleye, O., Bertucci, F., and Parmentier, É. (2018). Sound production mechanism in triggerfish (Balistidae): a synapomorphy. J. Exp. Biol. 221 (1), jeb168948. doi:10.1242/jeb.168948
Rice, A. N., Farina, S. C., Makowski, A. J., Kaatz, I. M., Lobel, P. S., Bemis, W. E., et al. (2022). Evolutionary patterns in sound production across fishes. Ichthyology Herpetology 110 (1), 1–12. doi:10.1643/i2020172
Rountree, R., and Bowers-Altman, J. (2002). Soniferous behaviour of the striped cusk-eel Ophidion marginatum. Bioacoustics 12 (2-3), 240–242. doi:10.1080/09524622.2002.9753709
Rountree, R. A., and Juanes, F. (2010). First attempt to use a remotely operated vehicle to observe soniferous fish behavior in the Gulf of Maine, Western Atlantic Ocean. Curr. Zool. 56 (1), 90–99. doi:10.1093/czoolo/56.1.90
Santiago, J. A., and Castro, J. J. (1997). Acoustic behaviour of Abudefduf luridus. J. Fish Biol. 51 (5), 952–959. doi:10.1006/jfbi.1997.0499
Santos, R. S., Porteiro, F. M., and Barreiros, J. P. (1997). Marine Fishes of the Azores annotated checklist and bibliography. Portugal: Universidade dos Açores.
Sethi, S. S., Jones, N. S., Fulcher, B. D., Picinali, L., Clink, D. J., Klinck, H., et al. (2020). Characterizing soundscapes across diverse ecosystems using a universal acoustic feature set. Proc. Natl. Acad. Sci. 117 (29), 17049–17055. doi:10.1073/pnas.2004702117
Shephard, S., Greenstreet, S. P., Piet, G. J., Rindorf, A., and Dickey-Collas, M. (2015). Surveillance indicators and their use in implementation of the marine strategy framework directive. ICES J. Mar. Sci. 72 (8), 2269–2277. doi:10.1093/icesjms/fsv131
Shertzer, K. W., and Williams, E. W. (2008). Fish assemblages and indicator species: reef fishes off the southeastern, Fishery Bulletin United States 106, 3.
Spalding, M. D., Fox, H. E., Allen, G. R., Davidson, N., Ferdaña, Z. A., Finlayson, M. A. X, et al. (2007). Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. Biosci. 57 (7), 573–583. doi:10.1641/B570707
Stowell, D. (2022). Computational bioacoustics with deep learning: a review and roadmap. PeerJ 10, e13152. doi:10.7717/peerj.13152
Stratoudakis, Y., Vieira, M., Marques, J. P., Amorim, M. C. P., Fonseca, P. J., and Quintella, B. R. (2024). Long-Term passive acoustic monitoring to support adaptive management in a sciaenid fishery (Tagus estuary, Portugal). Rev. Fish Biol. Fish. 34 (1), 491–510. doi:10.1007/s11160-023-09825-z
Tricas, T. C., and Boyle, K. S. (2014). Acoustic behaviors in Hawaiian coral reef fish communities. Mar. Ecol. Prog. Ser. 511, 1–16. doi:10.3354/meps10930
Vieira, M., Amorim, M. C. P., Marques, T. A., and Fonseca, P. J. (2022). Temperature mediates chorusing behaviour associated with spawning in the sciaenid Argyrosomus regius. Mar. Ecol. Prog. Ser. 697, 109–124. doi:10.3354/meps14128
Vieira, M., Fonseca, P. J., and Amorim, M. C. P. (2021). Fish sounds and boat noise are prominent soundscape contributors in an urban European estuary. Mar. Pollut. Bull. 172, 112845. doi:10.1016/j.marpolbul.2021.112845
Whittaker, R. J., and Fernández-Palacios, J. M. (2007). Island biogeography: ecology, evolution, and conservation. Oxford University Press.
Williams, B., Belvanera, S. M., Sethi, S. S., Lamont, T. A., Jompa, J., Prasetya, M., et al. (2024). Unlocking the soundscape of coral reefs with artificial intelligence: pretrained networks and unsupervised learning win out. bioRxiv 2024-02. doi:10.1101/2024.02.02.578582
Wilson, K. C., Semmens, B. X., Pattengill-Semmens, C. V., and McCoy, C. (2020). Potential for grouper acoustic competition and partitioning at a multispecies spawning site off Little Cayman, Cayman Islands. Mar. Ecol. Prog. Ser. 634, 127–146. doi:10.3354/meps13181
Keywords: soundscape ecology, unidentified fish sounds, Portugal, passive acoustic monitoring, bioacoustics, fish communities
Citation: Vieira M, Ríos N, Muñoz-Duque S, Pereira J, Carriço R, Fernandez M, Monteiro JG, Pais MP, Quintella BR, Silva G, Silva RP, Fonseca PJ and Amorim MCP (2024) Cross-referencing unidentified fish sound data sets to unravel sound sources: a case study from the Temperate Northern Atlantic. Front. Remote Sens. 5:1377206. doi: 10.3389/frsen.2024.1377206
Received: 26 January 2024; Accepted: 16 July 2024;
Published: 07 August 2024.
Edited by:
Lucia Di Iorio, UMR5110 Centre de Formation et de Recherche surles Environnements Méditerranéens (CEFREM), FranceCopyright © 2024 Vieira, Ríos, Muñoz-Duque, Pereira, Carriço, Fernandez, Monteiro, Pais, Quintella, Silva, Silva, Fonseca and Amorim. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel Vieira, bWF2aWVpcmFAY2llbmNpYXMudWxpc2JvYS5wdA==